Back to Journals » Drug Design, Development and Therapy » Volume 20
mTOR Signaling Pathway in Antiphospholipid Syndrome: Pathogenesis and Potential Therapeutic Targets
Authors Huo R, Wei C, Yang Y, Meng D, Yang Y, Huang X ![]()
Received 25 December 2025
Accepted for publication 11 March 2026
Published 1 April 2026 Volume 2026:20 591631
DOI https://doi.org/10.2147/DDDT.S591631
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Rongxiu Huo,* Chengcheng Wei,* Yanting Yang,* Danli Meng, Yang Yang, Xinxiang Huang
Department of Rheumatology and Immunology, Guangxi Academy of Medical Sciences, The People’s Hospital of Guangxi Zhuang Autonomous Region, Nanning, 530016, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xinxiang Huang, Department of Rheumatology and Immunology, Guangxi Academy of Medical Sciences, The People’s Hospital of Guangxi Zhuang Autonomous Region, 6 Taoyuan Road, Qingxiu District, Nanning City, Guangxi Zhuang Autonomous Region, 530016, People’s Republic of China, Email [email protected]
Abstract: Antiphospholipid syndrome (APS) is an autoimmune disorder characterized by thrombosis in arteries, veins or small blood vessels, and/or obstetric APS (OAPS), as well as persistent positive antiphospholipid antibodies. In recent years, some authors have proposed that the pathogenesis of APS is closely related to activation of vascular endothelial cells, immune cells, and complement activation. However, further exploration is still needed. Previous studies have shown that the mammalian target of rapamycin (mTOR) is associated with pro-inflammatory and pro-coagulant processes. This indicates that the activation of the mTOR signaling pathway may function as an intermediate mediator, causing immune disorders, thereby leading to thrombosis and OAPS. Therefore, we should correctly understand the potential pathogenic role of the mTOR signaling pathway in APS, which will be more conducive to clinicians’ understanding of the pathogenesis of this disease and the search for new therapeutic targets. We hope this can open up a new window for the management of APS.
Keywords: antiphospholipid syndrome, mTOR, pathogenesis, target, thrombosis, OAPS
Introduction
Antiphospholipid syndrome (APS) is an autoimmune disorder characterized by the formation of arterial and/or venous thrombosis, various obstetric complications, and persistent positivity of antiphospholipid antibodies (aPLs).1 For the overall incidence and prevalence of APS, their estimates range from 1 to 2 cases per 100,000 people and from 40 to 50 cases per 100,000 people, respectively.1 Regarding thrombosis, it can occur in any tissue or organ’s microvessels, including those in the heart, lungs, kidneys, and other organs. The most common sites are the deep veins of the lower extremities and the cerebral artery circulation. In obstetric APS (OAPS), it can lead to premature birth, miscarriage, preeclampsia, premature rupture of membranes, and fetal distress, with severe cases posing a threat to life and having a significant impact on the patient’s health and quality of life.2–4
Thrombosis in APS was initially understood through the interference of aPLs with the regulation of the coagulation and fibrinolysis systems,5 but this was not comprehensive. With further research, aPLs are not only related to thrombosis but also closely associated with obstetric complications such as recurrent miscarriage (RSM). At present, the pathogenesis of APS is still unclear. Scholars have proposed various pathogenic mechanisms for this disease, including endothelial cell activation, complement activation and complement-mediated damage, platelet activation, imbalance of anticoagulation and fibrinolysis systems, immune cells (neutrophils, T cells, monocytes), and procoagulant effects of extracellular vesicles,1,5–8 leading to procoagulant and pro-inflammatory consequences, and ultimately resulting in thrombosis and OAPS. The 16th International Congress on Antiphospholipid Antibodies reviewed the treatment trends of APS and reported on the progress of its treatment, including the use of anticoagulant and antiplatelet drugs, adjuvant therapy, targeted therapy, complement inhibition in patients with APS.9 Although research on the pathogenesis and treatment has achieved gratifying results in recent years, there is still a lack of a clear pathogenesis and more precise targeted treatment at present.
Mammalian target of rapamycin (mTOR) plays a significant role in cell metabolism, growth and survival. Studies in animal models have shown that overactivation of mTOR signaling pathway is associated with autoimmune diseases, such as psoriasis, etc.10,11 In 2014, the role of mTOR signaling pathway activation in APS was first described.12 Although research is still ongoing, the mechanism of its action in the occurrence and development of APS has not been fully elucidated. In addition, the excessive activation of mTOR signaling pathway is associated with immune disorders, such as the activation of immune cells, platelets and endothelial cells, and the secretion of pro-inflammatory and pro-coagulant factors,13 which disrupts the balance of the immune system and is closely related to APS thrombosis or OAPS, but further exploration is still needed. Therefore, the purpose of this article is to review the role of mTOR in APS, thereby further understanding the potential pathogenesis of APS and seeking potential therapeutic targets for the treatment of APS.
mTOR
mTOR is a serine/threonine kinase and belongs to the phosphatidylinositol 3-kinase (PI3K) -related kinase family.14 It is a highly conserved protein, composed of 2549 amino acids, with a molecular weight of approximately 289 kD.15 It is an important component of the PI3K/protein kinase B (AKT)/mTOR pathway and one of the most significant pathways.16 The protein kinase complex formed by mTOR includes mTORC1 and mTORC2.17 The main function of mTORC1 is to sense nutrients and growth factor signals to promote anabolic metabolism, such as protein synthesis and lipid synthesis, and inhibit catabolic pathways, such as lysosomal biosynthesis and autophagy, to create nutrients through autophagy to promote cell survival.18 Under favorable conditions, mTORC1 can initiate the biosynthetic pathway for cell growth and proliferation by phosphorylating the ribosomal protein S6 kinase (S6K) and eukaryotic initiation factor 4E.19 The main functions of mTORC2 are closely related to cell survival, metabolism and cytoskeletal organization.18 Under normal circumstances, the balance of mTOR plays a significant role in immune balance. Research has found that a reduction in its activation may lead to immune deficiency, impaired development, proliferation and migration of lymphocytes, while an increase in activation is associated with autoimmune diseases.10,20
Importance of mTOR Signaling Pathway in Immune Function
The mTOR signaling pathway plays a crucial regulatory role in the differentiation, activation and functional characteristics of immune cells. Therefore, the role of the mTOR signaling pathway in the development and function of immune cells after activation has received extensive attention. For instance, in monocytes and macrophages, mTORC1 plays a role in regulating the balance between pro-inflammatory and anti-inflammatory effects. The protein encoded by the tuberous sclerosis 2 gene (TSC2) is an upstream inhibitory factor of mTORC1. The loss of TSC2 in monocytes leads to the activation of the mTORC1 signaling pathway, which not only enhances the production of IL-10 and reduces the production of pro-inflammatory cytokines such as IL-6, IL-12, IL-23 and TNF-α in response to lipopolysaccharide stimulation,21,22 but also may induce the transformation of monocytes into other phenotypes of macrophages. In macrophages, continuous activation of mTORC1 can promote their own hypertrophy and proliferation, which is related to the formation of granulomas in the body.23 This indicates that the excessive activation of the mTORC1 signaling pathway in macrophages is associated with the occurrence and development of immune pathology. In adaptive immunity, activated dendritic cells can typically present antigens through T-cell receptors, thereby initiating the activation, proliferation and differentiation of CD4+ and CD8+ T cells. The high level of mTOR is associated with an active metabolic state, while the low level of mTOR is related to the quiescent state of T cells and B cells.24 During the initial CD4+ activation process, the high activity of mTOR in T cells supports the expression of key transcription factors necessary for their differentiation into T helper (Th) 1, Th2 and Th17 cells. When mTOR signaling is absent, T cells differentiate into cells that produce regulatory T (Treg) cells and T follicular helper (Tfh) cells.24 Study has shown that rapamycin inhibits mTOR or the deletion of the mTOR gene can suppress the differentiation of Th1, Th2 and Th17 effector cells, while promoting the differentiation of Treg cells.25 This is related to the increase in the expression of forkhead box P3 (FOXP3) after the mTOR signaling pathway is inhibited. Study has shown that when the mTOR gene is knocked out in mice, they exhibit an excessive number of FOXP3+ Treg cells, and these cells outnumber Th1 and Th2 cells.26 However, in another study, when the TORC1 gene was specifically knocked out in FOXP3+ Treg cells, it led to a severe loss of Treg inhibitory activity in the body and the development of a fatal early-onset inflammatory disease. Notably, the activation of TORC1 is still necessary for the function of Treg cells.27 The germinal centers of Tfh cells are activated, thereby generating antibodies.24 Additionally, in T cells stimulated by cytokines, mTOR regulates the transition of the cell cycle from the G1 phase to the S phase, thereby making mTOR more active.24 For instance, IL-12, after binding to the receptors on T cells, activates the mTOR signaling pathway in T cells, and then drives the development of Th1 cells by stimulating the production of IFN-γ.28 Regarding the role of the mTOR signaling pathway in B cells, for instance, its function in B cell development, differentiation and function is less studied compared to that in T cells. In mouse models, when the mTOR post-transcriptional process is disrupted, there is a partial blockage of B cell development from large pre-B cells to small pre-B cells, and the proliferation response in the B cell mitotic signal is impaired.29 At the same time, it can also reduce the survival, proliferation, oxidative and glycolytic metabolic capabilities of pre-B cells, causing an obstruction at the early pre-B cell stage, accompanied by the loss of immature and mature peripheral B cells, impairment of plasma cell function, and thus disrupting the early stage of antibody production.30,31 The above indicates that the mTOR signaling pathway plays a crucial role in maintaining immune balance by regulating immune cells. If it is over-activated, it may trigger the occurrence of autoimmune diseases. In Table 1, we summarize the effects of excessive activation of the mTOR signaling pathway on immune cells.
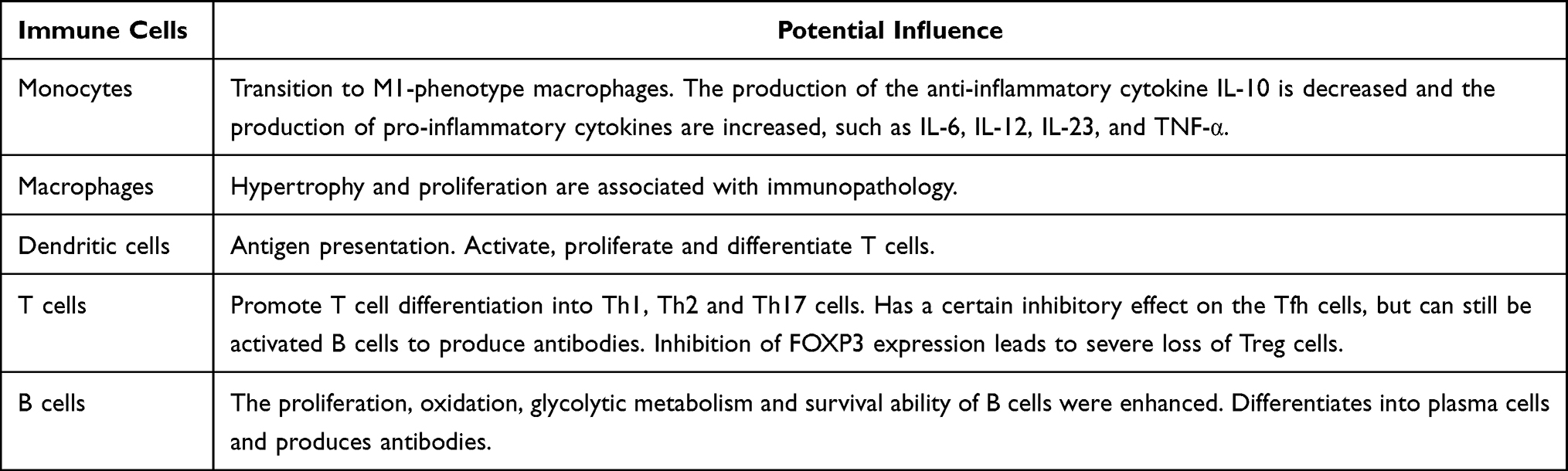 |
Table 1 Influence of mTOR Excessive Activation on Various Immune Cells |
Potential Mechanisms of mTOR Signaling Pathway in the Pathogenesis of OAPS
Firstly, we understand the role of the mTOR signaling pathway in normal pregnancy. Although there are some differences in the biological functions of mTORC1 and mTORC2, they are of great significance in normal pregnancy. For instance, the mTORC2 signaling pathway mainly participates in the regulation of placental inflammation, the transport of trace nutrients, and angiogenesis.32 Study has shown that inhibiting the mTORC2 signaling pathway can down-regulate the expression of cell division cycle protein 42, leading to reduced placental amino acid transfer and slower fetal growth, which is associated with intrauterine growth restriction (IUGR).33 The mTORC1 signaling pathway can also regulate the uptake of folic acid by human placental trophoblast cells, providing a direct link between placental function, gene methylation, and fetal programming.34 Maternal inflammatory conditions are associated with RSM. Study has shown that inhibiting the mTORC1 signaling pathway can suppress the activation of inflammasomes and improve complications related to reproductive immunity.35 In addition, the mTOR signaling pathway plays an active role in normal pregnancy by regulating autophagy, as autophagy helps regulate chorionization, trophoblast invasion, and immune action at the maternal-fetal interface.36,37 Usually, during normal pregnancy, the mother can enhance autophagy through self-regulation and promote the process of decidualization, which is beneficial for the growth of embryo implantation.38 However, during the decidualization process, there is a significant consumption of energy. In this situation, the maternal-fetal interface experiences hypoxia and low nutritional status. This environment can further stimulate autophagy, rescue ROS-induced cell apoptosis, promote trophoblast invasion, spiral artery remodeling, and placental angiogenesis.38 However, excessive autophagy also brings certain negative effects. The mTOR signaling pathway also regulates the migration and invasion functions of extravillous trophoblast cells (EVT) by modulating autophagy, balancing energy sources and responding to nutritional/oxygen stress, thereby contributing to placental development.39 Furthermore, the mTOR signaling pathway can regulate the infiltration, enrichment and functional regulation of decidual immune cells through autophagy.37 For instance, inhibiting the excessive activation of the mTOR signaling pathway and enhancing autophagy can reduce the secretion of insulin-like growth factor 2 in the trophoblast, decrease the expression of killing receptors such as CD16, natural killer (NK) G2D, NKP30 and NKP46, thereby inhibiting the toxicity of decidual NK (dNK) cells and reducing the miscarriage rate.36 At the same time, it promotes the polarization of decidual macrophages towards anti-inflammatory M2-like macrophages, thereby reducing the loss of the fetus.40 Study has shown that if mTOR is blocked, decidual macrophages will polarize towards the M2 phenotype, which may promote the polarization of macrophages towards the pro-inflammatory M1 phenotype, leading to fetal rejection and increasing the probability of fetal loss.41 These immune cells facilitate maternal-fetal immune tolerance during pregnancy and play a crucial role in promoting endometrial decidualization, trophoblast invasion, placental development, vascular remodeling, and fetal growth.38,41,42
In APS, study has shown that anti-cardiolipin antibodies (aCL) can activate the mTOR signaling pathway.43 In this case, Tfh cells promote B cell activation and differentiate into plasma cells, thereby generating more pathogenic antibodies. These antibodies, after binding to self-antigens, can trigger complement activation, leading to the production of pro-inflammatory fragments C5a and C3a.44 When the C5a receptor on immune cells binds to C5a, it can trigger the release of pro-inflammatory mediators, attract and chemoattract a large number of monocytes to infiltrate the decidua, and possibly cause macrophages to polarize towards the pro-inflammatory M1 phenotype, resulting in an explosive inflammatory response. These inflammatory cells and inflammatory mediators act on the placenta, thereby triggering placental inflammation,45 ultimately leading to fetal damage. In addition, studies have shown that aPLs can promote the activation of mTOR signaling pathway and inhibit autophagy,46,47 which leads to pregnancy loss. Specifically, the activation of the mTOR signaling pathway can promote the nuclear translocation of retinoic acid receptor-related orphan receptor (ROR)γt, regulate the differentiation of Th17 cells, promote the differentiation and function of effector T cells, and inhibit the expansion of the Treg cell population.48,49 The reduction in the number of Treg cells is associated with insufficient EVT infiltration and poor spiral artery remodeling, which leads to unstable placental development and results in shallow placental implantation.50 At the same time, it can further cause an overly strong Th17 cell response, which can induce the activation of dNK cells and the secretion of pro-inflammatory cytokines (such as IFN-γ and TNF-α), perforin and granzymes, thereby causing damage to the target cells.51 This may damage the placental vascular endothelial cells, lead to the formation of placental microthrombi, and subsequently result in OAPS. Excessive Th17 cells and the cytokines they secrete can also cause neutrophils to be chemoattracted, leading to uncontrolled infiltration of neutrophils at the maternal-fetal interface.52 This may result in a decrease in essential angiogenic factors and vascular endothelial growth factors, ultimately leading to abnormal placental formation and fetal death. Study in mice have shown that an increase in the ratio of Th17/Treg cells can lead to the occurrence of miscarriage.49 After using mTOR signaling pathway inhibitors (rapamycin), the protein levels of mTOR and RORγt can be reduced, the expression of FOXP3 can be increased, and the imbalance between Treg and Th17 cells can be corrected.53 In patients with recurrent implantation failure (RIF), the ratio of Th17/Treg was elevated. Low-dose rapamycin treatment was administered to induce the expansion of Treg cells, thereby correcting the imbalance of Th17/Treg ratio by regulating the Th17/Treg axis, and improving the reproductive outcomes of the patients.54 In summary, in APS patients, aPLs activate the mTOR signaling pathway, leading to an imbalance in various immune cells, such as B cells, monocytes, dNK cells, Treg/Th17 cells and their related cytokines, as well as enhanced cytotoxicity of dNK cells and imbalance in T cell phenotypes. Through continuous amplification of the inflammatory response, the regulation of immune homeostasis at the maternal-fetal interface is disrupted. This not only inhibits the remodeling of uterine spiral arteries but also damages the placenta and reduces the number of fetal capillaries, leads to placental microthrombosis, and ultimately results in OAPS (Figure 1).
 |
Figure 1 Potential pathogenesis of mTOR signaling pathway activation in OAPS. In OAPS, when aPLs bind to various cells, they can activate the PI3K-AKT-mTOR pathway. Under the regulation of S6RP, this leads to the activation of multiple immune cells and a series of consequences, such as B cells producing more autoantibodies and activating the complement pathway. Th0 cells differentiate into Th1, Th2 and Th17 cells, inhibit Treg cells, and cause an imbalance between Treg and Th17 cells. Although it can partially inhibit Tfh cells, it further activates B cells, transforming monocytes into M1 type macrophages and secreting various inflammatory cytokines. At the same time, it increases the expression of killing receptors such as CD16, NKG2D, NKP30 and NKP46, and enhances the toxicity of dNK cells. Taking all these factors into account, it can cause an imbalance in the immune response at the maternal-fetal interface, leading to placental inflammation. The damage to the placental vascular endothelial cells inhibits the remodeling of the uterine spiral arteries, interferes with the normal EVT invasion, and ultimately results in abnormal placental formation and fetal absorption. Abbreviations: OAPS, obstetric antiphospholipid syndrome; PI3K, phosphatidylinositol 3-kinase; mTOR, mammalian target of rapamycin; Akt, protein kinase B; S6RP, S6 kinase ribosomal protein; RORγt, retinoic acid receptor-related orphan receptor γt; P, phosphorylation; IL, interleukin; TNF, tumor necrosis factor; IFN, interferon; aPL, antiphospholipid antibody; Th, T helper; EVT, extra-villous trophoblast; Treg, regulatory T; NK, natural killer; dNK, decidual natural killer. |
Potential Mechanisms of mTOR Signaling Pathway in Thrombosis in APS
In APS, the mTOR signaling pathway is closely related to thrombus formation. Firstly, under normal circumstances, when the body is injured, tissue factor (TF) can activate factor VII to initiate intracellular coagulation, promoting the release of thrombin, converting fibrinogen into fibrin, and by activating other coagulation factors (V, VIII, and XI), maintaining the stability of the fibrin clot and further amplifying the coagulation process,55 thereby playing a role in hemostasis. The study has found that anti-β2GPI antibodies can bind to monocytes and activate the mTOR signaling pathway within monocytes, thereby activating the cells themselves. On the other hand, it can increase the expression and activity of tissue factor (TF) and IL-8.56 TF is the main initiating factor for thrombosis, which also indicates that the activation of the mTOR signaling pathway in monocytes is closely related to its pro-inflammatory and pro-coagulant phenotypes.57 APLs can trigger an inflammatory response in vascular endothelial cells. Additionally, due to changes in hemodynamics, the mTOR signaling pathway activates monocytes, leading to an increase in the expression of adhesion molecules. Monocytes can adhere to endothelial cells through adhesion molecules, causing chronic and persistent inflammation in the endothelial cells and resulting in their damage. At the same time, the activation of the mTOR signaling pathway in monocytes can transform them into pro-inflammatory macrophages, generating a large amount of inflammatory factors and TF.58 These inflammatory cytokines (such as IFN-γ and TNF-α) cause T cells to differentiate into Th1 type cells,58 further aggravating endothelial cell damage. Additionally, the activation of the mTOR signaling pathway in T cells promotes the nuclear translocation of RORγt, while at this time the expression of FOXP3 decreases. As a result, the number of Treg cells is low, unable to control the proliferation of IL-17+ T cells and generating excessive IL-17. IL-17 can induce endothelial cells to secrete pro-inflammatory cytokines and chemokines, which are related to the promotion of leukocyte recruitment and endothelial cell invasion.59,60 The activation of the mTOR signaling pathway in monocytes/macrophages and T cells can trigger endothelial cell damage, which may not only cause arterial/venous thrombosis but even induce thrombotic microangiopathy.61–63
Anti-β2GPI antibodies can also cause platelet activation through mTORC1-mediated Fcγ receptor IIa.64 Platelet activation can lead to an increase in thromboxane A2 production, an increase in glycoprotein IIb/IIIa expression, and an increase in platelet-derived microparticles formation, thereby triggering thrombosis.7 In another study, anti-β2GP1 antibodies enhanced platelet activation by activating the platelet Akt/mTORC2 signaling pathway, which was consistent with the excessive activation of APS platelets upon stimulation, manifested as an increase in mRNA levels related to platelet activation, thereby inducing thrombosis.65 After activation, platelets can express P-selectin on their own surface and bind to receptors on monocytes, forming a complex.66,67 This complex, under the mediation of chemokines, increases the binding of platelet factor 4 to the surface of monocytes and causes the platelets to accumulate on the endothelial cells,68,69 which is related to thrombosis. In addition, in an in vitro model, IgG isolated from the blood of female APS patients with previous OAPS and vascular thrombosis was able to activate mTOR in human umbilical vein endothelial cells.46 Regarding the mTOR signaling pathway in aPLs-positive patients, there have been studies reporting that mTOR is involved in the development of aPLs endothelial cell damage, dysfunction, and proliferation.70 When the above-mentioned damage occurs to the vascular endothelial cells, their collagen is exposed in the circulation and, under the action of monocytes, forms platelet-rich thrombi.71 Meanwhile, after activation by various receptors and ligands between endothelial cells, monocytes, and platelets, they can reduce the expression of anticoagulant molecules and adhesion molecules (such as E-selectin, etc.), thereby leading to an increase in vascular endothelial cell inflammatory response and promoting coagulation, causing imbalance in the anticoagulation and fibrinolysis systems, and making the fibrin network denser, with lower permeability and reduced susceptibility to dissolution.7,71 From the above, it can be seen that the activation of the mTOR signaling pathway in APS can trigger platelet activation, imbalance of the anticoagulation and fibrinolysis systems, and an increase in the level of fibrinogen in the circulation, which may be related to the formation of arterial and venous thrombosis.72 In severe cases, microvascular lesions can occur, affecting occlusive vascular lesions of small blood vessels in multiple organs such as the kidneys, lungs, brain, heart and liver, resulting in catastrophic APS.7 We have summarized the potential role of mTOR signaling pathway activation in thrombosis formation in APS in Figure 2 as discussed above.
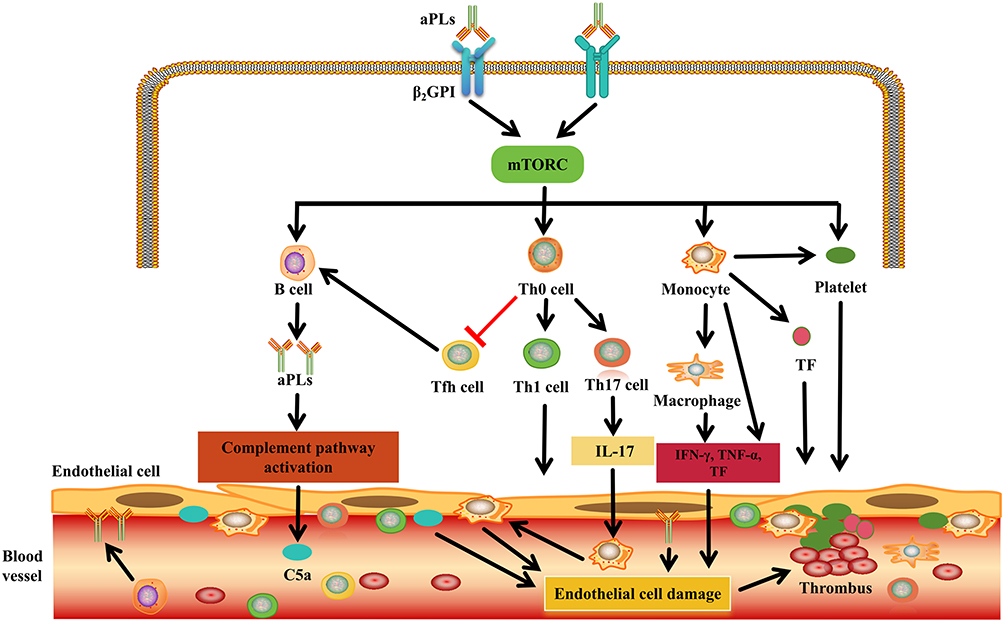 |
Figure 2 Potential pathogenesis of mTOR signaling pathway activation in thrombosis in APS. In APS, anti-β2GPI antibody binds to cells through β2GPI protein and activates mTOR signaling pathway in a variety of immune cells, including B cells, T cells, monocytes and platelets. The activation of these cells is related to the formation of autoantibodies, the activation of the complement pathway to produce the pro-inflammatory fragment C5a, and the secretion of pro-inflammatory cytokines and pro-coagulation factors. Under the combined effect of these factors, endothelial cells can be damaged, and monocytes, platelets and tissue factors gather at the damaged site, further leading to increased expression of adhesion molecules and enhanced inflammatory response, ultimately resulting in the formation of a thrombus. mTOR, mammalian target of rapamycin. Abbreviations: IL, interleukin; TNF, tumor necrosis factor; IFN, interferon; aPL, antiphospholipid antibody; Th, T helper; β2GPI, β2-glycoprotein I; Th, T helper; C5a, Complement 5a; TF, Tissue factor. |
The Potential of Targeting mTOR Signaling Pathway in the Treatment of APS
The pathogenesis of APS is rather complex, and the specific treatment plan is still unclear. The current strategy emphasizes anticoagulation as the foundation of treatment, and the standard protocol usually includes vitamin K antagonists and low-molecular-weight heparin. It is necessary to monitor the international normalized ratio and assess the risk of bleeding.73 Other treatments include direct oral anticoagulants (dabigatran), or direct factor Xa inhibitors (rivaroxaban, edoxaban or apixaban), and low-dose aspirin (LDA).73 Symptomatic treatment includes hydroxychloroquine (HCQ), statins and vitamin D.73 Targeted treatments include rituximab, belimumab, anti-TNF drugs, complement inhibitors, etc.9,73 In the context of OAPS, the existing evidence supports the use of LDA and heparin to reduce the miscarriage rate, and HCQ is of great significance in increasing the live birth rate and reducing preterm birth.74 Although these drugs have achieved certain therapeutic effects in the aspects of APS thrombosis and OAPS, the specific pathogenesis of APS is not clear, and it is currently impossible to cure it completely. Therefore, the current demand for potential therapies is highlighted. In the above description, we have mentioned that the excessive activation of the mTOR signaling pathway in APS patients leads to immune disorders, which are related to the occurrence and development of APS. Eventually, it may result in OAPS or thrombosis, and even pose a threat to life. Therefore, targeting the mTOR signaling pathway is of vital importance for improving the outcomes of APS patients.
In vitro study has revealed that the mTOR signaling pathway is involved in the activation of platelets in APS.60 After treating with mTOR signaling pathway inhibitors (rapamycin or everolimus), the mTOR signaling pathway in monocytes and endothelial cells was inhibited, thereby suppressing platelet aggregation induced by aPLs and blocking the interaction between anti-β2GPI antibodies and the low-affinity Fcγ receptor IIa on the platelet surface, thereby preventing platelet activation.60 This is also closely related to the inhibition of thrombosis. In another study, anti-β2GPI antibodies activated monocytes by activating the mTOR signaling pathway and increased the expression of TF and IL-8.56 After co-culturing monocytes with the mTOR signaling pathway inhibitor rapamycin (100 nM), their expression was downregulated, indicating that the inhibition of the mTOR signaling pathway may be beneficial for preventing and treating inflammatory thrombosis in patients with APS mediated by aPLs.56 In addition, when anti-β 2GPI antibodies are co-cultured with human umbilical vein endothelial cells, the β2GPI/anti-β2GPI complex can activate the PI3K/AKT/mTOR signaling pathway in endothelial cells and inhibit autophagy, inducing the expression of inflammatory cytokines, leading to inflammation, damage and dysfunction of endothelial cells.75 After rapamycin was added, the autophagy of endothelial cells was activated, which inhibited their apoptosis and downregulated the expression of pro-inflammatory factors IL-1β, IL-6 and intercellular adhesion molecule-1, thereby alleviating the damage of endothelial cells.75
In animal models, researchers injected the representative marker antibody against β2 GPI, which is characteristic of APS patients, into mice to create an APS mouse model and induce thrombosis in the carotid arteries of the APS mice.76 After using the mTOR signaling pathway inhibitor (rapalink-1), the levels of the anti-β2GPI antibody and aCL decreased, while the infiltration of macrophages was reduced and autophagy was enhanced, significantly preventing the occurrence and progression of the thrombus plaque area.76 In OAPS, the activation of the mTOR signaling pathway plays a crucial role. It may weaken the invasive ability of trophoblast cells, disrupt spiral artery remodeling and cause abnormal decidualization of the endometrium, all of which can lead to miscarriage.77 Targeting the mTOR signaling pathway can regulate the balance between immune cells, autophagy and cytokines in experimental models, thereby facilitating pregnancy.49 In the rat model of pregnancy loss induced by aCL-IgG isolated from the serum of APS patients, they exhibited decreased fetal weight and increased fetal absorption rate.78 After treatment with hyperoside (40 mg/kg) targeting the mTOR signaling pathway, the highly activated mTOR and S6K in the mouse placenta were inhibited, enhancing autophagy and anti-inflammatory levels, reducing IL-1β and IL-8 levels, and improving the pregnancy outcome (increased fetal weight and decreased absorption rate).78 Similarly, in a mouse model with a predisposition to miscarriage, treatment with the mTOR signaling pathway inhibitor, epimedium glycoside (50 mg/kg), led to an increase in the expansion of Treg cells, a reduction in Th1 cells, and a decrease in the expression of pro-inflammatory factors, thereby reducing placental inflammation and preventing spontaneous miscarriage in the mice.79
In terms of clinical application, it mainly focuses on the treatment of thrombosis. Currently, there is still a lack of relevant research in this area. Mexico reported a case of a patient with APS and myocardial infarction who underwent stent implantation during hospitalization. After the operation, the patient was treated with aspirin, clopidogrel, and warfarin and was discharged in good condition. After the operation, the patient experienced angina pectoris twice, and coronary angiography showed varying degrees of internal occlusion. Subsequently, stents coated with sirolimus were implanted, and the patient remained asymptomatic. Repeated coronary angiography showed no stent stenosis, thrombosis, or intimal hyperplasia.80 In another case, it was also found that APS was combined with coronary microcirculation obstruction. Through histological examination of endomyocardial biopsy, it was confirmed that there was coronary obstructive microvascular disease. After three intravenous injections of 1000mg methylprednisolone, this patient began to receive sirolimus (1mg per day) treatment. The clinical condition improved and during the 12-month follow-up visit, the patient reported no discomfort.81 The activation of the mTOR signaling pathway is associated with vascular damage, especially in the kidneys. A case of refractory lupus nephritis secondary to APS was reported in China. The renal biopsy met the WHO III grade criteria for lupus nephritis and was accompanied by acute tubular injury. Immunofluorescence confirmed the activation of the mTOR signaling pathway. After treatment with the mTOR signaling pathway inhibitor sirolimus for 6 months, the patient achieved complete remission and maintained a disease-free remission for the next 3.5 years.82 In addition, the endothelial cells of the renal vessels in APS patients have activated the mTORC signaling pathway. After using sirolimus, the renal function of aPL-positive kidney transplant recipients was protected, reducing the proliferation of blood vessels and preventing recurrence.70 In the study, 10 patients received sirolimus treatment, among whom 7 (70%) had normal kidney graft function 144 months after transplantation, while only 3 (11%) of the 27 untreated patients did.70 The authors also found that the activation of the mTOR signaling pathway was also observed in the autopsy vessels of patients with catastrophic APS.70
Through the above retrospective summary of studies from in vitro research to animal experiments and then to clinical applications, it has been demonstrated that targeting the mTOR signaling pathway has significant benefits in the treatment of APS, especially in the aspect of thrombosis. However, there are also some limitations regarding the efficacy of targeting the mTOR signaling pathway for patients with ASP at present. Firstly, APS is a heterogeneous disease. Currently, the pathogenesis of the mTOR signaling pathway in APS has not discussed the heterogeneity (such as triple aPL positive type and single aPL positive type; primary and secondary APS), and there is a lack of stratification of mechanisms/treatment significance based on disease phenotypes. Secondly, in the current research, more emphasis is placed on preclinical studies (in vitro studies and animal experiments), while clinical applications are mainly in individual cases. Preclinical studies may differ from those in humans. Therefore, we hope that in the future, there will be more research to explore large-scale prospective studies to investigate the efficacy of mTOR signaling pathway inhibitors in treating patients with APS, both in terms of thrombosis and OAPS. At the same time, this will be beneficial for us to assess its safety and efficacy, further supporting our conclusion and making our conclusion more universal.
Potential Adverse Effects of Targeting mTOR Signaling Pathway
With the increasing understanding of the disease, mTOR signaling pathway inhibitor therapy is more and more widely used. Currently, they have been approved for use in immunosuppressive therapy in cancer patients and for preventing rejection reactions after allogeneic organ transplantation.83 There are also some diseases that are activated through the mTOR signaling pathway, such as inflammatory bowel disease, renal cell carcinoma, prevention of Alzheimer’s disease, delaying aging, and prevention of other age-related diseases.38,84–86 It is also considered that mTOR signaling pathway inhibitors can be used for treatment. In the field of rheumatic diseases, in systemic lupus erythematosus, rheumatoid arthritis, ankylosing spondylitis, psoriatic arthritis, systemic sclerosis and osteoarthritis, with the further study of the pathogenesis, it has been found that the activation of the mTOR signaling pathway plays an important role in these diseases.87 Therefore, the mTOR signaling pathway may become a potential favorable therapeutic target for this autoimmune disease, and it is expected to extend the life expectancy by blocking the mTOR signaling pathway. Although there are potential advantages, blocking the mTOR signaling pathway may also cause some adverse reactions. We have summarized these adverse reactions in Table 2. 88–103
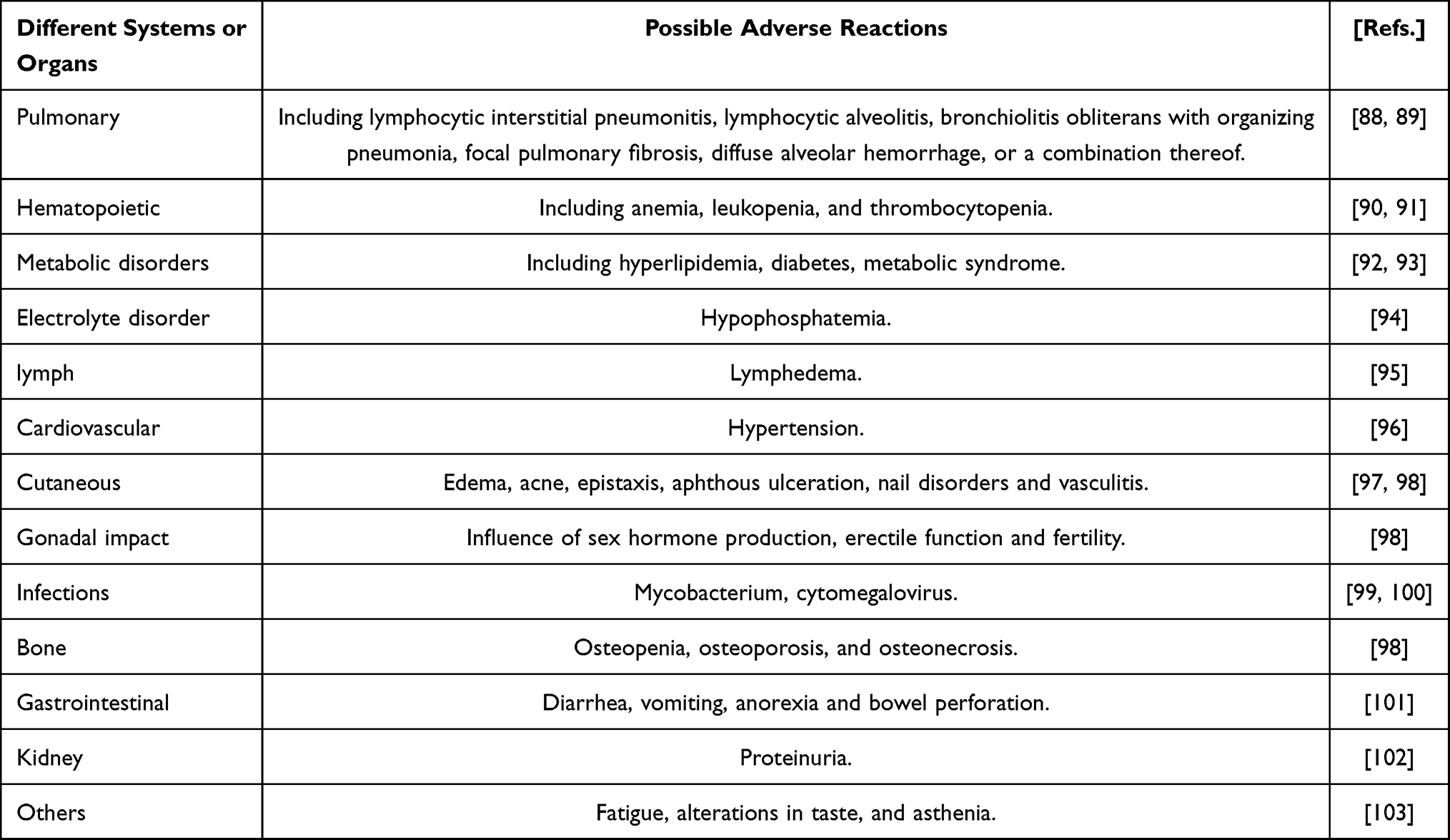 |
Table 2 Possible Adverse Reactions of mTOR Inhibition |
Conclusion
Thrombosis and OAPS caused by APS not only seriously affect the quality of life of patients, but also increase their economic burden. However, the specific pathogenesis behind these conditions remains unclear. The immune disorder caused by activation of the mTOR signaling pathway is a potentially important factor in the pathogenesis of APS thrombosis and OAPS. Targeting the mTOR signaling pathway plays a crucial role in regulating immune balance. Therefore, it is a promising and beneficial drug in the treatment of APS, but attention should also be paid to the related adverse reactions. However, at present, there are still insufficient relevant clinical studies on targeted mTOR signaling pathway therapy in APS. Most of the research focuses on preclinical studies. We expect that in the future, there will be more large-scale prospective clinical studies to further confirm the efficacy and deeper mechanisms (such as the pathogenesis related to the mTOR signaling pathway in the heterogeneity of APS). Especially stratifying the mechanisms according to disease phenotypes is of great significance for treatment. At the same time, exploring more about the optimal drug dosage and safety of mTOR signaling pathway inhibitors in APS is expected to further open up new treatment windows for APS.
Funding
No funding was received.
Disclosure
The authors declare that they have no competing interests.
References
1. Knight JS, Branch DW, Ortel TL. Antiphospholipid syndrome: advances in diagnosis, pathogenesis, and management. BMJ. 2023;380:e069717. doi:10.1136/bmj-2021-069717
2. Walter IJ, Klein Haneveld MJ, Lely AT, Bloemenkamp KWM, Limper M, Kooiman J. Pregnancy outcome predictors in antiphospholipid syndrome: a systematic review and meta-analysis. Autoimmun Rev. 2021;20(10):102901. doi:10.1016/j.autrev.2021.102901
3. Hubben A, McCrae KR. How to diagnose and manage antiphospholipid syndrome. Hematology Am Soc Hematol Educ Program. 2023;2023(1):606–13. doi:10.1182/hematology.2023000493
4. Lu C, Gao R, Qing P, et al. Single-cell transcriptome analyses reveal disturbed decidual homoeostasis in obstetric antiphospholipid syndrome. Ann Rheum Dis. 2024;83(5):624–637. doi:10.1136/ard-2023-224930
5. Chaturvedi S, Alluri R, McCrae KR. Extracellular Vesicles in the Antiphospholipid Syndrome. Semin Thromb Hemost. 2018;44(5):493–504. doi:10.1055/s-0037-1599081
6. Knight JS, Kanthi Y. Mechanisms of immunothrombosis and vasculopathy in antiphospholipid syndrome. Semin Immunopathol. 2022;44(3):347–362. doi:10.1007/s00281-022-00916-w
7. Stańczewska A, Szewczyk-Golec K, Hołyńska-Iwan I. Antiphospholipid Syndrome—Diagnostic and Methodologic Approach. Metabolites. 2025;15(8):500. doi:10.3390/metabo15080500
8. Patriarcheas V, Tsamos G, Vasdeki D, et al. Antiphospholipid Syndrome: a Comprehensive Clinical Review. J Clin Med. 2025;14(3):733. doi:10.3390/jcm14030733
9. Cohen H, Cuadrado MJ, Erkan D, et al. 16th International Congress on Antiphospholipid Antibodies Task Force Report on Antiphospholipid Syndrome Treatment Trends. Lupus. 2020;29(12):1571–1593. doi:10.1177/0961203320950461
10. Marnett LJ. Lipid peroxidation-DNA damage by malondialdehyde. Mutat Res. 1999;424(1–2):83–95. doi:10.1016/s0027-5107(99)00010-x
11. Roy T, Banang-Mbeumi S, Boateng ST, et al. Dual targeting of mTOR/IL-17A and autophagy by fisetin alleviates psoriasis-like skin inflammation. Front Immunol. 2023;13:1075804. doi:10.3389/fimmu.2022.1075804
12. Eikelboom JW, Weitz JI. The mTORC pathway in the antiphospholipid syndrome. N Engl J Med. 2014;371(4):369–371. doi:10.1056/NEJMe1406870
13. Ji L, Zhang Z, Perl A. The mTOR pathway in the antiphospholipid syndrome. Med Rev. 2023;3(6):511–513. doi:10.1515/mr-2023-0033
14. Mita MM, Mita A, Rowinsky EK. Mammalian target of rapamycin: a new molecular target for breast cancer. Clin Breast Cancer. 2003;4(2):126–137. doi:10.3816/cbc.2003.n.018
15. Keith CT, Schreiber SL. PIK-related kinases: DNA repair, recombination, and cell cycle checkpoints. Science. 1995;270(5233):50–51. doi:10.1126/science.270.5233.50
16. Yilmaz OH, Morrison SJ. The PI-3kinase pathway in hematopoietic stem cells and leukemia-initiating cells: a mechanistic difference between normal and cancer stem cells. Blood Cells Mol Dis. 2008;41(1):73–76. doi:10.1016/j.bcmd.2008.02.004
17. Yang M, Lu Y, Piao W, Jin H. The Translational Regulation in mTOR Pathway. Biomolecules. 2022;12(6):802. doi:10.3390/biom12060802
18. Kim YC, Guan KL. mTOR: a pharmacologic target for autophagy regulation. J Clin Invest. 2015;125(1):25–32. doi:10.1172/JCI73939
19. Jung S, Gámez-Díaz L, Proietti M, Grimbacher B. “Immune TOR-opathies,” a Novel Disease Entity in Clinical Immunology. Front Immunol. 2018;9:966. doi:10.3389/fimmu.2018.00966
20. Lai ZW, Kelly R, Winans T, et al. Sirolimus in patients with clinically active systemic lupus erythematosus resistant to, or intolerant of, conventional medications: a single-arm, open-label, Phase 1/2 trial. Lancet. 2018;391(10126):1186–1196. doi:10.1016/S0140-6736(18)30485-9
21. Weichhart T, Costantino G, Poglitsch M, et al. The TSC-mTOR signaling pathway regulates the innate inflammatory response. Immunity. 2008;29(4):565–577. doi:10.1016/j.immuni.2008.08.012
22. Corazza M, Oton-Gonzalez L, Scuderi V, et al. Tissue cytokine/chemokine profile in vulvar lichen sclerosus: an observational study on keratinocyte and fibroblast cultures. J Dermatol Sci. 2020;100(3):223–226. doi:10.1016/j.jdermsci.2020.09.006
23. Linke M, Pham HT, Katholnig K, et al. Chronic signaling via the metabolic checkpoint kinase mTORC1 induces macrophage granuloma formation and marks sarcoidosis progression. Nat Immunol. 2017;18(3):293–302. doi:10.1038/ni.3655
24. Panwar V, Singh A, Bhatt M, et al. Multifaceted role of mTOR (mammalian target of rapamycin) signaling pathway in human health and disease. Signal Transduct Target Ther. 2023;8(1):375. doi:10.1038/s41392-023-01608-z
25. Delgoffe GM, Kole TP, Cotter RJ, Powell JD. Enhanced interaction between Hsp90 and raptor regulates mTOR signaling upon T cell activation. Mol Immunol. 2009;46(13):2694–2698. doi:10.1016/j.molimm.2009.05.185
26. Delgoffe GM, Kole TP, Zheng Y, et al. The mTOR kinase differentially regulates effector and regulatory T cell lineage commitment. Immunity. 2009;30(6):832–844. doi:10.1016/j.immuni.2009.04.014
27. Zeng H, Yang K, Cloer C, Neale G, Vogel P, Chi H. mTORC1 couples immune signals and metabolic programming to establish T-cell function. Nature. 2013;499:485–490. doi:10.1038/nature12297
28. Kusaba H, Ghosh P, Derin R, et al. Interleukin-12-induced interferon-gamma production by human peripheral blood T cells is regulated by mammalian target of rapamycin (mTOR). J Biol Chem. 2005;280(2):1037–1043. doi:10.1074/jbc.M405204200
29. Zhang S, Readinger JA, DuBois W, et al. Constitutive reductions in mTOR alter cell size, immune cell development, and antibody production. Blood. 2011;117(4):1228–1238. doi:10.1182/blood-2010-05-287821
30. Iwata TN, Ramírez JA, Tsang M, et al. Conditional Disruption of Raptor Reveals an Essential Role for mTORC1 in B Cell Development, Survival, and Metabolism. J Immunol. 2016;197(6):2250–2260. doi:10.4049/jimmunol.1600492
31. Gaudette BT, Jones DD, Bortnick A, Argon Y, Allman D. mTORC1 coordinates an immediate unfolded protein response-related transcriptome in activated B cells preceding antibody secretion. Nat Commun. 2020;11(1):723. doi:10.1038/s41467-019-14032-1
32. Rosario FJ, Kelly AC, Gupta MB, Powell TL, Cox L, Jansson T. Mechanistic Target of Rapamycin Complex 2 Regulation of the Primary Human Trophoblast Cell Transcriptome. Front Cell Dev Biol. 2021;9:670980. doi:10.3389/fcell.2021.670980
33. Jansson T, Castillo-Castrejon M, Gupta MB, Powell TL, Rosario FJ. Down-regulation of placental Cdc42 and Rac1 links mTORC2 inhibition to decreased trophoblast amino acid transport in human intrauterine growth restriction. Clin Sci. 2020;134(1):53–70. doi:10.1042/CS20190794
34. Rosario FJ, Powell TL, Jansson T. Mechanistic target of rapamycin (mTOR) regulates trophoblast folate uptake by modulating the cell surface expression of FR-α and the RFC. Sci Rep. 2016;6:31705. doi:10.1038/srep31705
35. Lee S, Shin J, Kim JS, Shin J, Lee SK, Park HW. Targeting TBK1 Attenuates LPS-Induced NLRP3 Inflammasome Activation by Regulating of mTORC1 Pathways in Trophoblasts. Front Immunol. 2021;12:743700. doi:10.3389/fimmu.2021.743700
36. Tan HX, Yang SL, Li MQ, Wang HY. Autophagy suppression of trophoblast cells induces pregnancy loss by activating decidual NK cytotoxicity and inhibiting trophoblast invasion. Cell Commun Signal. 2020;18(1):73. doi:10.1186/s12964-020-00579-w
37. Goudarzi ST, Vousooghi N, Verdi J, Mehdizadeh A, Aslanian-Kalkhoran L, Yousefi M. Autophagy genes and signaling pathways in endometrial decidualization and pregnancy complications. J Reprod Immunol. 2024;163:104223. doi:10.1016/j.jri.2024.104223
38. Li MY, Shen HH, Cao XY, et al. Targeting a mTOR/autophagy axis: a double-edged sword of rapamycin in spontaneous miscarriage. Biomed Pharmacother. 2024;177:116976. doi:10.1016/j.biopha.2024.116976
39. Nakashima A, Yamanaka-Tatematsu M, Fujita N, et al. Impaired autophagy by soluble endoglin, under physiological hypoxia in early pregnant period, is involved in poor placentation in preeclampsia. Autophagy. 2013;9(3):303–316. doi:10.4161/auto.22927
40. Liu T, Wang L, Liang P, et al. USP19 suppresses inflammation and promotes M2-like macrophage polarization by manipulating NLRP3 function via autophagy. Cell Mol Immunol. 2021;18(10):2431–2442. doi:10.1038/s41423-020-00567-7
41. Rao VA, Kurian NK, Cytokines RKA. NK cells and regulatory T cell functions in normal pregnancy and reproductive failures. Am J Reprod Immunol. 2023;89(2):e13667. doi:10.1111/aji.13667
42. Zhang X, Li Y, Huang C, et al. The role of decidual natural killer cell-derived soluble factors in early pregnancy. Am J Reprod Immunol. 2021;86(5):e13477. doi:10.1111/aji.13477
43. Wei A, Xiao H, Xu G, et al. Hyperoside Protects Human Umbilical Vein Endothelial Cells Against Anticardiolipin Antibody-Induced Injury by Activating Autophagy. Front Pharmacol. 2020;11:762. doi:10.3389/fphar.2020.00762
44. Dijkstra DJ, Joeloemsingh JV, Bajema IM, Trouw LA. Complement activation and regulation in rheumatic disease. Semin Immunol. 2019;45:101339. doi:10.1016/j.smim.2019.101339
45. Niyonzima N, Rahman J, Kunz N, et al. Mitochondrial C5aR1 activity in macrophages controls IL-1β production underlying sterile inflammation. Sci Immunol. 2021;6(66):eabf2489. doi:10.1126/sciimmunol.abf2489
46. Rodríguez CM, Velásquez-Berrío M, Rúa C, et al. Antiphospholipid Antibodies From Women With Pregnancy Morbidity and Vascular Thrombosis Induce Endothelial Mitochondrial Dysfunction, mTOR Activation, and Autophagy. Front Physiol. 2021;12:706743. doi:10.3389/fphys.2021.706743
47. Mulla MJ, Weel IC, Potter JA, et al. Antiphospholipid Antibodies Inhibit Trophoblast Toll-Like Receptor and Inflammasome Negative Regulators. Arthritis Rheumatol. 2018;70(6):891–902. doi:10.1002/art.40416
48. Chang S, Yin T, He F, Ding J, Shang Y, Yang J. CaMK4 promotes abortion-related Th17 cell imbalance by activating AKT/mTOR signaling pathway. Am J Reprod Immunol. 2020;84(6):e13315. doi:10.1111/aji.13315
49. Chapman NM, Chi H. mTOR signaling, Tregs and immune modulation. Immunotherapy. 2014;6(12):1295–1311. doi:10.2217/imt.14.84
50. Minton K. Vitamin D shuts down T cell-mediated inflammation. Nat Rev Immunol. 2022;22(1):1. doi:10.1038/s41577-021-00663-3
51. Shields CA, McCalmon M, Ibrahim T, et al. Placental ischemia-stimulated T-helper 17 cells induce preeclampsia-associated cytolytic natural killer cells during pregnancy. Am J Physiol Regul Integr Comp Physiol. 2018;315(2):R336–R343. doi:10.1152/ajpregu.00061.2018
52. Wang W, Sung N, Gilman-Sachs A, Kwak-Kim J. T helper (Th) cell profiles in pregnancy and recurrent pregnancy losses: th1/Th2/Th9/Th17/Th22/Tfh cells. Front Immunol. 2020;11:2025. doi:10.3389/fimmu.2020.02025
53. Gao L, Dong Y, Lin R, Meng Y, Wu F, Jia L. The imbalance of Treg/Th17 cells induced by perinatal bisphenol A exposure is associated with activation of the PI3K/Akt/mTOR signaling pathway in male offspring mice. Food Chem Toxicol. 2020;137:111177. doi:10.1016/j.fct.2020.111177
54. Royster GD, Harris JC, Nelson A, et al. Rapamycin Corrects T Regulatory Cell Depletion and Improves Embryo Implantation and Live Birth Rates in a Murine Model. Reprod Sci. 2019;26(12):1545–1556. doi:10.1177/1933719119828110
55. Maugeri N, Brambilla M, Camera M, et al. Human polymorphonuclear leukocytes produce and express functional tissue factor upon stimulation. J Thromb Haemost. 2006;4(6):1323–1330. doi:10.1111/j.1538-7836.2006.01968.x
56. Xia L, Zhou H, Wang T, et al. Activation of mTOR is involved in anti-β2GPI/β2GPI-induced expression of tissue factor and IL-8 in monocytes. Thromb Res. 2017;157:103–110. doi:10.1016/j.thromres.2017.05.023
57. Salet DM, Bekkering S, Middeldorp S, van den Hoogen LL. Targeting thromboinflammation in antiphospholipid syndrome. J Thromb Haemost. 2023;21(4):744–757. doi:10.1016/j.jtha.2022.12.002
58. Tektonidou MG. Cardiovascular disease risk in antiphospholipid syndrome: thrombo-inflammation and atherothrombosis. J Autoimmun. 2022;128:102813. doi:10.1016/j.jaut.2022.102813
59. Benagiano M, Borghi MO, Romagnoli J, et al. Interleukin-17/Interleukin-21 and Interferon-γ producing T cells specific for β2 Glycoprotein I in atherosclerosis inflammation of systemic lupus erythematosus patients with antiphospholipid syndrome. Haematologica. 2019;104(12):2519–2527. doi:10.3324/haematol.2018.209536
60. Robert M, Miossec P, Hot A. The Th17 Pathway in Vascular Inflammation: culprit or Consort? Front Immunol. 2022;13:888763. doi:10.3389/fimmu.2022.888763
61. Drummer C, Saaoud F, Shao Y, et al. Trained Immunity and Reactivity of Macrophages and Endothelial Cells. Arterioscler Thromb Vasc Biol. 2021;41(3):1032–1046. doi:10.1161/ATVBAHA.120.315452
62. Lu MJ, Zhang JQ, Nie ZY, et al. Monocyte/macrophage-mediated venous thrombus resolution. Front Immunol. 2024;15:1429523. doi:10.3389/fimmu.2024.1429523
63. Sharma SB, Melvin WJ, Audu CO, et al. The histone methyltransferase MLL1/KMT2A in monocytes drives coronavirus-associated coagulopathy and inflammation. Blood. 2023;141(7):725–742. doi:10.1182/blood.2022015917
64. Hollerbach A, Müller-Calleja N, Ritter S, et al. Platelet Activation by Antiphospholipid Antibodies Depends on Epitope Specificity and is Prevented by mTOR Inhibitors. Thromb Haemost. 2019;119(7):1147–1153. doi:10.1055/s-0039-1685453
65. Tang Z, Shi H, Chen C, et al. Activation of Platelet mTORC2/Akt Pathway by Anti-β2GP1 Antibody Promotes Thrombosis in Antiphospholipid Syndrome. Arterioscler Thromb Vasc Biol. 2023;43(10):1818–1832. doi:10.1161/ATVBAHA.123.318978
66. Gawaz MP, Loftus JC, Bajt ML, Frojmovic MM, Plow EF, Ginsberg MH. Ligand bridging mediates integrin alpha IIb beta 3 (platelet GPIIB-IIIA) dependent homotypic and heterotypic cell-cell interactions. J Clin Invest. 1991;88(4):1128–1134. doi:10.1172/JCI115412
67. Silverstein RL, Asch AS, Nachman RL. Glycoprotein IV mediates thrombospondin-dependent platelet-monocyte and platelet-U937 cell adhesion. J Clin Invest. 1989;84(2):546–552. doi:10.1172/JCI114197
68. Han Z, Liu Q, Li H, et al. The role of monocytes in thrombotic diseases: a review. Front Cardiovasc Med. 2023;10:1113827. doi:10.3389/fcvm.2023.1113827
69. Moore KL, Stults NL, Diaz S, et al. Identification of a specific glycoprotein ligand for P-selectin (CD62) on myeloid cells. J Cell Biol. 1992;118(2):445–456. doi:10.1083/jcb.118.2.445
70. Canaud G, Bienaimé F, Tabarin F, et al. Inhibition of the mTORC pathway in the antiphospholipid syndrome. N Engl J Med. 2014;371(4):303–312. doi:10.1056/NEJMoa1312890
71. Álvarez D, Morales-Prieto DM, Cadavid ÁP. Interaction between endothelial cell-derived extracellular vesicles and monocytes: a potential link between vascular thrombosis and pregnancy-related morbidity in antiphospholipid syndrome. Autoimmun Rev. 2023;22(4):103274. doi:10.1016/j.autrev.2023.103274
72. Hur WS, Kawano T, Mwiza JMN, et al. Mice expressing nonpolymerizable fibrinogen have reduced arterial and venous thrombosis with preserved hemostasis. Blood. 2024;143(2):105–117. doi:10.1182/blood.2023020805
73. Zhu QN, Qi XB, Ren SW, et al. Novel advances on pathophysiological mechanisms, clinical manifestations, and treatment of antiphospholipid syndrome. Front Immunol. 2025;16:1639065. doi:10.3389/fimmu.2025.1639065
74. Murvai VR, Galiș R, Panaitescu A, et al. Antiphospholipid syndrome in pregnancy: a comprehensive literature review. BMC Pregnancy Childbirth. 2025;25(1):337. doi:10.1186/s12884-025-07471-w
75. Zhang G, He C, Wu Q, et al. Impaired Autophagy Induced by oxLDL/β2GPI/anti-β2GPI Complex through PI3K/AKT/mTOR and eNOS Signaling Pathways Contributes to Endothelial Cell Dysfunction. Oxid Med Cell Longev. 2021;2021:6662225. doi:10.1155/2021/6662225
76. Mu F, Jiang Y, Ao F, Wu H, You Q, Chen Z. RapaLink-1 plays an antithrombotic role in antiphospholipid syndrome by improving autophagy both in vivo and vitro. Biochem Biophys Res Commun. 2020;525(2):384–391. doi:10.1016/j.bbrc.2020.02.084
77. Lv S, Wang N, Lv H, et al. The Attenuation of Trophoblast Invasion Caused by the Downregulation of EZH2 Is Involved in the Pathogenesis of Human Recurrent Miscarriage. Mol Ther Nucleic Acids. 2019;14:377–387. doi:10.1016/j.omtn.2018.12.011
78. Wei A, Song Y, Ni T, et al. Hyperoside attenuates pregnancy loss through activating autophagy and suppressing inflammation in a rat model. Life Sci. 2020;254:117735. doi:10.1016/j.lfs.2020.117735
79. Peng F, Han Z, Chen H, Zhang Q, Liu C, Liang X. The Effects of Treatment with Icariin on Immune Tolerance in the Recurrent Spontaneous Abortion Mice. Reprod Sci. 2023;30(9):2794–2804. doi:10.1007/s43032-023-01185-0
80. Mora-Ramírez M, González-Pacheco H, Amezcua-Guerra LM. Stents Coated With Mammalian Target of Rapamycin Inhibitors (mTOR) Appear to Be the Best Choice in Patients With Antiphospholipid Syndrome and Myocardial Infarction. J Clin Rheumatol. 2016;22(5):281. doi:10.1097/RHU.0000000000000409
81. Sartorelli S, De Luca G, Campochiaro C, et al. Successful use of sirolimus in a patient with cardiac microangiopathy in primary antiphospholipid syndrome. Scand J Rheumatol. 2019;48(6):515–516. doi:10.1080/03009742.2019.1574022
82. Zhang D, Sun F, Ye S. Successful treatment of sirolimus in a Chinese patient with refractory LN and APS: a case report. Ther Adv Musculoskelet Dis. 2022;14:1759720X221079253. doi:10.1177/1759720X221079253
83. Wojciechowski D, Wiseman A. Long-Term Immunosuppression Management: opportunities and Uncertainties. Clin J Am Soc Nephrol. 2021;16(8):1264–1271. doi:10.2215/CJN.15040920
84. Afzal O, Altamimi ASA, Mubeen B, et al. mTOR as a Potential Target for the Treatment of Microbial Infections, Inflammatory Bowel Diseases, and Colorectal Cancer. Int J Mol Sci. 2022;23(20):12470. doi:10.3390/ijms232012470
85. Chauhan A, Semwal DK, Mishra SP, Goyal S, Marathe R, Semwal RB. Combination of mTOR and MAPK Inhibitors-A Potential Way to Treat Renal Cell Carcinoma. Med Sci. 2016;4(4):16. doi:10.3390/medsci4040016
86. Davoody S, Asgari Taei A, Khodabakhsh P, Dargahi L. mTOR signaling and Alzheimer’s disease: what we know and where we are? CNS Neurosci Ther. 2024;30(4):e14463. doi:10.1111/cns.14463
87. Perl A. Activation of mTOR (mechanistic target of rapamycin) in rheumatic diseases. Nat Rev Rheumatol. 2016;12(3):169–182. doi:10.1038/nrrheum.2015.172
88. Champion L, Stern M, Israël-Biet D, et al. Brief communication: sirolimus-associated pneumonitis: 24 cases in renal transplant recipients. Ann Intern Med. 2006;144(7):505–509. doi:10.7326/0003-4819-144-7-200604040-00009
89. Eren MF, Ay Eren A, Sayan M, et al. The Impact of Everolimus and Radiation Therapy on Pulmonary Fibrosis. Medicina. 2020;56(7):348. doi:10.3390/medicina56070348
90. Augustine JJ, Knauss TC, Schulak JA, Bodziak KA, Siegel C, Hricik DE. Comparative effects of sirolimus and mycophenolate mofetil on erythropoiesis in kidney transplant patients. Am J Transplant. 2004;4(12):2001–2006. doi:10.1111/j.1600-6143.2004.00612.x
91. Sofroniadou S, Goldsmith D. Mammalian target of rapamycin (mTOR) inhibitors: potential uses and a review of haematological adverse effects. Drug Saf. 2011;34(2):97–115. doi:10.2165/11585040-000000000-00000
92. D’Elia JA, Weinrauch LA. Hyperglycemia and Hyperlipidemia with Kidney or Liver Transplantation: a Review. Biology. 2023;12(9):1185. doi:10.3390/biology12091185
93. Granata S, Mercuri S, Troise D, Gesualdo L, Stallone G, Zaza G. mTOR-inhibitors and post-transplant diabetes mellitus: a link still debated in kidney transplantation. Front Med. 2023;10:1168967. doi:10.3389/fmed.2023.1168967
94. Jebali M, Elaidi R, Brizard M, et al. Biological toxicities as surrogate markers of efficacy in patients treated with mTOR inhibitors for metastatic renal cell carcinoma. BMC Cancer. 2017;17(1):27. doi:10.1186/s12885-016-2993-7
95. Ersoy A, Koca N. Everolimus-induced lymphedema in a renal transplant recipient: a case report. Exp Clin Transplant. 2012;10(3):296–298. doi:10.6002/ect.2011.0139
96. Reis F, Parada B, Teixeira de Lemos E, et al. Hypertension induced by immunosuppressive drugs: a comparative analysis between sirolimus and cyclosporine. Transplant Proc. 2009;41(3):868–873. doi:10.1016/j.transproceed.2009.02.005
97. Mahé E, Morelon E, Lechaton S, et al. Cutaneous adverse events in renal transplant recipients receiving sirolimus-based therapy. Transplantation. 2005;79(4):476–482. doi:10.1097/01.tp.0000151630.25127.3a
98. Zaza G, Tomei P, Ria P, Granata S, Boschiero L, Lupo A. Systemic and nonrenal adverse effects occurring in renal transplant patients treated with mTOR inhibitors. Clin Dev Immunol. 2013;2013:403280. doi:10.1155/2013/403280
99. Singh P, Subbian S. Harnessing the mTOR Pathway for Tuberculosis Treatment. Front Microbiol. 2018;9:70. doi:10.3389/fmicb.2018.00070
100. Mallat SG, Tanios BY, Itani HS, et al. CMV and BKPyV Infections in Renal Transplant Recipients Receiving an mTOR Inhibitor-Based Regimen Versus a CNI-Based Regimen: a Systematic Review and Meta-Analysis of Randomized, Controlled Trials. Clin J Am Soc Nephrol. 2017;12(8):1321–1336. doi:10.2215/CJN.13221216
101. Lebranchu Y, Thierry A, Toupance O, et al. Efficacy on renal function of early conversion from cyclosporine to sirolimus 3 months after renal transplantation: concept study. Am J Transplant. 2009;9(5):1115–1123. doi:10.1111/j.1600-6143.2009.02615.x
102. Mjörnstedt L, Schwartz Sørensen S, von Zur Mühlen B, et al. Renal function three years after early conversion from a calcineurin inhibitor to everolimus: results from a randomized trial in kidney transplantation. Transpl Int. 2015;28(1):42–51. doi:10.1111/tri.12437
103. Hudes G, Carducci M, Tomczak P, et al. Temsirolimus, interferon alfa, or both for advanced renal-cell carcinoma. N Engl J Med. 2007;356(22):2271–2281. doi:10.1056/NEJMoa066838
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Coexistence of Congenital Dysfibrinogenemia and Antiphospholipid Syndrome in Pregnancy: A Case Report
Wang Z, Mao S, Wang S, Zhang W, Jia C, Zhu T
International Journal of Women's Health 2025, 17:2885-2890
Published Date: 4 September 2025