Back to Journals » International Journal of Nanomedicine » Volume 21
Nanoparticles Targeting the Tumor Microenvironment for the Treatment of Osteosarcoma: Recent Progress and Perspectives
Authors Liang G, Wang W, Li C, Zhong B, Zhao L, Zhang Z, Liu J
Received 5 November 2025
Accepted for publication 6 February 2026
Published 16 February 2026 Volume 2026:21 579152
DOI https://doi.org/10.2147/IJN.S579152
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Eng San Thian
Guohui Liang,1 Wei Wang,1 Chao Li,2 Binlong Zhong,1 Lei Zhao,1 Zhicai Zhang,1 Jianxiang Liu1
1Department of Orthopedics, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430022, People’s Republic of China; 2Department of Orthopedics, Zigong Fourth People’s Hospital, Zigong, 643000, People’s Republic of China
Correspondence: Jianxiang Liu, Department of Orthopedics, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, No. 1277, Jiefang Avenue, Wuhan, Hubei, 430022, People’s Republic of China, Email [email protected]
Abstract: Osteosarcoma (OS) is the most common primary malignant bone tumor, with two incidence peaks: one in adolescence and the other in older individuals. Despite significant research, patient prognosis has not substantially improved in recent decades because of a limited understanding of its pathogenesis and a lack of innovation in treatment approaches. The tumor microenvironment (TME) is a rapidly evolving area of cancer therapy, offering critical insights into the dynamics of osteosarcoma development at the cellular and molecular levels. This study also provides valuable guidance for the development of novel therapeutic strategies. To date, a broad array of nanomedicines have been engineered to target specific ligands within the OS TME. Compared with conventional chemotherapeutic agents, these nanomedicines can substantially enhance drug delivery efficiency while minimizing off-target side effects. In this review, we focus on nanomedicines that target the TME of osteosarcoma. We first explore the core components of the OS TME, which include osteoblasts, mesenchymal stem cells, the vascular microenvironment, and immune cells. Subsequently, we delve into the latest advances and biomedical applications of nanodrug delivery systems engineered specifically for targeting the TME of osteosarcoma. The findings of this review aim to contribute to improved treatment options and outcomes for osteosarcoma patients.
Keywords: osteosarcoma, tumor microenvironment, targeting, nanomedicine, nanoparticles
Introduction
Osteosarcoma is the most common malignant primary bone tumor, but it is generally rare. The global annual incidence is approximately 1 to 3 cases per million people.1 The incidence of osteosarcoma exhibited a bimodal distribution across all age groups. The first peak age of onset occurs in children and adolescents, which may be associated with the rapid bone growth and remodeling during this period.2–4 The second peak arises in the elderly population aged over 60 years, among whom osteosarcoma is mostly secondary to benign bone lesions.5,6 Characterized by early metastasis and tumor heterogeneity, approximately 15–20% of patients present with lung metastasis at the time of diagnosis, which is correlated with poor prognosis.7 The somatic genome of OS is highly complex and exhibits extensive intratumoral heterogeneity. It is characterized by chromosomal aneuploidy, gene mutations, copy number alterations, structural variants, and genomic instability.4 These factors contribute to the complex tumor microenvironment of OS. For instance, the oncogene MDM2 can promote the polarization of macrophages toward the M2 phenotype, whereas mutant p53 can form a complex with heat shock protein 90, thereby inhibiting MDM2 and participating in the regulation of the TME.8
The current standard treatment regimen consists of surgical resection combined with adjuvant chemotherapy. First-line chemotherapy agents include doxorubicin (DOX), cisplatin (CDDP), and methotrexate (MTX).9,10 This regimen has substantially improved the prognosis of osteosarcoma patients, increasing the 5-year survival rate of those with localized OS to 60%–70%. However, it exhibits limited efficacy in patients with metastatic or recurrent disease, with their 5-year survival rate remaining at merely approximately 20%.11,12 Moreover, chemotherapeutic agents are plagued by drawbacks such as poor targeting ability, severe systemic toxic effects, and the development of drug resistance, thus highlighting an urgent need for the development of novel therapeutic strategies.4
Recently, nano-targeted strategies tailored to the tumor microenvironment have attracted extensive attention from researchers. The TME of osteosarcoma is a highly specialized and sophisticated niche, composed of osteoblasts, stromal cells, immune cells, endothelial cells, and noncellular components such as the extracellular matrix (ECM) and inorganic minerals. OS cells interact with the TME through cytokines, chemokines, growth factors, and extracellular vesicles, all of which collectively contribute to tumor initiation, proliferation, apoptosis, invasion, and metastasis.13 Compared with conventional chemotherapeutic agents, nano-targeted drugs exhibit the following advantages: (1) enhanced specificity and selectivity, which elevate the local drug concentration within tumor tissues; (2) improved protection of drugs against degradation, coupled with high drug-loading efficiency; (3) increased solubility of hydrophobic drugs; (4) capacity for co-delivery of multiple therapeutic agents, thereby serving as a promising platform for combinatorial therapy.14,15 In this study, we first summarize the major components of the osteosarcoma microenvironment, including bone cells, stem cells, the vascular microenvironment, and immune cells. We then discuss in depth the nano-targeted strategies developed for the osteosarcoma microenvironment, aiming to provide valuable insights for technological advances in this field. The structure and coverage of this review are illustrated in Figure 1.
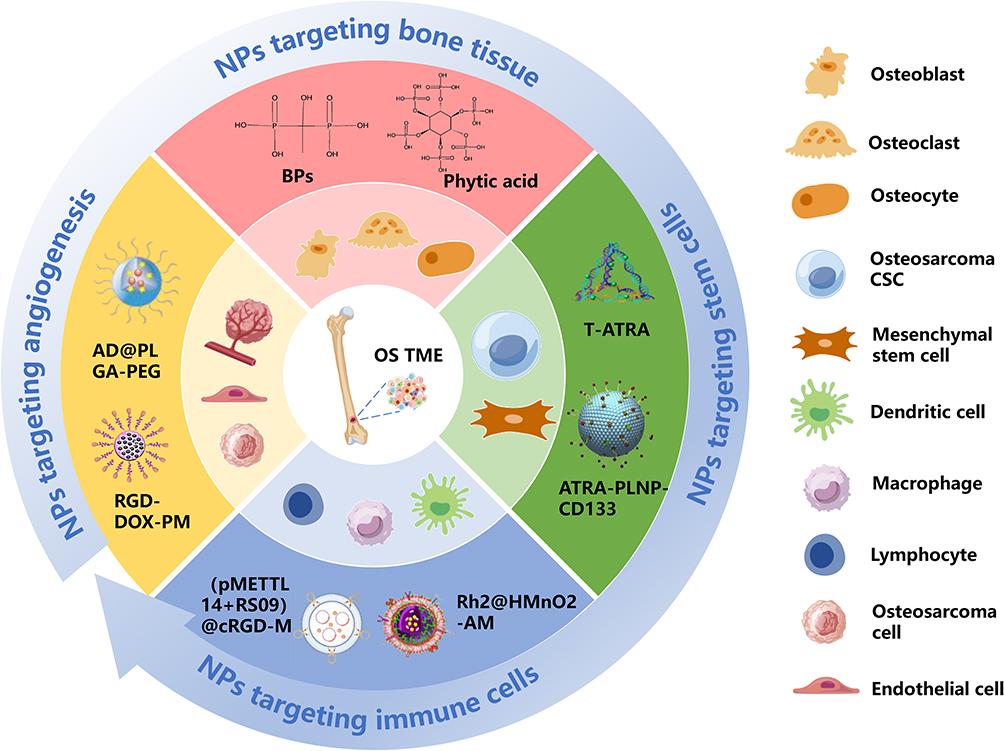 |
Figure 1 Nanoparticles targeting the TME of osteosarcoma. |
Tumor Microenvironment in Osteosarcoma
The osteosarcoma tumor microenvironment (TME) consists of both cellular and noncellular components. The cellular components include bone cells (osteoblasts, osteoclasts, and osteocytes), stromal cells (mesenchymal stem cells and fibroblasts), vascular cells (endothelial cells and pericytes), and immune cells (macrophages, dendritic cells, lymphocytes, natural killer cells, and neutrophils). Noncellular components primarily include the extracellular matrix (ECM) and inorganic minerals.16 Under normal physiological conditions, these components work together to maintain bone homeostasis through sophisticated intercellular communication and paracrine signaling. However, the presence of osteosarcoma cells disrupts this balance, inducing bone remodeling into a “vicious cycle” that creates an immunosuppressive and hypoxic environment; promotes neovascularization; and facilitates the growth, invasion, and metastasis of osteosarcoma.17,18 Herein, we outline the specific details of each component in the osteosarcoma microenvironment during disease progression and their crosstalk. Based on the biological characteristics of key components in the OS TME, such as the bone-targeting property of osteoclasts and the polarization plasticity of immune cells, we systematically summarize the corresponding nanoparticle design strategies.
Bone Cells
The bone cells that constitute the osteosarcoma microenvironment include osteoclasts, osteoblasts, and osteocytes. Under normal physiological conditions, osteoclasts resorb old bone, whereas osteoblasts produce new bone. These cells interact in a highly coordinated manner to maintain bone remodeling homeostasis.19 Most bone destruction in OS is osteolytic, which is closely associated with the activation of osteoclasts.1 Osteoclasts (OCs) are the only cells in bone tissue capable of resorbing bone, and their primary source after birth is hematopoietic stem cells (HSCs).20 Key regulatory factors, such as macrophage colony-stimulating factor (M-CSF) and receptor activator of nuclear factor-kappa B ligand (RANKL), control osteoclast differentiation. M-CSF promotes the survival and proliferation of osteoclast precursors by activating its cohomologous receptor, c-Fms. As a member of the tumor necrosis factor (TNF) superfamily, RANKL promotes the differentiation of osteoclast precursors into osteoclasts by activating its homologous receptor, RANK.21
Osteosarcoma cells form a “vicious cycle” with osteoclasts by producing cytokines such as parathyroid hormone-related protein (PTHrP), transforming growth factor-beta (TGFβ), and interleukin 11 (IL-11). These cytokines stimulate osteoclast activation by inducing RANKL expression on osteoblasts, which leads to osteolysis and the release of growth factors (such as BMP, TGFβ, IGF1, or FGF) from the bone matrix, further promoting osteosarcoma cell proliferation.13,22 Ohba et al reported an increase in the number of osteoclasts (OCs) in the bones of mice inoculated with OS cells in situ compared with control mice, suggesting that OS can stimulate osteoclast differentiation. Endo-Munoz et al reported that the expression of TRACP5 (a biomarker on the surface of OCs) was significantly lower in OS patients than in healthy controls, with this phenomenon being more pronounced in patients with metastatic lesions. These findings indicate that the loss of osteoclasts may enhance OS metastasis.23 Zoledronic acid, an osteoclast inhibitor, has been shown to induce apoptosis, inhibit cell proliferation, and reduce the metastatic potential of OS.24 Osteoclasts play a role in the onset of OS and could serve as potential therapeutic targets. However, understanding the seemingly contradictory roles of osteoclasts in osteosarcoma is complex. One perspective is that in the early stages of disease, osteoclasts promote OS growth by inducing osteolysis and releasing growth factors from the bone matrix. These growth factors, in turn, are thought to prevent OS metastasis. As the disease progresses, genetic and/or epigenetic alterations in OS cells enable them to inhibit osteoclast production, and a reduction in osteoclast numbers may facilitate metastasis and tumor spread.25 The specific mechanism is as follows: In the early stage of the disease, RANKL secreted by osteoblasts and osteocytes in the bone matrix can bind to the RANK receptor on the surface of OC precursor cells. This binding activates the RANKL-RANK signaling pathway, thereby inducing the differentiation and maturation of OCs. Mature OCs can degrade the bone matrix by secreting enzymes such as cathepsin K (CTSK) and matrix metalloproteinase 9 (MMP9), which in turn releases growth factors including TGFβ. These factors promote the proliferation of OS cells through signaling pathways such as the TGF-β-Smad pathway, while providing a “trophic microenvironment” for OS cells to suppress their metastasis.25 Following genetic/epigenetic alterations, OS cells gain the ability to secrete factors such as VEGF, urokinase-type plasminogen activator (uPA), and matrix metalloproteinase 1 (MMP1). These secreted factors inhibit the differentiation of OC precursor cells, leading to a reduction in OC numbers.23
Osteoblasts primarily originate from bone marrow mesenchymal stem cells (MSCs). Osteosarcoma can disrupt normal bone remodeling, leading to the formation of Codman’s triangle or Sunburst periosteal reactions, which are closely associated with osteoblast activity. The differentiation of MSCs into osteoblasts involves several key steps. The transcription factors RUNX2 and transforming growth factor-beta (TGF-β1) play crucial roles in the early stages of osteoblast differentiation by promoting the differentiation of MSCs into preosteoblasts. Osterix (also known as SP7) is predominantly involved in osteoblast differentiation downstream of RUNX2 and drives the differentiation of preosteoblasts into mature osteoblasts.26,27 Recent studies have revealed that osteoblast-derived extracellular vesicles (OB-EVs) may participate in cross-communication between osteosarcoma cells and the TME, reducing the invasiveness and viability of osteosarcoma cells.28 Additionally, MET overexpression has been shown to induce the transformation of primary human osteoblasts into osteosarcoma cells, suggesting that osteoblasts could serve as progenitor cells for osteosarcoma.29
Osteocytes are mature, mineralized cells derived from osteoblasts and are the most abundant cell type in human bones, with an estimated 42 billion present.30 Osteocytes serve as the “coordinators” of bone tissue, maintaining the physiological responses of bone to mechanical and metabolic demands. They play crucial roles in regulating, initiating, and terminating bone remodeling.19 Cytokines such as CXCL1 and CXCL2, which are produced by osteocytes, have been implicated in driving the metastasis of breast cancer cells.31 Additionally, prostate cancer cells can induce a positive feedback loop, promoting osteocytes to produce GDF15 and enhancing the expression of EGR1 in tumor cells, thereby facilitating metastasis.32 Sottnik et al reported increased expression of osteocyte markers (DMP1, MEPE, and PHEX) in the osteoblast subtypes of osteosarcoma, suggesting that osteocytes may be the origin of osteosarcoma cells.33 However, the mechanisms by which osteoblasts and osteocytes contribute to the initiation and progression of osteosarcoma remain unclear. Further research is necessary to explore their roles in osteosarcoma development before these cells can be targeted for therapeutic purposes.
Mesenchymal Stem Cells (MSCs)
Mesenchymal stem cells (MSCs) are important components of the osteosarcoma tumor microenvironment. MSCs contribute to osteosarcoma growth, recurrence, metastasis, and drug resistance through complex crosstalk mechanisms. MSCs may also serve as the origin of osteosarcoma cells. For example, silencing the retinoblastoma (Rb) tumor suppressor gene and overexpressing c-MYC in MSCs has been linked to osteosarcoma tumorigenesis.34 Osteosarcoma cells secrete cytokines such as TGF-β, stromal-derived factor-1 (SDF-1), and PDGF, which attract MSCs to the tumor site.35 MSCs secrete various factors, such as IL-6, VEGF, CCL5, and CXCL10, which promote osteosarcoma growth, metastasis, and angiogenesis.36,37 The TGF-β secreted by MSCs is also thought to be involved in immune escape.38 MSCs are also involved in the resistance of osteosarcoma to chemotherapy. IL-6 secreted by MSCs can activate the STAT3 signaling pathway, thereby promoting the expression of MDR-1 (multidrug resistance gene 1) and MRP (multidrug resistance-associated protein), which further increases the survival rate of OS cells treated with DOX or cisplatin.39
MSCs can also serve as both receptors for and donors of extracellular vesicles (EVs) within the TME. These EVs mediate intercellular communication by transporting a diverse array of bioactive molecules, including microRNAs (eg, miR-21, miR-34a), proteins (eg, PDGFR-β, TIMP-1, TIMP-2) and metabolites such as lactate and glutamate.40 Research has shown that EVs derived from OS cells can induce the secretion of matrix metalloproteinase 1 (MMP1), VEGF, and intercellular adhesion molecule 1 (ICAM-1) from MSCs, promoting tumor angiogenesis and metastasis.41 MSC-derived exosomes can also promote osteosarcoma progression by inducing oncogenic autophagy.42 Extracellular vesicles derived from MSCs can carry microRNA-208a, which enhances the viability of osteosarcoma cells and promotes metastasis by regulating PDCD4.43 In contrast, miR-206 significantly inhibits the proliferation, migration, and invasion of osteosarcoma cells while promoting their apoptosis by targeting TRA2B.44 Lee et al reported that low concentrations of AD-MSCs inhibited osteosarcoma growth, whereas high concentrations of AD-MSCs promoted osteosarcoma growth.45 Exosomes derived from mesenchymal stem cells (MSCs) can also serve as nanoparticle carriers for the targeted delivery of therapeutic agents. The role of MSCs in osteosarcoma growth is complex and contradictory and potentially influenced by various factors, including the source of the MSCs, the tumor type, and the tumor microenvironment.37
Vascular Microenvironment
Like other invasive tumors, osteosarcoma creates a dense network of microvessels to ensure a sufficient supply of nutrients and oxygen, a process that is highly complex and regulated by various cytokines, signaling pathways, and intercellular interactions. Endothelial progenitor cells (EPCs) play a significant role in tumor angiogenesis and work synergistically with mesenchymal stem cells and hematopoietic stem cells to drive tumor progression.46 A rat model study confirmed that compared with the injection of tumor cells alone, the coinjection of EPCs with osteosarcoma cells significantly increased the number of microvessels.47 There are two theories regarding the source of EPCs in the tumor microenvironment: one posits that EPCs reside within the tumor, whereas the other suggests that they are recruited from the bone marrow, with the latter being the dominant source.46,48 Owing to the intense metabolic activity of osteosarcoma cells, a hypoxic and acidic environment often forms within the tumor. Hypoxia primarily works through hypoxia-inducible factor 1-alpha (HIF-1α), which is a transcriptional regulator that promotes the expression of various proangiogenic genes, including VEGF, PDGF, and FGF.49,50 VEGF, a key angiogenic factor, binds to its receptor, VEGFR, triggering the production of matrix metalloproteinases (MMPs) and urokinase plasminogen activators, which degrade the extracellular matrix to promote metastasis and induce endothelial cell migration and neovascularization.51–53 High expression of VEGF and VEGFR in OS is positively correlated with tumor stage and metastasis.54 A systematic review by Chen et al reached the same conclusion, suggesting that VEGF expression could serve as a prognostic biomarker for OS patients.55 Nanoparticle-based targeted delivery systems have been developed to target VEGFR and its ligands, as well as to inhibit HIF-1α for anti-angiogenic therapy.
Exosomes secreted by osteosarcoma cells also carry key molecules such as EWSAT1, which increases the sensitivity and reactivity of vascular endothelial cells, further promoting OS angiogenesis.56 Another study revealed that the lncRNA OIP5-AS1, which is carried by exosomes from osteosarcoma cells, regulates angiogenesis, autophagy, and apoptosis through the miR-153-ATG5 axis.57 Integrins are a class of critical cell surface receptors that promote angiogenesis at tumor sites by mediating cell–extracellular matrix interactions and facilitating intracellular signal transduction.58 Several antiangiogenic agents, including VEGF monoclonal antibodies (such as bevacizumab), tyrosine kinase inhibitors (apatinib, sorafenib, pazopanib and regorafenib), and human recombinant endostatin (Endostar), have been developed to target angiogenesis in tumors.59 However, these therapies are still in the early stages of development and require further clinical validation.
Immune Cells
Osteosarcoma tissues are infiltrated by many immune cells, which interact to create an immunosuppressive microenvironment that supports tumor growth, migration, apoptosis, and drug resistance. Recent technological advances in immunohistochemistry, transcriptomics, and genomics have facilitated the study of tumor heterogeneity and its various subpopulations. The immune microenvironment of osteosarcoma is predominantly populated by myeloid cells, such as monocytes, tumor-associated macrophages (TAMs), dendritic cells (DCs), and neutrophils, whereas lymphocytes, such as CD8+ T cells, CD4+ T cells, regulatory T cells (Tregs), and natural killer (NK) cells, are relatively underrepresented.17,60 Zhou et al conducted scRNA-seq analysis on 11 osteosarcoma patients and reported that in the myeloid lineage, monocytes and macrophages accounted for 70–80% of the cells, dendritic cells (DCs) represented 5%, and neutrophils were present in small numbers.61 The tumor immune microenvironment holds great potential for guiding new drug target development, making it crucial to explore this area further.
Tumor-Associated Macrophages (TAMs)
TAMs are among the most abundant immune cells within the osteosarcoma TME, potentially constituting >50% of the total immune cell population.62 TAMs can be recruited from the periphery or generated through in situ proliferation of tissue-resident macrophages.63 TAMs exhibit heterogeneity and plasticity and can exhibit both antitumor and protumor characteristics, depending on the microenvironment and the stimuli to which they are exposed. TAMs are classified into two subtypes: M1-type and M2-type. M1-type TAMs have antitumor properties; they promote the secretion of inflammatory factors that trigger antitumor immunity, produce reactive oxygen species (ROS) to induce tumor tissue damage, and can directly phagocytose tumor cells.64,65 M2-type TAMs, as important tumor-associated anti-inflammatory macrophages, increase the expression of anti-inflammatory factors (such as IL-10 and TGF-β), VEGF, and extracellular matrix remodeling cytokines, thereby promoting immune evasion, angiogenesis, and metastasis.65 A small subset of M2-TAMs exhibit increased expression levels of M1-TAM marker genes, suggesting that M1-TAMs and M2-TAMs can interconvert within the OS microenvironment.61 Induction of M1 polarization has provided valuable insights for the development of novel nanoparticle-based therapeutics.
A study by Dumars et al revealed that vascular density was significantly increased in patients with metastatic OS, and there was a correlation between M2-TAMs and CD146+ vascular cells, suggesting that M2-TAMs are involved in promoting neovascularization.66 TAMs secrete various proangiogenic factors, including vascular endothelial growth factor (VEGF), matrix metalloproteinase 9 (MMP-9), and fibroblast growth factor (FGF).67 It has been reported that OS cells secrete IL-34, which stimulates endothelial cell proliferation and recruits M2 macrophages to tumors, thereby promoting OS growth and metastasis.68 TAMs are also associated with immunosuppression through several mechanisms. First, TAMs express programmed death ligand-1 (PD-L1) and cytotoxic T lymphocyte-associated antigen-4 (CTLA-4), which inhibit T lymphocyte activity.69 Additionally, CD163+ M2-TAMs secrete immunosuppressive cytokines (IL-10 and TGF-β), which inhibit T-cell activation and contribute to T-cell exhaustion.70 Furthermore, TAMs stimulate the differentiation of regulatory T cells (Tregs) and their recruitment into tumor tissues, suppressing T lymphocyte-mediated antitumor immunity.71 TAMs promote OS metastasis primarily by secreting cytokines and chemokines. For example, the chemokine CCL18 derived from M2-TAMs significantly promotes OS proliferation and metastasis through the EP300/UCA1/Wnt/β-catenin pathway.72 TAMs can also upregulate COX-2, activate the COX-2-STAT3 axis, and promote OS metastasis by facilitating epithelial‒mesenchymal transition (EMT).73
Existing evidence shows that M2-TAMs are highly infiltrated in primary osteosarcoma tissues. All-trans retinoic acid (ATRA) has been shown to reduce the colony-forming ability of mouse osteosarcoma cells and inhibit tumor formation by suppressing M2 polarization of TAMs.74 Another study revealed that localized osteosarcoma tissues are rich in M1-TAMs, whereas metastatic osteosarcoma tissues have a greater presence of M2-TAMs. This finding suggests that the ratio of M1 to M2 TAMs may be linked to osteosarcoma metastasis, with M1 polarization inhibiting OS metastasis.66 Emilie et al reported that the TAM density in primary localized osteosarcoma is approximately twice that in metastatic osteosarcoma and that the antitumor effect of macrophages outweighs their tumor-promoting effect, resulting in overall inhibition of osteosarcoma metastasis.75 Interestingly, the prognosis of osteosarcoma patients appears to be less related to the proportions of M1 and M2 TAM subtypes and more related to the total number of TAMs. Patients with higher TAM counts tend to have a lower likelihood of metastasis, which may be linked to the dual role of TAMs—both the M1 and M2 subtypes may inhibit OS growth under certain conditions.76,77
Lymphocytes
Lymphocytes, which include T cells, B cells, and natural killer (NK) cells, play critical roles in the immune response against osteosarcoma. These cells originate from hematopoietic stem cells in the bone marrow, but their maturation and specific functions vary. T lymphocytes can be classified on the basis of their functions into cytotoxic T lymphocytes (CTLs, CD8+), which directly kill tumor or infected cells; helper T lymphocytes (THLs, CD4+), which assist other immune cells and enhance immune responses; and regulatory T cells (Tregs), which suppress excessive immune responses. B lymphocytes mature in the bone marrow, and their main function is to produce antibodies. NK cells mature in the bone marrow and peripheral lymphoid tissues (such as the spleen and lymph nodes) and are characterized by their ability to kill abnormal cells directly in the body without the need for prior sensitization.
On the basis of their receptor characteristics, T lymphocytes can be further divided into αβT and γδT lymphocytes. αβT lymphocyte receptors activate T cells by recognizing antigenic peptide–MHC complexes. Cytotoxic T lymphocytes can recognize tumor antigens presented by MHC class I molecules on the surface of tumor cells, which activate downstream immune receptor tyrosine activation motifs (ITAMs), initiating immune killing.78 Evidence suggests that OS may evade cytotoxic T lymphocyte attacks by inhibiting the expression of MHC class I molecules.79 After recognizing the antigenic peptide-MHC complex, CD4+ T lymphocytes can promote the release of cytokines such as IL-2 and IFN-γ from antigen-presenting cells, thereby exerting antitumor effects.80 PD-1 is a major inhibitory receptor expressed on T cells, and its binding to PD-L1 can induce T-cell exhaustion and suppress immune responses.81 Studies have shown that OS cells can evade immune surveillance by secreting PD-L1.82 Casanova et al reported that infiltration of CD4+ T cells improved the prognosis of OS patients and that patients with greater infiltration of CD8+ T cells had significantly longer overall survival (OS) and progression-free survival (PFS).83 The activation of Toll-like receptor 4 (TLR4) can suppress OS progression. Richert et al reported that activating TLR4 in a mouse model induced osteosarcoma regression by stimulating CD8+ T cells and promoting the polarization of TAMs toward the M1 phenotype.84 Furthermore, macrophages may suppress T lymphocyte function through complex interactions. For example, Liu et al reported that C5_IFIT1+ macrophages interact with Treg cells via ligand‒receptor interactions, leading to CD8+ T-cell exhaustion.85
NK cells are effector cells of the innate immune system and are highly sensitive to the downregulation of MHC expression on cell surfaces, making them particularly effective in eliminating tumor cells that reduce MHC expression to evade immune surveillance.86 The primary mechanisms by which NK cells kill target cells include the release of cytotoxic granules containing perforin, granzymes, and granulysin to induce apoptosis; the production of death ligands such as Fas ligand (FasL) and TNF-related apoptosis-inducing ligand (TRAIL); and the secretion of cytokines such as TNF-α and IFN-γ to stimulate adaptive immunity.87 Several studies have highlighted the potential of NK cells in OS treatment. For example, Rademacher et al reported that IL-12 overexpression in sarcomas promotes NK cell-mediated immune modulation, triggering a specific immune response and significantly controlling tumor growth.88 Furthermore, Yamato et al demonstrated that blocking FSTL1 significantly restores NK cell activity, inhibits OS cell growth in vivo, and notably extends overall survival in mice.89
Dendritic Cells (DCs)
Dendritic cells (DCs) are highly efficient antigen-presenting cells derived from the bone marrow that play a critical role in initiating and coordinating T-cell-mediated antitumor immune responses.90 On the basis of their phenotype and function, DCs are classified into four distinct subtypes: classical DCs (cDCs), including cDC1s and cDC2s; plasmacytoid DCs (pDCs); and monocyte-derived DCs. cDC1s cross-present exogenous antigens and activate CD8+ T cells via MHC class I molecules, thereby triggering cellular immune responses against tumor cells. cDC2s primarily activate CD4+ T cells and, through the secretion of cytokines and costimulatory molecules such as CD40L, help induce the differentiation of Th1, Th2, or Th17 cells. Monocyte-derived DCs are involved mainly in inflammatory responses and can promote immune reactions by activating both CD4+ and CD8+ T cells.91 Plasmacytoid DCs produce type I interferons (IFN-Is) and can differentiate into antigen-presenting cells when stimulated by viruses or self-DNA.92
Evidence suggests that the number of cDC2s is significantly greater in lung metastases than in primary and recurrent lesions.62 DCs express glutamate receptor 4 (GMR-4), which regulates the expression of IL-23 and related cytokines, such as IL-12, to drive osteosarcoma development. Kansara et al reported that treatments with GMR-4 agonists and IL-23 antagonists were as effective as those with DOX, indicating that these pathways could serve as new therapeutic targets for osteosarcoma.93 The chemokine receptor CCR7 guides DCs from peripheral tissues to lymph nodes and regulates T-cell activation. Additionally, CCR7 modulates DC survival, chemotaxis, and migration speed through the activation of three signaling pathways (PI3K/Akt, MAPK, and RhoA), which are closely linked to tumor metastasis.94 Single-cell RNA sequencing (scRNA-seq) of osteosarcoma cells revealed that mature immune-regulatory DCs (mregDCs) can induce tumor immune tolerance by recruiting Tregs.82
Nanoparticle Applications in Osteosarcoma
The specific tumor microenvironment (TME) of osteosarcoma significantly hinders the efficient delivery of traditional chemotherapy drugs. Issues such as multidrug resistance and off-target drug effects further complicate treatment, making conventional therapies less effective. In recent years, nanotechnology-based targeted therapies have attracted considerable attention, as they provide new opportunities for overcoming these challenges. NPs designed to mimic biological agents exhibit high biocompatibility and low immunogenicity, while ligand-modified nanocarriers can specifically bind to biomarkers on osteosarcoma cells, thereby reducing off-target effects and enhancing treatment efficacy. In this section, we focus on the different nanoparticle-based drug delivery systems developed to target the components of the osteosarcoma microenvironment.
Targeted Bone Tissue
Bones consist of both organic and inorganic components, which together form the body’s framework, maintaining stability and ensuring proper posture and movement. Inorganic components account for approximately 60%-70% of bone mass, primarily in the form of hydroxyapatite (Ca10(PO4)6(OH)2; HAp), whereas organic components make up approximately 30%-40%, with type I collagen being the predominant protein and the remainder consisting of small amounts of water.95 The surface of hydroxyapatite is rich in hydroxyl and phosphate groups, which can strongly bind to drugs through covalent bonding or electrostatic interactions, facilitating high accumulation in bone tissue and making it an ideal target for drug delivery. To harness these properties, nanoparticles based on bisphosphonates, oligopeptides, tetracyclines, and phytic acid have been developed and have shown significant efficacy in preclinical studies.
Bisphosphonates
Bisphosphonates (BPs) were first reported over half a century ago in rat models for the treatment of osteoporosis.96 In recent years, further research has revealed their effectiveness in treating hypercalcemia, multiple myeloma, Paget’s disease of bone in adults, and both primary and secondary bone tumors.97 BPs are highly selective for bone, and their properties are closely related to their molecular structure. The P-C-P backbone of BPs contains two terminal phosphate groups, which chelate calcium ions on the surface of hydroxyapatite in a bidentate structure. Additionally, the R1 and R2 side chains influence their binding affinity.98 BPs are classified into three generations on the basis of their chemical structure. First-generation BPs, which lack nitrogen atoms, are metabolized into nonhydrolyzable ATP analogs, affecting osteoclast energy metabolism. Examples include etidronate and clodronate. Second-generation BPs contain a nitrogen group, enabling them to target farnesyl pyrophosphate synthase (FPPS) and inhibit the mevalonate pathway, leading to cell death. Members of this generation include alendronate (ALN), pamidronate, neridronate, and ibandronate. Third-generation BPs, which contain nitrogen heterocycles, function similarly to second-generation BPs, with zoledronic acid (ZOL) and risedronate as key examples.99
Recently, several BP-modified nanoparticle drug delivery systems have been developed to target the bone microenvironment. Mitchell et al designed BP-modified lipid nanoparticles (LNPs) for the bone-targeted delivery of mRNA-based therapeutics (Figure 2A).100 Segal et al conjugated ALN with the antiangiogenic agent TNP-470 and N-(2-hydroxypropyl) methacrylamide (HPMA) copolymers. In a mouse osteosarcoma xenograft model, the HPMA copolymer-ALN-TNP-470 conjugate demonstrated greater antitumor activity and lower organ toxicity than did the free combination of ALN and TNP-470, which specifically inhibited tumor-induced neovascularization.101 Nguyen et al designed a targeted nanoparticle composed of ALN and DOX. In vitro experiments revealed that the nanoparticles significantly accumulated in hydroxyapatite crystals and exhibited greater cytotoxicity than free DOX did (Figure 2B–D).102
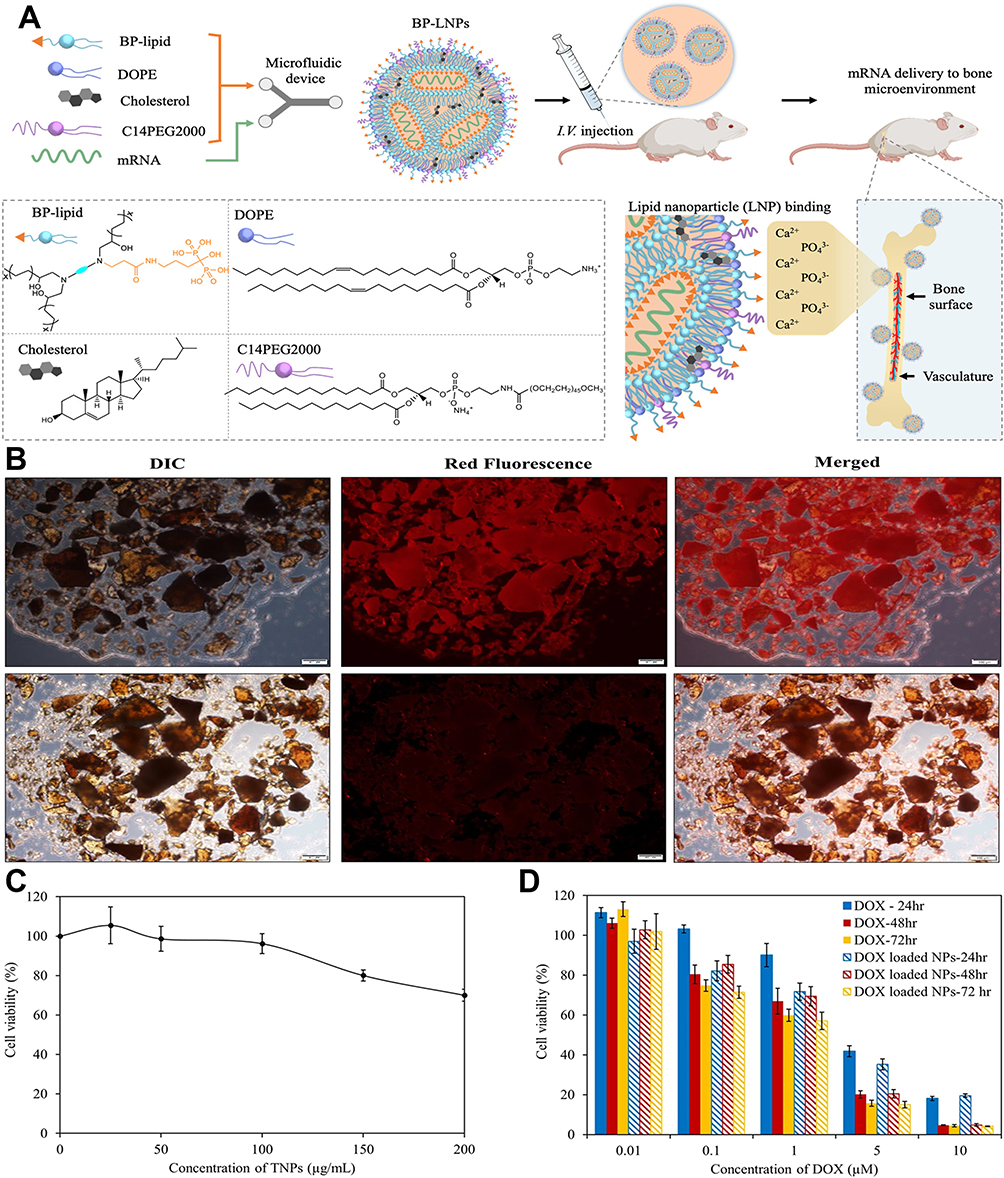 |
Figure 2 BP-modified nanoparticles are used for targeting bone tissue. (A) Schematic diagram of the preparation process of BP-LNPs. Following intravenous administration, BP-LNPs can chelate Ca2+ in bones, allowing them to target bone tissue.100 Copyright 2022, Journal of the American Chemical Society. (B) Representative fluorescence images of HAp crystals after incubation with RhB-labeled TNPs (upper panel) showing the interaction between HAp and targeted NPs. The lower panel represents nontargeted PEGylated NPs. (C and D) Comparison of the cytotoxicity between nanoparticles loaded with DOX and free DOX. Within a shorter incubation period, nanomedicines demonstrated greater toxic effects than did free DOX. Scale bar represents 100 μm for each panel.102 Copyright 2016, Scientific Reports. |
Oligopeptides
Osteopontin and bone sialoprotein are noncollagen proteins in bone that exhibit strong affinity for hydroxyapatite owing to the repetitive L-Asp and L-Glu sequences in their structures.103 The mechanism behind this affinity involves ionic interactions between the carboxyl groups of peptides, which carry negative charges, and the calcium ions in hydroxyapatite (HAP). The strength of this interaction correlates with the number of exposed amino acid residues.104 It has been reported that the optimal HA binding affinity occurs with Asp repeat sequences of 6 or 8 residues.105 After modifying nanoparticles (NPs) with Asp-rich peptides, bone accumulation can be increased by more than 10-fold.106 Inspired by this, researchers are designing Asp-rich oligopeptides as new ligands for bone targeting. Wang et al developed an aspartic acid octapeptide-modified dendritic platinum-copper alloy nanoparticle (Asp-DPCN), which demonstrated significantly greater affinity for hydroxyapatite and ex vivo bone fragments than nontargeted Gly-DPCN did, along with improved accumulation efficiency in bone tumors.107 Additionally, a hydrophilic D-aspartic acid octapeptide-modified nanomicelle was designed for conjugating DOX for targeted osteosarcoma therapy, achieving a binding rate of up to 91% with Hap.108
Other Bone-Targeting Materials
Tetracycline molecules also demonstrate a strong affinity for hydroxyapatite (HAP), making them promising bone-targeting ligands. Compared with bisphosphonates (BPs), tetracyclines may offer superior oral bioavailability and fewer side effects. Wang et al developed bone-targeting nanoparticles (TC-PLGA NPs) using poly(lactic-co-glycolic acid) (PLGA) and tetracycline (TC). These nanoparticles exhibited significantly greater bone-targeting efficiency than did PLGA NPs without TC.109 Phytic acid (PA), with its molecular structure containing phosphate groups, also has a high affinity for HAP. Additionally, PA has inherent anticancer properties and excellent biocompatibility. Zhou et al prepared PA-coated platinum nanoparticles (PA/PtNPs), and the results indicated that PA significantly enhanced the accumulation of PA/PtNPs in tumor-associated bone lesions, effectively inhibiting bone tumor growth and osteolysis.110
Targeted Immune Cells
In recent years, researchers have increasingly focused on the tumor microenvironment and its immune components to develop new immunotherapies aimed at improving the prognosis of OS patients. Nanotechnology has demonstrated significant potential in this emerging field, providing novel approaches for modulating the immune microenvironment of OS. For example, nanoparticles can promote the polarization of macrophages from the M2 phenotype to the M1 phenotype, thus initiating an immune response. When combined with immune checkpoint inhibitors (such as anti-PD-1 antibodies), these nanoparticles enhance immune activation at the tumor site while significantly minimizing off-target effects. Certain nanomaterials can also improve the presentation of tumor antigens to DCs and T lymphocytes, thereby increasing antitumor immunity. Moreover, owing to their unique optical and physical properties, these nanomaterials can be integrated with therapies such as photothermal therapy (PTT), photodynamic therapy (PDT), and radiation therapy to achieve more effective tumor-killing effects. Herein, we will introduce a variety of nanoparticles targeting immune cells within the OS immune microenvironment (Table 1).
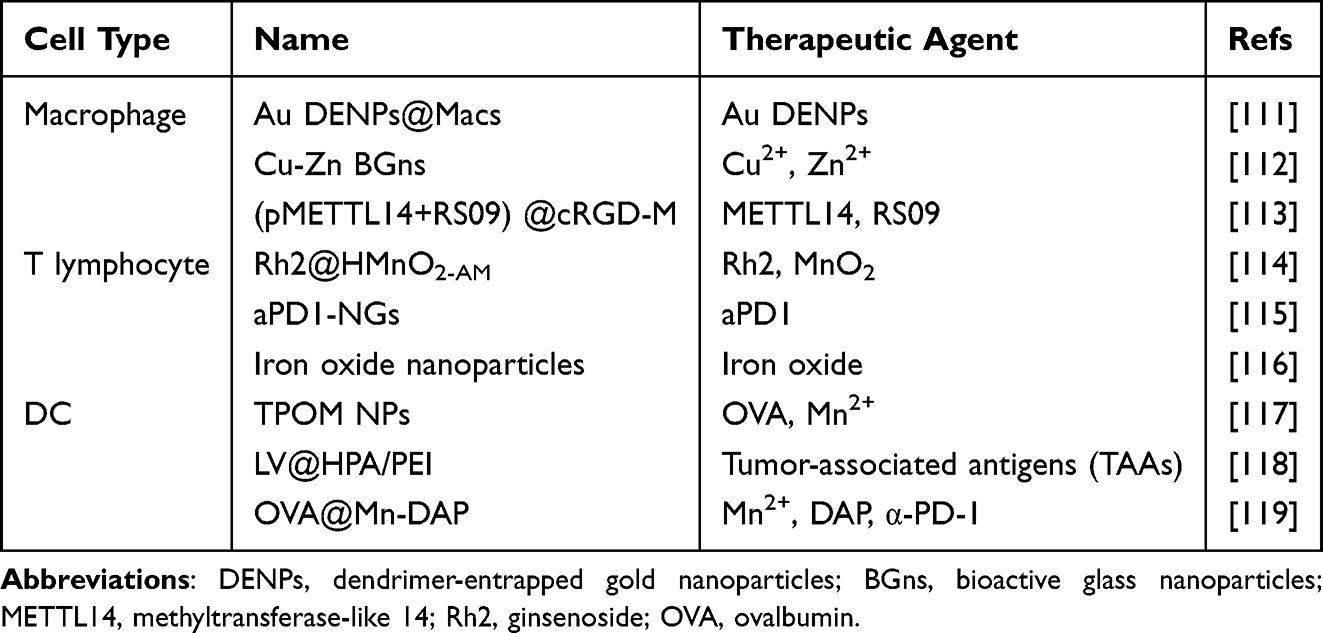 |
Table 1 Summary of Nanoparticles Targeting Immune Cells |
Regulating Macrophages
Macrophages are a key component of the OS tumor microenvironment, and their polarization status plays a crucial role in reshaping the immune microenvironment and regulating local immune responses. Several nanoparticle-based therapies have been developed to modulate the polarization of TAMs. Mifamurtide, a synthetic bacterial cell wall analog, interacts with Toll-like receptor 4 (TLR4) on the cell surface, triggering the activation of the innate immune system and increasing the antitumor immune response of macrophages.120 Owing to its significant efficacy, mifamurtide was approved for use in Europe in 2009. A recent prospective clinical study demonstrated that combining mifamurtide with conventional chemotherapy improved survival rates in young patients with localized OS.121 High concentrations of Zn2+ and Cu2+ have been shown to promote the secretion of anti-inflammatory cytokines and induce M1 macrophage activation.112,122 Huo et al developed a novel bioactive glass nanoparticle (Cu-Zn BGn) doped with copper and zinc ions, which regulates M1/M2 macrophage polarization by alternately activating the NOD-like receptor pathway and inhibiting the MAPK signaling pathway (Figure 3A).112 Researchers have also engineered macrophages with gold nanoparticles (Au DENPs@Macs) as a therapeutic and diagnostic platform for osteosarcoma (Figure 3B). They synthesized dendrimer-entrapped gold nanoparticles (Au DENPs) and demonstrated their potent ability to drive M1 macrophage polarization. As shown in Figure 3C–E, treatment with Au DENPs markedly up-regulated the canonical M1 markers CD86, iNOS and TNF-α. After being combined with the Au DENPs, engineered macrophages enhanced tumor apoptosis and demonstrated excellent CT imaging performance. The Au DENPs@MACs group exhibited the highest apoptosis rate (Figure 3F). Subsequent Western blot analyses (Figure 3G and H) corroborated this finding, revealing the highest level of activated cleaved Caspase-3 in the Au DENPs@MACs group and underscoring its superior pro-apoptotic effect.111
 |
Figure 3 Nanoparticles regulate the polarization of macrophages. (A) Synthesis of the PADM@CZ hydrogel and its bidirectional immunomodulatory effects on macrophages.112 The upward red arrow indicates that the quantity of ROS is on the rise. The green upward arrow indicates that the number of M2 type macrophages is on the rise. The downward green arrow indicates that the number of MAPK and M1 type macrophages is decreasing. Copyright 2023, Advanced Science. (B) Preparation of the Au DENPs@Macs and their application in osteosarcoma treatment and CT imaging. The red arrows indicate that the quantities of CD86, iNOS and TNF-α are increasing. (C–E) Increased expression of CD86, inducible nitric oxide synthase (iNOS), and TNF-α, which are characteristic of M1 polarization in macrophages, was observed via flow cytometry, immunofluorescence staining, etc. (***P<0.001) (F) The highest apoptosis rate was observed in the Au DENPs@MACs group by flow cytometry. (G and H) Western blot analysis revealed the highest expression of cleaved caspase-3 and the best proapoptotic effect in the Au DENPs@MACs group (*P<0.05, ***P<0.001). Scale bar represents 100 μm for each panel.111 Copyright 2021, Chemical Engineering Journal. |
Huang et al developed cRGD peptide-modified macrophage membrane-coated nanovesicles for targeted OS therapy. These nanovesicles not only enhance targeting efficiency but also facilitate the delivery of METTL14 and TLR4 agonists, denoted as pMETTL14+RS09@cRGD-M. METTL14 (methyltransferase-like 14) downregulates TICAM2 expression and inhibits the TLR4 signaling pathway in macrophages. RS09, a TLR4 agonist, has the potential to induce M1 polarization of TAMs, thereby exerting antitumor effects (Figure 4A). Experimental results demonstrated that (pMETTL14+RS09) @cRGD-M nanovesicles exhibit significant antitumor efficacy both in vitro and in vivo (Figure 4B and C) and promote M1 polarization of macrophages (Figure 4D and E).113
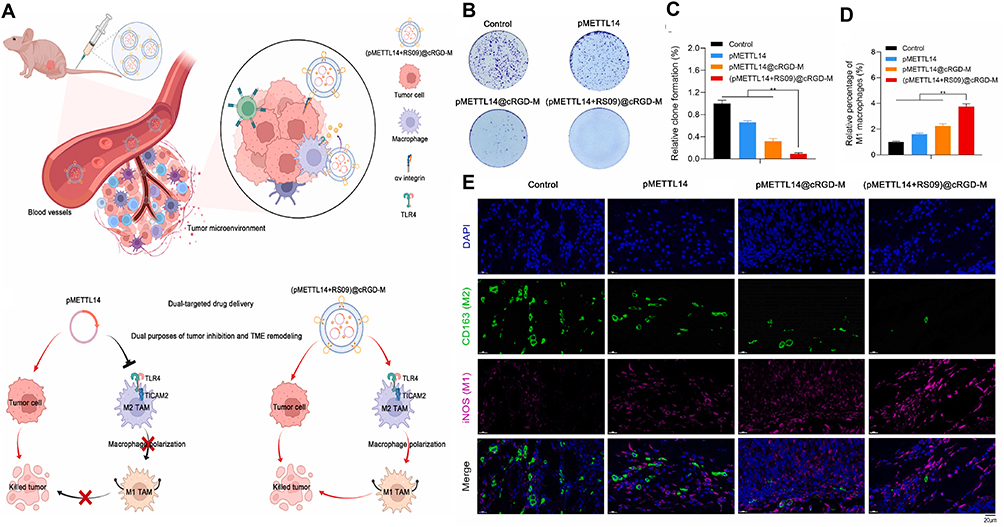 |
Figure 4 (pMETTL14+RS09)@cRGD-M-targeting regimen and therapeutic effect analysis. (A) The dual-targeting mechanism of pMETTL14+RS09@cRGD-M can not only kill tumors but also induce M1 polarization of macrophages. (B and C) Plate cloning experiment study (pMETTL14+RS09)@cRGD-M on the inhibitory effect of tumor cell proliferation (**p < 0.01). (D and E) Immunofluorescence staining was used to observe the number and proportion of M1 (iNOS) and M2 (CD163) macrophages in tumor tissues of the control, pMETTL14, pMETTL14@cRGD-M, and pMETTL14+RS09@cRGD-M groups (**p < 0.01) Scale bar represents 20 μm for each panel.113 Copyright 2022, Bioactive Materials. |
Regulating T Lymphocytes
T lymphocytes are essential components of adaptive immunity and play a crucial role in the antitumor immune response in OS. Nanomaterials have been shown to effectively promote T lymphocyte activation, proliferation, and infiltration into tumor tissues. Fu et al designed hollow manganese dioxide (HMnO2) nanoparticles modified with alendronate (ALD) and coated them with tumor cell membranes to deliver ginsenoside Rh2 for OS therapy, denoted as Rh2@HMnO2-AM (Figure 5A). The combination of ALD and the tumor cell membrane enhances bone-targeting capabilities, whereas ginsenoside Rh2 inhibits tumor cell growth and induces apoptosis.123 The experimental results demonstrated that the Rh2@HMnO2-AM nanoparticles triggered immunogenic cell death, leading to T-cell infiltration and activation (Figure 5B), while also promoting the expression of BAX and BCL-2 to induce tumor apoptosis (Figure 5C). In vivo studies further revealed significant inhibition of tumor growth in the mouse model (Figure 5D).114 Research indicates that calcium ions play pivotal roles in T-cell activation and proliferation. Increased Ca2+ signaling induces the nuclear translocation of T-cell nuclear factor (NFAT), which promotes the expression of proliferation-associated genes, such as IL-2, facilitating T-cell proliferation and differentiation, thus strengthening the immune response.124,125 In this context, calcium phosphate nanocarriers have been developed to modulate intracellular calcium levels, thereby increasing T-cell receptor (TCR) signaling and promoting cytokine secretion and the proliferation of CD8+ cytotoxic T cells.126
 |
Figure 5 Preparation of Rh2@HMnO2-AM and the immune-chemodynamic synergistic osteosarcoma therapy mediated by it. (A) Schematic diagram of the mechanism of the synthesis and treatment of osteosarcoma by Rh2@HMnO2-AM. (B) Representative immunofluorescence staining of CD3+ and CD8+ T cells in tumor tissues. (C) Western blot analysis of BAX, BCL-2 and Caspase 3 in the different groups. (D) Photographs of tumors from the mice in each group after 15 days of drug treatment. Scale bar represents 100 μm for each panel.114 Copyright 2022, Bioactive Materials. |
Chimeric antigen receptor (CAR) T-cell therapy represents a groundbreaking advancement in cancer treatment, as it allows tumor cells to be recognized and targeted without the need for MHC molecules. This approach effectively addresses the challenge posed by tumor cells that downregulate MHC expression to evade immune surveillance.127 While CAR-T-cell therapy has shown great success in treating hematologic malignancies, it faces significant challenges in solid tumors, including physical barriers that hinder CAR-T-cell infiltration, the absence of uniformly expressed tumor-specific antigens, and the immunosuppressive tumor microenvironment.128,129 A key factor contributing to the immunosuppressive tumor microenvironment is the expression of PD-L1 on tumor cells, which binds to PD-L1 on cytotoxic T lymphocytes (CTLs), thereby hijacking the immune checkpoint pathway and suppressing T-cell-mediated antitumor immunity.130 To address this issue, Chen et al developed nanogels (aPD1-NGs) capable of carrying immune checkpoint inhibitors anchored to the surface of CAR-T cells. Their results demonstrated that this strategy significantly inhibited tumor growth.115 One of the challenges in applying CAR-T-cell therapy to OS is the difficulty in monitoring CAR-T-cell accumulation within the tumor. To overcome this, iron oxide nanoparticles have been used to label CAR-T cells, enabling noninvasive detection of labeled CAR-T cells via magnetic resonance imaging (MRI), magnetic particle imaging (MPI), and photoacoustic tomography (PAT).116
Regulating Dendritic Cells (DCs)
Upon activation, DCs are specialized antigen-presenting cells that present tumor antigens to T lymphocytes, triggering cytotoxic T lymphocyte responses that kill tumor cells.131,132 Liu et al developed a titanium carbide-based TPOM nanoplatform designed for immune therapy triggered by photothermal therapy. This nanoplatform is composed of titanium carbide and manganese ions integrated with ovalbumin, facilitating cross-talk between innate and adaptive immune responses. Upon near-infrared laser irradiation, dying tumor cells release mt-DNA and tumor-derived antigens, while TPOM nanoparticles breakdown to release manganese ions and ovalbumin. More importantly, the released mt-DNA and Mn2+ synergistically activate the STING pathway, stimulating innate immunity in immature DCs, whereas ovalbumin and tumor-derived antigens enhance adaptive immunity. In an OS model, the TPOM nanoplatform was shown to promote antigen presentation, enhancing DC maturation and increasing the infiltration of cytotoxic T lymphocytes into tumor tissues.117 Furthermore, a polyethyleneimine (PEI)-modified aluminum hydroxide nanoparticle (LV@HPA/PEI) was developed, which promotes DC maturation and activation by increasing the expression of costimulatory molecules (such as CD80 and CD86), thereby improving their antigen cross-presentation ability.118
In recent years, cancer vaccines, as a crucial branch of immunotherapy, have garnered significant attention from scientists. As nanocarriers for tumor antigens and adjuvants, cancer vaccines can specifically target antigen-presenting cells and lymph nodes, triggering robust tumor-specific immune responses.133–135 Nanomaterials hold great promise in this field, with studies demonstrating that combining tumor vaccines with nanoparticles can substantially increase their efficacy.136,137 For example, Zhao et al developed a nanoparticle-based vaccine composed of ovalbumin (OVA), Mn2+, and meso-2,6-diaminopimelic acid (DAP) (OVA@Mn-DAP) (Figure 6A). In this formulation, OVA serves as a model protein antigen, Mn2+ provides imaging capabilities, and DAP activates the nucleotide oligomerization binding domain 1 (Nod1) signaling pathway, thereby increasing the cross-presentation of antigens by DCs.138,139 The experimental results revealed that this nanoparticle vaccine effectively promoted DC maturation (Figure 6B and C) and increased the expression of interleukin-12 (IL-12) and tumor necrosis factor-α (TNF-α) (Figure 6D and E), thereby increasing the immune response. Additionally, OVA@Mn-DAP can be tracked in vivo via magnetic resonance imaging (MRI) and fluorescence imaging, revealing its prolonged retention in the lymph nodes (Figure 6F–I). This suggests its potential as a preventive vaccine.119
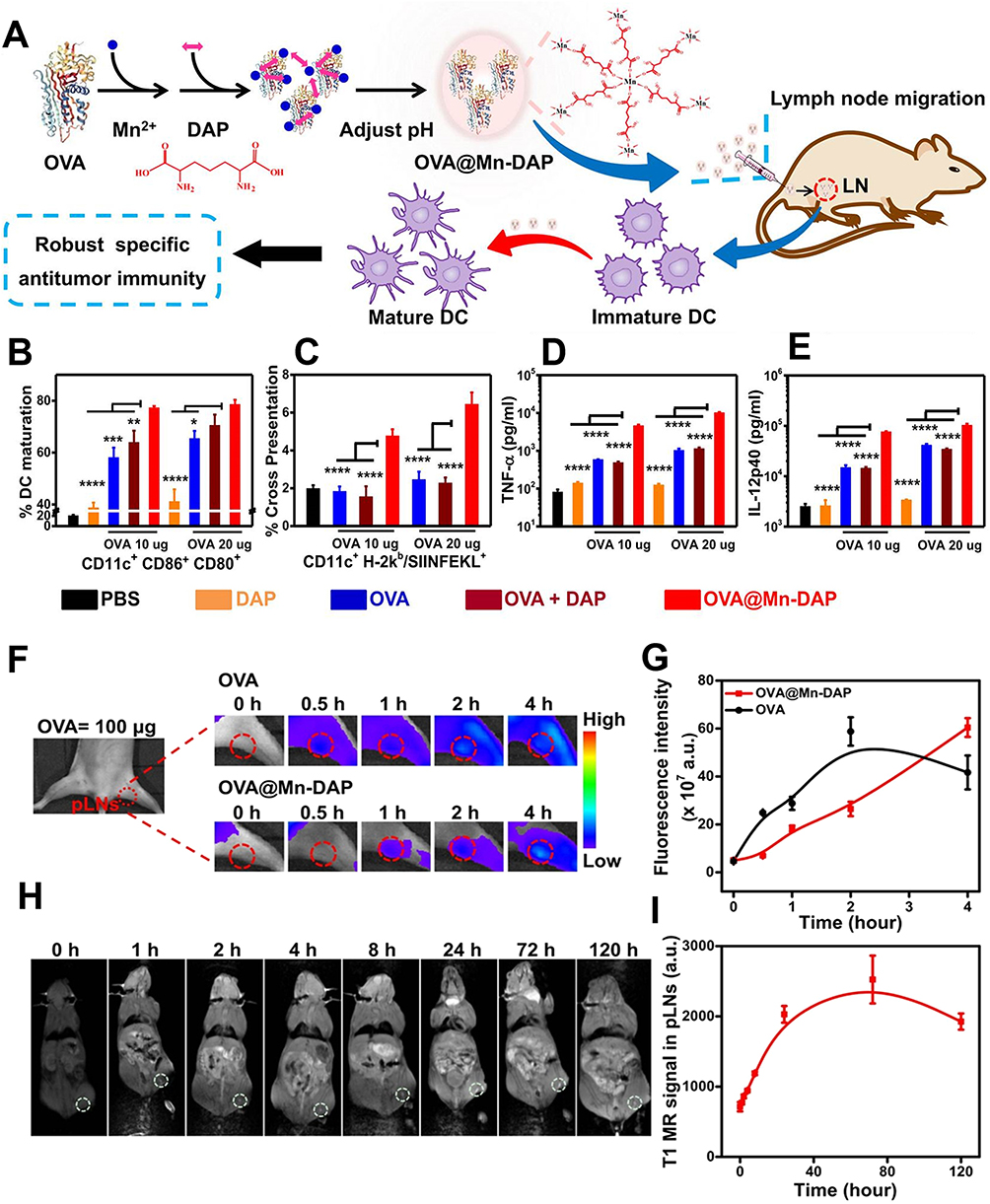 |
Figure 6 Nanovaccines participate in antitumor immunity by regulating DCs. (A) Schematic illustration of the synthesis and function of OVA@Mn-DAP nanoparticles. (B and C) Statistical data of DC maturation (B) and OVA antigen cross-presentation (C) in DCs treated with PBS, OVA, DAP, OVA + DAP, or OVA@Mn-DAP. CD11cCD80CD86 are markers of mature DCs. (D and E) TNF-α (H) and IL-12 p40 (I) were secreted from DCs after treatment with the various formulations indicated. (F and G) In vivo fluorescence images (F) and quantified fluorescence signals at the popliteal lymph nodes (pLNs) (G) after the injection of free OVA-Cy5.5 or the OVA-Cy5.5@Mn-DAP nanovaccine. (H and I) In vivo T1-weighted MR images (H) and quantified MR signals at the pLNs (I) (*P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. The red and white colored dotted circle represent the area where the popliteal lymph nodes (pLNs) are located).119 Copyright 2019, ACS Nano. |
Targeting Osteosarcoma Cells
Tumor cells often express specific biomarkers, creating opportunities for the development of nanotechnology-based targeting strategies. When nanoparticles are modified with high-affinity ligands (such as aptamers and peptides), they can selectively target these biomarkers and deliver therapeutic agents directly to tumor cells. For example, prostate-specific membrane antigen (PSMA) is overexpressed on the surface of most prostate cancer cells, and aptamers targeting PSMA have been developed for treating bone-metastatic prostate cancer.140 Human epidermal growth factor receptor-2 (HER2) is overexpressed in approximately 20%-30% of breast cancer patients, and trastuzumab, which targets HER2, can kill tumor cells, significantly improving patient prognosis.141,142 Research has also revealed that OS cells express a variety of specific biomarkers on their surface, including integrins, folate receptors, lectin receptors, CD44, and EphA2 (adrenaline α2 receptor). This review provides a detailed overview of nanodrug delivery systems targeting OS cell-specific biomarkers (Table 2).
 |
Table 2 Summary of Nanoparticles Targeting OS Cells |
Targeting Integrins
Integrins are a class of transmembrane glycoprotein adhesion molecules that primarily mediate interactions between cells and the extracellular matrix (ECM). They are expressed not only on tumor cells but also on tumor-associated host cells, such as endothelial cells and fibroblasts, where they regulate tumor cell survival, proliferation, and metastasis through signal transduction.152 The pivotal role of integrins in tumor biology makes them attractive targets for cancer therapy.
Arg-Gly-Asp (RGD) is a cell-adhesive peptide that specifically targets and binds to αvβ3 and αvβ5 integrins expressed on osteosarcoma cells.153,154 This strong affinity of RGD for osteosarcoma cells has led to the development of various RGD-modified nanoparticles, which have demonstrated promising tumor-suppressive effects.
Fang et al designed an RGD-modified DOX-loaded polymeric micelle (RGD-DOX-PM). Owing to their high affinity for integrins on MG-63 OS cells and efficient drug-loading capacity, these nanoparticles exhibited enhanced tumor targeting ability and cytotoxicity in vitro. In the experiment, MG-63 cells were divided into two groups: one group received pretreatment (ie, blocking αvβ3 and αvβ5 integrins on the cell membrane), whereas the other group did not. Compared with pretreated cells, untreated cells exhibited greater uptake of RGD-DOX-PM, and the inhibition of osteosarcoma cell proliferation was approximately six times greater.143 A cyclic RGD-modified, MEG3-loaded engineered exosome (cRGD-Exo-MEG3) was designed for the treatment of osteosarcoma. Compared with exosomes without cyclic RGD modification, cRGD-Exo-MEG3 demonstrated more effective tumor accumulation and stronger tumor-suppressive effects both in vitro and in vivo.144 Hu et al developed an RGD-modified, mitochondria-targeting ligand (TPP)-containing smart nanocomposite material (AIBI@H-mMnO2-TPP@PDA-RGD, AHTPR) for synergistically enhanced low-temperature photothermal therapy/thermodynamic therapy (LPTT/TDT) (Figure 7A and B). RGD modification allows AHTPR nanoparticles to target osteosarcoma cells. The acidic tumor microenvironment disrupts the PDA shell, and through the targeting ability of TPP, the nanoparticles accumulate in the mitochondria. After near-infrared light irradiation, AIBI decomposes and releases large amounts of free radicals. Moreover, MnO2 depletes intracellular GSH, significantly reducing the consumption of free radicals. Ultimately, the accumulation of free radicals in the mitochondria triggers tumor apoptosis (Figure 7C).145
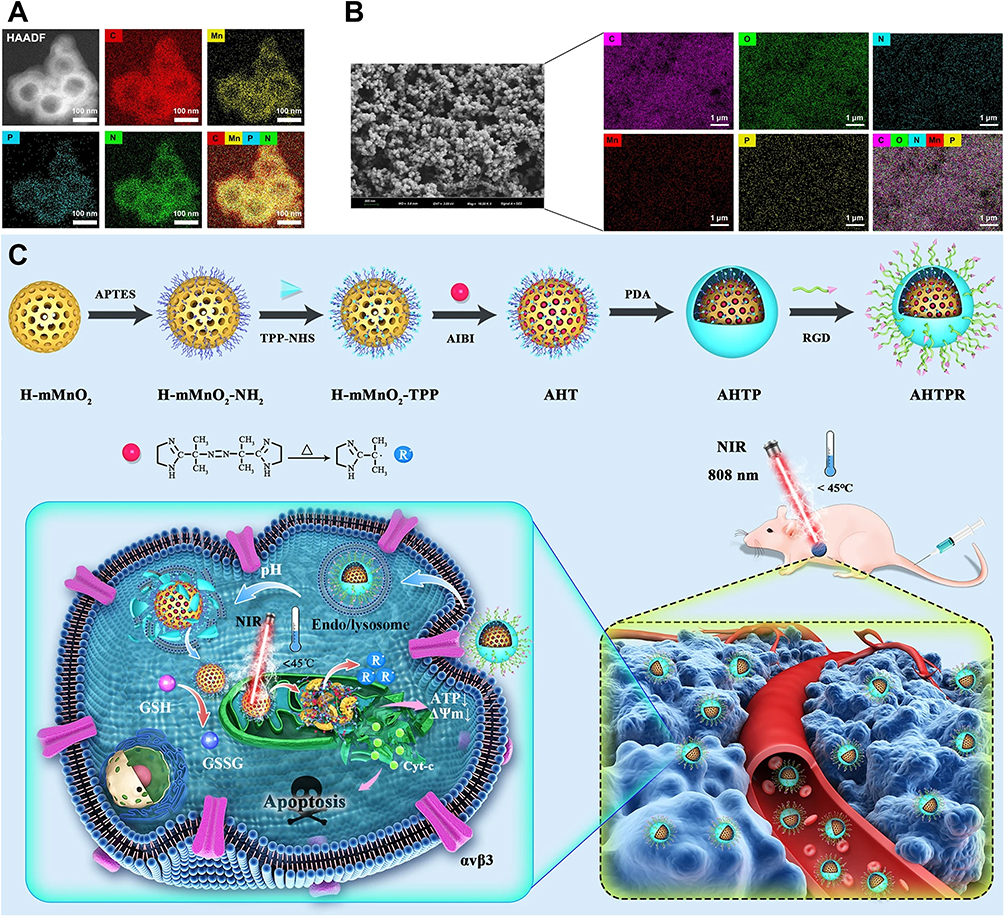 |
Figure 7 Design of AHTPR nanoparticles targeting integrins. (A and B) TEM mapping and SEM mapping of the AHTPR NPs. (C) Schematic illustration of the synthesis procedures of AHTPR NPs and the application of the NPs in synergistic mitochondrion-targeted LPTT/TDT.145 Different colored alphabets are abbreviations for chemical elements. The white arrows indicate that the quantities of ATP and ΔΨm in the mitochondria are showing a downward trend. The blue solid circle marked with “R” represents the oxygen-independent free radicals generated after AHTPR NPs act on mitochondria. Copyright 2021, Journal of Nanobiotechnology. |
Targeting VEGFR
Studies have shown that vascular endothelial growth factor receptor (VEGFR) is highly expressed on the surface of OS cells and may be involved in OS metastasis and immune evasion.155,156 Wu et al developed a biomimetic nanodrug system, V-RZCD, designed for VEGFR-targeted therapy for OS. V-RZCD consists of two components: a metal‒organic framework made from Ca2+ and zoledronic acid, which serves as the core and carries DOX, whereas VEGF-modified red blood cell membranes form the outer shell. The results showed that V-RZCD specifically binds to highly expressed VEGF receptors on osteosarcoma cells, significantly inhibiting OS cell proliferation while also reducing osteolysis.146
Targeting CD44
CD44 is an adhesion molecule that is highly expressed on the surface of OS cells. It binds to hyaluronic acid (HA) and mediates interactions between cells and the extracellular matrix, playing a key role in tumor proliferation, metastasis, and the development of multidrug resistance.157–159 HA, which serves as a targeting ligand for CD44, has been utilized in targeted drug delivery systems for OS cells. For example, a nanogel composed of cisplatin (CDDP) and crosslinked with HA, known as CDDPHANG, has been designed for the delivery of DOX in OS therapy. In addition to its direct antitumor effects, CDDP helps stabilize drug-loaded nanogels, extending their circulation time in the bloodstream. Under the acidic conditions of the TME, the nanogel disassembles rapidly, releasing both DOX and CDDP, thus demonstrating a synergistic antitumor effect.147 Xu et al developed hyaluronic acid/polyethylene glycol/nanohydroxyapatite nanoparticles (HA-PEG-nHA-ZOL NPs) loaded with zoledronic acid (ZOL), which have significant therapeutic potential. The specific binding of HA to CD44, combined with the acid-sensitive release of PEG, allows for targeted delivery to OS cells and the controlled release of ZOL. This approach not only extends the drug’s action time but also reduces toxicity to normal cells. In vivo experiments demonstrated that local injection of these nanoparticles triggered vascular inflammation in the tumor, inducing tumor necrosis and apoptosis.148
Peptide-Based Targeting
Targeted peptide-modified nanoparticles can specifically bind to biomarkers on the surface of OS cells, significantly enhancing their targeting efficiency. Peptides offer several advantages, including ease of synthesis, low immunogenicity, high biocompatibility and cost-effectiveness.160,161 As a result, targeted peptide-modified nanoparticles have been widely studied to improve drug accumulation in OS cells.
EphA2, a receptor tyrosine kinase, is overexpressed on both primary and metastatic osteosarcoma cells and may play a role in tumor invasion and metastasis.162 YSA (YSAYPDSVPMMS) is a 12-amino acid peptide that specifically targets EphA2, making it an ideal ligand for modifying nanoparticles.163,164 Haghiralsadat et al utilized YSA peptide-modified polyethylene glycolylated liposomes, referred to as YSA-L-DOX, to develop targeted nanovesicles for DOX delivery. The results showed that the YSA conjugation efficiency exceeded 98%, with no effect on the drug-loading efficiency of the polyethylene glycolylated liposomes. Moreover, compared with free DOX, YSA-L-DOX exhibited nearly twice the toxicity to Saos-2 cells.149
Cell surface vimentin is a key biomarker in OS, with its high expression closely linked to metastatic phenotypes.165,166 The peptide VATANST (STP) can specifically bind to vimentin expressed on the surface of OS cells, enabling targeted delivery to OS. On this basis, STP-modified, DOX-loaded mPEG-P(Phe-co-Cys) nanoparticles (Figure 8A) were developed. These nanoparticles facilitate controlled release of DOX, increasing its anti-osteosarcoma effects while significantly reducing DOX-induced side effects.150 Li et al also developed STP-modified nanogels (STP-PEG-P(LP-co-LC)) for targeted delivery of SHK to OS cells, denoted as STP-NG/SHK (Figure 8B). With the help of STP, STP-NG/SHK exhibited stronger tumor-suppressive effects than did NG/SHK or SHK alone, with these effects mediated via RIP1- and RIP3-dependent necroptosis (Figure 8C and D). Additionally, in mouse models, STP-NG/SHK significantly reduces lung metastasis while decreasing systemic toxicity.151
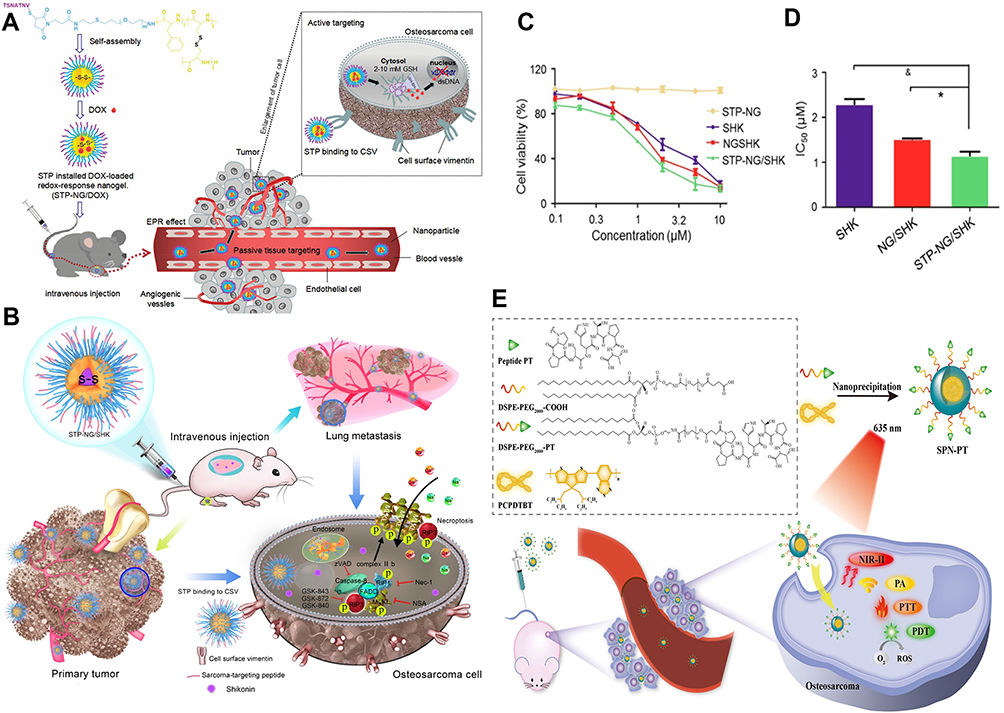 |
Figure 8 Design of nanoparticle schemes for targeted peptide modification. (A) Schematic illustration of STP-NP/DOX development and its antitumor mechanisms.150 The red dotted circle represents the area where the drug interacts with the tumor after being injected intravenously. The black-colored dotted square box represents the area where the drug interacts with the tumor cells. Copyright 2020, Drug Delivery. (B) Schematic illustration of STP-NG/SHK and RIP1- and RIP3-dependent cell necroptosis in osteosarcoma and lung metastasis. (C) The cell viability of 143B cells was detected. (D) IC50s of SHK, NG/SHK, and STP-NG/SHK after 24 h (*P < 0.05, &P < 0.001).151 The blue-colored circles represent the targeted, loaded nanogel, STP-NG/SHK. Copyright 2018, Theranostics. (E) Schematic illustration of the preparation process of SPN-PT and NIR-II fluorescence/NIR-I PA imaging and targeted PTT/PDT against osteosarcoma.167 Copyright 2022, Journal of Nanobiotechnology. |
The peptide PT (PPSHTPT) was designed to mimic the properties of natural osteocalcin proteins and has been shown to target osteosarcoma cell lines.168,169 Semiconductor polymer nanoparticles (SPNs) are a novel class of organic optical nanomaterials that are widely utilized in fluorescence imaging, PTT, and PDT.170–172 Yuan et al designed and fabricated a peptide-based SPN (SPN-PT) (Figure 8E) for OS-targeted PTT/PDT. Compared with the controls, the PT-modified nanoparticles were rapidly internalized by OS cells, completing uptake within 4 hours. They also enhanced NIR-II fluorescence and photoacoustic signals, providing high-sensitivity, high-quality tumor imaging.167
Targeting the Vascular Microenvironment
Neovascularization in OS is a complex and adaptive process that plays a vital role in tumor growth and metastasis. Currently, several nanoparticle drug delivery systems with antiangiogenic properties have been developed to target and disrupt the angiogenesis process (Table 3).
 |
Table 3 Summary of Nanoparticles Targeting Angiogenesis and Stem Cells |
As one of the most extensively studied proangiogenic factors, VEGF has become a prominent target for the development of nanoparticle-based drugs. Kovach et al conjugated VEGF antibodies and CD80 to iron oxide nanoparticles to achieve a synergistic therapeutic effect. CTLA-4 is expressed on OS cells, and CD80, as its ligand, binds to CTLA-4, inducing OS apoptosis. Experimental results showed that a combination of 1 μg/mL CD80 and VEGF antibodies significantly reduced osteoblast proliferation and induced cell death.173 As previously mentioned, the binding of VEGF to its receptor, VEGFR, triggers the release of matrix metalloproteinases, which promote angiogenesis. To address this, Li et al developed an amphiphilic LMWH-U (LHU) nanoparticle system that covalently combines low-molecular-weight heparin (LMWH) and ursolic acid (UA) to inhibit matrix metalloproteinases. They also modified LHU with DSPE-PEG-AA to obtain A-LHU nanoparticles, improving drug delivery to the tumor. This nanoparticle demonstrated strong antiangiogenic effects both in vitro and in vivo, significantly inhibiting tumor growth in a mouse model.174
Zhao et al designed poly(lactic-co-glycolic acid)-polyethylene glycol (PLGA-PEG) nanoparticles for the codelivery of albendazole (ABZ) and DOX to treat OS, named AD@PLGA-PEG NPs. The results showed that the AD@PLGA-PEG NPs promoted mitochondria-mediated oxidative stress and induced tumor apoptosis by increasing ROS production. More importantly, it significantly downregulates the expression of hypoxia-inducible factor-1α (HIF-1α), thereby inhibiting the expression of its downstream protein VEGF.175 These findings highlight the potential of targeting the hypoxic TME for OS therapy.
Targeting Cancer Stem Cells
Cancer stem cells (CSCs) are the primary tumor-initiating cells and are closely associated with tumor recurrence, metastasis, and multidrug resistance.182,183 Therefore, targeting and eliminating OS stem cells could significantly improve the therapeutic outcomes of OS treatment. Herein, we will summarize the latest research advances in nanoparticles targeting CSCs for osteosarcoma therapy (Table 3). All-trans retinoic acid (ATRA) can target OS stem cells and inhibit the progression of OS. For example, ATRA effectively suppresses the stemness of OS cells by blocking the M2 polarization of TAMs.74 Additionally, ATRA inhibits tumor initiation and differentiation by blocking the Wnt signaling pathway in CSCs.184,185 Zhang et al designed a tetrahedral framework nucleic acid (tFNA) to load ATRA into OS cells, referred to as T-ATRA (Figure 9A). Compared with ATRA alone, these novel nanoparticles significantly enhanced the ability to eliminate OS stem cells through the Wnt signaling pathway (Figure 9B–F). Experimental results also demonstrated that T-ATRA effectively inhibited OS cell proliferation, metastasis, and invasion, significantly prolonging the survival of OS model mice (Figure 9G–J).176 Lu et al developed graphene oxide (GO)-based nanoparticles loaded with ginsenoside Rg3 (PEG-GO-FA/ICG-Rg3) to improve the photodynamic therapy of osteosarcoma. After near-infrared laser treatment, PEG-GO-FA/ICG-Rg3 significantly increased the suppression of OS cell growth and migration while also inhibiting the stemness of OS cell-derived CSCs.177
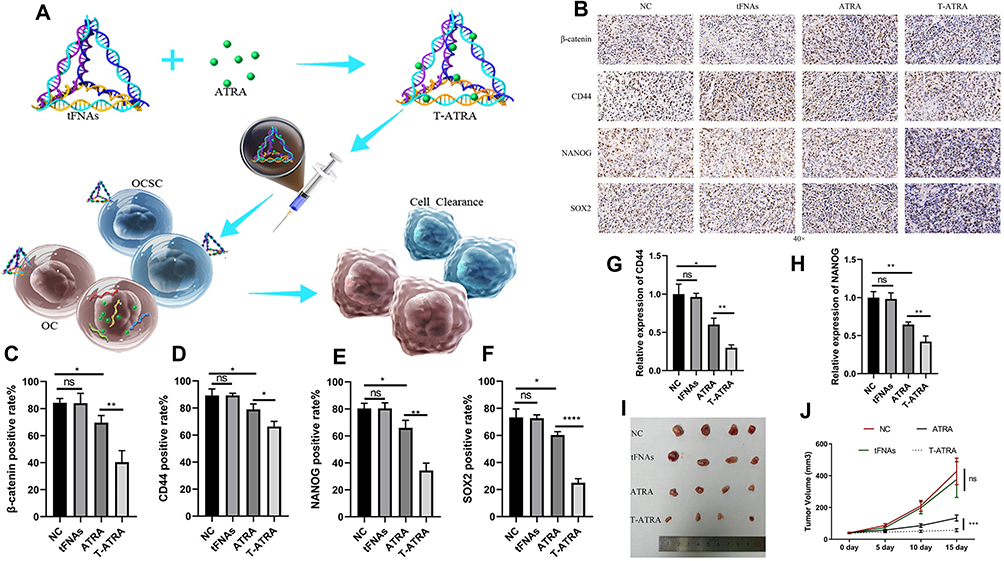 |
Figure 9 Retinoic acid carried by tetrahedral framework nucleic acids is used to promote the clearance of osteosarcoma stem cells. (A) Schematic illustration of T-ATRA and its antiosteosarcoma mechanism. (B) β-Catenin, CD44, NANOG and SOX2 protein immunohistochemistry. (C) Immunohistochemical quantification of the β-catenin protein. (D) Immunohistochemical quantification of the CD44 protein. (E) Immunohistochemical quantification of NANOG protein. (F) Immunohistochemical quantification of the SOX2 protein. (G) Quantification of CD44 protein of WB. (H) Quantification of NANOG protein of WB. (I) Subcutaneous tumorigenesis assay of OS in nude mice. (J) Changes in the volume of tumors in each group (n = 4). The Wnt/β-catenin signaling pathway is considered a key signaling pathway in osteosarcoma stem cells. CD44, NANOG and SOX2 are considered markers closely related to osteosarcoma stem cells (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, “ns” indicates no statistically significant difference).176 Copyright 2024, ACS Applied Materials & Interfaces. |
Studies have shown that CD133 is a specific biomarker of OS stem cells.186 CD133-expressing OS cells exhibit stem cell-like properties, such as high proliferative capacity, multilineage differentiation potential and elevated levels of drug resistance proteins.187,188 Gui et al developed a CD133 aptamer-labeled lipid‒polymer nanoparticle for the targeted delivery of all-trans retinoic acid (ATRA) to OS stem cells. Compared with free ATRA and nontargeted nanoparticles, ATRA-PLNP-CD133 demonstrated stronger targeting and therapeutic effects, confirming that CD133 aptamers can effectively deliver nanoparticles to OS stem cells.178
Similarly, Ni et al developed CD133-modified salinomycin-loaded PLGA nanoparticles (Ap-SAL-NPs), which effectively deliver salinomycin to Saos-2 CD133+ cells. Compared with SAL-NPs and salinomycin alone, Ap-SAL-NPs significantly reduced the proportion of CD133+ OS CSCs, indicating selective toxicity to CD133+ OS CSCs.179 CD271 is another surface biomarker of OS stem cells.189,190 Consequently, Tian et al conjugated a CD271 monoclonal antibody with polyethylene glycol-modified hollow gold nanoparticles (HGN) to create a novel nanomaterial (HGNs-PEG-CD271) for specific photothermal ablation of OS CSCs. Following NIR laser irradiation, the percentage of apoptotic cells in the HGN-PEG-CD271 group was significantly greater than that in the HGN-PEG group, and the expression of γ-H2AX (a marker of DNA double-strand breaks) was significantly greater in the former. This finding suggests that HGN-PEG-CD271 may target and kill OS stem cells through apoptotic pathways and DNA double-strand damage.180
Targeting MSCs
MSCs play a crucial role in regulating the OS microenvironment by secreting various cytokines, growth factors, chemokines, and MMPs, all of which are essential for tumor progression.191,192 The acidic microenvironment created by tumor cell metabolism further stimulates MSCs to secrete protumor mediators, thereby driving tumor progression.193,194 Research suggests that this process may be linked to the activation of the NF-κB pathway, which contributes to the enhanced protumor phenotype of MSCs.195 Curcumin, an NF-κB inhibitor, can promote OS apoptosis; however, its chemical instability and low bioavailability limit its therapeutic potential.196,197 To enhance curcumin delivery, Gemma et al developed spherical polymer nanoparticles (SPN-curc) capable of loading curcumin. Compared with free curcumin, SPN-curc significantly reduced the secretion of the inflammatory cytokines IL-6 and IL-8, effectively preventing the induction of OS stemness and reducing OS migration and invasion induced by MSCs181 (Table 3).
Additionally, MSCs are widely utilized as drug delivery carriers owing to their unique ability to migrate to and colonize the tumor stroma.198,199 For example, Duchi et al designed fluorescence nanoparticles (FNPs) coated with the photosensitizer tetra-phenyl sulfonated porphyrin (TPPS) and loaded them into MSCs for targeted photodynamic therapy of osteosarcoma. Owing to the tumor-homing ability of MSCs, TPPS@FNPs can specifically accumulate at the tumor site. When exposed to 405 nm laser light, reactive oxygen species (ROS) are generated, leading to rapid OS cell death.200 Similarly, Lenna et al used tetra-sulfonated aluminum phthalocyanine (AlPcS4) as a photosensitizer in combination with FNPs to create AlPcS4@FNPs. These nanoparticles could be internalized by MSCs within one hour and retained for at least three days. After light irradiation, the AlPcS4@FNPs loaded in MSCs induced OS cell death and reduced the tumor burden compared with those in the control group (PBS and free AlPcS4). More importantly, compared with mice injected with free AlPcS4@FNPs, the AlPcS4@FNPs@MSC group demonstrated a smaller drug distribution range, confirming the tumor-targeting ability of MSCs.201 Wei et al isolated exosomes from bone marrow mesenchymal stem cells (BM-MSCs) as nanocarriers for DOX in OS therapy. BM-MSC-derived exosomes efficiently delivered DOX to OS tissues, inhibited OS cell proliferation and migration both in vitro and in vivo, and significantly reduced cardiotoxicity. Further studies revealed that the targeting ability of these exosomes stems from the interaction between the CXCR4 protein expressed on OS cells and the SDF-1 protein expressed on both exosomes and MSCs.202
Conclusions and Future Perspectives
In this review, we systematically summarize the components of the OS TME and their pivotal roles in the initiation and progression of OS. Notably, during the early stages of disease, the emergence of OS cells disrupts bone homeostasis. Through the secretion of cytokines, growth factors, and the RANKL-RANK pathway, tumor cells and osteoclasts form a “vicious cycle” that significantly accelerates the early progression of OS. MSCs, a major source of OS cells, also coordinate signaling pathways within the TME by secreting extracellular vesicles and various cytokines, contributing to tumor-promoting activities. Hypoxic and acidic microenvironments stimulate the expression of hypoxia-inducible factor (HIF) and VEGF, which, in turn, promote angiogenesis. These newly formed blood vessels supply oxygen and nutrients to tumor cells while providing pathways for OS metastasis. TAMs, lymphocytes, dendritic cells, and other immune cells together create an immunosuppressive microenvironment, helping OS cells evade immune surveillance and participate in tumor maintenance and migration. The complexity of the TME presents challenges in understanding the pathophysiological mechanisms of OS, but it also guides the exploration of new therapeutic strategies.
Nanoparticles developed for targeting the tumor microenvironment of OS have demonstrated significant therapeutic potential, offering new hope for OS patients. However, numerous unresolved issues remain before their successful translation from laboratory research to clinical application, and future studies need to focus on specific directions for targeted investigations. Firstly, the safety of nanoparticles has not been fully guaranteed. Studies have shown that some NPs can accumulate in the body for a long time, inducing chronic toxicity and impairing the functions of organs such as the liver and kidneys.203 Meanwhile, since some OS-related biomarkers are not exclusive to tumor cells, single-ligand targeted nanoparticles are prone to off-target effects, which further exacerbates safety risks. Therefore, the identification and validation of OS-specific TME-targeting biomarkers represent one of the key research directions. In particular, the exploration of specific targets such as certain integrins, chemokine receptors, or extracellular matrix components can provide precise anchors for targeted delivery. Additionally, the development of multi-ligand nanoparticles that simultaneously target multiple TME components can also effectively improve targeting specificity and reduce off-target risks.
Secondly, nanoparticles face a high rate of immune clearance and are easily recognized and eliminated by the reticuloendothelial system and macrophages, which greatly limits their in vivo circulation time and therapeutic efficacy. Studies have reported that immune clearance can be reduced through surface modifications such as PEGylation and optimization of physicochemical properties—for example, maintaining the particle size between 10~100 nm to balance circulatory stability and tumor tissue permeability,204,205 or selecting neutral nanoparticles with the lowest affinity for macrophages.206 Meanwhile, the research and application of theranostic nanoplatforms are worthy of focused exploration. By integrating imaging and therapeutic functions, these platforms can enable patient stratification, real-time monitoring of TME regulation processes, and accurate evaluation of treatment responses, thereby providing support for personalized therapy.
Furthermore, interpatient and intratumoral heterogeneity in OS is a critical challenge affecting therapeutic outcomes. The development of adaptive or personalized TME-targeting nanoplatforms, which customize delivery systems based on the TME characteristics of individual patients, is expected to overcome the therapeutic bottlenecks caused by heterogeneity. Finally, the high production costs, insufficient large-scale industrial production capacity, and the lack of unified international regulatory guidelines leading to difficulties in consistent quality evaluation collectively hinder the clinical translation of nano-targeted drugs. In summary, nanotechnology-based drug delivery strategies hold tremendous potential for osteosarcoma treatment. Despite the numerous challenges inherent in translating these approaches from the laboratory to the clinic, we firmly believe they represent the advancing direction of modern medicine.
Abbreviations
OS, Osteosarcoma; TME, tumor microenvironment; DOX, doxorubicin; ECM, extracellular matrix; MDM2, Mouse Double Minute 2 Homolog; TAMs, Tumor-associated macrophages; CTLs, cytotoxic T lymphocytes; OCs, Osteoclasts; HSCs, hematopoietic stem cells; RANKL, receptor activator of nuclear factor-kappa B ligand; TNF, tumor necrosis factor; MSCs, mesenchymal stem cells; BMSCs, bone marrow stromal cells; EVs, extracellular vesicles; MMP, matrix metalloproteinase; EPCs, Endothelial progenitor cells; HIF, hypoxia-inducible factor; DCs, dendritic cells; Tregs, regulatory T cells; VEGF, vascular endothelial growth factor; VEGFR, vascular endothelial growth factor receptor; PD-L1, programmed death ligand-1; ATRA, All-trans retinoic acid; TLR4, Toll-like receptor 4; BPs, Bisphosphonates; ALN, alendronate; ZA/ZOL, zoledronic acid; LNPs, lipid nanoparticles; HAP, hydroxyapatite; PA, Phytic acid; PTT, photothermal therapy; PDT, photodynamic therapy; MRI, magnetic resonance imaging, MPI, magnetic particle imaging; PAT, photoacoustic tomography; PEI, polyethyleneimine; OVA, ovalbumin; DENPs, dendrimer-entrapped gold nanoparticles; BGns, bioactive glass nanoparticles; METTL14, methyltransferase-like 14; Rh2, ginsenoside; PSMA, prostate-specific membrane antigen; HER2, Human epidermal growth factor receptor-2; HA, hyaluronic acid; CDDP, cisplatin; HSFs, human skin fibroblasts; VIM, vimentin; SHK, shikonin; CSCs, Cancer stem cells; GO, graphene oxide.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
All authors declare that they have no competing interests in this work.
References
1. Kansara M, Teng MW, Smyth MJ, Thomas DM. Translational biology of osteosarcoma. Nat Rev Cancer. 2014;14(11):722–28. doi:10.1038/nrc3838
2. Le Vu B, de Vathaire F, Shamsaldin A, et al. Radiation dose, chemotherapy and risk of osteosarcoma after solid tumours during childhood. Int J Cancer. 1998;77(3):370–377. doi:10.1002/(SICI)1097-0215(19980729)77:3<370::AID-IJC11>3.0.CO;2-C
3. Cotterill SJ, Wright CM, Pearce MS, Craft AW. Stature of young people with malignant bone tumors. Pediatr Blood Cancer. 2004;42(1):59–63. doi:10.1002/pbc.10437
4. Gianferante DM, Mirabello L, Savage SA. Germline and somatic genetics of osteosarcoma — connecting aetiology, biology and therapy. Nat Rev Endocrinol. 2017;13(8):480–491. doi:10.1038/nrendo.2017.16
5. Mirabello L, Troisi RJ, Savage SA. Osteosarcoma incidence and survival rates from 1973 to 2004: data from the surveillance, epidemiology, and end results program. Cancer. 2009;115(7):1531–1543. doi:10.1002/cncr.24121
6. Smith J, Botet JF, Yeh SD. Bone sarcomas in Paget disease: a study of 85 patients. Radiology. 1984;152(3):583–590. doi:10.1148/radiology.152.3.6235535
7. Chiesa AM, Spinnato P, Miceli M, Facchini G. Radiologic assessment of osteosarcoma lung metastases: state of the art and recent advances. Cells. 2021;10(3):553. doi:10.3390/cells10030553
8. Shen Y, Huang S, Chen G, Wang G, Sui L. Involvement of TP53 in osteosarcoma - challenges and prospects. Front Oncol. 2025;15:1605080. doi:10.3389/fonc.2025.1605080
9. Ritter J, Bielack SS. Osteosarcoma. Ann Oncol. 2010;21:vii320–5. doi:10.1093/annonc/mdq276
10. Tian H, Cao J, Li B, et al. Managing the immune microenvironment of osteosarcoma: the outlook for osteosarcoma treatment. Bone Res. 2023;11(1):11. doi:10.1038/s41413-023-00246-z
11. Ji Y, Harris MA, newton LM, et al. Osteosarcoma cells depend on MCL-1 for survival, and osteosarcoma metastases respond to MCL-1 antagonism plus regorafenib in vivo. BMC Cancer. 2024;24(1):1350. doi:10.1186/s12885-024-13088-7
12. Link MP, Goorin AM, Miser AW, et al. The effect of adjuvant chemotherapy on relapse-free survival in patients with osteosarcoma of the extremity. N Engl J Med. 1986;314(25):1600–1606. doi:10.1056/NEJM198606193142502
13. Alfranca A, Martinez-Cruzado L, Tornin J, et al. Bone microenvironment signals in osteosarcoma development. Cell Mol Life Sci. 2015;72(16):3097–3113. doi:10.1007/s00018-015-1918-y
14. Gu W, Wu C, Chen J, Xiao Y. Nanotechnology in the targeted drug delivery for bone diseases and bone regeneration. Int J Nanomed. 2013;8:2305–2317. doi:10.2147/IJN.S44393
15. Amiryaghoubi N, Fathi M, Barar J, Omidian H, Omidi Y. Advanced nanoscale drug delivery systems for bone cancer therapy. Biochimica et Biophysica Acta. 2023;1869(6):166739. doi:10.1016/j.bbadis.2023.166739
16. Corre I, Verrecchia F, Crenn V, Redini F, Trichet V. The osteosarcoma microenvironment: a complex but targetable ecosystem. Cells. 2020;9(4):976. doi:10.3390/cells9040976
17. Dutour A, Pasello M, Farrow L, et al. Microenvironment matters: insights from the FOSTER consortium on microenvironment-driven approaches to osteosarcoma therapy. Cancer Metastasis Rev. 2025;44(2):44. doi:10.1007/s10555-025-10257-3
18. Lamoureux F, Richard P, Wittrant Y, et al. Therapeutic relevance of osteoprotegerin gene therapy in osteosarcoma: blockade of the vicious cycle between tumor cell proliferation and bone resorption. Cancer Res. 2007;67(15):7308–7318. doi:10.1158/0008-5472.CAN-06-4130
19. Bolamperti S, Villa I, Rubinacci A. Bone remodeling: an operational process ensuring survival and bone mechanical competence. Bone Res. 2022;10(1):48. doi:10.1038/s41413-022-00219-8
20. Jacome-Galarza CE, Percin GI, Muller JT, et al. Developmental origin, functional maintenance and genetic rescue of osteoclasts. Nature. 2019;568(7753):541–545. doi:10.1038/s41586-019-1105-7
21. Xu F, Teitelbaum SL. Osteoclasts: new insights. Bone Res. 2013;1(1):11–26. doi:10.4248/BR201301003
22. Lamora A, Talbot J, Mullard M, Brounais-Le Royer B, Redini F, Verrecchia F. TGF-β signaling in bone remodeling and osteosarcoma progression. J Clin Med. 2016;5(11):96. doi:10.3390/jcm5110096
23. Endo-Munoz L, Cumming A, Rickwood D, et al. Loss of osteoclasts contributes to development of osteosarcoma pulmonary metastases. Cancer Res. 2010;70(18):7063–7072. doi:10.1158/0008-5472.CAN-09-4291
24. Lu KH, Lu EWH, Lin CW, Yang JS, Yang SF. New insights into molecular and cellular mechanisms of zoledronate in human osteosarcoma. Pharmacol Ther. 2020;214:107611. doi:10.1016/j.pharmthera.2020.107611
25. Endo-Munoz L, Evdokiou A, Saunders NA. The role of osteoclasts and tumour-associated macrophages in osteosarcoma metastasis. Biochimica et Biophysica Acta. 2012;1826(2):434–442. doi:10.1016/j.bbcan.2012.07.003
26. Nishimura R, Wakabayashi M, Hata K, et al. Osterix regulates calcification and degradation of chondrogenic matrices through matrix metalloproteinase 13 (MMP13) expression in association with transcription factor Runx2 during endochondral ossification. J Biol Chem. 2012;287(40):33179–33190. doi:10.1074/jbc.M111.337063
27. Juárez P, Guise TA. TGF-β in cancer and bone: implications for treatment of bone metastases. Bone. 2011;48(1):23–29. doi:10.1016/j.bone.2010.08.004
28. Ponzetti M, Ucci A, Puri C, et al. Effects of osteoblast-derived extracellular vesicles on aggressiveness, redox status and mitochondrial bioenergetics of MNNG/HOS osteosarcoma cells. Front Oncol. 2022;12. doi:10.3389/fonc.2022.983254
29. Patanè S, Avnet S, Coltella N, et al. MET overexpression turns human primary osteoblasts into osteosarcomas. Cancer Res. 2006;66(9):4750–4757. doi:10.1158/0008-5472.CAN-05-4422
30. Buenzli PR, Sims NA. Quantifying the osteocyte network in the human skeleton. Bone. 2015;75:144–150. doi:10.1016/j.bone.2015.02.016
31. Dwivedi A, Kiely PA, Hoey DA. Mechanically stimulated osteocytes promote the proliferation and migration of breast cancer cells via a potential CXCL1/2 mechanism. Biochem Biophys Res Commun. 2021;534:14–20. doi:10.1016/j.bbrc.2020.12.016
32. Wang W, Yang X, Dai J, Lu Y, Zhang J, Keller ET. Prostate cancer promotes a vicious cycle of bone metastasis progression through inducing osteocytes to secrete GDF15 that stimulates prostate cancer growth and invasion. Oncogene. 2019;38(23):4540–4559. doi:10.1038/s41388-019-0736-3
33. Sottnik JL, Campbell B, Mehra R, Behbahani-Nejad O, Hall CL, Keller ET. Osteocytes serve as a progenitor cell of osteosarcoma. J Cell Biochem. 2014;115(8):1420–1429. doi:10.1002/jcb.24793
34. Wang J, Wu P, Chen PC, Lee C, Chen W, Hung S. Generation of osteosarcomas from a combination of Rb silencing and c‐myc overexpression in human mesenchymal stem cells. Stem Cells Transl Med. 2017;6(2):512–526. doi:10.5966/sctm.2015-0226
35. Sun Z, Wang S, Zhao RC. The roles of mesenchymal stem cells in tumor inflammatory microenvironment. J Hematol Oncol. 2014;7(1):14. doi:10.1186/1756-8722-7-14
36. Chaturvedi P, Gilkes DM, Wong CCL, et al. Hypoxia-inducible factor–dependent breast cancer–mesenchymal stem cell bidirectional signaling promotes metastasis. J Clin Invest. 2013;123(1):189–205. doi:10.1172/JCI69244
37. Zheng Y, Wang G, Chen R, Hua Y, Cai Z. Mesenchymal stem cells in the osteosarcoma microenvironment: their biological properties, influence on tumor growth, and therapeutic implications. Stem Cell Res Ther. 2018;9(1):22. doi:10.1186/s13287-018-0780-x
38. Chang AI, Schwertschkow AH, Nolta JA, Wu J. Involvement of mesenchymal stem cells in cancer progression and metastases. Curr Cancer Drug Targets. 2015;15(2):88–98. doi:10.2174/1568009615666150126154151
39. Tu B, Zhu J, Liu S, et al. Mesenchymal stem cells promote osteosarcoma cell survival and drug resistance through activation of STAT3. Oncotarget. 2016;7(30):48296–48308. doi:10.18632/oncotarget.10219
40. Vallabhaneni KC, Penfornis P, Dhule S, et al. Extracellular vesicles from bone marrow mesenchymal stem/stromal cells transport tumor regulatory microRNA, proteins, and metabolites. Oncotarget. 2014;6(7):4953–4967. doi:10.18632/oncotarget.3211
41. Mannerström B, Kornilov R, Abu-Shahba AG, et al. Epigenetic alterations in mesenchymal stem cells by osteosarcoma-derived extracellular vesicles. Epigenetics. 2019;14(4):352–364. doi:10.1080/15592294.2019.1585177
42. Huang Y, Liu W, He B, et al. Exosomes derived from bone marrow mesenchymal stem cells promote osteosarcoma development by activating oncogenic autophagy. J Bone Oncol. 2020;21:100280. doi:10.1016/j.jbo.2020.100280
43. Qin F, Tang H, Zhang Y, Zhang Z, Huang P, Zhu J. Bone marrow-derived mesenchymal stem cell-derived exosomal microRNA-208a promotes osteosarcoma cell proliferation, migration, and invasion. J Cell Physiol. 2020;235(5):4734–4745. doi:10.1002/jcp.29351
44. Zhang H, Wang J, Ren T, et al. Bone marrow mesenchymal stem cell-derived exosomal miR-206 inhibits osteosarcoma progression by targeting TRA2B. Cancer Lett. 2020;490:54–65. doi:10.1016/j.canlet.2020.07.008
45. Lee SW, Jeon TJ, Biswal S. Effect of local treatment with adipose tissue-derived mesenchymal stem cells in the early tumorigenesis of osteosarcoma. Oncol Rep. 2015;33(3):1381–1387. doi:10.3892/or.2015.3711
46. Marçola M, Rodrigues CE. Endothelial progenitor cells in tumor angiogenesis: another brick in the wall. Stem Cells Int. 2015;2015:832649. doi:10.1155/2015/832649
47. An R, Schmid R, Klausing A, et al. Proangiogenic effects of tumor cells on endothelial progenitor cells vary with tumor type in an in vitro and in vivo rat model. FASEB J. 2018;32(10):5587–5601. doi:10.1096/fj.201800135RR
48. Beck B, Driessens G, Goossens S, et al. A vascular niche and a VEGF-Nrp1 loop regulate the initiation and stemness of skin tumours. Nature. 2011;478(7369):399–403. doi:10.1038/nature10525
49. Lv X, Li J, Zhang C, et al. The role of hypoxia-inducible factors in tumor angiogenesis and cell metabolism. Genes Dis. 2016;4(1):19–24. doi:10.1016/j.gendis.2016.11.003
50. Liao D, Johnson RS. Hypoxia: a key regulator of angiogenesis in cancer. Cancer Metastasis Rev. 2007;26(2):281–290. doi:10.1007/s10555-007-9066-y
51. Ohba T, Cates JMM, Cole HA, et al. Autocrine VEGF/VEGFR1 signaling in a subpopulation of cells associates with aggressive osteosarcoma. Mol Cancer Res. 2014;12(8):1100–1111. doi:10.1158/1541-7786.MCR-14-0037
52. van Hinsbergh VWM, Koolwijk P. Endothelial sprouting and angiogenesis: matrix metalloproteinases in the lead. Cardiovasc Res. 2008;78(2):203–212. doi:10.1093/cvr/cvm102
53. Lugano R, Ramachandran M, Dimberg A. Tumor angiogenesis: causes, consequences, challenges and opportunities. Cell Mol Life Sci. 2020;77(9):1745–1770. doi:10.1007/s00018-019-03351-7
54. Lammli J, Fan M, Rosenthal HG, et al. Expression of vascular endothelial growth factor correlates with the advance of clinical osteosarcoma. Int Orthop. 2012;36(11):2307–2313. doi:10.1007/s00264-012-1629-z
55. Chen D, Zhang YJ, Wei ZK, Wang WC. A systematic review of vascular endothelial growth factor expression as a biomarker of prognosis in patients with osteosarcoma. Tumor Biol. 2013;34(3):1895–1899. doi:10.1007/s13277-013-0733-z
56. Tao SC, Huang JY, Wei ZY, Li ZX, Guo SC. EWSAT1 acts in concert with exosomes in osteosarcoma progression and tumor-induced angiogenesis: the “double stacking effect. Adv Biosyst. 2020;4(9):2000152. doi:10.1002/adbi.202000152
57. Li Y, Lin S, Xie X, Zhu H, Fan T, Wang S. Highly enriched exosomal lncRNA OIP5-AS1 regulates osteosarcoma tumor angiogenesis and autophagy through miR-153 and ATG5. Am J Transl Res. 2021;13(5):4211–4223.
58. Nisato RE, Tille J-C, Jonczyk A, Goodman SL, Pepper MS. αvβ3 and αvβ5 integrin antagonists inhibit angiogenesis in vitro. Angiogenesis. 2003;6(2):105–119. doi:10.1023/B:AGEN.0000011801.98187.f2
59. Liu Y, Huang N, Liao S, et al. Current research progress in targeted anti‐angiogenesis therapy for osteosarcoma. Cell Prolif. 2021;54(9):e13102. doi:10.1111/cpr.13102
60. Toda Y, Kohashi K, Yamada Y, et al. PD-L1 and IDO1 expression and tumor-infiltrating lymphocytes in osteosarcoma patients: comparative study of primary and metastatic lesions. J Cancer Res Clin Oncol. 2020;146(10):2607–2620. doi:10.1007/s00432-020-03242-6
61. Zhou Y, Yang D, Yang Q, et al. Single-cell RNA landscape of intratumoral heterogeneity and immunosuppressive microenvironment in advanced osteosarcoma. Nat Commun. 2020;11(1):6322. doi:10.1038/s41467-020-20059-6
62. Yu Y, Zhang H, Ren T, et al. Development of a prognostic gene signature based on an immunogenomic infiltration analysis of osteosarcoma. J Cell Mol Med. 2020;24(19):11230–11242. doi:10.1111/jcmm.15687
63. Cersosimo F, Lonardi S, Bernardini G, et al. Tumor-associated macrophages in osteosarcoma: from mechanisms to therapy. Int J Mol Sci. 2020;21(15):5207. doi:10.3390/ijms21155207
64. Ahirwar DK, Charan M, Mishra S, et al. Slit2 inhibits breast cancer metastasis by activating M1-like phagocytic and anti-fibrotic macrophages. Cancer Res. 2021;81(20):5255–5267. doi:10.1158/0008-5472.CAN-20-3909
65. Szulc-Kielbik I, Kielbik M. Tumor-associated macrophages: reasons to be cheerful, reasons to be fearful. In: Klink M, Szulc-Kielbik I, editors. Interaction of Immune and Cancer Cells. Cham: Springer International Publishing; 2022:107–140. doi:10.1007/978-3-030-91311-3_4
66. Dumars C, Ngyuen JM, Gaultier A, et al. Dysregulation of macrophage polarization is associated with the metastatic process in osteosarcoma. Oncotarget. 2016;7(48):78343–78354. doi:10.18632/oncotarget.13055
67. Coffelt SB, Hughes R, Lewis CE. Tumor-associated macrophages: effectors of angiogenesis and tumor progression. Biochim Biophys Acta. 2009;1796(1):11–18. doi:10.1016/j.bbcan.2009.02.004
68. Ségaliny AI, Mohamadi A, Dizier B, et al. Interleukin-34 promotes tumor progression and metastatic process in osteosarcoma through induction of angiogenesis and macrophage recruitment. Int J Cancer. 2015;137(1):73–85. doi:10.1002/ijc.29376
69. Noy R, Pollard JW. Tumor-associated macrophages: from mechanisms to therapy. Immunity. 2014;41(1):49–61. doi:10.1016/j.immuni.2014.06.010
70. Han Q, Shi H, Liu F. CD163(+) M2-type tumor-associated macrophage support the suppression of tumor-infiltrating T cells in osteosarcoma. Int Immunopharmacol. 2016;34:101–106. doi:10.1016/j.intimp.2016.01.023
71. Schlecker E, Stojanovic A, Eisen C, et al. Tumor-infiltrating monocytic myeloid-derived suppressor cells mediate CCR5-dependent recruitment of regulatory T cells favoring tumor growth. J Immunol. 2012;189(12):5602–5611. doi:10.4049/jimmunol.1201018
72. Su Y, Zhou Y, Sun YJ, et al. Macrophage-derived CCL18 promotes osteosarcoma proliferation and migration by upregulating the expression of UCA1. J Mol Med. 2019;97(1):49–61. doi:10.1007/s00109-018-1711-0
73. Han Y, Guo W, Ren T, et al. Tumor-associated macrophages promote lung metastasis and induce epithelial-mesenchymal transition in osteosarcoma by activating the COX-2/STAT3 axis. Cancer Lett. 2019;440–441:116–125. doi:10.1016/j.canlet.2018.10.011
74. Shao XJ, Xiang SF, Chen YQ, et al. Inhibition of M2-like macrophages by all-trans retinoic acid prevents cancer initiation and stemness in osteosarcoma cells. Acta Pharmacol Sin. 2019;40(10):1343–1350. doi:10.1038/s41401-019-0262-4
75. Buddingh EP, Kuijjer ML, Duim RAJ, et al. Tumor-infiltrating macrophages are associated with metastasis suppression in high-grade osteosarcoma: a rationale for treatment with macrophage activating agents. Clin Cancer Res. 2011;17(8):2110–2119. doi:10.1158/1078-0432.CCR-10-2047
76. Huang Q, Liang X, Ren T, et al. The role of tumor-associated macrophages in osteosarcoma progression - therapeutic implications. Cell Oncol Dordr. 2021;44(3):525–539. doi:10.1007/s13402-021-00598-w
77. Pahl JHW, Kwappenberg KMC, Varypataki EM, et al. Macrophages inhibit human osteosarcoma cell growth after activation with the bacterial cell wall derivative liposomal muramyl tripeptide in combination with interferon-γ. J Exp Clin Cancer Res. 2014;33(1):27. doi:10.1186/1756-9966-33-27
78. Caillens V, Boisel E, Ouksel A, Nugue M, Evnouchidou I, Saveanu L. Integrin linked kinase and threonine tyrosine kinase modulate TCR signaling. Sci Rep. 2025;15(1):14392. doi:10.1038/s41598-025-99331-y
79. Orrapin S, Moonmuang S, Udomruk S, Yongpitakwattana P, Pruksakorn D, Chaiyawat P. Unlocking the tumor-immune microenvironment in osteosarcoma: insights into the immune landscape and mechanisms. Front Immunol. 2024;15:1394284. doi:10.3389/fimmu.2024.1394284
80. Zeng J, Peng Y, Wang D, Ayesha K, Chen S. The interaction between osteosarcoma and other cells in the bone microenvironment: from mechanism to clinical applications. Front Cell Dev Biol. 2023;11:1123065. doi:10.3389/fcell.2023.1123065
81. Wang J, Yuan R, Song W, Sun J, Liu D, Li Z. PD-1, PD-L1 (B7-H1) and tumor-site immune modulation therapy: the historical perspective. J Hematol Oncol. 2017;10(1):34. doi:10.1186/s13045-017-0403-5
82. Liu W, Hu H, Shao Z, et al. Characterizing the tumor microenvironment at the single-cell level reveals a novel immune evasion mechanism in osteosarcoma. Bone Res. 2023;11(1):4. doi:10.1038/s41413-022-00237-6
83. Casanova JM, Almeida JS, Reith JD, et al. Tumor-infiltrating lymphocytes and cancer markers in osteosarcoma: influence on patient survival. Cancers. 2021;13(23):6075. doi:10.3390/cancers13236075
84. Richert I, Berchard P, Abbes L, et al. A TLR4 agonist induces osteosarcoma regression by inducing an antitumor immune response and reprogramming M2 macrophages to M1 macrophages. Cancers. 2023;15(18):4635. doi:10.3390/cancers15184635
85. Liu Y, Feng W, Dai Y, et al. Single-cell transcriptomics reveals the complexity of the tumor microenvironment of treatment-naive osteosarcoma. Front Oncol. 2021;11:709210. doi:10.3389/fonc.2021.709210
86. Sivori S, Pende D, Quatrini L, et al. NK cells and ILCs in tumor immunotherapy. Mol Aspects Med. 2021;80:100870. doi:10.1016/j.mam.2020.100870
87. Ramírez-Labrada A, Pesini C, Santiago L, et al. All about (NK Cell-Mediated) death in two acts and an unexpected encore: initiation, execution and activation of adaptive immunity. Front Immunol. 2022;13:896228. doi:10.3389/fimmu.2022.896228
88. Rademacher MJ, Cruz A, Faber M, et al. Sarcoma IL-12 overexpression facilitates NK cell immunomodulation. Sci Rep. 2021;11(1):8321. doi:10.1038/s41598-021-87700-2
89. Ogiwara Y, Nakagawa M, Nakatani F, Uemura Y, Zhang R, Kudo-Saito C. Blocking FSTL1 boosts NK immunity in treatment of osteosarcoma. Cancer Lett. 2022;537:215690. doi:10.1016/j.canlet.2022.215690
90. Marciscano AE, Anandasabapathy N. The role of dendritic cells in cancer and anti-tumor immunity. Semin Immunol. 2021;52:101481.
91. Wculek SK, Cueto FJ, Mujal AM, Melero I, Krummel MF, Sancho D. Dendritic cells in cancer immunology and immunotherapy. Nat Rev Immunol. 2020;20(1):7–24.
92. Guilliams M, Dutertre CA, Scott CL, et al. Unsupervised high-dimensional analysis aligns dendritic cells across tissues and species. Immunity. 2016;45(3):669–684. doi:10.1016/j.immuni.2016.08.015
93. Kansara M, Thomson K, Pang P, et al. Infiltrating myeloid cells drive osteosarcoma progression via GRM4 regulation of IL23. Cancer Discovery. 2019;9(11):1511–1519. doi:10.1158/2159-8290.CD-19-0154
94. Rodríguez-Fernández JL, Criado-García O. The chemokine receptor CCR7 uses distinct signaling modules with biased functionality to regulate dendritic cells. Front Immunol. 2020;11:528. doi:10.3389/fimmu.2020.00528
95. Niu Y, Du T, Liu Y. Biomechanical characteristics and analysis approaches of bone and bone substitute materials. J Funct Biomater. 2023;14(4):212.
96. Mühlbauer RC, Russell RG, Williams DA, Fleisch H. The effects of diphosphonates, polyphosphates, and calcitonin on “immobilisation osteoporosis” in rats. Eur J Clin Invest. 1971;1(5):336–344. doi:10.1111/j.1365-2362.1971.tb00640.x
97. Russell RGG. Bisphosphonates: mode of action and pharmacology. Pediatrics. 2007;119 Suppl 2(Supplement_2):S150–162. doi:10.1542/peds.2006-2023H
98. Abdalla AME, Xiao L, Ullah MW, Yu M, Ouyang C, Yang G. Current challenges of cancer anti-angiogenic therapy and the promise of nanotherapeutics. Theranostics. 2018;8(2):533–548. doi:10.7150/thno.21674
99. Wang Y, Wang C, Xia M, et al. Engineering small-molecule and protein drugs for targeting bone tumors. Mol Ther. 2024;32(5):1219–1237. doi:10.1016/j.ymthe.2024.03.001
100. Xue L, Gong N, Shepherd SJ, et al. Rational design of bisphosphonate lipid-like materials for mRNA delivery to the bone microenvironment. J Am Chem Soc. 2022;144(22):9926–9937. doi:10.1021/jacs.2c02706
101. Segal E, Pan H, Benayoun L, et al. Enhanced anti-tumor activity and safety profile of targeted nano-scaled HPMA copolymer-alendronate-TNP-470 conjugate in the treatment of bone malignances. Biomaterials. 2011;32(19):4450–4463. doi:10.1016/j.biomaterials.2011.02.059
102. Nguyen TDT, Pitchaimani A, Aryal S. Engineered nanomedicine with alendronic acid corona improves targeting to osteosarcoma. Sci Rep. 2016;6(1):36707. doi:10.1038/srep36707
103. Fujisawa R, Wada Y, Nodasaka Y, Kuboki Y. Acidic amino acid-rich sequences as binding sites of osteonectin to hydroxyapatite crystals. Biochim Biophys Acta. 1996;1292(1):53–60. doi:10.1016/0167-4838(95)00190-5
104. Zhao Z, Chen C, Xie C, Zhao Y. Design, synthesis and evaluation of liposomes modified with dendritic aspartic acid for bone-specific targeting. Chem Phys Lipids. 2020;226:104832. doi:10.1016/j.chemphyslip.2019.104832
105. Sekido T, Sakura N, Higashi Y, et al. Novel drug delivery system to bone using acidic oligopeptide: pharmacokinetic characteristics and pharmacological potential. J Drug Target. 2001;9(2):111–121. doi:10.3109/10611860108997922
106. Wu Y, Sun B, Tang Y, et al. Bone targeted nano-drug and nano-delivery. Bone Res. 2024;12(1):51. doi:10.1038/s41413-024-00356-2
107. Wang Y, Yang J, Liu H, et al. Osteotropic peptide-mediated bone targeting for photothermal treatment of bone tumors. Biomaterials. 2017;114:97–105. doi:10.1016/j.biomaterials.2016.11.010
108. Low SA, Yang J, Kopeček J. Bone-targeted acid-sensitive doxorubicin conjugate micelles as potential osteosarcoma therapeutics. Bioconjug Chem. 2014;25(11):2012–2020. doi:10.1021/bc500392x
109. Wang H, Liu J, Tao S, et al. Tetracycline-grafted PLGA nanoparticles as bone-targeting drug delivery system. Int J Nanomed. 2015;10:5671–5685. doi:10.2147/IJN.S88798
110. Zhou Z, Fan T, Yan Y, et al. One stone with two birds: phytic acid-capped platinum nanoparticles for targeted combination therapy of bone tumors. Biomaterials. 2019;194:130–138. doi:10.1016/j.biomaterials.2018.12.024
111. Yin F, Fan Y, Xu L, et al. Macrophages loaded with dendrimer-entrapped gold nanoparticles as a theranostic platform for CT imaging-guided combinational therapy of orthotopic osteosarcoma. Chem Eng J. 2021;417:129273. doi:10.1016/j.cej.2021.129273
112. Huo S, Liu S, Liu Q, et al. Copper–Zinc‐doped bilayer bioactive glasses loaded hydrogel with spatiotemporal immunomodulation supports MRSA‐infected wound healing. Adv Sci. 2023;11(5):2302674. doi:10.1002/advs.202302674
113. Huang X, Wang L, Guo H, Zhang W. Macrophage membrane-coated nanovesicles for dual-targeted drug delivery to inhibit tumor and induce macrophage polarization. Bioact Mater. 2022;23:69–79. doi:10.1016/j.bioactmat.2022.09.027
114. Fu L, Zhang W, Zhou X, Fu J, He C. Tumor cell membrane-camouflaged responsive nanoparticles enable MRI-guided immuno-chemodynamic therapy of orthotopic osteosarcoma. Bioact Mater. 2022;17:221–233. doi:10.1016/j.bioactmat.2022.01.035
115. Chen X, Gao M, An S, et al. Enhancing adoptive T cell therapy for solid tumor with cell-surface anchored immune checkpoint inhibitor nanogels. Nanomed Nanotechnol Biol Med. 2022;45:102591. doi:10.1016/j.nano.2022.102591
116. Kiru L, Zlitni A, Tousley AM, et al. In vivo imaging of nanoparticle-labeled CAR T cells. Proc Natl Acad Sci U S A. 2022;119(6):e2102363119. doi:10.1073/pnas.2102363119
117. Liu K, Liao Y, Zhou Z, et al. Photothermal-triggered immunogenic nanotherapeutics for optimizing osteosarcoma therapy by synergizing innate and adaptive immunity. Biomaterials. 2022;282:121383. doi:10.1016/j.biomaterials.2022.121383
118. Dong H, Wen ZF, Chen L, et al. Polyethyleneimine modification of aluminum hydroxide nanoparticle enhances antigen transportation and cross-presentation of dendritic cells. Int J Nanomed. 2018;13:3353–3365. doi:10.2147/IJN.S164097
119. Zhao H, Xu J, Li Y, et al. Nanoscale coordination polymer based nanovaccine for tumor immunotherapy. ACS Nano. 2019;13(11):13127–13135. doi:10.1021/acsnano.9b05974
120. Kleinerman ES, Murray JL, Snyder JS, Cunningham JE, Fidler IJ. Activation of tumoricidal properties in monocytes from cancer patients following intravenous administration of liposomes containing muramyl tripeptide phosphatidylethanolamine. Cancer Res. 1989;49(16):4665–4670.
121. Múdry P, Kýr M, Rohleder O, et al. Improved osteosarcoma survival with addition of mifamurtide to conventional chemotherapy – observational prospective single institution analysis. J Bone Oncol. 2021;28:100362. doi:10.1016/j.jbo.2021.100362
122. Štorkánová H, Oreská S, Špiritović M, et al. Plasma Hsp90 levels in patients with systemic sclerosis and relation to lung and skin involvement: a cross-sectional and longitudinal study. Sci Rep. 2021;11(1):1. doi:10.1038/s41598-020-79139-8
123. Kim HS, Lee EH, Ko SR, Choi KJ, Park JH, Im DS. Effects of ginsenosides Rg3 and Rh2 on the proliferation of prostate cancer cells. Arch Pharm Res. 2004;27(4):429–435. doi:10.1007/BF02980085
124. Trebak M, Kinet JP. Calcium signalling in T cells. Nat Rev Immunol. 2019;19(3):154–169. doi:10.1038/s41577-018-0110-7
125. Jakobs J, Bertram J, Rink L. Ca2+ signals are essential for T-cell proliferation, while Zn2+ signals are necessary for T helper cell 1 differentiation. Cell Death Discov. 2024;10(1):336. doi:10.1038/s41420-024-02104-1
126. Qin S, Hu Y, Luo H, Chu W, Deng R, Ma J. Metal ions and nanomaterials for targeted bone cancer immunotherapy. Front Immunol. 2025;16:1513834. doi:10.3389/fimmu.2025.1513834
127. Maus MV, Grupp SA, Porter DL, June CH. Antibody-modified T cells: cARs take the front seat for hematologic malignancies. Blood. 2014;123(17):2625–2635. doi:10.1182/blood-2013-11-492231
128. Zuo YH, Zhao XP, Fan XX. Nanotechnology-based chimeric antigen receptor T-cell therapy in treating solid tumor. Pharmacol Res. 2022;184:106454. doi:10.1016/j.phrs.2022.106454
129. Li H, Wang Z, Ogunnaike EA, et al. Scattered seeding of CAR T cells in solid tumors augments anticancer efficacy. Natl Sci Rev. 2021;9(3):nwab172. doi:10.1093/nsr/nwab172
130. Sharma P, Allison JP. Dissecting the mechanisms of immune checkpoint therapy. Nat Rev Immunol. 2020;20(2):75–76. doi:10.1038/s41577-020-0275-8
131. Bryant CE, Sutherland S, Kong B, Papadimitrious MS, Fromm PD, Hart DNJ. Dendritic cells as cancer therapeutics. Semin Cell Dev Biol. 2019;86:77–88. doi:10.1016/j.semcdb.2018.02.015
132. Gilboa E. DC-based cancer vaccines. J Clin Invest. 2007;117(5):1195–1203. doi:10.1172/JCI31205
133. Zhu G, Lynn GM, Jacobson O, et al. Albumin/vaccine nanocomplexes that assemble in vivo for combination cancer immunotherapy. Nat Commun. 2017;8(1):1954. doi:10.1038/s41467-017-02191-y
134. Melero I, Gaudernack G, Gerritsen W, et al. Therapeutic vaccines for cancer: an overview of clinical trials. Nat Rev Clin Oncol. 2014;11(9):509–524. doi:10.1038/nrclinonc.2014.111
135. Musetti S, Huang L. Nanoparticle-mediated remodeling of the tumor microenvironment to enhance immunotherapy. ACS Nano. 2018;12(12):11740–11755. doi:10.1021/acsnano.8b05893
136. Yang R, Xu J, Xu L, et al. Cancer cell membrane-coated adjuvant nanoparticles with mannose modification for effective anticancer vaccination. ACS Nano. 2018;12(6):5121–5129. doi:10.1021/acsnano.7b09041
137. Paulis LE, Mandal S, Kreutz M, Figdor CG. Dendritic cell-based nanovaccines for cancer immunotherapy. Curr Opin Immunol. 2013;25(3):389–395. doi:10.1016/j.coi.2013.03.001
138. Asano J, Tada H, Onai N, et al. Nucleotide oligomerization binding domain-like receptor signaling enhances dendritic cell-mediated cross-priming in vivo. J Immunol. 2009;184(2):736–745. doi:10.4049/jimmunol.0900726
139. Chen GY, Shaw MH, Redondo G, Núñez G. The innate immune receptor Nod1 protects the intestine from inflammation-induced tumorigenesis. Available from: https://aacrjournals.org/cancerres/article/68/24/10060/541834/The-Innate-Immune-Receptor-Nod1-Protects-the.
140. Hao Z, Fan W, Hao J, et al. Efficient delivery of micro RNA to bone-metastatic prostate tumors by using aptamer-conjugated atelocollagen in vitro and in vivo. Drug Deliv. 2016;23(3):874–881. doi:10.3109/10717544.2014.920059
141. Ishikawa T, Ichikawa Y, Shimizu D, et al. The role of HER-2 in breast cancer. J Surg Sci. 2014;2(1):4–9.
142. Kievit FM, Stephen ZR, Veiseh O, et al. Targeting of primary breast cancers and metastases in a transgenic mouse model using rationally designed multifunctional SPIONs. ACS Nano. 2012;6(3):2591–2601. doi:10.1021/nn205070h
143. Fang Z, Sun Y, Xiao H, et al. Targeted osteosarcoma chemotherapy using RGD peptide-installed doxorubicin-loaded biodegradable polymeric micelle. Biomed Pharmacother. 2017;85:160–168. doi:10.1016/j.biopha.2016.11.132
144. Huang X, Wu W, Jing D, et al. Engineered exosome as targeted lncRNA MEG3 delivery vehicles for osteosarcoma therapy. J Control Release. 2022;343:107–117. doi:10.1016/j.jconrel.2022.01.026
145. Hu H, Deng X, Song Q, et al. Mitochondria-targeted accumulation of oxygen-irrelevant free radicals for enhanced synergistic low-temperature photothermal and thermodynamic therapy. J Nanobiotechnology. 2021;19(1):390. doi:10.1186/s12951-021-01142-6
146. Wu X, Zhang X, Feng W, et al. A targeted erythrocyte membrane-encapsulated drug-delivery system with anti-osteosarcoma and anti-osteolytic effects. ACS Appl Mater Interfaces. 2021;13(24):27920–27933. doi:10.1021/acsami.1c06059
147. Zhang Y, Wang F, Li M, et al. Self‐stabilized hyaluronate nanogel for intracellular codelivery of doxorubicin and cisplatin to osteosarcoma. Adv Sci. 2018;5(5):1700821. doi:10.1002/advs.201700821
148. Xu Y, Qi J, Sun W, Zhong W, Wu H. Therapeutic effects of zoledronic acid-loaded hyaluronic Acid/Polyethylene Glycol/Nano-hydroxyapatite nanoparticles on osteosarcoma. Front Bioeng Biotechnol. 2022;10:897641. doi:10.3389/fbioe.2022.897641
149. Haghiralsadat F, Amoabediny G, Naderinezhad S, et al. EphA2 targeted doxorubicin-nanoliposomes for osteosarcoma treatment. Pharm Res. 2017;34(12):2891–2900. doi:10.1007/s11095-017-2272-6
150. Qiu R, Sun D, Bai Y, Li J, Wang L. Application of tumor-targeting peptide-decorated polypeptide nanoparticles with doxorubicin to treat osteosarcoma. Drug Deliv. 2020;27(1):1704–1717. doi:10.1080/10717544.2020.1856221
151. Li S, Zhang T, Xu W, et al. Sarcoma-targeting peptide-decorated polypeptide nanogel intracellularly delivers shikonin for upregulated osteosarcoma necroptosis and diminished pulmonary metastasis. Theranostics. 2018;8(5):1361–1375. doi:10.7150/thno.18299
152. Desgrosellier JS, Cheresh DA. Integrins in cancer: biological implications and therapeutic opportunities. Nat Rev Cancer. 2010;10(1):9–22. doi:10.1038/nrc2748
153. Martinez-Velez N, Xipell E, Jauregui P, et al. The oncolytic adenovirus Δ24-RGD in combination with cisplatin exerts a potent anti-osteosarcoma activity. J Bone Miner Res. 2014;29(10):2287–2296. doi:10.1002/jbmr.2253
154. Witlox AM, van Beusechem VW, Molenaar B, et al. Conditionally replicative adenovirus with tropism expanded towards integrins inhibits osteosarcoma tumor growth in vitro and in vivo. Clin Cancer Res. 2004;10(1):61–67. doi:10.1158/1078-0432.CCR-0609-03
155. Zheng B, Zhou C, Qu G, et al. VEGFR2 promotes metastasis and PD-L2 expression of human osteosarcoma cells by activating the STAT3 and RhoA-ROCK-LIMK2 pathways. Front Oncol. 2020;10:543562. doi:10.3389/fonc.2020.543562
156. Assi T, Watson S, Samra B, et al. Targeting the VEGF pathway in osteosarcoma. Cells. 2021;10(5):1240. doi:10.3390/cells10051240
157. Liu Y, Wu Y, Gu S, et al. Prognostic role of CD44 expression in osteosarcoma: evidence from six studies. Diagn Pathol. 2014;9(1):140. doi:10.1186/1746-1596-9-140
158. Ji H, Kong L, Wang Y, et al. CD44 expression is correlated with osteosarcoma cell progression and immune infiltration and affects the Wnt/β-catenin signaling pathway. J Bone Oncol. 2023;41:100487. doi:10.1016/j.jbo.2023.100487
159. Xiao Z, Wan J, Nur AA, et al. Targeting CD44 by CRISPR-Cas9 in multi-drug resistant osteosarcoma cells. Cell Physiol Biochem. 2018;51(4):1879–1893. doi:10.1159/000495714
160. Majerova P, Hanes J, Olesova D, Sinsky J, Pilipcinec E, Kovac A. Novel blood–brain barrier shuttle peptides discovered through the phage display method. Molecules. 2020;25(4):874. doi:10.3390/molecules25040874
161. Sun B, Li R, Ji N, et al. Brain-targeting drug delivery systems: the state of the art in treatment of glioblastoma. Mater Today Bio. 2025;30:101443. doi:10.1016/j.mtbio.2025.101443
162. Li A, Wang S, Nie J, et al. USP3 promotes osteosarcoma progression via deubiquitinating EPHA2 and activating the PI3K/AKT signaling pathway. Cell Death Dis. 2024;15(3):235. doi:10.1038/s41419-024-06624-7
163. Patil MA, Upadhyay AK, Hernandez-Lagunas L, et al. Targeted delivery of YSA-functionalized and non-functionalized polymeric nanoparticles to injured pulmonary vasculature. Artif Cells Nanomed Biotechnol. 2018;46(SUP3):S1059–66. doi:10.1080/21691401.2018.1528984
164. Liu Z, Tao Z, Zhang Q, et al. YSA-conjugated mesoporous silica nanoparticles effectively target EphA2-overexpressing breast cancer cells. Cancer Chemother Pharmacol. 2018;81(4):687–695. doi:10.1007/s00280-018-3535-6
165. Fasanya HO, Dopico PJ, Yeager Z, Fan ZH, Siemann DW. Using a combination of gangliosides and cell surface vimentin as surface biomarkers for isolating osteosarcoma cells in microfluidic devices. J Bone Oncol. 2021;28:100357. doi:10.1016/j.jbo.2021.100357
166. Satelli A, Mitra A, Cutrera JJ, et al. Universal marker and detection tool for human sarcoma circulating tumor cells. Cancer Res. 2014;74(6):1645–1650. doi:10.1158/0008-5472.CAN-13-1739
167. Yuan Y, Diao S, Ni X, et al. Peptide-based semiconducting polymer nanoparticles for osteosarcoma-targeted NIR-II fluorescence/NIR-I photoacoustic dual-model imaging and photothermal/photodynamic therapies. J Nanobiotechnology. 2022;20(1):44. doi:10.1186/s12951-022-01249-4
168. Zhou H, Yi W, Li A, et al. Specific small-molecule NIR-II fluorescence imaging of osteosarcoma and lung metastasis. Adv Healthc Mater. 2020;9(1):1901224. doi:10.1002/adhm.201901224
169. Ma Z, Qin H, Chen H, et al. Phage display-derived oligopeptide-functionalized probes for in vivo specific photoacoustic imaging of osteosarcoma. Nanomedicine. 2017;13(1):111–121. doi:10.1016/j.nano.2016.09.002
170. Liu S, Ou H, Li Y, et al. Planar and twisted molecular structure leads to the high brightness of semiconducting polymer nanoparticles for NIR-IIa fluorescence imaging. J Am Chem Soc. 2020;142(35):15146–15156. doi:10.1021/jacs.0c07193
171. Zhang W, Sun X, Huang T, et al. 1300 nm absorption two-acceptor semiconducting polymer nanoparticles for NIR-II photoacoustic imaging system guided NIR-II photothermal therapy. Chem Commun. 2019;55(64):9487–9490. doi:10.1039/C9CC04196F
172. Zhou W, Chen Y, Zhang Y, et al. Iodine-rich semiconducting polymer nanoparticles for CT/Fluorescence dual-modal imaging-guided enhanced photodynamic therapy. Small. 2020;16(5):e1905641. doi:10.1002/smll.201905641
173. Kovach AK, Gambino JM, Nguyen V, et al. Prospective preliminary in vitro investigation of a magnetic iron oxide nanoparticle conjugated with ligand CD80 and VEGF antibody as a targeted drug delivery system for the induction of cell death in rodent osteosarcoma cells. Biores Open Access. 2016;5(1):299–307. doi:10.1089/biores.2016.0020
174. Li Y, Wu Y, Huang L, et al. Sigma receptor-mediated targeted delivery of anti-angiogenic multifunctional nanodrugs for combination tumor therapy. J Control Release. 2016;228:107–119. doi:10.1016/j.jconrel.2016.02.044
175. Zhao TT, Zhou TJ, Zhang C, et al. Hypoxia inhibitor combined with chemotherapeutic agents for antitumor and antimetastatic efficacy against osteosarcoma. Mol Pharm. 2023;20(5):2612–2623. doi:10.1021/acs.molpharmaceut.3c00068
176. Zhang W, Li M, Zhao Z, et al. Tetrahedral framework nucleic acid-loaded retinoic acid promotes osteosarcoma stem cell clearance. ACS Appl Mater Interfaces. 2024;16(43):58452–58463. doi:10.1021/acsami.4c14440
177. Lu SL, Wang YH, Liu GF, et al. Graphene oxide nanoparticle–loaded ginsenoside Rg3 improves photodynamic therapy in inhibiting malignant progression and stemness of osteosarcoma. Front Mol Biosci. 2021;8:663089. doi:10.3389/fmolb.2021.663089
178. Gui K, Zhang X, Chen F, et al. Lipid-polymer nanoparticles with CD133 aptamers for targeted delivery of all-trans retinoic acid to osteosarcoma initiating cells. Biomed Pharmacother. 2019;111:751–764. doi:10.1016/j.biopha.2018.11.118
179. Ni M, Xiong M, Zhang X, et al. Poly(lactic-co-glycolic acid) nanoparticles conjugated with CD133 aptamers for targeted salinomycin delivery to CD133+ osteosarcoma cancer stem cells. Int J Nanomed. 2015;10:2537–2554. doi:10.2147/IJN.S78498
180. Tian J, Gu Y, Li Y, Liu T. CD271 antibody-functionalized HGNs for targeted photothermal therapy of osteosarcoma stem cells. Nanotechnology. 2020;31(30):305707. doi:10.1088/1361-6528/ab8593
181. Di Pompo G, Cortini M, Palomba R, et al. Curcumin-loaded nanoparticles impair the pro-tumor activity of acid-stressed MSC in an in vitro model of osteosarcoma. Int J Mol Sci. 2021;22(11):5760. doi:10.3390/ijms22115760
182. Kuşoğlu A, Biray Avcı Ç. Cancer stem cells: a brief review of the current status. Gene. 2019;681:80–85. doi:10.1016/j.gene.2018.09.052
183. Chen F, Zeng Y, Qi X, et al. Targeted salinomycin delivery with EGFR and CD133 aptamers based dual-ligand lipid-polymer nanoparticles to both osteosarcoma cells and cancer stem cells. Nanomed Nanotechnol Biol Med. 2018;14(7):2115–2127. doi:10.1016/j.nano.2018.05.015
184. Chu X, Tian W, Ning J, et al. Cancer stem cells: advances in knowledge and implications for cancer therapy. Signal Transduct Target Ther. 2024;9(1):170. doi:10.1038/s41392-024-01851-y
185. Liu X, Shan W, Li T, et al. Cellular retinol binding protein-1 inhibits cancer stemness via upregulating WIF1 to suppress Wnt/β-catenin pathway in hepatocellular carcinoma. BMC Cancer. 2021;21(1):1224. doi:10.1186/s12885-021-08967-2
186. Bashur L, Zhou G. CANCER STEM CELLS IN OSTEOSARCOMA. Case Orthop J. 2013;10(1):38–42.
187. Tirino V, Desiderio V, Paino F, et al. Human primary bone sarcomas contain CD133+ cancer stem cells displaying high tumorigenicity in vivo. FASEB J. 2011;25(6):2022–2030. doi:10.1096/fj.10-179036
188. Tirino V, Desiderio V, d’Aquino R, et al. Detection and characterization of CD133+ cancer stem cells in human solid tumours. PLoS One. 2008;3(10):e3469. doi:10.1371/journal.pone.0003469
189. Boiko AD, Razorenova OV, van de Rijn M, et al. Human melanoma-initiating cells express neural crest nerve growth factor receptor CD271. Nature. 2010;466(7302):133–137. doi:10.1038/nature09161
190. Tian J, Li X, Si M, Liu T, Li J. CD271+ osteosarcoma cells display stem-like properties. PLoS One. 2014;9(6):e98549. doi:10.1371/journal.pone.0098549
191. Ponte AL, Marais E, Gallay N, et al. The in vitro migration capacity of human bone marrow mesenchymal stem cells: comparison of chemokine and growth factor chemotactic activities. Stem Cells. 2007;25(7):1737–1745. doi:10.1634/stemcells.2007-0054
192. Cortini M, Avnet S, Baldini N. Mesenchymal stroma: role in osteosarcoma progression. Cancer Lett. 2017;405:90–99. doi:10.1016/j.canlet.2017.07.024
193. Massa A, Perut F, Chano T, et al. The effect of extracellular acidosis on the behaviour of mesenchymal stem cells in vitro. Eur Cell Mater. 2017;33:252–267. doi:10.22203/eCM.v033a19
194. Peppicelli S, Bianchini F, Toti A, Laurenzana A, Fibbi G, Calorini L. Extracellular acidity strengthens mesenchymal stem cells to promote melanoma progression. Cell Cycle. 2015;14(19):3088–3100. doi:10.1080/15384101.2015.1078032
195. Avnet S, Di Pompo G, Chano T, et al. Cancer-associated mesenchymal stroma fosters the stemness of osteosarcoma cells in response to intratumoral acidosis via NF-κB activation. Int J Cancer. 2017;140(6):1331–1345. doi:10.1002/ijc.30540
196. Karunaweera N, Raju R, Gyengesi E, Münch G. Plant polyphenols as inhibitors of NF-κB induced cytokine production—a potential anti-inflammatory treatment for Alzheimer’s disease? Front Mol Neurosci. 2015;8:24. doi:10.3389/fnmol.2015.00024
197. Chang Z, Xing J, Yu X. Curcumin induces osteosarcoma MG63 cells apoptosis via ROS/Cyto-C/Caspase-3 pathway. Tumour Biol. 2014;35(1):753–758. doi:10.1007/s13277-013-1102-7
198. Kidd S, Spaeth E, Dembinski JL, et al. Direct evidence of mesenchymal stem cell tropism for tumor and wounding microenvironments using in vivo bioluminescence imaging. Stem Cells. 2009;27(10):2614–2623. doi:10.1002/stem.187
199. Gao Z, Zhang L, Hu J, Sun Y. Mesenchymal stem cells: a potential targeted-delivery vehicle for anti-cancer drug loaded nanoparticles. Nanomed Nanotechnol Biol Med. 2013;9(2):174–184. doi:10.1016/j.nano.2012.06.003
200. Duchi S, Sotgiu G, Lucarelli E, et al. Mesenchymal stem cells as delivery vehicle of porphyrin loaded nanoparticles: effective photoinduced in vitro killing of osteosarcoma. J Control Release. 2013;168(2):225–237. doi:10.1016/j.jconrel.2013.03.012
201. Lenna S, Bellotti C, Duchi S, et al. Mesenchymal stromal cells mediated delivery of photoactive nanoparticles inhibits osteosarcoma growth in vitro and in a murine in vivo ectopic model. J Exp Clin Cancer Res. 2020;39(1):40. doi:10.1186/s13046-020-01548-4
202. Wei H, Chen F, Chen J, et al. Mesenchymal stem cell derived exosomes as nanodrug carrier of doxorubicin for targeted osteosarcoma therapy via SDF1-CXCR4 axis. Int J Nanomed. 2022;17:3483–3495. doi:10.2147/IJN.S372851
203. Wu K, Yu B, Li D, Tian Y, Liu Y, Jiang J. Recent advances in nanoplatforms for the treatment of osteosarcoma. Front Oncol. 2022;12:805978. doi:10.3389/fonc.2022.805978
204. Yu W, Liu R, Zhou Y, Gao H. Size-tunable strategies for a tumor targeted drug delivery system. ACS Cent Sci. 2020;6(2):100–116. doi:10.1021/acscentsci.9b01139
205. Wilhelm S, Tavares AJ, Dai Q, et al. Analysis of nanoparticle delivery to tumours. Nat Rev Mater. 2016;1(5):1–12. doi:10.1038/natrevmats.2016.14
206. Thorp EB, Boada C, Jarbath C, Luo X. Nanoparticle platforms for antigen-specific immune tolerance. Front Immunol. 2020;11:945. doi:10.3389/fimmu.2020.00945
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
GOx-Functionalized Platelet Membranes-Camouflaging Nanoreactors for Enhanced Multimodal Tumor Treatment
Du Y, Wang S, Luan J, Zhang M, Chen B, Shen Y
International Journal of Nanomedicine 2022, 17:2979-2993
Published Date: 7 July 2022
Camptothecin-Loaded and Manganese Dioxide-Coated Polydopamine Nanomedicine Used for Magnetic Resonance Imaging Diagnosis and Chemo-Photothermal Therapy for Lung Cancer

Su M, Chen Y, Jia L, Zhang Z
International Journal of Nanomedicine 2022, 17:6687-6705
Published Date: 28 December 2022
Drug Delivery System Targeting Cancer-Associated Fibroblast for Improving Immunotherapy
Zhang Z, Wang R, Chen L
International Journal of Nanomedicine 2025, 20:483-503
Published Date: 11 January 2025
Nanofibers in Glioma Therapy: Advances, Applications, and Overcoming Challenges

Zhou S, Zhang M, Wang J, Chen X, Xu Z, Yan Y, Li Y
International Journal of Nanomedicine 2025, 20:4677-4703
Published Date: 14 April 2025
Triphenylphosphine-Based Mitochondrial Targeting Nanocarriers: Advancing Cancer Therapy

Ali MS, Gowda BHJ, Shukla R, Kesharwani P
Clinical Pharmacology: Advances and Applications 2025, 17:119-141
Published Date: 10 June 2025