Back to Journals » International Journal of Nanomedicine » Volume 20
Nano-Drug Delivery System Targeting the Oxidative Stress Microenvironment: A Prospective Strategy for Intervertebral Disc Degeneration Treatment
Authors Wang Y, Wen P, Zhang BF, Hu S, Yang Z
Received 19 March 2025
Accepted for publication 29 July 2025
Published 25 September 2025 Volume 2025:20 Pages 11717—11780
DOI https://doi.org/10.2147/IJN.S529127
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Xing Zhang
Yidian Wang,* Pengfei Wen,* Bin-Fei Zhang, Shouye Hu, Zhi Yang
Department of Joint Surgery, Honghui Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhi Yang; Yidian Wang, Department of Joint Surgery, Honghui Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi, People’s Republic of China, Email [email protected]; [email protected]
Abstract: Intervertebral disc degeneration (IDD) is the main cause of low back pain (LBP), which imposes a heavy burden on individuals and society. At present, the treatment methods for IDD aim to control clinical symptoms, and it is difficult to fundamentally reverse IDD and reconstruct its mechanical function. Related studies have shown that oxidative stress is a key factor in promoting intervertebral disc (IVD) degeneration and impeding its repair, so antioxidant therapy may be effective in improving or reversing IDD progression. However, considering the non-targeted systemic cytotoxicity and limited bioavailability of antioxidant biopharmaceuticals, it is urgent to establish an efficient drug delivery system. With the development and innovation of nanomedicine and materials science, nano-drug delivery systems (NDDSs) have shown broad application prospects in various diseases. Relying on the characteristics of nanoscale materials, NDDSs can achieve good biodegradability, biocompatibility, targeted drug delivery, and controlled drug release, thereby enhancing the efficacy of antioxidant preparations. This review not only summarizes the research progress of NDDSs in the treatment of IDD, but also evaluates the advantages and challenges faced by various NDDSs. We anticipate that the perspectives articulated in this review will offer valuable insights for the precise management of IDD.
Keywords: intervertebral disc degeneration, oxidative stress, reactive oxygen species, drug delivery system, nanoparticles
Introduction
Low back pain (LBP) is considered as pain or discomfort in the area below the rib margin and above the buttock crease, and it is a common musculoskeletal disorder.1 With the intensification of global population aging, the incidence rate of LBP is on the rise.2 According to relevant studies, the global number of people suffering from LBP was 377.5 million in 1990, which increased to 577 million in 2017.3,4 In low- and middle-income countries, the prevalence of LBP is even higher. Based on limited data, the annual prevalence of LBP in adults is 57% and 67% in Africa and Latin America respectively,5,6 with a lifetime prevalence reaching up to 93%.7,8 LBP is the leading cause of disability, with a 54% increase in disability caused by LBP globally between 1990 and 2015.3,9 The LBP has also brought about a significant economic burden. According to estimates, the direct medical costs for LBP in the UK and the US are approximately £19.77 billion and $100 billion respectively, while in Japan it is 1.2 trillion yen.4,10 Although there are many factors contributing to LBP, IDD is the main cause, accounting for approximately 40% of symptomatic LBP.11,12
 |
Figure 1 The main pathogenic factors of IDD. |
 |
Figure 2 Different developmental stages of IDD and their manifestations. |
 |
Figure 3 Diagnosis and current IDD treatments. |
 |
Figure 4 Redox disorders and their pathogenic mechanisms in degenerative IVD. |
 |
Figure 5 Complex signalling networks in degenerating IVD cells and their interaction with the oxidative stress microenvironment. |
The intervertebral disc (IVD) is a fibrocartilaginous tissue located between the vertebrae, consisting primarily of the nucleus pulposus (NP), annulus fibrosus (AF), and cartilaginous endplate (CEP), which plays a crucial role in maintaining spinal stability, cushioning spinal pressure, and enhancing spinal mobility.13 In the development process of IDD, multiple risk factors are involved in the progression of IDD, including genetic susceptibility, aging, trauma, immune disorders, occupational exposure and abnormal non-physiological mechanical load14–17 (Figure 1). However, the exact pathogenic mechanism of IDD is unknown. Currently, IDD is considered to be a degenerative process involving molecules, cells, tissues and organs mediated by IVD cells in a specific genetic context, which impairs the normal tissue structure and biomechanical properties of the IVD, ultimately destroying its ability to withstand loading.8,18 The latest research indicates that the oxidative stress microenvironment plays a crucial role in the occurrence, development, and inhibition of regeneration in IDD.19 Oxidative stress is caused by an imbalance in intracellular and extracellular redox reactions, which induces cellular senescence, death, and imbalanced metabolism of the extracellular matrix (ECM) through various signaling cascades and intracellular signal transduction pathways, thereby participating in the progression of IDD.19 Current treatments for IDD are pharmacological and surgical to alleviate symptoms and reduce the incidence of disability, but both have the disadvantages of high complications, high costs and unknown efficacy.20 These methods can only act on the terminal stage of IDD and cannot delay or reverse the progression of IDD by improving the pathological changes of IVD. In addition, some recently emerging preclinical treatment methods, such as small molecule drugs, proteins, and nucleic acids, have shown excellent therapeutic effects in improving the survival and functional impairments of IVD cells.21 However, the short half-life and rapid elimination of these drugs limit their treatment efficacy.11 In this regard, approaches based on nano-drug delivery systems (NDDSs) may be promising for the treatment of IDD.22 Currently, NDDSs are mainly involved in the delivery of antioxidant agents through physical encapsulation (liposomes and exosomes, etc.) or chemical binding [inorganic and polymeric nanoparticles (NPs), etc.].23 Among them, some NDDSs possess excellent antioxidant capacity by themselves (polydopamine NPs, MnO2 NPs, fullerenes, and fullerols), and their combination with antioxidant agents can exert stronger synergistic antioxidant effects.24 Secondly, NDDSs also have the ability to control release, which mainly depends on physical conformation changes or chemical bond breaks of nanocarriers.25 In particular, some carriers can respond to internal (ROS, pH, enzymes, etc.) or external stimuli (temperature, light, magnetic field, etc.) to achieve precise controlled release of drugs in time and space.25,26 In addition, NDDSs can increase drug accumulation at the lesion through passive and active targeting capabilities, thereby improving efficacy and reducing side effects.27 These properties make NDDSs an effective method for improving the oxidative stress microenvironment within degenerating IVDs.
The aim of this review is to briefly review the changing pathophysiology of IDD and the limitations of current treatments. In addition, we also emphasized the oxidative stress microenvironment in degenerative IVDs and its role in IDD. Importantly, we demonstrated that biomedical engineering approaches can be used to ameliorate the oxidative stress microenvironment of IDD, thereby promoting regeneration and recovery of IDD, which may help advance the development of NDDSs in the treatment of IDD.
The Pathophysiology of IDD
Normal Structure and Function of IVD
The IVD is an integral part of the spinal tissue and biomechanical composition. The normal spine comprises a minimum of 23 highly hydrated IVDs that connect adjacent vertebrae, playing a crucial role in enhancing spinal mobility and optimizing load distribution.28 The complete IVD comprises NP, AF, and CEP on both sides.
NP is located between the center and posterior of IVD, serving as the core of IVD.29 In general, NP is a soft and elastic gel-like substance. The healthy IVD typically exhibits a relatively high-water content, ranging from approximately 80% to 90%, whereas degenerated IVDs demonstrate a reduction in water content to around 70–75%, or even lower.30,31 In terms of cellular composition, NP cells are a group of mixed cells at different stages of maturation, used to maintain the metabolic balance of ECM.4 Before the age of 10, the mixed cell population consisted of larger vacuolated notochord cells (NC) (mainly expressing CK-8, LGALS3, STMN2, RTN1, PRPH, CXCL12, IGF1, MAP1B, ISL1, CLDN1 and THBS2) and smaller chondrocyte-like cells (mainly expressing HIF1α, GLUT1, proteoglycans, type II collagen, KRT18/19, CA12 and CD24).31–34 In adulthood, these cells all differentiate into chondrocyte-like cells with reduced metabolic activity.35 Research has shown that NC contributes to the regulation of proteoglycan production and proliferation activity in IVD chondrocytes, suggesting a potential association between NC degradation and IDD.36 In terms of the organization composition of NP, 35–65% is composed of proteoglycans, 5–20% is composed of collagen II (Col II) and elastin protein.37 The high specific gravity and negative charge properties of proteoglycans contribute to the highly hydrated characteristics of NP tissue, while also facilitating the dispersion of loads.11,38 The mesh composed of Col II and elastic protein fibers is embedded in the gel formed by proteoglycans to maintain the elasticity of ECM.39
The AF is located on the periphery of NP, consisting of well-arranged concentric rings or lamellae composed of collagen fibers and proteoglycans.4 The lamellae are inclined approximately 30° from one vertebra to another and cross diagonally at an angle of >60°between adjacent lamellae, which helps to limit rotation and bending between adjacent vertebrae while providing sufficient resistance to axial loads.37,40 The AF can be further divided into an inner fibrocartilage region and an outer fibrous region, which have different cellular and tissue compositions.21 The inner region mainly consists of circular chondrocyte-like cells that produce proteoglycans and Col II, while the outer region contains elongated fibroblast-like cells that primarily produce Col I.21,41 Correspondingly, the organizational structure of both the internal and external regions has also undergone excessive changes. AF tends to gradually lose proteoglycans, Col II, and water content from the inside out, resulting in a large amount of Col deposition, which effectively restricts NP during axial compression, extension, and bending.42–44
The endplate is the boundary between the IVD and the adjacent vertebral bodies, including the bony endplate and CEP. The main cell type within the CEP is the chondrocyte, which primarily secretes Col II and proteoglycans.45 Compared to NP, the proportion of proteoglycans and Col II in CEP is relatively low, which can provide a solid mechanical barrier between IVD and vertebral body, thereby preventing the protrusion of NP tissue into the trabecular bone of the vertebral body.46,47 In addition, CEP is also the main channel for IVD to exchange substances with the external environment.48
The IVD is the largest avascular tissue in the body.49 Compared to the lifelong avascularity in NP, CEP and AF have abundant blood vessels during fetal and infant stages.50 However, with age, the blood vessels in CEP will completely regress, while only the outer layer of AF maintains vascularization.51 The limited distribution of blood vessels in the IVD primarily facilitates substance exchange through diffusion (for small molecular solutes such as glucose, oxygen, and lactate) or convection (for large molecular solutes such as growth factors, hormones, and proteoglycans).52,53 Therefore, IVD is in a hypoxic microenvironment. According to the study of canine IVD samples, the oxygen tension in the IVD (0.53–1.06 kPa) is lower compared to that in muscle (3.8±0.8 kPa) and brain (3.4±0.2 kPa), and it decreases as it gets closer to the center of the IVD.54,55 The hypoxic microenvironment makes NP cells partially dependent on anaerobic glycolysis to produce ATP, which contributes to the formation of an acidic microenvironment.56,57 Regarding the innervation within the IVD, the NP has no innervation, whereas the outer layer of the AF is innervated by branches of the sinus vertebral nerve.58
Degenerative IVD: Molecular, Tissue and Mechanical Functions
IDD is an age-related degenerative process that is influenced and accelerated by other unfavourable factors, such as genetic and environmental factors.59 The process of degeneration can be divided into three closely connected stages (Figure 2). Firstly, under specific genetic backgrounds, abnormal mechanical load induces functional impairment of IVD cells, leading to an imbalance in ECM metabolism. Subsequently, the microenvironment of IVD continued to deteriorate, leading to further decrease in the number and activity of IVD cells, resulting in a cascade reaction of damage. In the later stage, IVD loses its mechanical function and is accompanied by infiltration of nerves and blood vessels, eventually leading to IVD-related LBP.
Although the traditional view holds that IDD is an age-related disease, it cannot be denied that genetic factors may play a significant role in determining this condition.60 The existence of genetic susceptibility to IDD is supported by preliminary evidence from twins and family studies. In 1999, Sambrook et al evaluated the MRI characteristics of cervical and lumbar IVD in 172 identical twins and 154 dizygotic twins.61 The results indicated a heritability rate of 74% for lumbar IVD degeneration and 73% for cervical IVD degeneration. Another prospective study involving 116 identical twins found that the estimated heritability of IDD within 5 years was between 47–66%.62 In addition, in a 10-year longitudinal study involving 234 pairs of twins, Williams et al found that disc herniation has a genetic component across all age groups.63 Recently, genome-wide association studies (GWAS) have made significant contributions to the etiology of complex diseases and have indicated that genetic susceptibility to IDD may be widespread.64–66 In 2022, Bjornsdottir et al conducted a (58,854 cases, 922,958 controls) GWAS study on IDD and identified 41 genetic variants in 33 loci that were associated with cartilage, bone biology, and inflammatory processes.67
In fact, IVD had already undergone degradation in the first decade of life.68 The reduction of vacuolar NCs in the NP is considered to be the initiation process of IVD degeneration, because NCs can increase the anabolic and proliferative activity of NP cells.36,69,70 In all cases, early IDD is mainly characterized by an imbalance between anabolic and catabolic processes in the ECM, which is mainly caused by a phenotypic transition of IVD cells.71,72 In terms of anabolism, there was a significant reduction in the production of proteoglycans, along with a shift in the composition of their side chains from chondroitin sulphate to keratan sulphate, which led to diminished hydration of NP tissues.73,74 The balance of collagen production shifts from Col II to Col I, accelerating the process of IVD fibrosis.74 In terms of catabolism, the expression of matrix metalloproteinase (MMP)-1, MMP-2, MMP-3, MMP-13, MMP-14, a disintegrin and metalloproteinase with thrombospondin motifs (ADAMTS)-4 and ADAMTS-5 significantly increased.74–76 In contrast, the levels of tissue inhibitor of metalloproteinase (TIMP)-1 and TIMP-2 were decreased.76 These factors have led to a further decline in the quality of ECM.
Early IVD degeneration lasts for a considerable period of time (10 or 20 years or more), which may be accelerated by other unfavorable factors (genetics, trauma and abnormal loads) and enters the radical phase. During the radical phase, the microenvironment within the IVD is severely deteriorated, accompanied by inflammation and oxidative stress storms.77 NP cells secrete various cytokines, including tumor necrosis factor α (TNF-α), interleukin-1 β (IL-1β), IL-6, IL-8, IL-17 and prostaglandin E2 (PGE2).47,78 Simultaneously, NP cells stimulate the synthesis of chemokines, specifically CCL2, 3, 4, 5, 7, 10 and 13, which activate and recruit immune cells to infiltrate the IVD.22,32 This further exacerbates the cellular inflammatory response. In addition, degenerating IVDs undergo intense local oxidative stress, with large amounts of reactive oxygen species (ROS) being generated.19 Excessive ROS can attack the mitochondrial respiratory chain, endoplasmic reticulum, and inhibit the production of reductases, further promoting the accumulation of ROS.19,79,80 Inflammatory response and oxidative stress promote each other, thus forming a positive feedback loop and acting directly or indirectly on a dynamic signaling network with complex feedback loops composed of different signaling pathways.72,81,82 Aberrant activation or inhibition of these signaling networks may exacerbate NP dysfunction, including apoptosis, senescence, hypertrophy and calcification, which in turn exacerbate the imbalance of ECM synthesis and degradation.18,83 In addition, hypertrophy and calcification of CEP cells and sclerosis and occlusion of the bone marrow lumen will lead to loss of CEP permeability, which will result in impaired energy supply and accumulation of metabolic wastes within the IVD, ultimately accelerating IDD.84–86
When entering the late stage of IDD, IVD will gradually lose its mechanical function and result in IVD-related LBP. At this time, the normally active NP cells are almost completely damaged, and proteoglycan and Col II are replaced by Col I.87 As a result, the highly hydrated and elastic jelly-like NP is replaced by stiff fibrous tissue, which results in an inability to effectively distribute the load.87 Meanwhile, the imbalance in ECM metabolism leads to structural disorder in AF, resulting in a significant reduction in its ability to resist loads and the occurrence of cracks.87 Once the integrity of AF is compromised, internal NP tissue is exposed to the circulatory system.88 The immune system will recognize these “foreign” tissues and generate an autoimmune response.89 IgG antibodies and antigen-antibody complexes against proteoglycans and collagen were found in human degenerative IVD samples, providing direct evidence for autoimmune reactions.90,91 In addition, NP cells and immune cells secrete a large amount of IL-1, TNF-α, IL-8, and PGE2, which can stimulate the production of brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), and vascular endothelial growth factor (VEGF).92–94 These growth factors induce infiltration of blood vessels and nerves, leading to IVD-induced LBP.94,95 Moreover, the herniated NP can exert direct compression on the adjacent nerve roots, resulting in significant low back and leg pain.96 Meanwhile, the dysfunctional IVD causes spinal instability, leading to degeneration of surrounding small joints, formation of bone spurs, and hypertrophy of surrounding ligaments.97,98 This in turn results in spondylolisthesis, spinal stenosis, and even degenerative scoliosis.98
Diagnosis and Treatment of IDD
The initial diagnosis of IDD is mainly based on the patient’s symptoms and physical signs. Patients with IDD typically present with LBP. This type of pain is often characterized by an unclear location, primarily concentrated in the central region of the lower back or adjacent to the IVD, and worsens after prolonged standing or sitting. When the protruding NP directly compresses or the inflammatory response irritates the nerve root, LBP may worsen and radiate to one or both limbs. In addition to clinical symptoms and signs, the diagnosis of IDD is mainly based on X-ray, CT scan or MRI, and its severity is further assessed by IVD hydration status (Pfirrmann) or CEP Modic changes.99,100
Currently, the treatment of IDD includes conservative treatment and surgical treatment according to the patient’s symptoms (Figure 3). Conservative treatment includes drug therapy and non-drug therapy, which are mainly suitable for early-stage IDD patients and can alleviate symptoms and improve quality of life. When the degeneration is mild, bed rest, appropriate and regular exercise, weight loss, and physical therapy (such as cold compresses, electrical stimulation, ultrasound therapy, and traction) are simple and effective measures to improve symptoms.101–103 These methods are designed to reduce stress on the IVD, strengthen the low back muscles and improve spinal stability. When patients experience significant pain in the lower back, medication can be used for treatment, including nonsteroidal anti-inflammatory drugs (NSAIDs), opioid analgesics, muscle relaxants, benzodiazepines, antidepressants, corticosteroids and antiepileptic drugs.102 For some advanced patients, some invasive methods can be used, such as chemical nucleolysis, radiofrequency ablation, nucleotomy and nerve root blockade.21 Once conservative treatment fails, symptoms of nerve root compression are severe, cauda equina syndrome and severe spinal deformity or instability occur, surgical treatment is required. Surgical treatment mainly consists of interbody fusion, partial or total IVD removal and total disc replacement (TDA).103 Among them, interbody fusion has always been regarded as the surgical gold standard for symptomatic IVD protrusion and is suitable for the majority of patients with disc herniation.103,104 TDA has similar clinical results to interbody fusion, but is usually reserved for single-level situations without facet joint disease.104–106
Oxidative Stress and IDD
Oxidative Stress in IDD
ROS are a class of unstable and highly reactive molecules, including superoxide anion (O2−), hydroxyl radical (OH-), hydrogen peroxide (H2O2), and hypochlorite ion (OC1−).107 In addition, some nitrogen-containing compounds (nitric oxide and nitrogen dioxide) are also considered as members of the ROS superfamily due to their similar effects.108 As a byproduct of aerobic metabolism, the generation of ROS in cells is inevitable Although IVD cells are in a hypoxic environment due to lack of direct blood supply, aerobic metabolism is still an important way for IVD cells to obtain energy and thus generate ROS.80 In order to maintain redox balance, the human body has developed a set of ROS scavenging systems, which remove excess ROS to keep them at a level that is harmless to the organism.109 In normal levels, ROS is an important intracellular signaling molecule that participates in various physiological processes through intracellular signal transduction.110 As IVD degeneration progresses, aging, trauma, abnormal mechanical load, and smoking cause an imbalance between oxidants and antioxidants, resulting in oxidative stress, which in turn leads to damage to molecules, cells, and tissues111 (Figure 4). At the molecular level, biomacromolecules such as proteins, nucleic acids, lipids, and carbohydrates in cells are inevitably exposed to ROS and suffer from oxidative damage.111 Currently, there are reports indicating a significant elevation of advanced oxidative protein products (AOPPs), 8-oxoguanine (an oxidative product derived from guanine), malondialdehyde (MDA), and advanced glycation end products (AGEs) in degenerative human IVD tissue.112–115 At the cellular level, large amounts of ROS lead to restricted survival and altered metabolic phenotype of IVD cells by damaging cellular macromolecules, which in turn accelerates IDD progression.79 At the tissue level, oxidative stress causes damage to normal IVD tissue structure and biomechanical dysfunction, which in turn leads to corresponding clinical symptoms.116
Redox Imbalance Within the IVD
Increased ROS Production in Degenerative IVD
Mitochondria are the center of oxidative metabolism and the main site of ROS production, because 1–2% O2 is converted into ROS during oxidative phosphorylation, which depends on the mitochondrial electron transport chain (ETC).117,118 Indeed, under physiological conditions, 0.2–2% of the electrons do not follow the normal ETC transfer sequence (complex I/III/IV and complex II/III/IV with succinate as substrate), but rather leak directly from the ETC and interact with O2 to produce O2 -.119 The complexes I and III are the main sites where electrons transfer to O2 to generate O2-.120 For complex I, ROS are thought to originate from reduced flavin mononucleotide or N-1a and N-1b iron-sulfur clusters, whereas ROS for complex III are thought to originate from ubiquinone oxidation sites.121,122 When the activities of complex I and complex III are inhibited or the concentration of succinate is low, complex II becomes the main source of ROS.42 Interestingly, under hypoxic conditions, complex II switches its catalytic activity from succinate dehydrogenase to fumarate reductase, and this change is associated with increased ROS generation.123,124 Therefore, it is possible that complex II has a unique association with increased ROS production in degenerative IVD. In degenerative IVD, the adverse microenvironment can lead to abnormal opening of mitochondrial permeability transition pores (mPTP) in IVD cells, decreased membrane potential, and damage to the respiratory chain, ultimately leading to mitochondrial dysfunction.125,126 Mitochondrial dysfunction leads to massive electron leakage, which increases ROS production.127 A large amount of ROS attacks mitochondrial DNA, proteins, and membrane lipids, which further exacerbates mitochondrial damage, thus forming a vicious cycle.127,128
The NADPH oxidase (Nox) family comprises seven isoforms (Nox1, Nox2, Nox3, Nox4, Nox5, Duox1, and Duox2), which exhibit distinct expression patterns across various cell types and subcellular compartments.129 Nox generates O2− or H2O2 by catalyzing NADPH electron transfer.130,131 The significant increase in the NADPH/NADP+ ratio in degenerated IVD cells suggests that NOX is involved in the formation of the oxidative microenvironment in IDD.132,133 In IDD, the most extensively studied are Nox 2 and Nox 4.134–136 Chang et al found that 15% high cyclic stretch upregulated Nox 2 expression in AF cells, which led to a significant increase in ROS levels.137 In addition, 20% O2 increases the production of ROS by upregulating Nox4 in NP cells, thereby activating the p53-p21-Rb and p16-Rb pathways to induce cellular senescence.138 The endoplasmic reticulum (ER) is also an important source of ROS.139,140 Under the influence of nutritional deficiency, oxidative stress, and genetic mutations, the accumulation of unfolded/misfolded proteins leads to ER stress, which in turn activates the unfolded protein response (UPR).141,142 This phenomenon is widely observed in IDD.142,143 During the UPR process, the involvement of protein disulfide isomerase, endoplasmic reticulum oxidoreductin-1 and Nox complex can trigger ROS cascade reactions.42 In addition, ER stress leads to Ca2+ overload and increases intra-mitochondrial Ca2+ levels through the mitochondria-associated membrane (MAM), which in turn leads to mitochondrial dysfunction and increased ROS generation.139,144 In addition to the above pathways, xanthine oxidase (XO), uncoupled endothelial nitric oxide synthase, cytochrome P-450 monooxygenases, and cyclooxygenases are also sites of ROS generation,42,120 collectively contributing to endogenous ROS accumulation. In addition to endogenous pathways, external stimuli such as smoking, UV radiation, ionizing radiation, air pollution and drugs can also lead to the accumulation of ROS.145–147
Reduced Antioxidant Defense in Degenerative IVD
The complete intracellular antioxidant defence system mainly consists of antioxidant enzymes, non-enzymatic small molecules and redox signal transduction.116 Antioxidant enzymes are the first line of defense system, mainly including superoxide dismutase (SOD), glutathione peroxidase (GPX), catalase (CAT), and methionine sulfoxide reductase (Msr).148,149 SOD is an important antioxidant enzyme, which can be classified into Cu/Zn-SOD based on the types of metal ions it binds to.150 These two enzymes are prevalent in the mitochondria of NP cells and alleviate oxidative stress by efficiently catalysing the conversion of O2− to H2O2 and O2.151,152 Similarly, GPX and CAT are members of the antioxidant defence within the IVD, both of which can catalyze the decomposition of H2O2 into H2O.151 However, multiple studies have indicated a significant decrease in the expression of SOD, GSH, and CAT in degenerated IVD tissues and cells.153–155 In addition, Msr is an repair enzyme that can remove ROS by reducing the methionine residues in oxidized proteins.156 The expression of Msr is significantly reduced in aging AF cells, which makes AF cells vulnerable to oxidative stress damage.157 Following antioxidant enzymes, non-enzymatic small molecules are a class of compounds that do not rely on the enzyme system in the organism to scavenge free radicals and combat oxidative stress, and mainly include vitamins C and E, β-carotene, lipoic acid, ubiquinone, melatonin (MT), carotenoids, ascorbic acid and uric acid.97,158 They are widely present in the cytoplasm of IVD cells, and the reduction of their expression levels can also lead to oxidative stress in IVD. In addition to the classical antioxidant system, intracellular redox signaling is also an important way to maintain intracellular redox homeostasis, which mainly includes the Kelch-like ECH-associated protein 1 (Keap1) - Nuclear factor E2- related factor 2 (Nrf2) - Antioxidant response element (ARE) signaling pathway.159 When the intracellular ROS levels increase, Keap1 dissociates from CUL-E3 ligase, causing a conformational change in Keap1 and releasing Nrf2, which leads to the accumulation of Nrf2 in the cytoplasm and subsequent nuclear translocation.160 High levels of Nrf2 bind to the ARE, which activates the transcription of antioxidant genes, including heme oxygenase-1 (HO-1), GSH, SOD, and CAT, and thus exerts antioxidant effects.116,160 I In human degenerated IVD and aged NP cells, Nrf2 expression levels were significantly decreased and correlated with the severity of IDD.161,162 In addition, high levels of Nrf2 promote mitochondrial biosynthesis by up-regulating the expression of nuclear respiratory factor 1/2 (NRF-1/2), PGC-1α, and mitochondrial transcription factor A, as well as mitochondrial fusion by degrading the mitochondrial fission protein dynamin-related protein 1 (Drp1), which contribute to protection against oxidative stress.163,164
Role of Oxidative Stress in IDD
ROS is an important signaling molecule within cells, which means it can directly or indirectly interact with dynamic signal networks composed of different signaling pathways and complex feedback loops, thereby regulating the physiological and pathological processes of IDD151 (Figure 5). During this process, there are various phenotypic changes in IVD cells, which accelerate the progression of IDD.
Oxidative Stress and Cell Death
The balance of anabolic and catabolic metabolism within the IVD is dependent on the metabolic capacity conferred by a sufficient number of cells and normal cellular function. When excessive cell death occurs within the IVD, this balance is disrupted and promotes the progression of IDD. ROS are potent inducers of cell death, and here we focus on three cell death types, including apoptosis, pyroptosis, and ferroptosis.165,166
Apoptosis is a controlled cell death that includes two major pathways: intrinsic and extrinsic,167 the former of which is often associated with mitochondrial dysfunction. Intrinsic apoptosis depends on the abnormal activation of BCL-2 family proteins on the mitochondrial membrane,168 which leads to increased mitochondrial outer membrane permeability and leakage of cyt-c, ultimately inducing cell apoptosis by activating a series of caspases.169 ROS is involved in IVD cell apoptosis through the regulation of multiple signaling pathways, including phosphatidylinositol 3-kinase (PI3K)/protein kinase B (PKB/Akt), NF-κB, mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK), Sirtuin (SIRT) protein family, Keap1/Nrf2, and AMP-activated protein kinase (AMPK).170–173 The activation or inhibition of these signaling pathways leads to the differential expression of pro-apoptotic proteins (such as Bak, Bax, and Bok) and anti-apoptotic proteins (such as Bcl-XL, Bcl-2, Bcl-w, and Mcl-1), thereby inducing cell apoptosis.168 For example, when IVD cells were co-cultured with oxidants (H2O2 and TBHP) in vitro, a decrease in the expression of PI3K/Akt,174 AMPK,175 SIRT,114 and Nrf2162 and an increase in the expression of MAPK/ ERK176 and NF-κB177 were observed. This resulted in up-regulation of BAX, Caspase-9 and Caspase-3 expression and down-regulation of Bcl-2 expression, which in turn induced apoptosis. In fact, these signaling pathways are not isolated but intertwined with each other to form positive or negative feedback loops. For example, when the PI3K/Akt signaling pathway is activated, Akt can phosphorylate and inhibit the degradation of IκBα (a member of the IκB protein family and a specific inhibitor of NF-κB), thereby suppressing the activation of NF-κB.178 In specific circumstances, NF-κB can in turn promote the activation of Akt, forming a positive feedback loop that enhances cell survival capability.179 In addition, the roles of these signaling pathways in cell apoptosis are diverse and may depend on the cell type, stimulus type, and intracellular environment.180
Pyroptosis is a type of programmed cell death between apoptosis and necrosis that depends on the activation of inflammasomes.181 Inflammasomes are a class of protein complexes, the most widely studied of which is the NLRP3 inflammasome,182 which can sense pathogen-associated molecular patterns or damage-associated molecular patterns and then recruit and activate Caspase-1.182 The activated Caspase-1 can cleave and activate other proteins, such as Gasdermin D (GSDMD), IL-1β, and IL-18.183 After being cleaved, the N-terminal fragment of GSDMD forms pores on the cell membrane, leading to leakage of cellular contents and triggering a strong inflammatory response.184 Compared with non-degenerated cells, high levels of ROS and caspase-1 were observed in degenerated human IVD cells.185 In vitro, after NP cells were co-cultured with H2O2, the expression of ROS, NLRP3 inflammasome and caspase-1 in the cells was significantly upregulated, indicating that oxidative stress can effectively induce cell pyroptosis.186 In fact, oxidative stress is involved in the induction of cell death through various mechanisms, including activation of inflammasomes, mitochondrial dysfunction, enhanced inflammatory response, and instability of cell membrane.187 ROS can directly promote the activation of NLRP3 inflammasome, with mitochondrial ROS (mtROS) being the most important signal for NLRP3 inflammasome activation,188 which has also been demonstrated in cardiovascular disease.189 In addition, ROS can damage IVD cell mitochondria, leading to abnormal opening of mPTP and leakage of mitochondrial DNA (mtDNA).190 Released mtDNA promotes cell pyroptosis by activating the Toll-like receptor 9 (TLR9)-NF-κB-NLRP3 axis.191 Recent studies have also shown that thioredoxin-interacting protein (TXNIP) is a crucial bridging molecule for the activation of NLRP3 inflammasome.192 When ROS is overloaded in IVD cells, TXNIP dissociates from Thioredoxin (Trx) and binds to NLRP3, thereby initiating cell pyroptosis, but this process is inhibited by Nrf2 signaling.186,193 A possible explanation is that Nrf2 inhibits TXNIP function by up-regulating antioxidant proteins and regulating the balance of Trx/TXNIP complexes.194–196 In addition, the SIRT protein family is involved in the regulation of cellular pyroptosis.197 Overexpression of SIRT1 can improve IL-1β-induced mitochondrial dysfunction and NLRP3 inflammasome activation in NP cells by promoting PINK1/Parkin-mediated mitophagy, thereby alleviating NP cell pyroptosis.188
Ferroptosis is a form of iron-driven cell death characterized by intracellular iron accumulation and oxidative stress, resulting in lipid peroxidation and membrane damage.198 In fact, there is a vicious cycle of mutual promotion between oxidative stress and iron overload.199 Iron is a catalyst for many oxidation reactions, especially the Fenton Reaction, in which Fe2+ can combine with H2O2 to form OH-, accelerating the accumulation of intracellular ROS.200 Furthermore, iron overload can induce the depletion of antioxidant defense systems, rendering cells more vulnerable to oxidative stress-induced damage.201,202 On the other hand, oxidative stress can exacerbate iron overload. Oxidative stress can lead to the oxidation modification of ferritin, weakening its normal iron storage capacity and exacerbating the accumulation of iron within cells.203 Compared with the normal control group, the expression of GPX4 and ferritin in degenerated IVD tissues was significantly decreased, accompanied by increased oxidative stress and ferroptosis.204 In vitro, co-culture of oxidants with IVD cells also leads to oxidative stress, lipid peroxidation and iron overload in a dose-dependent manner.204–206 Multiple molecules and signaling pathways are involved in regulating iron death within the IVD. Ferroportin (FPN) is an iron efflux protein responsible for the translocation of intracellular iron to the extracellular space.207 In TBHP-induced human NP cells, FPN is severely dysfunctional, which leads to intercellular iron overload and ferroptosis.208 Nrf2 is also an important signaling molecule that regulates ferroptosis in IVD cells.194 In vitro, Nrf2 activation can improve Parkin-mediated mitophagy, which helps to alleviate mitochondrial dysfunction and oxidative stress, thereby inhibiting ferroptosis.209 Similarly, Nrf2 can also upregulate the expression of HO-1 and inhibit the nuclear translocation of NF-κB, thereby alleviating oxidative stress, inflammatory response, and ferroptosis in IVD cells.84,210,211 In addition, SIRT3 can also alleviate oxidative stress-induced ferroptosis in IVD cells by upregulating the expression of antioxidant genes HO-1, NQO1, SOD2, and GPX4.212
Oxidative Stress and Cellular Senescence
Cellular senescence is the process by which cells gradually lose their function and proliferative capacity during their life cycle, including replicative senescence and stress-induced senescence.213 The replicative senescence is caused by the shortening of telomeres and exacerbated by factors that promote the accumulation or inhibit the breakdown of cellular aging, with ROS being the most important factor inducing cell senescence.113 In both human and rat degenerated IVD tissues, the proportion of senescence-associated β-galactosidase (SA-β-gal) positive cells significantly increases, which is positively correlated with oxidative stress levels within the IVD.214 Cellular senescence not only leads to a decrease in the number of functionally normal IVD cells, but also deteriorates the IVD internal microenvironment through the paracrine effect of senescence-associated secretory phenotype (SASP).16 These SASP-associated proteins include ECM-degrading enzymes (MMPs and ADAMTS), pro-inflammatory cytokine factors (IL-1β, TNF-α, IL-7, IL-8), chemokines, and other biologically active substances, which can further exacerbate the mitochondrial dysfunction and the production of ROS, thus generating a vicious circle.39,215 More importantly, high levels of SASP-associated MMPs have been shown to induce autocrine ligand shedding, which renders senescent cells resistant to immune surveillance and clearance.216 Multiple signaling molecules are involved in oxidative stress-induced senescence of IVD cells. For example, ROS can promote the ubiquitin-dependent degradation of IκB through ROS-Hsp27-IκB kinase (IKK), leading to increased nuclear translocation of NF-κB (such as p65) and subsequent activation of downstream target genes to regulate cellular functions.19,217 Compared with normal NP cells, the p65 level in the nucleus of oxidant-induced degenerative NP cells increased, and induced cell senescence by regulating p53-p21-Rb and p16-Rb.218,219 The PI3K/Akt signaling pathway is an important intracellular signaling pathway that participates in regulating cell apoptosis, aging, growth, and metabolism.220 When the PI3K/Akt signaling pathway is activated, it activates downstream signaling molecules to alleviate mitochondrial dysfunction and intracellular ROS levels, while downregulating the p53-p21-Rb signaling axis to mitigate cellular senescence in IVD cells.174,220 Recently, Zhang et al found that high concentrations of lactic acid can induce senescence and oxidative stress in NP cells.221 Mechanistically, lactate can bind to lysine 39 and leucine 52 residues in the Akt PH domain to inhibit Akt kinase activity, thereby inducing NP cell senescence and oxidative stress by regulating p21/p27/cyclinD1 and Nrf2/HO-1 signaling. In addition, the SIRT protein family is also involved in regulating cellular aging in IVD. Activation of SIRT1 has been reported to ameliorate oxidative stress-induced cellular senescence and ECM degradation.222,223 Similarly, several studies have found that SIRT3 activation can improve mitochondrial function and SOD activity, and alleviate IVD cell senescence by inhibiting p16-Rb signaling.224,225 In addition, SIRT2 overexpression can also inhibit oxidative stress by upregulating the expression of SOD1/2 and inhibit NP cell senescence by downregulating the level of p53-p21-Rb.226
Oxidative Stress and Cellular Autophagy
Autophagy is an evolutionarily conserved self-degradation system that degrades and recycles intracellular proteins and damaged organelles to maintain cell homeostasis and function.227 Compared with healthy IVD tissues, the expression levels of autophagy-related genes (Atg) in degenerated IVD tissues were significantly changed, indicating that autophagy plays an important role in IDD.228,229 As a stress response system, almost all stress factors that affect cellular homeostasis can induce autophagy. In recent years, an increasing amount of evidence has shown that ROS is a crucial intracellular signaling molecule that affects autophagy.230,231 However, the relationship between autophagy and oxidative stress is not a simple upstream-downstream signaling regulation relationship, but rather a complex network composed of numerous signaling pathways and crossroads. Mammalian target of rapamycin (mTOR) is a serine/threonine protein kinase that responds to nutrient levels and growth signals.82 It is a negative regulator of autophagy initiation and its activity is regulated by PI3K-Akt, AMPK and MAPK.232–234 Excessive ROS down-regulated the ratios of p-PI3K/PI3K, p-Akt/Akt and p-mTOR/mTOR, resulting in the inhibition of the PI3K/Akt/mTOR signaling pathway, which in turn induced autophagy in NP cells.235 In addition, ROS can also downregulate the level of p-AMPK, leading to the downregulation of ULK1 phosphorylation and upregulation of mTOR phosphorylation, thereby inhibiting NP cell autophagy activation and autophagic flux.235 Interestingly, under oxidative stress conditions, the upregulation of p38MAPK phosphorylation promotes mTOR phosphorylation, thereby inhibiting autophagy in NP cells.236 In addition to being regulated by upstream signaling molecules, mTOR can also regulate autophagy through its target genes. Downregulation of mTOR signaling can lead to increased nuclear translocation of EB transcription factor (TFEB) and downregulation of p70S6K levels, thereby enhancing cellular autophagy to combat oxidative stress.237–239 Mitophagy is a type of cellular autophagy that plays a key role in maintaining mitochondrial quality control.240 PINK1/Parkin signaling pathway is an important signaling axis regulating mitophagy.116 In NP cells, upregulation of the PINK1/Parkin signaling pathway induces mitophagy, thereby delaying the progression of IDD by eliminating damaged mitochondria.162,188 In addition, the SIRT protein family and Nrf2 can also regulate mitochondrial autophagy in IVD cells.241,242
Oxidative Stress and Inflammation
The inflammatory response is a defence mechanism against pathogens, damaged cells or specific stimuli, which is induced by a stressor and leads to the expression of inflammatory cytokines through a series of intracellular signal transduction.243 As a complex immune response, inflammatory response can maintain tissue homeostasis by eliminating pathogens and repairing damaged tissues. However, excessive activation and persistence of inflammatory responses may lead to tissue damage and a variety of diseases. ROS can act as signaling molecules to activate the expression of inflammation-related genes, thereby initiating or exacerbating inflammatory responses.244 In vitro, co-culture of rat NP cells with H2O2 can lead to the expression of pro-inflammatory cytokines, such as IL-1β, IL-6, TNF-α, and nitric oxide synthase (iNOS).245 In addition, ROS can cause damage to intracellular biomacromolecules and organelles (such as mitochondria), which will trigger the activation and assembly of NLRP3, further aggravating the inflammatory response.246,247 ROS regulates inflammatory responses through a variety of signaling molecules, including PI3K/Akt, MAPK, Nrf2, and NF-κB, among which MAPK and NF-κB pathways play the most important roles.248,249 ROS can activate MAPK kinase, which in turn phosphorylates and activates downstream MAPK, leading to the expression of pro-inflammatory cytokines in IVD cells.249,250 In addition, oxidative stress can directly or indirectly activate NF-κB, promoting the expression of inflammatory genes such as IL-1, IL-6, TNF-α in IVD cells and exacerbating the inflammatory response.251,252 Cell damage caused by oxidative stress can trigger an inflammatory response, while inflammation can also exacerbate oxidative stress, forming a vicious cycle.19 For example, TNF-α, IL-1β and lipopolysaccharide (LPS) co-cultured with NP cells can lead to oxidative stress by promoting intracellular ROS production and inhibiting the expression of SOD1, SOD2, CAT and GPX3.248,253,254
Oxidative Stress and ECM Degradation
The complete biomechanical function of IVD depends on the integrity of its tissue structure. With the progression of IVD degeneration, there have been significant changes in the quality and composition of ECM. Due to the death and aging of a large number of IVD cells, the synthesis of ECM is significantly reduced.255 At the same time, the catabolic phenotype of IVD cells is enhanced, which further exacerbates the metabolic imbalance of ECM.139 As an important stressor, ROS can regulate cellular behavior through multiple signaling pathways, thereby influencing the expression and secretion of ECM proteins.77 According to the current study, excessive ROS can downregulate PI3K/Akt, SIRT, AMPK and Nrf2 and upregulate the expression of NF-κB and MAPK, which in turn promotes the degradation of ECM.18,151,225,256 In addition, ROS can disrupt the components of ECM through oxidation, such as causing cross-linking and degradation of collagen protein, affecting its structural stability and functional integrity.257 In conclusion, oxidative stress can lead to abnormal ECM metabolism and oxidative damage of related molecules, promoting the progression of IDD.
Oxidative Stress and Epigenetics
Epigenetics refers to the mechanisms that can influence gene expression without altering the DNA sequence, and these changes can be passed on to future generations.258 Common epigenetic regulations include DNA methylation, histone modification, and non-coding RNA (ncRNA) regulation.259 These changes not only play an important role in the normal physiological processes of cells, but are also involved in driving the pathological processes of various diseases.260 Oxidative stress and epigenetic regulation have a bidirectional control relationship and are involved in the progression of IDD.
DNA methylation is a crucial epigenetic modification involving the addition of a methyl group at specific positions on the DNA molecule, predominantly occurring at the 5′ carbon position of cytosine (C) bases, resulting in the formation of 5-methylcytosine (5-mC).261 This process is catalyzed by DNA methyltransferases (DNMTs), especially DNMT1, DNMT3A, and DNMT3B.262 DNA methylation induces gene silencing by interfering with the binding of transcription factors to highly methylated promoter regions, modifying chromatin structure, or reactivating the domains of methyl-binding proteins.261,263 During oxidative stress, ROS can affect DNA methylation through oxidative DNA damage formed by pyrimidine and 5-methylcytosine (5mC) hydroxylation.264 In addition, ROS can also affect DNA demethylation through DNA oxidation and hydroxymethylation mediated by ten-eleven translocation (TET).264 Meanwhile, DNA methylation can also affect the expression of antioxidant genes, such as SOD, GSH and CAT, affecting the antioxidant defence of cells.265,266 SLC40A1 is responsible for encoding the FPN, which is the only carrier capable of exporting intracellular Fe2+.267,268 Upregulated DNMT3B in degenerated NP cells inhibits SLC40A1 expression through methylation, thereby promoting ECM degradation, ferroptosis, and oxidative stress in NP cells.269 DNA methylation also participates in the regulation of various signaling pathways, including MAPK and NF-κB, which are involved in IVD cell survival, aging, inflammatory response, ECM metabolism, and oxidative stress.18,256 For example, DNMT3A activates the NF-κB pathway by inducing hypermethylation of the peroxisome proliferator-activated receptor γ (PPARγ) promoter, which promotes apoptosis and ECM degradation in NP cells.270 In addition, the hypermethylated genes MAPKAPK5 and PRKCZ, which are associated with the MAPK pathway, have been shown to regulate a wide range of catabolic molecules,42 but have not yet been correlated in IDD.
Histone modifications refer to covalent chemical modifications that occur on histone molecules.271 These modifications can affect the structure and function of chromatin, thereby regulating gene expression, mainly including acetylation, methylation, phosphorylation and ubiquitination.272 Oxidative stress is strongly associated with histone modification. Evidence suggests that oxidative stress has a direct impact on the function of chromatin remodeling proteins that act as histone modifying enzymes.273,274 Histone acetyltransferases (HATs) are responsible for adding acetyl groups to the lysine residues of histones, including various subtypes such as Gcn5, Tip60, p300/CBP.275–277 Among them, the activity of p300 can undergo significant changes after oxidation modification of key cysteine residues.278 Histone demethylases (HDMs) typically require Fe2+ as a cofactor, therefore the oxidation state of iron ions (Fe2+ or Fe3+) is a key factor affecting the activity of this enzyme.279,280 In this regard, O2− and OH- can oxidize Fe2+ to Fe3+ at the catalytic center of HDMs and suppress its activity.281,282 In addition, the imbalance of non-enzymatic antioxidant GSH oxidation and reduction levels can indirectly affect histone methylation.283 Histone modification also affects the redox state in IVD. ZRT/IRT-like Protein 4 (ZIP4) is a zinc transporter protein belonging to the ZIP family.284 The upregulation of ZIP4 enhanced inflammation and oxidative stress in NP cells, exacerbating ECM degradation.285 In terms of mechanism, ZIP4 upregulates the expression of histone deacetylase 4 (HDAC4), which in turn exerts its effects by regulating the expression levels of NF-κB, SIRT1, and FoxO3a.
ncRNA is a class of RNA molecules that do not encode proteins, mainly including microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs).286 ncRNAs exhibit differential expression in degenerated human IVD tissues and participate in almost all cellular functional processes, including ECM degradation, inflammatory response, cell apoptosis, cellular senescence, oxidative stress, and angiogenesis.287,288 In addition, N6-methyladenosine (m6A) is the most common RNA modification at the post-transcriptional level and is widely present in IDD.289,290 Chen et al found that m6A-methylated circGPATCH2L was recognized by the YTHDF2-RPL10-RNaseP/MRP complex and cleaved by endoribonuclease, which was beneficial to alleviate DNA damage accumulation and apoptosis in NP cells.291
Advantages of NDDSs for the Treatment of IDD
At present, the administration methods of drugs and bioactive agents for the treatment of IDD mainly include systemic administration and IVD in situ injection, but both have the disadvantages of poor curative effect and many side effects. Due to the non-vascular nature of IVDs, systemic administration makes it difficult to maintain adequate drug concentration and duration of action within the IVD, which results in limited bioavailability in local tissues.292 Systemic administration can also result in altered or impaired active units of the drug, especially as oral drugs are susceptible to degradation by digestive enzymes and intestinal flora as well as first-pass elimination by the liver. In addition, systemic administration may result in widespread distribution of the drug throughout the body, leading to potential off-target effects and toxic side effects on other organs with prolonged excessive exposure.293,294 For IVD in situ injection, the disadvantage is the poor injection dose and retention of drugs, which leads to limited absorption of drugs by target cells, thus reducing the therapeutic effect. However, increasing the number of injections may lead to intervertebral infection and accelerate the progression of IDD.295 Moreover, the administration of in situ injections may result in drug infiltration into the dural sac, thereby leading to central nervous system infections and other detrimental consequences.296
Nanomedicine refers to the application of nanoscience and technology to solve medical problems, which involves all aspects of diagnosis, prevention and treatment of disease.297 The reduction in size of nanomaterials gives them unique physical and chemical properties, which can be used to enhance existing medical technologies.298 NDDSs are at the forefront of nanomedicine applications and are currently used in the treatment of a wide range of diseases, including IDD.299 Compared with traditional drug delivery methods, NDDSs have the following advantages: (1) Through surface modification, NDDSs can carry specific ligands or antibodies that enable them to recognise and bind to diseased cells or tissues, enhancing drug accumulation at the site of the lesion.37 (2) NDDSs can be designed as controlled drug release systems, allowing drugs to be released at a constant rate or in response to specific stimuli, improving therapeutic efficacy and reducing toxicity.22 (3) With their nanoscale size, certain specific NDDSs can assist drugs in entering the interior of cells through micropores in the cell membrane, which is crucial for drugs that require intracellular action.37,300 (4) NDDSs generally have good biocompatibility and degradability, which can reduce damage to normal tissues and provide a superior cell microenvironment.292
 |
Table 1 Common Nano-Drug Delivery Systems in IDD Treatment |
Many NDDSs carrying therapeutic agents have been used to ameliorate the oxidative stress microenvironment of degenerating IVDs and to promote IVD repair.37 Table 1 summarises the advantages and limitations of some common NDDSs used to treat IDD. In addition, Table 2 lists several promising nanocarriers that have been used to treat IDD. At the same time, combined with research in other fields, we will explore new NDDSs that are more valuable and effective for the treatment of IDD in the future.
 |
Table 2 Recent Studies on Nano-Drug Delivery Systems for the Treatment of IDD. |
Major Types of NDDSs for IDD
Liposomes
Liposome is a kind of nanoscale double-layer spherical structure mainly composed of lipid materials such as phospholipids and cholesterol, which makes liposomes have the ability to transport drugs, genes or other active molecules.357 Compared with other NPs, liposomes have high biocompatibility, easily adjustable physicochemical properties, diverse functionalization modifications and mature preparation methods, making them the first NPs to be converted into clinical applications.358,359 However, to date, there have been few studies on the use of liposomes in the treatment of IDD. In several limited studies, liposomes have been described as carriers for small molecule drugs and ncRNA to improve cell apoptosis and ECM degradation in degenerative IVD.360,361 In fact, liposomes also have unique potential in responding to oxidative stress. Liposomes are composed of a hydrophilic core and one or more hydrophobic spaces surrounded by lipid bilayers, which makes liposomes a multifunctional drug delivery platform capable of carrying hydrophobic drugs and hydrophilic drugs at the same time.22 Currently, there is a wide variety of hydrophilic drugs (such as vitamin C, glutathione, water-soluble flavonoids)362–364and lipophilic drugs (such as vitamin E, retinol analogues, carotenoids, fat-soluble flavonoids and ubiquinone)365–367 that can be used to improve oxidative stress. In addition, liposomes can carry antioxidant enzymes to directly enhance the cellular antioxidant defense system. For example, Zhang et al developed a liposome encapsulating SOD, which released SOD to improve oxidative damage induced by sodium dextran sulfate and significantly alleviate symptoms of colitis in mice.368 These methods may be used in the future to improve the oxidative stress microenvironment in IDD.
Currently, many challenges remain for liposome-associated drug delivery in IDD, including enhancement of drug encapsulation, tissue targeting, prolonged circulation and reduction of biotoxicity, and these optimisations can help to improve the therapeutic efficacy of liposomal systems.369 Firstly, the ideal size of liposomes is usually between 50 and 200 nm, and excessively large particle sizes can diminish therapeutic efficacy by affecting pharmacokinetic and pharmacodynamic profiles.370 The encapsulation efficiency of drugs in lipid membranes can be influenced by the composition of liposomes, with cholesterol being an example of a component that may decrease this efficiency.371 Secondly, the encapsulation strategies for hydrophobic and hydrophilic drugs are also different, as they achieve high encapsulation efficiency by respectively binding with lipid bilayers and aqueous cores.372 In addition, the active loading strategy allows the introduction of a transmembrane pH gradient, which can drive drug molecules into empty vesicles.301 For enhanced tissue targeting, bioconjugation strategies can be employed to modify liposomes with small molecule ligands, nucleic acids, peptides, or monoclonal antibodies.373 In addition, smart stimulus-responsive liposomes can be constructed to target specific pathological environments, tissue sites and subcellular compartments by incorporating microenvironmental features (low pH, oxidative stress, and pro-catabolism) in degenerative IVDs.374,375 In particular, redox-responsive liposomes can rapidly release encapsulated antioxidants in a high ROS environment and effectively alleviate oxidative stress.376,377 In addition, liposomes are susceptible to clearance by the mononuclear phagocyte system,378 especially the large amount of immune cell infiltration in the late stage of IDD, which shortens their residence time in the IVD. One of the improvements is the use of biocompatible inert polymers to encapsulate liposomes (such as polyethylene glycol, PEG) to reduce their clearance by phagocytes.379 The degradation and biotoxicity of liposomes are also a concern, especially for some functional components used in the construction of functionalized liposomes.380 For example, cationic lipids can reduce cell mitosis and damage key cellular proteins.381,382 In summary, developing and improving liposomal carriers is a complex challenge that involves optimizing and balancing multiple parameters to achieve safe and effective drug delivery.
Polymeric Nanoparticles
Polymeric nanoparticles (PNPs) are typically nanoscale homogeneous spherical structures composed of biocompatible and biodegradable polymers, which can encapsulate hydrophilic and hydrophobic pharmaceutical drugs inside the particles or attach to the particle surface.383 According to the source, PNPs can be divided into natural polymers and synthetic polymers.384
Natural polymers typically come from plants or animals, including polysaccharides, polyphenols, fibroin proteins, gelatin (Gel), collagen protein and albumin.384,385 Their main advantages as drug delivery carriers are low biotoxicity, biodegradability, high biocompatibility, and unique bioactivity.386 Polysaccharides are common PNPs that naturally possess excellent antioxidant and anti-inflammatory properties.387,388 It was shown that carboxymethylated chitosan (CMCS) significantly ameliorated H2O2-induced mitochondrial dysfunction and ROS production in NP cells, which in turn inhibited ECM degradation and cell apoptosis.389 In addition, fucoidan can upregulate the expression of antioxidant enzymes in NP cells by activating the Nrf2 signaling pathway, thereby alleviating IL-1β-induced oxidative stress damage.390 Gelatin methacrylate (GelMA) microspheres functionalised with fucoidan can effectively preserve the ECM component of IVD and maintain hydration of NP tissues, thus preventing puncture-induced IDD in rats.390 Hyaluronic acid (HA), as an anionic glycosaminoglycan, is the main component of ECM.391 As a drug carrier, HA has excellent biocompatibility, biodegradability and mechanical properties.392 Meanwhile, existing evidence also suggests that HA can improve TBHP-induced mitochondrial dysfunction, oxidative stress, cell apoptosis, aging and ECM degradation by activating mitophagy.393 Importantly, HA can also alleviate IVD-induced pain by altering glycosylation and modulating key inflammatory signaling.393 Gallic Acid (GA) is a polyphenolic compound with inherent antioxidant properties.394 Recently, Chen et al developed a poly-GA-Mn (PGA-Mn) antioxidant NPs by combining GA with Mn2+ via a self-assembly method, and used a mitochondria-targeting peptide (TP04) as a surface modification (PGA-Mn-TP04).312 The rapid electron exchange between Mn2+ and GA enhances the ROS scavenging ability of PGA-Mn-TP04, effectively alleviating mitochondrial damage. In addition, PGA-Mn-TP04 also promotes mitochondrial fusion by targeting mitochondria to restore mitochondrial function, thereby alleviating NP cell apoptosis and ECM degradation. After in situ injection, PGA-Mn-TP04 can persist in the rat IVD for 13 days and effectively delay the progression of IDD (Figure 6). The natural polymer carrier has great potential in the treatment of IDD, but it lacks batch-to-batch consistency and the diverse functionality possessed by synthetic polymers.395
 |
Figure 6 Metallo phenolic nanoparticles targeting mitochondria for the treatment of IDD. (A) Schematic representation for the synthesis of the PGA-Mn-TP04. (B) OH- and (C) O2− scavenging efficiency of PGA-Mn with various concentrations. (D) H2O2 scavenging efficiency at different concentrations of PGA-Mn over time. (E) Cytosolic ROS (green fluorescence) and mitochondrial ROS (red fluorescence) levels in NP cells under different treatment conditions. (F) Transmission electron microscopy (TEM) images of mitochondria in NP cells under different treatment conditions. (G) Immunofluorescence staining of MMP-13 and COL2A1 in NP cells. (H) Imaging manifestations of PGA-Mn-TP04 in treating IDD in rat acupuncture model. (I) HE staining images of IVD at 4 and 8 weeks after surgery. (J) Safranin-O/Fast Green staining of different groups at 4 and 8 weeks. (*means p value < 0.05, **means p value < 0.01, ***means p value < 0.001, ****means p value < 0.0001). Reprinted with permission from Chen Q, Qian Q, Xu H, et al. Mitochondrial-Targeted Metal-Phenolic Nanoparticles to Attenuate Intervertebral Disc Degeneration: alleviating Oxidative Stress and Mitochondrial Dysfunction. ACS nano. 2024;18(12):8885–8905. Copyright © 2024, American Chemical Society.312 |
 |
Figure 7 PDA@CNO nanoparticles can mimic the activities of multiple antioxidant enzymes to alleviate oxidative stress in degenerated IVD. (A) Preparation of PDA@CNO. (B) ROS Scavenging Process of PDA@CNO. (C) PDA@CNO attenuates oxidative stress to protect NP cells. (D) Representative Micro-CT and MRI images of the caudal vertebrae of rats at 4 and 8 weeks. (E) Representative images of HE and Safranin O/fast green. Reprinted with permission from Wang J, Wu R, Liu Z, et al. Core-Shell Structured Nanozyme with PDA-Mediated Enhanced Antioxidant Efficiency to Treat Early Intervertebral Disc Degeneration. ACS Appl Mater Interfaces. 2024;16(4):5103–5119. Copyright © 2024, American Chemical Society.314 |
 |
Figure 8 MnO2 nanoparticles coated with TrkA overexpressing macrophage membranes alleviate oxidative stress and discogenic pain. (A) Preparation process of MnO2@TMNP nanoparticles. (B) Mechanism of MnO2@TMNP in the treatment of IDD. (C) Flowchart of the timing of in vivo experiments to evaluate the effects of MnO2@TMNP on disc regeneration. (D) Representative T2-weighted MRI images under different conditions. The red arrow marks the modeled section. (E) HE staining and Safranin O solid green staining of target segments of IVDs in different treatment groups. Reproduced from Yang W, Li K, Pan Q, et al. An Engineered Bionic Nanoparticle Sponge as a Cytokine Trap and Reactive Oxygen Species Scavenger to Relieve Disc Degeneration and Discogenic Pain. ACS nano. 2024;18(4):3053–3072. Copyright © 2024, The Authors. Published by American Chemical Society and licensed under CC-BY-NC-ND 4.0. https://creativecommons.org/licenses/by/4.0/.315 |
 |
Figure 9 Mitochondria-targeted carbon dots treat IDD by exerting antioxidant enzyme-like activity. (A) Preparation process of CD-PB-TPP nanoparticles. (B)Mechanism of CD-PB-TPP in the treatment of IDD. (C and D) X-ray and MRI images of rat tail in different treatment groups. (E) DHI of rat caudal IVD in different groups. (F) Mean gray value of the target intervertebral disc. (G) HE and Safranin O/fast green of the rat caudal IVD. (*means p value < 0.05, **means p value < 0.01, ***means p value < 0.001, ****means p value < 0.0001). Reproduced with permission from Shi Y, Bu W, Chu D, et al. Rescuing Nucleus Pulposus Cells from ROS Toxic Microenvironment via Mitochondria-Targeted Carbon Dot-Supported Prussian Blue to Alleviate Intervertebral Disc Degeneration. Adv Healthcare Mater. 2024;13(8):e2303206. Copyright © 2024, Wiley‐VCH.323 |
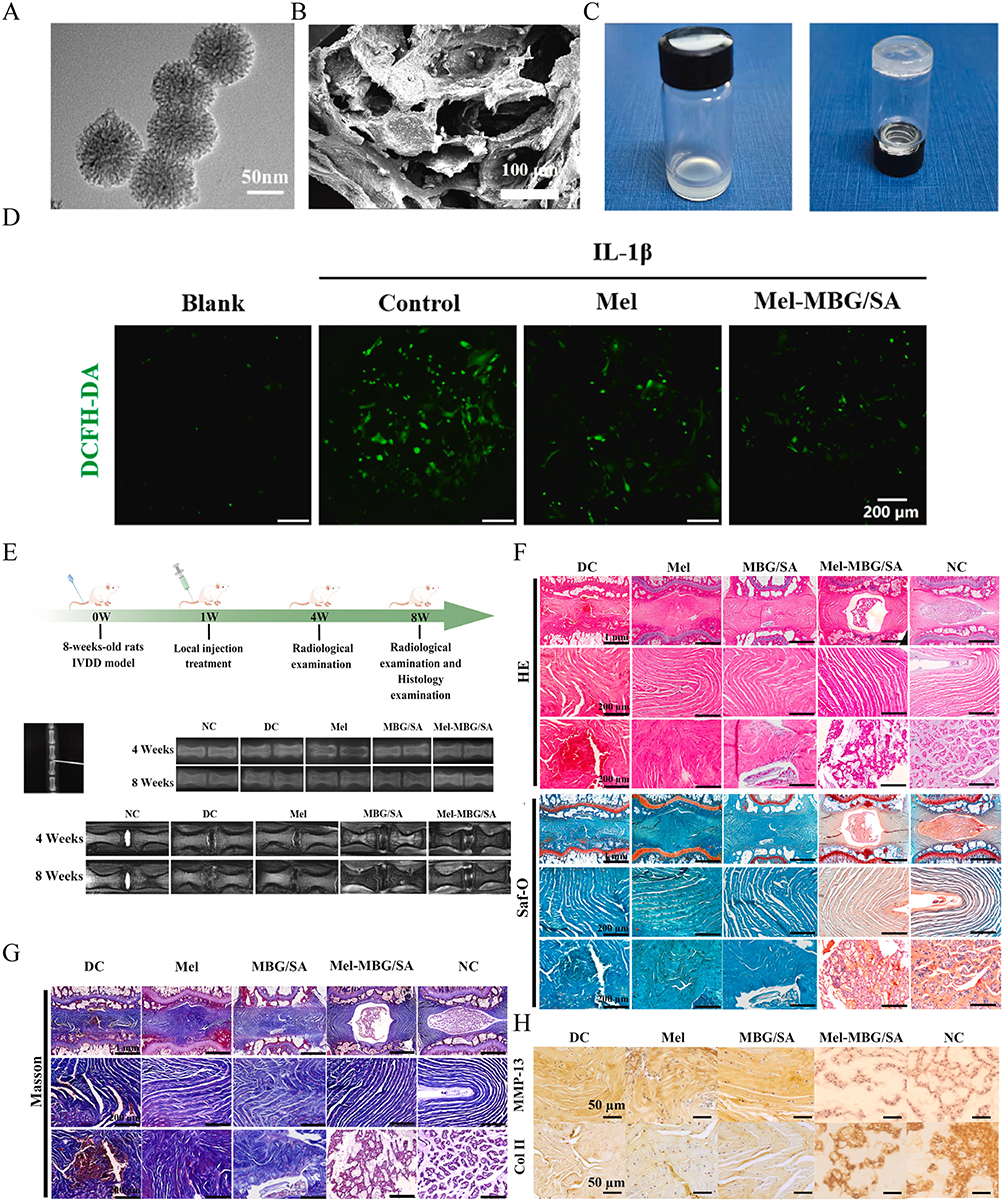 |
Figure 10 Injectable mesoporous bioactive glass/sodium alginate hydrogel loaded with melatonin for intervertebral disc regeneration. (A) TEM image of MBG nanoparticles. (B) Scanning electron microscopy (SEM) image of Mel-MBG/SA hydrogel. (C) Photos of the MBG/SA hydrogels before and after crosslinking. (D) Intracellular ROS levels in NP cells were monitored after different treatments using a ROS probe (DCFH-DA). (E) Schematic illustration of the animal experiment and X-ray and MRI images of the caudal vertebrae of rats after different treatments. (F) HE and Safranin O/fast green of the rat caudal IVD. (G) Masson staining of the rat caudal IVD. (H) Immunohistochemical staining images of COL-II and MMP-13. Reproduced with permission from Wu R, Huang L, Xia Q, et al. Injectable mesoporous bioactive glass/sodium alginate hydrogel loaded with melatonin for intervertebral disc regeneration. Mater Today Bio. 2023;22:100731. Under CC-BY-NC-ND 4.0. https://creativecommons.org/licenses/by/4.0/.327 |
 |
Figure 11 Injectable decellularized NP hydrogel can be used as a carrier for MSCs to repair degenerated IVD. (A) Schematic overview of the construction of LM. (B) TEM images of poly(allylamine hydrochloride)-coated MnO2 nanoparticles and LM. (C) Schematic overview of the construction of LMGDNPs.(D) Size distribution and microstructure of microspheres. (E) Cumulative release profile of LOX in LMGDNPs based on the BCA assay. (F) Lactate consumption efficiency of LOX-MnO2 nanozyme and microspheres. (**means p value < 0.01, ***means p value < 0.001, ****means p value < 0.0001, between LM and CTRL groups; ns, no significance, # means p value < 0.05, between LM and LMGDNP groups). (G) Dissolved oxygen profiles of H2O2 solution mixed with LOX-MnO2 nanozyme and microspheres. (**means p value < 0.01, ***means p value < 0.001, between LM and LMGDNP groups). (H) Fluorescence images showing changes in fluorescence intensity after delivery of DiR Iodide-labeled BMSCs via different vectors on days 1, 7, 14 and 21.(**means p value < 0.01, ***means p value < 0.001, GDNPs compared to cell pellets; #means p value < 0.05, ##means p value < 0.01, LMGDNPs compared to cell pellets). (I) Representative images of T2-weighted MRI of rat tails. (J) Histological images based on HE and SO staining. Reproduced with permission from Peng Y, Chen X, Zhang Q, et al. Enzymatically Bioactive Nucleus Pulposus Matrix Hydrogel Microspheres for Exogenous Stem Cells Therapy and Endogenous Repair Strategy to Achieve Disc Regeneration. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2024;11(10):e2304761. Copyright © 2023, Wiley‐VCH GmbH. Under CC-BY-NC-ND 4.0. https://creativecommons.org/licenses/by/4.0/.332 |
Synthetic polymers are synthesized by chemical methods, mainly including polyesters, polyamino acids, phosphate-based polymers, vinyl polymers and PEG, etc.302 As drug carriers, they have the advantages of high stability, simple synthesis, strong flexibility and diverse modified structures.396 Polyesters are polymers consisting of repeating ester groups, commonly including poly(lactic acid) (PLA), poly(glycolic acid) (PGA), poly(lactic-co-glycolic acid) (PLGA) and poly(ε-caprolactone) (PCL).302 PLGA is the most widely studied polyester in IDD.397 Bradykinin (BK) is a nonapeptide hormone that exerts pleiotropic effects by promoting cellular proliferation, attenuating oxidative stress, and inhibiting apoptotic processes.398,399 Based on this, Qiu et al developed BK/PLGA sustained-release microspheres.400 The results indicate that BK/PLGA sustained-release microspheres can inhibit TBHP-induced oxidative stress and mitochondrial dysfunction through the BK receptor 2 and its downstream PI3K/AKT pathway. Unfortunately, the average particle size of such microspheres is about 100 μm, which does not reach the nanoscale. Lim et al improved the process by synthesizing nanoscale PLGA particles and loading the anti-aging drug ABT263 into them (ABT/PLGA).313 After in situ injection of ABT/PLGA into acupuncture-induced IDD of rats for 6 weeks, cell senescence, inflammatory response and ECM degradation in IVD were significantly reduced. In addition, polydopamine (PDA) is a synthetic polymer that possesses the ability to scavenge free radicals and chelate metal ions.401 Based on this, Wang constructed a core-shell nanozyme (PDA@CNO) with co-doped NiO NPs (CNO) as the core and PDA as the shell314 (Figure 7). PDA@CNO can mimic the activities of multiple antioxidant enzymes to effectively scavenge O2−, H2O2, and OH−, which can protect NP cells from proliferation arrest, metabolic abnormalities, and inflammation, thereby reestablishing ECM homeostasis. Furthermore, it was shown that PDA@CNO NPs were able to attenuate iron overload through chelation and up-regulation of iron storage protein expression, which in turn attenuated lipid peroxidation-induced ferroptosis in NP cells.205 These results suggest that PDA may serve as an effective drug carrier for the treatment of IDD. Currently, synthetic polymers have broad prospects in drug delivery, but attention should be paid to the degradability and potential biological toxicity of synthetic polymers.402
Inorganic Nanoparticles
Inorganic NPs come from inorganic materials, mainly including metal NPs and non-metal NPs.403 Inorganic NPs have the advantages of adjustable particle size, enhanced stability, and large specific surface area, making them widely used in various industries such as healthcare, cosmetics, composite materials, and energy.404 In terms of drug delivery, drug molecules can be combined with NPs through electrostatic interactions, hydrophobic interactions, and covalent bonds to achieve drug delivery.22
Metal NPs usually include metal-based NPs and their oxides, including silver (Ag), gold (Au), copper (Cu), iron (Fe), zinc (Zn), Mn, cadmium (Cd) and cobalt (Co).404 The main purpose of synthesizing metal oxides is to improve their reactivity and efficiency.405 MnO2 is the oxidised form of Mn and its nanoscale form has unique chemical and physical properties of high specific surface area, low toxicity, high adsorption and biocompatibility.406 MnO2 can also change the inflammation and oxidative stress microenvironment of IVD cells at the genetic level to achieve IVD repair.335,407 During the repair process, its metabolite Mn2+ is discharged with body fluids, thus reducing the impact on the internal environment.408 Therefore, MnO2 NPs are an excellent drug carrier for the treatment of IDD. In the oxidative microenvironment of degenerating IVDs, excess ROS induces macrophage M1 polarisation, which in turn induces NGF production.409 The tropomyosin receptor kinase A (TrkA) belongs to the receptor tyrosine kinase family and is the specific receptor for NGF.410 Based on the above, Yang et al developed a nanomaterial encapsulating MnO2 nanoparticles with TrkA overexpressing macrophage cell membrane (TMNP) (MnO2@TMNP)315 (Figure 8). MnO2@TMNP can act as a “molecular sponge” to capture NGF, thereby alleviating IVD-derived pain caused by the infiltration of IVD nerve fibers. At the same time, MnO2@TMNP can also target macrophages to deliver MnO2 nanoparticles, which in turn effectively scavenge intracellular ROS and prevent M1 polarisation to improve IDD. In addition, NPs based on Prussian blue (PB) (C18Fe7N18) can mimic the activities of multiple antioxidant enzymes (CAT and SOD) to effectively remove ROS.411 Zhou et al injected PB NPs into the IVD of a rat IDD model.316 The results showed that PB NPs could stabilize SOD1 from ubiquitination-proteasome degradation, thereby improving mitochondrial structure to enhance antioxidant capacity and thus alleviate ROS-induced IDD. Recently, a new type of multifunctional NPs called metallo-phenols have been developed, which can effectively improve the oxidative stress microenvironment of degenerated IVDs.317 Such NPs are usually synthesized by self-assembly of metal ions (Cu, Mn, Fe) and phenolic compounds (catechol, gallic acid) in aqueous solution.317,412 The ortho-hydroxyl groups of phenolic compounds can form stable coordination bonds with metal ions, which promotes the aggregation of metal ions and phenolic molecules in solution and forms NPs.413 Metal-phenol NPs can effectively remove ROS in IVD cells by simulating the activities of various antioxidant enzymes through the active sites provided by the metal center.312,317,318 At the same time, its surface modification molecules can usually accurately target mitochondria to improve mitochondrial function, thereby inhibiting IVD cell death and ECM degradation and effectively delaying the progression of IDD.312,318
At present, the common inorganic non-metallic NPs in IDD treatment are mainly carbon-based NPs, including fullerenes, carbon nanotubes, graphene and carbon quantum dots.414 Fullerene is a hollow molecule composed of 60 carbon atoms and is currently the most widely used inorganic NPs.415 Due to its antioxidant capacity hundreds of times higher than other conventional antioxidants, it is called a “free radical sponge”.416 Xiao et al combined functionalized nano fullerene with cFlFlF peptide, which can specifically bind to formyl peptide receptor-1 expressed on activated macrophages, thereby effectively reducing oxidative stress, inflammatory response and pain in a mouse model of radiculopathy.417 Compared with the low water solubility and poor biocompatibility of fullerene, its polyhydroxy derivative fullerol has better biocompatibility and is the most widely studied.418 In 2014, Jin et al reported that fullerol could significantly improve H2O2-induced ROS production in NP cells, thereby alleviating acupuncture-induced IDD in rabbits.319 Subsequently, they further verified the antioxidant activity of fullerol in mouse bone marrow stromal cells, and the results showed that fullerol could reduce 23% of basal and 18% of IL-1β-induced intracellular ROS.419 These studies demonstrate the powerful antioxidant properties of fullerenes and their derivatives, providing a new option for targeted IVD delivery. In addition, carbon nanotubes (CNTs) are considered as new drug and gene delivery carriers due to their high surface-to-volume ratio, high strength, biocompatibility and easy functionalization.420,421 In 2022, Huang et al developed a cationic polymer-modified CNT for targeted delivery of LINC02569 (a ncRNA synthesized at an active enhancer) siRNA to NP cells.320 Compared with traditional siRNA delivery systems, CNTs can accumulate in NP cells and significantly reduce the inflammatory response and aging of NP cells by inhibiting p65 phosphorylation. The limitation of this study lies in the absence of in vivo testing for the role of CNT. Recently, carbon dots (CDs) have been widely used in nanomedicine due to their nanoscale size, high water solubility, and strong biocompatibility.422,423 Related studies have shown that CD has inherent enzyme-like activity and free radical scavenging ability, which can be further enhanced by surface modification and doping elements.424 For example, Wu et al and Sun et al designed NAC-modified CD (NAC-CD)321 and carbonized Mn-containing nanodots (MCD),322 respectively. The results showed that both NAC-CD and MCD had SOD- and CAT-like activities and strong total antioxidant capacity, which could effectively alleviate H2O2-induced mitochondrial dysfunction, cell death and ECM degradation in NP cells, and were verified in the rat IDD model. In addition, surface-modified CDs can easily cross the cell membrane and enter the organelles to exert targeted antioxidant effects.425 For example, Shi et al reported a triphenylphosphine (TPP, a mitochondrial affinity ligand) modified PB-loaded CD (CD-PB-TPP)323 (Figure 9). The results showed that CD-PB-TPP could effectively evade lysosomal phagocytosis to achieve effective mitochondrial targeting, thereby significantly reducing ROS in mitochondria and protecting NP cells from senescence, catabolism and inflammatory responses.
Polymer Micelles
Polymer micelle (PM) is a nanostructure formed by self-assembly of amphiphilic polymers in solution.426 The material typically comprises a hydrophobic core and a hydrophilic shell layer, with the former located at the center of the PM and the latter forming an outer layer.427 The shell determines the hydrophilicity, charge, and surface modification of the PM, while the core is used for drug encapsulation.428 PM can be classified as graft or block copolymers, the latter including diblock, triblock and multiblock copolymers.429 Currently, PM drug delivery systems aim to achieve targeted, controlled, and sustained release of hydrophobic drugs.430
In 2021, Yu et al designed a new amphiphilic copolymer PEG-PAPO, which can self-assemble into nanomicelles and load kartogenin (KGN) and apocynin (APO).324 APO is linked to the polymer chain via a phenolic ester bond, while KGN is physically embedded in the hydrophobic core of the micelle. The results showed that the micelles could achieve precise and controllable delivery of APO and KGN, thereby protecting human adipose-derived stem cells (ADSCs) from oxidative stress and promoting NP-like differentiation of ADSCs. This study provides a potential adjuvant for stem cell transplantation therapy for IDD. In another study, Chang et al synthesized a polyamine-based PM loaded with Runx1 mRNA (a cartilage anabolic factor).431 After the PM was injected into the degenerative IVD of rats, the intervertebral height and ECM content were significantly improved. Recently, researchers have used an emulsion-restricted assembly method to encapsulate coenzyme Q10 (CoQ10) (a mitochondria-targeted hydrophobic antioxidant) into lecithin micelles to achieve a water-soluble transformation of CoQ10.325 The results showed that CoQ10-loaded micelles had better bioactivity and could enhance the antioxidant capacity of rat bone marrow mesenchymal stem cells (BMSCs) by down-regulating NF-κB signaling. Furthermore, co-injection of the micelles with BMSCs maintained IVD height and alleviated IDD in a rat acupuncture model.
Nanofiber
Nanofibres are typically fabricated through phase separation, self-assembly, and electrostatic spinning using biodegradable natural polymers, synthetic polymers, and polymer blends.307 Currently, nanofibers used for drug delivery feature high specific surface area, high porosity, high biocompatibility, and easy modification, thus enabling high drug loading, sustained release properties, improved therapeutic index, and significantly reduced side effects.432–434 In addition, nanofibers have certain mechanical strength and structural characteristics similar to ECM,435,436 which help to improve the microenvironment within IVD, promote tissue repair and the interaction between cells and drugs. Therefore, nanofibers are a promising NDDSs for the treatment of IDD.
Initially, nanofibers were mainly used as tissue engineering scaffolds to repair AF.437 In 2011, researchers reported a PCL scaffold produced by electrospinning technology that can mimic the layered structure and mechanical function of AF.438 The scaffold can promote the adhesion, proliferation and ECM production of primary porcine AF cells. Subsequently, Martin et al combined PCL scaffolds with cell-seeded hydrogels to form disc-like angle-ply structures (DAPS).439 They found that only a few cells and ECM were deposited after implanting DAPS into rat IVD, which was significantly improved upon the addition of poly(ethyleneoxide). Although these nanofiber scaffolds demonstrate effective IVD repair, the minimally invasive implantation of the scaffolds remains a problem to be addressed. In 2019, Uysal et al reported an injectable collagen peptide-presenting nanofiber scaffold that can be easily implanted in IVD centers without damaging other healthy tissues.440 Injection of this scaffold into degenerated rabbit IVDs significantly induced glycosaminoglycan and collagen deposition. In pursuit of better biocompatibility and biodegradability, Liu et al combined Poly (ether carbonate urethane) urea (PECUU) electrospun nanofibre scaffolds with decellularized AF matrix (DAFM).441 Compared with PECUU nanofiber scaffolds, DAFM/PECUU electrospun scaffolds can significantly promote ECM production from AF-derived stem cells. In addition, nanofibers can also be surface modified to further endow the scaffold with biological functional properties.432,442 For example, Yu et al constructed a fucoidan-loaded PECUU nanofiber scaffold (F-PECUU).326 Compared with PECUU scaffolds, F-PECUU can significantly reduce LPS-induced inflammatory response and oxidative stress in AF cells, while promoting AF cell proliferation and ECM synthesis. In the rat AF defect model, the IVD height and NP hydration after F-PECUU treatment reached 75% and 85% of the sham-operated group, respectively.
Nanohydrogel
Nanohydrogel is a three-dimensional network structure formed by cross-linking hydrogel and nanomaterials through chemical or physical methods.309,443 Nanohydrogels not only possess the excellent biocompatibility, biodegradability, mechanical strength and adhesion properties of hydrogels, but also the unique properties of each of the different NPs.309,444 By adjusting the type of nanohydrogel skeleton material, cross-linking degree, type of nanoparticles and functionalization, minimally invasive implantation and in situ curing of hydrogels can be achieved,445–447 thereby establishing a protective microenvironment that is conducive to cell proliferation and ECM synthesis. Currently, nanohydrogels have been widely studied in IDD. They can be used as sustained-release systems for bioactive substances, templates for ECM deposition, and scaffolds for carrying cells.448 According to the source of nanohydrogel skeleton materials, we divide them into natural polymer hydrogels, synthetic polymer hydrogels and composite hydrogels.
The skeleton materials of natural hydrogels are usually composed of natural polymers from animals and plants, including Gel, chondroitin, alginate, HA and chitosan (CS).449 Due to their diverse range of material sources, excellent biocompatibility, and degradability,450 natural hydrogels have found extensive applications in IDD repair and drug delivery with remarkable efficacy. Alginate is a natural polymer derived from brown algae, and hydrogels based on it have strong biocompatibility, high porosity and water retention capacity.451 In 2023, Wu et al reported a sodium alginate (SA)-based injectable hydrogel for delivering MT-loaded mesoporous bioactive glass (MBG) NPs (MT-MBG/SA), which can continuously release MT to alleviate IL-1β-induced oxidative stress327 (Figure 10). Compared with the sham operation group, MT-MBG/SA treatment could significantly improve IVD height and NP water content. To enhance the mechanical capabilities of SA, the researchers added silk fibroin nanofibers to SA hydrogels to provide support for adjacent tissues and promote cell proliferation.328 In addition to SA, Gel is a widely used natural material, which is obtained by partial hydrolysis of collagen.452 In order to enhance the mechanical and biological properties of Gel, it is often combined with other polymers. For example, Gan et al combined dextran with Gel to construct a hydrogel with good biocompatibility and low cytotoxicity for the delivery of PLGA nanoparticles loaded with TGF-β3.329 The system can stably release TGF-β3 and successfully induce the NP-like differentiation of MSCs. In another study, researchers combined SA with Gel to construct a hydrogel that exhibited good cell adhesion, biocompatibility, and mechanical function.330 In addition, decellularized ECM (dECM) hydrogel is a new type of biomaterial that retains the natural structure of ECM by removing the cellular components and has injectability, biocompatibility and good mechanical function.453 In 2023, Peng et al reported a decellularized AF matrix hydrogel for the delivery of TrkA-loaded amphiphilic polycarbonate NPs.331 The system can significantly inhibit the expression of neurogenic mediators and nerve axon growth, thereby reducing mechanical hyperalgesia in the rat IVD protrusion model. Related studies have also shown that dECM contains a variety of growth factors and cell signaling molecules that help promote cell proliferation, differentiation and tissue regeneration.454 In 2024, in another study by Peng et al, they developed an injectable hydrogel microsphere by incorporating lactate oxidase (LOX)-MnO2 NPs (LM) into glucose-rich decellularized NP hydrogel (GDNP) through a microfluidic system (LM-GDNP).332 The results showed that LM-GDNP was able to ameliorate the microenvironment of nutrient deprivation, hypoxia, low pH and oxidative stress in degenerating IVDs and activate autophagy through transforming growth factor β2 overlapping transcript 1 (TGFB2-OT1) and its downstream pathway to promote cell survival and matrix regeneration. After co-injection of LM-GDNP with BMSCs into degenerated IVD in rats, the activity of BMSCs can persist and significantly improve the water content and tissue structure of degenerated IVD (Figure 11). Therefore, LM-GDNP is expected to serve as an ideal delivery system for exogenous stem cell therapy and endogenous repair. In addition, fibrin hydrogel333 and DNA hydrogel334 have also been used for IDD treatment and have shown good stability, controlled release effect and biocompatibility.
Synthetic hydrogels are obtained by processing natural products or other industrial raw materials and are highly customizable biomaterials.446 Compared with natural hydrogels, synthetic hydrogels have lower immunogenicity, better mechanical properties, adjustable biodegradability and easy modification.309 GelMA is a photosensitive injectable biohydrogel material formed by polymerization of Gel and methacrylate (MA) in phosphate buffer at 50°C.455 Compared with Gel, GelMA has significantly improved degradation rate, immunogenicity and mechanical properties.447,456 In addition, since GelMA hydrogel is very similar to natural ECM, it can provide excellent mechanical properties and bioactivity, thus providing a suitable microenvironment and corresponding biological matrix for the repair of degenerated IVD.457 Currently, GelMA is the most commonly used hydrogel material for repairing IDD.456 In a study by Xu et al, they constructed a MnO2/GelMA hydrogel with good biocompatibility and porous structure.335 The results showed that MnO2/GelMA hydrogel could improve IVD microenvironment by removing intracellular ROS through SIRT1/Nrf2 pathway. However, this study did not verify the effects of the hydrogel in vivo. In another study, Li et al combined CS NPs encapsulating black phosphorus quantum dots (BPQDs) with GelMA through microfluidic technology to construct engineered hydrogel microspheres (GM@CS-BP), which can continuously release highly reducing BPQDs to improve the oxidative and inflammatory microenvironment in IVD.336 In the rat IDD model, GM@CS-BP microspheres were able to reshape the ECM structure and function of degenerated IVD by providing mechanical support and antioxidant effects. In addition, some researchers combined TGF-β1337 and aspirin338 -loaded NPs with GelMA, which significantly improved the NP structure and biomechanical function by reducing the inflammatory response.
Composite hydrogels are hydrogel materials composed of two or more different types of polymers.458 This material combines the advantages of multiple polymers, enabling it to achieve performance that is difficult for a single polymer hydrogel to attain.459 In 2015, Nair et al reported a composite hydrogel composed of CS and poly(hydroxybutyrate-co-valerate) (CP) for loading chondroitin sulfate NPs.339 The hydrogel has mechanical properties similar to those of natural tissues and can withstand different stresses under dynamic conditions corresponding to daily activities such as lying down, sitting and standing. The chondroitin sulfate NPs carried by them can enhance the activity of MSCs and promote NP-like differentiation. However, this study did not verify the long-term effects of the hydrogel in vivo. In addition, researchers have constructed a PBNPs-loaded hydrogel (PBNPs@OBG) composed of oxidized hyaluronic acid (OHA), borax and Gel, which exhibits antibacterial activity, antioxidant capacity, biocompatibility, tissue adhesion, self-healing and suitable mechanical properties340 (Figure 12). Compared with the sham operation group, PBNPs@OBG could significantly improve the water content, intervertebral height and tissue integrity of degenerated IVD in rats. In another study, Nezadi et al used laponite (LAP) as a cross-linker to construct a hydrogel composed of quaternized CS (QCS) and Gel.341 The results show that the hydrogel-wrapped MgAl layered double hydroxide (LDH) NPs can improve the IVD microenvironment, thereby significantly promoting NP cell proliferation and ECM synthesis. In addition, DNA damage-inducible transcript 4 (DDIT4) is an evolutionarily conserved protein that is overexpressed when cells are subjected to oxidative stress and is associated with a variety of diseases.438,461 Ma et al constructed a hydrogel loaded with DDIT4 siRNA (siDDIT4) using fifth-generation polyamidoamine (PAMAM) and HA (siDDIT4@ P-HA).193 The results showed that siDDIT4@P-HA could alleviate H2O2-induced mitochondrial dysfunction and pyroptosis in NP cells by downregulating the ROS-TXNIP-NLRP3 signaling axis. In vivo, siDDIT4@P-HA continuously releases siDDIT4 for up to 35 days, which can significantly alleviate IDD in rats.
 |
Figure 12 Dual-dynamic-bond cross-linked injectable hydrogel of multifunction for intervertebral disc degeneration therapy. (A) Fabrication scheme of the PBNPs@OBG hydrogel. (B) Mitigation of oxidative stress microenvironment within NP cells and notable amelioration of IDD caused by PBNPs-dotted hydrogel.(C and D) X-ray and MRI images of rat tail in different treatment groups. (E) DHI of rat caudal IVD in different groups. (F) Pfirrmann grade of the target intervertebral disc. (G–I) Representative images of HE, Safranin-O/Fast and Alcian Blue staining. (*means p value < 0.05, **means p value < 0.01, ***means p value < 0.001, ****means p value < 0.0001). Reproduced with permission from Yang L, Yu C, Fan X, et al. Dual-dynamic-bond cross-linked injectable hydrogel of multifunction for intervertebral disc degeneration therapy. J Nanobiotechnol. 2022;20(1):433. Copyright © 2022, Springer Nature. Under CC-BY-NC-ND 4.0. https://creativecommons.org/licenses/by/4.0/.340 |
 |
Figure 13 Exosome-loaded temperature-sensitive decellularised ECM hydrogels for the treatment of IDD. (A) Images of NP tissue decellularization and preparation of hydrogels. (B) TEM analysis of exosomes. (C) SEM analysis of NP, dECM and dECM@exo. (D) Control-release curve of dECM@exo.(E) Fluorescence images of living (green) and dead (red) cells in each group. (F) X-ray images of the intervertebral disc at 0, 4, 8, and 16 weeks. (G) Differences in DHI between different treatment groups at each time period. (H) Images of HE and Safranin O/fast green staining from each group. (* means p value < 0.05). Reproduced with permission from Xing H, Zhang Z, Mao Q, et al. Injectable exosome-functionalized extracellular matrix hydrogel for metabolism balance and pyroptosis regulation in intervertebral disc degeneration. J Nanobiotechnol. 2021;19(1):264. Copyright © 2021, Springer Nature. Under CC-BY-NC-ND 4.0. https://creativecommons.org/licenses/by/4.0/.73 |
 |
Figure 14 NO-loaded ROS-responsive system for amelioration of oxidative stress microenvironment in degenerating IVDs. (A) NO-loaded ROS-responsive system for amelioration of oxidative stress microenvironment in degenerating IVDs. (B) Cumulative release of NO. (C)Cumulative release of l-Histidine. (D) Cumulative release of zinc ions. (E)X-ray images of rat tail disc in animal experiment. (F) quantitative analysis of DHI. (G) MRI images of rat tail disc in animal experiment. (H and I) The results of quantitative water content in the NP tissue and Modified Pfirrmann grading. (J and K) Representative images of HE, Safranin O/fast green staining, and immunohistochemical staining for Col-I and Col-II. (*means p value < 0.05, **means p value < 0.01, ***means p value < 0.001, ****means p value < 0.0001). Reproduced with permission from Feng Y, Su L, Chen Z, et al. Accurate Spatio-Temporal Delivery of Nitric Oxide Facilitates the Programmable Repair of Avascular Dense Connective Tissues Injury. Adv Healthc Mater. 2024;13(14):e2303740. Copyright © 2024, Wiley‐VCH GmbH.345 |
 |
Figure 15 pH-responsive NDDS targeting senescent NP cells for the treatment of IDD. (A) The preparation procedures of nanoparticles. (B) Mechanism of SP@NNPm targeting of senescent NP cells. (C) SP@NNPm effectively targets senescent NP cells and activates the PGC1α-NRF2/TFAM pathway to reduce oxidative stress and cellular senescence. (D) Representative X-ray images and T2-MRI images of discs from sham or IDD rats. (E) Statistical analysis of the DHI. (F and G) Statistical analysis of the Pfirmann grades and relative signal intensity of NP tissues. (H) Representative images of HE and Safranin O/fast green staining and statistical analysis. (*means p value < 0.05, **means p value < 0.01, ***means p value < 0.001, ****means p value < 0.0001). Reproduced with permission from Liu S, Li K, He Y, et al. PGC1α-Inducing Senomorphic Nanotherapeutics Functionalized with NKG2D-Overexpressing Cell Membranes for Intervertebral Disc Degeneration. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2024;11(22):e2400749. Copyright © 2024, Wiley‐VCH GmbH. Under CC-BY-NC-ND 4.0. https://creativecommons.org/licenses/by/4.0/.347 |
 |
Figure 16 Hydrogen Ion Capturing Hydrogel Microspheres for the treatment of IDD. (A) Construction of MNP composed of CAT and TGF-β via biomimetic mineralization. (B) Preparation of GMNP loaded with MNP via the microfluidic device. (C) The mechanism by which GMNPs improve the IDD microenvironment, inhibit inflammatory response and promote ECM synthesis. (D) Experimental design of rat IDD model. (E) X-ray and MRI images of rat IDD at different time points. (F) Statistical analysis of the DHI. (G) Quantitative analysis of the gray value of IVDs. (H) Schematic of the material composition in different groups. (I) Images of HE and Safranin O/fast green staining from each group. (*means p value < 0.05, Statistically different from the puncture group; #means p value < 0.05, Statistically different from the control group). Reproduced with permission from Zheng D, Chen W, Chen T, et al. Hydrogen Ion Capturing Hydrogel Microspheres for Reversing Inflammaging. Advanced Materials (Deerfield Beach, Fla). 2024;36(5):e2306105. Copyright © 2023, Wiley‐VCH GmbH.348 |
 |
Figure 17 Sustained and bioresponsive two-stage delivery of therapeutic miRNA by injectable hydrogel loaded with polymeric micelles to inhibit IVD fibrosis. (A) Schematic illustration for formation of miRNA/PGPC polyplex micelles. (B) Encapsulation of miRNA/PGPC polyplexes in PEG hydrogels in an injectable manner and molecular mechanism of MMP-2 silence in nucleus pulposus cells for fibrosis inhibition. (C) Injection sites in the IVDs of rabbits. Reproduced with permission from Feng G, Zha Z, Huang Y, et al.. Sustained and Bioresponsive Two-Stage Delivery of Therapeutic miRNA via Polyplex Micelle-Loaded Injectable Hydrogels for Inhibition of Intervertebral Disc Fibrosis. Adv Healthcare Mater. 2018;7(21):e1800623. Copyright © 2018, WILEY‐VCH Verlag GmbH & Co. KGaA, Weinheim.349 |
 |
Figure 18 Multi-responsive spermidine delivery system for improving oxidative stress microenvironment in degenerated IVD. (A) High magnification TEM images of PDA NPs. (B) TEM image of PDA NPs’ responsiveness in simulated media for 12 h. (C) In vitro release of SPD from PDA NPs. (D) Representative fluorescent images of NPCs stained with DCFH-DA (ROS probe). (E) RT-qPCR analyses of the relative expression of the GPX1, SOD1, and CAT genes. (F) Representative images of the general appearance of IVDs in different treatment groups. (G) MRI images of rat IDD at different time points. (H) Representative images of HE staining at different time points. (I) Representative images of Safranin O/fast green staining at different time points. (*means p value < 0.05, #means p value < 0.01). Reprinted from Int J Biol Macromol, 274, Wang D, Lu K, Zou G, Wu D, Cheng Y, Sun Y. Attenuating ntervertebral disc degeneration through spermidine-delivery nanoplatform based on polydopamine for persistent regulation of oxidative stress. 132881. Copyright 2024, with permission from Elsevier.353 |
 |
Figure 19 MnOx-functionalized temperature-sensitive nanohydrogels contribute to the removal of endogenous ROS and provide a promising stem cell delivery platform for IDD. (A) States of the CS hydrogel and CS@MnOx nanohydrogel at 4 °C and 37 °C. (B) Injectability of the hydrogel. (C) SEM images of the CS hydrogel and the CS@MnOx nanohydrogel. (D) Evaluation of H2O2 elimination by the CS hydrogel and CS@MnOx nanohydrogel in vitro. (E) Gross images, X-ray images, and MRI images of IDD repair after treatment. (F) Quantitative analysis of DHI on X-ray images. (G) Quantitative analysis of the relative MRI index in MRI images. (H) Evaluation of Pfirrmann grading on MR images. (I) HE, Safranin-O/Fast Green, and Alcian blue staining of IVDs from different groups. (*means p value < 0.05, **means p value < 0.01, ***means p value < 0.001, ****means p value < 0.0001). Reproduced with permission from Wang X, Yu L, Duan J, et al. Anti-Stress and Anti-ROS Effects of MnOx-Functionalized Thermosensitive Nanohydrogel Protect BMSCs for Intervertebral Disc Degeneration Repair. Adv Healthcare Mater. 2024;13(29):e2400343. Copyright © 2024, Wiley‐VCH GmbH.354 |
Exosomes
Exosomes (Exos) are tiny membrane vesicles produced by intracellular multivesicular bodies (MVBs), with a diameter of approximately 30–150 nanometers.462 Exos typically contain complex biomolecules, including proteins, lipids, DNA and different types of RNA, which can act as messaging molecules playing a key role in intercellular communication.463 In recent years, Exos have been used as carriers of genes and drugs to treat a variety of diseases due to their biocompatibility, low immunogenicity, intrinsic targeting ability, and enhanced intracellular efficiency.464,465 A large number of studies have explored the role of Exos in IDD, but these results show that Exos may play a “double-edged sword” role in IDD.466,467 Depending on the cellular source and contents, Exos can both accelerate IDD progression by promoting cellular senescence, inflammatory responses and vascularisation,468,469 while some Exos can also protect against IVD by promoting NP cell survival, proliferation and ECM synthesis.470,471 Currently, Exos can be loaded with small molecule drugs, proteins and nucleic acids through loading incubation, electroporation technology, ultrasonic treatment, extrusion and endogenous loading.472 However, low drug loading efficiency is a significant drawback.473 Therefore, Exos are often combined with other nanocarriers to enhance their delivery efficiency.473,474 For example, in 2021, Xing et al combined dECM with Exos derived from ADSCs (dECM@Exo) for the treatment of IDD73 (Figure 13). The hydrogel provides an environment for NP cell growth, while its sustained release of Exos inhibits NP cell pyroptosis and ECM degradation. In vivo experimental results showed that the dECM@Exo hydrogel system maintained the homeostasis of the early IVD microenvironment and improved IDD. Although we have gained some understanding of the role of Exos in IDD, the research on Exos as drug delivery carriers is still focused on the field of tumor research.475,476 Given the huge differences in pathophysiology between IDD and tumors, further research is needed on the effectiveness and safety of Exos as drug carriers in the future.
Special Stimuli-Responsive Drug Delivery System
Stimuli-responsive nano-drug delivery systems are smart drug carriers that can control drug release based on changes in the internal or external environment.477 The advantage of this system lies in its capacity to enhance treatment efficacy and minimize side effects by precisely delivering the drug to the disease site.25
Internal Stimuli-Responsive Drug Delivery System
Redox-Responsive Nano-Drug Delivery System
We have previously described the disturbance of redox state in degenerating IVDs, which can induce the transformation of physicochemical properties, structural changes, functional regulation and degradation of redox-responsive NDDSs, thereby achieving controlled and targeted drug release.478 Compared with other carriers, redox-responsive materials remain stable in normal tissues but target lesions, which can significantly reduce the biotoxicity of carriers and drugs and increase drug utilization.479 Secondly, it releases cargo under the stimulation of ROS. In particular, redox-responsive materials themselves can alleviate oxidative stress by eliminating the corresponding oxidative species.480 In addition, redox-responsive materials can precisely locate subcellular locations, including the cytoplasm, mitochondria, and nucleus.481,482 Redox-responsive materials include two drug release mechanisms: amphiphilic transition and bond cleavage.478 For the former, the transformation of NPs from hydrophobic to hydrophilic properties leads to the dissociation of polyphenylene sulfide, hydrophobic monosulfide, monosilane, or monotellurium polymer structures and subsequent cargo release.483 The functional bonds of the latter (such as diselenide bonds, disulfide bonds, arylboronic acid bonds, succinimide-thioether bonds, and tetrasulfide bonds) are completely broken under the action of ROS, resulting in the release of cargo.484
IL-1β is a major component of the inflammatory microenvironment of degenerating IVDs. Under oxidative stress conditions, it is produced by lysosomal-independent secretory autophagy in macrophages, in which tripartite motif containing 16 (TRIM16) plays an important role.342 Based on this, Zheng et al designed a ROS-responsive drug delivery system.342 They loaded water-soluble MR409 (TRIM16/LC3B reducing agent) into ROS-responsive vesicles composed of polyphenylene sulfide-PEG (PPS-PEG) amphiphilic polymers. These vesicles were then embedded in a hydrogel composed of PLGA and PEG for protection and controlled release. The system reaches a stable solid state at around 37°C, and can achieve rapid release of MR409 in the presence of H2O2 (more than 80% release in 40 hours), thereby significantly improving oxidative stress and inflammatory microenvironment. However, the degradation products of PLGA (lactic and glycolic acids) can reduce the local pH, which may lead to a deterioration of the IVD microenvironment.485 In another study, Yu et al constructed a diselenide block copolymer-based isoginkgetin (IGK)-loaded NPs (IGK@SeNPs), which showed excellent biocompatibility and low cytotoxicity.343 Compared with the control group, the system showed a cumulative release rate of 63% of IGK in H2O2 solution within 36 h, showing sensitive ROS responsiveness. With the combined action of Se-Se bond in diselenide and IGK, intracellular ROS are effectively eliminated. In addition, the researchers developed a rapamycin-loaded ROS-scavenging Gel hydrogel scaffold (Rapa@Gel).344 The hydrogel degraded slowly over 4 weeks without any obvious inflammatory response at the injection site, indicating that the hydrogel has excellent degradability and biocompatibility. In the presence of H2O2, rapamycin can achieve slow and sustained release and inhibit the inflammatory response by promoting M2 polarization of macrophages. Compared with traditional drugs, gas molecules can penetrate dense connective tissues more easily to reach damaged tissues due to their stronger permeability.486 Due to its unique biochemical properties, NO has the ability to antioxidant, anti-inflammatory, accelerate tissue regeneration and collagen deposition.487,488 Based on this, the researchers synthesised NO-loaded biometal-organic frameworks (BioMOFs) and encapsulated them in modified polyvinyl alcohol and PCL composite electrospun fibre membranes with excellent ROS responsiveness.345 The results showed that the system could respond to the high oxidative stress environment in damaged AF tissue to achieve sustained NO release, thereby repairing AF damage by scavenging ROS, inhibiting the expression of inflammatory cytokines, promoting macrophage M2 polarization, and alleviating mitochondrial dysfunction. In a rat model of AF defect, this systemic treatment effectively repaired AF and improved the height and water content of the degenerated IVD (Figure 14).
Although redox-responsive NDDSs show excellent efficacy in alleviating oxidative stress in degenerative IVDs, there are still some issues worth considering. Firstly, it is necessary to systematically detect the redox status of IVDs at different stages of degeneration to distinguish the different ROS levels in normal and degenerated IVDs. Secondly, the sensitivity of the material depends on the specific functional groups and structure of the polymer, which may require individualized design at different disease stages. In addition, redox-responsive NDDSs should have good biocompatibility, degradability, and low cytotoxicity, which can avoid causing additional damage. Finally, while ensuring adequate drug concentration in the IVD, the rate of drug release should be delayed as much as possible through solubility changes and bond cleavage, which can increase efficacy and patient compliance.
pH -Responsive Nano-Drug Delivery System
Compared with the pH of normal IVD (7.1), the pH of mildly, moderately, and severely degraded IVD is 6.8, 6.5, and 6.2, respectively.489,490 This observed variation in pH serves as the foundation for the application of pH-responsive NDDSs. pH-responsive NDDSs adjust the pH response range by changing the material structure to achieve targeted drug delivery at the lesion site.491,492 At present, the mainstream response mechanisms of pH-sensitive materials include pH-sensitive bond cleavage and chemical group protonation.493 pH-sensitive chemical bonds (including hydrazone bonds, imine bonds, ester bonds, amide bonds, metal coordination bonds, and non-covalent bonds) act as connectors between drugs and carriers, and undergo conformational changes in response to the pH value of the surrounding environment to achieve controlled release of drugs.483 The latter NPs contain ionizable functional groups (such as carboxylic acids and amines), which can undergo protonation and deprotonation at different pH values, resulting in changes in their physical properties or structures, thereby achieving drug release.493 Based on these properties, researchers loaded antioxidant drugs onto pH-responsive NDDSs to alleviate oxidative stress in degenerated IVDs.
Hydrogen (H2) is a promising antioxidant and anti-inflammatory substance, but its concentration in tissues is difficult to maintain for a long time.494 In 2023, Wang et al developed a pH-responsive hollow polydopamine (HPDA) loaded with ammonia borane (AB) H2 sustained-release system (AB@HPDA).346 AB releases H2 efficiently in an acid-responsive manner, while HPDA has the characteristics of large specific surface area, biocompatibility and strong permeability, which can effectively encapsulate AB. Compared with the sham operation group, AB@HPDA can significantly inhibit oxidative stress, inflammatory response and ECM degradation in acupuncture-induced degenerative IVD in rats. Peroxisome proliferator-activated receptor-ɣ coactivator 1α (PGC1α) activates nuclear respiratory factor-1/2 (NRF1/2) to control the expression of mitochondrial transcription factor A (TFAM), which serves as a key regulator of mitochondrial biogenesis and redox homeostasis.495,496 PGC1α is significantly downregulated in degenerative IVD and is involved in regulating cellular senescence in IVD.497 Researchers have developed a pH-responsive and senescent NP cell-targeted PGC1α inducer delivery system347 (Figure 15).They coated mesoporous silica (SiO2) NPs encapsulating PGC1α inducers (SP) with natural killer cell group 2D (NKG2D, which recognises NKG2D ligands up-regulated by senescent cells) overexpressing NP cell membranes (NNPm) (SP@NNPm), which in turn, through the dual-targeting mechanism of homologous membrane binding and receptor-ligand interactions targeting senescent NP cells. SP@NNPm treatment can significantly improve NP cell mitochondrial dysfunction and mtROS production through the PGC1α –NRF2/TFAM signaling axis, thereby inhibiting SASP-related paracrine effects and delaying the progression of IDD. TGF-β and CAT are important intracellular signalling molecules,152,498,499 yet their direct delivery to Degenerated IVD tissues results in rapid protein release and degradation. Zheng et al constructed a mineralized nanoparticle (MNP) loaded with GelMA microspheres (GMNP) composed of CAT and TGF-β through biomimetic mineralization and microfluidics technology348 (Figure 16). The calcium carbonate mineralized layer in GMNPs can respond to the acidic environment and release encapsulated TGF-β and CAT. In vitro, GMNPs can inhibit the overactivation of the TXNIP/NLRP3/IL-1β signaling axis, thereby alleviating oxidative stress, inflammatory response, and ECM degradation in NP cells. In the rat IDD model, GMNP treatment could continuously improve intervertebral disc height and water content within 8 weeks, demonstrating its ability to promote the regeneration of degenerated IVD.
The pH-responsive NDDSs utilize the subtle differences in pH between normal and degenerated intervertebral discs, which requires the pH-sensitive functional groups to exhibit sensitive responses to slight changes in acidic microenvironments. The acidic environment in various subcellular organelles is the most commonly used stimulus to trigger the release of drugs from pH-responsive NDDSs. It is important to be aware that some pH-sensitive nanomaterials can induce lysosomal membrane damage and the release of hydrolases, leading to cell autophagy and death.500,501 In addition, we must consider the biocompatibility and degradation properties of pH-responsive NDDSs, as well as the cytotoxicity of the residues.502 For example, the toxic mechanisms of SiO2 NPs include oxidative stress, inflammation, and cell apoptosis, and these toxic effects involve almost all organs in the human body.503 In addition, nanosized and negatively charged SiO2 NPs also induced immunosuppression by inhibiting lymphocyte proliferation and NK cell activity.504 Therefore, pH-responsive NDDSs are still a long way from the laboratory to clinical treatment.
Enzyme-Responsive Nano-Drug Delivery System
Enzyme-responsive NDDSs are an advanced drug delivery technology that utilizes the activity of specific enzymes to control the release of drugs. It usually consists of two parts, including enzyme-sensitive nanomaterials and therapeutic agents wrapped in biodegradable encapsulation materials.505 After being acted upon by specific enzymes, enzyme-sensitive materials release drugs through size shrinkage, surface charge conversion, surface ligand activation, and chemical bond cleavage, which helps increase bioavailability and reduce adverse side effects in the body.478 The titers of these enzymes are often positively correlated with the severity of IDD, thus achieving precise controlled release.83,506 In addition, the target enzymes are localized in different subcellular compartments in IVD cells, which facilitates selective targeted drug delivery.507
Protease can catalyze the hydrolysis of peptide bonds within protein molecules, thereby breaking down large molecular proteins into smaller peptides or amino acids.508 During IVD degeneration, the expression of multiple proteases is abnormally upregulated (including MMPs, ADAMTS, cathepsins, and hyaluronidase),76,391,509 which can act as regulatory enzymes of enzyme response NDDSs and mediate the precise release of drugs at the target site. miRNA-29a is an endogenous ncRNA that regulates the expression of multiple ECM components to inhibit abnormal fibrosis and collagen conversion,510 which may help alleviate NP fibrosis in degenerated IVD. In view of the lack of effective miRNA delivery systems, Feng et al developed a MMPs-responsive nucleic acid delivery system349 (Figure 17). The shell of the encapsulated miRNA-29a carrier is composed of cationic block copolymer micelles and hydrogel. In the presence of MMPs, miRNA-29a achieves a two-stage delivery in a responsive and sustained manner. In a rabbit IDD model, in situ injection of this system significantly inhibited NP fibrosis and increased IVD regeneration capacity. In addition to proteases, esterases can also be used in enzyme-responsive NDDSs.511 NP progenitor cells (NPPCs) derived from NP tissue are an ideal cell source for stem cell therapy of IDD.512 However, the hypoxic, acidic, and inflammatory microenvironment of IDD severely limits the survival and function of transplanted stem cells.513,514 To overcome these challenges, Xia et al prepared esterase-responsive ibuprofen copolymer nanomicelles PEG-polyisobutene (PIB), which were internalized by NPPCs to form PEG-PIB-premodified NPPCs.350 Under the stimulation of the acidic microenvironment of the degenerated IVD, the increased esterase activity in the transplanted stem cells led to the sustained release of ibuprofen, which in turn effectively inhibited NPPCs pyroptosis and increased its therapeutic effect through the COX2/NF-κB/Caspase-1 signaling pathway.
At present, a variety of enzyme-responsive NDDSs have been successfully developed for promoting the repair of degenerated IVDs with efficient drug delivery performance and good application prospects. However, no studies have attempted to alleviate oxidative stress in IDD. Prior to undertaking the task, it is imperative to quantitatively assess alterations in enzyme content within IVDs.
Multi-Stimulus-Responsive Nano-Drug Delivery System
There are complex microenvironmental changes in degenerating IVDs, including hypoxia, low pH, oxidative stress, and promotion of catabolism. Therefore, combining the metabolic characteristics of two or more microenvironments can achieve multi-stimulus responsive drug delivery, which will further improve the targeting and side effects of drug delivery.515 For example, embedding a redox-responsive cross-linked interlayer in pH-sensitive peptide micelles can trigger nanocarrier disassembly and release of encapsulated drugs in a reducing agent-rich environment.501 In addition, Li et al prepared cross-linked nanogels containing disulfide bonds, which showed pH, temperature and reduction responsiveness.516
As an important signaling molecule, H2S plays an important role in various diseases by participating in multiple pathophysiological processes.517,518 In terms of antioxidation, H2S has been proven to protect cells from oxidative stress by neutralizing ROS molecules, upregulating the expression of antioxidant enzymes, and modifying cysteine residues on key signaling molecules.519,520 Existing evidence suggests that H2S has a protective effect on IDD.521 However, traditional H2S donors (such as NaHS and Na2S) are difficult to provide continuous and stable H2S output.522 At the same time, the rapid removal of H2S donors from the lesion and the volatilization of H2S gas limit the therapeutic effect of H2S.523 Based on this, Zheng et al prepared a pH and enzyme dual-responsive H2S release system with low cytotoxicity and biocompatibility.351 They encapsulated JK1 (a H2S-releasing agent based on phosphoramidate thioester) into collagen hydrogel (Col-JK1). High levels of MMPs in IDD lead to collagen degradation and release of JK1, which is gradually degraded in a low pH environment to produce H2S. In the rat IDD model, Col-JK1 treatment can significantly improve the imaging and histological changes of IDD within 16 weeks. The advantages of MnO2 NPs as drug delivery carriers have been mentioned above, and they are highly sensitive to H2O2 and pH.524,525 Zhu et al reported a smart drug delivery system based on hollow MnO2 NPs encapsulating TGF-β3 (TGF-β3/MnO2), which can dissociate and release loaded TGF-β3 under low pH and H2O2 environment.352 Compared with simple TGF-β3 and MnO2 NPs, TGF-β3/MnO2 was superior in inhibiting H2O2-induced ECM degradation, ROS production, and apoptosis of NP cells. In vivo, TGF-β3/MnO2 ameliorates IDD progression and promotes regeneration in rats. In addition, spermidine (SPD) is a natural biogenic amine that participates in various physiological activities such as DNA synthesis, protein folding, and cell apoptosis.526 Importantly, SPD can reduce excessive accumulation of ROS by protecting mitochondrial function.527 Based on this, the researchers developed a smart drug delivery system that encapsulates SPD with PDA (SPD/PDA).353 After SPD/PDA was exposed to low pH and H2O2 environment, PDANPs dissociated and released SPD, thereby inhibiting ROS production and inducing macrophage M2 polarization, ultimately alleviating the progression of IDD (Figure 18).
Although these systems demonstrate targeted and controlled drug release advantages, their triggering conditions are too complex, which may lead to the failure of encapsulated drug release. To determine the feasibility of these strategies, modulation of responses to each stimulus needs to be demonstrated both in vitro and in vivo.
External Stimulus-Responsive Drug Delivery System
External stimulus-responsive NODDs can control drug release through a variety of external signals (such as light, temperature, magnetism, and ultrasound).528 Compared with internal stimuli that are complex and difficult to control, external stimuli can precisely control drug release in time and space through non-invasive means.528,529 Therefore, not only can potentially damage to normal tissues be minimized, but also stimulation can be added, removed, provided multiple times and continuously for drug delivery and treatment as needed for disease development. These advantages suggest that external stimuli-responsive drug delivery systems have good prospects in the treatment of IDD.
Thermo-Responsive Nano-Drug Delivery System
Thermo-responsive NDDSs control drug release based on the temperature difference between the lesion site and normal tissue, which requires that the properties of at least one component of the nanocarrier material change dramatically and nonlinearly with temperature.501 When the temperature is below or above a critical temperature, thermosensitive materials undergo structural changes, leading to the dissolution of drug encapsulation layers or the formation of pores, thereby releasing drugs.530 Currently, common temperature-sensitive materials include functionalized modified liposomes,531 poly(N-isopropylacrylamide) (PNIPAM),532 poly(amidoamine) (PAMAM),533 poly(2-oxazoline) (POxs)534 and poly[2-(2-methoxyethoxy) ethyl methacrylate] (PMEOMA).535
In 2024, Wang et al developed a temperature-sensitive hydrogel composed of CS and glycerophosphate (GP), in which MnOx NPs with excellent water dispersibility were uniformly incorporated (CS@MnOx)354 (Figure 19). The hydrogel is an injectable liquid at 4°C and cross-links into a cavity-rich hydrogel at 37°C. Co-culture of BMSCs with CS@MnOx significantly down-regulated ROS production and increased the expression of SOD and CAT to inhibit oxidative stress, thereby ameliorating apoptosis, senescence and NP-like differentiation of BMSCs. Compared with the sham surgery group, CS@MnOx +BMSCs treatment efficiently restores intervertebral height and IVD water content. In addition, there are a variety of thermo-responsive hydrogels for the treatment of IDD, including PNIPAM-based temperature-sensitive hydrogels,536–538 dECM temperature-sensitive hydrogels73 and PLGA-PEG-PLGA temperature-sensitive hydrogels.539 Although the size of these materials has not yet reached the nanoscale, the delivery of small molecule drugs, ncRNA and stem cells using them as carriers can effectively improve the microenvironment of degenerative IVDs and promote repair. Similarly, NPs can also be endowed with temperature-sensitive properties. For example, researchers have developed a thermo-responsive nanosphere loaded with diclofenac and cartocinine, whose diameter increases as the temperature decreases (650nm at 4°C and 305nm at 37°C).540 Therefore, local cold therapy can promote the release of drugs from nanospheres, thereby improving synovial inflammation and delaying the progression of osteoarthritis in rats. Furthermore, thermosensitive liposomes can achieve temperature-responsive drug release by undergoing phase transitions in their constituent lipids and associated conformational changes in lipid bilayers, rendering them extensively employed in malignancy research.541,542
Currently, the challenge of thermo-responsive NDDSs is to increase biocompatibility and reduce cytotoxicity while maintaining their thermosensitive properties. Because the thermo-responsive nanocarriers based on the mainstream thermosensitive material PNIPAM are not degradable in the body, which may bring therapeutic side effects.541
Light-Responsive Nano-Drug Delivery System
Light-responsive NDDSs are typically made of nanoscale carrier materials, which can encapsulate drugs and remain stable in the body.501 The surface of the nanocarrier is modified with photosensitizers or other photo-responsive components that are sensitive to light in a specific spectral range (UV, visible and near-infrared light).541 When the photosensitive carrier is stimulated by light of corresponding wavelengths, the photosensitive groups within it can be activated through photoisomerization, photolysis, photocrosslinking, photooxidation-reduction and photothermal triggering, thereby releasing drugs.37 Compared with other carriers, photosensitive carriers can be manipulated with high temporal and spatial precision by adjusting light intensity, emission wavelength, pulse length, exposure time and light direction.543,544
As early as 2014, Kumar et al constructed a photocurable, biocompatible, degradable and injectable synthetic polymer hydrogel.545 The hydrogel solidified under 2.5 minutes of UV irradiation and did not significantly increase the genotoxicity of BMSCs. Under hypoxic conditions, the photocurable hydrogel significantly enhances the NP cell-like differentiation ability of BMSCs. Coincidentally, Chen et al also prepared a photocrosslinked hydrogel that promotes NP-like differentiation of adipose stromal cells (ASC).546 This hydrogel uses UV-induced cross-linked GelMA as the backbone, and hyaluronic acid methacrylate (HAMA) is added in a certain proportion to improve its mechanical properties. In vivo, the combination of hydrogel composites and ASCs promoted the repair of degenerated IVD in rats while avoiding osteophyte formation caused by direct injection of stem cells. Recently, researchers have synthesized a biodegradable photocrosslinked hydrogel with collagen and tyramine-modified HA as the backbone and non-toxic riboflavin as the photocrosslinker.547 This hydrogel solidified after being exposed to blue light for 3 minutes, and culturing BMSCs in it significantly increased ECM synthesis.
Although these studies have shown that photoresponsive nanomaterials can be used to repair IDD, no studies have yet explored the use of photoresponsive drug carriers to improve the oxidative stress microenvironment of IDD. In addition, compared with the non-nanomaterials mentioned above, nanoscale carriers have the advantages of high specific surface area, excellent biodistribution and strong targeting.548 At present, commonly used photoresponsive NDDSs include metal NPs, metal sulfides, metal organic frameworks, carbon nanomaterials, semiconductor NPs, stimuli-responsive micelles, liposome NPs and various organic dyes,541,544,549 which may be used to repair the oxidative stress microenvironment in degenerative IVD in the future. In addition, for triggering light, UV light has low penetration depth and the risk of cell damage, which limits its clinical application.550,551 Near-infrared light with a wavelength of 700–1000 nm has strong penetration and low cytotoxicity, which may be more suitable as a trigger light.552 However, the lower energy of near-infrared light may not be sufficient to induce photochemical effects, which may lead to decreased drug release efficiency.549 Therefore, future attention should be paid to both the update of photoresponsive NDDSs and the optimization of triggering light.
Ultrasound-Responsive Nano-Drug Delivery System
With the development of modern medicine, ultrasound has become an indispensable and important means for the diagnosis and treatment of various diseases. Due to its non-invasive, cost-effective, safe, easy-to-operate, and highly penetrative characteristics, ultrasound has broad prospects in the field of drug delivery.553 Currently, ultrasound-responsive NDDSs induce drug release mainly through cavitation, mechanical effects and local thermal effects.554,555 Ultrasound also increases the permeability of cell membranes through the double-layer acoustic wave effect.556,557 In addition, both stable and inertial cavitation can form temporary holes in cell membranes through the “sonoporation” to increase drug delivery and thus improve efficacy.558,559 Moreover, related studies have shown that low-intensity pulsed ultrasound can promote ECM production in NP cells and reduce inflammatory responses, which shows potential in the treatment of IDD.560,561
Microbubbles are the most common ultrasound-responsive drug delivery method.562 Microbubbles are usually composed of a gas core and a surfactant shell, which can expand, shrink or even rupture under ultrasound pressure, thereby releasing drugs to the desired location.556 Common gas cores are usually inert and non-metabolizable high molecular weight perfluorocarbons (PFCs) or sulfur hexafluoride (SF), which prevent dissolution and improve bubble stability.553 In 2018, Shen et al constructed an ultrasound-responsive resveratrol (RES)-loaded PLGA nanobubble with PFC as the gas core and N-cadherin antibody as the surface modification.355 The NPs imploded under external ultrasound induction, which promoted drug delivery through the released microjets and effectively delayed IVD degeneration in rats. In addition to PFC and SF, NO563,564 and H2565 can also serve as the gas core of microbubbles, and these gases have been shown to effectively alleviate oxidative stress in IDD. In addition to microbubbles, nanodroplets can also be used for ultrasound-responsive NDDSs, which have similar structures and fabrication mechanisms to microbubbles.553 In 2020, Nguyen et al developed nanoscale phase-change PFC droplets loaded with simvastatin.356 Multiple exposures to high-intensity focused ultrasound (HIFU) can lead to a phase transition of nanodroplets into bubbles, thereby inducing the release of simvastatin. The nanodroplets exhibited high stability, low cytotoxicity, and controllable triggered release in an ex vivo rabbit IVD for at least 14 days. In addition to microbubbles and droplets, micelles, liposomes and mesoporous SiO2 NPs can also be used as ultrasound-responsive nanocarrier materials562,566 and carry antioxidant drugs for the treatment of IDD in the future.
Although ultrasound-responsive drug carriers have many advantages, there are still many challenges in balancing the sensitivity of ultrasound response and the stability of the carrier in vivo.567 In addition, bone has a significant attenuation effect on ultrasound, which may affect its penetration of the targeted IVD. This effect may be ameliorated by changing the frequency, gain compensation or the use of specialised probes.
Magnetic-Responsive Nano Drug-Delivery System
Magnetic-responsive NDDSs are an advanced drug delivery technology that utilizes magnetic NPs as carriers and controls drug release through an external magnetic field.568 At present, common magnetic NPs include iron oxide, cobalt oxide, nickel oxide and magnetic SiO2.37 Among them, superparamagnetic iron oxide (SPION) NPs are widely used due to their high level of superparamagnetism, biocompatibility, improved biodistribution, efficient cellular uptake, and minimal toxicity.569 In addition, some magnetic polymer composites have also been developed. For example, polysaccharides can be functionalized with other polymers or inorganic NPs to synthesize synergistic smart polysaccharide-based magnetic hydrogels with magnetic responsiveness and mechanical stability.570 For the external magnetic field, it mainly includes constant magnetic field and alternating magnetic field (AMF).571 The constant magnetic field can focus the magnetic field on the biological target, thereby guiding the accumulation of the injectable carrier at the target site.572 At the same time, the constant magnetic field can also induce the release of loaded drugs. For example, iron gel consisting of SPION NPs and Pluronic-F127 micelles will undergo aggregation of iron oxide NPs in response to an external magnetic field, which in turn releases the drug by squeezing the micelles.483 When magnetic NPs are in AMFs, they generate thermal effects through hysteresis loss and Néel relaxation,573 which can lead to detachment of the coating material or changes in the nanocarrier structure of magnetic NPs, such as increase in bilayer pores,574 disintegration of Fe3O4 cores,501 and deformation of the lattice of single-crystal nanoshells.575
Despite the numerous advantages of magnetic responsive NDDSs, their application is limited by the complexity of external magnetic field setup and the potential biotoxicity of magnetic NPs.570,576 Currently, the research on magnetic responsive NDDSs is mainly focused on the field of tumors,577 and there is no application in the treatment of IDD.
Conclusions and Future Perspectives
IDD is a common age-related musculoskeletal disorder, which is characterised by high morbidity, high disability and high healthcare costs. Numerous studies have shown that oxidative stress is an important factor in accelerating IDD. Oxidative stress not only affects the survival and metabolic characteristics of IVD cells by activating downstream signaling pathways, but also interferes with endogenous and exogenous repair by promoting the deterioration of the IVD microenvironment. The current primary objective in the treatment of IDD is to manage clinical symptoms rather than ameliorate pathological changes in degenerative IVD. Meanwhile, these conventional treatment modalities also exhibit limitations including significant invasiveness, numerous systemic adverse reactions, suboptimal patient adherence, and a heightened risk of recurrence. Although some emerging therapies (growth factors, cell therapies, and gene editing technologies) have been tested in preclinical and clinical studies over the past few decades, these approaches have not yet been truly applied in the clinic. The main obstacles leading to repair failure include the avascular structure of IVD, the harsh microenvironment, and the insufficient endogenous repair potential. Fortunately, rapid advances in nanomedicine and NDDSs have brought light to the treatment of IDD. These disruptive and rapidly developing approaches have the potential to address the challenges of current treatment approaches and provide innovative solutions for repairing and reversing IDD.
In this study, we conducted a comprehensive review of NDDSs that have the potential to enhance the oxidative stress microenvironment. We thoroughly analyzed their strengths, weaknesses, and future prospects for development. The ideal NDDSs should possess the advantages of biocompatibility, degradability, high drug encapsulation efficiency, controllable release, robust mechanical performance, low toxicity and facile clinical application. Based on the current study, we believe that nanohydrogels may be relatively more promising NDDSs for IDD repair. As NP repair and drug carrier, nanohydrogels not only have similar mechanical properties to natural tissues, but also provide ideal microenvironment to improve the proliferation, differentiation and signal transduction of endogenous or exogenous cells. In addition, nanohydrogels can achieve multifunctional properties by incorporating functionalized nanoparticles, such as improved controlled drug release, enhanced targeting, and increased cell adhesion. However, nanohydrogels also have limitations for the treatment of IDD and are not a perfect repair medium. For example, nanohydrogels have a better repair effect on NP, but the repair effect on AF is quite limited. In this regard, nanofiber scaffolds can better repair defective AF tissue. In addition, in order to better target lesion tissues to improve the efficacy of the drug and reduce side effects, stimuli-responsive drug delivery systems were introduced into drug delivery. In particular, redox-responsive NDDSs, which target diseased tissues or subcellular regions, can significantly improve the oxidative stress microenvironment in degenerative IVDs. However, stimuli-responsive drug delivery systems also face significant challenges in moving from the laboratory to clinical success. Internal stimuli-responsive NDDSs are dependent on changes in the microenvironment within the IVD and carrier-specific functional groups, demanding conditions that may limit their clinical application. As for external stimuli-responsive NDDSs, they often have difficulty in distinguishing between normal and diseased tissues, and therefore cannot be truly targeted. In summary, these finer vectors often involve limited application conditions or complex synthetic techniques, which make large-scale production and clinical application of vectors more difficult. Therefore, more active targeting strategies for identifying biomarkers in diseased tissues or cells should be introduced in the future. Furthermore, although many potential antioxidants have shown some efficacy in the treatment of IDD. However, it is undeniable that antioxidant therapy alone is not sufficient to halt or reverse the degenerative process. To achieve better clinical outcomes, NDDSs may need to be combined with other more cutting-edge treatments, such as NDDS combined with stem cells or gene therapy. These treatments will help to further enhance targeting and precision while encompassing the multiple pathological processes involved in IVD degeneration.
Although NDDSs have made great progress in treating IDD, they are still a long way from true clinical application, which involves many regulatory and ethical considerations. The progression of IDD is a chronic process, so the efficacy and safety of NDDSs need to be determined by long-term observation. However, the experimental periods of current studies are relatively short. Secondly, the good biocompatibility and biodegradability of nanocarriers is a prerequisite for their wide application, so minimizing immunogenicity and biotoxicity is the focus for further optimization. Currently, regeneration of NP is the current focus of attention, but future treatment should focus on synergistic recovery of NP, AF and CEP. In addition, the model animals currently used to evaluate the therapeutic effects of IDD are mainly rats and rabbits. The anatomical structure and tissue composition of these two animal IVDs differ greatly from humans,578 which may affect the accuracy of evaluation results. In comparison, the cell and ECM composition of sheep IVD is similar to that of humans, and may be an ideal experimental animal for studying IDD treatment.579,580 Finally, the relief of low back and leg pain is the main purpose of patients seeking medical treatment and the main concern of treatment effect. Therefore, it is crucial to establish animal models of IVD-induced pain and reliable evaluation criteria for future clinical applications.
In conclusion, continuous research on the oxidative stress microenvironment of degenerative IVD, development of antioxidant drugs or treatment methods, and optimization of drug delivery strategies may potentially achieve a comprehensive treatment for IDD.
Data Sharing Statement
Data sharing not applicable to this article as no data sets were generated or analysed during the current study.
Acknowledgments
This work was supported by the Cultivation Project of Xi’an Health and Construction Commission (Program No. 2024ms14) and the Innovative Talent Promotion Plan-Technology Innovation Team (Program No. 2024RS-CXTD-86).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wong CK, Mak RY, Kwok TS, et al. Prevalence, Incidence, and Factors Associated With Non-Specific Chronic Low Back Pain in Community-Dwelling Older Adults Aged 60 Years and Older: a Systematic Review and Meta-Analysis. J Pain. 2022;23(4):509–534.
2. Buchbinder R, van Tulder M, Öberg B, et al. Low back pain: a call for action. Lancet (London, England). 2018;391(10137):2384–2388. doi:10.1016/S0140-6736(18)30488-4
3. Hartvigsen J, Hancock MJ, Kongsted A, et al. What low back pain is and why we need to pay attention. Lancet (London, England). 2018;391(10137):2356–2367. doi:10.1016/S0140-6736(18)30480-X
4. Mohd Isa IL, Teoh SL, Mohd Nor NH, Mokhtar SA. Discogenic Low Back Pain: anatomy, Pathophysiology and Treatments of Intervertebral Disc Degeneration. Int J Mol Sci. 2022;24(1):208. doi:10.3390/ijms24010208
5. Morris LD, Daniels KJ, Ganguli B, Louw QA. An update on the prevalence of low back pain in Africa: a systematic review and meta-analyses. BMC Musculoskeletal Disorders. 2018;19(1):196. doi:10.1186/s12891-018-2075-x
6. Garcia JB, Hernandez-Castro JJ, Nunez RG, et al. Prevalence of low back pain in Latin America: a systematic literature review. Pain Physician. 2014;17(5):379–391. doi:10.36076/ppj.2014/17/379
7. Sharma S, Jensen MP, Pathak A, Sharma S, Pokharel M, Abbott JH. State of clinical pain research in Nepal: a systematic scoping review. Pain Reports. 2019;4(6):e788. doi:10.1097/PR9.0000000000000788
8. Wang Y, Hu S, Zhang W, Zhang B, Yang Z. Emerging role and therapeutic implications of p53 in intervertebral disc degeneration. Cell Death Discovery. 2023;9(1):433. doi:10.1038/s41420-023-01730-5
9. GBD 2015 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990-2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet. 2016;388(10053):1545–1602. doi:10.1016/S0140-6736(16)31678-6
10. Hall JA, Jowett S, Lewis M, Oppong R, Konstantinou K. The STarT Back stratified care model for nonspecific low back pain: a model-based evaluation of long-term cost-effectiveness. Pain. 2021;162(3):702–710. doi:10.1097/j.pain.0000000000002057
11. Kamali A, Ziadlou R, Lang G, et al.. Small molecule-based treatment approaches for intervertebral disc degeneration: current options and future directions. Theranostics. 2021;11(1):27–47. doi:10.7150/thno.48987
12. Bennett RL. The epigenetic underpinnings of lower back pain. Clinical and Translational Medicine. 2022;12(6):e868. doi:10.1002/ctm2.868
13. Yang S, Zhang F, Ma J, Ding W. Intervertebral disc ageing and degeneration: the antiapoptotic effect of oestrogen. Ageing Res Rev. 2020;57:100978. doi:10.1016/j.arr.2019.100978
14. Dickinson PJ, Bannasch DL. Current Understanding of the Genetics of Intervertebral Disc Degeneration. Frontiers in Veterinary Science. 2020;7:431. doi:10.3389/fvets.2020.00431
15. Theodore N, Ahmed AK, Fulton T, et al. Genetic Predisposition to Symptomatic Lumbar Disk Herniation in Pediatric and Young Adult Patients. Spine. 2019;44(11):E640–e649. doi:10.1097/BRS.0000000000002949
16. Song C, Zhou Y, Cheng K, et al. Cellular senescence - Molecular mechanisms of intervertebral disc degeneration from an immune perspective. Biomed Pharmacother. 2023;162:114711. doi:10.1016/j.biopha.2023.114711
17. Cannata F, Vadalà G, Ambrosio L, et al. Intervertebral disc degeneration: a focus on obesity and type 2 diabetes. Diabetes/Metab Res Rev. 2020;36(1):e3224. doi:10.1002/dmrr.3224
18. Zhang GZ, Liu MQ, Chen HW, et al. NF-κB signalling pathways in nucleus pulposus cell function and intervertebral disc degeneration. Cell Proliferation. 2021;54(7):e13057. doi:10.1111/cpr.13057
19. Wang Y, Cheng H, Wang T, Zhang K, Zhang Y, Kang X. Oxidative stress in intervertebral disc degeneration: molecular mechanisms, pathogenesis and treatment. Cell Proliferation. 2023;56(9):e13448. doi:10.1111/cpr.13448
20. Kloppenburg M, Berenbaum F. Osteoarthritis year in review 2019: epidemiology and therapy. Osteoarthritis Cartilage. 2020;28(3):242–248. doi:10.1016/j.joca.2020.01.002
21. Xin J, Wang Y, Zheng Z, Wang S, Na S, Zhang S. Treatment of Intervertebral Disc Degeneration. Orthopaedic Surgery. 2022;14(7):1271–1280. doi:10.1111/os.13254
22. Liu W, Ma Z, Wang Y, Yang J. Multiple nano-drug delivery systems for intervertebral disc degeneration: current status and future perspectives. Bioact Mater. 2023;23:274–299. doi:10.1016/j.bioactmat.2022.11.006
23. Mansour A, Romani M, Acharya AB, Rahman B, Verron E, Badran Z. Drug Delivery Systems in Regenerative Medicine: an Updated Review. Pharmaceutics. 2023;15(2):695. doi:10.3390/pharmaceutics15020695
24. Zhang S, Wang L, Kang Y, Wu J, Zhang Z. Nanomaterial-based reactive oxygen species scavengers for osteoarthritis therapy. Acta Biomater. 2023;162:1–19. doi:10.1016/j.actbio.2023.03.030
25. Yu H, Gao R, Liu Y, Fu L, Zhou J, Li L. Stimulus-Responsive Hydrogels as Drug Delivery Systems for Inflammation Targeted Therapy. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2024;11(1):e2306152. doi:10.1002/advs.202306152
26. Zhang Q, Wang Y, Wu D, Chen Z. Stimuli-responsive nanoscale drug delivery system for epilepsy theranostics. Acta Biomater. 2025;194:58–79. doi:10.1016/j.actbio.2025.01.046
27. Morici L, Allémann E, Rodríguez-Nogales C, Jordan O. Cartilage-targeted drug nanocarriers for osteoarthritis therapy. Int J Pharm. 2024;666:124843. doi:10.1016/j.ijpharm.2024.124843
28. Newell N, Little JP, Christou A, Adams MA, Adam CJ, Masouros SD. Biomechanics of the human intervertebral disc: a review of testing techniques and results. J Mech Behav Biomed Mater. 2017;69:420–434. doi:10.1016/j.jmbbm.2017.01.037
29. Samanta A, Lufkin T, Kraus P. Intervertebral disc degeneration-Current therapeutic options and challenges. Front Public Health. 2023;11:1156749. doi:10.3389/fpubh.2023.1156749
30. Yang B, Wendland MF, O’Connell GD. Direct Quantification of Intervertebral Disc Water Content Using MRI. Journal of Magnetic Resonance Imaging: JMRI. 2020;52(4):1152–1162. doi:10.1002/jmri.27171
31. Song C, Hu P, Peng R, Li F, Fang Z, Xu Y. Bioenergetic dysfunction in the pathogenesis of intervertebral disc degeneration. Pharmacol Res. 2024;202:107119. doi:10.1016/j.phrs.2024.107119
32. Risbud MV, Shapiro IM. Role of cytokines in intervertebral disc degeneration: pain and disc content. Nat Rev Rheumatol. 2014;10(1):44–56. doi:10.1038/nrrheum.2013.160
33. Hunter CJ, Matyas JR, Duncan NA. The notochordal cell in the nucleus pulposus: a review in the context of tissue engineering. Tissue Eng. 2003;9(4):667–677. doi:10.1089/107632703768247368
34. Rodrigues-Pinto R, Richardson SM, Hoyland JA. An understanding of intervertebral disc development, maturation and cell phenotype provides clues to direct cell-based tissue regeneration therapies for disc degeneration. European Spine Journal: Official Publication of the European Spine Society, the European Spinal Deformity Society, and the European Section of the Cervical Spine Research Society. 2014;23(9):1803–1814. doi:10.1007/s00586-014-3305-z
35. Peng H, Qiao J, Wang G, Shi W, Xia F, Qiao R. Dong B: a collagen-rich arch in the urochordate notochord coordinates cell shaping and multi-tissue elongation. Current Biology: CB. 2023;33(24):5390–5403.e5393. doi:10.1016/j.cub.2023.11.001
36. Erwin WM, Inman RD. Inman RD: notochord cells regulate intervertebral disc chondrocyte proteoglycan production and cell proliferation. Spine. 2006;31(10):1094–1099. doi:10.1097/01.brs.0000216593.97157.dd
37. Tang J, Luo Y, Wang Q, Wu J, Wei Y. Stimuli-Responsive Delivery Systems for Intervertebral Disc Degeneration. Int J Nanomed. 2024;19:4735–4757. doi:10.2147/IJN.S463939
38. Koroth J, Buko EO, Abbott R, et al. Macrophages and Intervertebral Disc Degeneration. Int J Mol Sci. 2023;24(2):1367. doi:10.3390/ijms24021367
39. Silwal P, Nguyen-Thai AM, Mohammad HA, et al. Cellular Senescence in Intervertebral Disc Aging and Degeneration: molecular Mechanisms and Potential Therapeutic Opportunities. Biomolecules. 2023;13(4):686. doi:10.3390/biom13040686
40. Zhang A, Cheng Z, Chen Y, Shi P, Gan W, Zhang Y. Emerging tissue engineering strategies for annulus fibrosus therapy. Acta Biomater. 2023;167:1–15. doi:10.1016/j.actbio.2023.06.012
41. Mohd Isa IL, Mokhtar SA, Fauzi MB, Devitt A, Devitt A, Pandit A. Intervertebral Disc Degeneration: biomaterials and Tissue Engineering Strategies toward Precision Medicine. Adv Healthcare Mater. 2022;11(13):e2102530. doi:10.1002/adhm.202102530
42. Chen X, Zhang A, Zhao K, et al. The role of oxidative stress in intervertebral disc degeneration: mechanisms and therapeutic implications. Ageing Res Rev. 2024;98:102323. doi:10.1016/j.arr.2024.102323
43. Molladavoodi S, McMorran J, Gregory D. Mechanobiology of annulus fibrosus and nucleus pulposus cells in intervertebral discs. Cell Tissue Res. 2020;379(3):429–444. doi:10.1007/s00441-019-03136-1
44. Raj PP. Intervertebral disc: anatomy-physiology-pathophysiology-treatment. Pain Practice: the Official Journal of World Institute of Pain. 2008;8(1):18–44. doi:10.1111/j.1533-2500.2007.00171.x
45. Ashinsky BG, Bonnevie ED, Mandalapu SA, et al. Intervertebral Disc Degeneration Is Associated With Aberrant Endplate Remodeling and Reduced Small Molecule Transport. J Bone Miner Res. 2020;35(8):1572–1581. doi:10.1002/jbmr.4009
46. Mwale F, Roughley P, Antoniou J. Distinction between the extracellular matrix of the nucleus pulposus and hyaline cartilage: a requisite for tissue engineering of intervertebral disc. Eur Cells Mater. 2004;8:58–63. doi:10.22203/eCM.v008a06
47. Sun K, Jiang J, Wang Y, et al. The role of nerve fibers and their neurotransmitters in regulating intervertebral disc degeneration. Ageing Res Rev. 2022;81:101733. doi:10.1016/j.arr.2022.101733
48. Zhang H, Yang X, Huang Y, et al. Reviving Intervertebral Discs: treating Degeneration Using Advanced Delivery Systems. Mol Pharmaceut. 2024;21(2):373–392. doi:10.1021/acs.molpharmaceut.3c00579
49. Zhang S, Wang P, Hu B, et al. Inhibiting Heat Shock Protein 90 Attenuates Nucleus Pulposus Fibrosis and Pathologic Angiogenesis Induced by Macrophages via Down-Regulating Cell Migration-Inducing Protein. Am J Pathol. 2023;193(7):960–976. doi:10.1016/j.ajpath.2023.03.014
50. Fournier DE, Kiser PK, Shoemaker JK, Battié MC, Séguin CA. Vascularization of the human intervertebral disc: a scoping review. JOR spine. 2020;3(4):e1123. doi:10.1002/jsp2.1123
51. Nerlich AG, Schaaf R, Wälchli B, Boos N. Temporo-spatial distribution of blood vessels in human lumbar intervertebral discs. Eur Spine J. 2007;16(4):547–555. doi:10.1007/s00586-006-0213-x
52. Wang Z, Chen H, Tan Q, et al. Inhibition of aberrant Hif1α activation delays intervertebral disc degeneration in adult mice. Bone Res. 2022;10(1):2. doi:10.1038/s41413-021-00165-x
53. De Geer CM. Intervertebral Disk Nutrients and Transport Mechanisms in Relation to Disk Degeneration: a Narrative Literature Review. Journal of Chiropractic Medicine. 2018;17(2):97–105. doi:10.1016/j.jcm.2017.11.006
54. Ejeskär A, Holm S. Oxygen tension measurements in the intervertebral disc. A methodological and experimental study. Upsala Journal of Medical Sciences. 1979;84(1):83–93. doi:10.3109/03009737909179143
55. De Santis V, Singer M. Tissue oxygen tension monitoring of organ perfusion: rationale, methodologies, and literature review. Br J Anaesth. 2015;115(3):357–365. doi:10.1093/bja/aev162
56. Madhu V, Hernandez-Meadows M, Boneski PK, et al. The mitophagy receptor BNIP3 is critical for the regulation of metabolic homeostasis and mitochondrial function in the nucleus pulposus cells of the intervertebral disc. Autophagy. 2023;19(6):1821–1843. doi:10.1080/15548627.2022.2162245
57. Johnston SN, Silagi ES, Madhu V, Nguyen DH, Shapiro IM, Risbud MV. GLUT1 is redundant in hypoxic and glycolytic nucleus pulposus cells of the intervertebral disc. JCI Insight. 2023;8(8). doi:10.1172/jci.insight.164883
58. Binch AL, Cole AA, Breakwell LM, et al. Nerves are more abundant than blood vessels in the degenerate human intervertebral disc. Arthritis Res Therapy. 2015;17(1):370. doi:10.1186/s13075-015-0889-6
59. Francisco V, Pino J, González-Gay M, et al. A new immunometabolic perspective of intervertebral disc degeneration. Nat Rev Rheumatol. 2022;18(1):47–60. doi:10.1038/s41584-021-00713-z
60. Hanaei S, Khoshnevisan A, Kepler CK, Rezaei N, Rezaei N. Genetic aspects of intervertebral disc degeneration. Reviews in the Neurosciences. 2015;26(5):581–606. doi:10.1515/revneuro-2014-0077
61. Sambrook PN, MacGregor AJ, Spector TD. Genetic influences on cervical and lumbar disc degeneration: a magnetic resonance imaging study in twins. Arthritis Rheum. 1999;42(2):366–372. doi:10.1002/1529-0131(199902)42:2<366::AID-ANR20>3.0.CO;2-6
62. Videman T, Battié MC, Ripatti S, Gill K, Manninen H, Kaprio J. Determinants of the progression in lumbar degeneration: a 5-year follow-up study of adult male monozygotic twins. Spine. 2006;31(6):671–678. doi:10.1097/01.brs.0000202558.86309.ea
63. Williams FM, Sambrook PN, Jones AF, Spector TD, MacGregor AJ, MacGregor AJ. Progression of lumbar disc degeneration over a decade: a heritability study. Ann Rheumatic Dis. 2011;70(7):1203–1207. doi:10.1136/ard.2010.146001
64. Williams FM, Bansal AT, van Meurs JB, et al. Novel genetic variants associated with lumbar disc degeneration in Northern Europeans: a meta-analysis of 4600 subjects. Ann Rheumatic Dis. 2013;72(7):1141–1148. doi:10.1136/annrheumdis-2012-201551
65. Wu F, Huang X, Zhang Z, Shao Z. A Meta-analysis Assessing the Association Between COL11A1 and GDF5 Genetic Variants and Intervertebral Disc Degeneration Susceptibility. Spine. 2020;45(11):E616–e623. doi:10.1097/BRS.0000000000003371
66. Mogensen MS, Karlskov-Mortensen P, Proschowsky HF, et al. Genome-wide association study in Dachshund: identification of a major locus affecting intervertebral disc calcification. The Journal of Heredity. 2011;102(1):S81–86. doi:10.1093/jhered/esr021
67. Bjornsdottir G, Stefansdottir L, Thorleifsson G, et al. Rare SLC13A1 variants associate with intervertebral disc disorder highlighting role of sulfate in disc pathology. Nat Commun. 2022;13(1):634. doi:10.1038/s41467-022-28167-1
68. TYK A, Lam TK, Peng Y, et al. Transformation of resident notochord-descendent nucleus pulposus cells in mouse injury-induced fibrotic intervertebral discs. Aging Cell. 2020;19(11):e13254. doi:10.1111/acel.13254
69. McCann MR, Séguin CA. Notochord Cells in Intervertebral Disc Development and Degeneration. Journal of Developmental Biology. 2016;4(1):3. doi:10.3390/jdb4010003
70. McCann MR, Bacher CA, Séguin CA. Exploiting notochord cells for stem cell-based regeneration of the intervertebral disc. J Cell Commun Signal. 2011;5(1):39–43. doi:10.1007/s12079-010-0116-9
71. Liao Z, Liu H, Ma L, et al. Engineering Extracellular Vesicles Restore the Impaired Cellular Uptake and Attenuate Intervertebral Disc Degeneration. ACS nano. 2021;15(9):14709–14724. doi:10.1021/acsnano.1c04514
72. Teng Y, Yu H, Wu C, et al. Nimbolide targeting SIRT1 mitigates intervertebral disc degeneration by reprogramming cholesterol metabolism and inhibiting inflammatory signaling. Acta Pharmaceutica Sinica B. 2023;13(5):2269–2280. doi:10.1016/j.apsb.2023.02.018
73. Xing H, Zhang Z, Mao Q, et al. Injectable exosome-functionalized extracellular matrix hydrogel for metabolism balance and pyroptosis regulation in intervertebral disc degeneration. J Nanobiotechnol. 2021;19(1):264. doi:10.1186/s12951-021-00991-5
74. Oichi T, Taniguchi Y, Oshima Y, Tanaka S, Saito T. Pathomechanism of intervertebral disc degeneration. JOR spine. 2020;3(1):e1076. doi:10.1002/jsp2.1076
75. Luo L, Gong J, Wang Z, et al. Injectable cartilage matrix hydrogel loaded with cartilage endplate stem cells engineered to release exosomes for non-invasive treatment of intervertebral disc degeneration. Bioact Mater. 2022;15:29–43. doi:10.1016/j.bioactmat.2021.12.007
76. Vo NV, Hartman RA, Yurube T, Jacobs LJ, Sowa GA, Kang JD. Expression and regulation of metalloproteinases and their inhibitors in intervertebral disc aging and degeneration. Spine J. 2013;13(3):331–341. doi:10.1016/j.spinee.2012.02.027
77. Kang L, Zhang H, Jia C, Zhang R, Shen C. Targeting Oxidative Stress and Inflammation in Intervertebral Disc Degeneration: therapeutic Perspectives of Phytochemicals. Front Pharmacol. 2022;13:956355. doi:10.3389/fphar.2022.956355
78. Kang L, Zhang H, Jia C, Zhang R, Shen C. Epigenetic modifications of inflammation in intervertebral disc degeneration. Ageing Res Rev. 2023;87:101902. doi:10.1016/j.arr.2023.101902
79. Wen P, Zheng B, Ma T, Hao L, Zhang Y, Zhang Y. The role of ageing and oxidative stress in intervertebral disc degeneration. Front Mol Biosci. 2022;9:1052878. doi:10.3389/fmolb.2022.1052878
80. Feng C, Yang M, Liu C, et al. ROS: crucial Intermediators in the Pathogenesis of Intervertebral Disc Degeneration. Oxid Med Cell Longev. 2017;2017(1):5601593. doi:10.1155/2017/5601593
81. Shi ZW, Zhu L, Song ZR, Liu TJ, Hao DJ. Roles of p38 MAPK signalling in intervertebral disc degeneration. Cell Proliferation. 2023;56(8):e13438. doi:10.1111/cpr.13438
82. Chen HW, Zhou JW, Zhang GZ, Luo ZB, Li L, Kang XW. Emerging role and therapeutic implication of mTOR signalling in intervertebral disc degeneration. Cell Proliferation. 2023;56(1):e13338. doi:10.1111/cpr.13338
83. Liang H, Luo R, Li G, Zhang W, Song Y, Yang C. The Proteolysis of ECM in Intervertebral Disc Degeneration. Int J Mol Sci. 2022;23(3):1715.
84. Yang G, Liu X, Chen F, Wang W, Shao Y, Cui X. Astaxanthin suppresses oxidative stress and calcification in vertebral cartilage endplate via activating Nrf-2/HO-1 signaling pathway. Int immunopharmacol. 2023;119:110159. doi:10.1016/j.intimp.2023.110159
85. Aigner T, Gresk-otter KR, Fairbank JC, von der Mark K, Urban JP. Variation with age in the pattern of type X collagen expression in normal and scoliotic human intervertebral discs. Calcified Tissue International. 1998;63(3):263–268. doi:10.1007/s002239900524
86. Grant MP, Epure LM, Bokhari R, Roughley P, Antoniou J, Mwale F. Human cartilaginous endplate degeneration is induced by calcium and the extracellular calcium-sensing receptor in the intervertebral disc. Eur Cells Mater. 2016;32:137–151. doi:10.22203/eCM.v032a09
87. Kritschil R, Scott M, Sowa G, Vo N. Role of autophagy in intervertebral disc degeneration. J Cell Physiol. 2022;237(2):1266–1284. doi:10.1002/jcp.30631
88. Bermudez-Lekerika P, Crump KB, Tseranidou S, et al. Immuno-Modulatory Effects of Intervertebral Disc Cells. Front Cell Develop Biol. 2022;10:924692. doi:10.3389/fcell.2022.924692
89. Wu XT, Wang YX, Sun HH, Feng M, Sun -H-H. Update on the roles of macrophages in the degeneration and repair process of intervertebral discs. Joint Bone Spine. 2023;90(3):105514. doi:10.1016/j.jbspin.2022.105514
90. Sun Z, Liu B, Luo ZJ. The Immune Privilege of the Intervertebral Disc: implications for Intervertebral Disc Degeneration Treatment. Int J Med Sci. 2020;17(5):685–692. doi:10.7150/ijms.42238
91. Capossela S, Schläfli P, Bertolo A, et al. Degenerated human intervertebral discs contain autoantibodies against extracellular matrix proteins. Eur Cells Mater. 2014;27:251–263. doi:10.22203/eCM.v027a18
92. Lisiewski LE, Jacobsen HE, Viola DCM, Kenawy HM, Kiridly DN, Chahine NO. Intradiscal inflammatory stimulation induces spinal pain behavior and intervertebral disc degeneration in vivo. FASEB j. 2024;38(1):e23364. doi:10.1096/fj.202300227R
93. Staszkiewicz R, Gładysz D, Gralewski M, et al. Usefulness of Detecting Brain-Derived Neurotrophic Factor in Intervertebral Disc Degeneration of the Lumbosacral Spine. Medical Science Monitor: International Medical Journal of Experimental and Clinical Research. 2023;29:e938663. doi:10.12659/MSM.938663
94. Damle SR, Krzyzanowska AK, Korsun MK, et al. Inducing Angiogenesis in the Nucleus Pulposus. Cells. 2023;12(20):2488. doi:10.3390/cells12202488
95. Lyu FJ, Cui H, Mc Cheung K, Cao X, Iatridis JC, Zheng Z. Painful intervertebral disc degeneration and inflammation: from laboratory evidence to clinical interventions. Bone Res. 2021;9(1):7. doi:10.1038/s41413-020-00125-x
96. Yu P, Mao F, Chen J, et al. Characteristics and mechanisms of resorption in lumbar disc herniation. Arthritis Res Therapy. 2022;24(1):205. doi:10.1186/s13075-022-02894-8
97. Cheng Z, Xiang Q, Wang J, Zhang Y. The potential role of melatonin in retarding intervertebral disc ageing and degeneration: a systematic review. Ageing Res Rev. 2021;70:101394. doi:10.1016/j.arr.2021.101394
98. Vergroesen PP, Kingma I, Emanuel KS, et al.. Mechanics and biology in intervertebral disc degeneration: a vicious circle. Osteoarthritis Cartilage. 2015;23(7):1057–1070. doi:10.1016/j.joca.2015.03.028
99. Haughton V. Imaging intervertebral disc degeneration. J Bone Joint Surg Am Vol. 2006;88(2):15–20. doi:10.2106/JBJS.F.00010
100. Tamagawa S, Sakai D, Nojiri H, Sato M. Imaging Evaluation of Intervertebral Disc Degeneration and Painful Discs-Advances and Challenges in Quantitative MRI. Diagnostics. 2022;12(3):707.
101. Zhang AS, Xu A, Ansari K, Anderson G, Alsoof D, Daniels AH. Lumbar Disc Herniation: diagnosis and Management. Am J Med. 2023;136(7):645–651. doi:10.1016/j.amjmed.2023.03.024
102. Kreiner DS, Hwang SW, Easa JE, et al. An evidence-based clinical guideline for the diagnosis and treatment of lumbar disc herniation with radiculopathy. Spine J. 2014;14(1):180–191.
103. Hlubek RJ, Mundis Jr GM. Treatment for Recurrent Lumbar Disc Herniation. Current Reviews in Musculoskeletal Medicine. 2017;10(4):517–520. doi:10.1007/s12178-017-9450-3
104. Latka D, Miekisiak G, Jarmuzek P, Lachowski M, Kaczmarczyk J. Treatment of lumbar disc herniation with radiculopathy. Clinical practice guidelines endorsed by The Polish Society of Spinal Surgery. Neurologia i neurochirurgia polska. 2016;50(2):101–108.
105. Sköld C, Tropp H, Berg S. Five-year follow-up of total disc replacement compared to fusion: a randomized controlled trial. European Spine Journal: Official Publication of the European Spine Society, the European Spinal Deformity Society, and the European Section of the Cervical Spine Research Society. 2013;22(10):2288–2295. doi:10.1007/s00586-013-2926-y
106. Berg S, Tropp HT, Leivseth G. Disc height and motion patterns in the lumbar spine in patients operated with total disc replacement or fusion for discogenic back pain. Results from a randomized controlled trial. Spine J. 2011;11(11):991–998. doi:10.1016/j.spinee.2011.08.434
107. Cheung EC, Vousden KH. The role of ROS in tumour development and progression. Nat Rev Cancer. 2022;22(5):280–297.
108. Yang S, Lian G. ROS and diseases: role in metabolism and energy supply. Mol Cell Biochem. 2020;467(1–2):1–12.
109. Bai R, Guo J, Ye XY, Xie Y, Xie T. Oxidative stress: the core pathogenesis and mechanism of Alzheimer’s disease. Ageing Res Rev. 2022;77:101619. doi:10.1016/j.arr.2022.101619
110. Reczek CR, Chandel NS. ROS-dependent signal transduction. Curr Opin Cell Biol. 2015;33:8–13. doi:10.1016/j.ceb.2014.09.010
111. Li Y, Chen L, Gao Y, Zou X, Wei F. Oxidative Stress and Intervertebral Disc Degeneration: pathophysiology, Signaling Pathway, and Therapy. Oxid Med Cell Longev. 2022;2022:1984742. doi:10.1155/2022/1984742
112. Xiang Q, Cheng Z, Wang J, et al. Allicin Attenuated Advanced Oxidation Protein Product-Induced Oxidative Stress and Mitochondrial Apoptosis in Human Nucleus Pulposus Cells. Oxid Med Cell Longev. 2020;2020:6685043. doi:10.1155/2020/6685043
113. Jitjumnong M, Chalermkitpanit P, Suantawee T, Dechsupa S, Vajarintarangoon L, Honsawek S. Telomere Shortening and Increased Oxidative Stress in Lumbar Disc Degeneration. Int J Mol Sci. 2022;23(17):1.
114. Song Y, Li S, Geng W, et al. Sirtuin 3-dependent mitochondrial redox homeostasis protects against AGEs-induced intervertebral disc degeneration. Redox Biol. 2018;19:339–353. doi:10.1016/j.redox.2018.09.006
115. Yang F, Zhu D, Wang Z, et al. Role of Advanced Glycation End Products in Intervertebral Disc Degeneration: mechanism and Therapeutic Potential. Oxid Med Cell Longev. 2022;2022(1):7299005. doi:10.1155/2022/7299005
116. Wang DK, Zheng HL, Zhou WS, et al. Mitochondrial Dysfunction in Oxidative Stress-Mediated Intervertebral Disc Degeneration. Orthopaedic Surgery. 2022;14(8):1569–1582.
117. Palma FR, Gantner BN, Sakiyama MJ, et al.. ROS production by mitochondria: function or dysfunction? Oncogene. 2024;43(5):295–303.
118. Guo Y, Guan T, Shafiq K, et al. Mitochondrial dysfunction in aging. Ageing Res Rev. 2023;88:101955.
119. Zhao RZ, Jiang S, Zhang L, Yu ZB. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). IntJ Mol Med. 2019;44(1):3–15. doi:10.3892/ijmm.2019.4188
120. Fukai T, Ushio-Fukai M. Cross-Talk between NADPH Oxidase and Mitochondria: role in ROS Signaling and Angiogenesis. Cells. 2020;9(8). doi:10.3390/cells9081849
121. Okoye CN, Koren SA, Wojtovich AP. Mitochondrial complex I ROS production and redox signaling in hypoxia. Redox Biol. 2023;67:102926. doi:10.1016/j.redox.2023.102926
122. Read AD, Bentley RE, Archer SL, Dunham-Snary KJ. Mitochondrial iron-sulfur clusters: structure, function, and an emerging role in vascular biology. Redox Biol. 2021;47:102164. doi:10.1016/j.redox.2021.102164
123. Hadrava Vanova K, Kraus M, Neuzil J, Rohlena J. Mitochondrial complex II and reactive oxygen species in disease and therapy. Redox Report: Communications in Free Radical Research. 2020;25(1):26–32. doi:10.1080/13510002.2020.1752002
124. Paddenberg R, Ishaq B, Goldenberg A, et al. Essential role of complex II of the respiratory chain in hypoxia-induced ROS generation in the pulmonary vasculature. Am J Physiol Lung Cell Mol Physiol. 2003;284(5):L710–719. doi:10.1152/ajplung.00149.2002
125. Song Y, Liang H, Li G, et al. The NLRX1-SLC39A7 complex orchestrates mitochondrial dynamics and mitophagy to rejuvenate intervertebral disc by modulating mitochondrial Zn(2+) trafficking. Autophagy. 2024;20(4):809–829. doi:10.1080/15548627.2023.2274205
126. Mao J, Wu Q, Shang Q, et al. SIRT5-related desuccinylation modification of AIFM1 protects against compression-induced intervertebral disc degeneration by regulating mitochondrial homeostasis. Exp Mol Med. 2023;55(1):253–268. doi:10.1038/s12276-023-00928-y
127. Song Y, Lu S, Geng W, et al. Mitochondrial quality control in intervertebral disc degeneration. Exp Mol Med. 2021;53(7):1124–1133. doi:10.1038/s12276-021-00650-7
128. Chen C, Zhou Y, Hu C, Wang Y, Yan Z, Wu R. Mitochondria and oxidative stress in ovarian endometriosis. Free Radic Biol Med. 2019;136:22–34. doi:10.1016/j.freeradbiomed.2019.03.027
129. Brandes RP, Weissmann N, Schröder K. Nox family NADPH oxidases: molecular mechanisms of activation. Free Radic Biol Med. 2014;76:208–226.
130. Bedard K, Krause KH. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev. 2007;87(1):245–313. doi:10.1152/physrev.00044.2005
131. Pecchillo Cimmino T, Ammendola R, Cattaneo F, Esposito G. NOX Dependent ROS Generation and Cell Metabolism. Int J Mol Sci. 2023;24(3):2086. doi:10.3390/ijms24032086
132. Li Z, Chen S, Huang D, Peng Y, Chen S, Ma K. TIGAR impedes compression-induced intervertebral disc degeneration by suppressing nucleus pulposus cell apoptosis and autophagy. J Cell Physiol. 2020;235(2):1780–1794. doi:10.1002/jcp.29097
133. Jiang LB, Cao L, Ma YQ, et al. TIGAR mediates the inhibitory role of hypoxia on ROS production and apoptosis in rat nucleus pulposus cells. Osteoarthritis Cartilage. 2018;26(1):138–148. doi:10.1016/j.joca.2017.10.007
134. Liu C, Liu YJ, Ai X, et al. A positive feedback loop between EZH2 and NOX4 regulates nucleus pulposus cell senescence in age-related intervertebral disc degeneration. Cell Division. 2020;15(1):2. doi:10.1186/s13008-020-0060-x
135. Dai X, Chen Y, Yu Z, et al. Advanced oxidation protein products induce annulus fibrosus cell senescence through a NOX4-dependent, MAPK-mediated pathway and accelerate intervertebral disc degeneration. PeerJ. 2022;10:e13826. doi:10.7717/peerj.13826
136. Liu Q, Tan Z, Xie C, Ling L, Hu H. Oxidative stress as a critical factor might involve in intervertebral disc degeneration via regulating NOXs/FOXOs. Journal of Orthopaedic Science: Official Journal of the Japanese Orthopaedic Association. 2023;28(1):105–111. doi:10.1016/j.jos.2021.09.010
137. Chang HI, Chen CN, Huang KY. Mechanical Stretch-Induced NLRP3 Inflammasome Expression on Human Annulus Fibrosus Cells Modulated by Endoplasmic Reticulum Stress. Int J Mol Sci. 2022;23(14):4.
138. Feng C, Zhang Y, Yang M, Liu H, Huang B, Zhou Y. Oxygen-Sensing Nox4 Generates Genotoxic ROS to Induce Premature Senescence of Nucleus Pulposus Cells through MAPK and NF- κ B Pathways. Oxid Med Cell Longev. 2017;2017(1):7426458. doi:10.1155/2017/7426458
139. Yao D, Chen E, Li Y, et al. The role of endoplasmic reticulum stress, mitochondrial dysfunction and their crosstalk in intervertebral disc degeneration. Cell. Signalling. 2024;114:110986. doi:10.1016/j.cellsig.2023.110986
140. Cui X, Zhang Y, Lu Y, Xiang M. ROS and Endoplasmic Reticulum Stress in Pulmonary Disease. Front Pharmacol. 2022;13:879204. doi:10.3389/fphar.2022.879204
141. Ren J, Bi Y, Sowers JR, Hetz C, Zhang Y. Endoplasmic reticulum stress and unfolded protein response in cardiovascular diseases. Nat Rev Cardiol. 2021;18(7):499–521. doi:10.1038/s41569-021-00511-w
142. Oakes SA, Papa FR. The role of endoplasmic reticulum stress in human pathology. Annual Review of Pathology. 2015;10(1):173–194. doi:10.1146/annurev-pathol-012513-104649
143. Huang J, Zhou Q, Ren Q, Luo L, Ji G, Zheng T. Endoplasmic reticulum stress associates with the development of intervertebral disc degeneration. Front Endocrinol (Lausanne). 2022;13:1094394. doi:10.3389/fendo.2022.1094394
144. Miglioranza Scavuzzi B, Holoshitz J. Endoplasmic Reticulum Stress, Oxidative Stress, and Rheumatic Diseases. Antioxidants (Basel, Switzerland). 2022;11(7). doi:10.3390/antiox11071306
145. Niemann B, Rohrbach S, Miller MR, Newby DE, Fuster V, Kovacic JC. Oxidative Stress and Cardiovascular Risk: obesity, Diabetes, Smoking, and Pollution: part 3 of a 3-Part Series. Journal of the American College of Cardiology. 2017;70(2):230–251. doi:10.1016/j.jacc.2017.05.043
146. Caliri AW, Tommasi S, Besaratinia A. Relationships among smoking, oxidative stress, inflammation, macromolecular damage, and cancer. Mutation Research Reviews in Mutation Research. 2021;787:108365. doi:10.1016/j.mrrev.2021.108365
147. Cheng W, Shi X, Zhang J, et al. Role of PI3K-AKT Pathway in Ultraviolet Ray and Hydrogen Peroxide-Induced Oxidative Damage and Its Repair by Grain Ferments. Foods (Basel, Switzerland). 2023;12(4):3.
148. Wang Q, Yu Q, Wu M. Antioxidant and neuroprotective actions of resveratrol in cerebrovascular diseases. Front Pharmacol. 2022;13:948889. doi:10.3389/fphar.2022.948889
149. Li S. Novel insight into functions of ascorbate peroxidase in higher plants: more than a simple antioxidant enzyme. Redox Biol. 2023;64:102789. doi:10.1016/j.redox.2023.102789
150. Zelko IN, Mariani TJ, Folz RJ. Superoxide dismutase multigene family: a comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic Biol Med. 2002;33(3):337–349. doi:10.1016/S0891-5849(02)00905-X
151. Xiang Q, Zhao Y, Lin J, Jiang S, Li W. The Nrf2 antioxidant defense system in intervertebral disc degeneration: molecular insights. Exp Mol Med. 2022;54(8):1067–1075. doi:10.1038/s12276-022-00829-6
152. Xiao L, Xu SJ, Liu C, Wang J, Hu B, Xu HG. Sod2 and catalase improve pathological conditions of intervertebral disc degeneration by modifying human adipose-derived mesenchymal stem cells. Life Sci. 2021;267:118929. doi:10.1016/j.lfs.2020.118929
153. Hou G, Lu H, Chen M, Yao H, Zhao H. Oxidative stress participates in age-related changes in rat lumbar intervertebral discs. Arch Gerontol Geriatrics. 2014;59(3):665–669. doi:10.1016/j.archger.2014.07.002
154. Zhang Q, Li J, Li Y, et al. Bmi deficiency causes oxidative stress and intervertebral disc degeneration which can be alleviated by antioxidant treatment. J Cell & Mol Med. 2020;24(16):8950–8961. doi:10.1111/jcmm.15528
155. Tamagawa S, Sakai D, Nojiri H, et al. SOD2 orchestrates redox homeostasis in intervertebral discs: a novel insight into oxidative stress-mediated degeneration and therapeutic potential. Redox Biol. 2024;71:103091. doi:10.1016/j.redox.2024.103091
156. Chandran S, Binninger D. Role of Oxidative Stress, Methionine Oxidation and Methionine Sulfoxide Reductases (MSR) in Alzheimer’s Disease. Antioxidants (Basel, Switzerland). 2023;13(1). doi:10.3390/antiox13010021
157. Gruber HE, Watts JA, Hoelscher GL, et al. Mitochondrial gene expression in the human annulus: in vivo data from annulus cells and selectively harvested senescent annulus cells. Spine J. 2011;11(8):782–791. doi:10.1016/j.spinee.2011.06.012
158. Bastani NE, Sakhi AK, Karlsen A, et al. Reduced antioxidant defense and increased oxidative stress in spinal cord injured patients. Arch Phys Med Rehabil. 2012;93(12):2223–2228.e2222. doi:10.1016/j.apmr.2012.06.021
159. Liu S, Pi J, Zhang Q. Signal amplification in the KEAP1-NRF2-ARE antioxidant response pathway. Redox Biol. 2022;54:102389. doi:10.1016/j.redox.2022.102389
160. Wang L, Zhang X, Xiong X, et al. Nrf2 Regulates Oxidative Stress and Its Role in Cerebral Ischemic Stroke. Antioxidants (Basel, Switzerland). 2022;11(12):2.
161. Pan C, Hou W, Deng X, et al. The Pivotal Role of Nrf2 Signal Axis in Intervertebral Disc Degeneration. J Inflamm Res. 2023;16:5819–5833. doi:10.2147/JIR.S432575
162. Kang L, Liu S, Li J, Tian Y, Xue Y, Liu X. The mitochondria-targeted anti-oxidant MitoQ protects against intervertebral disc degeneration by ameliorating mitochondrial dysfunction and redox imbalance. Cell Proliferation. 2020;53(3):e12779. doi:10.1111/cpr.12779
163. Da W, Chen Q, Shen B. The current insights of mitochondrial hormesis in the occurrence and treatment of bone and cartilage degeneration. Biol Res. 2024;57(1):37. doi:10.1186/s40659-024-00494-1
164. Lin Z, Xu G, Lu X, et al. Chondrocyte-targeted exosome-mediated delivery of Nrf2 alleviates cartilaginous endplate degeneration by modulating mitochondrial fission. J Nanobiotechnol. 2024;22(1):281. doi:10.1186/s12951-024-02517-1
165. Foo J, Bellot G, Pervaiz S, Alonso S. Mitochondria-mediated oxidative stress during viral infection. Trends in Microbiology. 2022;30(7):679–692. doi:10.1016/j.tim.2021.12.011
166. Zheng D, Liu J, Piao H, Zhu Z, Wei R, Liu K. ROS-triggered endothelial cell death mechanisms: focus on pyroptosis, parthanatos, and ferroptosis. Front Immunol. 2022;13:1039241. doi:10.3389/fimmu.2022.1039241
167. Newton K, Strasser A, Kayagaki N, Dixit VM. Cell death. Cell. 2024;187(2):235–256. doi:10.1016/j.cell.2023.11.044
168. Czabotar PE, Garcia-Saez AJ. Mechanisms of BCL-2 family proteins in mitochondrial apoptosis. Nat Rev Mol Cell Biol. 2023;24(10):732–748. doi:10.1038/s41580-023-00629-4
169. Sahoo G, Samal D, Khandayataray P, Murthy MK. A Review on Caspases: key Regulators of Biological Activities and Apoptosis. Molecular Neurobiology. 2023;60(10):5805–5837. doi:10.1007/s12035-023-03433-5
170. Kumari S, Dhapola R, Reddy DH. Apoptosis in Alzheimer’s disease: insight into the signaling pathways and therapeutic avenues. Apoptosis: an International Journal on Programmed Cell Death. 2023;28(7–8):943–957. doi:10.1007/s10495-023-01848-y
171. Verdin E, Hirschey MD, Finley LW, Haigis MC. Sirtuin regulation of mitochondria: energy production, apoptosis, and signaling. Trends Biochem Sci. 2010;35(12):669–675. doi:10.1016/j.tibs.2010.07.003
172. Cantoni O, Zito E, Guidarelli A, Fiorani M, Ghezzi P, Mitochondrial ROS. ER Stress, and Nrf2 Crosstalk in the Regulation of Mitochondrial Apoptosis Induced by Arsenite. Antioxidants (Basel, Switzerland). 2022;11(5). doi:10.3390/antiox11051034
173. Turco MC, Romano MF, Petrella A, Bisogni R, Tassone P, Venuta S. NF-kappaB/Rel-mediated regulation of apoptosis in hematologic malignancies and normal hematopoietic progenitors. Leukemia. 2004;18(1):11–17. doi:10.1038/sj.leu.2403171
174. Huang ZN, Wang ZY, Cheng XF, et al. Melatonin alleviates oxidative stress-induced injury to nucleus pulposus-derived mesenchymal stem cells through activating PI3K/Akt pathway. Journal of Orthopaedic Translation. 2023;43:66–84. doi:10.1016/j.jot.2023.10.002
175. Zhang Z, Wang C, Lin J, et al. Therapeutic Potential of Naringin for Intervertebral Disc Degeneration: involvement of Autophagy Against Oxidative Stress-Induced Apoptosis in Nucleus Pulposus Cells. Am J Chin Med;2018:1–20. doi:10.1142/S0192415X18500805
176. Zhu P, Wu X, Ni L, et al. Inhibition of PP2A ameliorates intervertebral disc degeneration by reducing annulus fibrosus cells apoptosis via p38/MAPK signal pathway. Biochim Biophys Acta Mol Basis Dis. 2024;1870(1):166888. doi:10.1016/j.bbadis.2023.166888
177. Xiao H, Wang K, Peng L, Yin Z. Laquinimod attenuates oxidative stress-induced mitochondrial injury and alleviates intervertebral disc degeneration by inhibiting the NF-κB signaling pathway. Int immunopharmacol. 2024;131:111804. doi:10.1016/j.intimp.2024.111804
178. Chen J, Luo Y, Li Y, Chen D, Yu B, He J. Chlorogenic Acid Attenuates Oxidative Stress-Induced Intestinal Epithelium Injury by Co-Regulating the PI3K/Akt and IκBα/NF-κB Signaling. Antioxidants (Basel, Switzerland). 2021;10(12):1.
179. Guo Q, Jin Y, Chen X, et al. NF-κB in biology and targeted therapy: new insights and translational implications. Signal Transduction and Targeted Therapy. 2024;9(1):53. doi:10.1038/s41392-024-01757-9
180. Strasser A, O’Connor L, Dixit VM. Apoptosis signaling. Annu Rev Biochem. 2000;69(1):217–245. doi:10.1146/annurev.biochem.69.1.217
181. Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: mechanisms and diseases. Signal Transduction and Targeted Therapy. 2021;6(1):128. doi:10.1038/s41392-021-00507-5
182. Sharma BR, Kanneganti TD. NLRP3 inflammasome in cancer and metabolic diseases. Nat Immunol. 2021;22(5):550–559. doi:10.1038/s41590-021-00886-5
183. Huang Y, Xu W, Zhou R. NLRP3 inflammasome activation and cell death. Cell Mol Immunol. 2021;18(9):2114–2127. doi:10.1038/s41423-021-00740-6
184. Burdette BE, Esparza AN, Zhu H, Wang S. Gasdermin D in pyroptosis. Acta Pharmaceutica Sinica B. 2021;11(9):2768–2782. doi:10.1016/j.apsb.2021.02.006
185. Bai Z, Liu W, He D, et al. Protective effects of autophagy and NFE2L2 on reactive oxygen species-induced pyroptosis of human nucleus pulposus cells. Aging. 2020;12(8):7534–7548. doi:10.18632/aging.103109
186. Ma H, Xie C, Chen Z, et al. MFG-E8 alleviates intervertebral disc degeneration by suppressing pyroptosis and extracellular matrix degradation in nucleus pulposus cells via Nrf2/TXNIP/NLRP3 axis. Cell Death Discovery. 2022;8(1):209. doi:10.1038/s41420-022-01002-8
187. Chen Y, Luo X, Xu B, Bao X, Jia H, Yu B. Oxidative Stress-Mediated Programmed Cell Death: a Potential Therapy Target for Atherosclerosis. Cardiovasc Drugs Ther. 2024;38(4):819–832. doi:10.1007/s10557-022-07414-z
188. Ma Z, Tang P, Dong W, et al. SIRT1 alleviates IL-1β induced nucleus pulposus cells pyroptosis via mitophagy in intervertebral disc degeneration. Int immunopharmacol. 2022;107:108671. doi:10.1016/j.intimp.2022.108671
189. Tian K, Yang Y, Zhou K, et al. The role of ROS-induced pyroptosis in CVD. Frontiers in Cardiovascular Medicine. 2023;10:1116509. doi:10.3389/fcvm.2023.1116509
190. Zhao Y, Liu B, Xu L, et al. ROS-Induced mtDNA Release: the Emerging Messenger for Communication between Neurons and Innate Immune Cells during Neurodegenerative Disorder Progression. Antioxidants. 2021;10(12):1917.
191. Lu P, Zheng H, Liu C, Duan L, Sun T. Mitochondrial DNA induces nucleus pulposus cell pyroptosis via the TLR9-NF-κB-NLRP3 axis. J Transl Med. 2023;21(1):389. doi:10.1186/s12967-023-04266-5
192. Choi EH, Park SJ. TXNIP: a key protein in the cellular stress response pathway and a potential therapeutic target. Exp Mol Med. 2023;55(7):1348–1356. doi:10.1038/s12276-023-01019-8
193. Ma M, Zhang C, Zhong Zet al. siRNA incorporated in slow-release injectable hydrogel continuously silences DDIT4 and regulates nucleus pulposus cell pyroptosis through the ROS/TXNIP/NLRP3 axis to alleviate intervertebral disc degeneration. Bone & Joint Research. 2024;13(5):247–260. doi:10.1302/2046-3758.135.BJR-2023-0320.R1
194. Wu J, Han W, Zhang Y, et al. Glutamine Mitigates Oxidative Stress-Induced Matrix Degradation, Ferroptosis, and Pyroptosis in Nucleus Pulposus Cells via Deubiquitinating and Stabilizing Nrf2. Antioxid. Redox Signaling. 2024;41(4–6):278–295. doi:10.1089/ars.2023.0384
195. Li T, Yang J, Tan A, Chen H. Irisin suppresses pancreatic β cell pyroptosis in T2DM by inhibiting the NLRP3-GSDMD pathway and activating the Nrf2-TrX/TXNIP signaling axis. Diabetol Metab Syndr. 2023;15(1):239. doi:10.1186/s13098-023-01216-5
196. Li Z, Deng H, Guo X, et al. Effective dose/duration of natural flavonoid quercetin for treatment of diabetic nephropathy: a systematic review and meta-analysis of rodent data. Phytomedicine: International Journal of Phytotherapy and Phytopharmacology. 2022;105:154348. doi:10.1016/j.phymed.2022.154348
197. Jin X, Fu W, Zhou J, Shuai N, Yang Y, Wang B. Oxymatrine attenuates oxidized low‑density lipoprotein‑induced HUVEC injury by inhibiting NLRP3 inflammasome‑mediated pyroptosis via the activation of the SIRT1/Nrf2 signaling pathway. IntJ Mol Med. 2021;48(4). doi:10.3892/ijmm.2021.5020
198. Lei G, Zhuang L, Gan B. Targeting ferroptosis as a vulnerability in cancer. Nat Rev Cancer. 2022;22(7):381–396. doi:10.1038/s41568-022-00459-0
199. Hu Y, He B, Cao Q, et al. Crosstalk of ferroptosis and oxidative stress in infectious diseases. Front Mol Biosci. 2023;10:1315935. doi:10.3389/fmolb.2023.1315935
200. Galaris D, Barbouti A, Pantopoulos K. Iron homeostasis and oxidative stress: an intimate relationship. Biochim Biophys Acta Mol Cell Res. 2019;1866(12):118535. doi:10.1016/j.bbamcr.2019.118535
201. Yang Y, Wu Q, Shan X, et al. Ginkgolide B attenuates cerebral ischemia-reperfusion injury via inhibition of ferroptosis through disrupting NCOA4-FTH1 interaction. J Ethnopharmacol. 2024;318(Pt B):116982. doi:10.1016/j.jep.2023.116982
202. Liu Y, Jiang Y, Zhang L, Cheng W. GPX4: the hub of lipid oxidation, ferroptosis, disease and treatment. Biochim. Biophys. Acta, Rev. Cancer. 2023;1878(3):188890. doi:10.1016/j.bbcan.2023.188890
203. Rudeck M, Volk T, Sitte N, Grune T. Ferritin oxidation in vitro: implication of iron release and degradation by the 20S proteasome. IUBMB Life. 2000;49(5):451–456. doi:10.1080/152165400410317
204. Yang RZ, Xu WN, Zheng HL, et al. Involvement of oxidative stress-induced annulus fibrosus cell and nucleus pulposus cell ferroptosis in intervertebral disc degeneration pathogenesis. J Cell Physiol. 2021;236(4):2725–2739. doi:10.1002/jcp.30039
205. Yang X, Chen Y, Guo J, et al. Polydopamine Nanoparticles Targeting Ferroptosis Mitigate Intervertebral Disc Degeneration Via Reactive Oxygen Species Depletion, Iron Ions Chelation, and GPX4 Ubiquitination Suppression. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2023;10(13):e2207216. doi:10.1002/advs.202207216
206. Wang W, Jing X, Du T, et al. Iron overload promotes intervertebral disc degeneration via inducing oxidative stress and ferroptosis in endplate chondrocytes. Free Radic Biol Med. 2022;190:234–246. doi:10.1016/j.freeradbiomed.2022.08.018
207. Ganz T. Systemic iron homeostasis. Physiol Rev. 2013;93(4):1721–1741. doi:10.1152/physrev.00008.2013
208. Lu S, Song Y, Luo R, et al. Ferroportin-Dependent Iron Homeostasis Protects against Oxidative Stress-Induced Nucleus Pulposus Cell Ferroptosis and Ameliorates Intervertebral Disc Degeneration In Vivo. Oxid Med Cell Longev. 2021;2021:6670497. doi:10.1155/2021/6670497
209. Wang H, Liu X, Yang H, et al. Activation of the Nrf-2 pathway by pinocembrin safeguards vertebral endplate chondrocytes against apoptosis and degeneration caused by oxidative stress. Life Sci. 2023;333:122162. doi:10.1016/j.lfs.2023.122162
210. Zhu J, Sun R, Yan C, et al. Hesperidin mitigates oxidative stress-induced ferroptosis in nucleus pulposus cells via Nrf2/NF-κB axis to protect intervertebral disc from degeneration. Cell Cycle (Georgetown, Tex). 2023;22(10):1196–1214. doi:10.1080/15384101.2023.2200291
211. Li C, Zhang Y, Deng Y, et al. Fisetin suppresses ferroptosis through Nrf2 and attenuates intervertebral disc degeneration in rats. Eur J Pharmacol. 2024;964:176298. doi:10.1016/j.ejphar.2023.176298
212. Zhu J, Sun R, Sun K, et al. The deubiquitinase USP11 ameliorates intervertebral disc degeneration by regulating oxidative stress-induced ferroptosis via deubiquitinating and stabilizing Sirt3. Redox Biol. 2023;62:102707. doi:10.1016/j.redox.2023.102707
213. Di Micco R, Krizhanovsky V, Baker D, d’Adda Di Fagagna F. d’Adda di Fagagna F: cellular senescence in ageing: from mechanisms to therapeutic opportunities. Nat Rev Mol Cell Biol. 2021;22(2):75–95. doi:10.1038/s41580-020-00314-w
214. Kim KW, Chung HN, Ha KY, Lee JS, Kim YY. Senescence mechanisms of nucleus pulposus chondrocytes in human intervertebral discs. Spine J. 2009;9(8):658–666. doi:10.1016/j.spinee.2009.04.018
215. Wang F, Shi R, Wang XH, Wu XT, Wu X-T. Aging and age related stresses: a senescence mechanism of intervertebral disc degeneration. Osteoarthritis Cartilage. 2016;24(3):398–408. doi:10.1016/j.joca.2015.09.019
216. Beck J, Turnquist C, Horikawa I, Harris C. Targeting cellular senescence in cancer and aging: roles of p53 and its isoforms. Carcinogenesis. 2020;41(8):1017–1029. doi:10.1093/carcin/bgaa071
217. Bhattacharyya S, Dudeja PK, Tobacman JK. ROS, Hsp27, and IKKbeta mediate dextran sodium sulfate (DSS) activation of IkappaBa, NFkappaB, and IL-8. Inflammatory Bowel Diseases. 2009;15(5):673–683. doi:10.1002/ibd.20821
218. Li P, Gan Y, Wang Let al. 17beta-estradiol Attenuates TNF-α-Induced Premature Senescence of Nucleus Pulposus Cells through Regulating the ROS/NF-κB Pathway. Int J Bio Sci. 2017;13(2):145–156. doi:10.7150/ijbs.16770
219. Han Y, Zhou CM, Shen H, et al. Attenuation of ataxia telangiectasia mutated signalling mitigates age-associated intervertebral disc degeneration. Aging Cell. 2020;19(7):e13162. doi:10.1111/acel.13162
220. Fruman DA, Chiu H, Hopkins BD, Bagrodia S, Cantley LC, Abraham RT. The PI3K Pathway in Human Disease. Cell. 2017;170(4):605–635. doi:10.1016/j.cell.2017.07.029
221. Zhang Y, Liu L, Qi Y, et al. Lactic acid promotes nucleus pulposus cell senescence and corresponding intervertebral disc degeneration via interacting with Akt. Cellular and Molecular Life Sciences: CMLS. 2024;81(1):24. doi:10.1007/s00018-023-05094-y
222. Yao C, Guo G, Huang R, et al. Manual therapy regulates oxidative stress in aging rat lumbar intervertebral discs through the SIRT1/FOXO1 pathway. Aging. 2022;14(5):2400–2417. doi:10.18632/aging.203949
223. Song Y, Wang Z, Liu L, Qian Y, Zhang H, Qian Y. 1,4-Dihydropyridine (DHP) suppresses against oxidative stress in nucleus pulposus via activating sirtuin-1. Biomed Pharmacother. 2020;121:109592. doi:10.1016/j.biopha.2019.109592
224. Wang J, Nisar M, Huang C, et al. Small molecule natural compound agonist of SIRT3 as a therapeutic target for the treatment of intervertebral disc degeneration. Exp Mol Med. 2018;50(11):1–14.
225. Shen J, Lan Y, Ji Z, Liu H. Sirtuins in intervertebral disc degeneration: current understanding. Molecular Medicine (Cambridge, Mass). 2024;30(1):44. doi:10.1186/s10020-024-00811-0
226. Yang M, Peng Y, Liu W, Zhou M, Meng Q, Yuan C. Sirtuin 2 expression suppresses oxidative stress and senescence of nucleus pulposus cells through inhibition of the p53/p21 pathway. Biochem Biophys Res Commun. 2019;513(3):616–622. doi:10.1016/j.bbrc.2019.03.200
227. Debnath J, Gammoh N, Ryan KM. Autophagy and autophagy-related pathways in cancer. Nat Rev Mol Cell Biol. 2023;24(8):560–575. doi:10.1038/s41580-023-00585-z
228. Gruber HE, Hoelscher GL, Ingram JA, Bethea S, Hanley Jr EN. Autophagy in the Degenerating Human Intervertebral Disc: in Vivo Molecular and Morphological Evidence, and Induction of Autophagy in Cultured Annulus Cells Exposed to Proinflammatory Cytokines-Implications for Disc Degeneration. Spine. 2015;40(11):773–782. doi:10.1097/BRS.0000000000000865
229. Ye W, Xu K, Huang D, et al. Age-related increases of macroautophagy and chaperone-mediated autophagy in rat nucleus pulposus. Connective Tissue Res. 2011;52(6):472–478. doi:10.3109/03008207.2011.564336
230. Filomeni G, De Zio D, Cecconi F. Oxidative stress and autophagy: the clash between damage and metabolic needs. Cell Death Differ. 2015;22(3):377–388. doi:10.1038/cdd.2014.150
231. Gu Y, Han J, Jiang C, Zhang Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res Rev. 2020;59:101036. doi:10.1016/j.arr.2020.101036
232. Xu K, He Y, Moqbel SAA, Zhou X, Wu L, Bao J. SIRT3 ameliorates osteoarthritis via regulating chondrocyte autophagy and apoptosis through the PI3K/Akt/mTOR pathway. Int J Biol Macromol. 2021;175:351–360. doi:10.1016/j.ijbiomac.2021.02.029
233. Li Y, Mei J, Wu W, et al. High fat diet-induced obesity leads to depressive and anxiety-like behaviors in mice via AMPK/mTOR-mediated autophagy. Exp Neurol. 2022;348:113949. doi:10.1016/j.expneurol.2021.113949
234. Zhang Q, Wang X, Cao S, et al. Berberine represses human gastric cancer cell growth in vitro and in vivo by inducing cytostatic autophagy via inhibition of MAPK/mTOR/p70S6K and Akt signaling pathways. Biomed Pharmacother. 2020;128:110245. doi:10.1016/j.biopha.2020.110245
235. Li Z, Wang J, Deng X, Huang D, Shao Z, Ma K. Compression stress induces nucleus pulposus cell autophagy by inhibition of the PI3K/AKT/mTOR pathway and activation of the JNK pathway. Connective Tissue Res. 2021;62(3):337–349. doi:10.1080/03008207.2020.1736578
236. Zhang S, Liang W, Abulizi Y, et al. Quercetin Alleviates Intervertebral Disc Degeneration by Modulating p38 MAPK-Mediated Autophagy. Biomed Res. Int. 2021;2021(1):6631562. doi:10.1155/2021/6631562
237. Xie C, Shi Y, Chen Z, et al. Apigenin Alleviates Intervertebral Disc Degeneration via Restoring Autophagy Flux in Nucleus Pulposus Cells. Front Cell Develop Biol. 2021;9:787278. doi:10.3389/fcell.2021.787278
238. Chen K, Lv X, Li W, Yu F, Lin J, Xiao D. Autophagy Is a Protective Response to the Oxidative Damage to Endplate Chondrocytes in Intervertebral Disc: implications for the Treatment of Degenerative Lumbar Disc. Oxid Med Cell Longev. 2017;2017(1):4041768. doi:10.1155/2017/4041768
239. Tong T, Liu Z, Zhang H, et al. Age-dependent expression of the vitamin D receptor and the protective effect of vitamin D receptor activation on H(2)O(2)-induced apoptosis in rat intervertebral disc cells. J Steroid Biochem Mol Biol. 2019;190:126–138. doi:10.1016/j.jsbmb.2019.03.013
240. Picca A, Faitg J, Ferrucci L, D’Amico D, D’Amico D. Mitophagy in human health, ageing and disease. Nat Metab. 2023;5(12):2047–2061. doi:10.1038/s42255-023-00930-8
241. Xu WN, Yang RZ, Zheng HL, Yu W, Zheng XF, Li B. PGC-1α acts as an mediator of Sirtuin2 to protect annulus fibrosus from apoptosis induced by oxidative stress through restraining mitophagy. Int J Biol Macromol. 2019;136:1007–1017. doi:10.1016/j.ijbiomac.2019.06.163
242. Hu S, Zhang C, Qian T, Bai Y, Wang X, Jin H. Promoting Nrf2/Sirt3-Dependent Mitophagy Suppresses Apoptosis in Nucleus Pulposus Cells and Protects against Intervertebral Disc Degeneration. Oxid Med Cell Longev. 2021;2021(1):6694964. doi:10.1155/2021/6694964
243. Zhao Y, Simon M, Seluanov A, Gorbunova V. DNA damage and repair in age-related inflammation. Nat Rev Immunol. 2023;23(2):75–89. doi:10.1038/s41577-022-00751-y
244. Ansari MY, Ahmad N, Haqqi TM. Oxidative stress and inflammation in osteoarthritis pathogenesis: role of polyphenols. Biomed Pharmacother. 2020;129:110452. doi:10.1016/j.biopha.2020.110452
245. Cheng X, Lin J, Chen Z, et al. CB2-mediated attenuation of nucleus pulposus degeneration via the amelioration of inflammation and oxidative stress in vivo and in vitro. Molecular Medicine (Cambridge, Mass). 2021;27(1):92. doi:10.1186/s10020-021-00351-x
246. Lin Q, Li S, Jiang N, et al. PINK1-parkin pathway of mitophagy protects against contrast-induced acute kidney injury via decreasing mitochondrial ROS and NLRP3 inflammasome activation. Redox Biol. 2019;26:101254. doi:10.1016/j.redox.2019.101254
247. Wan L, Bai X, Zhou Q, et al. The advanced glycation end-products (AGEs)/ROS/NLRP3 inflammasome axis contributes to delayed diabetic corneal wound healing and nerve regeneration. Int J Bio Sci. 2022;18(2):809–825. doi:10.7150/ijbs.63219
248. Xu G, Lu X, Liu S, et al. MSC-Derived Exosomes Ameliorate Intervertebral Disc Degeneration By Regulating the Keap1/Nrf2 Axis. Stem Cell Rev Rep. 2023;19(7):2465–2480. doi:10.1007/s12015-023-10570-w
249. Wang H, Jiang Z, Pang Z, Zhou T, Gu Y. Acacetin Alleviates Inflammation and Matrix Degradation in Nucleus Pulposus Cells and Ameliorates Intervertebral Disc Degeneration in vivo. Drug Des Devel Ther. 2020;14:4801–4813. doi:10.2147/DDDT.S274812
250. Xie T, Gu X, Pan R, Huang W, Dong S. Evodiamine ameliorates intervertebral disc degeneration through the Nrf2 and MAPK pathways. Cytotechnology. 2024;76(2):153–166. doi:10.1007/s10616-023-00605-y
251. Lu G, Zhang C, Li K, Fu M, Lyu C, Quan Z. Sinomenine Ameliorates IL-1β-Induced Intervertebral Disc Degeneration in Rats Through Suppressing Inflammation and Oxidative Stress via Keap1/Nrf2/NF-κB Signaling Pathways. J Inflamm Res. 2023;16:4777–4791. doi:10.2147/JIR.S430423
252. Zhao K, An R, Xiang Q, et al. Acid-sensing ion channels regulate nucleus pulposus cell inflammation and pyroptosis via the NLRP3 inflammasome in intervertebral disc degeneration. Cell Proliferation. 2021;54(1):e12941. doi:10.1111/cpr.12941
253. Yu H, Zhang Z, Wei F, et al. Hydroxytyrosol Ameliorates Intervertebral Disc Degeneration and Neuropathic Pain by Reducing Oxidative Stress and Inflammation. Oxid Med Cell Longev. 2022;2022(1):2240894. doi:10.1155/2022/2240894
254. Chen D, Jiang X, Zou H. hASCs-derived exosomal miR-155-5p targeting TGFβR2 promotes autophagy and reduces pyroptosis to alleviate intervertebral disc degeneration. Journal of Orthopaedic Translation. 2023;39:163–176. doi:10.1016/j.jot.2023.02.004
255. Cao G, Yang S, Cao J, et al. The Role of Oxidative Stress in Intervertebral Disc Degeneration. Oxid Med Cell Longev. 2022;2022(1):2166817. doi:10.1155/2022/2166817
256. Zhang HJ, Liao HY, Bai DY, Wang ZQ, Xie XW. MAPK /ERK signaling pathway: a potential target for the treatment of intervertebral disc degeneration. Biomed Pharmacother. 2021;143:112170. doi:10.1016/j.biopha.2021.112170
257. Cannizzo ES, Clement CC, Morozova K, et al. Age-related oxidative stress compromises endosomal proteostasis. Cell Rep. 2012;2(1):136–149. doi:10.1016/j.celrep.2012.06.005
258. Wu Z, Zhang W, Qu J, Liu GH. Emerging epigenetic insights into aging mechanisms and interventions. Trends Pharmacol Sci. 2024;45(2):157–172. doi:10.1016/j.tips.2023.12.002
259. Gu M, Ren B, Fang Y, et al. Epigenetic regulation in cancer. MedComm. 2024;5(2):e495. doi:10.1002/mco2.495
260. Hogg SJ, Beavis PA, Dawson MA, Johnstone RW. Targeting the epigenetic regulation of antitumour immunity. Nat Rev Drug Discov. 2020;19(11):776–800. doi:10.1038/s41573-020-0077-5
261. Younesian S, Yousefi AM, Ghaffari SH, Bashash D, Bashash D. The DNA Methylation in Neurological Diseases. Cells. 2022;11(21). doi:10.3390/cells11213439
262. Yousefi PD, Suderman M, Langdon R, Whitehurst O, Davey Smith G, Relton CL. DNA methylation-based predictors of health: applications and statistical considerations. Nat Rev Genet. 2022;23(6):369–383. doi:10.1038/s41576-022-00465-w
263. Mattei AL, Bailly N, Meissner A. DNA methylation: a historical perspective. Trends in Genetics: TIG. 2022;38(7):676–707. doi:10.1016/j.tig.2022.03.010
264. Guillaumet-Adkins A, Yañez Y, Peris-Diaz MD, Calabria I, Palanca-Ballester C, Sandoval J. Epigenetics and Oxidative Stress in Aging. Oxid Med Cell Longev. 2017;2017(1):9175806. doi:10.1155/2017/9175806
265. Sundar V, Ramasamy T, Doke M, Samikkannu T. Psychostimulants influence oxidative stress and redox signatures: the role of DNA methylation. Redox Report: Communications in Free Radical Research. 2022;27(1):53–59. doi:10.1080/13510002.2022.2043224
266. Cyr AR, Hitchler MJ, Domann FE. Regulation of SOD2 in cancer by histone modifications and CpG methylation: closing the loop between redox biology and epigenetics. Antioxid. Redox Signaling. 2013;18(15):1946–1955. doi:10.1089/ars.2012.4850
267. Chen X, Zhang R, Yu C, et al. Identification of HPCAL1 as a specific autophagy receptor involved in ferroptosis. Autophagy. 2023;19(1):54–74. doi:10.1080/15548627.2022.2059170
268. Nemeth E, Ganz T. Hepcidin-Ferroportin Interaction Controls Systemic Iron Homeostasis. Int J Mol Sci. 2021;22(12):6493. doi:10.3390/ijms22126493
269. Chen J, Yang X, Li Q, et al. Inhibiting DNA methyltransferase DNMT3B confers protection against ferroptosis in nucleus pulposus and ameliorates intervertebral disc degeneration via upregulating SLC40A1. Free Radic Biol Med. 2024;220:139–153. doi:10.1016/j.freeradbiomed.2024.05.007
270. Cheng P, Wei HZ, Chen HW, Wang ZQ, Mao P, Zhang HH. DNMT3a-mediated methylation of PPARγ promote intervertebral disc degeneration by regulating the NF-κB pathway. J Cell & Mol Med. 2024;28(2):e18048. doi:10.1111/jcmm.18048
271. Nagaraju GP, Dariya B, Kasa P, Peela S, El-Rayes BF. Epigenetics in hepatocellular carcinoma. Semi Cancer Biol. 2022;86(Pt 3):622–632. doi:10.1016/j.semcancer.2021.07.017
272. Bajbouj K, Al-Ali A, Ramakrishnan RK, Saber-Ayad M, Hamid Q. Histone Modification in NSCLC: molecular Mechanisms and Therapeutic Targets. Int J Mol Sci. 2021;22(21):11701. doi:10.3390/ijms222111701
273. Ito F, Yamada Y, Shigemitsu A, et al. Role of Oxidative Stress in Epigenetic Modification in Endometriosis. Reproductive Sciences (Thousand Oaks, Calif). 2017;24(11):1493–1502. doi:10.1177/1933719117704909
274. Zheng X, Sawalha AH. The Role of Oxidative Stress in Epigenetic Changes Underlying Autoimmunity. Antioxid. Redox Signaling. 2022;36(7–9):423–440. doi:10.1089/ars.2021.0066
275. Chen Q, Yang B, Liu X, Zhang XD, Zhang L, Liu T. Histone acetyltransferases CBP/p300 in tumorigenesis and CBP/p300 inhibitors as promising novel anticancer agents. Theranostics. 2022;12(11):4935–4948. doi:10.7150/thno.73223
276. Haque ME, Jakaria M, Akther M, Cho DY, Kim IS, Choi DK. The GCN5: its biological functions and therapeutic potentials. Clinical Science (London, England: 1979). 2021;135(1):231–257. doi:10.1042/CS20200986
277. Mir US, Bhat A, Mushtaq A, Pandita S, Altaf M, Pandita TK. Role of histone acetyltransferases MOF and Tip60 in genome stability. DNA Repair. 2021;107:103205. doi:10.1016/j.dnarep.2021.103205
278. Vasudevarao MD, Mizar P, Kumari S, et al. Naphthoquinone-mediated inhibition of lysine acetyltransferase KAT3B/p300, basis for non-toxic inhibitor synthesis. J Biol Chem. 2014;289(11):7702–7717. doi:10.1074/jbc.M113.486522
279. Hou H, Yu H. Structural insights into histone lysine demethylation. Curr Opin Struct Biol. 2010;20(6):739–748. doi:10.1016/j.sbi.2010.09.006
280. Yang J, Chen S, Yang Y, et al. Jumonji domain-containing protein 6 protein and its role in cancer. Cell Proliferation. 2020;53(2):e12747. doi:10.1111/cpr.12747
281. Niu Y, DesMarais TL, Tong Z, Yao Y, Costa M. Oxidative stress alters global histone modification and DNA methylation. Free Radic Biol Med. 2015;82:22–28. doi:10.1016/j.freeradbiomed.2015.01.028
282. Polytarchou C, Pfau R, Hatziapostolou M, Tsichlis PN. The JmjC domain histone demethylase Ndy1 regulates redox homeostasis and protects cells from oxidative stress. Mol Cell Biol. 2008;28(24):7451–7464. doi:10.1128/MCB.00688-08
283. García-Giménez JL, Garcés C, Romá-Mateo C, Pallardó FV. Oxidative stress-mediated alterations in histone post-translational modifications. Free Radic Biol Med. 2021;170:6–18. doi:10.1016/j.freeradbiomed.2021.02.027
284. Liu M, Zhang Y, Yang J, et al. ZIP4 Increases Expression of Transcription Factor ZEB1 to Promote Integrin α3β1 Signaling and Inhibit Expression of the Gemcitabine Transporter ENT1 in Pancreatic Cancer Cells. Gastroenterology. 2020;158(3):679–692.e671. doi:10.1053/j.gastro.2019.10.038
285. Shen M, Li K, Wang L, et al. ZIP4 upregulation aggravates nucleus pulposus cell degradation by promoting inflammation and oxidative stress by mediating the HDAC4-FoxO3a axis. Aging. 2024;16(1):685–700. doi:10.18632/aging.205412
286. Winkle M, El-Daly SM, Fabbri M, Calin GA. Noncoding RNA therapeutics - challenges and potential solutions. Nat Rev Drug Discov. 2021;20(8):629–651. doi:10.1038/s41573-021-00219-z
287. Guo HY, Guo MK, Wan ZY, Song F, Wang H-Q. Wang HQ: emerging evidence on noncoding-RNA regulatory machinery in intervertebral disc degeneration: a narrative review. Arthritis Res Therapy. 2020;22(1):270. doi:10.1186/s13075-020-02353-2
288. Li H, Tian L, Li J, et al. The Roles of circRNAs in Intervertebral Disc Degeneration: inflammation, Extracellular Matrix Metabolism, and Apoptosis. Analytical Cellular Pathology (Amsterdam). 2022;2022:9550499. doi:10.1155/2022/9550499
289. Li G, Ma L, He S, et al. WTAP-mediated m(6)A modification of lncRNA NORAD promotes intervertebral disc degeneration. Nat Commun. 2022;13(1):1469. doi:10.1038/s41467-022-28990-6
290. Xu X, Shen L, Qu Y, et al. Experimental validation and comprehensive analysis of m6A methylation regulators in intervertebral disc degeneration subpopulation classification. Sci Rep. 2024;14(1):8417. doi:10.1038/s41598-024-58888-w
291. Chen Z, Song J, Xie L, et al. N6-methyladenosine hypomethylation of circGPATCH2L regulates DNA damage and apoptosis through TRIM28 in intervertebral disc degeneration. Cell Death Differ. 2023;30(8):1957–1972. doi:10.1038/s41418-023-01190-5
292. Colella F, Garcia JP, Sorbona M, et al. Drug delivery in intervertebral disc degeneration and osteoarthritis: selecting the optimal platform for the delivery of disease-modifying agents. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2020;328:985–999. doi:10.1016/j.jconrel.2020.08.041
293. Guo Z, Ye J, Cheng X, et al. Nanodrug Delivery Systems in Antitumor Immunotherapy. Biomater Res. 2024;28:0015. doi:10.34133/bmr.0015
294. Zhou D, Zhou F, Sheng S, Wei Y, Chen X, Su J. Intra-articular nanodrug delivery strategies for treating osteoarthritis. Drug Discovery Today. 2023;28(3):103482. doi:10.1016/j.drudis.2022.103482
295. Guterl CC, See EY, Blanquer SB, et al. Challenges and strategies in the repair of ruptured annulus fibrosus. Eur Cells Mater. 2013;25:1–21. doi:10.22203/eCM.v025a01
296. Krut Z, Pelled G, Gazit D, Gazit Z. Stem Cells and Exosomes: new Therapies for Intervertebral Disc Degeneration. Cells. 2021;10(9):2241. doi:10.3390/cells10092241
297. Fan D, Cao Y, Cao M, Wang Y, Cao Y, Gong T. Nanomedicine in cancer therapy. Signal Transduction and Targeted Therapy. 2023;8(1):293.
298. Cheng J, Huang H, Chen Y, Wu R. Nanomedicine for Diagnosis and Treatment of Atherosclerosis. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2023;10(36):e2304294. doi:10.1002/advs.202304294
299. Hu Y, Yang R, Liu S, Song Z, Wang H. The Emerging Roles of Nanocarrier Drug Delivery System in Treatment of Intervertebral Disc Degeneration-Current Knowledge, Hot Spots, Challenges and Future Perspectives. Drug Des Devel Ther. 2024;18:1007–1022. doi:10.2147/DDDT.S448807
300. Qiu Z, Xu T, Wang L, Meng N, Jin H, Xu B. Novel Nano-Drug Delivery System for Brain Tumor Treatment. Cells. 2022;11(23):3761. doi:10.3390/cells11233761
301. Jiang Y, Li W, Wang Z, Lu J. Lipid-Based Nanotechnology: liposome. Pharmaceutics. 2023;16(1):34. doi:10.3390/pharmaceutics16010034
302. Beach MA, Nayanathara U, Gao Y, et al. Polymeric Nanoparticles for Drug Delivery. Chem Rev. 2024;124(9):5505–5616. doi:10.1021/acs.chemrev.3c00705
303. Floyd TG, Gurnani P, Rho JY. Characterisation of polymeric nanoparticles for drug delivery. Nanoscale. 2025;17(13):7738–7752. doi:10.1039/D5NR00071H
304. Luther DC, Huang R, Jeon T, et al. Delivery of drugs, proteins, and nucleic acids using inorganic nanoparticles. Adv. Drug Delivery Rev. 2020;156:188–213.
305. Ghezzi M, Pescina S, Padula C, et al. Polymeric micelles in drug delivery: an insight of the techniques for their characterization and assessment in biorelevant conditions. J Control Release. 2021;332:312–336. doi:10.1016/j.jconrel.2021.02.031
306. Perumal S, Atchudan R, Lee W. A Review of Polymeric Micelles and Their Applications. Polymers. 2022;14(12):2510. doi:10.3390/polym14122510
307. Hiwrale A, Bharati S, Pingale P, Rajput A. Nanofibers: a current era in drug delivery system. Heliyon. 2023;9(9):e18917. doi:10.1016/j.heliyon.2023.e18917
308. Zhou S, Zhang M, Wang J, Chen X, Xu Z, Yan Y. Nanofibers in Glioma Therapy: advances, Applications, and Overcoming Challenges. Int J Nanomed. 2025;20:4677–4703. doi:10.2147/IJN.S510363
309. Quazi MZ, Park N. Nanohydrogels: advanced Polymeric Nanomaterials in the Era of Nanotechnology for Robust Functionalization and Cumulative Applications. Int J Mol Sci. 2022;23(4):1943. doi:10.3390/ijms23041943
310. Liang Y, Duan L, Lu J, Xia J. Engineering exosomes for targeted drug delivery. Theranostics. 2021;11(7):3183–3195. doi:10.7150/thno.52570
311. Meng W, He C, Hao Y, Wang L, Zhu G, Zhu G. Prospects and challenges of extracellular vesicle-based drug delivery system: considering cell source. Drug Delivery. 2020;27(1):585–598. doi:10.1080/10717544.2020.1748758
312. Chen Q, Qian Q, Xu H, et al. Mitochondrial-Targeted Metal-Phenolic Nanoparticles to Attenuate Intervertebral Disc Degeneration: alleviating Oxidative Stress and Mitochondrial Dysfunction. ACS nano. 2024;18(12):8885–8905. doi:10.1021/acsnano.3c12163
313. Lim S, An SB, Jung M, et al. Local Delivery of Senolytic Drug Inhibits Intervertebral Disc Degeneration and Restores Intervertebral Disc Structure. Adv Healthcare Mater. 2022;11(2):e2101483. doi:10.1002/adhm.202101483
314. Wang J, Wu R, Liu Z, et al. Core-Shell Structured Nanozyme with PDA-Mediated Enhanced Antioxidant Efficiency to Treat Early Intervertebral Disc Degeneration. ACS Appl Mater Interfaces. 2024;16(4):5103–5119. doi:10.1021/acsami.3c15938
315. Yang W, Li K, Pan Q, et al. An Engineered Bionic Nanoparticle Sponge as a Cytokine Trap and Reactive Oxygen Species Scavenger to Relieve Disc Degeneration and Discogenic Pain. ACS nano. 2024;18(4):3053–3072. doi:10.1021/acsnano.3c08097
316. Zhou T, Yang X, Chen Z, et al. Prussian Blue Nanoparticles Stabilize SOD1 from Ubiquitination-Proteasome Degradation to Rescue Intervertebral Disc Degeneration. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2022;9(10):e2105466. doi:10.1002/advs.202105466
317. Zhou H, Qian Q, Chen Q, et al. Enhanced Mitochondrial Targeting and Inhibition of Pyroptosis with Multifunctional Metallopolyphenol Nanoparticles in Intervertebral Disc Degeneration. Small (Weinheim an der Bergstrasse, Germany). 2024;20(13):e2308167. doi:10.1002/smll.202308167
318. Zhou H, He J, Liu R, et al. Microenvironment-responsive metal-phenolic network release platform with ROS scavenging, anti-pyroptosis, and ECM regeneration for intervertebral disc degeneration. Bioact Mater. 2024;37:51–71. doi:10.1016/j.bioactmat.2024.02.036
319. Yang X, Jin L, Yao L, Shen FH, Shimer AL, Li X. Antioxidative nanofullerol prevents intervertebral disk degeneration. Int J Nanomed. 2014;9:2419–2430. doi:10.2147/IJN.S60853
320. Huang Y, Yang J, Liu X, et al. Cationic Polymer Brush-Modified Carbon Nanotube-Meditated eRNA LINC02569 Silencing Attenuates Nucleus Pulposus Degeneration by Blocking NF-κB Signaling Pathway and Alleviate Cell Senescence. Front Cell Develop Biol. 2021;9:837777.
321. Wu S, Shi Y, Jiang L, et al. N-Acetylcysteine-Derived Carbon Dots for Free Radical Scavenging in Intervertebral Disc Degeneration. Adv Healthcare Mater. 2023;12(24):e2300533.
322. Sun K, Yan C, Dai X, et al. Catalytic Nanodots-Driven Pyroptosis Suppression in Nucleus Pulposus for Antioxidant Intervention of Intervertebral Disc Degeneration. Advanced Materials (Deerfield Beach, Fla). 2024;36(19):e2313248. doi:10.1002/adma.202313248
323. Shi Y, Bu W, Chu D, et al. Rescuing Nucleus Pulposus Cells from ROS Toxic Microenvironment via Mitochondria-Targeted Carbon Dot-Supported Prussian Blue to Alleviate Intervertebral Disc Degeneration. Adv Healthcare Mater. 2024;13(8):e2303206. doi:10.1002/adhm.202303206
324. Yu C, Li D, Wang C, et al. Injectable kartogenin and apocynin loaded micelle enhances the alleviation of intervertebral disc degeneration by adipose-derived stem cell. Bioact Mater. 2021;6(10):3568–3579. doi:10.1016/j.bioactmat.2021.03.018
325. Sun J, Yang F, Wang L, et al. Delivery of coenzyme Q10 loaded micelle targets mitochondrial ROS and enhances efficiency of mesenchymal stem cell therapy in intervertebral disc degeneration. Bioact Mater. 2023;23:247–260. doi:10.1016/j.bioactmat.2022.10.019
326. Yu Q, Han F, Yuan Z, et al. Fucoidan-loaded nanofibrous scaffolds promote annulus fibrosus repair by ameliorating the inflammatory and oxidative microenvironments in degenerative intervertebral discs. Acta Biomater. 2022;148:73–89. doi:10.1016/j.actbio.2022.05.054
327. Wu R, Huang L, Xia Q, et al. Injectable mesoporous bioactive glass/sodium alginate hydrogel loaded with melatonin for intervertebral disc regeneration. Mater Today Bio. 2023;22:100731. doi:10.1016/j.mtbio.2023.100731
328. Li M, Wu Y, Li H, et al. Nanofiber reinforced alginate hydrogel for leak-proof delivery and higher stress loading in nucleus pulposus. Carbohydr Polym. 2023;299:120193. doi:10.1016/j.carbpol.2022.120193
329. Gan Y, Li S, Li P, et al. A Controlled Release Codelivery System of MSCs Encapsulated in Dextran/Gelatin Hydrogel with TGF- β 3-Loaded Nanoparticles for Nucleus Pulposus Regeneration. Stem Cells International. 2016;2016(1):9042019. doi:10.1155/2016/9042019
330. Zamboni F, Ren G, Culebras M, O’Driscoll J, Ryan EJ, Collins MN. Curcumin encapsulated polylactic acid nanoparticles embedded in alginate/gelatin bioinks for in situ immunoregulation: characterization and biological assessment. Int J Biol Macromol. 2022;221:1218–1227. doi:10.1016/j.ijbiomac.2022.09.014
331. Peng Y, Chen X, Rao Z, et al. Multifunctional annulus fibrosus matrix prevents disc-related pain via inhibiting neuroinflammation and sensitization. Acta Biomater. 2023;170:288–302. doi:10.1016/j.actbio.2023.08.028
332. Peng Y, Chen X, Zhang Q, et al. Enzymatically Bioactive Nucleus Pulposus Matrix Hydrogel Microspheres for Exogenous Stem Cells Therapy and Endogenous Repair Strategy to Achieve Disc Regeneration. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2024;11(10):e2304761. doi:10.1002/advs.202304761
333. Wang D, Zhang L, He D, et al. A natural hydrogel complex improves intervertebral disc degeneration by correcting fatty acid metabolism and inhibiting nucleus pulposus cell pyroptosis. Mater Today Bio. 2024;26:101081. doi:10.1016/j.mtbio.2024.101081
334. Qingxin S, Kai J, Dandan Z, et al. Programmable DNA hydrogel provides suitable microenvironment for enhancing autophagy-based therapies in intervertebral disc degeneration treatment. J Nanobiotechnol. 2023;21(1):350. doi:10.1186/s12951-023-02109-5
335. Xu B, Huang M, Li J, et al. The MnO(2)/GelMA Composite Hydrogels Improve the ROS Microenvironment of Annulus Fibrosus Cells by Promoting the Antioxidant and Autophagy through the SIRT1/NRF2 Pathway. Gels (Basel, Switzerland). 2024;10(5). doi:10.3390/gels10050333.
336. Li Z, Cai F, Tang J, et al. Oxygen metabolism-balanced engineered hydrogel microspheres promote the regeneration of the nucleus pulposus by inhibiting acid-sensitive complexes. Bioact. Mater. 2023;24:346–360.
337. Zhou L, Cai F, Zhu H, et al. Immune-defensive microspheres promote regeneration of the nucleus pulposus by targeted entrapment of the inflammatory cascade during intervertebral disc degeneration. Bioact Mater. 2024;37:132–152. doi:10.1016/j.bioactmat.2024.03.020
338. Liu Y, Du J, Peng P, et al. Regulation of the inflammatory cycle by a controllable release hydrogel for eliminating postoperative inflammation after discectomy. Bioact Mater. 2021;6(1):146–157. doi:10.1016/j.bioactmat.2020.07.008
339. Nair MB, Baranwal G, Vijayan P, Keyan KS, Jayakumar R. Composite hydrogel of chitosan-poly(hydroxybutyrate-co-valerate) with chondroitin sulfate nanoparticles for nucleus pulposus tissue engineering. Colloids Surf. B. 2015;136:84–92. doi:10.1016/j.colsurfb.2015.08.026
340. Yang L, Yu C, Fan X, et al. Dual-dynamic-bond cross-linked injectable hydrogel of multifunction for intervertebral disc degeneration therapy. J Nanobiotechnol. 2022;20(1):433. doi:10.1186/s12951-022-01633-0
341. Nezadi M, Keshvari H, Shokrolahi F, Shokrollahi P. Injectable, self-healing hydrogels based on gelatin, quaternized chitosan, and laponite as localized celecoxib delivery system for nucleus pulpous repair. Int J Biol Macromol. 2024;266(Pt 2):131337.
342. Zheng Q, Shen H, Tong Z, et al. A thermosensitive, reactive oxygen species-responsive, MR409-encapsulated hydrogel ameliorates disc degeneration in rats by inhibiting the secretory autophagy pathway. Theranostics. 2021;11(1):147–163. doi:10.7150/thno.47723
343. Yu H, Teng Y, Ge J, et al. Isoginkgetin-loaded reactive oxygen species scavenging nanoparticles ameliorate intervertebral disc degeneration via enhancing autophagy in nucleus pulposus cells. J Nanobiotechnol. 2023;21(1):99. doi:10.1186/s12951-023-01856-9
344. Bai J, Zhang Y, Fan Q, et al. Reactive Oxygen Species-Scavenging Scaffold with Rapamycin for Treatment of Intervertebral Disk Degeneration. Adv. Healthcare Mater. 2020;9(3):e1901186. doi:10.1002/adhm.201901186
345. Feng Y, Su L, Chen Z, et al. Accurate Spatio-Temporal Delivery of Nitric Oxide Facilitates the Programmable Repair of Avascular Dense Connective Tissues Injury. Adv Healthc Mater. 2024;13(14):e2303740. doi:10.1002/adhm.202303740
346. Wang W, Xiao B, Qiu Yet al. pH-Responsive Delivery of H2 through Ammonia Borane-Loaded Hollow Polydopamine for Intervertebral Disc Degeneration Therapy. Oxid Med Cell Longev. 2023;2023:7773609. doi:10.1155/2023/7773609
347. Liu S, Li K, He Y, et al. PGC1α-Inducing Senomorphic Nanotherapeutics Functionalized with NKG2D-Overexpressing Cell Membranes for Intervertebral Disc Degeneration. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2024;11(22):e2400749. doi:10.1002/advs.202400749
348. Zheng D, Chen W, Chen T, et al. Hydrogen Ion Capturing Hydrogel Microspheres for Reversing Inflammaging. Advanced Materials (Deerfield Beach, Fla). 2024;36(5):e2306105. doi:10.1002/adma.202306105
349. Feng G, Zha Z, Huang Y, et al.. Sustained and Bioresponsive Two-Stage Delivery of Therapeutic miRNA via Polyplex Micelle-Loaded Injectable Hydrogels for Inhibition of Intervertebral Disc Fibrosis. Adv Healthcare Mater. 2018;7(21):e1800623. doi:10.1002/adhm.201800623
350. Xia KS, Li DD, Wang CG, et al. An esterase-responsive ibuprofen nano-micelle pre-modified embryo derived nucleus pulposus progenitor cells promote the regeneration of intervertebral disc degeneration. Bioact Mater. 2023;21:69–85. doi:10.1016/j.bioactmat.2022.07.024
351. Zheng Z, Chen A, He H, et al. pH and enzyme dual-responsive release of hydrogen sulfide for disc degeneration therapy. J Mat Chem B. 2019;7(4):611–618. doi:10.1039/C8TB02566E
352. Zhu L, Yang Y, Yan Z, et al. Controlled Release of TGF-β3 for Effective Local Endogenous Repair in IDD Using Rat Model. Int J Nanomed. 2022;17:2079–2096. doi:10.2147/IJN.S358396
353. Wang D, Lu K, Zou G, Wu D, Cheng Y, Sun Y. Attenuating ntervertebral disc degeneration through spermidine-delivery nanoplatform based on polydopamine for persistent regulation of oxidative stress. Int J Biol Macromol. 2024;274(Pt 1):132881. doi:10.1016/j.ijbiomac.2024.132881
354. Wang X, Yu L, Duan J, et al. Anti-Stress and Anti-ROS Effects of MnOx-Functionalized Thermosensitive Nanohydrogel Protect BMSCs for Intervertebral Disc Degeneration Repair. Adv Healthcare Mater. 2024;13(29):e2400343. doi:10.1002/adhm.202400343
355. Shen J, Zhuo N, Xu S, Song Z, Hao J, Guo X. Resveratrol delivery by ultrasound-mediated nanobubbles targeting nucleus pulposus cells. Nanomedicine (London, England). 2018;13(12):1433–1446. doi:10.2217/nnm-2018-0019
356. Nguyen K, Pan HY, Haworth K, et al. Multiple-Exposure Drug Release from Stable Nanodroplets by High-Intensity Focused Ultrasound for a Potential Degenerative Disc Disease Treatment. Ultrasound Med Biol. 2019;45(1):160–169. doi:10.1016/j.ultrasmedbio.2018.09.014
357. Raza F, Evans L, Motallebi M, et al. Liposome-based diagnostic and therapeutic applications for pancreatic cancer. Acta Biomater. 2023;157:1–23. doi:10.1016/j.actbio.2022.12.013
358. Gao Y, Liu X, Chen N, Yang X, Tang F. Recent Advance of Liposome Nanoparticles for Nucleic Acid Therapy. Pharmaceutics. 2023;15(1):178. doi:10.3390/pharmaceutics15010178
359. Cheng R, Liu L, Xiang Y, et al. Advanced liposome-loaded scaffolds for therapeutic and tissue engineering applications. Biomaterials. 2020;232:119706. doi:10.1016/j.biomaterials.2019.119706
360. Chang H, Cai F, Zhang Y, et al. Silencing Gene-Engineered Injectable Hydrogel Microsphere for Regulation of Extracellular Matrix Metabolism Balance. Small Methods. 2022;6(4):e2101201. doi:10.1002/smtd.202101201
361. Banala RR, Vemuri SK, Dar GH, et al. Efficiency of dual siRNA-mediated gene therapy for intervertebral disc degeneration (IVDD). Spine J. 2019;19(5):896–904. doi:10.1016/j.spinee.2018.10.016
362. Liu X, Wang P, Zou YX, Luo ZG, Tamer TM. Co-encapsulation of Vitamin C and β-Carotene in liposomes: storage stability, antioxidant activity, and in vitro gastrointestinal digestion. Food Research International (Ottawa, Ont). 2020;136:109587. doi:10.1016/j.foodres.2020.109587
363. Stone WL, Mukherjee S, Smith M, Das SK. Therapeutic uses of antioxidant liposomes. Methods in Molecular Biology (Clifton, NJ). 2002;199:145–161.
364. Wang L, Wang L, Wang X, Lu B, Zhang J. Preparation of blueberry anthocyanin liposomes and changes of vesicle properties, physicochemical properties, in vitro release, and antioxidant activity before and after chitosan modification. Food Sci Nutr. 2022;10(1):75–87. doi:10.1002/fsn3.2649
365. Xu T, Zhang J, Jin R, Wang X, Yuan C. Physicochemical properties, antioxidant activities and in vitro sustained release behaviour of co-encapsulated liposomes as vehicle for vitamin E and β-carotene. J Sci Food Agric. 2022;102(13):5759–5767. doi:10.1002/jsfa.11925
366. Zhao G, Hu C, Xue Y. In vitro evaluation of chitosan-coated liposome containing both coenzyme Q10 and alpha-lipoic acid: cytotoxicity, antioxidant activity, and antimicrobial activity. J Cosmet Dermatol. 2018;17(2):258–262. doi:10.1111/jocd.12369
367. Chen J, Chen J, Yu P, et al. A novel quercetin encapsulated glucose modified liposome and its brain-target antioxidative neuroprotection effects. Molecules. 2024;29(3):607.
368. Zhang C, Hu Y, Yuan Y, et al. Liposome-embedded SOD attenuated DSS-induced ulcerative colitis in mice by ameliorating oxidative stress and intestinal barrier dysfunction. Food Funct. 2023;14(9):4392–4405. doi:10.1039/D2FO03312G
369. Tan C, Wang J, Sun B. Biopolymer-liposome hybrid systems for controlled delivery of bioactive compounds: recent advances. Biotechnol Adv. 2021;48:107727. doi:10.1016/j.biotechadv.2021.107727
370. Guimarães D, Cavaco-Paulo A, Nogueira E. Design of liposomes as drug delivery system for therapeutic applications. Int J Pharm. 2021;601:120571. doi:10.1016/j.ijpharm.2021.120571
371. Maritim S, Boulas P, Lin Y. Comprehensive analysis of liposome formulation parameters and their influence on encapsulation, stability and drug release in glibenclamide liposomes. Int J Pharm. 2021;592:120051. doi:10.1016/j.ijpharm.2020.120051
372. Sarfraz M, Afzal A, Yang T, et al. Development of Dual Drug Loaded Nanosized Liposomal Formulation by A Reengineered Ethanolic Injection Method and Its Pre-Clinical Pharmacokinetic Studies. Pharmaceutics. 2018;10(3):151. doi:10.3390/pharmaceutics10030151
373. Large DE, Abdelmessih RG, Fink EA, Auguste DT. Liposome composition in drug delivery design, synthesis, characterization, and clinical application. Adv Drug Delivery Rev. 2021;176:113851. doi:10.1016/j.addr.2021.113851
374. Wang C, Lan X, Zhu L, et al. Construction Strategy of Functionalized Liposomes and Multidimensional Application. Small (Weinheim an der Bergstrasse, Germany). 2024;20(25):e2309031. doi:10.1002/smll.202309031
375. Ding Q, Tang W, Li X, et al. Mitochondrial-targeted brequinar liposome boosted mitochondrial-related ferroptosis for promoting checkpoint blockade immunotherapy in bladder cancer. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2023;363:221–234. doi:10.1016/j.jconrel.2023.09.024
376. Liu J, Wu Z, Liu Y, et al. ROS-responsive liposomes as an inhaled drug delivery nanoplatform for idiopathic pulmonary fibrosis treatment via Nrf2 signaling. J Nanobiotechnol. 2022;20(1):213. doi:10.1186/s12951-022-01435-4
377. Li Z, Zhu H, Liu H, et al. Synergistic dual cell therapy for atherosclerosis regression: ROS-responsive Bio-liposomes co-loaded with Geniposide and Emodin. J Nanobiotechnol. 2024;22(1):129. doi:10.1186/s12951-024-02389-5
378. Zahednezhad F, Saadat M, Valizadeh H, Zakeri-Milani P, Baradaran B. Liposome and immune system interplay: challenges and potentials. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2019;305:194–209. doi:10.1016/j.jconrel.2019.05.030
379. Tenchov R, Curtze AE, Zhou Q, Zhou Q. Lipid Nanoparticles─From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS nano. 2021;15(11):16982–17015. doi:10.1021/acsnano.1c04996
380. Strachan JB, Dyett BP, Nasa Z, Valery C, Conn CE. Toxicity and cellular uptake of lipid nanoparticles of different structure and composition. J Colloid Interface Sci. 2020;576:241–251. doi:10.1016/j.jcis.2020.05.002
381. Lv H, Zhang S, Wang B, Cui S, Yan J. Toxicity of cationic lipids and cationic polymers in gene delivery. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2006;114(1):100–109. doi:10.1016/j.jconrel.2006.04.014
382. Knudsen KB, Northeved H, Kumar PE, et al. In vivo toxicity of cationic micelles and liposomes. Nanomedicine: Nanotechnology, Biology, and Medicine. 2015;11(2):467–477. doi:10.1016/j.nano.2014.08.004
383. Begines B, Ortiz T, Pérez-Aranda M, et al. Polymeric Nanoparticles for Drug Delivery: recent Developments and Future Prospects. Nanomaterials (Basel, Switzerland). 2020;10(7). doi:10.3390/nano10071403
384. El-Say KM, El-Sawy HS. Polymeric nanoparticles: promising platform for drug delivery. Int J Pharm. 2017;528(1–2):675–691. doi:10.1016/j.ijpharm.2017.06.052
385. Zhang M, Ma H, Wang X, Yu B, Cong H, Shen Y. Polysaccharide-based nanocarriers for efficient transvascular drug delivery. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2023;354:167–187. doi:10.1016/j.jconrel.2022.12.051
386. Barclay TG, Day CM, Petrovsky N, Garg S. Review of polysaccharide particle-based functional drug delivery. Carbohydr Polym. 2019;221:94–112. doi:10.1016/j.carbpol.2019.05.067
387. Yuan Q, Yuan Y, Zheng Y, et al. Anti-cerebral ischemia reperfusion injury of polysaccharides: a review of the mechanisms. Biomed Pharmacother. 2021;137:111303. doi:10.1016/j.biopha.2021.111303
388. Fang CY, Lou DY, Zhou LQ, et al. Natural products: potential treatments for cisplatin-induced nephrotoxicity. Acta Pharmacol Sin. 2021;42(12):1951–1969. doi:10.1038/s41401-021-00620-9
389. He B, Tao H, Liu S, Wei A. Protective effect of carboxymethylated chitosan on hydrogen peroxide-induced apoptosis in nucleus pulposus cells. Molecular Medicine Reports. 2015;11(3):1629–1638. doi:10.3892/mmr.2014.2942
390. Li Y, Tian X, He W, et al. Fucoidan-functionalized gelatin methacryloyl microspheres ameliorate intervertebral disc degeneration by restoring redox and matrix homeostasis of nucleus pulposus. Int J Biol Macromol. 2023;250:126166. doi:10.1016/j.ijbiomac.2023.126166
391. Yamamoto T, Suzuki S, Fujii T, et al. Efficacy of hyaluronic acid on intervertebral disc inflammation: an in vitro study using notochordal cell lines and human disc cells. J Orthopaedic Res. 2021;39(10):2197–2208. doi:10.1002/jor.24933
392. Vasvani S, Kulkarni P, Rawtani D. Hyaluronic acid: a review on its biology, aspects of drug delivery, route of administrations and a special emphasis on its approved marketed products and recent clinical studies. Int J Biol Macromol. 2020;151:1012–1029. doi:10.1016/j.ijbiomac.2019.11.066
393. Zhang F, Wang S, Gao M, et al. Hyaluronic acid ameliorates intervertebral disc degeneration via promoting mitophagy activation. Front Bioeng Biotechnol. 2022;10:1057429. doi:10.3389/fbioe.2022.1057429
394. Xu Y, Tang G, Zhang C, Wang N, Feng Y. Gallic Acid and Diabetes Mellitus: its Association with Oxidative Stress. Molecules (Basel, Switzerland). 2021;26(23):7115. doi:10.3390/molecules26237115
395. Rahimi M, Charmi G, Matyjaszewski K, Banquy X, Pietrasik J. Recent developments in natural and synthetic polymeric drug delivery systems used for the treatment of osteoarthritis. Acta Biomater. 2021;123:31–50. doi:10.1016/j.actbio.2021.01.003
396. Pontes AP, Welting TJM, Rip J, Creemers LB. Polymeric Nanoparticles for Drug Delivery in Osteoarthritis. Pharmaceutics. 2022;14(12):2639. doi:10.3390/pharmaceutics14122639
397. Gorth DJ, Mauck RL, Chiaro JA, et al. IL-1ra delivered from poly(lactic-co-glycolic acid) microspheres attenuates IL-1β-mediated degradation of nucleus pulposus in vitro. Arthritis Res Therapy. 2012;14(4):R179. doi:10.1186/ar3932
398. Cicardi M, Zuraw BL. Angioedema Due to Bradykinin Dysregulation. The Journal of allergy and Clinical Immunology in Practice. 2018;6(4):1132–1141. doi:10.1016/j.jaip.2018.04.022
399. Kayashima Y, Smithies O, Kakoki M. The kallikrein-kinin system and oxidative stress. Curr Opin Nephrol Hypertens. 2012;21(1):92–96. doi:10.1097/MNH.0b013e32834d54b1
400. Qiu X, Ma C, Luo Z, et al. Bradykinin protects nucleus pulposus cells from tert-butyl hydroperoxide-induced damage and delays intervertebral disc degeneration. Int immunopharmacol. 2024;134:112161. doi:10.1016/j.intimp.2024.112161
401. Wang Z, Zou Y, Li Y, Cheng Y. Metal-Containing Polydopamine Nanomaterials: catalysis, Energy, and Theranostics. Small (Weinheim an der Bergstrasse, Germany). 2020;16(18):e1907042. doi:10.1002/smll.201907042
402. Zielińska A, Carreiró F, Oliveira AM, et al. Polymeric Nanoparticles: production, Characterization, Toxicology and Ecotoxicology. Molecules (Basel, Switzerland). 2020;25(16):3731. doi:10.3390/molecules25163731
403. Amaldoss MJN, Yang JL, Koshy P, Unnikrishnan A, Sorrell CC. Inorganic nanoparticle-based advanced cancer therapies: promising combination strategies. Drug Discovery Today. 2022;27(12):103386. doi:10.1016/j.drudis.2022.103386
404. Mohammadpour R, Dobrovolskaia MA, Cheney DL, Greish KF, Ghandehari H. Subchronic and chronic toxicity evaluation of inorganic nanoparticles for delivery applications. Adv Drug Delivery Rev. 2019;144:112–132. doi:10.1016/j.addr.2019.07.006
405. Unnikrishnan G, Joy A, Megha M, Kolanthai E, Senthilkumar M. Exploration of inorganic nanoparticles for revolutionary drug delivery applications: a critical review. Discover Nano. 2023;18(1):157. doi:10.1186/s11671-023-03943-0
406. Wen J, Yang K, Sun S. MnO(2)-Based nanosystems for cancer therapy. Chemical Communications (Cambridge, England). 2020;56(52):7065–7079. doi:10.1039/D0CC02782K
407. Zhang W, Yang M, Sun T, Zhang J, Zhao Y. Can Manganese Dioxide Microspheres be Used as Intermediaries to Alleviate Intervertebral Disc Degeneration With Strengthening Drugs? Front Bioeng Biotechnol. 2022;10:866290. doi:10.3389/fbioe.2022.866290
408. Li C, Zhao Z, Luo Y, et al. Macrophage-Disguised Manganese Dioxide Nanoparticles for Neuroprotection by Reducing Oxidative Stress and Modulating Inflammatory Microenvironment in Acute Ischemic Stroke. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2021;8(20):e2101526. doi:10.1002/advs.202101526
409. Nakawaki M, Uchida K, Miyagi M, et al. Changes in Nerve Growth Factor Expression and Macrophage Phenotype Following Intervertebral Disc Injury in Mice. J Orthopaedic Res. 2019;37(8):1798–1804. doi:10.1002/jor.24308
410. Lee S, Hwang C, Marini S, et al. NGF-TrkA signaling dictates neural ingrowth and aberrant osteochondral differentiation after soft tissue trauma. Nat Commun. 2021;12(1):4939. doi:10.1038/s41467-021-25143-z
411. Song X, Ding Q, Wei W, et al. Peptide-Functionalized Prussian Blue Nanomaterial for Antioxidant Stress and NIR Photothermal Therapy against Alzheimer’s Disease. Small. 2023;19(41):e2206959. doi:10.1002/smll.202206959
412. Lan X, Zhuo J, Luo L, et al. Metal-phenolic networks derived CN-FeC hollow nanozyme with robust peroxidase-like activity for total antioxidant capacity detection. Colloids Surf. B. 2024;234:113640. doi:10.1016/j.colsurfb.2023.113640
413. Chen J, Pan S, Zhou J, et al. Assembly of Bioactive Nanoparticles via Metal-Phenolic Complexation. Adv Mater. 2022;34(10):e2108624. doi:10.1002/adma.202108624
414. Chatterjee N, Kumar P, Kumar K, Misra SK. What makes carbon nanoparticle a potent material for biological application? Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2022;14(3):e1782. doi:10.1002/wnan.1782
415. Borisenkova AA, Bolshakova OI, Titova AV, et al. Fullerene C(60) Conjugate with Folic Acid and Polyvinylpyrrolidone for Targeted Delivery to Tumor Cells. Int J Mol Sci. 2024;25(10):5350. doi:10.3390/ijms25105350
416. Zhang G, Fang H, Chang S, et al. Fullerene [60] encapsulated water-soluble supramolecular cage for prevention of oxidative stress-induced myocardial injury. Mater Today Bio. 2023;21:100693. doi:10.1016/j.mtbio.2023.100693
417. Xiao L, Huang R, Zhang Y, et al. A New Formyl Peptide Receptor-1 Antagonist Conjugated Fullerene Nanoparticle for Targeted Treatment of Degenerative Disc Diseases. ACS Appl. Mater. Interfaces. 2019;11(42):38405–38416. doi:10.1021/acsami.9b11783
418. Pei Y, Cui F, Du X, et al. Antioxidative nanofullerol inhibits macrophage activation and development of osteoarthritis in rats. Int J Nanomed. 2019;14:4145–4155. doi:10.2147/IJN.S202466
419. Liu Q, Jin L, Shen FH, Balian G, Li XJ. Fullerol nanoparticles suppress inflammatory response and adipogenesis of vertebral bone marrow stromal cells--a potential novel treatment for intervertebral disc degeneration. Spine J. 2013;13(11):1571–1580. doi:10.1016/j.spinee.2013.04.004
420. Zare H, Ahmadi S, Ghasemi A, et al. Carbon Nanotubes: smart Drug/Gene Delivery Carriers. Int J Nanomed. 2021;16:1681–1706. doi:10.2147/IJN.S299448
421. Raphey VR, Henna TK, Nivitha KP, Sabu C, Pramod K, Pramod K. Advanced biomedical applications of carbon nanotube. Mater Sci Eng C Mater Biol Appl. 2019;100:616–630. doi:10.1016/j.msec.2019.03.043
422. Geng H, Chen J, Tu K, et al. Carbon dot nanozymes as free radicals scavengers for the management of hepatic ischemia-reperfusion injury by regulating the liver inflammatory network and inhibiting apoptosis. J Nanobiotechnol. 2023;21(1):500.
423. Jaleel JA, Pramod K. Artful and multifaceted applications of carbon dot in biomedicine. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2018;269:302–321.
424. Kong J, Zhou F. Preparation and Application of Carbon Dots Nanozymes. Antioxidants. 2024;13(5):535.
425. Gong N, Ma X, Ye X, et al. Carbon-dot-supported atomically dispersed gold as a mitochondrial oxidative stress amplifier for cancer treatment. Nature Nanotechnol. 2019;14(4):379–387. doi:10.1038/s41565-019-0373-6
426. Bodratti AM, Alexandridis P. Amphiphilic block copolymers in drug delivery: advances in formulation structure and performance. Expert Opin Drug Delivery. 2018;15(11):1085–1104. doi:10.1080/17425247.2018.1529756
427. Hari SK, Gauba A, Shrivastava N, Tripathi RM, Jain SK, Pandey AK. Polymeric micelles and cancer therapy: an ingenious multimodal tumor-targeted drug delivery system. Drug Delivery Transl Res. 2023;13(1):135–163. doi:10.1007/s13346-022-01197-4
428. Movassaghian S, Merkel OM, Torchilin VP. Applications of polymer micelles for imaging and drug delivery. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2015;7(5):691–707. doi:10.1002/wnan.1332
429. Hwang D, Ramsey JD, Kabanov AV. Polymeric micelles for the delivery of poorly soluble drugs: from nanoformulation to clinical approval. Adv Drug Delivery Rev. 2020;156:80–118. doi:10.1016/j.addr.2020.09.009
430. Kotta S, Aldawsari HM, Badr-Eldin SM, Nair AB, Yt K. Progress in Polymeric Micelles for Drug Delivery Applications. Pharmaceutics. 2022;14(8):1636. doi:10.3390/pharmaceutics14081636
431. Chang CC, Tsou HK, Chang HH, et al. Runx1 Messenger RNA Delivered by Polyplex Nanomicelles Alleviate Spinal Disc Hydration Loss in a Rat Disc Degeneration Model. Int J Mol Sci. 2022;23(1):565.
432. Pattnaik S, Swain K, Ramakrishna S. Optimal delivery of poorly soluble drugs using electrospun nanofiber technology: challenges, state of the art, and future directions. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2023;15(2):e1859. doi:10.1002/wnan.1859
433. Anup N, Chavan T, Chavan S, et al. Reinforced electrospun nanofiber composites for drug delivery applications. J Biomed Mater Res Part A. 2021;109(10):2036–2064. doi:10.1002/jbm.a.37187
434. Wildy M, Lu P. Electrospun Nanofibers: shaping the Future of Controlled and Responsive Drug Delivery. Materials. 2023;16(22):7062. doi:10.3390/ma16227062
435. Jiang Z, Zheng Z, Yu S, et al. Nanofiber Scaffolds as Drug Delivery Systems Promoting Wound Healing. Pharmaceutics. 2023;15(7):1829. doi:10.3390/pharmaceutics15071829
436. Formica FA, Öztürk E, Hess SC, et al. A Bioinspired Ultraporous Nanofiber-Hydrogel Mimic of the Cartilage Extracellular Matrix. Adv Healthcare Mater. 2016;5(24):3129–3138. doi:10.1002/adhm.201600867
437. Li C, Chen J, Lv Y, et al. Recent Progress in Electrospun Nanofiber-Based Degenerated Intervertebral Disc Repair. ACS Biomater Sci Eng. 2022;8(1):16–31. doi:10.1021/acsbiomaterials.1c00970
438. Koepsell L, Zhang L, Neufeld D, Fong H, Deng Y. Electrospun nanofibrous polycaprolactone scaffolds for tissue engineering of annulus fibrosus. Macromol biosci. 2011;11(3):391–399. doi:10.1002/mabi.201000352
439. Martin JT, Milby AH, Chiaro JA, et al. Translation of an engineered nanofibrous disc-like angle-ply structure for intervertebral disc replacement in a small animal model. Acta Biomater. 2014;10(6):2473–2481. doi:10.1016/j.actbio.2014.02.024
440. Uysal O, Arslan E, Gulseren G, et al. Collagen Peptide Presenting Nanofibrous Scaffold for Intervertebral Disc Regeneration. ACS Appl. Bio Mater. 2019;2(4):1686–1695. doi:10.1021/acsabm.9b00062
441. Liu C, Xiao L, Zhang Y, Zhao Q, Xu H. Regeneration of annulus fibrosus tissue using a DAFM/PECUU-blended electrospun scaffold. J biomater sci Poly ed. 2020;31(18):2347–2361. doi:10.1080/09205063.2020.1812038
442. Madhukiran D, Jha A, Kumar M, Ajmal G, Bonde GV, Mishra B. Electrospun nanofiber-based drug delivery platform: advances in diabetic foot ulcer management. Expert Opin Drug Delivery. 2021;18(1):25–42. doi:10.1080/17425247.2021.1823966
443. Makhathini SS, Kondiah PJ, Kharodia ME, et al. Biomedicine Innovations and Its Nanohydrogel Classifications. Pharmaceutics. 2022;14(12):2839. doi:10.3390/pharmaceutics14122839
444. Damiri F, Fatimi A, Santos ACP, Varma RS, Berrada M. Smart stimuli-responsive polysaccharide nanohydrogels for drug delivery: a review. J Mat Chem B. 2023;11(44):10538–10565.
445. Desai SU, Srinivasan SS, Kumbar SG, Moss IL. Hydrogel-Based Strategies for Intervertebral Disc Regeneration: advances, Challenges and Clinical Prospects. Gels (Basel, Switzerland). 2024;10(1). doi:10.3390/gels10010062
446. Zheng K, Du D. Recent advances of hydrogel-based biomaterials for intervertebral disc tissue treatment: a literature review. J Tissue Eng Regener Med. 2021;15(4):299–321. doi:10.1002/term.3172
447. Chen X, Jing S, Xue C, Guan X. Progress in the Application of Hydrogels in Intervertebral Disc Repair: a Comprehensive Review. Current Pain and Headache Reports. 2024;28(12):1333–1348.
448. Liu Y, Zhao Z, Guo C, et al. Application and development of hydrogel biomaterials for the treatment of intervertebral disc degeneration: a literature review. Front Cell Develop Biol. 2023;11:1286223.
449. Cao H, Duan L, Zhang Y, Cao J, Zhang K. Current hydrogel advances in physicochemical and biological response-driven biomedical application diversity. Signal Transduction and Targeted Therapy. 2021;6(1):426.
450. Mikhail AS, Morhard R, Mauda-Havakuk M, Arrichiello A, Wood BJ. Hydrogel drug delivery systems for minimally invasive local immunotherapy of cancer. Adv Drug Delivery Rev. 2023;202:115083.
451. Shaikh MAJ, Alharbi KS, Almalki WH, et al. Sodium alginate based drug delivery in management of breast cancer. Carbohydr Polym. 2022;292:119689.
452. Lai P, Daear W, Löbenberg R, Prenner EJ. Overview of the preparation of organic polymeric nanoparticles for drug delivery based on gelatine, chitosan, poly(d,l-lactide-co-glycolic acid) and polyalkylcyanoacrylate. Colloids Surf B. 2014;118:154–163. doi:10.1016/j.colsurfb.2014.03.017
453. Serna JA, Rueda-Gensini L, Céspedes-Valenzuela DN, Cifuentes J, Cruz JC, Muñoz-Camargo C. Recent Advances on Stimuli-Responsive Hydrogels Based on Tissue-Derived ECMs and Their Components: towards Improving Functionality for Tissue Engineering and Controlled Drug Delivery. Polymers. 2021;13(19):3263. doi:10.3390/polym13193263
454. Dikici S, Aldemir Dikici B, MacNeil S, Claeyssens F. Decellularised extracellular matrix decorated PCL PolyHIPE scaffolds for enhanced cellular activity, integration and angiogenesis. Biomater Sci. 2021;9(21):7297–7310. doi:10.1039/D1BM01262B
455. Kurian AG, Singh RK, Patel KD, Lee JH, Kim HW. Multifunctional GelMA platforms with nanomaterials for advanced tissue therapeutics. Bioactive Mater. 2022;8:267–295.
456. Lv B, Lu L, Hu L, et al. Recent advances in GelMA hydrogel transplantation for musculoskeletal disorders and related disease treatment. Theranostics. 2023;13(6):2015–2039. doi:10.7150/thno.80615
457. Li P, Zhang M, Chen Z, Tian B, Kang X. Tissue-Engineered Injectable Gelatin-Methacryloyl Hydrogel-Based Adjunctive Therapy for Intervertebral Disc Degeneration. ACS omega. 2023;8(15):13509–13518. doi:10.1021/acsomega.3c00211
458. Ao Y, Zhang E, Liu Y, Yang L, Li J, Wang F. Advanced Hydrogels With Nanoparticle Inclusion for Cartilage Tissue Engineering. Front Bioeng Biotechnol. 2022;10:951513.
459. Zhang P, Qi J, Zhang R, et al. Recent advances in composite hydrogels: synthesis, classification, and application in the treatment of bone defects. Biomater Sci. 2024;12(2):308–329.
460. Chen Q, He J, Liu H, et al. Small extracellular vesicles-transported lncRNA TDRKH-AS1 derived from AOPPs-treated trophoblasts initiates endothelial cells pyroptosis through PDIA4/DDIT4 axis in preeclampsia. J Transl Med. 2023;21(1):496.
461. Zhidkova EM, Lylova ES, Grigoreva DD, et al. Nutritional Sensor REDD1 in Cancer and Inflammation: friend or Foe? Int J Mol Sci. 2022;23(17):9686. doi:10.3390/ijms23179686
462. Han QF, Li WJ, Hu KS, et al. Exosome biogenesis: machinery, regulation, and therapeutic implications in cancer. Mol Cancer. 2022;21(1):207. doi:10.1186/s12943-022-01671-0
463. Arya SB, Collie SP, Parent CA. The ins-and-outs of exosome biogenesis, secretion, and internalization. Trends Cell Biol. 2024;34(2):90–108. doi:10.1016/j.tcb.2023.06.006
464. Lu M, Huang Y. Bioinspired exosome-like therapeutics and delivery nanoplatforms. Biomaterials. 2020;242:119925. doi:10.1016/j.biomaterials.2020.119925
465. Dad HA, Gu TW, Zhu AQ, Huang LQ, Peng LH. Plant Exosome-like Nanovesicles: emerging Therapeutics and Drug Delivery Nanoplatforms. Molecular Therapy: the Journal of the American Society of Gene Therapy. 2021;29(1):13–31. doi:10.1016/j.ymthe.2020.11.030
466. Fan C, Wang W, Yu Z, et al. M1 macrophage-derived exosomes promote intervertebral disc degeneration by enhancing nucleus pulposus cell senescence through LCN2/NF-κB signaling axis. J Nanobiotechnol. 2024;22(1):301. doi:10.1186/s12951-024-02556-8
467. Li W, Zhang S, Wang D, et al. Exosomes Immunity Strategy: a Novel Approach for Ameliorating Intervertebral Disc Degeneration. Front Cell Develop Biol. 2021;9:822149. doi:10.3389/fcell.2021.822149
468. Sun Z, Zhao H, Liu B, et al. AF cell derived exosomes regulate endothelial cell migration and inflammation: implications for vascularization in intervertebral disc degeneration. Life Sci. 2021;265:118778. doi:10.1016/j.lfs.2020.118778
469. CC C, Chen J, Wang WL, Xie L, Shao CQ, Zhang YX. Inhibition of the P53/P21 Pathway Attenuates the Effects of Senescent Nucleus Pulposus Cell-Derived Exosomes on the Senescence of Nucleus Pulposus Cells. Orthopaedic Surgery. 2021;13(2):583–591. doi:10.1111/os.12886
470. Guo Z, Su W, Zhou R, et al. Exosomal MATN3 of Urine-Derived Stem Cells Ameliorates Intervertebral Disc Degeneration by Antisenescence Effects and Promotes NPC Proliferation and ECM Synthesis by Activating TGF- β. Oxid Med Cell Longev. 2021;2021(1):5542241. doi:10.1155/2021/5542241
471. Xia C, Zeng Z, Fang B, et al. Mesenchymal stem cell-derived exosomes ameliorate intervertebral disc degeneration via anti-oxidant and anti-inflammatory effects. Free Radic Biol Med. 2019;143:1–15. doi:10.1016/j.freeradbiomed.2019.07.026
472. Zeng H, Guo S, Ren X, Wu Z, Liu S, Yao X. Current Strategies for Exosome Cargo Loading and Targeting Delivery. Cells. 2023;12(10):1416. doi:10.3390/cells12101416
473. Sadeghi S, Tehrani FR, Tahmasebi S, Shafiee A, Hashemi SM. Exosome engineering in cell therapy and drug delivery. Inflammopharmacology. 2023;31(1):145–169. doi:10.1007/s10787-022-01115-7
474. Marquez CA, Oh CI, Ahn G, Shin WR, Kim YH, Ahn JY. Synergistic vesicle-vector systems for targeted delivery. J Nanobiotechnol. 2024;22(1):6. doi:10.1186/s12951-023-02275-6
475. Zhang Y, Liu Q, Zhang X, et al. Recent advances in exosome-mediated nucleic acid delivery for cancer therapy. J Nanobiotechnol. 2022;20(1):279. doi:10.1186/s12951-022-01472-z
476. Kar R, Dhar R, Mukherjee S, et al. Exosome-Based Smart Drug Delivery Tool for Cancer Theranostics. ACS Biomater Sci Eng. 2023;9(2):577–594. doi:10.1021/acsbiomaterials.2c01329
477. Peng S, Xiao F, Chen M, Gao H. Tumor-Microenvironment-Responsive Nanomedicine for Enhanced Cancer Immunotherapy. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2022;9(1):e2103836. doi:10.1002/advs.202103836
478. He Q, Chen J, Cai S, et al. Tumor microenvironment responsive drug delivery systems. Asian J Pharm Sci. 2020;15(4):416–448. doi:10.1016/j.ajps.2019.08.003
479. Zhao H, Li Y, Chen J, et al. Environmental stimulus-responsive mesoporous silica nanoparticles as anticancer drug delivery platforms. Colloids Surf. B. 2024;234:113758. doi:10.1016/j.colsurfb.2024.113758
480. Ozawa R, Saita M, Sakaue S, et al. Redox injectable gel protects osteoblastic function against oxidative stress and suppresses alveolar bone loss in a rat peri-implantitis model. Acta Biomater. 2020;110:82–94. doi:10.1016/j.actbio.2020.04.003
481. Yu H, Jin F, Liu D, et al. ROS-responsive nano-drug delivery system combining mitochondria-targeting ceria nanoparticles with atorvastatin for acute kidney injury. Theranostics. 2020;10(5):2342–2357. doi:10.7150/thno.40395
482. Cheng R, Feng F, Deng C, Feijen J, Zhong Z, Zhong Z. Glutathione-responsive nano-vehicles as a promising platform for targeted intracellular drug and gene delivery. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2011;152(1):2–12. doi:10.1016/j.jconrel.2011.01.030
483. Zhang M, Hu W, Cai C, Wu Y, Li J, Dong S. Advanced application of stimuli-responsive drug delivery system for inflammatory arthritis treatment. Mater Today Bio. 2022;14:100223. doi:10.1016/j.mtbio.2022.100223
484. Abed HF, Abuwatfa WH, Husseini GA. Redox-Responsive Drug Delivery Systems: a Chemical Perspective. Nanomaterials (Basel, Switzerland). 2022;12(18). doi:10.3390/nano12183183
485. Danhier F, Ansorena E, Silva JM, Coco R, Le Breton A, Préat V. PLGA-based nanoparticles: an overview of biomedical applications. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2012;161(2):505–522. doi:10.1016/j.jconrel.2012.01.043
486. Gong W, Xia C, He Q. Therapeutic gas delivery strategies. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2022;14(1):e1744. doi:10.1002/wnan.1744
487. Andrabi SM, Sharma NS, Karan A, Shahriar SMS, Cordon B, Xie J. Nitric Oxide: physiological Functions, Delivery, and Biomedical Applications. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2023;10(30):e2303259. doi:10.1002/advs.202303259
488. Jiang H, Ji P, Shang X, Zhou Y. Connection between Osteoarthritis and Nitric Oxide: from Pathophysiology to Therapeutic Target. Molecules. 2023;28(4):1683.
489. Kitano T, Zerwekh JE, Usui Y, Edwards ML, Flicker PL, Mooney V. Biochemical changes associated with the symptomatic human intervertebral disk. Clin Orthopaedics Related Res. 1993;1993(293):372–377.
490. Gilbert HTJ, Hodson N, Baird P, Richardson SM, Hoyland JA. Acidic pH promotes intervertebral disc degeneration: acid-sensing ion channel −3 as a potential therapeutic target. Sci Rep. 2016;6(1):37360. doi:10.1038/srep37360
491. Gannimani R, Walvekar P, Naidu VR, Aminabhavi TM, Govender T. Govender T: acetal containing polymers as pH-responsive nano-drug delivery systems. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2020;328:736–761. doi:10.1016/j.jconrel.2020.09.044
492. Verkhovskii RA, Ivanov AN, Lengert EV, Tulyakova KA, Shilyagina NY, Ermakov AV. Current Principles, Challenges, and New Metrics in pH-Responsive Drug Delivery Systems for Systemic Cancer Therapy. Pharmaceutics. 2023;15(5):1566. doi:10.3390/pharmaceutics15051566
493. Ding H, Tan P, Fu S, et al. Preparation and application of pH-responsive drug delivery systems. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2022;348:206–238. doi:10.1016/j.jconrel.2022.05.056
494. Ohsawa I, Ishikawa M, Takahashi K, et al. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nature Med. 2007;13(6):688–694. doi:10.1038/nm1577
495. Taherzadeh-Fard E, Saft C, Akkad DA, et al.. PGC-1alpha downstream transcription factors NRF-1 and TFAM are genetic modifiers of Huntington disease. Molecular Neurodegeneration. 2011;6(1):32. doi:10.1186/1750-1326-6-32
496. Ma L, Wang Y, Li X, et al. Tom70-regulated mitochondrial biogenesis via TFAM improves hypoxia-induced dysfunction of pulmonary vascular endothelial cells and alleviates hypoxic pulmonary hypertension. Respir Res. 2023;24(1):310. doi:10.1186/s12931-023-02631-y
497. Shi PZ, Wang JW, Wang PC, et al. Urolithin a alleviates oxidative stress-induced senescence in nucleus pulposus-derived mesenchymal stem cells through SIRT1/PGC-1α pathway. World Journal of Stem Cells. 2021;13(12):1928–1946. doi:10.4252/wjsc.v13.i12.1928
498. Massagué J, Sheppard D. TGF-β signaling in health and disease. Cell. 2023;186(19):4007–4037. doi:10.1016/j.cell.2023.07.036
499. Wu T, Li X, Zhu Z, et al. Krüppel like factor 10 prevents intervertebral disc degeneration via TGF-β signaling pathway both in vitro and in vivo. Journal of Orthopaedic Translation. 2021;29:19–29. doi:10.1016/j.jot.2021.04.003
500. Sun T, Jiang C. Stimuli-responsive drug delivery systems triggered by intracellular or subcellular microenvironments. Adv Drug Delivery Rev. 2023;196:114773. doi:10.1016/j.addr.2023.114773
501. Mura S, Nicolas J, Couvreur P. Stimuli-responsive nanocarriers for drug delivery. Nature Mater. 2013;12(11):991–1003. doi:10.1038/nmat3776
502. Fang H, Xu S, Wang Y, Yang H, Su D. Endogenous stimuli-responsive drug delivery nanoplatforms for kidney disease therapy. Colloids Surf. B. 2023;232:113598. doi:10.1016/j.colsurfb.2023.113598
503. Huang Y, Li P, Zhao R, et al. Silica nanoparticles: biomedical applications and toxicity. Biomed Pharmacother. 2022;151:113053. doi:10.1016/j.biopha.2022.113053
504. Ignacio RM, Kim DH, Sajo ME, et al. Immunotoxicity of silicon dioxide nanoparticles with different sizes and electrostatic charge. Int J Nanomed. 2014;9(Suppl 2):183–193. doi:10.2147/IJN.S57934
505. Hu J, Zhang G, Liu S. Enzyme-responsive polymeric assemblies, nanoparticles and hydrogels. Chem Soc Rev. 2012;41(18):5933–5949. doi:10.1039/c2cs35103j
506. Millward-Sadler SJ, Costello PW, Freemont AJ, Hoyland JA. Regulation of catabolic gene expression in normal and degenerate human intervertebral disc cells: implications for the pathogenesis of intervertebral disc degeneration. Arthritis Res Therapy. 2009;11(3):R65. doi:10.1186/ar2693
507. Hu Q, Katti PS, Gu Z. Enzyme-responsive nanomaterials for controlled drug delivery. Nanoscale. 2014;6(21):12273–12286. doi:10.1039/C4NR04249B
508. Soh WT, Zhang J, Hollenberg MD, et al. Protease allergens as initiators-regulators of allergic inflammation. Allergy. 2023;78(5):1148–1168. doi:10.1111/all.15678
509. Chen D, Jiang X. Correlation Between Proteolytic Enzymes and Microangiogenesis in Degenerative Intervertebral Disc Nucleus. Journal of Investigative Surgery: the Official Journal of the Academy of Surgical Research. 2021;34(6):679–684. doi:10.1080/08941939.2019.1679921
510. Lin IT, Lin YH, Lian WS, Wang FS, Wu RW. MicroRNA-29a Mitigates Laminectomy-Induced Spinal Epidural Fibrosis and Gait Dysregulation by Repressing TGF-β1 and IL-6. Int J Mol Sci. 2023;24(11). doi:10.3390/ijms24119158
511. Shahriari M, Zahiri M, Abnous K, Taghdisi SM, Ramezani M, Alibolandi M. Enzyme responsive drug delivery systems in cancer treatment. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2019;308:172–189. doi:10.1016/j.jconrel.2019.07.004
512. Gao B, Jiang B, Xing W, Xie Z, Zou W, Zou W. Discovery and Application of Postnatal Nucleus Pulposus Progenitors Essential for Intervertebral Disc Homeostasis and Degeneration. Advanced Science (Weinheim, Baden-Wurttemberg, Germany). 2022;9(13):e2104888. doi:10.1002/advs.202104888
513. Zhang W, Sun T, Li Z. Application of stem cells in the repair of intervertebral disc degeneration. Stem Cell Res Ther. 2022;13(1):70. doi:10.1186/s13287-022-02745-y
514. Ekram S, Salim A, Khan I, Khan I. Regulating the fate of stem cells for regenerating the intervertebral disc degeneration. World Journal of Stem Cells. 2021;13(12):1881–1904. doi:10.4252/wjsc.v13.i12.1881
515. Zhang YM, Liu YH, Liu Y. Cyclodextrin-Based Multistimuli-Responsive Supramolecular Assemblies and Their Biological Functions. Advanced Materials (Deerfield Beach, Fla). 2020;32(3):e1806158. doi:10.1002/adma.201806158
516. Li Z, Jiang Y, Zhao H, Liu L. Ca(2+)-Chelation-Induced Fabrication of Multistimuli-Responsive Charged Nanogels from Phospholipid-Polymer Conjugates and Use for Drug/Protein Loading. Langmuir: the ACS Journal of Surfaces and Colloids. 2022;38(21):6612–6622. doi:10.1021/acs.langmuir.2c00464
517. Kolluru GK, Shackelford RE, Shen X, Dominic P, Kevil CG. Sulfide regulation of cardiovascular function in health and disease. Nat Rev Cardiol. 2023;20(2):109–125. doi:10.1038/s41569-022-00741-6
518. Sun HJ, Wu ZY, Nie XW, Wang XY, Bian JS. Implications of hydrogen sulfide in liver pathophysiology: mechanistic insights and therapeutic potential. J Adv Res. 2021;27:127–135. doi:10.1016/j.jare.2020.05.010
519. Scammahorn JJ, Nguyen ITN, Bos EM, Van Goor H, Joles JA. Fighting Oxidative Stress with Sulfur: hydrogen Sulfide in the Renal and Cardiovascular Systems. Antioxidants (Basel, Switzerland). 2021;10(3). doi:10.3390/antiox10030373
520. Greabu M, Totan A, Miricescu D, Radulescu R, Virlan J, Calenic B. Calenic B: hydrogen Sulfide, Oxidative Stress and Periodontal Diseases: a Concise Review. Antioxidants (Basel, Switzerland). 2016;5(1). doi:10.3390/antiox5010003
521. Xu D, Jin H, Wen J, et al. Hydrogen sulfide protects against endoplasmic reticulum stress and mitochondrial injury in nucleus pulposus cells and ameliorates intervertebral disc degeneration. Pharmacol Res. 2017;117:357–369. doi:10.1016/j.phrs.2017.01.005
522. Kang J, Li Z, Organ CL, et al. Xian M: pH-Controlled Hydrogen Sulfide Release for Myocardial Ischemia-Reperfusion Injury. J Am Chem Soc. 2016;138(20):6336–6339. doi:10.1021/jacs.6b01373
523. Wu J, Chen A, Zhou Y, et al. Novel H(2)S-Releasing hydrogel for wound repair via in situ polarization of M2 macrophages. Biomaterials. 2019;222:119398. doi:10.1016/j.biomaterials.2019.119398
524. Zhou ZH, Liang SY, Zhao TC, et al. Overcoming chemotherapy resistance using pH-sensitive hollow MnO(2) nanoshells that target the hypoxic tumor microenvironment of metastasized oral squamous cell carcinoma. J Nanobiotechnol. 2021;19(1):157. doi:10.1186/s12951-021-00901-9
525. Sisakhtnezhad S, Rahimi M, Mohammadi S. Biomedical applications of MnO(2) nanomaterials as nanozyme-based theranostics. Biomed Pharmacother. 2023;163:114833. doi:10.1016/j.biopha.2023.114833
526. Chamoto K, Zhang B, Tajima M, Honjo T, Fagarasan S. Spermidine - an old molecule with a new age-defying immune function. Trends Cell Biol. 2024;34(5):363–370. doi:10.1016/j.tcb.2023.08.002
527. Niu C, Jiang D, Guo Y, et al. Spermidine suppresses oxidative stress and ferroptosis by Nrf2/HO-1/GPX4 and Akt/FHC/ACSL4 pathway to alleviate ovarian damage. Life Sci. 2023;332:122109. doi:10.1016/j.lfs.2023.122109
528. Xing Y, Zeng B, Yang W. Light responsive hydrogels for controlled drug delivery. Front Bioeng Biotechnol. 2022;10:1075670. doi:10.3389/fbioe.2022.1075670
529. Yuan Z, Gottsacker C, He X, Waterkotte T, Park YC. Repetitive drug delivery using Light-Activated liposomes for potential antimicrobial therapies. Adv Drug Delivery Rev. 2022;187:114395. doi:10.1016/j.addr.2022.114395
530. Liu Y, Chen L, Shi Q, Ma H, Ma H. Tumor Microenvironment-Responsive Polypeptide Nanogels for Controlled Antitumor Drug Delivery. Front Pharmacol. 2021;12:748102. doi:10.3389/fphar.2021.748102
531. Nemoto R, Fujieda K, Hiruta Y, et al. Liposomes with temperature-responsive reversible surface properties. Colloids Surf. B. 2019;176:309–316. doi:10.1016/j.colsurfb.2019.01.007
532. Nagase K. Thermoresponsive interfaces obtained using poly(N-isopropylacrylamide)-based copolymer for bioseparation and tissue engineering applications. Adv Colloid Interface Sci. 2021;295:102487. doi:10.1016/j.cis.2021.102487
533. Tono Y, Kojima C, Haba Y, et al. Thermosensitive properties of poly(amidoamine) dendrimers with peripheral phenylalanine residues. Langmuir: the ACS Journal of Surfaces and Colloids. 2006;22(11):4920–4922. doi:10.1021/la060066t
534. Mathivanan N, Paramasivam G, Vergaelen M, Rajendran J, Hoogenboom R, Sundaramurthy A. Hydrogen-Bonded Multilayer Thin Films and Capsules Based on Poly(2-n-propyl-2-oxazoline) and Tannic Acid: investigation on Intermolecular Forces, Stability, and Permeability. Langmuir: the ACS Journal of Surfaces and Colloids. 2019;35(45):14712–14724. doi:10.1021/acs.langmuir.9b02938
535. Yuan W, Zou H, Shen J. Amphiphilic graft copolymers with ethyl cellulose backbone: synthesis, self-assembly and tunable temperature-CO2 response. Carbohydr Polym. 2016;136:216–223. doi:10.1016/j.carbpol.2015.09.052
536. Malonzo C, Chan SC, Kabiri A, et al. A papain-induced disc degeneration model for the assessment of thermo-reversible hydrogel-cells therapeutic approach. J Tissue Eng Regener Med. 2015;9(12):E167–176. doi:10.1002/term.1667
537. Guo W, Douma L, Hu MH, et al. Hyaluronic acid-based interpenetrating network hydrogel as a cell carrier for nucleus pulposus repair. Carbohydr Polym. 2022;277:118828. doi:10.1016/j.carbpol.2021.118828
538. Huang Y, Huang L, Li L, et al. MicroRNA-25-3p therapy for intervertebral disc degeneration by targeting the IL-1β/ZIP8/MTF1 signaling pathway with a novel thermo-responsive vector. Ann Translat Med. 2020;8(22):1500. doi:10.21037/atm-20-6595
539. Chen Q, Wang J, Xia Q, et al. Treatment outcomes of injectable thermosensitive hydrogel containing bevacizumab in intervertebral disc degeneration. Front Bioeng Biotechnol. 2022;10:976706. doi:10.3389/fbioe.2022.976706
540. Kang ML, Kim JE, Im GI. Thermoresponsive nanospheres with independent dual drug release profiles for the treatment of osteoarthritis. Acta Biomater. 2016;39:65–78. doi:10.1016/j.actbio.2016.05.005
541. Mi P. Stimuli-responsive nanocarriers for drug delivery, tumor imaging, therapy and theranostics. Theranostics. 2020;10(10):4557–4588. doi:10.7150/thno.38069
542. Li J, Gao Y, Liu S, et al. Aptamer-functionalized quercetin thermosensitive liposomes for targeting drug delivery and antitumor therapy. Biomedical Materials (Bristol, England). 2022;17(6):065003. doi:10.1088/1748-605X/ac8c75
543. Municoy S, Álvarez Echazú MI, Antezana PE, et al. Stimuli-Responsive Materials for Tissue Engineering and Drug Delivery. Int J Mol Sci. 2020;21(13):4724. doi:10.3390/ijms21134724
544. Linsley CS, Wu BM. Recent advances in light-responsive on-demand drug-delivery systems. Therapeutic Delivery. 2017;8(2):89–107. doi:10.4155/tde-2016-0060
545. Kumar D, Gerges I, Tamplenizza M, Lenardi C, Forsyth NR, Liu Y. Three-dimensional hypoxic culture of human mesenchymal stem cells encapsulated in a photocurable, biodegradable polymer hydrogel: a potential injectable cellular product for nucleus pulposus regeneration. Acta Biomater. 2014;10(8):3463–3474. doi:10.1016/j.actbio.2014.04.027
546. Chen P, Ning L, Qiu P, et al. Photo-crosslinked gelatin-hyaluronic acid methacrylate hydrogel-committed nucleus pulposus-like differentiation of adipose stromal cells for intervertebral disc repair. J Tissue Eng Regener Med. 2019;13(4):682–693. doi:10.1002/term.2841
547. Lin CL, Su YW, Kuo CH, Tu TY, Tsai JC, Shyong YJ. BMSC loaded photo-crosslinked hyaluronic acid/collagen hydrogel incorporating FG4592 for enhanced cell proliferation and nucleus pulposus differentiation. Int J Biol Macromol. 2024;273(Pt 1):132828. doi:10.1016/j.ijbiomac.2024.132828
548. Ramasamy T, Ruttala HB, Munusamy S, Chakraborty N, Kim JO. Nano drug delivery systems for antisense oligonucleotides (ASO) therapeutics. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2022;352:861–878. doi:10.1016/j.jconrel.2022.10.050
549. Tao Y, Chan HF, Shi B, Li M, Leong KW. Light: a Magical Tool for Controlled Drug Delivery. Adv Funct Mater. 2020;30(49). doi:10.1002/adfm.202005029
550. Agiba AM, Arreola-Ramírez JL, Carbajal V, Segura-Medina P. Light-Responsive and Dual-Targeting Liposomes: from Mechanisms to Targeting Strategies. Molecules (Basel, Switzerland). 2024;29(3):636. doi:10.3390/molecules29030636
551. Rapp TL, DeForest CA. Targeting drug delivery with light: a highly focused approach. Adv Drug Delivery Rev. 2021;171:94–107. doi:10.1016/j.addr.2021.01.009
552. Shin Y, Husni P, Kang K, et al. Recent Advances in pH- or/and Photo-Responsive Nanovehicles. Pharmaceutics. 2021;13(5):725. doi:10.3390/pharmaceutics13050725
553. Shakya G, Cattaneo M, Guerriero G, Prasanna A, Fiorini S, Supponen O. Ultrasound-responsive microbubbles and nanodroplets: a pathway to targeted drug delivery. Adv Drug Delivery Re. 2024;206:115178. doi:10.1016/j.addr.2023.115178
554. Chandan R, Mehta S, Banerjee R. Ultrasound-Responsive Carriers for Therapeutic Applications. ACS Biomater Sci Eng. 2020;6(9):4731–4747. doi:10.1021/acsbiomaterials.9b01979
555. Barzegar-Fallah A, Gandhi K, Rizwan SB, Slatter TL, Reynolds JNJ. Harnessing Ultrasound for Targeting Drug Delivery to the Brain and Breaching the Blood-Brain Tumour Barrier. Pharmaceutics. 2022;14(10):2231. doi:10.3390/pharmaceutics14102231
556. Entzian K, Aigner A. Drug Delivery by Ultrasound-Responsive Nanocarriers for Cancer Treatment. Pharmaceutics. 2021;13(8):1135. doi:10.3390/pharmaceutics13081135
557. Negishi Y, Omata D, Iijima H, et al. Enhanced laminin-derived peptide AG73-mediated liposomal gene transfer by bubble liposomes and ultrasound. Mol Pharmaceut. 2010;7(1):217–226. doi:10.1021/mp900214s
558. Chapla R, Huynh KT, Schutt CE. Microbubble-Nanoparticle Complexes for Ultrasound-Enhanced Cargo Delivery. Pharmaceutics. 2022;14(11):2396. doi:10.3390/pharmaceutics14112396
559. Yang Y, Li Q, Guo X, Tu J, Zhang D. Mechanisms underlying sonoporation: interaction between microbubbles and cells. Ultrason Sonochem. 2020;67:105096. doi:10.1016/j.ultsonch.2020.105096
560. Kobayashi Y, Sakai D, Iwashina T, Iwabuchi S, Mochida J. Low-intensity pulsed ultrasound stimulates cell proliferation, proteoglycan synthesis and expression of growth factor-related genes in human nucleus pulposus cell line. Eur Cells Mater. 2009;17:15–22. doi:10.22203/eCM.v017a02
561. Zhang X, Hu Z, Hao J, Shen J. Low Intensity Pulsed Ultrasound Promotes the Extracellular Matrix Synthesis of Degenerative Human Nucleus Pulposus Cells Through FAK/PI3K/Akt Pathway. Spine. 2016;41(5):E248–254. doi:10.1097/BRS.0000000000001220
562. Al-Jawadi S, Thakur SS. Ultrasound-responsive lipid microbubbles for drug delivery: a review of preparation techniques to optimise formulation size, stability and drug loading. Int J Pharm. 2020;585:119559. doi:10.1016/j.ijpharm.2020.119559
563. Liang Z, Chen H, Gong X, et al. Ultrasound-Induced Destruction of Nitric Oxide-Loaded Microbubbles in the Treatment of Thrombus and Ischemia-Reperfusion Injury. Front Pharmacol. 2021;12:745693. doi:10.3389/fphar.2021.745693
564. Sutton JT, Raymond JL, Verleye MC, Pyne-Geithman GJ, Holland CK. Pulsed ultrasound enhances the delivery of nitric oxide from bubble liposomes to ex vivo porcine carotid tissue. Int J Nanomed. 2014;9:4671–4683. doi:10.2147/IJN.S63850
565. He Y, Zhang B, Chen Y, et al. Image-Guided Hydrogen Gas Delivery for Protection from Myocardial Ischemia-Reperfusion Injury via Microbubbles. ACS Appl. Mater. Interfaces. 2017;9(25):21190–21199. doi:10.1021/acsami.7b05346
566. Amin MU, Ali S, Tariq I, et al. Ultrasound-Responsive Smart Drug Delivery System of Lipid Coated Mesoporous Silica Nanoparticles. Pharmaceutics. 2021;13(9):1396. doi:10.3390/pharmaceutics13091396
567. Xia H, Zhao Y, Tong R. Ultrasound-Mediated Polymeric Micelle Drug Delivery. Adv Exp Med Biol. 2016;880:365–384.
568. Araújo EV, Carneiro SV, Neto DMA, et al. Advances in surface design and biomedical applications of magnetic nanoparticles. Adv Colloid Interface Sci. 2024;328:103166. doi:10.1016/j.cis.2024.103166
569. Mok H, Zhang M. Superparamagnetic iron oxide nanoparticle-based delivery systems for biotherapeutics. Expert Opin Drug Delivery. 2013;10(1):73–87. doi:10.1517/17425247.2013.747507
570. Fragal EH, Fragal VH, Silva EP, et al. Magnetic-responsive polysaccharide hydrogels as smart biomaterials: synthesis, properties, and biomedical applications. Carbohydr. Polym. 2022;292:119665. doi:10.1016/j.carbpol.2022.119665
571. Dadfar SM, Roemhild K, Drude NI, et al. Iron oxide nanoparticles: diagnostic, therapeutic and theranostic applications. Adv Drug Delivery Rev. 2019;138:302–325. doi:10.1016/j.addr.2019.01.005
572. Millart E, Lesieur S, Faivre V. Superparamagnetic lipid-based hybrid nanosystems for drug delivery. Expert Opin Drug Delivery. 2018;15(5):523–540. doi:10.1080/17425247.2018.1453804
573. Tong S, Quinto CA, Zhang L, Mohindra P, Bao G. Size-Dependent Heating of Magnetic Iron Oxide Nanoparticles. ACS nano. 2017;11(7):6808–6816. doi:10.1021/acsnano.7b01762
574. Bringas E, Ö K, Quach DV, et al. Triggered release in lipid bilayer-capped mesoporous silica nanoparticles containing SPION using an alternating magnetic field. Chemical Communications (Cambridge, England). 2012;48(45):5647–5649. doi:10.1039/c2cc31563g
575. Hu SH, Chen SY, Liu DM, Hsiao CS. Core/Single-Crystal-Shell Nanospheres for Controlled Drug Release via a Magnetically Triggered Rupturing Mechanism. Advanced Materials (Deerfield Beach, Fla). 2008;20(14):2690–2695. doi:10.1002/adma.200800193
576. Xu Y, Zheng H, Schumacher D, Liehn EA, Slabu I, Rusu M. Recent Advancements of Specific Functionalized Surfaces of Magnetic Nano- and Microparticles as a Theranostics Source in Biomedicine. ACS Biomater Sci Eng. 2021;7(6):1914–1932. doi:10.1021/acsbiomaterials.0c01393
577. Dinakar YH, Karole A, Parvez S, Jain V, Mudavath SL. Organ-restricted delivery through stimuli-responsive nanocarriers for lung cancer therapy. Life Sci. 2022;310:121133. doi:10.1016/j.lfs.2022.121133
578. Wang Y, Kang J, Guo X, et al. Intervertebral Disc Degeneration Models for Pathophysiology and Regenerative Therapy -Benefits and Limitations. Journal of Investigative Surgery: the Official Journal of the Academy of Surgical Research. 2022;35(4):935–952. doi:10.1080/08941939.2021.1953640
579. Poletto DL, Crowley JD, Tanglay O, Walsh WR, Pelletier MH. Preclinical in vivo animal models of intervertebral disc degeneration. Part 1: a systematic review. JOR spine. 2023;6(1):e1234. doi:10.1002/jsp2.1234
580. Beckstein JC, Sen S, Schaer TP, Vresilovic EJ, Elliott DM. Comparison of animal discs used in disc research to human lumbar disc: axial compression mechanics and glycosaminoglycan content. Spine. 2008;33(6):E166–173. doi:10.1097/BRS.0b013e318166e001
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.