Back to Journals » OncoTargets and Therapy » Volume 9
miR-486-5p suppresses prostate cancer metastasis by targeting Snail and regulating epithelial–mesenchymal transition
Authors Zhang XG, Zhang T, Yang K, Zhang MH, Wang KM
Received 15 July 2016
Accepted for publication 14 September 2016
Published 8 November 2016 Volume 2016:9 Pages 6909—6914
DOI https://doi.org/10.2147/OTT.S117338
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr William C. Cho
Xiaoguang Zhang,1 Tong Zhang,2 Kuo Yang,3 Minghao Zhang,1 Keming Wang1
1Department of Urology, Tianjin Third Central Hospital, Tianjin, 2Department of Urology, Provincial Hospital Affiliated to Shandong University, Jinan, 3Tianjin Institute of Urology, Tianjin, People’s Republic of China
Abstract: The most common cause of death from prostate cancer (PCa) is metastases. There is an increasing body of evidence that microRNAs play an important role in the development of PCa by regulating target genes involved in tumor metastasis. Here, we identified that expression of miR-486-5p was decreased in metastatic C4-2 cells compared to non-metastatic LNCaP cells. Further validation in clinical samples showed that miR-486-5p expression was significantly decreased in metastatic PCa tissues compared to localized PCa tissues. Functional studies demonstrated that increased miR-486-5p expression can suppress cell migration and the invasive ability of C4-2 cells. Moreover, Snail, a key regulator of the epithelial–mesenchymal transition, was verified as a target gene of miR-486-5p. In conclusion, these findings suggest that miR-486-5p plays a suppressive role in mediating the migration and invasion of PCa by directly suppressing the protein expression of Snail and may provide a potential therapeutic target for the disease.
Keywords: microRNA, prostate cancer, metastasis, EMT
Introduction
Prostate cancer (PCa) is the most common cancer and the second leading cause of cancer-related deaths among North American males.1 Most deaths from this disease are related to metastases. However, the mechanisms underlying PCa metastasis are not fully understood. Further understanding of the machinery driving this process is urgently needed to improve PCa treatment and patient survival rates.
Metastasis is a multi-step process of interrelated events, which is thought to be driven by changes in the gene expression of cancer cells and the interaction between cancer cell and its microenvironment.2 In view of this, it is likely that genes and their upstream regulators, found to be differentially expressed in closely related metastatic and non-metastatic PCa, may play a critical role in metastasis. Such genes and/or their regulators may provide new targets for the therapy of metastatic disease.
MicroRNAs (miRNAs) are a class of single-stranded noncoding RNAs, which act as posttranscriptional regulators of gene expression.3 Many studies have demonstrated that some miRNAs may play important roles in cancer cell invasion, migration, and metastasis by regulating the expression of their target genes.4 Reduced miR-486-5p expression has been observed in multiple types of tumors, such as gastric cancer, lung cancer, colorectal carcinoma, and glioblastoma.5–11 In these cancers, it may play a suppressive role in tumor progression by regulating tumor proliferation and metastasis. However, miR-486-5p was observed to be upregulated and can enhance tumor growth and survival of some malignancies, such as renal cancer and chronic myeloid leukemia, indicating a context-dependent role of it in cancer progression.12,13 To date, the function of miR-486-5p in PCa, especially its role in PCa metastasis, is not clear. In this study, we tested the expression of miR-486-5p in non-metastatic and metastatic PCa cell lines and clinical samples, investigated the roles of miR-486-5p in the regulation of PCa cell migration and invasion, and explored the underlying mechanisms regulated by miR-486-5p.
Materials and methods
Clinical samples
All patients provided written informed consent and a total of 28 fresh PCa specimens were collected from Tianjin Third Central Hospital and Tianjin Institute of Urology. In all, 17 primary PCa tissues were obtained from radical prostatectomy. Lymph node metastasis tissues were obtained from 11 patients with PCa. Following surgical resection, samples were immediately frozen in liquid nitrogen. This study was approved by the research ethics committee of Tianjin Third Central Hospital and Tianjin Institute of Urology.
Cell culture
LNCaP and C4-2 cells were cultured in a Roswell Park Memorial Institute-1640 medium (Thermo Fisher Scientific, MA, USA) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific) at 37°C under a humidified atmosphere containing 5% CO2.
Cell transfection
C4-2 cells were transfected with miR-486-5p mimics or scramble (Sangon Biotech Co., Ltd., Shanghai, People’s Republic of China) for 48 hours by Lipofectamine 2000 (Thermo Fisher Scientific) in accordance with the manufacturer’s instructions.
RNA extraction and real-time PCR analysis
The total RNA from cultured cells and clinical PCa tissues were extracted using TRIzol reagent (Thermo Fisher Scientific). miRNA-specific complementary DNA was synthesized using a TaqMan MicroRNA Reverse Transcription Kit and miRNA-specific RT primers from a TaqMan MicroRNA Assay (Thermo Fisher Scientific). The miR-486-5p expression was measured by TaqMan miRNA Assays according to the manufacturer’s instructions (Thermo Fisher Scientific). The expression of miR-486-5p was normalized to U6 small nuclear RNA.
Western blot
The total protein from the cultured cells was prepared using radioimmunoprecipitation assay lysis buffer. Protein concentration was determined using the bicinchoninic acid protein assay (Pierce, Rockford, IL, USA). For immunoblotting, the following antibodies were used: anti-E-cadherin antibody (Abcam, Cambridge, UK), anti-N-cadherin antibody (Santa Cruz Biotechnology Inc., Dallas, TX, USA), anti-Snail antibody (Santa Cruz Biotechnology Inc.), and anti-actin (Sigma-Aldrich Co., St Louis, MO, USA). Signals were detected by Super Signal West Pico Chemiluminescent Substrate Kit (Pierce).
Cell migration assay
C4-2 cells were plated onto six-well plates. Once the cells reached confluence, the cells were rinsed with prostate-specific antigen (PSA). Straight scratches were created using P200 pipette tips. Cells were incubated in a 0.1% fetal bovine serum medium afterward. Images of wounds were taken immediately and after 24 hours of incubation. Wound closure was quantified by measuring the areas covered by cells.
Invasion assay
Modified Boyden chambers (BD Biosciences, San Jose, CA, USA) were used according to the manufacturer’s manual for the tumor cell invasion assay. In brief, C4-2 cells were seeded to the upper part of the chambers. In all, 5% fetal bovine serum was added to the lower chambers. The cells were incubated at 37°C for 22 hours. The cells that were attached to the upper surface of the chamber were wiped off using a cotton swab. The cells on the lower surface were fixed with paraformaldehyde and stained with crystal violet. The cells were counted under a light microscope.
Luciferase reporter gene assay
The 3′-untranslated region (UTR) sequence of SNAI1 predicted to interact with miR-486-5p was synthesized and cloned downstream of the Renilla luciferase gene in psiCHECK-2 vector (Promega, Madison, WI, USA). Plasmid DNAs were co-transfected into C4-2 cells with miR-486-5p mimics or scramble sequence. Luciferase activates were measured 48 hours post-transfection using Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer’s instructions. Renilla luciferase activities were normalized to the internal control firefly luciferase activity.
Statistical analysis
Data were presented as mean ± SD. Statistical significance was established using the Student’s t-test. Statistical analyses were performed using SPSS, version 17.0 (SPSS Inc., Chicago, IL, USA). The significance level was set at 0.05 to indicate statistical significance (P<0.05).
Results
Expression of miR-486-5p was downregulated in the metastatic PCa cell line and clinical metastatic PCa tissues
At first, miR-486-5p expression was examined in a non-metastatic LNCaP cell line and its metastatic derivative cell line C4-2. Quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) results demonstrated that the metastatic C-4 cell line showed significantly decreased expression of miR-486-5p than the non-metastatic LNCaP cells (P<0.01), which indicates an association of miR-486-5p expression with PCa metastasis (Figure 1A). To validate this association, the expression of miR-486-5p was further examined in 28 fresh PCa specimens, including 17 primary PCa cases and 11 metastatic PCa cases. The qRT-PCR results showed that the expression of miR-486-5p in metastatic PCa tissues was significantly lower than that in localized PCa tissues (P<0.01), which is consistent with the findings in cell line models (Figure 1B).
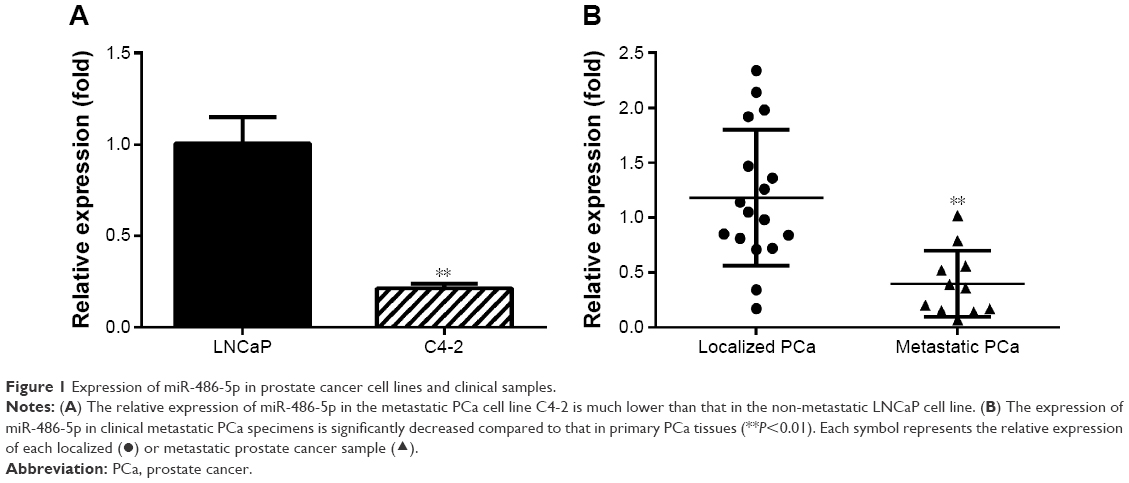 | Figure 1 Expression of miR-486-5p in prostate cancer cell lines and clinical samples. |
miR-486-5p suppresses PCa cell migration and invasion
To study the function of miR-486-5p in PCa, the mimics of miR-486-5p and scramble sequence (negative control) were transfected into C4-2 cells, which were known to express relatively low levels of miR-486-5p. qRT-PCR confirmed the overexpression of miR-486-5p in C4-2 cells transfected with miR-486-5p mimics (Figure 2A). The wound healing assay and Matrigel (Boyden chamber) invasive assay showed that increased miR-486-5p expression significantly suppressed C4-2 cell migration and invasion (Figure 2B and C).
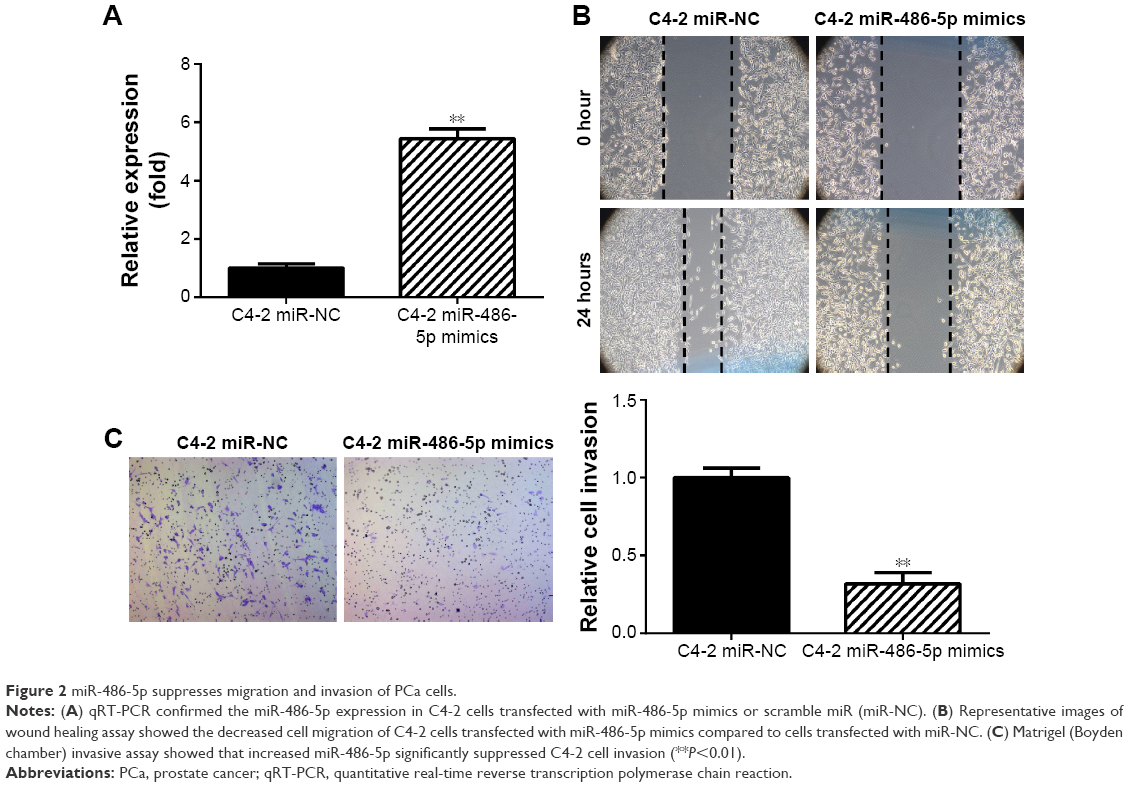 | Figure 2 miR-486-5p suppresses migration and invasion of PCa cells. |
miR-486-5p targets SNAI1 and inhibits epithelial–mesenchymal transition
To study the mechanisms underlying miR-486-5p-regulated PCa metastasis, a number of potential target genes of miR-486-5p were identified using TargetScan Release 7.0 (Figure 3A). Further validation of the protein expression of these candidates in LNCaP and C4-2 using Western blot demonstrated that Snail, a protein encoded by the SNAI1 gene, showed increased expression in C4-2 cells compared to LNCaP cells (Figure 3B). In C4-2 cells transfected with miR-486-5p mimics, the expression of Snail was decreased compared to that transfected with scramble sequence. This result suggested that SNAI1 is likely a target gene of miR-486-5p in PCa. Moreover, the expression of the epithelial marker E-cadherin was increased and the expression of N-cadherin was decreased in the miR-486-5p-overexpressed C4-2 cells compared to the negative control cells (Figure 3C). The direct regulation of miR-486-5p was further validated using a luciferase reporter assay by cloning the SNAI1 3′-UTR sequence into a psiCHECK-2 vector. We observed that the miR-486-5p-overexpressed C4-2 cells showed an evidently suppressed luciferase activity compared to the negative control cells (Figure 3D). Furthermore, a significant negative correlation between miR-486-5p and Snail expression was observed in the clinical samples (r=−0.56 and P<0.01). These results indicated that the overexpression of miR-486-5p may reverse epithelial–mesenchymal transition (EMT) by suppressing Snail expression.
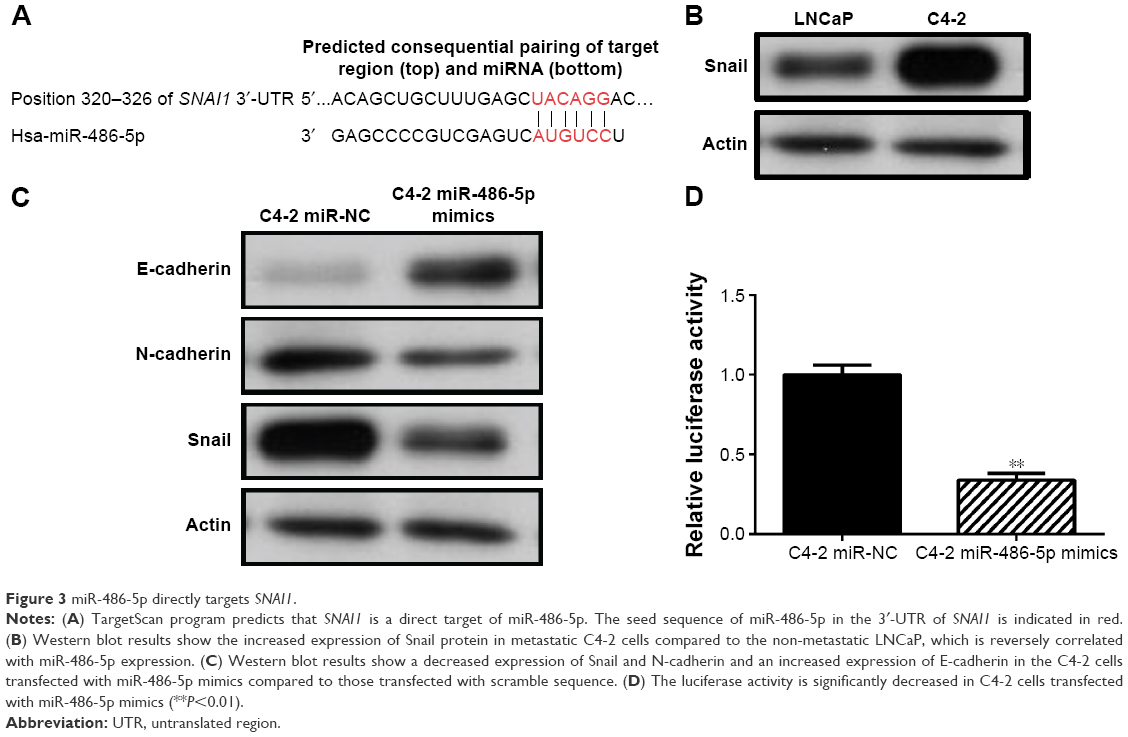 | Figure 3 miR-486-5p directly targets SNAI1. |
Discussion
Most PCa deaths are due to metastases. However, the mechanisms underlying PCa metastasis remain largely unclear. There is an urgent need to identify reliable metastatic biomarkers and effective therapeutic targets, which may lead to improved management of the disease. A number of miRNAs have been reported to be deregulated and play an important role in PCa development and progression by regulating the expression of key target genes. It has been observed that miR-486-5p is downregulated in several types of tumors and may play a suppressive role in tumor progression. Recently, Song et al14 reported a significant downregulation of miR-486-5p in PCa tissues compared to benign tissues using high-throughput Illumina sequencing and qRT-PCR methods, which suggest that miR-486-5p may act as a tumor suppressor in PCa. However, previous studies of miR-486-5p in PCa mainly focused on primary disease and lacked functional assays.14 In this study, we demonstrated the downregulation of miR-486-5p in the metastatic PCa cell line compared to the non-metastatic cell line and further validated the finding in clinical PCa samples. We did not observe a significant correlation between miR-486-5p, androgen receptor, and PSA expression. In addition, functional analysis showed that an increased miR-486-5p can suppress PCa cell migration and invasion, suggesting a metastasis-suppressing role in PCa.
EMT is a crucial mechanism that guides proper cell movement during several phases of embryonic development15 and is also commonly observed in the process of cancer metastasis.16 Such transition from a non-invasive to an invasive phenotype is characterized by decreased expressions of epithelial cell adhesion molecules, such as E-cadherin, and increased expressions of mesenchymal markers, such as N-cadherin and Vimentin.16 Such changes in cell phenotype and behavior are mediated by a number of key transcription factors, including Snail.17,18
It has been reported that Snail can repress the expression of epithelial genes like E-cadherin by binding to E-box DNA sequences through their carboxy-terminal zinc-finger domains followed by recruitment of the PRC2.19,20 As a key EMT transcription factor, Snail can also activate other transcription factors and markers that contribute to the mesenchymal phenotype, such as TWIST, ZEB1, ZEB2, and N-cadherin.21 In this study, we predicted that Snail may be directly regulated by miR-486-5p and validated the direct binding of miR-486-5p and 3′-UTR of SNAI1. Western blot analyses confirmed that Snail expression was decreased in C4-2 PCa cells transfected with miR-486-5p mimics when compared with the cells transfected with scramble sequence. Furthermore, E-cadherin expression was significantly increased and N-cadherin expression was decreased in miR-486-5p-overexpressed C4-2 cells. These data suggested that miR-486-5p may directly control Snail protein expression and further regulate EMT and cell invasion in PCa.
Conclusion
This study demonstrated that miR-486-5p is downregulated in metastatic PCa cell lines and clinical samples compared to non-metastatic ones. Functional studies showed that miR-486-5p suppresses PCa cell migration and invasion by directly regulating Snail protein expression. The findings suggest that miR-486-5p plays a metastasis-suppressor role in PCa and may provide a potential therapeutic target for the disease.
Acknowledgment
This work was supported by Tianjin Science and Technique Foundation (13RCGFSY19100).
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015;65(1):5–29. | ||
Fidler IJ. The pathogenesis of cancer metastasis: the ‘seed and soil’ hypothesis revisited. Nat Rev Cancer. 2003;3(6):453–458. | ||
Lin S, Gregory RI. MicroRNA biogenesis pathways in cancer. Nat Rev Cancer. 2015;15(6):321–333. | ||
Croce CM. Causes and consequences of microRNA dysregulation in cancer. Nat Rev Genet. 2009;10(10):704–714. | ||
Navon R, Wang H, Steinfeld I, Tsalenko A, Ben-Dor A, Yakhini Z. Novel rank-based statistical methods reveal microRNAs with differential expression in multiple cancer types. PLoS One. 2009;4(11):e8003. | ||
Peng Y, Dai Y, Hitchcock C, et al. Insulin growth factor signaling is regulated by microRNA-486, an underexpressed microRNA in lung cancer. Proc Natl Acad Sci U S A. 2013;110(37):15043–15048. | ||
Pang W, Tian X, Bai F, et al. Pim-1 kinase is a target of miR-486-5p and eukaryotic translation initiation factor 4E, and plays a critical role in lung cancer. Mol Cancer. 2014;13:240. | ||
Chen H, Ren C, Han C, Wang D, Chen Y, Fu D. Expression and prognostic value of miR-486-5p in patients with gastric adenocarcinoma. PLoS One. 2015;10(3):e0119384. | ||
Oh HK, Tan AL, Das K, et al. Genomic loss of miR-486 regulates tumor progression and the OLFM4 antiapoptotic factor in gastric cancer. Clin Cancer Res. 2011;17(9):2657–2667. | ||
Gal H, Pandi G, Kanner AA, et al. MIR-451 and imatinib mesylate inhibit tumor growth of glioblastoma stem cells. Biochem Biophys Res Commun. 2008;376(1):86–90. | ||
Liu C, Li M, Hu Y, et al. miR-486-5p attenuates tumor growth and lymphangiogenesis by targeting neuropilin-2 in colorectal carcinoma. Onco Targets Ther. 2016;9:2865–2871. | ||
Wang LS, Li L, Chu S, et al. MicroRNA-486 regulates normal erythropoiesis and enhances growth and modulates drug response in CML progenitors. Blood. 2015;125(8):1302–1313. | ||
Goto K, Oue N, Shinmei S, et al. Expression of is a potential prognostic factor after nephrectomy in advanced renal cell carcinoma. Mol Clin Oncol. 2013;1(2):235–240. | ||
Song C, Chen H, Wang T, Zhang W, Ru G, Lang J. Expression profile analysis of microRNAs in prostate cancer by next-generation sequencing. Prostate. 2015;75(5):500–516. | ||
Hay ED. An overview of epithelio-mesenchymal transformation. Acta Anat. 1995;154(1):8–20. | ||
Kalluri R, Weinberg RA. The basics of epithelial-mesenchymal transition. J Clin Invest. 2009;119(6):1420–1428. | ||
Peinado H, Quintanilla M, Cano A. Transforming growth factor beta-1 induces snail transcription factor in epithelial cell lines: mechanisms for epithelial mesenchymal transitions. J Biol Chem. 2003;278(23):21113–21123. | ||
Yang J, Mani SA, Donaher JL, et al. Twist, a master regulator of morphogenesis, plays an essential role in tumor metastasis. Cell. 2004;117(7):927–939. | ||
Peinado H, Olmeda D, Cano A. Snail, Zeb and bHLH factors in tumour progression: an alliance against the epithelial phenotype? Nat Rev Cancer. 2007;7(6):415–428. | ||
Barrallo-Gimeno A, Nieto MA. The Snail genes as inducers of cell movement and survival: implications in development and cancer. Development. 2005;132(14):3151–3161. | ||
Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–196. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.