Back to Journals » International Journal of Nanomedicine » Volume 13
Nano-selenium and its nanomedicine applications: a critical review
Authors Hosnedlova B ![]() , Kepinska M
, Kepinska M ![]() , Skalickova S
, Skalickova S ![]() , Fernandez C, Ruttkay-Nedecky B, Peng Q, Baron M, Melcova M
, Fernandez C, Ruttkay-Nedecky B, Peng Q, Baron M, Melcova M ![]() , Opatrilova R
, Opatrilova R ![]() , Zidkova J, Bjørklund G
, Zidkova J, Bjørklund G ![]() , Sochor J, Kizek R
, Sochor J, Kizek R
Received 21 November 2017
Accepted for publication 12 January 2018
Published 10 April 2018 Volume 2018:13 Pages 2107—2128
DOI https://doi.org/10.2147/IJN.S157541
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Thomas Webster
Bozena Hosnedlova,1 Marta Kepinska,2 Sylvie Skalickova,3 Carlos Fernandez,4 Branislav Ruttkay-Nedecky,3 Qiuming Peng,5 Mojmir Baron,1 Magdalena Melcova,6 Radka Opatrilova,3 Jarmila Zidkova,6 Geir Bjørklund,7 Jiri Sochor,1 Rene Kizek2,3
1Department of Viticulture and Enology, Faculty of Horticulture, Mendel University in Brno, Lednice, Czech Republic; 2Department of Biomedical and Environmental Analyses, Faculty of Pharmacy, Wroclaw Medical University, Wroclaw, Poland; 3Central Laboratory, Faculty of Pharmacy, University of Veterinary and Pharmaceutical Sciences Brno, Brno, Czech Republic; 4School of Pharmacy and Life Sciences, Robert Gordon University, Aberdeen, UK; 5State Key Laboratory of Metastable Materials Science and Technology, Yanshan University, Qinhuangdao, People’s Republic of China; 6Department of Biochemistry and Microbiology, University of Chemistry and Technology, Prague, Czech Republic; 7Council for Nutritional and Environmental Medicine, Rana, Norway
Abstract: Traditional supplements of selenium generally have a low degree of absorption and increased toxicity. Therefore, it is imperative to develop innovative systems as transporters of selenium compounds, which would raise the bioavailability of this element and allow its controlled release in the organism. Nanoscale selenium has attracted a great interest as a food additive especially in individuals with selenium deficiency, but also as a therapeutic agent without significant side effects in medicine. This review is focused on the incorporation of nanotechnological applications, in particular exploring the possibilities of a more effective way of administration, especially in selenium-deficient organisms. In addition, this review summarizes the survey of knowledge on selenium nanoparticles, their biological effects in the organism, advantages, absorption mechanisms, and nanotechnological applications for peroral administration.
Keywords: nanoparticles, biomedicine, drug delivery, oxidative stress, anticancer effect, antimicrobial activity, protective effect
Methodology of the review
To carry out this study, different databases such as Web of Science, PubMed, MEDLINE, and Google Scholar were employed. The findings of research studies (original articles, experimental studies) chosen from more than 1,000 viewed scientific publications were compared. The emphasis was put on original articles, while more than 100 review articles were not included. The search was based on phrases such as selenium nanoparticles and antioxidant activity/antimicrobial properties/antiviral effect/antibacterial activity/anticancer agent, selenium nanoparticles and synthesis, nano-selenium and bioavailability/toxicity/safety/drug delivery/orthopedic implants/protective effects/oxidative stress/metal intoxication/immunomodulation/reproduction/growth/gastrointestinal tract/hair, selenium nanoparticles and animals, advantages of nanoparticles, and effects of nano-selenium. The review includes research findings from the years 2000–2017.
Nanotechnological modifications
Nanoparticle (NP) systems appear to be a promising alternative to peroral drug delivery1 as well as nutritional supplements.2 The effects of bioactive compound supplements – omega-3 and omega-6 fatty acids, probiotics, prebiotics, vitamins, and minerals – in nanoparticulate preparations have already been dealt by numerous studies.3–16 Some applications of NPs in nutrition and medicine have already been approved for clinical use, and many others are at different stages of their development.17
The incentive to incorporate nanotechnology in nutrition has the following advantages: taste and smell, administration and solubility, protection against oxidation and enzymatic degradation, prolongation of residence time, and effective passage through the gastrointestinal tract enhancing the bioavailability of supplemented substances.18 The use of most perorally administered supplements carries a number of drawbacks such as insufficient residence time, low permeability and solubility, and instability during the production process (temperature, presence of oxygen, light) or in the gastrointestinal tract environment (pH, enzymes, presence of other nutrients), which can reduce the activity and potential health benefits of supplements.19 Thus, it is necessary to develop more appropriate products to ensure their physiological and therapeutic effect.20
The use of NPs is a promising alternative for the peroral application of supplements that could solve the mentioned shortcomings of traditional forms of supplements and preserve their health benefits.1 The main advantages of NPs are shown in Figure 1.
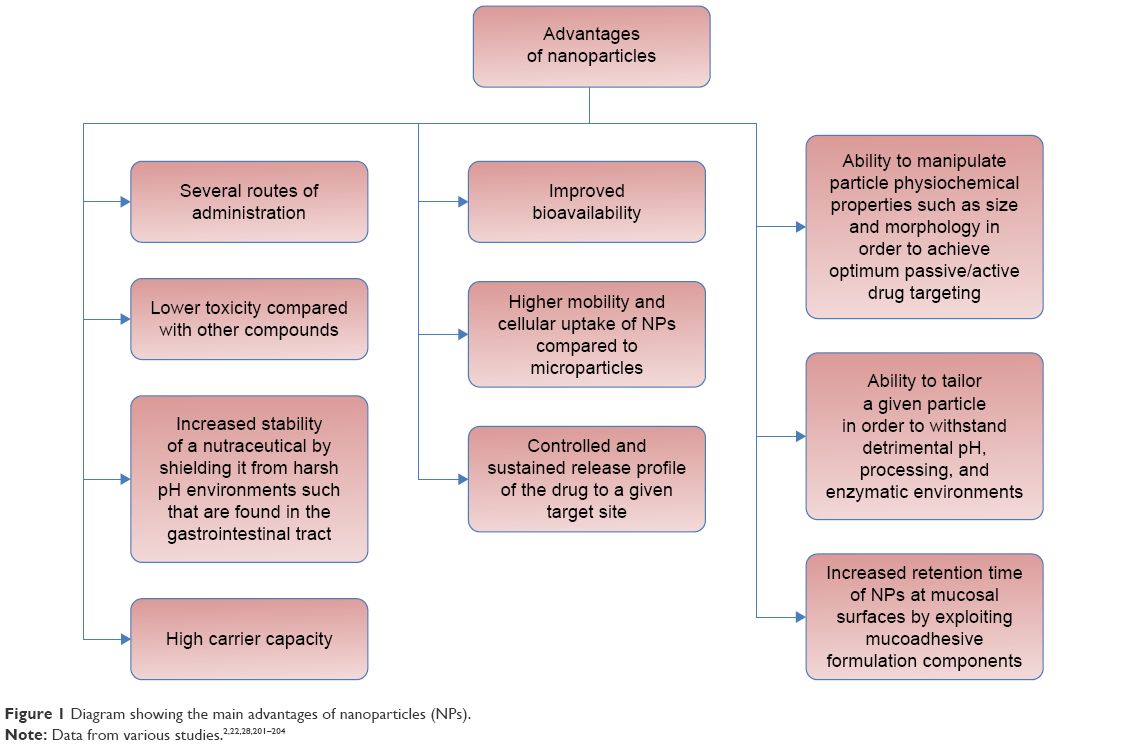 | Figure 1 Diagram showing the main advantages of nanoparticles (NPs). |
Selenium NPs as a food additive
The nanoform of selenium attracts even more attention, thanks to its high bioavailability and lower toxicity than inorganic and organic forms,21–23 where inorganic compounds are more toxic than organic ones.24
The biological properties of selenium nanoparticles (SeNPs) depend on their size: smaller particles have a greater activity.25 Particle size affects the cellular intake of NPs; for example, in vitro absorption of 0.1 μm particles was found to be 2.5 and 6 times higher compared to 1 and 10 μm particles, respectively.26 With respect to this fact, in the preparation of dietary supplements, an appropriate particle size and morphology, as well as encapsulation material, should be chosen.27 A system based on the administration of food supplements encapsulated into NPs represents an important potential to improve the bioavailability of supplements with the possibility of modifying their properties, such as resistance to adverse pH, digestion, and enzymatic cleavage.28
The advantage of nano-selenium (Nano-Se) is the possibility of using selenium in zero oxidation state (Se0), which presents low toxicity and excellent bioavailability compared to other oxidation states (Se+IV, Se+VI);22,25 however, it is very unstable and easily transformed into an inactive form. Although, its stabilization can be achieved by encapsulation into suitable nano-vehicles, for example, chitosan (CS)15 (Figure 2).
 | Figure 2 Scheme of preparation of selenium nanoparticles (SeNPs). |
Nanoscale selenium has a very wide range of biomedical applications. Its effect on the reduction of oxidative stress is very well known.12,29 Gao et al30 demonstrated the antioxidant properties of hollow spherical SeNPs reducing the risk of selenium toxicity. Besides its use as an antioxidant with reduced risk of toxicity, Nano-Se also possesses potential as a chemopreventive agent.22 The results of numerous studies indicate that Nano-Se can be more helpful in cancer chemoprevention as a potential anticancer drug,9,31–34 as well as an anticancer drug delivery carrier.32,35,36 Many studies have shown the antimicrobial effect37–39 and antifungal activity of Nano-Se.40 In addition, its protective effects against metal intoxication were well documented.41–43 Moreover, the immunostimulatory effect of nanoscale selenium was confirmed.8,44 Last but not least, Nano-Se has beneficial effects on a number of physiological functions.13,45–47
Besides these unique abilities of Nano-Se, its antiprotozoal effect was described. Based on in vitro and in vivo studies, biogenic SeNPs can be considered as a novel therapeutic agent for the treatment of localized lesions typical of cutaneous leishmaniasis caused by Leishmania major.48 The antileishmanial activities of SeNPs against Leishmania infantum were also described. SeNPs have more growth-inhibitory effect on promastigotes than selenium dioxide (SeO2), while the IC50 (half minimal [50%] inhibitory concentration) was determined to be 25 and 50 μg·mL−1, respectively.49
The main effects of the SeNPs found in various scientific studies are summarized in Figure 3.
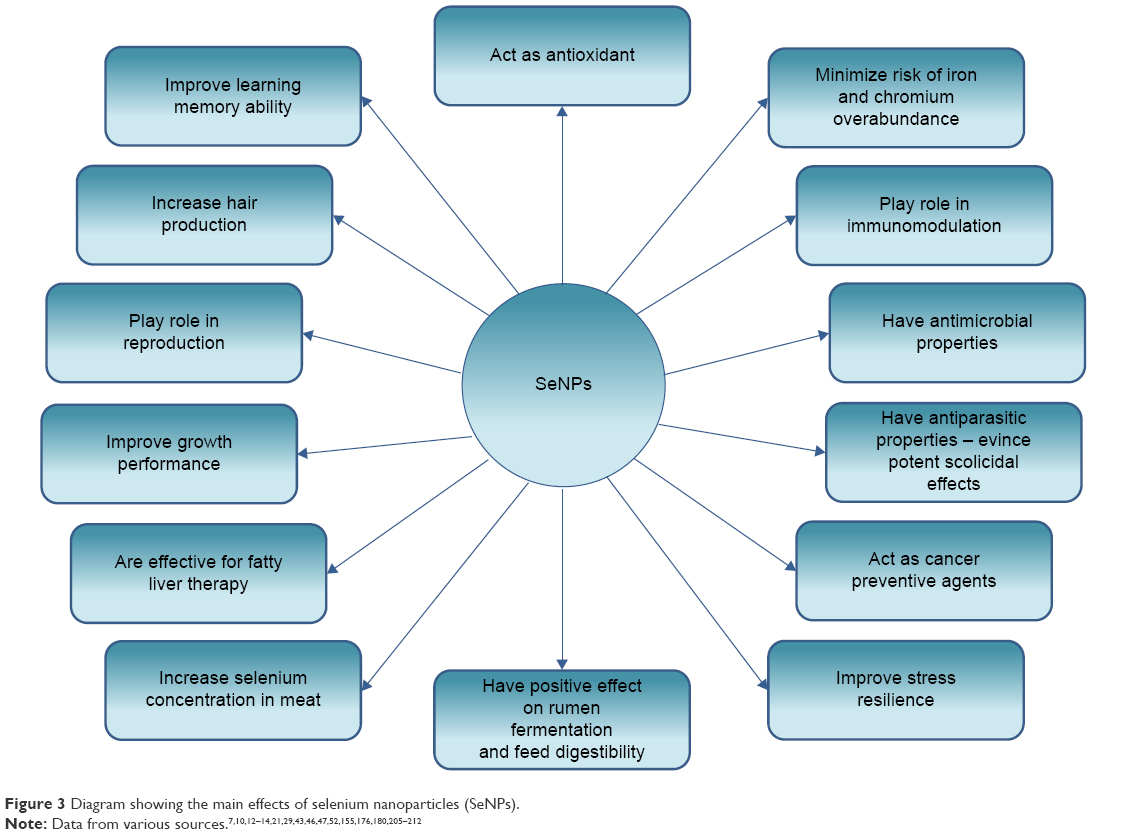 | Figure 3 Diagram showing the main effects of selenium nanoparticles (SeNPs). |
Synthesis of SeNPs
SeNPs can be synthesized chemically,50 or using physical procedures,51 or can even be obtained by biological way – using microorganisms or plant extracts, the so-called green synthesis.52,53
Chemical synthesis
In terms of chemical synthesis, SeNPs are usually prepared by reduction of selenious acid solution by ascorbic acid in the presence of polysaccharides (Figure 2) such as CS, glucomannan, acacia gum, or carboxymethylcellulose.50 CS is positively charged, biocompatible, non-immunogenic, nontoxic, pH sensitive, and biodegradable, and is therefore a suitable component for oral administration for a wide range of biomedical and nutritional applications.54–56 It has been extensively examined in the pharmaceutical industry due to its potential in the development of medication delivery systems.57 In the molecular structure of polysaccharides, reactive amino, hydroxyl, or carboxyl groups are present, which have a substantial effect on the formation, stabilization, and growth of SeNPs.50 After the synthesis of NPs, their ex situ characterization is performed using different biophysical methods including electron microscopy. The obtained monodisperse spherical selenium particles are very stable in solution16 and can be used as dietary additive.14
The effects of selenium encapsulation into CS NPs were investigated by Zhang et al.2 The study showed that encapsulation of selenium compounds into CS NPs is an effective way of delivering selenium to cells in which selenium retention increases, while reducing the risk of DNA damage. The development of NP–selenium compound systems may, in the case of low selenium level, significantly improve the bioavailability of selenium and facilitate the expression of selenoproteins.
Another example for chemical preparation of SeNPs is ionic liquid-induced synthesis with sodium selenosulfate as selenium precursor, in the presence of polyvinyl alcohol stabilizer, which can produce spherical SeNPs in the size range of 76–150 nm.58
Physical synthesis
Among the physical techniques for the preparation of SeNPs, synthesis by pulsed laser ablation (PLA) or deposition was described in a study.51 Generally, physical methods have distinct advantages over the chemical ones, since these often require a final calcination step, which makes them unsuitable for certain applications. Furthermore, with sputtering and laser ablation, the stoichiometry of the material is maintained.59 In PLA, the size of the nanoclusters can be controlled by laser parameters, such as fluence, wavelength, and pulse duration, as well as by ambient gas conditions, such as pressure and flow parameters.60 However, physical techniques are not widely applied for producing SeNPs.
Biological synthesis
Originally, SeNPs, like other NPs, were synthesized by various chemical methods. However, the high cost of production and the presence of toxic by-products led to the development of novel methods of NPs synthesis.61 Biological organisms such as plants, fungi, or bacteria have the ability to convert some toxic metal ions to less toxic forms including metal precipitants or NPs.62–64 Thanks to this advantage, researches focused on their use for synthesizing NPs with an ecofriendly approach.65–68
As for green synthesis, SeNPs were synthesized, for example, using the following plant extracts: aqueous extract of Allium sativum,69 tea extract,70 Clausena dentata plant leaf extract,71 Undaria pinnatifida polysaccharide solutions,31 and Terminalia arjuna leaf extract.42
Biosynthesis of nanomaterials using plant extracts has more advantages than other biological methods because it is inexpensive and does not require any special conditions.52
NPs synthesis using bacteria is more effective than chemical synthesis, thanks to the following advantages: 1) high purity of selenium spheres (which are relatively regular and uniform, and their size depends on the bacterium), 2) cheaper and faster production process, and 3) better possibility to control the parameters.72
Microorganisms are capable of synthesizing metal NPs.53 Different bacterial strains are able to reduce Se+IV (selenite) and (or) Se+VI (selenate) to less toxic Se0 with the formation of SeNPs. The biogenic SeNPs have exhibited promising application perspectives in the field of medicine, biosensors, and environmental remediation.73 Interestingly, two halotolerant Bacillus megaterium strains (BSB6 and BSB12), isolated from saline mangrove habitat without selenium contamination, were found to be capable of reducing Se+IV to elemental selenium, even in the presence of high salt concentrations.74
The synthesis of SeNPs by macro- or microorganisms, due to the diversity of reducing enzymes in organisms, involves morphological and shape changes of the particles. By changing the redox state, the reducing enzymes of microorganisms convert metal ions (Se−II) to SeNPs without charge (Se0). The biological activity of SeNPs includes their protective role against DNA oxidation.53 It was found that certain anaerobic bacteria respire toxic selenium oxyanions and as a result cause extracellular accumulation of elemental selenium (Se0). The spectral properties differ considerably from those of amorphous Se0 formed by chemical oxidation of hydrogen selenide (H2Se) and of black, vitreous Se0 formed chemically by reduction of selenite with ascorbate. The microbial synthesis of Se0 nanospheres results in unique, complex, with compact nanostructural arrangement of Se atoms. This probably reflects a wide diversity of enzymes involved in the dissimilatory reduction that are slightly different in various bacteria. Remarkably, these conditions cannot be achieved by current methods of chemical synthesis.66
Different types of bacteria have been used for the biosynthesis of SeNPs, such as the species of phylum Proteobacteria (Escherichia coli ATCC 35218,75 recombinant E. coli,76 Ralstonia eutropha,77 Enterobacter cloacae Z0206,78 Pseudomonas aeruginosa ATCC 27853,79 Klebsiella pneumoniae,80,81 Pantoea agglomerans,25 Zooglea ramigera,82 Rhodopseudomonas palustris strain N,83 Shewanella sp. HN-41,84 Azoarcus sp. CIB,85 Burkholderia fungorum,86 Stenotrophomonas maltophilia39), Firmicutes (Lactobacillus casei,72,81,87 Lactobacillus acidophilus [LA-5],72 Lactobacillus helveticus [LH-B02],72 Enterococcus faecalis,53 Streptococcus thermophilus,72 Staphylococcus carnosus,88 Bacillus sp. MSh-1,89 Bacillus subtilis,65 Bacillus mycoides SelTE01,38 Bacillus licheniformis JS290), Actinobacteria (Streptomyces sp. ES2-5,73 Bifidobacterium BB-1272), and Cyanobacteria (Arthrospira [Spirulina] platensis91).
For in vivo synthesis of Nano-Se, unicellular eukaryotic organisms, such as the protozoa Tetrahymena thermophila SB210,92 yeast Saccharomyces cerevisiae,93 and a genetically modified Pichia pastoris,94 or even multicellular organisms, such as a fungus from the phylum of Ascomycota (Aspergillus terreus95), were used.
Problems with traditional forms of oral supplementation of selenium and potential benefits of SeNPs
The bioavailability of various chemical forms of selenium has been the subject of many studies. For instance, Mahan et al stated that the degree of selenium absorption is much lower in ruminants than in nonruminants; after oral supplementation, the selenium uptake in sheep was only 34%, whereas in pigs 85%.96 In ruminants, the microbial digestion in the rumen and reticulum precedes its assimilation in the abomasum and microbial digestion in the small intestine.97,98 Ruminal microorganisms partially transform selenium supplements into insoluble forms, such as elemental selenium and selenides, which are unavailable to animals, and thus, the absorption of selenium from the digestive tract is reduced.99
To avoid this negative phenomenon of reduced selenium absorption due to microbial digestion in ruminants and improve the availability of this microelement, polymer NP systems with in vivo potential have been proposed.
This technology was implemented using the Eudragit® RL and Eudragit® RS polymers.100 These nonbiodegradable polymer NPs were already used in 2005 to encapsulate the lipophilic cyclic undecapeptide cyclosporin A (CyA) for oral administration in rabbits for the prevention of graft rejection and the treatment of autoimmune diseases. CyA was encapsulated by nanoprecipitation in mentioned NP polymers, and its relative bioavailability from both preparations ranged from 20% to 35%.101 The design of selenium in the nanoform for oral administration to ruminants was proposed as late as 2010 by Romero-Pérez et al,100 who monitored the in vitro effect of sodium selenite (Na2SeO3) encapsulated in polymeric NPs by nanoprecipitation and emulsion-evaporation methods using two different solvents – ethanol and acetone. Nanoprecipitated NPs were spherical and had a greater particle size variability; on the contrary, the NPs produced by the emulsion-evaporation technique were both spherical and irregular in shape and homogeneous in size. The size of NPs ranged from 30 to 200 nm. Higher particle size, zeta potential, and polydispersity index were found for the NPs produced by the nanoprecipitation method using ethanol as the solvent of polymers. The release of selenium from NPs was monitored in vitro at different pH values and was higher in the strongly acidic environment (pH less than 4), which is a prerequisite for providing better accessibility of this element in the abomasum. At pH values lower than 4, selenium release increases by 62%100 compared to pH 6. The pH in the rumen ranges from 5.5 to 6.5. Thus, the sheep rumen essentially does not absorb selenium.97 On the contrary, the pH in the abomasum is less than 4; under this condition, the largest amount of selenium is released from the particles. NPs increase the bioavailability of this element because at pH 6 only a small amount of selenium is released (by rumen microorganisms) and the remaining, bigger part, is released then in the more acidic environment of the gastrointestinal tract, which guarantees its better availability.100
SeNPs evince, in addition to better selenium availability, much lower toxicity compared to other selenium compounds.22,23 For example, in mice, SeNPs showed much lower toxicity measured by median lethal dose (50%, LD50), liver impairment, and short-term toxicity.23
Mechanism of passage of NPs through intestinal mucosa
Oral administration of NPs is considered to be the most appropriate and cost-effective method of supplementation. However, absorption of NPs may also be made more difficult by the presence of absorption barriers in the digestive tract. It is necessary to overcome two barriers in the gastrointestinal tract for the absorption of NPs, which are mucus covering the intestinal mucosa and the intestinal mucosa.1
The intestinal epithelium is composed of a series of specialized cells, primarily enterocytes, goblet cells, and M cells. One of the main functions of enterocytes is to control the transition of macromolecules and allow the absorption of nutrients. The goblet cells secrete mucus consisting of a high-molecular-weight glycoprotein (mainly mucin) suspended in the electrolyte solution,102 which covers with the adherent layer the mucous membrane of the intestine. Their primary function is to protect the intestinal mucosa from potential pathogens or chemicals and maintain a different pH between the lumen and the mucosal surface of the intestine.1
NPs can theoretically pass through the intestinal epithelium in two ways: paracellular (between adjacent cells) or transcellular (through the cells) (Figure 4).102 Under physiological conditions, the first way is restricted by a narrow region of intercellular spaces and by the tightness of the junctions between the epithelial cells (pore diameter is between 0.3 and 1.0 nm).103 Transcellular transport of NPs takes place through a process called transcytosis, which starts with endocytosis in the apical membrane of the cells. Subsequently, the NPs are transported through the cells and released on the basolateral pole.104
 | Figure 4 Diagram of nanoparticle transport across the intestinal mucosa of the small intestine. Particles pass either paracellularly, that is, between adjacent cells, or by a transcellular pathway that has been more explored. The transcellular transition takes place via normal enterocytes or M cells. A, paracellular transport; B, transcellular transport: transcytosis through M cell; C1, transcytosis through enterocyte, first mechanism; C2, transcytosis through enterocyte, second mechanism; 1, apical membrane of enterocyte; 2, the basolateral membrane of the enterocyte. M cells form a row of vesicles before transfer from subepithelial dome dendritic cells (SDDCs) to T- or B-lymphocytes (B). Uptake of nanoparticles by microvilli of enterocytes is often followed by endosome formation, microvesicular bodies (MVBs) genesis, and their fusion with lysosomes, and then the particles are transported to lamina propria (C1). Another mechanism of enterocyte utilization for transepithelial transport involves uptake into endosomes, MVB formation, fusion with Golgi apparatus, and exosomal transfer to lamina propria (C2). This is followed by transport into the bloodstream and the lymphatic system.102,214 |
Intestinal epithelial cells are capable of transferring NPs with mineral elements, although their capacity is limited. Transcellular transport begins with endocytosis (pinocytosis or macropinocytosis). It is an active process requiring energy for the internalization of NPs. Macropinocytosis is actin-dependent but is not regulated by receptors, and a large amount of fluid with particles smaller than 5 μm can be internalized through it.105 The absorption of NPs is influenced by electric charge, surface hydrophobicity, and size.106 The epithelial cell membrane of the gastrointestinal tract is composed of lipids, so that hydrophobic NPs have a higher absorption efficiency than hydrophilic particles. The absorption of NPs of 100 nm in the gastrointestinal tract is 15–250 times higher than that of larger-sized NPs.107
Application of SeNPs through oral administration
Nano-Se as an antioxidant
Nano-Se possesses better antioxidant capability than other chemical forms of selenium while reducing the risk of selenium toxicity. Wang et al22 demonstrated the antioxidant properties of SeNPs that evinced lower toxicity than selenomethionine (SeMet).
Zhang et al108 studied the effect of elemental Nano-Se on the activity of glutathione peroxidase (GPx) in the liver of weanling pigs (Duroc × Landrace × Yorkshire) in comparison to inorganic form of selenium. The animals had significantly higher activities of GPx at a concentration range of 0.50 and 1.0 mg Se·kg−1 diet in the form of Nano-Se than Na2SeO3.
Effect of SeNPs on reproductive performance
Oxidative stress affects the fertility potential of spermatozoa by lipid peroxidation which can lead to sperm dysfunction.109 Selenium deficiency leads to the occurrence of abnormal mitochondria in goat spermatozoa, and Nano-Se supplementation increases selenium concentration in testes, and testicular and semen GPx activity. In addition, it has a protective effect on membrane integrity and the tight arrayment of the midpiece of the mitochondria. To study the effects of elemental Nano-Se in the diet on the testes ultrastructure, ejaculate quality and GPx activity in goats were investigated. The animals were administered with Nano-Se in the declared selenium dose of 0.3 mg·kg−1 of the feed dry matter (DM) for 12 weeks from weaning to sexual maturity. The results showed that the level of selenium in testes and the activity of GPx and ATPase in the ejaculate significantly increased in the group supplemented with nanoscale selenium compared to the control group. The quality of the ejaculate (volume, density, motility, and pH) was not affected by the addition of selenium, but the percentage of abnormal spermatozoa in the control group of goats was significantly higher compared to the SeNPs group. Using transmission electron microscopy, it was found that in selenium-deficient goats the sperm plasma membrane was damaged, and there were abnormalities in the mitochondria midpiece of spermatozoa.13
Use of Nano-Se for increasing hair follicle development and fetal growth
The study of Wu et al46 demonstrated the importance of maternal administration of selenium at nano size for improving the hair follicle development and promoting the growth of fetus. This was attributed to the influencing antioxidant status in the fetal skin. An increase in the antioxidant defence results in a decrease of reactive oxygen species (ROS) generation, leading to upregulation of IGF-1 and its receptor (IGF-1R) which are crucial for the improvement of both properties.
In cashmere goats, SeNPs have a positive effect on the amount of wool. The administration of 0.5 mg·kg−1 diet (containing 1,500 mg Se·kg−1) in the period from 30 days before conception to 110 days of gestation was manifested in their fetuses with significantly higher expression levels of the GPx, IGF-1, and IGF-1R genes in skin. In addition, increased GPx and superoxide dismutase (SOD) activities, and IGF-1 and selenium concentrations in both skin and blood serum at 110 days were observed. Moreover, a significantly lower malondialdehyde (MDA) production in both skin and serum which affected the antioxidant status in skin of the fetuses, and significantly increased the number of their secondary hair follicles, was observed. The low level of ROS probably upregulates IGF-1 and IGF-1R, which favorably influences the development of hair follicles in the fetus. In addition, the weights of fetus and placenta were significantly higher in SeNPs-receiving group than those in control group.46
Antiviral and antibacterial effects of SeNPs
SeNPs have attracted substantial attention due to their unique antimicrobial activity.53,110–114 Selenium is an essential trace element regulated by cellular redox homeostasis115–117 and is an integral component of selenoproteins controlling some crucial biological processes, such as ROS elimination and specific enzyme modulation.118,119 Selenium deficiency can result in susceptibility to viral infections. The antiviral capability of SeNPs, together with other advantages such as low toxicity and excellent activity, has recently attracted increased attention. The mortality of the H1N1 influenza virus-infected selenium-deficient mice was 3 times higher compared to those receiving Na2SeO3 at the dose of 0.5 mg Se·kg−1, and mice with low serum selenium concentrations showed a marked reduction in body weight (BW) and lower levels of TNF-α and IFN-γ.120 For improving the immune response in the body, the administration of SeNPs can also be an efficient realizable approach.
Li et al121 synthesized oseltamivir (OTV) surface-modified SeNPs (Se@OTV) with superior antiviral properties and restriction on drug resistance. Although the clinical application of OTV itself as an effective antiviral agent is ordinarily limited by the appearance of drug-resistant viruses, OTV decoration of SeNPs evidently inhibited H1N1 influenza virus infection and showed less toxicity. Se@OTV interfered with the entry of H1N1 into host cells through inhibiting the activity of influenza virus glycoproteins – hemagglutinin and neuraminidase. Modified NPs were able to prevent H1N1 from infecting Madin-Darby canine kidney cells and block chromatin condensation and DNA fragmentation. In addition, they inhibited the generation of ROS as well as the activation of phosphorylation of cellular tumor antigen p53 and Akt. Thus, ROS play a pivotal role in the antiviral action. The presence of H1N1 virus increased the intracellular ROS generation from 100% (control) to 380%, and OTV and SeNPs slightly inhibited their production to 270% and 210%, respectively, whereas Se@OTV markedly decreased the ROS generation (120%). It follows that Se@OTV are promising efficient antiviral pharmaceuticals against highly infectious respiratory disease caused by H1N1,121 which belongs to influenza A-type viruses.122
SeNPs also show excellent antibacterial activity. With SeNPs biosynthesized by R. eutropha, 99% inhibition of growth of P. aeruginosa, S. aureus, E. coli, and Streptococcus pyogenes at the concentrations of 100, 100, 250, and 100 μg·mL−1, respectively, was observed. Moreover, the concentration of 500 μg·mL−1 of SeNPs was found to inhibit the growth of pathogenic fungi Aspergillus clavatus. The antimicrobial efficacy of SeNPs can be comparable with commercially available antibiotic ampicillin.77 SeNPs synthesized by another bacterial species E. faecalis can be used as an antistaphylococcal element to effectively prevent and treat S. aureus infections.53 SeNPs with application of bioactive glass scaffolds (45S5Bioglass®) and poly(lactic-co-glycolic acid) (PLGA) (45S5Bioglass®/SeNPs and 45S5Bioglass®/PLGA/SeNPs) showed a considerable antibacterial activity against Gram-positive bacteria, S. aureus and Staphylococcus epidermidis.114 In another study, Yang et al123 introduced the antibacterial effect of Qe/CdSe/ZnS (quercetin/cadmium selenide/zinc sulfide) nanoparticles (QCZNPs) against drug-resistant E. coli and B. subtilis in vitro. QCZNPs showed markedly more effective antibacterial activities than Qe or CdSe NPs. The in vitro study of Wang and Webster37 showed that the selenium coatings on polycarbonate medical devices significantly inhibited S. aureus growth to 27% compared with an uncoated polycarbonate surface after 72 hours.
The antimicrobial activity of SeNPs depends on the way they are synthesized. Piacenza et al38 evaluated the antimicrobial efficacy of spherical biogenic selenium nanostructures embedded in organic material produced by B. mycoides SelTE01 in comparison with two different chemical SeNP classes produced using L-cysteine or ascorbic acid. The antimicrobial activity was tested on P. aeruginosa and S. aureus biofilms grown onto hydroxyapatite-coated clinical devices and surfaces. After 6 or 24 hours of Na2SeO3 exposure, biogenic SeNPs evinced the same effective antibiofilm activity against both tested strains at 0.078 and 0.3125 mg·mL−1, respectively. On the contrary, chemically synthesized SeNPs at the highest tested concentration (2.5 mg·mL−1) showed only moderate antimicrobial activity. Cremonini et al39 reported that SeNPs synthesized by Gram-negative S. maltophilia and Gram-positive B. mycoides achieved much stronger antimicrobial effects compared with synthetically prepared SeNPs. Both biogenic types of NPs were active at low minimum inhibitory concentrations against a number of clinical isolates of P. aeruginosa. It was interesting that after an exposure of human dendritic cells and fibroblasts to all three SeNPs, the following phenomena were not observed: 1) loss of cell viability, 2) increased release of ROS, and 3) significant increase in the secretion of pro-inflammatory and immunostimulatory cytokines. Therefore, SeNPs appear to be reliable candidates for safe medical applications, either alone or in combination with traditional antibiotics, to inhibit the growth of P. aeruginosa clinical isolates.
Anticancer effects of SeNPs
SeNPs have also showed remarkable anticancer activity52,123–128 and exhibit high potential in cancer chemotherapy and as drug carriers.129–131 The anticancer effects of SeNPs are mediated through their ability to inhibit the growth of cancer cells through induction of cell cycle arrest at S phase.32 The induction of the cell cycle arrest at the S phase is mediated by deregulation of the eIF3 protein complex.132 A recent study revealed that cell membrane plays an important role in SeNPs-induced toxicity in cancer cells. SeNPs treatment changes the biomechanical properties of cancer cells, in particular remarkably decreases the adhesion force and Young’s modulus.133 Besides unique anticancer efficacy, SeNPs have been proved to present a better selectivity between normal and cancer cells than Se+IV at similar concentrations.31 SeNPs can be internalized selectively by cancer cells through endocytosis and induce cell apoptosis by triggering apoptotic signal transduction pathways.134–136
Nano-Se as an anticancer drug
Nano-Se possesses higher anticancer efficacy than other selenium compounds.119 A key mechanism for the chemopreventive effect is the induction of glutathione S-transferase (GST) by selenium.22 The activity of GST in the case of Nano-Se administration increased much earlier and more markedly than in the case of SeMet and selenite.22,137
Yanhua et al127 suggested that novel selenium-substituted hydroxyapatite NPs (SeHAN) could be a new promising anticancer agent to provide both survival advantage and lower toxicity in nude mice model of human hepatocellular carcinoma. The overall survival rate of nude mice in the control, pure hydroxyapatite, and SeHAN groups was 50.00%, 76.92%, and 100.00%, respectively. Blood biochemical studies showed that SeHAN group had significantly lower toxicity on the liver and kidney functions.
In highly metastatic breast cancer mice model, a better prognosis could be achieved by oral administration of SeNP-enriched Lactobacillus brevis. Both components are immunostimulators, and enhanced immune response in cancer-affected mice. Moreover, lactic acid bacteria (LAB) can reduce selenium ions to elemental SeNPs and deposit them in intracellular spaces. When LAB in combination with SeNPs were administered to cancer-bearing mice, a significant increase in natural killer cell cytotoxicity and delayed-type hypersensitivity responses, as well as a high level of IFN-γ and IL-17, compared to the control mice or mice that received L. brevis alone, was observed. In addition, an extended life span and a decline in the tumor metastasis to the liver were recorded in this group compared to the other two groups of mice.44
Chen et al31 reported that SeNPs fabricated in U. pinnatifida polysaccharide solutions induced mitochondria-mediated apoptosis in A375 human melanoma cells. Treatment of this cancer cell line with Nano-Se resulted in a dose-dependent cell apoptosis manifested by DNA fragmentation and phosphatidylserine translocation.
Sonkusre et al34 introduced biologically synthesized SeNPs by B. licheniformis JS2 and developed a method for extraction and purification of intracellular NPs. These neutral-charged, non-agglomerating SeNPs at a concentration as low as 2 μg Se·mL−1 were efficacious in inhibiting proliferation and inducing caspase-independent necrosis to human prostate adenocarcinoma cells (PC3) without causing any significant toxicity to human peripheral blood mononuclear cells.
Luo et al32 examined the in vitro antiproliferative effects of SeNPs (Nano-Se0, 10–40 μmol·L−1) on HeLa (human cervical carcinoma) cells and MDA-MB-231 (human breast carcinoma) cells by optical microscopic inspection and MTT assay. Nano-Se0 effectively inhibited the growth of cells of both the cancer cell lines in a dose-dependent manner. The morphology analysis with atomic force microscopy showed that the HeLa cells treated with 10 μmol·L−1 Nano-Se0 were rough and shrunken with truncated lamellipodia at the terminal part of the cells. Flow cytometric analysis demonstrated that HeLa cells were arrested at S phase of the cell cycle after exposed to mentioned amount of Nano-Se0 (10 μmol·L−1).
It was found that anticancer activity of SeNPs correlates with the level of ERα in breast cancer cells. SeNP-induced cell death and expression of apoptotic markers (pp38, Bax, and cytochrome c) were significantly higher in ERα-positive cells (MCF-7) but not in ERα-negative cells (MDA-MB-231).33
Nano-Se as an anticancer drug delivery carrier
Besides their direct anticancer effects, SeNPs have been appointed as potential anticancer drug delivery carriers.32,35 A key factor that usually contributes to nanomaterial-based drug cytotoxicity is cellular uptake.35 The nanosize of these materials allows an efficient uptake by various cell types and selective drug accumulation at target sites.26 Like other chemotherapeutics, the effective cytotoxicity of nanomaterial-based drugs usually requires a high level of accumulation within the cancer cells.138 Nanomaterials tend to accumulate in cancer cells through a passive targeting process128 and often serve as “nanocarriers” for chemotherapeutics.35,139–147 Moreover, the usage of various surface decorators enhances the cellular uptake and anticancer efficacy of nanomaterials;35,128,135 for example, Yang et al128 developed a simple and solution-phase method for functionalization of SeNPs with Spirulina polysaccharides (SPS). In addition, they found that SPS surface decoration significantly enhanced the cellular uptake and cytotoxicity of SeNPs against several cancer cell lines. The action mechanism of SPS-SeNPs was grounded in inhibition of cancer cell growth through induction of apoptosis, as evidenced by an increase in sub-G1 cell population, DNA fragmentation, chromatin condensation, and phosphatidylserine translocation. In another study, Yang et al148 designed and synthesized folic acid-conjugated SeNPs (FA@SeNPs) as cancer-targeting agents; the anticancer efficacy of FA@SeNPs was synergistically enhanced by radioactive 125I seeds, and these NPs inhibited colony formation ability, which showed that functionalized SeNPs can be used as a radiation sensitizer for 125I seeds for cancer therapy.
Chan et al129 reported that, based on the Auger-electron effect and Compton effect of Se atoms, cancer-targeted SeNPs in combination with 125I seeds achieve synergetic effects for inhibition of cancer cell growth and colony formation by inducing cell apoptosis and cell cycle arrest. In addition, these could be used as a safe and effective agent for next-generation cancer chemoradiotherapy in clinical applications. The action mechanism showed the activation of intracellular ROS overproduction to regulate p53-mediated DNA damage apoptotic signaling pathways and MAPKs phosphorylation as well as preventing the self-repair of cancer cells simultaneously.
SeNPs can be used as a carrier of 5-fluorouracil to achieve anticancer synergism, as introduced by Liu et al.35 A panel of five human cancer cell lines (A375, MCF-7, HepG2, Colo201, and PC3) evinced susceptibility to 5-fluorouracil surface-functionalized SeNPs (5-FU-SeNPs), with IC50 values ranging from 6.2 to 14.4 μM. Remarkably, despite this potency, the 5-FU-SeNPs possess great selectivity between cancer and normal cells. Induction of apoptosis in A375 human melanoma cells by 5-FU-SeNPs was evidenced by accumulation of sub-G1 cell population, DNA fragmentation, and nuclear condensation.
By comparing the effect of chemically prepared SeNPs on the behavior of cancer cells with that of other inorganic and organic selenospecies, similar alterations in terms of cell viability, proliferation, migration, and cell cycle arrest at the S-G2/M phase after exposure of hepatocarcinoma (HepG2) cells to SeNPs and selenocystine (SeCys2) were observed. In contrast, cells exposed to Se+IV showed evident signs of toxicity such as strong induction of apoptosis and a significant population of cells in the sub-G1 phase compared to control cells. Se+IV, SeMet, and seleno-methylselenocysteine (Se-MetSeCys) did not evince any significant differences in comparison to non-treated cells. While SeNPs only partially inhibited the Cdk1 expression, Se+IV and SeCys2 reduced drastically its expression, which can be toxic for healthy cells. Cdk1 plays an essential role in cell cycle progression and mitosis entry, and its inhibition induces cell cycle arrest. These findings may pave the way for use of Cdk1-targeting SeNPs in mitotic cell death for cancer therapy.36
Nano-Se as a promising orthopedic implant material and an agent reducing bone cancer cell functions
Currently used metallic orthopedic implants possess certain problems such as poor prolonged osseointegration, stress shielding,149 and wear debris-associated bone cell death.150 However, the greatest concern is corrosion as a result of continuous tissue exposure to metal. This limits orthopedic implant efficacy, especially in patients receiving implants due to bone cancer.151 Unfortunately, current orthopedic materials are not designed to prevent the occurrence or reoccurrence of cancer.150 Selenium, due to its chemopreventive and chemotherapeutic properties,150,152–154 appears to be a promising anticancer biocompatible orthopedic implant material.151 Moreover, SeNPs may be used to effectively prevent and treat S. aureus155 and S. epidermidis114 infections, which are one of the leading causes of implant failure.155 In addition, selenium did not evince inhibitory effect on the osteoblastic cell growth.156 Perla and Webster151 reported increased osteoblast adhesion on particulate surfaces of the compacts made from selenium compared with nonparticulate wrought titanium sheets. Moreover, osteoblast density was further increased on the surfaces of the selenium compacts with nanometer particles. Tran et al150 fabricated titanium orthopedic material coated with a high-density selenium nanoclusters-doped surface. This novel biomaterial inhibited, compared to traditional untreated titanium, cancerous bone cell proliferation while promoting healthy bone cell functions (including adhesion, proliferation, alkaline phosphatase activity, and calcium deposition).
SeNPs precipitated on a common orthopedic tissue engineering material poly-L-lactic acid evinced, without simultaneous use of chemotherapeutics or pharmaceutical agents, selectively decreased long-term osteosarcoma cell density while promoting healthy, noncancerous, osteoblast functions (for instance, up to 2 times higher alkaline phosphatase activity on selenium-coated tissue culture plates compared to osteoblasts grown on typical tissue culture plates), which is an impetus for follow-up studies to replace tumorous bone tissue with healthy bone tissue.157
Effect of SeNPs on oxidative stress parameters
The effect of Nano-Se on oxidative stress parameters was compared with the effect of organically bound selenium in a study. Nano-Se was found to have a comparable efficiency in increasing plasma GPx activity in mice as SeMet but exhibited much lower toxicity assessed on the basis of LD50, acute liver injury, and short-term toxicity. The results of the study showed that the nanoscale selenium can be administered as an antioxidant with a reduced risk of selenium toxicity.22 Upregulation of selenoenzymes by elemental Nano-Se is also comparable to the effect of selenite and Se-MetSeCys, again with a significant reduction of acute toxicity.23,158,159
When comparing the effect of SeNPs (red selenium) (orally 1 mg·kg−1 BW) and inorganic selenium (Na2SeO3) (orally 1 mg Se·kg−1 BW) on antioxidant activities of neutrophils and hematological parameters in sheep for 30 days, it was found that on the 30th day thiobarbituric acid-reactive substance levels were significantly higher in both groups than in control animals, in contrary to the expected decrease in their levels. There were no significant differences between the packed cell volume and red blood cell count between the experimental groups and control group. The white blood cell count in the Nano-Se group showed a significant increase on the 20th and 30th days, and in the Na2SeO3 group on day 20, compared to the control group. There was also a significant increase in neutrophil count and a significant decrease in lymphocyte count on day 10 in the Nano-Se group compared to the second experimental group and the control group, and on the 20th and 30th days in both the experimental groups compared to the control group.12
The effect of SeNPs on heat shock proteins (HSPs) and HSP90 gene expression as additional oxidative stress parameters was investigated. Increased oxygen metabolism induces the formation of ROS.160 Intensive training of trotter horses can lead to oxidative stress, the formation of ROS, and consequently lipid, protein, and DNA damage.161 In addition to adaptive changes in protective enzymes (SOD, catalase [CAT],162 GPx163), oxidative stress to cells is known to induce increased production of stress or HSPs.162 Expression of HSPs is an adaptive mechanism against the disruption of cellular homeostasis164 and integrity165 during physical exercise.
A study of the effect of oral administration of SeNPs at 0.5 mg·kg−1 concentration for 10 days on the expression of genes encoding HSP90 during intensive training in donkeys was performed.160
To assess the expression of the HSP90 gene in gluteus medius muscle, the total RNA was amplified by semiquantitative reverse transcription polymerase chain reaction. The results showed that serum concentration of selenium increased and the expression of the HSP90 gene decreased during rest after workload in the control group. As a response to intense exercise, in the experimental group with the 10-day administration of the SeNPs, both HSP90 expression and serum selenium concentration significantly increased. The induction of HSP resulting from SeNPs administration protected the cells from otherwise lethal stress levels. The result may explain the positive effect of short-term oral supplementation of SeNPs to donkeys on cell stability under stress conditions such as intensive training.160 Excessive dose of the antioxidants in combination with exercise may result in increased oxidative stress and impair exercise-induced adaptation, including direct induction of HSPs.166 Kojouri et al160 showed that oral supplementation of SeNPs in donkeys increased the expression of HSP90 in the gluteal muscle after 24 hours of rest compared to the control group. Expression of HSP90 induced by supplementation of SeNPs can alter the buffering capacity of the cells in response to stressful conditions and protects them from further damage. The epigenetic effect of SeNPs and mechanism of their action on gene expression remains misunderstood.
In addition, the effect of SeNPs on blood urea nitrogen (BUN) in donkeys was investigated. It is known that urea in high concentrations causes oxidative stress and DNA lesions in cells.167 A study of the effect of oral administration of SeNPs on changes in BUN, creatinine, and total protein during intensive training in donkeys revealed that serum selenium concentration was significantly increased after SeNPs supplementation.29 The creatinine concentration in the experimental and control groups was significantly increased in 2 hours of rest after training and rapidly declined in 72 hours of rest after workout in the experimental group. A similar pattern was obtained with changes in BUN in the control group: its concentration was significantly increased in 2 hours of rest after training compared to the group which was dosed with SeNPs. These findings can explain the positive effects of SeNPs supplementation on serum changes in BUN levels and creatinine in response to intense training of donkeys. The positive effect of SeNPs could be related to the incorporation of selenium into proteins, such as selenocysteine (SeCys) and its preventive role in oxidative tissue damage.29
Protective effects of Nano-Se
SeNPs in prevention of cisplatin (CIS)-induced reproductive toxicity
CIS, an anticancer alkylating agent,168 is widely used for cancer treatment.127,169–172 Despite its abundant clinical use, CIS possesses many side effects. It is known that CIS induces DNA adducts173 and forms DNA cross-links168 which interfere with cellular metabolism, such as DNA replication and transcription, triggering cell death.
Rezvanfar et al168 demonstrated that Nano-Se can be, due to its strong antioxidant potential, useful to prevent CIS-induced gonadotoxicity. Coadministration of SeNPs significantly improved the serum testosterone, sperm quality, and spermatogenesis and reduced CIS-induced free radical toxic stress and spermatic DNA damage in male rats.
Protective effect of Nano-Se against polycyclic aromatic hydrocarbons
In mice exposed to oxidative stress induced by polycyclic aromatic hydrocarbon – 7,12-dimethylbenz(a)anthracene (DMBA), a known immunotoxin and carcinogen,174,175 which were fed with lamb meat supplemented with selenium in the nanoform, the protective effect of Nano-Se against DMBA-induced immunotoxicity was observed. Compared to the control group of mice, also exposed to DMBA, survival of 2 times higher amount of leucocytes (of which 3 times higher amount of phagocytes) was found, and their recovery in the bone marrow was twice higher, and the regenerative capacity of granulopoiesis was 4 times higher. The study showed the functional dietary benefits of lamb meat enriched with selenium obtained by feeding lambs SeNPs.176
Use of SeNPs for minimization of risk of iron overabundance
An in vivo study compared the effect of SeNPs and Na2SeO3 (both were administered at a dose of 1 mg·kg−1 BW for 30 days) on iron homeostasis and expression of genes coding for transferrin and transferrin receptor in sheep.7 The study showed that at the beginning of the administration selenium decreased the serum iron concentration and increased the total iron-binding capacity (TIBC). In addition, after 30 days, a reduction of the iron level in blood serum was observed. The group of animals dosed with SeNPs showed a significantly increased TIBC level after 20 days compared to the Na2SeO3 and control groups, and significantly reduced iron serum level in comparison to the control group. At the beginning, the selenium administration stimulated the expression of the transferrin gene as well as the transferrin receptor gene, but at the end it suppressed their expression. This particular phenomenon was especially observed in animals that received Na2SeO3. This effect could be related to hypoferremia and cellular internalization of iron. Expression of both genes was significantly increased in the SeNPs group after 20 days compared to the Na2SeO3 group.
SeNPs in treatment of heavy metal intoxication
SeNPs were found to be a promising agent to check the chronic toxicity caused due to heavy metals exposure. Prasad and Selvaraj42 studied the effect of SeNPs, synthesized using T. arjuna leaf extract, on human lymphocytes treated with arsenite (As+III). Studies on cell viability using MTT assay and DNA damage using comet assay revealed protective effect of SeNPs against As+III-induced cell death and DNA damage. This approach could be used in future for minimizing arsenic-induced ROS-mediated toxic hazard, especially in an area with arsenic-contaminated groundwater and prevalence of arsenicosis.
The protective ability of SeNPs was also found against hexavalent chromium-induced thyrotoxicity.43 Toxic effect as a result of oxidative damage provoked by an intraperitoneal administration of single dose of potassium dichromate (K2Cr2O7; 60 μg·kg−1 BW) to rats was manifested by significant decrease in free T3 (triiodothyronine) and T4 (thyroxine) and glutathione levels, and significant increases in CAT, SOD, and MDA in the chromium-treated group compared to the controls. SeNPs administration resulted in correcting the hormonal levels as well as oxidative stress biomarkers compared to the K2Cr2O7-treated group.
Selenium also evinces protective effect against cadmium-induced nephrotoxicity.41 Two-week oral administration of Na2SeO3 to rats treated with repeated intraperitoneal injection of Cd at sublethal dose (1.50 mg Cd·kg−1 BW for 14 days) reduced lipid peroxidation and restored GPx and SOD activities in the kidneys. Selenium supplementation facilitated renal Cd accumulation in this group compared to Cd-exposed rats without selenium intake. Interestingly, X-ray diffraction analysis carried out on kidney fractions revealed CdSe and/or cadmium sulfide (CdS) NPs (about 62 nm in size). This indicates that Cd may induce the biosynthesis of red fluorescent CdSe and CdS NPs in the kidneys. The reduction of Cd-induced renal toxicity through selenium administration seems to lie in its ability to bind Cd in nanosized insoluble and fluorescent complexes. This implies that Cd complexation with Se or S at a nanoscale level could reduce oxidative stress induced by cadmium in the kidneys.
Nano-Se as an immunostimulator
Chronic oxidative stress reduces the respiratory burst response of neutrophils.177 Nano-Se, possessing an immunostimulating potential,44 evinces, in comparison with Na2SeO3, a stronger and faster effect in supporting the antioxidant defense system, based on increased chemotactic activity and respiratory burst activities of neutrophils.8 The effect of nanoscale selenium and Na2SeO3 (both supplements were administered at 1 mg·kg−1 BW, per os for 30 days) on neutrophils’ characteristics in sheep was compared. To determine the chemotactic activity and respiratory burst activities of the neutrophils, the leading front assay and the nitroblue tetrazolium (NBT) test (it is based on the reduction of the oxidant substances inside the phagocytic cells by NBT revealed by color change) were carried out on heparinized blood samples. It was found that in the Nano-Se group and the Na2SeO3 group, on the 10th, 20th, and 30th days, the neutrophil chemotactic activity increased significantly compared to its basal value. In contrast to the Nano-Se group, in animals supplemented with an inorganic form of selenium, chemotactic activity at day 30 compared to day 20 decreased significantly. In the SeNPs group, a significant increase was observed after 10, 20, and 30 days compared to the control group and at 10 days as compared to the Na2SeO3 group (increase on day 10 was 24% in Nano-Se group and 5% in the group administered with Na2SeO3, compared to basal levels). In the Na2SeO3 group, a significant increase in chemotactic activity was observed after 20 days compared to the control group. Respiratory burst activity value showed a significant increase in both groups from day 0 to day 30.
Effect of Nano-Se on microbial fermentation, nutrients digestibility, and probiotics support
The nanoscale selenium has also shown positive effects on rumen fermentation and increased nutrient conversion. A study of the effect of Nano-Se and selenium yeast (Se-yeast) supplements (both at a dose of 4 g·kg−1 feed DM to meet the need for 4 mg Se) on food digestibility, ruminant fermentation, and urine purine derivatives in sheep14 demonstrated a decrease in ruminal pH, ammonia nitrogen concentration, molar amount of propionate, and acetate/propionate ratio. In addition, an increase in total ruminal volatile fatty acids in the Nano-Se and Se-yeast groups was also observed. Selenium intake significantly improved the in situ degradation of ruminal amylase-treated neutral detergent fiber (aNDF) from Leymus chinensis and crude protein (CP) from soybean meal. The digestibility of DM, organic matter, CP, ethereal extract, aNDF, and acid detergent fiber throughout the digestive tract and urinary excretion of purine derivatives were also significantly affected by the addition of selenium. Ruminal fermentation was improved and feed conversion efficiency increased with Nano-Se compared to Se-yeast. Nano-Se can be considered as a preferentially available source of selenium for ruminants.14
Kheradmand et al40 investigated the effect of Nano-Se on antifungal activity of probiotics on Candida albicans, usually a commensal organism in humans, which can become pathogenic especially in immunocompromised individuals. After exposure to SeNPs-enriched Lactobacillus spp. (Lactobacillus plantarum and Lactobacillus johnsonii), a greater decline in viability of C. albicans than after its exposure to non-Se-enriched LAB was observed. This indicates an increase in the antifungal activity of both bacterial strains by means of selenium. This phenomenon could potentially be used in anti-Candida probiotic formulations in future.
Nano-Se in treatment of metabolic disorders
Recently, promising nanocarriers for oral delivery of antidiabetic supplement to potentiate its curative effect were developed. Yin et al45 designed selenium-coated nanostructured lipid carriers (SeNLCs) for enhancing the oral bioavailability and strengthening the hypoglycemic action of berberine, an antidiabetic phytomedicine. Berberine-loaded SeNLCs administered orally to rats greatly enhanced the bioavailability of berberine 6.6 times and evinced significantly higher hypoglycemic effect than berberine solution as well as berberine-loaded nanostructured lipid carriers. Both properties were improved due to better sustained drug release and intestinal absorption of selenium nanocarriers.
Nano-Se has also been shown to be efficacious in the treatment of fatty liver disease (FLD). Hepatic steatosis (FLD) or fatty liver is a sign of a metabolic disorder affecting 50% of dairy cows immediately after calving, caused by ectopic fat deposition in the liver.178 Fatty liver can cause an inflammatory response in the liver that can lead to permanent liver damage and even to death. Without proper treatment, the mortality may reach up to 25%.179 Apart from changes in dietetic arrangement, the application of Nano-Se may be effective for the therapy of this disease.180 Hegedüs et al47 reported a study of the effect of Nano-Se on FLD therapy in male Wistar rats which showed a low level of inflammation and free radical release in sick animals compared to the control group. This was confirmed by transmethylation ability and histological analysis of samples.47 Bioactive nanoscale selenium appears to be effective, but confirmation of its therapeutic effect requires further experiments.180
Safety and toxicity concerns of orally delivered SeNPs for use as food additives and drug carriers
Despite the growing interest of scientists on SeNPs and reporting of their wide range of positive effects, there are some concerns about their toxicity and objections to their use in clinical practice.
Many studies have demonstrated that the administration of selenium can prevent cancer and reduce its incidence.35,181,182 Moreover, another potential of selenium was conclusively showed in the fight against cancer – through combination with chemotherapeutic and hormonal agents.183,184 Numerous investigations have shown that both tissue and cell distribution profiles of anticancer drugs could be improved by nanotechnology.35,185,186 Nanosized anticancer drugs displayed increased antitumor efficiency and reduced serious side effects.187 It was revealed that selenium can sensitize cancer cells to conventionally used anticancer drugs. For example, Hu et al183 reported that selenium sensitized hormone-refractory prostate cancer cells to apoptosis induced by anticancer drug paclitaxel (Taxol) through enhancing multiple caspases. Li et al184 showed that the combination of doxorubicin with selenium enhanced apoptosis in the MCF-7 human breast cancer cell line.
However, selenium evinces a narrow margin between beneficial and toxic effects. An effective dose of selenium as an anticancer agent approaches the toxicity limit, which substantially limits its clinical application. Nevertheless, the beneficial and toxic effects of selenium on health are greatly dependent not only on its concentration but also on its chemical form.35 While some studies reported that exceeding the tolerable upper intake level of 400 μg·day−1 in humans can lead to selenosis,179,180 Reid et al181,182,188 observed no obvious symptoms of selenosis in patients receiving 1,600 μg Se·day−1 in the form of selenized yeast with SeMet as the major selenium species. Even in those with an intake of 3,200 Se·day−1, only some symptoms of selenium toxicity did occur.
Studies performed with Nano-Se revealed that elemental selenium at nanoscale is much less toxic in comparison with the organic selenium compounds, such as SeMet22 and Se-MetSeCys,23 and exhibited comparable efficacy to them in upregulating selenoenzymes and tissue selenium levels.
The in vitro free radical-scavenging efficiency of SeNPs was also found higher than organic and inorganic selenocompounds.22,159 For instance, based on the data of LD50, the in vivo toxicity of SeNPs was about 4–6 times lower than SeMet and Se-MetSeCys.22,23,159 The higher toxicity of selenite, SeCys2, and SeO2 is associated with their ability to initiate the oxidation of the thiol groups of proteins,189 which may lead to a change in the activity of essential enzymes containing the sulfhydryl group.190 Selenite, compared to the same dose of Nano-Se, more markedly reduces hepatic GPx level and increases the production of MDA which is the product of lipid peroxidation as well as reducing the activity of antioxidant enzymes SOD and CAT in the liver, than the same dose of nanoscale selenium.159 Wang et al investigated the effect of SeMet in mice which, compared to those with Nano-Se supplementation, had strongly elevated levels of liver enzymes such as alanine aminotransferase, aspartate aminotransferase, and lactate dehydrogenase in the blood, over a long period of time, indicating acute severe liver injury.22 Acute toxicity due to Nano-Se occurs at a much higher dose compared to organically bound selenium forms. The median lethal dose (LD50) is 92.1 mg Se·kg−1 for Nano-Se, 25.6 mg Se·kg−1 for SeMet,22 and 14.6 mg Se·kg−1 for Se-MetSeCys.23
Studies showed that Nano-Se has a similar bioavailability in rats and much less acute toxicity in mice compared with selenite.191 The dose of red elemental Nano-Se that causes acute toxicity was approximately seven times that of Na2SeO3: the LD50 was about 113 and 16 mg Se·kg−1 BW, respectively. The reaction ratio of red elemental Nano-Se with glutathione in vitro was one-tenth of that of Na2SeO3.192
Moreover, the design of SeNPs-loaded CS microspheres (SeNPs-M) can offer a new way for further development of SeNPs with a higher efficacy and better biosafety.115 Based on acute toxicity test, SeNPs-M were much safer than selenite; the LD50 was around 18-fold of selenite. Furthermore, SeNPs-M possessed a strong antioxidant activity, as evidenced by a dramatic increase in selenium retention and also in the activities of GPx, SOD, and CAT. Oral administration of SeNPs-M can be considered as an effective way to supply selenium.
Mittal et al193 synthetized bimetallic (Ag-Se) NPs which were used as anticancer agents for Dalton lymphoma cells. The Ag-Se NPs evinced strong anticancer activity at a lower concentration. In vitro viability of cells of the cancer line was reduced to 20% at 50 μg·mL−1 Ag-SeNPs.
Nano-Se is also less toxic than high-selenium protein. A study of the subchronic toxicity of these selenium forms performed in Sprague-Dawley rats191 (in both genders) fed with diets containing individual compounds at different concentrations (0, 2, 3, 4, and 5 mg·kg−1 Se) for 13 weeks showed significant abnormal changes in BW, hematology, clinical chemistry, relative organ weights, and histopathology parameters at the doses of 4 and 5 mg·kg−1 Se. The toxicity was more pronounced in the selenite and high-selenium protein groups than in the Nano-Se group, at the dose of 3 mg·kg−1 Se. Significant growth inhibition and degeneration of hepatic cells were found in the selenite and high-selenium protein groups. On the other hand, no changes attributable to administration of Nano-Se at the dose of 3 mg·kg−1 Se were found. No-observed-adverse-effect level (NOAEL; the highest experimental point that is without adverse effect) of Nano-Se in male and female rats was considered to be 3 mg Se·kg−1 (equivalent to 0.22 and 0.33 mg·kg−1 BW·day−1 for males and females, respectively). On the contrary, the NOAELs of selenite and high-selenium protein in both genders were considered to be 2 mg Se·kg−1 (equivalent to 0.14 and 0.20 mg·kg−1 BW·day−1 for males and females, respectively).191
Therefore, it can be assumed that SeNPs have a much wider margin between beneficial and toxic effects than other selenocompounds and could serve as a suitable potential chemopreventive agent with reduced risk of toxicity.35
Higher IC50 of the SeNPs (41.5 ± 0.9 μg·mL−1) compared to SeO2 (6.7 ± 0.8 μg·mL−1) confirmed the lower cytotoxicity of the biogenic SeNPs on MCF-7 cell line.89
Regarding testing toxicity in mammalian models, Benko et al194 compared the toxicity of different selenium species in mice which were administered for 14 days at concentrations of 0.5, 5, and 50 mg Se·kg−1 food, corresponding to an estimated 4, 40, and 400 mg Se·kg−1 BW·day−1, respectively. A study on 14-day murine subacute toxicity showed that toxicity was more pronounced when an inorganic selenium (Na2SeO4, NaHSeO3) was applied than after subacute application of Sel-Plex (a natural source of organic selenium consisting of predominantly selenoaminoacids [SeCys, SeMet] produced by S. cerevisiae), Nano-Se (synthetized by yogurt strains L. acidophilus, S. thermophilus, and L. casei), or LactoMicroSe (selenium-enriched yogurt powder – L. acidophilus, S. thermophilus, and L. casei). The toxicity of selenium species decreased in the following order: selenate > selenite > Nano-Se > Sel-Plex > LactoMicroSe. He et al195 reported a study on toxicity of Nano-Se in rats showing that supranutritional levels of SeNPs (at doses of 0.2, 0.4, 0.8, 2.0, 4.0, or 8.0 mg Se·kg−1 BW administered orally each day for 14 consecutive days) had no obvious toxic effects, and could be considered as potential candidates for cancer chemoprevention, although doses greater than 2.0 mg Se·kg−1 BW induced chronic toxicity.
Shakibaie et al196 investigated the acute and subacute toxicity of the biogenic SeNPs synthetized by Bacillus sp. MSh-1and SeO2 in mice. The biogenic SeNPs were much less (26-fold) toxic than the SeO2. The toxicological evaluation showed that the LD50 values of SeO2 and SeNPs were 7.3 and 198.1 mg·kg−1, respectively. No biochemical changes were observed from the administration of 2.5, 5, and 10 mg SeNPs·kg−1, but a dose of 20 mg·kg−1 was accompanied with signs of toxicity including lower BW and changes in clinical chemistry and hematological parameters.
On the other side, studies on the toxicity of Nano-Se to aquatic organisms with relatively contradictory results appeared. Li et al137 found that a 10-day exposure to SeNPs at a dosage of 100 μg Se·L−1 compared to the same amount of Na2SeO3 in Medaka (Oryzias latipes) fish showed a greater toxicity due to hyperaccumulation. Gallego-Gallegos et al197 evaluated the toxicity of SeNPs using 10-day waterborne and dietary exposures to larvae of Chironomus dilutus, a common benthic midge frequently used as a test organism for assessing the toxicity of sedimentary substances. They reported that even the lowest Se0 and SeNPs concentrations tested (2.81 μg·L−1 and 8.89 μg·g−1 dry weight, respectively) resulted in selenium bioaccumulation especially as SeMet. Inhibition of larval growth at higher concentrations due to both dietary and waterborne exposure was also observed.
However, both previous toxicological studies used chemically prepared NPs. Newer research works showed that the toxicity of Nano-Se also depends on the method of preparation; for instance, biogenic Nano-Se in comparison with that chemogenic is less toxic. Mal et al198 investigated the toxicity of biogenic Nano-Se formed by anaerobic granular sludge biofilms on zebrafish (Danio rerio) embryos in comparison with selenite and chemogenic Nano-Se. The biogenic Nano-Se formed by granular sludge biofilms showed an LC50 (50% lethal concentration) value of 1.77 mg·L−1, which was 3.2-fold less toxic to zebrafish embryos than selenite (LC50 = 0.55 mg·L−1) and 10-fold less toxic than chemogenic, bovine serum albumin-stabilized Nano-Se (LC50 = 0.16 mg·L−1). Interestingly, chemically synthesized Nano-Se particles of small (25–80 nm) and large (50–250 nm) size showed comparable toxicity on zebrafish embryos.
Moreover, a study of Khiralla and El-Deeb199 also did not show any high toxicity of SeNPs biosynthesized by B. licheniformis assayed using larvae of Artemia salina, which have been suggested as a model organism in toxicity assessment of NPs.200 No toxicity on Artemia larvae was demonstrated by SeNPs up to 100 μg·mL−1.
Furthermore, the method of extraction of biogenic NPs substantially affects the toxicity of SeNPs. Sonkusre et al34 reported a novel method for the extraction and purification of intracellular SeNPs from the Gram-positive bacteria B. licheniformis JS2. These neutral-charged, non-agglomerating NPs at a very low concentration as low as 2 μg Se·mL−1 were efficacious in inhibiting proliferation and excellent in inducing caspase-independent necrosis of human prostate adenocarcinoma cells (PC3) without causing any significant toxicity to human peripheral blood mononuclear cells.
Conclusion
Selenium is an important essential element that interferes through selenoproteins in many physiological processes of the organism and affects the production and reproductive properties. By providing adequate supply of selenium in the diet, it is possible to effectively prevent health problems from its deficiency. Due to its high bioavailability, low toxicity and affordability, selenium in its nanoform appears to be the most appropriate for supplementation, especially in ruminants, in which traditionally used selenium compounds exhibit very low absorption in the digestive tract.
Future perspective is the possibility of global application of nanoscale selenium in nutrition as well as in clinical medicine. The development of new NP systems for the transport of selenium in the organism, with the possibility of modifying physicochemical properties of the particles, greater stability in the gastrointestinal tract, and allowing controlled release of selenium, offers a significant dietary and therapeutic potential. Although there are currently some concerns about the use of SeNPs for therapeutic purposes, many scientific studies suggested that many of these generally prevailing doubts have not been confirmed. However, it is still necessary to carry out further preclinical studies in animal models.
Acknowledgments
This work was supported by the project for conceptual development of the research organization. The authors would like to thank The European Technology Platform for Nanomedicine for their cooperation.
Author contributions
Bozena Hosnedlova performed the literature search, wrote the manuscript, and drew figures. Marta Kepinska, Sylvie Skalickova, Carlos Fernandez, Branislav Ruttkay-Nedecky, Qiuming Peng, Mojmir Baron, Magdalena Melcova, Radka Opatrilova, Jarmila Zidkova, Geir Bjørklund, and Jiri Sochor participated in writing and correction of the manuscript. Rene Kizek conceived the idea for this topic, proposed the concept, performed the literature search, and participated in writing and critical revision of the manuscript. All authors contributed toward data analysis, drafting and revising the paper and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Ensign LM, Cone R, Hanes J. Oral drug delivery with polymeric nanoparticles: the gastrointestinal mucus barriers. Adv Drug Deliv Rev. 2012;64(6):557–570. | ||
Zhang S, Luo Y, Zeng H, et al. Encapsulation of selenium in chitosan nanoparticles improves selenium availability and protects cells from selenium-induced DNA damage response. J Nutr Biochem. 2011;22(12):1137–1142. | ||
Anal AK, Singh H. Recent advances in microencapsulation of probiotics for industrial applications and targeted delivery. Trends Food Sci Technol. 2007;18(5):240–251. | ||
Gökmen V, Mogol BA, Lumaga RB, Fogliano V, Kaplun Z, Shimoni E. Development of functional bread containing nanoencapsulated omega-3 fatty acids. J Food Eng. 2011;105(4):585–591. | ||
Hadrup N, Loeschner K, Skov K, et al. Effects of 14-day oral low dose selenium nanoparticles and selenite in rat – as determined by metabolite pattern determination. PeerJ. 2016;4:e2601. | ||
Hu CH, Li YL, Xiong L, Zhang HM, Song J, Xia MS. Comparative effects of nano elemental selenium and sodium selenite on selenium retention in broiler chickens. Anim Feed Sci Technol. 2012;177(3):204–210. | ||
Kojouri GA, Jahanabadi S, Shakibaie M, Ahadi AM, Shahverdi AR. Effect of selenium supplementation with sodium selenite and selenium nanoparticles on iron homeostasis and transferrin gene expression in sheep: a preliminary study. Res Vet Sci. 2012;93(1):275–278. | ||
Kojouri GA, Sadeghian S, Mohebbi A, Dezfouli MRM. The effects of oral consumption of selenium nanoparticles on chemotactic and respiratory burst activities of neutrophils in comparison with sodium selenite in sheep. Biol Trace Elem Res. 2012;146(2):160–166. | ||
Luo Y, Teng Z, Wang Q. Development of zein nanoparticles coated with carboxymethyl chitosan for encapsulation and controlled release of vitamin D3. J Agric Food Chem. 2012;60(3):836–843. | ||
Mahmoudvand H, Harandi MF, Shakibaie M, et al. Scolicidal effects of biogenic selenium nanoparticles against protoscolices of hydatid cysts. Int J Surg. 2014;12(5):399–403. | ||
Rashidi L, Khosravi-Darani K. The applications of nanotechnology in food industry. Crit Rev Food Sci Nutr. 2011;51(8):723–730. | ||
Sadeghian S, Kojouri GA, Mohebbi A. Nanoparticles of selenium as species with stronger physiological effects in sheep in comparison with sodium selenite. Biol Trace Elem Res. 2012;146(3):302–308. | ||
Shi LG, Yang RJ, Yue WB, et al. Effect of elemental nano-selenium on semen quality, glutathione peroxidase activity, and testis ultrastructure in male Boer goats. Anim Reprod Sci. 2010;118(2):248–254. | ||
Xun W, Shi L, Yue W, Zhang C, Ren Y, Liu Q. Effect of high-dose nano-selenium and selenium–yeast on feed digestibility, rumen fermentation, and purine derivatives in sheep. Biol Trace Elem Res. 2012;150(1–3):130–136. | ||
Zhai X, Zhang C, Zhao G, Stoll S, Ren F, Leng X. Antioxidant capacities of the selenium nanoparticles stabilized by chitosan. J Nanobiotechnology. 2017;15(1):4. | ||
Zhang J, Wang H, Bao Y, Zhang L. Nano red elemental selenium has no size effect in the induction of seleno-enzymes in both cultured cells and mice. Life Sci. 2004;75(2):237–244. | ||
Wacker MG. Nanotherapeutics – product development along the “nanomaterial” discussion. J Pharm Sci. 2014;103(3):777–784. | ||
Chen L, Remondetto GE, Subirade M. Food protein-based materials as nutraceutical delivery systems. Trends Food Sci Technol. 2006;17(5):272–283. | ||
Agrawal U, Sharma R, Gupta M, Vyas SP. Is nanotechnology a boon for oral drug delivery? Drug Discov Today. 2014;19(10):1530–1546. | ||
McClements DJ. Nanoemulsions versus microemulsions: terminology, differences, and similarities. Soft Matter. 2012;8(6):1719–1729. | ||
Shi L, Xun W, Yue W, et al. Effect of sodium selenite, Se-yeast and nano-elemental selenium on growth performance, Se concentration and antioxidant status in growing male goats. Small Ruminant Res. 2011;96(1):49–52. | ||
Wang H, Zhang J, Yu H. Elemental selenium at nano size possesses lower toxicity without compromising the fundamental effect on selenoenzymes: comparison with selenomethionine in mice. Free Radic Biol Med. 2007;42(10):1524–1533. | ||
Zhang J, Wang X, Xu T. Elemental selenium at nano size (Nano-Se) as a potential chemopreventive agent with reduced risk of selenium toxicity: comparison with Se-methylselenocysteine in mice. Toxicol Sci. 2008;101(1):22–31. | ||
Fajt Z, Drabek J, Steinhauser L, Svobodova Z. The significance of pork as a source of dietary selenium – an evaluation of the situation in the Czech Republic. Neuro Endocrinol Lett. 2009;30 Suppl 1:17–21. | ||
Torres SK, Campos VL, León CG, et al. Biosynthesis of selenium nanoparticles by Pantoea agglomerans and their antioxidant activity. J Nanopart Res. 2012;14(11):1236. | ||
Desai MP, Labhasetwar V, Walter E, Levy RJ, Amidon GL. The mechanism of uptake of biodegradable microparticles in Caco-2 cells is size dependent. Pharm Res. 1997;14(11):1568–1573. | ||
De Jong WH, Borm PJ. Drug delivery and nanoparticles: applications and hazards. Int J Nanomedicine. 2008;3(2):133–149. | ||
Yao M, McClements DJ, Xiao H. Improving oral bioavailability of nutraceuticals by engineered nanoparticle-based delivery systems. Curr Opin Food Sci. 2015;2:14–19. | ||
Kojouri GA, Sharifi S. Preventing effects of nano-selenium particles on serum concentration of blood urea nitrogen, creatinine, and total protein during intense exercise in donkey. J Equine Vet Sci. 2013;33(8):597–600. | ||
Gao X, Zhang J, Zhang L. Hollow sphere selenium nanoparticles: their in vitro anti hydroxyl radical effect. Adv Mater. 2002;14(4):290–293. | ||
Chen T, Wong YS, Zheng W, Bai Y, Huang L. Selenium nanoparticles fabricated in Undaria pinnatifida polysaccharide solutions induce mitochondria-mediated apoptosis in A375 human melanoma cells. Colloids Surf B Biointerfaces. 2008;67(1):26–31. | ||
Luo H, Wang F, Bai Y, Chen T, Zheng W. Selenium nanoparticles inhibit the growth of HeLa and MDA-MB-231 cells through induction of S phase arrest. Colloids Surf B Biointerfaces. 2012;94:304–308. | ||
Vekariya KK, Kaur J, Tikoo K. ERα signaling imparts chemotherapeutic selectivity to selenium nanoparticles in breast cancer. Nanomedicine. 2012;8(7):1125–1132. | ||
Sonkusre P, Nanduri R, Gupta P, Cameotra SS. Improved extraction of intracellular biogenic selenium nanoparticles and their specificity for cancer chemoprevention. J Nanomed Nanotechnol. 2014;5(2):1. | ||
Liu W, Li X, Wong YS, et al. Selenium nanoparticles as a carrier of 5-fluorouracil to achieve anticancer synergism. ACS Nano. 2012;6(8):6578–6591. | ||
Estevez H, Garcia-Lidon JC, Luque-Garcia JL, Camara C. Effects of chitosan-stabilized selenium nanoparticles on cell proliferation, apoptosis and cell cycle pattern in HepG2 cells: comparison with other selenospecies. Colloids Surf B Biointerfaces. 2014;122:184–193. | ||
Wang Q, Webster TJ. Nanostructured selenium for preventing biofilm formation on polycarbonate medical devices. J Biomed Mater Res A. 2012;100(12):3205–3210. | ||
Piacenza E, Presentato A, Zonaro E, et al. Antimicrobial activity of biogenically produced spherical Se-nanomaterials embedded in organic material against Pseudomonas aeruginosa and Staphylococcus aureus strains on hydroxyapatite-coated surfaces. Microb Biotechnol. 2017;10(4):804–818. | ||
Cremonini E, Zonaro E, Donini M, et al. Biogenic selenium nanoparticles: characterization, antimicrobial activity and effects on human dendritic cells and fibroblasts. Microb Biotechnol. 2016;9(6):758–771. | ||
Kheradmand E, Rafii F, Yazdi MH, Sepahi AA, Shahverdi AR, Oveisi MR. The antimicrobial effects of selenium nanoparticle-enriched probiotics and their fermented broth against Candida albicans. Daru. 2014;22(1):48. | ||
Trabelsi H, Azzouz I, Ferchichi S, Tebourbi O, Sakly M, Abdelmelek H. Nanotoxicological evaluation of oxidative responses in rat nephrocytes induced by cadmium. Int J Nanomedicine. 2013;8:3447–3453. | ||
Prasad KS, Selvaraj K. Biogenic synthesis of selenium nanoparticles and their effect on As (III)-induced toxicity on human lymphocytes. Biol Trace Elem Res. 2014;157(3):275–283. | ||
Hassanin KM, Abd El-Kawi SH, Hashem KS. The prospective protective effect of selenium nanoparticles against chromium-induced oxidative and cellular damage in rat thyroid. Int J Nanomedicine. 2013;8:1713–1720. | ||
Yazdi MH, Mahdavi M, Setayesh N, Esfandyar M, Shahverdi AR. Selenium nanoparticle-enriched Lactobacillus brevis causes more efficient immune responses in vivo and reduces the liver metastasis in metastatic form of mouse breast cancer. Daru. 2013;21(1):33. | ||
Yin J, Hou Y, Yin Y, Song X. Selenium-coated nanostructured lipid carriers used for oral delivery of berberine to accomplish a synergic hypoglycemic effect. Int J Nanomedicine. 2017;12:8671–8680. | ||
Wu X, Yao J, Yang Z, et al. Improved fetal hair follicle development by maternal supplement of selenium at nano size (Nano-Se). Livest Sci. 2011;142(1):270–275. | ||
Hegedüs V, Prokisch J, Fébel H, et al. Nanoselenium treatment in fatty liver. Z Gastroenterol. 2012;50(05):A29. | ||
Beheshti N, Soflaei S, Shakibaie M, et al. Efficacy of biogenic selenium nanoparticles against Leishmania major: in vitro and in vivo studies. J Trace Elem Med Biol. 2013;27(3):203–207. | ||
Soflaei S, Dalimi A, Abdoli A, et al. Anti-leishmanial activities of selenium nanoparticles and selenium dioxide on Leishmania infantum. Comp Clin Path. 2014;23(1):15–20. | ||
Zhang SY, Zhang J, Wang HY, Chen HY. Synthesis of selenium nanoparticles in the presence of polysaccharides. Mater Lett. 2004;58(21):2590–2594. | ||
Quintana M, Haro-Poniatowski E, Morales J, Batina N. Synthesis of selenium nanoparticles by pulsed laser ablation. Appl Surf Sci. 2002;195(1):175–186. | ||
Ramamurthy CH, Sampath KS, Arunkumar P, et al. Green synthesis and characterization of selenium nanoparticles and its augmented cytotoxicity with doxorubicin on cancer cells. Bioprocess Biosyst Eng. 2013;36(8):1131–1139. | ||
Shoeibi S, Mashreghi M. Biosynthesis of selenium nanoparticles using Enterococcus faecalis and evaluation of their antibacterial activities. J Trace Elem Med Biol. 2017;39:135–139. | ||
Agnihotri SA, Mallikarjuna NN, Aminabhavi TM. Recent advances on chitosan-based micro- and nanoparticles in drug delivery. J Control Release. 2004;100(1):5–28. | ||
Bezerra MA, Santelli RE, Oliveira EP, Villar LS, Escaleira LA. Response surface methodology (RSM) as a tool for optimization in analytical chemistry. Talanta. 2008;76(5):965–977. | ||
Saini D, Fazil M, Ali MM, Baboota S, Ameeduzzafar A, Ali J. Formulation, development and optimization of raloxifene-loaded chitosan nanoparticles for treatment of osteoporosis. Drug Deliv. 2015;22(6):823–836. | ||
Rinaudo M. Chitin and chitosan: properties and applications. Prog Polym Sci. 2006;31(7):603–632. | ||
Langi B, Shah C, Singh K, Chaskar A, Kumar MS, Bajaj PN. Ionic liquid-induced synthesis of selenium nanoparticles. Mater Res Bull. 2010;45(6):668–671. | ||
Singh RK, Narayan J. Pulsed-laser evaporation technique for deposition of thin films: physics and theoretical model. Phys Rev B. 1990;41(13):8843. | ||
Marine W, Patrone L, Luk’yanchuk B, Sentis M. Strategy of nanocluster and nanostructure synthesis by conventional pulsed laser ablation. Appl Surf Sci. 2000;154:345–352. | ||
Ankamwar B, Chaudhary M, Sastry M. Gold nanotriangles biologically synthesized using tamarind leaf extract and potential application in vapor sensing. Synth React Inorg Met Org Chem. 2005;35(1):19–26. | ||
Suresh K, Prabagaran SR, Sengupta S, Shivaji S. Bacillus indicus sp. nov., an arsenic-resistant bacterium isolated from an aquifer in West Bengal, India. Int J Syst Evol Microbiol. 2004;54(Pt 4):1369–1375. | ||
Song JY, Kim BS. Rapid biological synthesis of silver nanoparticles using plant leaf extracts. Bioprocess Biosyst Eng. 2009;32(1):79–84. | ||
Bhainsa KC, D’souza SF. Extracellular biosynthesis of silver nanoparticles using the fungus Aspergillus fumigatus. Colloids Surf B Biointerfaces. 2006;47(2):160–164. | ||
Wang T, Yang L, Zhang B, Liu J. Extracellular biosynthesis and transformation of selenium nanoparticles and application in H2O2 biosensor. Colloids Surf B Biointerfaces. 2010;80(1):94–102. | ||
Oremland RS, Herbel MJ, Blum JS, et al. Structural and spectral features of selenium nanospheres produced by Se-respiring bacteria. Appl Environ Microbiol. 2004;70(1):52–60. | ||
Mandal D, Bolander ME, Mukhopadhyay D, Sarkar G, Mukherjee P. The use of microorganisms for the formation of metal nanoparticles and their application. Appl Microbiol Biotechnol. 2006;69(5):485–492. | ||
Ingale AG, Chaudhari AN. Biogenic synthesis of nanoparticles and potential applications: an eco-friendly approach. J Nanomed Nanotechnol. 2013;4(165):1–7. | ||
Ezhuthupurakkal PB, Polaki LR, Suyavaran A, Subastri A, Sujatha V, Thirunavukkarasu C. Selenium nanoparticles synthesized in aqueous extract of Allium sativum perturbs the structural integrity of calf thymus DNA through intercalation and groove binding. Mater Sci Eng C Mater Biol Appl. 2017;74:597–608. | ||
Zhang W, Zhang J, Ding D, et al. Synthesis and antioxidant properties of Lycium barbarum polysaccharides capped selenium nanoparticles using tea extract. Artif Cells Nanomed Biotechnol. 2017:1–8. | ||
Sowndarya P, Ramkumar G, Shivakumar M. Green synthesis of selenium nanoparticles conjugated Clausena dentata plant leaf extract and their insecticidal potential against mosquito vectors. Artif Cells Nanomed Biotechnol. 2016;45(8):1490–1495. | ||
Eszenyi P, Sztrik A, Babka B, Prokisch J. Elemental, nano-sized (100–500 nm) selenium production by probiotic lactic acid bacteria. Int J Biosci Biochem Bioinforma. 2011;1(2):148–152. | ||
Tan Y, Yao R, Wang R, Wang D, Wang G, Zheng S. Reduction of selenite to Se(0) nanoparticles by filamentous bacterium Streptomyces sp. ES2-5 isolated from a selenium mining soil. Microb Cell Fact. 2016;15(1):157. | ||
Mishra RR, Prajapati S, Das J, Dangar TK, Das N, Thatoi H. Reduction of selenite to red elemental selenium by moderately halotolerant Bacillus megaterium strains isolated from Bhitarkanika mangrove soil and characterization of reduced product. Chemosphere. 2011;84(9):1231–1237. | ||
Kora AJ, Rastogi L. Bacteriogenic synthesis of selenium nanoparticles by Escherichia coli ATCC 35218 and its structural characterisation. IET Nanobiotechnol. 2016;11(2):179–184. | ||
Kim EB, Seo JM, Kim GW, Lee SY, Park TJ. In vivo synthesis of europium selenide nanoparticles and related cytotoxicity evaluation of human cells. Enzyme Microb Technol. 2016;95:201–208. | ||
Srivastava N, Mukhopadhyay M. Green synthesis and structural characterization of selenium nanoparticles and assessment of their antimicrobial property. Bioprocess Biosyst Eng. 2015;38(9):1723–1730. | ||
Song D, Li X, Cheng Y, et al. Aerobic biogenesis of selenium nanoparticles by Enterobacter cloacae Z0206 as a consequence of fumarate reductase mediated selenite reduction. Sci Rep. 2017;7(1):3239. | ||
Kora AJ, Rastogi L. Biomimetic synthesis of selenium nanoparticles by Pseudomonas aeruginosa ATCC 27853: an approach for conversion of selenite. J Environ Manage. 2016;181:231–236. | ||
Fesharaki PJ, Nazari P, Shakibaie M, et al. Biosynthesis of selenium nanoparticles using Klebsiella pneumoniae and their recovery by a simple sterilization process. Braz J Microbiol. 2010;41(2):461–466. | ||
Sasidharan S, Balakrishnaraja R. Comparison studies on the synthesis of selenium nanoparticles by various microorganisms. Int J Pure Appl Biosci. 2014;2(1):112–117. | ||
Srivastava N, Mukhopadhyay M. Biosynthesis and structural characterization of selenium nanoparticles mediated by Zooglea ramigera. Powder Technol. 2013;244:26–29. | ||
Li B, Liu N, Li Y, et al. Reduction of selenite to red elemental selenium by Rhodopseudomonas palustris strain N. PLoS One. 2014;9(4):e95955. | ||
Tam K, Ho CT, Lee JH, et al. Growth mechanism of amorphous selenium nanoparticles synthesized by Shewanella sp. HN-41. Biosci Biotechnol Biochem. 2010;74(4):696–700. | ||
Fernández-Llamosas H, Castro L, Blázquez ML, Díaz E, Carmona M. Biosynthesis of selenium nanoparticles by Azoarcus sp. CIB. Microb Cell Fact. 2016;15(1):109. | ||
Khoei NS, Lampis S, Zonaro E, Yrjälä K, Bernardi P, Vallini G. Insights into selenite reduction and biogenesis of elemental selenium nanoparticles by two environmental isolates of Burkholderia fungorum. N Biotechnol. 2017;34:1–11. | ||
Cavalu S, Prokisch J, Laslo V, Vicas S. Preparation, structural characterisation and release study of novel hybrid microspheres entrapping nanoselenium, produced by green synthesis. IET Nanobiotechnol. 2016;11(4):426–432. | ||
Estevam EC, Griffin S, Nasim MJ, et al. Natural selenium particles from Staphylococcus carnosus: hazards or particles with particular promise? J Hazard Mater. 2017;324:22–30. | ||
Forootanfar H, Adeli-Sardou M, Nikkhoo M, et al. Antioxidant and cytotoxic effect of biologically synthesized selenium nanoparticles in comparison to selenium dioxide. J Trace Elem Med Biol. 2014;28(1):75–79. | ||
Sonkusre P, Cameotra SS. Biogenic selenium nanoparticles induce ROS-mediated necroptosis in PC-3 cancer cells through TNF activation. J Nanobiotechnology. 2017;15(1):43. | ||
Zinicovscaia I, Chiriac T, Cepoi L, et al. Selenium uptake and assessment of the biochemical changes in Arthrospira (Spirulina) platensis biomass during the synthesis of selenium nanoparticles. Can J Microbiol. 2017;63(1):27–34. | ||
Cui YH, Li LL, Zhou NQ, et al. In vivo synthesis of nano-selenium by Tetrahymena thermophila SB210. Enzyme Microb Technol. 2016;95:185–191. | ||
Zhang L, Li D, Gao P. Expulsion of selenium/protein nanoparticles through vesicle-like structures by Saccharomyces cerevisiae under microaerophilic environment. World J Microbiol Biotechnol. 2012;28(12):3381–3386. | ||
Elahian F, Reiisi S, Shahidi A, Mirzaei SA. High-throughput bioaccumulation, biotransformation, and production of silver and selenium nanoparticles using genetically engineered Pichia pastoris. Nanomedicine. 2017;13(3):853–861. | ||
Zare B, Babaie S, Setayesh N, Shahverdi AR. Isolation and characterization of a fungus for extracellular synthesis of small selenium nanoparticles. Nanomed J. 2013;1(1):13–19. | ||
Mahan DC, Cline TR, Richert B. Effects of dietary levels of selenium-enriched yeast and sodium selenite as selenium sources fed to growing-finishing pigs on performance, tissue selenium, serum glutathione peroxidase activity, carcass characteristics, and loin quality. J Anim Sci. 1999;77(8):2172–2179. | ||
National Research Council. Nutrient Requirements of Beef Cattle: Seventh Revised Edition. Washington, DC: The National Academies Press; 2000. | ||
National Research Council. Selenium in Nutrition: Revised Edition. Washington, DC: The National Academies Press; 1983. | ||
Spears JW. Trace mineral bioavailability in ruminants. J Nutr. 2003;133(5):1506S–1509S. | ||
Romero-Pérez A, García-García E, Zavaleta-Mancera A, et al. Designing and evaluation of sodium selenite nanoparticles in vitro to improve selenium absorption in ruminants. Vet Res Commun. 2010;34(1):71–79. | ||
Ubrich N, Schmidt C, Bodmeier R, Hoffman M, Maincent P. Oral evaluation in rabbits of cyclosporin-loaded Eudragit RS or RL nanoparticles. Int J Pharm. 2005;288(1):169–175. | ||
des Rieux A, Fievez V, Garinot M, Schneider Y-J, Préat V. Nanoparticles as potential oral delivery systems of proteins and vaccines: a mechanistic approach. J Control Release. 2006;116(1):1–27. | ||
Nellans HN. (B) Mechanisms of peptide and protein absorption: (1) Paracellular intestinal transport: modulation of absorption. Adv Drug Deliv Rev. 1991;7(3):339–364. | ||
Shakweh M, Ponchel G, Fattal E. Particle uptake by Peyer’s patches: a pathway for drug and vaccine delivery. Expert Opin Drug Deliv. 2004;1(1):141–163. | ||
Buono C, Anzinger JJ, Amar M, Kruth HS. Fluorescent pegylated nanoparticles demonstrate fluid-phase pinocytosis by macrophages in mouse atherosclerotic lesions. J Clin Invest. 2009;119(5):1373–1381. | ||
Plapied L, Duhem N, des Rieux A, Préat V. Fate of polymeric nanocarriers for oral drug delivery. Curr Opin Colloid Interface Sci. 2011;16(3):228–237. | ||
Bergin IL, Witzmann FA. Nanoparticle toxicity by the gastrointestinal route: evidence and knowledge gaps. Int J Biomed Nanosci Nanotechnol. 2013;3(1–2):163–210. | ||
Zhang H, Xia M, Hu C. Effect of nano-selenium on the activities of glutathione peroxidase and type-I deiodinase in the liver of weanling pigs. Sheng Wu Yi Xue Gong Cheng Xue Za Zhi. 2007;24(1):153–156. | ||
Badade ZG, More K, Narshetty J. Oxidative stress adversely affects spermatogenesis in male infertility. Biomed Res. 2011;22(3):323–328. | ||
Cheng Z, Zhi X, Sun G, et al. Sodium selenite suppresses hepatitis B virus transcription and replication in human hepatoma cell lines. J Med Virol. 2016;88(4):653–663. | ||
Jackman JA, Lee J, Cho NJ. Nanomedicine for infectious disease applications: innovation towards broad-spectrum treatment of viral infections. Small. 2016;12(9):1133–1139. | ||
Men X, Xu W, Zhu X, Ma W. Extraction, selenium-nanoparticle preparation and anti-virus bioactivity determination of polysaccharides from Caulerpa taxifolia. Zhong Yao Cai. 2009;32(12):1891–1894. | ||
Ramya S, Shanmugasundaram T, Balagurunathan R. Biomedical potential of actinobacterially synthesized selenium nanoparticles with special reference to anti-biofilm, anti-oxidant, wound healing, cytotoxic and anti-viral activities. J Trace Elem Med Biol. 2015;32:30–39. | ||
Stevanović M, Filipović N, Djurdjević J, Lukić M, Milenković M, Boccaccini A. 45S5Bioglass®-based scaffolds coated with selenium nanoparticles or with poly(lactide-co-glycolide)/selenium particles: processing, evaluation and antibacterial activity. Colloids Surf B Biointerfaces. 2015;132:208–215. | ||
Bai K, Hong B, He J, Hong Z, Tan R. Preparation and antioxidant properties of selenium nanoparticles-loaded chitosan microspheres. Int J Nanomedicine. 2017;12:4527–4539. | ||
Erkekoğlu P, Aşçi A, Ceyhan M, et al. Selenium levels, selenoenzyme activities and oxidant/antioxidant parameters in H1N1-infected children. Turk J Pediatr. 2013;55(3):271–282. | ||
Li YH, Li XL, Wong YS, et al. The reversal of cisplatin-induced nephrotoxicity by selenium nanoparticles functionalized with 11-mercapto-1-undecanol by inhibition of ROS-mediated apoptosis. Biomaterials. 2011;32(34):9068–9076. | ||
Li Y, Li X, Zheng W, Fan C, Zhang Y, Chen T. Functionalized selenium nanoparticles with nephroprotective activity, the important roles of ROS-mediated signaling pathways. J Mater Chem B Mater Biol Med. 2013;1(46):6365–6372. | ||
Li Y, Lin Z, Zhao M, et al. Multifunctional selenium nanoparticles as carriers of HSP70 siRNA to induce apoptosis of HepG2 cells. Int J Nanomedicine. 2016;11:3065–3076. | ||
Yu L, Sun L, Nan Y, Zhu LY. Protection from H1N1 influenza virus infections in mice by supplementation with selenium: a comparison with selenium-deficient mice. Biol Trace Elem Res. 2011;141(1–3):254–261. | ||
Li Y, Lin Z, Guo M, et al. Inhibitory activity of selenium nanoparticles functionalized with oseltamivir on H1N1 influenza virus. Int J Nanomedicine. 2017;12:5733–5743. | ||
Yang J, Shim SM, Nguyen TQ, et al. Poly-γ-glutamic acid/chitosan nanogel greatly enhances the efficacy and heterosubtypic cross-reactivity of H1N1 pandemic influenza vaccine. Sci Rep. 2017;7:44839. | ||
Yang X, Zhang W, Zhao Z, et al. Quercetin loading CdSe/ZnS nanoparticles as efficient antibacterial and anticancer materials. J Inorg Biochem. 2017;167:36–48. | ||
Jia X, Liu Q, Zou S, Xu X, Zhang L. Construction of selenium nanoparticles/β-glucan composites for enhancement of the antitumor activity. Carbohydr Polym. 2015;117:434–442. | ||
Liao W, Yu Z, Lin Z, et al. Biofunctionalization of selenium nanoparticle with Dictyophora indusiata polysaccharide and its antiproliferative activity through death-receptor and mitochondria-mediated apoptotic pathways. Sci Rep. 2015;5:18629. | ||
Liao W, Zhang R, Dong C, Yu Z, Ren J. Novel walnut peptide-selenium hybrids with enhanced anticancer synergism: facile synthesis and mechanistic investigation of anticancer activity. Int J Nanomedicine. 2016;11:1305–1321. | ||
Yanhua W, Hao H, Li Y, Zhang S. Selenium-substituted hydroxyapatite nanoparticles and their in vivo antitumor effect on hepatocellular carcinoma. Colloids Surf B Biointerfaces. 2016;140:297–306. | ||
Yang F, Tang Q, Zhong X, et al. Surface decoration by Spirulina polysaccharide enhances the cellular uptake and anticancer efficacy of selenium nanoparticles. Int J Nanomedicine. 2012;7:835–844. | ||
Chan L, He L, Zhou B, et al. Cancer-targeted selenium nanoparticles sensitize cancer cells to continuous γ radiation to achieve synergetic chemo-radiotherapy. Chem Asian J. 2017;12(23):3053–3060. | ||
Bao P, Chen SC, Xiao KQ. Dynamic equilibrium of endogenous selenium nanoparticles in selenite-exposed cancer cells: a deep insight into the interaction between endogenous SeNPs and proteins. Mol Biosyst. 2015;11(12):3355–3361. | ||
Bao P, Chen Z, Tai RZ, Shen HM, Martin FL, Zhu YG. Selenite-induced toxicity in cancer cells is mediated by metabolic generation of endogenous selenium nanoparticles. J Proteome Res. 2015;14(2):1127–1136. | ||
Lopez-Heras I, Sanchez-Diaz R, Anunciação DS, Madrid Y, Luque-Garcia JL, Camara C. Effect of chitosan-stabilized selenium nanoparticles on cell cycle arrest and invasiveness in hepatocarcinoma cells revealed by quantitative proteomics. J Nanosci Nanotechnol. 2014;5(5):1. | ||
Pi J, Yang F, Jin H, et al. Selenium nanoparticles induced membrane bio-mechanical property changes in MCF-7 cells by disturbing membrane molecules and F-actin. Bioorg Med Chem Lett. 2013;23(23):6296–6303. | ||
Zhang Y, Li X, Huang Z, Zheng W, Fan C, Chen T. Enhancement of cell permeabilization apoptosis-inducing activity of selenium nanoparticles by ATP surface decoration. Nanomedicine. 2013;9(1):74–84. | ||
Wu H, Zhu H, Li X, et al. Induction of apoptosis and cell cycle arrest in A549 human lung adenocarcinoma cells by surface-capping selenium nanoparticles: an effect enhanced by polysaccharide–protein complexes from Polyporus rhinocerus. J Agric Food Chem. 2013;61(41):9859–9866. | ||
Pi J, Jin H, Liu R, et al. Pathway of cytotoxicity induced by folic acid modified selenium nanoparticles in MCF-7 cells. Appl Microbiol Biotechnol. 2013;97(3):1051–1062. | ||
Li H, Zhang J, Wang T, Luo W, Zhou Q, Jiang G. Elemental selenium particles at nano-size (Nano-Se) are more toxic to Medaka (Oryzias latipes) as a consequence of hyper-accumulation of selenium: a comparison with sodium selenite. Aquat Toxicol. 2008;89(4):251–256. | ||
Wong HL, Bendayan R, Rauth AM, Xue HY, Babakhanian K, Wu XY. A mechanistic study of enhanced doxorubicin uptake and retention in multidrug resistant breast cancer cells using a polymer-lipid hybrid nanoparticle system. J Pharmacol Exp Ther. 2006;317(3):1372–1381. | ||
Yang X, Grailer JJ, Pilla S, Steeber DA, Gong S. Tumor-targeting, pH-responsive, and stable unimolecular micelles as drug nanocarriers for targeted cancer therapy. Bioconjug Chem. 2010;21(3):496–504. | ||
Sanpui P, Chattopadhyay A, Ghosh SS. Induction of apoptosis in cancer cells at low silver nanoparticle concentrations using chitosan nanocarrier. ACS Appl Mater Interfaces. 2011;3(2):218–228. | ||
Sahu A, Kasoju N, Bora U. Fluorescence study of the curcumin–casein micelle complexation and its application as a drug nanocarrier to cancer cells. Biomacromolecules. 2008;9(10):2905–2912. | ||
Lin LS, Cong ZX, Li J, et al. Graphitic-phase C3N4 nanosheets as efficient photosensitizers and pH-responsive drug nanocarriers for cancer imaging and therapy. J Mater Chem B. 2014;2(8):1031–1037. | ||
Kano MR, Bae Y, Iwata C, et al. Improvement of cancer-targeting therapy, using nanocarriers for intractable solid tumors by inhibition of TGF-β signaling. Proc Natl Acad Sci U S A. 2007;104(9):3460–3465. | ||
Choi KY, Yoon HY, Kim JH, et al. Smart nanocarrier based on PEGylated hyaluronic acid for cancer therapy. ACS Nano. 2011;5(11):8591–8599. | ||
Cho HS, Dong Z, Pauletti GM, et al. Fluorescent, superparamagnetic nanospheres for drug storage, targeting, and imaging: a multifunctional nanocarrier system for cancer diagnosis and treatment. ACS Nano. 2010;4(9):5398–5404. | ||
Fan X, Jiao G, Zhao W, Jin P, Li X. Magnetic Fe3O4–graphene composites as targeted drug nanocarriers for pH-activated release. Nanoscale. 2013;5(3):1143–1152. | ||
Aryal S, Grailer JJ, Pilla S, Steeber DA, Gong SQ. Doxorubicin conjugated gold nanoparticles as water-soluble and pH-responsive anticancer drug nanocarriers. J Mater Chem. 2009;19(42):7879–7884. | ||
Yang Y, Xie Q, Zhao Z, et al. Functionalized selenium nanosystem as radiation sensitizer of 125I seeds for precise cancer therapy. ACS Appl Mater Interfaces. 2017;9(31):25857–25869. | ||
Tran P, Webster TJ. Enhanced osteoblast adhesion on nanostructured selenium compacts for anti-cancer orthopedic applications. Int J Nanomedicine. 2008;3(3):391–396. | ||
Tran PA, Sarin L, Hurt RH, Webster TJ. Titanium surfaces with adherent selenium nanoclusters as a novel anticancer orthopedic material. J Biomed Mater Res A. 2010;93(4):1417–1428. | ||
Perla V, Webster TJ. Better osteoblast adhesion on nanoparticulate selenium – a promising orthopedic implant material. J Biomed Mater Res A. 2005;75(2):356–364. | ||
Zhuo H, Smith AH, Steinmaus C. Selenium and lung cancer: a quantitative analysis of heterogeneity in the current epidemiological literature. Cancer Epidemiol Biomarkers Prev. 2004;13(5):771–778. | ||
Wei WQ, Abnet CC, Qiao YL, et al. Prospective study of serum selenium concentrations and esophageal and gastric cardia cancer, heart disease, stroke, and total death. Am J Clin Nutr. 2004;79(1):80–85. | ||
Hiraoka K, Komiya S, Hamada T, Zenmyo M, Inoue A. Osteosarcoma cell apoptosis induced by selenium. J Orthop Res. 2001;19(5):809–814. | ||
Tran PA, Webster TJ. Selenium nanoparticles inhibit Staphylococcus aureus growth. Int J Nanomedicine. 2011;6:1553–1558. | ||
Holinka J, Pilz M, Kubista B, Presterl E, Windhager R. Effects of selenium coating of orthopaedic implant surfaces on bacterial adherence and osteoblastic cell growth. Bone Joint J. 2013;95(5):678–682. | ||
Stolzoff M, Webster TJ. Reducing bone cancer cell functions using selenium nanocomposites. J Biomed Mater Res A. 2016;104(2):476–482. | ||
Bao-hua X, Zi-rong X, Mei-sheng X, Cai-hong H, Yue-song D, Li X. Effect of nano red elemental selenium on GPx activity of broiler chick kidney cells in vitro. Wuhan Univ J Nat Sci. 2003;8(4):1161–1166. | ||
Zhang J, Wang H, Yan X, Zhang L. Comparison of short-term toxicity between Nano-Se and selenite in mice. Life Sci. 2005;76(10):1099–1109. | ||
Kojouri GA, Faramarzi P, Ahadi AM, Parchami A. Effect of selenium nanoparticles on expression of HSP90 gene in myocytes after an intense exercise. J Equine Vet Sci. 2013;33(12):1054–1056. | ||
Kinnunen S, Hyyppä S, Lappalainen J, et al. Exercise-induced oxidative stress and muscle stress protein responses in trotters. Eur J Appl Physiol. 2005;93(4):496–501. | ||
Khassaf M, McArdle A, Esanu C, et al. Effect of vitamin C supplements on antioxidant defense and stress proteins in human lymphocytes and skeletal muscle. J Physiol. 2003;549(Pt 2):645–652. | ||
Ji LL. Antioxidant enzyme response to exercise and aging. Med Sci Sports Exerc. 1993;25(2):225–231. | ||
Kinnunen S, Hyyppä S, Oksala N, et al. α-Lipoic acid supplementation enhances heat shock protein production and decreases post exercise lactic acid concentrations in exercised standardbred trotters. Res Vet Sci. 2009;87(3):462–467. | ||
Fischer CP, Hiscock NJ, Basu S, et al. Vitamin E isoform-specific inhibition of the exercise-induced heat shock protein 72 expression in humans. J Appl Physiol (1985). 2006;100(5):1679–1687. | ||
Childs A, Jacobs C, Kaminski T, Halliwell B, Leeuwenburgh C. Supplementation with vitamin C and N-acetyl-cysteine increases oxidative stress in humans after an acute muscle injury induced by eccentric exercise. Free Radic Biol Med. 2001;31(6):745–753. | ||
Zhang Z, Dmitrieva NI, Park JH, Levine RL, Burg MB. High urea and NaCl carbonylate proteins in renal cells in culture and in vivo, and high urea causes 8-oxoguanine lesions in their DNA. Proc Natl Acad Sci U S A. 2004;101(25):9491–9496. | ||
Rezvanfar MA, Rezvanfar MA, Shahverdi AR, et al. Protection of cisplatin-induced spermatotoxicity, DNA damage and chromatin abnormality by selenium nano-particles. Toxicol Appl Pharmacol. 2013;266(3):356–365. | ||
Vogelzang NJ, Rusthoven JJ, Symanowski J, et al. Phase III study of pemetrexed in combination with cisplatin versus cisplatin alone in patients with malignant pleural mesothelioma. J Clin Oncol. 2003;21(14):2636–2644. | ||
Scagliotti GV, Parikh P, Von Pawel J, et al. Phase III study comparing cisplatin plus gemcitabine with cisplatin plus pemetrexed in chemotherapy-naive patients with advanced-stage non-small-cell lung cancer. J Clin Oncol. 2008;26(21):3543–3551. | ||
Ozols RF, Bundy BN, Greer BE, et al. Phase III trial of carboplatin and paclitaxel compared with cisplatin and paclitaxel in patients with optimally resected stage III ovarian cancer: a Gynecologic Oncology Group study. J Clin Oncol. 2003;21(17):3194–3200. | ||
Mitsudomi T, Morita S, Yatabe Y, et al. Gefitinib versus cisplatin plus docetaxel in patients with non-small-cell lung cancer harbouring mutations of the epidermal growth factor receptor (WJTOG3405): an open label, randomised phase 3 trial. Lancet Oncol. 2010;11(2):121–128. | ||
Zdraveski ZZ, Mello JA, Farinelli CK, Essigmann JM, Marinus MG. MutS preferentially recognizes cisplatin-over oxaliplatin-modified DNA. J Biol Chem. 2002;277(2):1255–1260. | ||
Page TJ, O’brien S, Holston K, MacWilliams PS, Jefcoate CR, Czuprynski CJ. 7,12-Dimethylbenz[a]anthracene-induced bone marrow toxicity is p53-dependent. Toxicol Sci. 2003;74(1):85–92. | ||
Page TJ, MacWilliams PS, Suresh M, Jefcoate CR, Czuprynski CJ. 7–12 Dimethylbenz[a]anthracene-induced bone marrow hypocellularity is dependent on signaling through both the TNFR and PKR. Toxicol Appl Pharmacol. 2004;198(1):21–28. | ||
Ungvári É, Monori I, Megyeri A, et al. Protective effects of meat from lambs on selenium nanoparticle supplemented diet in a mouse model of polycyclic aromatic hydrocarbon-induced immunotoxicity. Food Chem Toxicol. 2014;64:298–306. | ||
Amer J, Fibach E. Chronic oxidative stress reduces the respiratory burst response of neutrophils from beta-thalassaemia patients. Br J Haematol. 2005;129(3):435–441. | ||
Ferguson JD. Nutrition and Reproduction in Dairy Herds: Proceedings of the Intermountain Nutrition Conference, Salt Lake City, UT, 2001. Logan, UT: Utah State University; 2001. | ||
Ametaj BN. A new understanding of the causes of fatty liver in dairy cows. Adv Dairy Technol. 2005;17:97–112. | ||
Sarkar B, Bhattacharjee S, Daware A, Tribedi P, Krishnani K, Minhas P. Selenium nanoparticles for stress-resilient fish and livestock. Nanoscale Res Lett. 2015;10(1):371. | ||
Popova NV. Perinatal selenium exposure decreases spontaneous liver tumorogenesis in CBA mice. Cancer Lett. 2002;179(1):39–42. | ||
Prokopczyk B, Rosa JG, Desai D, et al. Chemoprevention of lung tumorigenesis induced by a mixture of benzo(a)pyrene and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone by the organoselenium compound 1,4-phenylenebis (methylene) selenocyanate. Cancer Lett. 2000;161(1):35–46. | ||
Hu H, Li GX, Wang L, Watts J, Combs GF Jr, Lü J. Methylseleninic acid enhances taxane drug efficacy against human prostate cancer and down-regulates antiapoptotic proteins Bcl-XL and survivin. Clin Cancer Res. 2008;14(4):1150–1158. | ||
Li S, Zhou Y, Wang R, Zhang H, Dong Y, Ip C. Selenium sensitizes MCF-7 breast cancer cells to doxorubicin-induced apoptosis through modulation of phospho-Akt and its downstream substrates. Mol Cancer Ther. 2007;6(3):1031–1038. | ||
Min KH, Park K, Kim YS, et al. Hydrophobically modified glycol chitosan nanoparticles-encapsulated camptothecin enhance the drug stability and tumor targeting in cancer therapy. J Controlled Release. 2008;127(3):208–218. | ||
Yang J, Lee CH, Ko HJ, et al. Multifunctional magneto-polymeric nanohybrids for targeted detection and synergistic therapeutic effects on breast cancer. Angew Chem Int Ed. 2007;119(46):8992–8995. | ||
Cao S, Durrani FA, Rustum YM. Selective modulation of the therapeutic efficacy of anticancer drugs by selenium containing compounds against human tumor xenografts. Clin Cancer Res. 2004;10(7):2561–2569. | ||
Reid ME, Stratton MS, Lillico AJ, et al. A report of high-dose selenium supplementation: response and toxicities. J Trace Elem Med Biol. 2004;18(1):69–74. | ||
Kim TS, Yun BY, Kim IY. Induction of the mitochondrial permeability transition by selenium compounds mediated by oxidation of the protein thiol groups and generation of the superoxide. Biochem Pharmacol. 2003;66(12):2301–2311. | ||
Spallholz JE, Hoffman DJ. Selenium toxicity: cause and effects in aquatic birds. Aquat Toxicol. 2002;57(1–2):27–37. | ||
Jia X, Li N, Chen J. A subchronic toxicity study of elemental Nano-Se in Sprague-Dawley rats. Life Sci. 2005;76(17):1989–2003. | ||
Gao X, Zhang J, Zhang L. Acute toxicity and bioavailability of nano red elemental selenium. Wei Sheng Yan Jiu. 2000;29(1):57–58. | ||
Mittal AK, Kumar S, Banerjee UC. Quercetin and gallic acid mediated synthesis of bimetallic (silver and selenium) nanoparticles and their antitumor and antimicrobial potential. J Colloid Interface Sci. 2014;431:194–199. | ||
Benko I, Nagy G, Tanczos B, et al. Subacute toxicity of nano-selenium compared to other selenium species in mice. Environ Toxicol Chem. 2012;31(12):2812–2820. | ||
He Y, Chen S, Liu Z, Cheng C, Li H, Wang M. Toxicity of selenium nanoparticles in male Sprague–Dawley rats at supranutritional and nonlethal levels. Life Sci. 2014;115(1):44–51. | ||
Shakibaie M, Shahverdi AR, Faramarzi MA, Hassanzadeh GR, Rahimi HR, Sabzevari O. Acute and subacute toxicity of novel biogenic selenium nanoparticles in mice. Pharm Biol. 2013;51(1):58–63. | ||
Gallego-Gallegos M, Doig LE, Tse JJ, Pickering IJ, Liber K. Bioavailability, toxicity and biotransformation of selenium in midge (Chironomus dilutus) larvae exposed via water or diet to elemental selenium particles, selenite, or selenized algae. Environ Sci Technol. 2012;47(1):584–592. | ||
Mal J, Veneman WJ, Nancharaiah YV, et al. A comparison of fate and toxicity of selenite, biogenically, and chemically synthesized selenium nanoparticles to zebrafish (Danio rerio) embryogenesis. Nanotoxicology. 2017;11(1):87–97. | ||
Khiralla GM, El-Deeb BA. Antimicrobial and antibiofilm effects of selenium nanoparticles on some foodborne pathogens. Lebenson Wiss Technol. 2015;63(2):1001–1007. | ||
Rajabi S, Ramazani A, Hamidi M, Naji T. Artemia salina as a model organism in toxicity assessment of nanoparticles. Daru. 2015;23(1):20. | ||
Nair HB, Sung B, Yadav VR, Kannappan R, Chaturvedi MM, Aggarwal BB. Delivery of antiinflammatory nutraceuticals by nanoparticles for the prevention and treatment of cancer. Biochem Pharmacol. 2010;80(12):1833–1843. | ||
Peng D, Zhang J, Liu Q, Taylor EW. Size effect of elemental selenium nanoparticles (Nano-Se) at supranutritional levels on selenium accumulation and glutathione S-transferase activity. J Inorg Biochem. 2007;101(10):1457–1463. | ||
Singh R, Lillard JW Jr. Nanoparticle-based targeted drug delivery. Exp Mol Pathol. 2009;86(3):215–223. | ||
Gelperina S, Kisich K, Iseman MD, Heifets L. The potential advantages of nanoparticle drug delivery systems in chemotherapy of tuberculosis. Am J Respir Crit Care Med. 2005;172(12):1487–1490. | ||
Cihalova K, Chudobova D, Michalek P, et al. Staphylococcus aureus and MRSA growth and biofilm formation after treatment with antibiotics and SeNPs. Int J Mol Sci. 2015;16(10):24656–24672. | ||
Dwivedi S, AlKhedhairy AA, Ahamed M, Musarrat J. Biomimetic synthesis of selenium nanospheres by bacterial strain JS-11 and its role as a biosensor for nanotoxicity assessment: a novel Se-bioassay. PLoS One. 2013;8(3):e57404. | ||
Qin F, Ye Y, Yao X. Effects of nano-selenium on the capability of learning memory and the activity of Se-protein of mice. Wei Sheng Yan Jiu. 2008;37(4):502–504. | ||
Singh N, Saha P, Rajkumar K, Abraham J. Biosynthesis of silver and selenium nanoparticles by Bacillus sp. JAPSK2 and evaluation of antimicrobial activity. Der Pharm Lett. 2014;6(1):175–181. | ||
Tran PA, Taylor E, Sarin L, Hurt RH, Webster TJ. Novel anti-cancer, antibacterial coatings for biomaterial applications: selenium nanoclusters. In: MRS Online Proceedings Library Archive. Boston, MA:Cambridge University Press. 2009. | ||
Wang Q, Webster TJ. Short communication: inhibiting biofilm formation on paper towels through the use of selenium nanoparticles coatings. Int J Nanomedicine. 2013;8:407–411. | ||
Xia MS, Zhang HM, Hu CH. Effect of nano-selenium on meat quality of pigs. J Zhejiang Univ Sci B. 2005;31:263–268. | ||
Yang J, Huang K, Qin S, Wu X, Zhao Z, Chen F. Antibacterial action of selenium-enriched probiotics against pathogenic Escherichia coli. Dig Dis Sci. 2009;54(2):246–254. | ||
Feng Y, Su J, Zhao Z, et al. Differential effects of amino acid surface decoration on the anticancer efficacy of selenium nanoparticles. Dalton Trans. 2014;43(4):1854–1861. | ||
Menter DG, Patterson SL, Logsdon CD, Kopetz S, Sood AK, Hawk ET. Convergence of nanotechnology and cancer prevention: are we there yet? Cancer Prev Res (Phila). 2014;7(10):973–992. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.