Back to Journals » International Journal of Nanomedicine » Volume 21
Nanomaterial-Induced Bacterial Ferroptosis-Like Death: A Novel Strategy to Counteract Antimicrobial Resistance
Authors Wu X, Wang Y, Zhang T, Li Z, Du Y ![]() , Gao J
, Gao J ![]()
Received 2 October 2025
Accepted for publication 20 December 2025
Published 9 January 2026 Volume 2026:21 571655
DOI https://doi.org/10.2147/IJN.S571655
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Eng San Thian
Xiaoyue Wu,1– 3,* Yanying Wang,1,* Tinglin Zhang,2,4 Zhaoshen Li,1,2,4,5 Yiqi Du,1,2,4,5 Jie Gao1,2,4,5
1Department of Gastroenterology, Shanghai Changhai Hospital, Naval Medical University, Shanghai, 200433, People’s Republic of China; 2Changhai Clinical Research Unit, Shanghai Changhai Hospital, Naval Medical University, Shanghai, 200433, People’s Republic of China; 3The 925th Hospital of PLA Joint Logistics Support Force CN, Guiyang, 550000, People’s Republic of China; 4Shanghai Key Laboratory of Nautical Medicine and Translation of Drugs and Medical Devices, Shanghai, 200433, People’s Republic of China; 5National Key Laboratory of Immunology and Inflammation, Naval Medical University, Shanghai, 200433, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jie Gao, Changhai Clinical Research Unit, Shanghai Changhai Hospital, Naval Medical University, Shanghai, 200433, People’s Republic of China, Tel/Fax +86-21-31166666, Email [email protected] Yiqi Du, Department of Gastroenterology, Shanghai Changhai Hospital, Naval Medical University, Shanghai, 200433, People’s Republic of China, Tel/Fax +86-021-31161344, Email [email protected]
Abstract: The escalating crisis of antimicrobial resistance (AMR) poses a profound threat to global public health, necessitating the development of alternative antibacterial strategies. Ferroptosis, an iron-dependent form of regulated cell death driven by lipid peroxidation, has recently emerged as a promising strategy for combating drug-resistant bacteria. In this review, we systematically summarize how nanomaterials precisely regulate bacterial iron metabolism through chemical kinetics, photodynamics, sonodynamics, and nanoenzyme catalytic pathways, which promote iron storage and reactive oxygen species (ROS) generation, destroy the antioxidant defense system, and eventually lead to ferroptosis-like lipid peroxidation and membrane damage. In addition, this study focused on the synergistic activation of the host immune response by nanomaterials through the regulation of macrophage polarization and the cytokine network, thereby enhancing bacterial clearance and tissue repair. As an emerging nonantibiotic antimicrobial method, nanomaterial-mediated bacterial ferroptosis provides new insights and approaches to address the challenge of drug-resistant infections.
Keywords: ferroptosis-like death, nanomaterials, antibacterial strategy, drug resistance, chemotherapeutic therapy, immune regulation, synergistic therapy
Introduction
Global antimicrobial use has increased by more than 46% since the beginning of the 21st century, and the overuse of antibiotics in humans and animals and inadequate infection prevention and control measures have led to a significant increase in antimicrobial resistance (AMR), which has become one of the most devastating crises in global public health.1–3 At present, the annual mortality rate of AMR is approximately 7.7 million worldwide each year; AMR has become one of the leading causes of death worldwide, especially for vulnerable groups such as newborns, elderly individuals and cancer patients.4–6 With the increasing severity of global aging, a decline in immune function increases the risk of infection with various chronic diseases. Therefore, an increase in drug-resistant pathogen infection also significantly increases the risk of death and medical expenditure related to chronic diseases in elderly individuals.7–9 Nearly half of the pathogens causing surgical site infections in cancer patients are resistant to standard prophylactic antibiotics, resulting in drug failure and increased mortality.10–12 Traditional antibiotics kill bacteria by inhibiting specific targets, such as those involved in cell wall synthesis, and interfering with protein synthesis. However, bacteria are facing the increasingly serious problem of bacterial resistance, and their effects on dense biofilms are generally poor.13,14 In summary, traditional antibacterial strategies can no longer meet current antibacterial needs, and the development of new antibacterial methods is particularly urgent.
Ferroptosis is a new type of programmed cell death that has been gradually discovered and clearly defined since the beginning of the 21st century. Morphologically, ferroptosis is characterized by a reduction in mitochondrial volume, an increase in membrane density, and a reduction in or disappearance of cristae, but the nucleus is normal in size, and there is no chromatin condensation or apoptotic body formation.15–17 These features are quite different from traditional modes of cell death, such as apoptosis, necrosis, and autophagy.18,19 The core mechanism of ferroptosis involves the production of reactive oxygen species (ROS) that are dependent on iron ions to promote lipid peroxidation, especially the oxidation of polyunsaturated fatty acids (PUFAs), leading to cell membrane damage and cell death.20–22 Ferroptosis has been confirmed to be widely involved in the occurrence and development of tumors, neurodegenerative diseases, ischemia‒reperfusion injury and other pathological processes.23,24 In recent years, bacteria have been shown to have a similar ferroptosis-like phenotype, which shares key features with mammalian cell ferroptosis in the core mechanism.25 All of these processes include iron accumulation, lipid peroxidation, glutathione (GSH) depletion and antioxidant enzyme activity reduction, leading to a burst of ROS in a variety of ways, leading to PUFA lipid peroxidation and eventually causing cell membrane damage and DNA oxidation.26 This means that the ferroptosis-like death pathway can be triggered specifically by regulating iron metabolism, inducing lipid peroxidation, and destroying the antioxidant system, and it does not need to act on the targets of traditional antibiotics to effectively kill drug-resistant bacteria.26
On the basis of the aforementioned mechanisms, ferroptosis-like death has considerable potential as a novel antibacterial strategy.27 However, achieving efficient and precise regulation of ferroptosis-like death remains a key challenge in current research.28 Nanomaterials exhibit distinct advantages in mediating ferroptosis-like death for their potent antibacterial effects: 1) Nanomaterials can act as “iron reservoirs” to release iron ions in the infected microenvironment. These iron pools, in combination with increased membrane permeability, promote iron ion influx, leading to bacterial iron overload and ferroptosis-like death.25 2) Through chemodynamic therapy (CDT), photodynamic therapy (PDT) and sonodynamic therapy (SDT), nanomaterials can catalyze the generation of large amounts of reactive oxygen species (ROS) and trigger lipid peroxidation, further enhancing their antibacterial effects.29,30 3) With unique advantages such as small size effects and high specific surface areas, nanomaterials enable efficient drug loading and easy penetration of biofilms and cell walls.31 4) Through surface modifications and intelligent design, nanomaterials can achieve specific bacterial targeting to promote high accumulation of therapeutic agents at the infection site.32 Nanomaterials can penetrate the physical barrier of biofilms, reduce the risk of infection recurrence, and achieve synergistic treatment through multitarget and intelligent design. Therefore, ferroptosis induced by nanomaterials shows great potential in the fight against drug-resistant bacterial infections.33
Although ferroptosis has immense potential in the antibacterial field, comprehensive reviews systematically elaborating on the mechanisms by which nanomaterials overcome bacterial resistance through the induction of ferroptosis-like death are lacking. This review aims to systematically outline the mechanisms and research progress regarding how nanomaterials trigger ferroptosis-like death in bacteria via multiple mechanisms, such as by modulating bacterial iron metabolism, inducing ROS bursts, disrupting antioxidant defenses, and activating host immune responses. By combining the three elements of “nanomaterials - bacterial ferroptosis-like death - host immune regulation”, this review systematically explains the dual functions of nanomaterials in direct bactericidal and immune synergy. This review describes a variety of bactericidal mechanisms, including chemodynamic, photodynamic, sonodynamic, and nanoenzyme-mediated therapies, with an emphasis on their immunomodulatory potential, such as macrophage polarization, neutrophil function recovery, and immune memory formation. We comprehensively discuss the current challenges and future directions, providing a theoretical basis for the development of efficient and precise nanoantibacterial strategies to promote the clinical translation of bacterial ferroptosis-like death therapy.
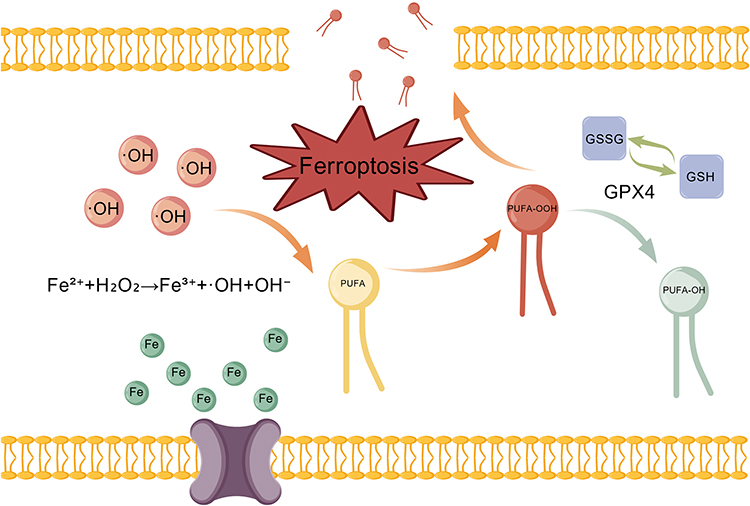 |
Figure 1 Core chemical reaction mechanism of ferroptosis. Ferrous iron (Fe2⁺) generates highly reactive hydroxyl radicals (OH) via the Fenton reaction. These radicals initiate a lipid peroxidation chain reaction in polyunsaturated fatty acids (PUFAs). Lipid hydroperoxides (PUFA-OOH) can be reduced to harmless lipid alcohols (PUFA-OH) by the glutathione (GSH)/glutathione peroxidase 4 (GPX4) system, thereby suppressing ferroptosis. When lipid hydroperoxides (PUFA-OOH) are overproduced or the antioxidant capacity is compromised, the accumulation of lipid hydroperoxides (PUFA-OOH) damages the membrane and ultimately culminates in ferroptosis. Created with https:BioGDP.com. |
Molecular Mechanisms of Ferroptosis
Definition and Mechanism of Ferroptosis
Ferroptosis is an iron-dependent regulatory form of cell death characterized by lipid peroxidation. The core of its chemical reaction mechanism is the peroxidation of polyunsaturated fatty acids (PUFAs).17,34 This process is initiated by the iron-mediated generation of reactive oxygen species (ROS) and is continuously enhanced in the absence of effective cellular antioxidant defenses, eventually leading to cell membrane breakdown.35,36 As shown in Figure 1, ferroptosis begins with the accumulation of intracellular free iron (Fe2+), which converts H2O2 to highly reactive hydroxyl radical (˙OH) via the Fenton reaction via the following formula: Fe2++H2O2→Fe3++˙OH+OH−.37,38 ˙OH can quickly attack the PUFA on the cell membrane and organelle membrane and extract hydrogen to form lipid free radicals.39 Lipid free radicals rapidly react with oxygen to form lipid peroxyl radicals, which in turn capture the hydrogen atoms of adjacent lipid molecules to form lipid hydroperoxides (PUFA-OOH) and generate new lipid free radicals˙, thus forming a chain propagation reaction.40 The rate of this propagation process depends on the dissociation energy of the C‒H bond, with hydrogen at the diallyl site being the most easily extracted; therefore, phospholipid molecules with high PUFA contents (PUFA‒PLs) are more susceptible to peroxidation.41 PUFA-OOH can be reduced to a harmless lipoalcohol (PUFA-OH) by glutathione peroxidase 4 (GPX4), thus interrupting the chain reaction.42 Once the formation of PUFA-OOH is not reduced in time, the accumulation of PUFA-OOH reaches a certain threshold, the integrity of the cell membrane is destroyed, and it eventually leads to cell death.37 Compared with traditional methods of cell death, such as apoptosis and necrosis, ferroptotic cells present unique morphological characteristics, such as reduced mitochondrial volume, increased membrane density, decreased crista structure, and left the nucleus intact. These characteristics are directly related to the chemical reaction mechanism driven by lipid peroxidation.24 In addition, lipoxygenases (LOXs), a class of iron-containing enzymes, directly catalyze the peroxidation of PUFA-PLs independent of the Fenton reaction, which is another important pathway for lipid peroxidation.43 This enzymatic reaction acts in concert with the nonenzymatic Fenton reaction to regulate the chemical process of ferroptosis.22,42 Although ferroptosis was initially characterized in mammalian cells, recent evidence suggests that microorganisms can undergo ferroptosis-like death. The core biochemical pathways of this process are highly similar to those of mammalian ferroptosis. A study by Kwun and Lee confirmed that in Saccharomyces cerevisiae (S. cerevisiae) and Vibrio vulnificus (V. vulnificus), treatment with gold nanoparticles (AuNPs) similarly resulted in intracellular iron accumulation and hydroxyl radical generation and induced a marked increase in the levels of malondialdehyde (MDA), the terminal product of lipid peroxidation.26
Regulation of Iron Metabolism
Iron is the core catalyst of ferroptosis, and cellular homeostasis precisely regulates sensitivity to ferroptosis through a variety of mechanisms.44 After binding to transferrin, extracellular Fe3+ enters the cell through endocytosis mediated by transferrin receptor 1 (TFR1), is reduced to Fe2+ by prostate transmembrane antigen 3 (STEAP3) in vivo, and is released into the cytoplasm via divalent metal transporter 1 (DMT1) to form a labile iron pool (LIP).45,46 Iron in LIP can directly participate in the Fenton reaction and produce ˙OH to promote lipid peroxidation.47 Excess iron in cells is usually stored in ferritin, which is composed of FTH1 and FTL subunits and has the activity of iron oxidase. Ferritin can convert Fe2+ to Fe3+ and store it in its protein cavity to avoid oxidative damage caused by excessive Fe2+.48,49 In iron deficiency, nuclear receptor coactivator 4 (NCOA4) mediates ferritinophagy, promotes the degradation of ferritin, releases iron ions, and increases the risk of ferroptosis.50,51 Ferroportin (FPN) is the only iron export protein, and its expression is regulated by hepcidin.52,53 Downregulation or functional inhibition of FPN can lead to intracellular iron accumulation and promote ferroptosis.54 Similarly, in microbial ferroptosis-like death, an increase in intracellular iron levels, which mediates ROS generation via the Fenton reaction, ultimately leading to ferroptosis-like death, is often observed.55
Regulation of Lipid Metabolism
The key to the regulation of ferroptosis by lipid metabolism is the synthesis and peroxidation of PUFA-PLs.56 PUFAs such as arachidonic acid (AA) and adrenal acid (AdA) are catalyzed by acyl-coenzyme A synthetase long-chain family member 4 (ACSL4) to form acyl-coenzyme A esters (PUFA-CoAs), which are then esterified to phosphatidylethanolamine (PE) by lysophosphatidylcholine acyltransferase 3 (LPCAT3). Thus, peroxidable PUFA-PLs are formed.21,57 These PUFA-PLs are the main substrates of lipid peroxidation, and members of the lipoxygenase (LOX) family, such as 12-LOX and 15-LOX, directly promote the ferroptosis process by catalyzing the peroxidation of PUFA-PLs.58 In contrast, membrane-bound O-acyltransferase family members (MBOAT1/2) inhibit ferroptosis by introducing monounsaturated fatty acids (MUFAs) into phospholipids and reducing the PUFA‒PL ratio.59 Exogenous MUFAs, such as oleic acid, are integrated into the membrane via ACSL3, dissubstituting PUFAs, and significantly reducing lipid ROS levels and ferroptosis sensitivity.60 Phospholipases iPLA2β and PAFAH2, which can hydrolyze oxidized phospholipids and remove lipid peroxides, are also considered important mechanisms for inhibiting ferroptosis.61,62 Thus, the remodeling of lipid metabolism not only affects the initiation of ferroptosis but also affects its rate and cell fate.
Notably, bacterial membranes are generally less susceptible to oxidation due to differences in polyunsaturated fatty acid (PUFA) integration capacity. Most bacterial and fungal membranes are composed of poorly oxidizable saturated or monounsaturated lipids, although exceptions exist in species such as Vibrio and Saccharomyces cerevisiae, which can incorporate exogenous PUFAs into membrane phospholipids, enabling lipid peroxidation and ferroptosis-like death.63
Antioxidant Defense System
There are a variety of antioxidant systems in cells that resist ferroptosis, including the GPX4-GSH and FSP1-CoQ10 pathways, of which the GPX4-GSH pathway is the classic anti-ferroptosis central pathway.64,65 GPX4 uses GSH as a reducing agent to reduce toxic PUFA-OOH to nontoxic PUFA-OH.66,67 The activity of GPX4 is critically dependent on the level of GSH and its own selenocysteine (Sec) active center. Selenium deficiency or mutation can lead to the loss of GPX4 function and aggravate ferroptosis.68 Any factor that causes GSH depletion or directly inhibits GPX4 activity induces ferroptosis.69,70 Independent of GPX4, the FSP1-CoQ10 pathway was also identified as a GPX4-independent ferroptosis inhibition pathway.71 Ferroptosis inhibitor protein 1 (FSP1) utilizes NADPH to reduce CoQ10 to CoQ10H2 at the cell membrane. As a lipid-soluble antioxidant, CoQ10H2 can directly capture lipid peroxyl radicals and interrupt the chain propagation of lipid peroxidation, thereby inhibiting ferroptosis.41,65 In addition to the core mechanisms mentioned above, multiple signaling pathways and transcription factors are involved in the regulation of ferroptosis. NRF2 is a key transcription factor that regulates the antioxidative stress response, and its activation can upregulate the expression of multiple ferroptosis-related genes, such as SLC7A11, GPX4, and FTH1,72,73 whereas p53 can inhibit and increase sensitivity to ferroptosis by inhibiting the expression of SLC7A11.74
In contrast to mammalian cells, bacteria lack the canonical GPX4 enzyme and often rely on alternative antioxidant defenses. For example, in Saccharomyces cerevisiae, a GPX4-like protein (GPx3) may partially fulfill a similar role, although its function and regulation differ.75,76 GSH depletion remains a critical trigger for ferroptosis-like death in bacteria.77 Once the antioxidant capacity is overwhelmed, it leads to membrane damage and cell death.
Nanomaterials Induce Bacterial Ferroptosis-Like Death
Although bacteria and eukaryotic cells exhibit mechanistic differences, both systems exhibit remarkable similarities in the core biochemical pathways of ferroptosis-like cell death. Under specific stimuli, they commonly exhibit intracellular iron accumulation, depletion of the glutathione system, and a consequent burst of reactive oxygen species.78–80 These events ultimately lead to lethal lipid peroxidation through iron-dependent Fenton reactions81. Such mechanistic parallels, and distinctions offer a novel perspective for addressing antibiotic resistance.82
 |
Figure 2 Construction of nanomaterials with a multifunctional synergistic system that interferes with (A) iron metabolism, (B) increases ROS production and (C) disrupts antioxidant systems to induce bacterial ferroptosis-like death. |
As shown in Figure 2, The core design strategy for the use of nanomaterials to induce bacterial ferroptosis-like death should involve the construction of multifunctional synergistic systems capable of disrupting iron metabolism, destroying the antioxidant system, and increasing ROS generation. 1) Disrupting Iron Metabolism: The material composition should incorporate iron-based active centers that enable the controlled release of Fe2⁺/Fe3⁺ ions in the bacterial microenvironment, triggering the Fenton reaction to generate ˙OH.83,84 2) Destroying the Antioxidant System: Introducing GSH-depleting units can disrupt bacterial antioxidant defense.85 3) Increasing ROS generation: Integrating physical stimulation or enzyme-mimicking components can increase the efficiency of cascaded ROS production.86,87 For example, sonodynamic therapy utilizes ultrasound to activate piezoelectric materials or sonosensitizers, thereby increasing ROS generation.88 Physical stimulation is a key strategy for enhancing the ferroptosis effect; it not only increases ROS production but also employs other mechanisms to induce ferroptosis; for example, photodynamic therapy can use near-infrared light to directly disrupt bacterial membrane structures and promote iron influx, whereas sonodynamics can increase material permeability.89 The rational design of nanomaterials can achieve synergistic induction of bacterial ferroptosis-like death through multiple mechanisms, thereby reversing bacterial drug resistance.
Nanomaterials Disturb Iron Metabolism in Bacteria
Disorders of iron metabolism constitute the core mechanism of bacterial ferroptosis-like death induced by nanomaterials.90 In recent years, a variety of nanomaterials have achieved multidimensional intervention in bacterial iron metabolism pathways through ingenious molecular design and functionalization strategies, which provides an important basis for the development of new antibacterial regimens.55,91 As shown in Table 1, we summarize the various ways in which different materials interfere with the iron metabolic pathways of bacteria.
 |
Table 1 Effects of the Multidimensional Intervention of Nanomaterials on Bacterial Iron Metabolic Pathways |
Nanomaterials that directly provide iron ions usually achieve iron loading and delivery via carriers. Zhou et al developed iron-loaded lipid nanoparticles (Fe-LNPs), which take advantage of the good biocompatibility and membrane fusion ability of liposomes to significantly improve the intracellular delivery efficiency and bioavailability of iron ions and maintain the intracellular iron concentration with the help of a sustained release effect by encapsulating ferric ammonium citrate (FAC) in the phospholipid bilayer. Moreover, the Fe-LNPs also significantly inhibited the proliferation of S. aureus and promoted wound healing. Using lipids as drug delivery carriers to trigger ferroptosis-like death through iron transport to achieve antibacterial effects may become a promising strategy.91 In another study, sustained Fe2+ release was utilized to effectively kill methicillin-resistant Staphylococcus aureus (MRSA). In this research, the ferrous sulfate (FeSO4)-embedded hyaluronic acid (HA) network and uniformly loaded Fe2+ hydrogel showed good sustained release performance in a Staphylococcus aureus infection model, with a 78.3% iron ion release rate within 12 h, indicating its potential for long-term and accurate iron delivery.55 In addition, Zhao et al loaded ultrasmall Prussian blue nanoparticles (UPBNPs) into the pores of mesoporous calcium silicate nanoparticles (MCSNs) to construct UPBNP-MCn composites. The mesoporous structure of MCSNs effectively prevents the aggregation of UPBNPs and regulates the release kinetics of iron ions, which not only results in simple iron loading but also creates a sustainable iron supply microenvironment. The Fe2+/Fe3+ released from UPBNP-MCSNs can invade the bacterial interior and catalyze the massive generation of ROS through the Fenton reaction, thereby initiating a ferroptosis-like pathway based on oxidative damage.92 The nanosystem, through the complete delivery pathway from “iron loading” to sustained and controllable “iron release” and subsequently to bacterial “iron overload,” achieves efficient iron transport and activates its antibacterial function.
By converting adverse factors in the pathological microenvironment (such as the acidic environment) into a “switch” that triggers ferrous ion release, the antibacterial precision is significantly enhanced. In the MOF-based antibacterial strategy, the MIL-100(Fe) nanomaterials studied by Yu et al underwent gradual degradation in the simulated wound microenvironment (pH 5.5). The MIL-100(Fe)@IR775 composite material formed after loading the photosensitizer IR775 not only enhanced the photothermal conversion efficiency of the material but also enhanced the ability to release iron ions. The sustained release of iron ions triggered by this acid and its reduction to Fe2+ provides the basis for the subsequent Fenton reaction to efficiently generate ˙OH and initiate lipid peroxidation.93 Additionally, in an acidic environment, Xu et al reported an intelligent delivery system based on a glycyrrhizic acid (GA) hydrogel, in which FeS nanoparticles were generated by an in situ reaction in response to the unique weakly acidic microenvironment (pH 5.0--6.0) of diabetic wounds. At this pH, the specific dissolution of FeS results in the sustained and simultaneous release of Fe2+ and H2S, which greatly promotes ferroptosis-like death in bacteria.94 To increase the precision of nanomaterial targeting, Wang et al designed an antibacterial composite material (Pal-ACPs) that targets the SSTR2 receptor on macrophage membranes, which utilizes the high affinity of the ZnO component for SSTR2 to significantly promote intracellular Fe2⁺ accumulation, thereby inducing ferroptosis in intracellular Staphylococcus aureus. These findings provide a new strategy for nanotherapeutic approaches against intracellular pathogens.95 Receptor-targeting strategies promote the active uptake of iron by bacteria, thereby achieving precise elimination of pathogens and providing a new perspective for nanotherapeutic strategies against intracellular pathogens.
Through transcriptome technology, several studies have revealed the regulatory effects of nanomaterials on iron homeostasis-related genes. The ZnO QOD-PTZ constructed by Shao et al can respond intelligently to bacterial microacid and amidase environments to trigger the controlled release of PTZ. This system significantly upregulated iron transport genes such as TonB, ExbD, and SufE, driving a large amount of iron ion influx. The induction of the ferroptosis-like pathway by iron enrichment synergistically enhances lipid peroxidation and GSH depletion, rapidly destroys membrane integrity and DNA stability, and achieves efficient synergistic sterilization.96 Sun et al reported that the expression of iron uptake genes (ygeO and yhfL) was significantly upregulated and that the expression of an iron storage gene (floX) was downregulated in E. coli O157: H7 treated with FeSO4, suggesting an imbalance in intracellular iron homeostasis. The study further revealed that the cinnamaldehyde nanoemulsion (CALNO) prepared by phaco emulsification had a small particle size and uniform distribution characteristics, which could effectively damage the integrity of the bacterial membrane and promote Fe ion influx, creating a synergistic effect with FeSO4. Combined treatment not only aggravated the expression changes in ferroptosis-like death-related genes but also effectively inhibited the expression of genes related to biofilm formation (such as csgA and luxS). This change in the expression profile disrupts intracellular iron homeostasis and drives the accumulation of free Fe2+, which in turn catalyzes a burst of lipid peroxidation to coordinate with ROS, ultimately inducing membrane breakdown and bacterial death.97 In addition, Xue et al constructed an Fe-doped titanite “nanosword” to destroy the bacterial membrane barrier in an alkaline microenvironment, resulting in a 75% reduction in the cell membrane stiffness of S. aureus (Figure 3). This weakening of mechanical properties makes it easier for the nanostructure to physically penetrate the bacterial membrane, and the Fe-doped titanite “nanosword” actively regulates the bacterial iron uptake system by triggering oxidative stress. Transcriptome analysis revealed that the expression of ABC transporter genes related to metal ion transport (such as IUJ41_07030, adcA) was significantly upregulated, indicating that the bacteria actively accelerated the influx of Fe2+ into the environment in response to stress.98 The aforementioned studies demonstrated that nanomaterials can not only facilitate iron influx but also actively regulate the expression of bacterial genes related to iron metabolism, enabling the precise activation of the bacterial ferroptosis pathway and synergistically leading to iron metabolic dysregulation, ultimately resulting in ferroptosis.
 |
Figure 3 Induction of bacterial ferroptosis-like death via iron metabolism dysregulation. (A) Illustration of the antibacterial mechanism: membrane damage by the nanostructure facilitates Fe2⁺ uptake, leading to oxidative stress and ferroptosis-like death. (B) Characterization of the nanostructure surface. The red dashed line depicts the cross-sectional morphology of the MH. (C) Visualization of bacterial membrane damage and intracellular iron accumulation posttreatment. EDS analysis within the red dashed boxes revealed 0.21% Fe inside the MH-group bacteria. (D) Transcriptomic profiling revealed the upregulation of ABC transporter-related genes. The white arrows highlight the upregulated gene IUJ41_02080. Reprinted from Xue Y, Zhang L, Liu F et al. Alkaline “nanoswords” coordinate ferroptosis-like bacterial death for antibiosis and osseointegration. ACS Nano. 2023;17(3):2711–2724, Copyright 2023, with permission from the American Chemical Society.98 Note: **P < 0.01. |
In summary, nanomaterials have evolved from “passive” iron donors to “active” regulators of bacterial iron metabolism. These strategies not only focus on simple ion release from nanoreservoirs but also emphasize intelligent and microenvironment-responsive iron release. Transcriptomic evidence confirms that these interventions disrupt iron homeostasis at the genetic level. This precise interference in iron metabolism not only supplies the essential catalyst for the Fenton reaction but also lays the material foundation for amplifying the ferroptosis effect via subsequent ROS bursts and antioxidant system collapse, constituting the first step of the synergistic antibacterial strategy.
Nanomaterials Induce a Burst of ROS in Bacteria
The generation of ROS is the key driving mechanism in the process of nanomaterial-induced bacterial ferroptosis-like death.99 A variety of physicochemical pathways are involved in this process, including chemodynamic therapy, photodynamic therapy, sonodynamic therapy, and nanoenzyme catalysis.100 These mechanisms not only act independently but also often have synergistic effects to promote oxidative stress and lipid peroxidation in bacteria, eventually leading to ferroptosis-like death of bacteria.100 As described in Table 2, they jointly promote oxidative stress and lipid peroxidation in bacteria and eventually lead to ferroptosis-like death of bacteria.
 |
Table 2 Different Pathways by Which Nanomaterials Induce Bursts of Reactive Oxygen Species (ROS) in Bacteria |
Chemodynamic Therapy (CDT)
As an emerging antibacterial strategy, chemodynamic therapy (CDT) has shown potential in combating drug-resistant bacterial infections by catalytically generating highly reactive free radicals from endogenous hydrogen peroxide (H2O2), which effectively triggers ferroptosis-like death in bacteria. Sun et al developed the bacteria-specific artificial biocatalyst MoS2/Fe@MBA@HA (MFBH) for the treatment of bacterial endophthalmitis. This material promotes the reduction of Fe3+ to Fe2+ when the exposed Mo4+ is used as an electron donor. Moreover, the sulfur vacancy enhances the affinity for H2O2 and continuously generates many hydroxyl radicals (˙OH) with strong oxidation, thus achieving an efficient Fenton catalytic reaction in the infected microenvironment, leading to lipid peroxidation, GSH depletion, and energy metabolism disorders and finally inducing ferroptosis-like death in S. aureus.101 To overcome the limitations of traditional CDT in practical applications, such as insufficient H2O2 and slow Fe3+/Fe2+ cycling, Zheng et al constructed an oxygen vacancy-rich [(BiFe)0.9(BaTi)0.1O3-X] (BFBT) nanoreactor through defect engineering, which significantly enhanced the efficiency of chemical kinetics, demonstrating that the abundant oxygen vacancies in the material can act as electron donors to effectively regulate the local electron density of adjacent Fe sites, greatly accelerating the valence cycle of the Fe3+/Fe2+ and enabling the Fenton reaction to continue and carry out efficiently. More importantly, the system provides sufficient substrates for CDT to self-supply H2O2 through an ultrasound-driven piezoelectric effect, forming a unique self-recycling catalytic mechanism to induce ferroptosis-like death.102 Similarly, Wang et al developed SPEEK/PDA-PS/PLGA@Fe (SP-PFe), which is a unique chemodynamic treatment strategy that overcomes the dependence of the Fenton reaction on H2O2 and O2 (Figure 4A). The key lies in the construction of an Fe2+-mediated catalytic cascade system independent of external energy. In the acidic microenvironment of the infection site, ˙SO4− was generated, while the Fe2+ released from the implants catalyzed the generation of ROS. ˙SO4− not only attacked the bacterial membrane but also further reacted with H2O to generate ˙OH, which triggered a strong oxidative storm locally and eventually led to ferroptosis-like death to treat infected bone defects.103 In summary, the above studies effectively overcome the key limitations of traditional chemical therapy through structural regulation and reaction pathway innovation and achieve efficient and specific induction of ferroptosis-like death. These strategies not only increase the efficiency of the Fenton reaction but also expand the application scenarios of CDT, fully demonstrate the unique advantages of nanomaterials in the anti-infection treatment of “low dose, low toxicity, and resistance”, and provide important references for the development of antibacterial nanomedicins in the future.
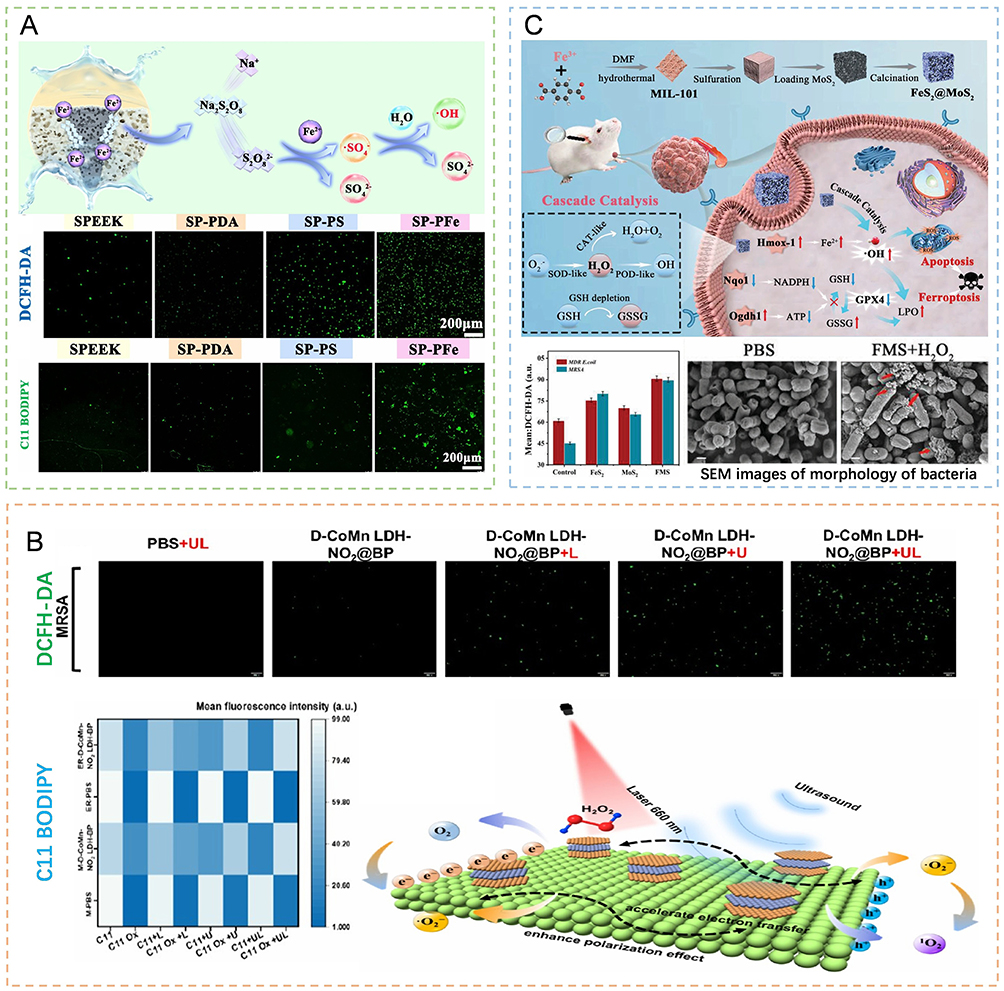 |
Figure 4 Diverse physicochemical pathways that induce oxidative stress and lipid peroxidation in bacteria, ultimately leading to bacterial ferroptosis-like death. (A) SP-Pfe-mediated chemodynamic therapy elicits a marked increase in intracellular ROS and lipid peroxidation, culminating in bacterial ferroptosis-like death. Reprinted from Wang Z, Huang Y, He S et al. Oxygen-independent sulfate radical and Fe2+-modified implants for fast sterilization and osseointegration of infectious bone defects. ACS Nano. 2025;19(19):18,804–18823, Copyright 2025, with permission from the American Chemical Society.103 (B) In response to laser and ultrasound irradiation, D-CoMn LDH-NO2@BP facilitates PDT/SDT-driven ROS accumulation and lipid peroxidation, causing bacterial ferroptosis-like death. Reprinted from An X, Chen B, Yuan Y et al. Laser-ultrasound responsive nanoreactor to remodel the microenvironment of antibiotic-resistant bacterial infection for diabetic wound healing. Nano Lett. Published online August 28, 2025, Copyright 2025, with permission from the American Chemical Society.108 (C) The catalytic activity of the FeS2@MoS2 nanozymes generates a surge of ROS, inducing bacterial ferroptosis-like death and concomitant membrane damage. The red upward arrow in the mechanism denotes an increase, whereas the blue downward arrow represents a decrease. The red arrows in the SEM image indicate the damaged cell membranes of the bacteria after treatment. Reprinted from Sun M, Wang L, Zhuo Y et al. Multienzyme activity of MIL-101 (Fe)-derived cascade nanoenzymes for antitumor and antimicrobial therapy. Small 2024;20(17): e2309593, Copyright 2023, with permission from John Wiley and Sons.109 |
Photodynamic Therapy (PDT) and Photothermal Therapy (PTT)
In recent years, researchers have focused on constructing a variety of photoactive materials to achieve efficient and specific anti-infective effects through photodynamic therapy and photothermal therapy, which provides new treatment ideas for drug-resistant bacteria and related biofilm infections. The CuSA-COF monatomic material developed by Wu et al not only generated large amounts of singlet oxygen (1O2), hydroxyl radicals (˙OH) and protons (H+) under a 635 nm laser but also formed an acidic microenvironment via the accumulation of protons in the pores, thereby self-enhancing peroxidase-like activity through the “metal-protonation” strategy. Finally, it can effectively eliminate MRSA and its biofilms through a lipid peroxidation-mediated ferroptosis-like mechanism and significantly promote wound healing in vivo, resulting in excellent dual antibacterial and tissue repair functions.104 To further expand the application of COF materials in PDT, Sun’s team constructed two single-atom catalysts, sp2c-COF-Ir-ppy2 and sp2c-COF-Ru-bpy2. These nanomaterials not only have excellent visible light absorption and charge separation properties but also exhibit efficient peroxidase-like activity. Under the stimulation of red light, the Ru site generates 1O2 through the energy transfer pathway, whereas the Ir site catalyzes ˙OH production in the presence of H2O2. The two synergistically form an ROS storm, which not only directly causes oxidative damage to bacteria but also consumes glutathione and inactivates antioxidant enzymes, thereby disrupting the bacterial antioxidant defense system and triggering ferroptosis-like death.105 In addition to COF nanomaterials, multifunctional hydrogel systems also perform well in the synergistic induction of bacterial ferroptosis-like death. The FCL-ECMH hydrogel developed by Lu et al is able to responsively release iron ions in the infected microenvironment and undergo photothermal transformation under excitation with 808 nm near-infrared light. Local heating not only directly destroys the bacterial structure but also enhances iron influx, which promotes the Fenton reaction to catalyze the conversion of endogenous H2O2 to ˙OH and induces ferroptosis-like death in bacteria. This system achieves the synergistic effect of photothermal therapy and chemodynamic therapy and has significant advantages in destroying biofilms and regulating bacterial iron metabolism, providing an innovative strategy for anti-infection treatment.106 These strategies not only effectively improve antibacterial efficiency but also promote tissue repair, reflecting the broad prospects of light-controlled nanomaterials in anti-infection treatment.
Sonodynamic Therapy (SDT) and Sonodynamic Therapy (SPT)
Sonodynamic catalysis generates ROS via ultrasound-activated sonosensitizers because of their excellent tissue penetration ability and spatiotemporal controllability. To further enhance the sonodynamic effect, Li et al constructed an organic‒inorganic heterojunction structure, TOMPE, by introducing manganese ions into the TiO2 lattice and modifying it with emodin (ED). This design effectively inhibits the rapid recombination of electron‒hole pairs in TiO2 and reduces the energy band gap of the material through an imbalance in the interfacial charge distribution, thereby promoting the generation of more charge carriers and increasing ROS production under ultrasonication. The generated ROS not only consume glutathione but also cause lipid peroxidation and DNA damage, eventually leading to ferroptosis-like death in MRSA.107 To induce bacterial ferroptosis-like death more efficiently, An et al developed a laser-ultrasound dual-response nanoreactor, D-CoMn LDH-NO2@BP, to achieve synergistic treatment with PDT and SDT. Under 660 nm laser excitation, significant electron polarization occurs at the interface between black phosphorus (BP) and defective layered double hydroxide (LDH), which improves the generation efficiency of singlet oxygen (1O2) (Figure 4B). Ultrasound stimulation not only enhances charge separation at the interface but also induces an acoustic current through the piezoelectric effect, which jointly promotes the massive generation of ˙O2−. In a diabetic wound model, this strategy increased the healing rate 3.18-fold, demonstrating excellent synergistic antibacterial potential.108 Unlike the above strategy of relying on exogenous sensitizers, Zheng et al innovatively used Ba/Ti-doped BFBT nanocrystals as voltage sonosensitizers for SPT. Under ultrasonication, the noncentral symmetric crystal structure is deformed, generating a strong internal electric field, which effectively drives charge separation and inhibits electron‒hole recombination. BFBT plays an “engine” role in the ferroptosis-like death process by enabling efficient electron reduction of O2 to ˙O2−, self-supplying reaction substrates for chemical treatment and achieving sustained antibacterial effects.102 In summary, from constructing heterojunctions to enhancing the separation of electrons and holes, combining multimodal catalysis to increase ROS production, and developing H2O2-supplied piezoelectric sonosensitizing agents, these strategies have deepened the understanding of the mechanism of the sonodynamic induction of bacterial ferroptosis-like death.
Nanoenzyme Catalysis
Nanoenzyme catalysis, which enables continuous generation of ROS by mimicking native enzyme activity, has shown important potential in antiinfective therapy induction strategies in recent years.112 The FeS2@MoS2(FMS) nanoenzyme developed by Sun et al has three mimicking activities, namely, superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT), and thus involves a self-cycling ROS generation pathway (Figure 4C). This material first converts the O2− produced by bacterial metabolism to H2O2 via SOD activity and then converts H2O2 to highly toxic ˙OH via POD-like activity to induce ferroptosis-like death in bacteria. Moreover, the CAT-like activity under neutral conditions can decompress H2O2 to produce O2, which is a continuous substrate for the SOD reaction, forming a self-enhancing catalytic cycle. This process effectively disrupts the bacterial REDOX balance and has been shown experimentally to inhibit MRSA biofilms and promote wound healing.109 To cope with the multiple challenges associated with diabetic wounds, such as high glucose levels, susceptibility to infection, and angiogenesis disorders, researchers have designed a cascade nanoenzyme system that is more pathogen-specific.113,114 Recently, a FeS@Au nanoenzyme was constructed to achieve synergistic intervention of “blood glucose regulation-sterilization-angiogenesis”. Among them, gold nanoclusters (aunCs) exhibit glucose oxidase-like activity and catalyze the production of gluconic acid and H2O2 from local glucose in wounds, providing substrates for subsequent reactions while reducing glucose and acidifying the microenvironment. The POD-like activity of FeS nanoparticles is enhanced under acidic conditions, which can efficiently convert H2O2 to ˙OH and induce ferroptosis-like death in drug-resistant bacteria. Moreover, H2S released by FeS can upregulate hypoxia-inducible factor-1 (HIF-1) and promote the expression of vascular endothelial growth factor (VEGF) and other related genes, thereby enhancing tissue repair ability and alleviating the damage caused by ROS in normal cells.110 In terms of intelligent ROS regulation, the Fe3O4/MXene (FM) heterojunction developed by You et al has a “bidirectional” function: on the one hand, it has CAT-like and SOD-like activities to degrade hydrogen peroxide, alleviate oxidative stress, and protect host cells from oxidative damage. On the other hand, it continuously releases Fe2+/Fe3+ through unique interface defects, triggering ferroptosis-like death within bacteria and leading to ROS bursts and lipid peroxidation. In a diabetic rat model, FM-loaded gelatin methylacrylamide (GFM) combined with photothermal therapy effectively eliminates S. aureus and promotes healing, demonstrating the intelligent potential of nanozymes in the dynamic regulation of ROS and ferroptosis-like death induction.111 ROS are the core effector molecules of bacterial ferroptosis-like death induced by nanomaterials. Researchers need to rationally design the composition, structure and surface properties of nanomaterials to achieve efficient and controllable generation of ROS through multiple mechanisms and then trigger ferroptosis-like death in bacteria, which provides a new strategy for the treatment of drug-resistant bacterial infections.
The various physicochemical pathways reviewed (CDT, PDT/PTT, SDT, and nanozyme catalysis) do not operate in isolation but rather form a synergistic “ROS-generating toolkit.” A common theme is overcoming the limitations of single-modality therapies, such as using defect engineering or constructing cascade systems to address insufficient H2O2 and slow Fe3⁺/Fe2⁺ cycling in CDT. Potent synergies exist between these mechanisms: for example, localized heating from PTT not only directly damages bacteria but also enhances membrane permeability, facilitating iron influx and Fenton reaction efficiency; similarly, SDT-induced sonoporation can enhance tissue penetration and the cellular uptake of materials. Crucially, these ROS-generating mechanisms are tightly coupled with iron dysregulation. The Fe2⁺ released by nanomaterials or promoted for uptake is the core catalyst for CAT- and POD-like activity; ROS storms can further damage bacterial membranes, accelerating iron influx and thereby establishing a self-reinforcing positive feedback loop for ferroptosis. Therefore, integrating multiple ROS-generating modalities with iron delivery capabilities is key to designing highly efficient and rapid-acting nanoantibacterial agents.
Nanomaterials Disrupt Bacterial Antioxidant Defense Systems
In the process of bacterial resistance to external threats and maintenance of cellular homeostasis, the antioxidant defense system plays a crucial role in scavenging excessive ROS, repairing oxidative damage, and maintaining normal metabolic function.115,116 Shen et al first introduced a ferroptosis-like mechanism into the field of bacterial research in 2020 and reported that synthetic aqueous ferrous polysulfide (Fe(II)Snaq) could efficiently induce ferroptosis-like death in bacteria. The key to this process is that the polysulfide in Fe(II)Snaq can oxidize with GSH in bacteria to form oxidized glutathione (GSSG), which directly depletes GSH stores and impairs antioxidant defense. Moreover, Fe2+ in the system promotes ROS accumulation and lipid peroxidation through the Fenton reaction, synergistically promoting the ferroptosis-like death process. Notably, polysulfide not only effectively counteracts the inhibitory effects of GSH but also enhances the stability of Fe2+ and prevents its oxidative inactivation, thereby continuously catalyzing the ROS burst. This dual synergistic effect enabled Fe(II) Snaq to maintain a high antibacterial rate of more than 90% in complex media.117 Following a similar GSH depletion strategy, Hu et al constructed biomimetic membrane-coated particles mFe-CA, further expanding the possibility of ferroptotic applications in antimicrobial applications. Cinnamaldehyde (CA), an α- and β-unsaturated aldehyde, reacts specifically with the sulfhydryl group of GSH, irreversibly consuming GSH in bacteria and converting it to GSSG, thereby disrupting REDOX homeostasis in bacteria. The depletion of GSH aggravates the Fenton reaction and the accumulation of hydroxyl free radicals (˙OH), which further enhances lipid peroxidation, inhibits the activity of the respiratory chain complex, and finally induces ferroptosis-like death in MRSA.118
In addition to depleting GSH directly, some nanomaterials produce ROS in large quantities by mediating the Fenton reaction, thereby indirectly depleting GSH and destroying the antioxidant defense system of bacteria. For example, FeSO4-loaded hyaluronic acid hydrogels can continuously release Fe2+ in the infected microenvironment, catalyze the formation of ˙OH through the Fenton reaction, and trigger severe oxidative stress, leading to intracellular ROS accumulation and lipid peroxidation. Transcriptome analysis revealed that genes related to GSH metabolism (such as glnA and metE) were significantly upregulated by Fe2+, indicating that the bacteria tried to activate antioxidant defense to cope with oxidative damage. The downregulation of genes involved in the cell membrane synthesis, such as yidC and secY, supported loss of membrane integrity. Notably, the addition of exogenous GSH or its precursor N-acetylcysteine (NAC) significantly inhibited the Fe2+-induced ROS burst and bacterial death, confirming the critical role of GSH system exhaustion in this bactericidal mechanism.55 Similarly, the CuFeS2 nanoenzyme developed by Wang et al not only efficiently produces ROS under photoexcitation but also significantly consumes GSH within bacteria, destroying their antioxidant capacity and leading to REDOX imbalance. This process was reversed by the ferroptosis inhibitor ferrostatin-1, which further confirmed that ferroptosis-like death is induced by interference with the GSH metabolic pathway.77
Furthermore, by influencing the expression of antioxidant enzymes, nanomaterials provide a new direction for the precise intervention in the bacterial antioxidant system. Wu et al reported that the combination of the photodynamic and Fenton reactions of CuSA-COF results in the production of a large amount of ROS, which directly attack the bacterial antioxidant system and lead to the rapid depletion of GSH, thereby inactivating antioxidant enzymes. The experiments revealed significant downregulation of antioxidant protein expression by approximately 57%, accompanied by massive accumulation of 4-HNE, a lipid peroxidation product, indicating aggravated oxidative damage, which eventually led to ferroptosis-like death.104 The sp2c-COF-Ir-ppy2 and sp2c-COF-Ru-bpy2 constructed by Sun et al simulated POD and glutathione oxidase activity under red light or H2O2 activation, respectively, rapidly oxidized GSH and significantly downregulated antioxidant enzyme expression in gram-negative bacteria. Thus, the bacterial antioxidant defense system is systematically destroyed, and irreversible ferroptosis-like death is induced.105
Disruption of the bacterial antioxidant defense system, particularly targeting glutathione (GSH), ultimately triggers ferroptosis-like death. Iron dysregulation and ROS bursts are powerful drivers of GSH depletion; GSH depletion cripples the function of antioxidant enzymes, rendering the cell more vulnerable to lipid peroxidation and amplifying the damage signals initiated by iron and ROS. Thus, the collapse of the antioxidant system is not an isolated endpoint but a critical amplifier that translates iron overload and ROS stress into irreversible cell death.
Integration of Synergistic Mechanisms: Toward a Unified Antibacterial Strategy
The efficacy of nanomaterials in inducing bacterial ferroptosis-like death does not rely on maximizing a single mechanism but rather stems from the exquisite synergy and amplification between the three core events: iron metabolism interference, ROS burst, and antioxidant defense collapse. Iron as the foundation and catalyst: Through intelligent delivery and active regulation, nanomaterials induce iron overload within bacteria. This directly provides the reactant for the Fenton reaction and forms the chemical basis for initiating lipid peroxidation. ROS as the effector executioner: ROS generated via various pathways (CDT, PDT, SDT, and nanozyme catalysis) launch direct attacks on lipids, proteins, and DNA. This process is catalyzed not only by iron but also through membrane damage, facilitating further iron influx and establishing a positive feedback loop. The Antioxidant system collapses as the “Terminal Switch”. By depleting key antioxidants such as GSH directly or indirectly, nanomaterials disarm the bacterial repair machinery for lipid peroxidation damage. This prevents the reduction of accumulated lipid hydroperoxides, ultimately leading to the irreversible loss of cell membrane integrity. Consequently, future research should focus not only on developing new nanomaterials but also on the rational design of multifunctional synergistic systems. An ideal nanoplatform should be capable of triggering these three core events simultaneously or sequentially, forming a self-propagating “lethal triangle” cycle.
Synergistic Therapy Involving Immune Responses and Nanomaterial-Induced Bacterial Ferroptosis-Like Death
Nanomaterial-induced bacterial ferroptosis-like death not only directly kills pathogens but also modulates the host immune system by regulating macrophage polarization, cytokine networks, neutrophil function, and lymphocyte immune memory, thereby constructing a multidimensional synergistic therapeutic system of “bactericidal-immune regulation-promoted repair”.
Macrophage Polarization
Macrophages, as central players in host immune defense, play pivotal roles in regulating the balance between infection and repair through their dynamic polarization states.119 In response to microenvironmental signals, macrophages can polarize into the proinflammatory bactericidal M1 phenotype or the anti-inflammatory, tissue-reparative M2 phenotype.120 Regulating this polarization is crucial for maintaining the equilibrium between pathogen clearance and anti-inflammatory repair.119 In this context, the TOMPE nanoplatform demonstrates excellent temporal immunomodulatory capabilities both in vitro and in vivo, guiding macrophage polarization to respond to different stages of infection. Under ultrasound triggering, the TOMPE nanoplatform releases manganese ions (Mn2⁺), which activate the cAMP/cGMP-PKG signaling pathway, significantly upregulate the expression of the M1 marker iNOS, enhance the phagocytic capacity of macrophages against MRSA, and achieve rapid early bactericidal effects. As infection is controlled, the loaded emodin promotes the transformation of macrophages to the M2 type, with an increase in CD206 and a decrease in iNOS. Metabolomic analysis confirmed that this treatment downregulates inflammation-related pathways such as cAMP/cGMP-PKG, thereby controlling the overall inflammatory response and promoting the osteogenic differentiation of bone marrow mesenchymal stem cells, synergistically facilitating tissue repair.107 In another study (Figure 5A), the FCL-ECMH hydrogel significantly promoted macrophage polarization toward the M2 phenotype by activating the JAK-STAT and PI3K/Akt/mTOR signaling pathways, manifested as increased CD206 expression, decreased CCR7, increased secretion of IL-10, and reduced TNF-α. Furthermore, in animal models of diabetic wounds and bone defects, FCL-ECMH combined with near-infrared light irradiation effectively cleared bacteria, promoted wound healing, and enhanced bone tissue regeneration.106 A growing body of research now focuses on the role of nanomaterials in regulating the immune microenvironment, particularly macrophage polarization.102 In a comparable approach, plant extracellular vesicles (C-evs) within CC-TFP hydrogels enriched with diverse bioactive molecules can be internalized by macrophages to directly modulate their functional state. Experimental results demonstrated that CC-TFP treatment significantly reduces M1 marker expression (iNOS and CD86) in RAW264.7 macrophages while upregulating the M2 marker CD206, accompanied by improved neovascularization and collagen deposition, thereby accelerating wound healing repair.121 In summary, nanomaterials intelligently regulate macrophage polarization by precisely intervening in key signaling pathways, such as the cAMP/cGMP-PKG, JAK-STAT, and PI3K/Akt/mTOR pathways, to achieve synergy between infection clearance and tissue regeneration.
 |
Figure 5 Nanomaterials induce bacterial ferroptosis-like death while modulating macrophage polarization and cytokine secretion to promote tissue repair. (A) FCL-ECMH facilitates macrophage polarization toward the M2 phenotype. This process involves the activation of the JAK-STAT and PI3K/Akt signaling pathways, as evidenced by increased phosphorylation levels of STAT6 and Akt, ultimately enhancing the repair of infected bone defects and new bone formation. Reprinted from Lu R, Luo Z, Zhang Y, Chen J, Zhang Y, Zhang C. A multifunctional tissue-engineering hydrogel aimed at regulating bacterial ferroptosis-like death and overcoming infection toward bone remodeling. Adv Sci 2024;11(30):e2309820, Open access.106 (B) COT-mediated programmable thermal CO bubble therapy effectively treats bacterial infections and modulates inflammatory responses. Following NIR irradiation (0.3 W/cm2) of COT surfaces, macrophages exhibit significant downregulation of the proinflammatory cytokine TNF-α and upregulation of the anti-inflammatory cytokine IL-10. Furthermore, the gene expression levels of key soft tissue repair-related factors (COL-1, CTGF, VEGF, TGF-β1, and α-SMA) were significantly elevated at 4 and 7 days post-implantation. Reprinted from Ding T, Zhang L, Chen J, Ma D, Han J, Han Y. “thermal bubbles”: photothermally triggered by a carbon monoxide nanocontainer for antibiosis and immune modulation therapy. Nano Today. 2025;63:102758, Copyright 2025, with permission from Elsevier.122 Notes: *P<0.05, **P<0.01, ***P<0.001. |
Networked Regulation of Cytokines
In research on nanomaterial regulation of immune responses, the MRMN microneedle system, by continuously releasing riboflavin with the assistance of near-infrared light, significantly downregulates the phosphorylation levels of key NF-κB pathway proteins p65 and IκBα, thereby inhibiting overactivation of this pathway. This blockade of upstream core signaling events effectively reduces the gene expression and secretion of key proinflammatory cytokines such as TNF-α and IL-6 while promoting the production of anti-inflammatory cytokines such as IL-4 and IL-10, successfully shifting macrophage polarization toward the M2 type.123 Moreover, the immunomodulatory function of these nanomaterials is also reflected in their synergistic activation of repair pathways; the ETN@Fe7S8 composite material, while inducing bacterial ferroptosis, also inhibits the NF-κB pathway, downregulates TNF-α and IL-6, and upregulates IL-10 and Arg-1, thereby alleviating inflammation. The released polysulfides significantly promote the expression of VEGFA in human umbilical vein endothelial cells and activate the PI3K/AKT pathway (as evidenced by increased phosphorylation of PI3K and AKT), thereby significantly enhancing endothelial cell tubulation and migration capabilities and promoting wound angiogenesis.124 These findings indicate that nanomaterials can synergistically promote anti-inflammatory effects and tissue regeneration by simultaneously regulating the NF-κB inflammatory pathway and the PI3K/AKT repair pathway. The COT light-responsive carbon monoxide nanocarrier demonstrated the ability to achieve dose-dependent immunomodulation through external stimuli (Figure 5B). After clearing bacteria in the antibacterial mode (0.7 W/cm2), switching to the immunomodulatory mode (0.3 W/cm2) significantly improved the immune microenvironment around the implant. Immunofluorescence analysis revealed that this treatment significantly reduced the expression of proinflammatory factors (TNF-α, IL-6, and iNOS) while increasing the gene expression of anti-inflammatory cytokines (IL-10 and TGF-β) and tissue repair factors (COL-1, CTGF, VEGF, TGF-β1, and α-SMA).122 This spatiotemporal regulatory capability highlights the advantages of intelligent nanoplatforms in dynamic immune intervention. The immunomodulatory effects of nanomaterials not only rely on external stimuli but also exhibit intrinsic dose-dependent characteristics. The dose-dependent bidirectional regulatory effect of the ZnO nanoparticles further reveals the complexity of immunomodulation. Low-dose ZnONPs exert anti-inflammatory effects by increasing autophagy (increasing the LC3-II/I ratio and decreasing p62) and inhibiting proinflammatory factors (COX-2, iNOS, IL-6, and TNF-α). Conversely, high-dose ZnONPs promote the secretion of proinflammatory cytokines (IL-1β, IL-6, and TNF-α) by increasing ferroptosis-like death (increased ACSL4 and elevated intracellular iron). Notably, the ferroptosis inhibitor Fer-1 not only reverses the proinflammatory response induced by high-dose ZnONPs but also alleviates the resulting acute lung injury and enhances the antibacterial efficacy of ZnONPs in vitro and in vivo, suggesting an association between the ferroptosis process and inflammatory responses.125 Through precise intervention in key immune signaling pathways and influencing the expression of inflammatory and repair-related factors, nanomaterials achieve a multifaceted “antibacterial–anti-inflammatory–prorepair” regulatory mechanism, revealing a refined strategy for promoting infectious wound healing by targeting the immunometabolic network.
 |
Figure 6 Nanomaterial-induced bacterial ferroptosis-like death potentiates neutrophil-mediated bactericidal activity and elicits B-cell-dependent long-term immunological memory, enhancing antibacterial efficacy. (A) FGO@MN treatment enhances neutrophil function (CD54 upregulation) by promoting chemotaxis toward macrophages, improving bacterial phagocytosis, and suppressing NETosis (reduced Cit-H3). Scale bars: 20 µm. Reprinted from Zhu W, Mei J, Zhang X et al. Photothermal Nanozyme-Based Microneedle Patch against Refractory Bacterial Biofilm Infection via Iron-Actuated Janus Ion Therapy. Adv Mater. 2022;34(51):e2207961. Copyright 2022, with permission from John Wiley and Sons.126 (B) COF-based single-atom catalysts induce ferroptosis-like death, resulting in increased CD19⁺/CD21⁺/CD45⁺ B cells. This process assists the host in secondary defense against infection and promotes the development of long-term memory immunity. Reprinted from Sun B, Wang X, Ye Z et al. Designing single-atom active sites on sp2-carbon-linked covalent organic frameworks to induce ferroptosis-like bacteria for robust anti-infection therapy. Adv Sci 2023;10(13): e2207507,. Open access.105 Notes: **P < 0.01; ***P < 0.001; ****P < 0.0001. |
Neutrophil Functional Recovery
Neutrophils, as core innate immune effector cells, clear pathogens during early bacterial infection via chemotaxis, phagocytosis, and neutrophil extracellular trap (NET) formation.127 However, the biofilm microenvironment (BME) often suppresses their function through mechanisms such as iron deprivation. FGO@MN, developed by Zhu et al, exhibits peroxidase-like activity activated in the BME (acidic, high H2O2) (Figure 6A). FGO@MN catalyzes the production of highly toxic ˙OH, which synergizes with the intracellular iron overload caused by FGO-released iron ions to induce bacterial ferroptosis-like death, fundamentally disrupting the biofilm structure. Concurrently, iron ions released from FGO@MN degradation in the BME effectively reversed the BME-induced “iron starvation” state in neutrophils, directly promoting their comprehensive functional recovery. Experiments confirmed that iron replenishment significantly upregulates the proinflammatory phenotype marker CD54 on neutrophils, facilitating their repolarization from a suppressed state to an activated state. FGO@MN-treated neutrophils exhibit significantly enhanced chemotactic capacity for macrophage recruitment, with upregulated chemokines CCL2 and CCL3, promoting immune cell recruitment to the infection site, alongside a substantial increase in direct bacterial phagocytosis efficiency. Crucially, restored intracellular iron levels inhibit NET formation (reduced Cit-H3, MPO), a critical shift, as excessive NETs exacerbate tissue damage and impede repair; thus, NET inhibition creates a more favorable tissue repair microenvironment. Ultimately, via this synergistic antibacterial–immune mechanism, FGO@MN eradicates biofilms while driving the infectious microenvironment from proinflammatory destruction toward prorepair.126 This dual strategy, which induces bacterial ferroptosis-like death while simultaneously revitalizing neutrophil function, synergistically enhances antibacterial efficacy and offers new insights into nanomaterial-mediated regulation of host immune responses.
Lymphocyte-Mediated Long-Term Immunity
In addition to direct pathogen killing, nanomaterial-induced bacterial ferroptosis-like death can activate the host adaptive immune response, establishing long-lasting, pathogen-specific immune memory. Sun et al constructed single-atom catalysts based on sp2-carbon-linked covalent organic frameworks (sp2c-COF-Ir-ppy2 and sp2c-COF-Ru-bpy2) (Figure 6B). In a mouse model of MRSA abscess, these bacteria not only cleared bacteria efficiently via ferroptosis-like death but also significantly promoted the formation of specific immune memory in the host, reducing the risk of reinfection. Compared with control mice, mice treated with Ir or Ru SACs presented a significantly greater proportion of CD19⁺/CD21⁺/CD45⁺ B cells in their blood, indicating activated B-cell-mediated immunity and suggestive of induced adaptive immunity. Upon rechallenge with homologous MRSA without further treatment, these mice effectively resist infection, demonstrating a long-term immune memory effect. Transcriptomic and immunohistochemical analyses indicated that Ir and Ru SACs treatment also significantly downregulated the expression of proinflammatory cytokines (TNF-α, IL-1β, and IL-6), promoted M2 macrophage polarization (increased CD206⁺, decreased CD80⁺), and increased vascular endothelial growth factor (VEGF) and CD31 expression, thereby accelerating angiogenesis and tissue repair, ultimately achieving synergistic infection control and wound healing.105 The establishment of this nanomaterial-mediated immune memory is potentially linked to antigenic components released during bacterial ferroptosis-like death, such as lipid peroxidation-modified membrane proteins and nucleic acid fragments. These components may act as “danger signals” recognized and processed by antigen-presenting cells, subsequently activating T cells and assisting B-cell differentiation into plasma cells and memory B cells. Nanomaterial-induced bacterial ferroptosis not only clears the current infection but also provides the host with long-term protection against reinfection by initiating adaptive immunity.
 |
Figure 7 Nanomaterial-induced bacterial ferroptosis orchestrates a temporal switch in immune polarization through distinct signaling pathways. During the initial stage, nanomaterials and bacterial signals activate proinflammatory pathways such as cAMP/cGMP-PKG, promoting macrophage polarization to the M1 phenotype and increasing iNOS and proinflammatory cytokine expression to ensure effective pathogen clearance. In the subsequent resolution phase, pathways such as the JAK-STAT and PI3K/Akt/mTOR pathways are activated, whereas NF-κB signaling is downregulated. This promotes a shift to the M2 phenotype, which secretes anti-inflammatory mediators such as IL-10 and TGF-β to promote tissue repair. (+, activation; -, inhibition). Created with https:BioGDP.com. |
In summary, while inducing bacterial ferroptosis, nanomaterials can achieve temporal and networked regulation of innate and adaptive immunity. As presented in Figure 7, while nanomaterials induce bacterial ferroptosis-like death, they simultaneously stimulate macrophages to activate pathways such as the cAMP/cGMP-PKG pathway, which promotes early proinflammatory (M1) responses and enhances bactericidal activity. As infection is controlled, nanomaterials drive macrophage polarization toward the M2 phenotype by activating repair pathways such as JAK-STAT and PI3K/Akt/mTOR and secrete anti-inflammatory factors such as IL-10 and TGF-β while downregulating NF-κB activity, collectively shifting the immune microenvironment from proinflammatory to repair. By generating components such as iron ions, nanomaterials can directly restore neutrophil function and enhance their chemotaxis and phagocytic capacity while suppressing excessive NET formation to prevent tissue damage. Furthermore, by triggering bacterial ferroptosis-like death, nanomaterials initiate a cascade that activates antigen-specific B cells, driving the production of memory B cells to establish long-term immune memory. This integrated “bactericidal–immune regulation–promoted repair–immune memory” approach therefore constitutes a transformative strategy in combating infections.
Summary and Prospects
In recent years, several treatment strategies have been developed to address the issue of antimicrobial resistance.128 The use of nanomaterials to induce bacterial ferroptosis-like death has emerged as an attractive emerging nonantibiotic antimicrobial strategy to combat AMR.25 This review systematically summarizes the latest progress in this field and reveals the precise interventions for bacterial iron metabolism, ROS homeostasis and the antioxidant defense system, with the advantages of designability, multimechanism cooperation and strong targeting of nanomaterials. Thus, it triggers iron dependent, lipid peroxidation-driven programmed cell death, which is especially suitable for fighting multidrug-resistant bacterial infections. In the regulation of iron metabolism, nanomaterials can not only serve as efficient delivery carriers of iron ions but also promote the active uptake of iron by bacteria through surface functionalization to achieve iron hyperaccumulation. In terms of ROS generation, the synergistic action of chemical dynamics, photodynamics, sonodynamics and nanoenzyme catalysis significantly enhances the intensity of oxidative stress, overcoming the limitations of a single mechanism. In addition, by directly or indirectly depleting GSH and inhibiting the activities of antioxidant enzymes, nanomaterials effectively collapse the antioxidant defense system of bacteria, further promoting lipid peroxidation and cell membrane damage. Notably, nanomaterials can also regulate the host immune response to achieve the dual goals of synergistic immune sterilization and tissue repair.
 |
Figure 8 Evolution of treatment strategies for antibiotic resistance. (A) Compared with traditional antibacterial methods, which have been based mainly on antibiotics in the past, (B) with the current use of nanomaterials to induce bacterial ferroptosis-like death, nanomaterials face many challenges from basic research to clinical application. (C) Future advances will be achieved through AI-assisted interdisciplinary collaboration to build smart nanoplatforms to achieve the precise arrival and targeted aggregation of materials in the infectious microenvironment and synergize with immune functions. This cutting-edge antimicrobial strategy is expected to provide a breakthrough solution for overcoming drug resistance. |
Unlike the mechanism of action of traditional antibiotics, the current research progress is reflected mainly in Figure 8, reflecting the transition from “single sterilization” to “comprehensive regulation”. On the one hand, researchers have constructed a variety of composite nanoplatforms to implement kinetic therapy through various physical means. These nanomaterials can not only be specifically activated in the infection microenvironment to efficiently produce ROS but also enhance the synergy of physical mechanisms to achieve multiple antibacterial effects. On the other hand, the research horizon has expanded from direct sterilization to active remodeling of the host immune microenvironment. Both the recovery of neutrophil function and the polarization of the macrophage phenotype reflect the integration of “antibacterial, anti-inflammatory, and repair” agents in the process of inducing bacterial ferroptosis-like death by nanomaterials, the establishment of immunological memory opens new avenues for tackling chronic infections. Future research should prioritize the rational design of intelligent, microenvironment-responsive nanomaterials capable of greater spatiotemporal control. The integration of artificial intelligence design tools as well as interdisciplinary collaborations offers great potential for developing advanced multifunctional nanodrug delivery systems, facilitating therapeutic diagnostics and combination therapies.129,130 The integration of artificial intelligence in nanomaterial design and interdisciplinary collaboration will be crucial in accelerating the development of these sophisticated next-generation antimicrobial agents.
Despite promising prospects, this field still faces some challenges. The regulation of the complex in vivo environment is a key bottleneck. Current studies mostly verify the mechanism in vitro or in simplified animal models. Therefore, ensuring that nanomaterials can induce bacterial ferroptosis-like death without damaging surrounding normal tissues and cells is the premise of clinical transformation, which requires more detailed studies on the degradation metabolism, in vivo distribution and potential off-target effects of nanomaterials. Furthermore, the immunological mechanisms triggered by bacterial ferroptosis-like death have not been fully elucidated. A deeper investigation into their underlying molecular mechanisms is needed. Addressing these knowledge gaps will provide a theoretical foundation for the rational design of next-generation nanomaterials.
In summary, this novel antibacterial strategy, which is based on nanomaterial-induced bacterial ferroptosis-like death, fundamentally overcomes the limitations of traditional antibiotics, which rely on single targets. The key translational advantage lies in its ability to operate through multiple mechanisms and leverage the unique infection microenvironment for spatially controlled activation and targeted delivery, thereby minimizing off-target effects. This approach not only circumvents existing bacterial resistance mechanisms but also integrates potent bactericidal activity, immunomodulation, and tissue repair into a unified therapeutic framework. It holds particular promise for treating chronic infections, biofilm-associated infections, and patients with compromised immunity. With the ongoing optimization of intelligent nanoplatforms and advancements in preclinical studies, this strategy is poised to shift the anti-infective paradigm from mere “pathogen killing” to holistic “systemic regulation,” offering a transformative solution to the global crisis of antimicrobial resistance.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (82470594), National Natural Science Foundation of China (82573571), National Science and Technology Major Project of the Ministry of Science and Technology of China (2025ZD0545300), Shanghai 2025 Basic Research Plan Natural Science Foundation (25ZR1401393), and National Key Laboratory 2024 Annual Basic Medicine Innovation Open Project (JCKFKT-MS-006).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Browne AJ, Chipeta MG, Haines-Woodhouse G, et al. Global antibiotic consumption and usage in humans, 2000–18: a spatial modelling study. The Lancet Planetary Health. 2021;5(12):e893–24. doi:10.1016/S2542-5196(21)00280-1
2. Laxminarayan R, Matsoso P, Pant S, et al. Access to effective antimicrobials: a worldwide challenge. Lancet Lond Engl. 2016;387(10014):168–175. doi:10.1016/S0140-6736(15)00474-2
3. Conesa A, Garofolo G, Pasquale AD, Cammà C. Monitoring AMR in campylobacter jejuni from Italy in the last 10 years (2011–2021): microbiological and WGS data risk assessment. EFSA J. 2022;20:e200406. doi:10.2903/j.efsa.2022.e200406
4. Okeke IN, De Kraker MEA, Van Boeckel TP, et al. The scope of the antimicrobial resistance challenge. Lancet. 2024;403(10442):2426–2438. doi:10.1016/S0140-6736(24)00876-6
5. Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet Lond Engl. 2022;399(10325):629–655. doi:10.1016/S0140-6736(21)02724-0.
6. GBD 2019 Antimicrobial Resistance Collaborators. Global mortality associated with 33 bacterial pathogens in 2019: a systematic analysis for the global burden of disease study 2019. Lancet Lond Engl. 2022;400(10369):2221–2248. doi:10.1016/S0140-6736(22)02185-7.
7. Statista. World population by age and region 2024. Available from: https://www.statista.com/statistics/265759/world-population-by-age-and-region/.
8. Bodro M, Gudiol C, Garcia-Vidal C, et al. Epidemiology, antibiotic therapy and outcomes of bacteremia caused by drug-resistant ESKAPE pathogens in cancer patients. Support Care Cancer off J Multinatl Assoc Support Care Cancer. 2014;22(3):603–610. doi:10.1007/s00520-013-2012-3
9. Signing AT, Marbou WJT, Penlap Beng V, Kuete V. Antibiotic resistance profile of uropathogenic bacteria in diabetic patients at the bafoussam regional hospital, west Cameroon region. Cureus. 2020;12(7):e9345. doi:10.7759/cureus.9345
10. Nanayakkara AK, Boucher HW, Fowler VG, Jezek A, Outterson K, Greenberg DE. Antibiotic resistance in the patient with cancer: escalating challenges and paths forward. CA Cancer J Clin. 2021;71(6):488–504. doi:10.3322/caac.21697
11. Zheng Y, Chen Y, Yu K, et al. Fatal infections among cancer patients: a population-based study in the United States. Infect Dis Ther. 2021;10(2):871–895. doi:10.1007/s40121-021-00433-7
12. Teillant A, Gandra S, Barter D, Morgan DJ, Laxminarayan R. Potential burden of antibiotic resistance on surgery and cancer chemotherapy antibiotic prophylaxis in the USA: a literature review and modelling study. Lancet Infect Dis. 2015;15(12):1429–1437. doi:10.1016/S1473-3099(15)00270-4
13. Mallari P, Rostami LD, Alanko I, et al. The next frontier: unveiling novel approaches for combating multidrug-resistant bacteria. Pharm Res. 2025;42(6):859–889. doi:10.1007/s11095-025-03871-x
14. Y Lai, T Zhang, X Yin, et al. An antibiotic-free platform for eliminating persistent helicobacter pylori infection without disrupting gut microbiota. Acta Pharm Sin B. 2024;14(7):3184–3204. doi:10.1016/j.apsb.2024.03.014
15. Dolma S, Lessnick SL, Hahn WC, Stockwell BR. Identification of genotype-selective antitumor agents using synthetic lethal chemical screening in engineered human tumor cells. Cancer Cell. 2003;3(3):285–296. doi:10.1016/s1535-6108(03)00050-3
16. Yang WS, Stockwell BR. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells. Chem Biol. 2008;15(3):234–245. doi:10.1016/j.chembiol.2008.02.010
17. Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–1072. doi:10.1016/j.cell.2012.03.042
18. Su LJ, Zhang JH, Gomez H, et al. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid Med Cell Longevity. 2019;2019:5080843. doi:10.1155/2019/5080843
19. Xie Y, Hou W, Song X, et al. Ferroptosis: process and function. Cell Death Differ. 2016;23(3):369–379. doi:10.1038/cdd.2015.158
20. Wong-Ekkabut J, Xu Z, Triampo W, Tang IM, Tieleman DP, Monticelli L. Effect of lipid peroxidation on the properties of lipid bilayers: a molecular dynamics study. Biophys J. 2007;93(12):4225–4236. doi:10.1529/biophysj.107.112565
21. Kagan VE, Mao G, Qu F, et al. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat Chem Biol. 2017;13(1):81–90. doi:10.1038/nchembio.2238
22. Yang WS, Kim KJ, Gaschler MM, Patel M, Shchepinov MS, Stockwell BR. Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc Natl Acad Sci U S A. 2016;113(34):E4966–4975. doi:10.1073/pnas.1603244113
23. Stockwell BR. Ferroptosis turns 10: emerging mechanisms, physiological functions, and therapeutic applications. Cell. 2022;185(14):2401–2421. doi:10.1016/j.cell.2022.06.003
24. Jiang X, Stockwell BR, Conrad M. Ferroptosis: mechanisms, biology and role in disease. Nat Rev Mol Cell Biol. 2021;22(4):266–282. doi:10.1038/s41580-020-00324-8
25. Dai W, Shu R, Yang F, et al. Engineered bio-heterojunction confers extra- and intracellular bacterial ferroptosis and hunger-triggered cell protection for diabetic wound repair. Adv Mater. 2024;36(9):2305277. doi:10.1002/adma.202305277
26. Kwun MS, Lee DG. Ferroptosis-like death in microorganisms: a novel programmed cell death following lipid peroxidation. J Microbiol Biotechnol. 2023;33(8):992–997. doi:10.4014/jmb.2307.07002
27. Hong L, Chen X, Liu Y, Liang H, Zhao Y, Guo P. The relationship between ferroptosis and respiratory infectious diseases: a novel landscape for therapeutic approach. Front Immunol. 2025;16:1550968. doi:10.3389/fimmu.2025.1550968
28. Guo R, Fang X, Shang K, Wen J, Ding K. Induction of ferroptosis: a new strategy for the control of bacterial infections. Microbiol Res. 2024;284:127728. doi:10.1016/j.micres.2024.127728
29. Badran Z, Rahman B, De Bonfils P, Nun P, Coeffard V, Verron E. Antibacterial nanophotosensitizers in photodynamic therapy: an update. Drug Discovery Today. 2023;28(4):103493. doi:10.1016/j.drudis.2023.103493
30. Liu H, Nan Z, Zhao C, et al. Emerging synergistic strategies for enhanced antibacterial sonodynamic therapy: advances and prospects. Ultrason Sonochem. 2025;116:107288. doi:10.1016/j.ultsonch.2025.107288
31. Liu Q, Zou J, Chen Z, He W, Wu W. Current research trends of nanomedicines. Acta Pharm Sin B. 2023;13(11):4391–4416. doi:10.1016/j.apsb.2023.05.018
32. Yin X, Lai Y, Zhang X, et al. Targeted sonodynamic therapy platform for holistic integrative helicobacter pylori therapy. Appl Sci. 2024;12(2):2408583. doi:10.1002/advs.202408583
33. Wang Y, Yang Y, Shi Y, Song H, Yu C. Antibiotic‐free antibacterial strategies enabled by nanomaterials: progress and perspectives. Adv Mater. 2020;32(18):1904106. doi:10.1002/adma.201904106
34. Dixon SJ, Stockwell BR. The role of iron and reactive oxygen species in cell death. Nat Chem Biol. 2014;10(1):9–17. doi:10.1038/nchembio.1416
35. Feng S, Tang D, Wang Y, et al. The mechanism of ferroptosis and its related diseases. Mol Biomed. 2023;4(1):33. doi:10.1186/s43556-023-00142-2
36. Stockwell BR, Friedmann Angeli JP, Bayir H, et al. Ferroptosis: a regulated cell death nexus linking metabolism, redox biology, and disease. Cell. 2017;171(2):273–285. doi:10.1016/j.cell.2017.09.021
37. Conrad M, Pratt DA. The chemical basis of ferroptosis. Nat Chem Biol. 2019;15(12):1137–1147. doi:10.1038/s41589-019-0408-1
38. Ayala A, Muñoz MF, Argüelles S. Lipid peroxidation: production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid Med Cell Longevity. 2014;2014:360438. doi:10.1155/2014/360438
39. Catalá A. Lipid peroxidation of membrane phospholipids generates hydroxy-alkenals and oxidized phospholipids active in physiological and/or pathological conditions. Chem Phys Lipids. 2009;157(1):1–11. doi:10.1016/j.chemphyslip.2008.09.004
40. Gaschler MM, Stockwell BR. Lipid peroxidation in cell death. Biochem Biophys Res Commun. 2017;482(3):419–425. doi:10.1016/j.bbrc.2016.10.086
41. Alves F, Lane D, Nguyen TPM, Bush AI, Ayton S. In defence of ferroptosis. Signal Transduction Targeted Ther. 2025;10(1):2. doi:10.1038/s41392-024-02088-5
42. Shah R, Shchepinov MS, Pratt DA. Resolving the role of lipoxygenases in the initiation and execution of ferroptosis. ACS Cent Sci. 2018;4(3):387–396. doi:10.1021/acscentsci.7b00589
43. Wenzel SE, Tyurina YY, Zhao J, et al. PEBP1 wardens ferroptosis by enabling lipoxygenase generation of lipid death signals. Cell. 2017;171(3):628–641.e26. doi:10.1016/j.cell.2017.09.044
44. Galy B, Conrad M, Muckenthaler M. Mechanisms controlling cellular and systemic iron homeostasis. Nat Rev Mol Cell Biol. 2024;25(2):133–155. doi:10.1038/s41580-023-00648-1
45. Andrews NC, Schmidt PJ. Iron homeostasis. Annu Rev Physiol. 2007;69:69–85. doi:10.1146/annurev.physiol.69.031905.164337
46. Levina A, Lay PA. Transferrin cycle and clinical roles of citrate and ascorbate in improved iron metabolism. ACS Chem Biol. 2019;14(5):893–900. doi:10.1021/acschembio.8b01100
47. Chen X, Yu C, Kang R, Tang D. Iron metabolism in ferroptosis. Front Cell Dev Biol. 2020;8:590226. doi:10.3389/fcell.2020.590226
48. Bou-Abdallah F. The iron redox and hydrolysis chemistry of the ferritins. Biochim Biophys Acta. 2010;1800(8):719–731. doi:10.1016/j.bbagen.2010.03.021
49. Arosio P, Levi S. Cytosolic and mitochondrial ferritins in the regulation of cellular iron homeostasis and oxidative damage. Biochim Biophys Acta. 2010;1800(8):783–792. doi:10.1016/j.bbagen.2010.02.005
50. Jin L, Yu B, Wang H, et al. STING promotes ferroptosis through NCOA4-dependent ferritinophagy in acute kidney injury. Free Radic Biol Med. 2023;208:348–360. doi:10.1016/j.freeradbiomed.2023.08.025
51. Gryzik M, Asperti M, Denardo A, Arosio P, Poli M. NCOA4-mediated ferritinophagy promotes ferroptosis induced by erastin, but not by RSL3 in HeLa cells. Biochim Biophys Acta, Mol Cell Res. 2021;1868(2):118913. doi:10.1016/j.bbamcr.2020.118913
52. Li Y, Zeng X, Lu D, Yin M, Shan M, Gao Y. Erastin induces ferroptosis via ferroportin-mediated iron accumulation in endometriosis. Hum Reprod Oxf Eng. 2021;36(4):951–964. doi:10.1093/humrep/deaa363
53. Collins JF, Wessling-Resnick M, Knutson MD. Hepcidin regulation of iron transport. J Nutr. 2008;138(11):2284–2288. doi:10.3945/jn.108.096347
54. Donovan A, Lima CA, Pinkus JL, et al. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell Metab. 2005;1(3):191–200. doi:10.1016/j.cmet.2005.01.003
55. Wang Z, Li H, Zhou W, et al. Ferrous sulfate-loaded hydrogel cures staphylococcus aureus infection via facilitating a ferroptosis-like bacterial cell death in a mouse keratitis model. Biomaterials. 2022;290:121842. doi:10.1016/j.biomaterials.2022.121842
56. Dixon SJ, Winter GE, Musavi LS, et al. Human haploid cell genetics reveals roles for lipid metabolism genes in nonapoptotic cell death. ACS Chem Biol. 2015;10(7):1604–1609. doi:10.1021/acschembio.5b00245
57. Doll S, Proneth B, Tyurina YY, et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat Chem Biol. 2017;13(1):91–98. doi:10.1038/nchembio.2239
58. Yang X, Liu J, Wang C, et al. miR-18a promotes glioblastoma development by down-regulating ALOXE3-mediated ferroptotic and anti-migration activities. Oncogenesis. 2021;10(2):15. doi:10.1038/s41389-021-00304-3
59. Liang D, Feng Y, Zandkarimi F, et al. Ferroptosis surveillance independent of GPX4 and differentially regulated by sex hormones. Cell. 2023;186(13):2748–2764.e22. doi:10.1016/j.cell.2023.05.003
60. Magtanong L, Ko PJ, To M, et al. Exogenous monounsaturated fatty acids promote a ferroptosis-resistant cell state. Cell Chem Biol. 2019;26(3):420–432.e9. doi:10.1016/j.chembiol.2018.11.016
61. Zhang Q, Sun T, Yu F, et al. PAFAH2 suppresses synchronized ferroptosis to ameliorate acute kidney injury. Nat Chem Biol. 2024;20(7):835–846. doi:10.1038/s41589-023-01528-7
62. Sun WY, Tyurin VA, Mikulska-Ruminska K, et al. Phospholipase iPLA2β averts ferroptosis by eliminating a redox lipid death signal. Nat Chem Biol. 2021;17(4):465–476. doi:10.1038/s41589-020-00734-x
63. Conrad M, Kagan VE, Bayir H, et al. Regulation of lipid peroxidation and ferroptosis in diverse species. Genes Dev. 2018;32(9–10):602–619. doi:10.1101/gad.314674.118
64. Weaver K, Skouta R. The selenoprotein glutathione peroxidase 4: from molecular mechanisms to novel therapeutic opportunities. Biomedicines. 2022;10(4):891. doi:10.3390/biomedicines10040891
65. Bersuker K, Hendricks JM, Li Z, et al. The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis. Nature. 2019;575(7784):688–692. doi:10.1038/s41586-019-1705-2
66. Forcina GC, Dixon SJ. GPX4 at the crossroads of lipid homeostasis and ferroptosis. Proteomics. 2019;19(18):e1800311. doi:10.1002/pmic.201800311
67. Seibt TM, Proneth B, Conrad M. Role of GPX4 in ferroptosis and its pharmacological implication. Free Radic Biol Med. 2019;133:144–152. doi:10.1016/j.freeradbiomed.2018.09.014
68. Ingold I, Berndt C, Schmitt S, et al. Selenium utilization by GPX4 is required to prevent hydroperoxide-induced ferroptosis. Cell. 2018;172(3):409–422.e21. doi:10.1016/j.cell.2017.11.048
69. Geng N, Shi BJ, Li SL, et al. Knockdown of ferroportin accelerates erastin-induced ferroptosis in neuroblastoma cells. Eur Rev Med Pharmacol Sci. 2018;22(12):3826–3836. doi:10.26355/eurrev_201806_15267
70. Yang WS, SriRamaratnam R, Welsch ME, et al. Regulation of ferroptotic cancer cell death by GPX4. Cell. 2014;156(1–2):317–331. doi:10.1016/j.cell.2013.12.010
71. Doll S, Freitas FP, Shah R, et al. FSP1 is a glutathione-independent ferroptosis suppressor. Nature. 2019;575(7784):693–698. doi:10.1038/s41586-019-1707-0
72. Wu KC, Cui JY, Klaassen CD. Beneficial role of Nrf2 in regulating NADPH generation and consumption. Toxicol Sci. 2011;123(2):590–600. doi:10.1093/toxsci/kfr183
73. Anandhan A, Dodson M, Schmidlin CJ, Liu P, Zhang DD. Breakdown of an ironclad defense system: the critical role of NRF2 in mediating ferroptosis. Cell Chem Biol. 2020;27(4):436–447. doi:10.1016/j.chembiol.2020.03.011
74. Jiang L, Kon N, Li T, et al. Ferroptosis as a p53-mediated activity during tumour suppression. Nature. 2015;520(7545):57–62. doi:10.1038/nature14344
75. Kho CW, Lee PY, Bae KH, et al. Glutathione peroxidase 3 of saccharomyces cerevisiae regulates the activity of methionine sulfoxide reductase in a redox state-dependent way. Biochem Biophys Res Commun. 2006;348(1):25–35. doi:10.1016/j.bbrc.2006.06.067
76. Manfredini V, Roehrs R, Peralba MCR, et al. Glutathione peroxidase induction protects Saccharomyces cerevisiae sod1deltasod2delta double mutants against oxidative damage. Braz J Med Biol Res. 2004;37(2):159–165. doi:10.1590/s0100-879x2004000200001
77. Wang H, Guo J, Yang Y, et al. CuFeS2 nanozyme regulating ROS/GSH redox induces ferroptosis-like death in bacteria for robust anti-infection therapy. Mater Des. 2024;239:112809. doi:10.1016/j.matdes.2024.112809
78. Latunde-Dada GO. Ferroptosis: role of lipid peroxidation, iron and ferritinophagy. Biochim Biophys Acta Gen Subj. 2017;1861(8):1893–1900. doi:10.1016/j.bbagen.2017.05.019
79. Cao JY, Dixon SJ. Mechanisms of ferroptosis. Cell Mol Life Sci CMLS. 2016;73(11–12):2195–2209. doi:10.1007/s00018-016-2194-1
80. Zhao X, Drlica K. Reactive oxygen species and the bacterial response to lethal stress. Curr Opin Microbiol. 2014;21:1–6. doi:10.1016/j.mib.2014.06.008
81. Yang WS, Stockwell BR. Ferroptosis: death by lipid peroxidation. Trends Cell Biol. 2016;26(3):165–176. doi:10.1016/j.tcb.2015.10.014
82. Tanouchi Y, Lee AJ, Meredith H, You L. Programmed cell death in bacteria and implications for antibiotic therapy. Trends Microbiol. 2013;21(6):265–270. doi:10.1016/j.tim.2013.04.001
83. Cheng B, Li D, Li C, Zhuang Z, Wang P, Liu G. The application of biomedicine in chemodynamic therapy: from material design to improved strategies. Bioeng. 2023;10(8):925. doi:10.3390/bioengineering10080925
84. Chen S, Huang F, Mao L, et al. High Fe-loading single-atom catalyst boosts ROS production by density effect for efficient antibacterial therapy. Nano Micro Lett. 2024;17(1):32. doi:10.1007/s40820-024-01522-1
85. Mourenza Á, Gil JA, Mateos LM, Letek M. Oxidative stress-generating antimicrobials, a novel strategy to overcome antibacterial resistance. Antioxid. 2020;9(5):361. doi:10.3390/antiox9050361
86. Zhu L, You Y, Zhu M, et al. Ferritin-hijacking nanoparticles spatiotemporally directing endogenous ferroptosis for synergistic anticancer therapy. Adv Mater. 2022;34(51):e2207174. doi:10.1002/adma.202207174
87. Li C, Hua C, Chu C, et al. A photothermal-responsive multi-enzyme nanoprobe for ROS amplification and glutathione depletion to enhance ferroptosis. Biosens Bioelectron. 2025;278:117384. doi:10.1016/j.bios.2025.117384
88. Zhang H, Wang Y, Jiang M, et al. Inherently anti-metastatic peptide hydrogels for sonodynamic-amplified ferroptosis in cancer therapy. Mater Today Bio. 2025;32:101688. doi:10.1016/j.mtbio.2025.101688
89. Jin Y, Zhang Y, Xue C, Du S, Yao J. Phototriggered biofilm nanodisruptor with genetic modulation for treating drug-resistant bacterial infections. ACS Appl Mater Interfaces. 2025;17(23):34370–34386. doi:10.1021/acsami.5c03333
90. Shao DH, Yao YF, Wang DN. Ferroptosis and iron-based therapies in pseudomonas aeruginosa infections: from pathogenesis to treatment. Virulence. 2025;16(1):2553787. doi:10.1080/21505594.2025.2553787
91. Zhou Y, Cai CY, Wang C, et al. Ferric-loaded lipid nanoparticles inducing ferroptosis-like cell death for antibacterial wound healing. Drug Delivery. 2023;30(1):1–8. doi:10.1080/10717544.2022.2152134
92. Zhao X, Wang Y, Zhu T, et al. Mesoporous calcium-silicate nanoparticles loaded with prussian blue promotes enterococcus faecalis ferroptosis-like death by regulating bacterial redox pathway ROS/GSH. Int J Nanomed. 2022;17:5187–5205. doi:10.2147/IJN.S382928
93. Yu Z, Fu X, Lucas T, et al. MOF-enhanced phototherapeutic wound dressings against drug-resistant bacteria. Adv Healthcare Mater. 2025;14(1):e2402418. doi:10.1002/adhm.202402418
94. Xu Z, Xu Z, Gu J, et al. In situ formation of ferrous sulfide in glycyrrhizic acid hydrogels to promote healing of multi-drug resistant staphylococcus aureus-infected diabetic wounds. J Colloid Interface Sci. 2023;650(Pt B):1918–1929. doi:10.1016/j.jcis.2023.07.141
95. Wang L, Li G, Hui A, et al. Palygorskite-based antibacterial composites induced intracellular S. aureus ferroptosis through SSTR2. Int J Biol Macromol. 2025;311(Pt 3):143897. doi:10.1016/j.ijbiomac.2025.143897
96. Shao W-B, Luo R-S, Huang Y-W, et al. Construction of phenothiazine-decorated ZnO quantum dots with intelligent response to bacterial pH/amidase microenvironment for inducing bacterial ferroptosis-like death. Chem Eng J. 2025;509:161352. doi:10.1016/j.cej.2025.161352
97. Sun J, Shen Q-J, Pan J-N, Zheng X, Yu T, Zhou -W-W. Ferrous sulfate combined with ultrasound emulsified cinnamaldehyde nanoemulsion to cause ferroptosis in escherichia coli O157:H7. Ultrason Sonochem. 2024;106:106884. doi:10.1016/j.ultsonch.2024.106884
98. Xue Y, Zhang L, Liu F, et al. Alkaline “nanoswords” coordinate ferroptosis-like bacterial death for antibiosis and osseointegration. ACS Nano. 2023;17(3):2711–2724. doi:10.1021/acsnano.2c10960
99. fa YH, Zou T, zhang TQ, et al. Ferroptosis: mechanisms and links with diseases. Signal Transduction Targeted Ther. 2021;6(1):49. doi:10.1038/s41392-020-00428-9
100. Zhang T, Zheng Y, Chen T, et al. Biomaterials mediated 3R (remove-remodel-repair) strategy: holistic management of helicobacter pylori infection. J Nanobiotechnol. 2025;23(1):475. doi:10.1186/s12951-025-03455-2
101. Sun C, Jiang Y, Zhang S, et al. A tailored artificial biocatalyst for bacterial endophthalmitis therapy via enhanced ferroptosis‐like death. Adv Sci. 2025;12(33):e04601. doi:10.1002/advs.202504601
102. Zheng F, Wan X, Zhang Y, et al. A multimodal defect-rich nanoreactor triggers sono-piezoelectric tandem catalysis and iron metabolism disruption for implant infections. Sci Adv. 2025;11(11):eads8694. doi:10.1126/sciadv.ads8694
103. Wang Z, Huang Y, He S, et al. Oxygen-independent sulfate radical and Fe2+-modified implants for fast sterilization and osseointegration of infectious bone defects. ACS Nano. 2025;19(19):18804–18823. doi:10.1021/acsnano.5c04147
104. Wu T, Han F, Mei J, et al. Photoactive metal-covalent organic framework nanozymes with enhanced peroxidase-mimicking activity for eliminating drug-resistant bacterial infections. J Colloid Interface Sci. 2025;699(Pt 1):138178. doi:10.1016/j.jcis.2025.138178
105. Sun B, Wang X, Ye Z, et al. Designing single-atom active sites on sp2 -carbon linked covalent organic frameworks to induce bacterial ferroptosis-like for robust anti-infection therapy. Adv Sci Weinh Baden-Wurtt Ger. 2023;10(13):e2207507. doi:10.1002/advs.202207507
106. Lu R, Luo Z, Zhang Y, Chen J, Zhang Y, Zhang C. A multifunctional tissue-engineering hydrogel aimed to regulate bacterial ferroptosis-like death and overcoming infection toward bone remodeling. Adv Sci Weinh Baden-Wurtt Ger. 2024;11(30):e2309820. doi:10.1002/advs.202309820
107. Li Z, Lu Y, Song J, et al. An emodin-mediated multifunctional nanoplatform with augmented sonodynamic and immunoregulation for osteomyelitis therapy. J Colloid Interface Sci. 2025;684(Pt 2):122–137. doi:10.1016/j.jcis.2025.01.094
108. An X, Chen B, Yuan Y, et al. Laser-ultrasound responsive nanoreactor to remodel the microenvironment of antibiotic-resistant bacterial infection for diabetic wound healing. Nano Lett. 2025. doi:10.1021/acs.nanolett.5c02548
109. Sun M, Wang L, Zhuo Y, et al. Multi-enzyme activity of MIL-101 (Fe)-derived cascade nano-enzymes for antitumor and antimicrobial therapy. Small Weinh Bergstr Ger. 2024;20(17):e2309593. doi:10.1002/smll.202309593
110. Yin Y, Guo W, Chen Q, et al. A single H2S-releasing nanozyme for comprehensive diabetic wound healing through multistep intervention. ACS Appl Mater Interfaces. 2025;17(12):18134–18149. doi:10.1021/acsami.5c00889
111. You W, Cai Z, Xiao F, et al. Biomolecular microneedle initiates Fe3O4/MXene heterojunction-mediated nanozyme-like reactions and bacterial ferroptosis to repair diabetic wounds. Adv Sci Weinh Baden-Wurtt Ger. 2025;12(11):e2417314. doi:10.1002/advs.202417314
112. Wang X, Liu T, Chen M, et al. An erythrocyte-templated iron single-atom nanozyme for wound healing. Adv Sci Weinh Baden-Wurtt Ger. 2024;11(6):e2307844. doi:10.1002/advs.202307844
113. Fan Z, Liang C, Zhang J, et al. Multimodal synergistic strategies for diabetic wound healing using glucose oxidase nanocomposites: therapeutic mechanisms and nanomaterial design. Int J Nanomed. 2025;20:5727–5762. doi:10.2147/IJN.S515057
114. Dawi J, Tumanyan K, Tomas K, et al. Diabetic foot ulcers: pathophysiology, immune dysregulation, and emerging therapeutic strategies. Biomedicines. 2025;13(5):1076. doi:10.3390/biomedicines13051076
115. Antelmann H. Oxidative stress responses and redox signalling mechanisms in bacillus subtilis and staphylococcus aureus. In: Tang YW, Sussman M, Liu D, Poxton I, Schwartzman J, editors. Molecular Medical Microbiology.
116. Acierno C, Barletta F, Nevola R, et al. Metabolic rewiring of bacterial pathogens in response to antibiotic pressure-a molecular perspective. Int J Mol Sci. 2025;26(12):5574. doi:10.3390/ijms26125574
117. Shen X, Ma R, Huang Y, et al. Nano-decocted ferrous polysulfide coordinates ferroptosis-like death in bacteria for anti-infection therapy. Nano Today. 2020;35:100981. doi:10.1016/j.nantod.2020.100981
118. Hu H, Hua SY, Lin X, et al. Hybrid biomimetic membrane coated particles-mediated bacterial ferroptosis for acute MRSA pneumonia. ACS Nano. 2023;17(12):11692–11712. doi:10.1021/acsnano.3c02365
119. Yan L, Wang J, Cai X, et al. Macrophage plasticity: signaling pathways, tissue repair, and regeneration. Medcomm. 2024;5(8):e658. doi:10.1002/mco2.658
120. Xu B, Liu Y, Li N, Geng Q. Lactate and lactylation in macrophage metabolic reprogramming: current progress and outstanding issues. Front Immunol. 2024;15:1395786. doi:10.3389/fimmu.2024.1395786
121. Ding H, Liu R, Ruan H, et al. Rapidly gelling sprayable hydrogels integrating carbon quantum dots and plant extracellular vesicles enable multifunctional wound care via photothermal management, immunomodulation, and ferroptosis. Small Weinh Bergstr Ger. 2025;21(35):e2503202. doi:10.1002/smll.202503202
122. Ding T, Zhang L, Chen J, Ma D, Han J, Han Y. “thermal bubbles”: photothermally triggered by a carbon monoxide nanocontainer for antibiosis and immune modulation therapy. Nano Today. 2025;63:102758. doi:10.1016/j.nantod.2025.102758
123. Zhou J, Zhang L, Wei Y, et al. Photothermal iron-based riboflavin microneedles for the treatment of bacterial keratitis via ion therapy and immunomodulation. Adv Healthcare Mater. 2024;13(26):e2304448. doi:10.1002/adhm.202304448
124. Chen M, Liu T, Wang X, et al. Comprehensive wound healing using ETN@Fe7S8 complex by positively regulating multiple programmed phases. J Nanobiotechnol. 2025;23(1):342. doi:10.1186/s12951-025-03396-w
125. Geng S, Hao P, Wang D, et al. Zinc oxide nanoparticles have biphasic roles on mycobacterium-induced inflammation by activating autophagy and ferroptosis mechanisms in infected macrophages. Microbiol Pathogen. 2023;180:106132. doi:10.1016/j.micpath.2023.106132
126. Zhu W, Mei J, Zhang X, et al. Photothermal Nanozyme-Based Microneedle Patch against Refractory Bacterial Biofilm Infection via Iron-Actuated Janus Ion Therapy. Adv Mater. 2022;34(51):e2207961. doi:10.1002/adma.202207961
127. Sirit IS, Peek RM. Decoding the ability of helicobacter pylori to evade immune recognition and cause disease. Cell Mol Gastroenterol Hepatol. 2025;19(5):101470. doi:10.1016/j.jcmgh.2025.101470
128. Makvandi P, Song H, Yiu CKY, et al. Bioengineered materials with selective antimicrobial toxicity in biomedicine. Mil Med Res. 2023;10(1):8. doi:10.1186/s40779-023-00443-1
129. Liu GY, Yu D, Fan MM, et al. Antimicrobial resistance crisis: could artificial intelligence be the solution? Mil Med Res. 2024;11(1):7. doi:10.1186/s40779-024-00510-1
130. He W, Wu X, Jin Z, et al. Generative artificial intelligence in medical imaging: current landscape, challenges, and future directions. Interdisciplinary Med. 2025;2025:e20250024. doi:10.1002/INMD.20250024
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Cu-Cy Nps@ZIF-8@HA-Mediated Photodynamic Therapy for Cutaneous Squamous Cell Carcinoma
Li N, Wan X, Hou X, Zhang J, Zhang C, Sun Y, Wang F, Kong S, Yang C, Jiang G
International Journal of Nanomedicine 2026, 21:560049
Published Date: 8 January 2026