Back to Journals » Journal of Blood Medicine » Volume 14
MicroRNAs as Quality Assessment Tool in Stored Packed Red Blood Cell in Blood Banks
Authors Mulatie Z, Aynalem M ![]() , Getawa S
, Getawa S ![]()
Received 29 November 2022
Accepted for publication 1 February 2023
Published 8 February 2023 Volume 2023:14 Pages 99—106
DOI https://doi.org/10.2147/JBM.S397139
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Zewudu Mulatie,1 Melak Aynalem,2 Solomon Getawa2
1Department of Medical Laboratory Sciences, College of Medicine and Health Sciences, Wollo University, Desie, Ethiopia; 2Department of Hematology and Immunohematology, School of Biomedical and Laboratory Sciences, College of Medicine and Health Sciences, University of Gondar, Gondar, Ethiopia
Correspondence: Zewudu Mulatie, Department of Medical Laboratory Sciences, College of Medicine and Health Sciences, Wollo University, P.O.box: 1145, Desie, Ethiopia, Tel +251945274251, Email [email protected]
Abstract: Micro-ribonucleic acids are control gene expression in cells. They represent the changed cellular states that occur can be employed as biomarkers. Red blood cells alter biochemically and morphologically while they are being stored, which could be detrimental to transfusion. The effect of storage on the erythrocyte transcriptome is not mostly investigated. Because adult erythrocytes lack a nucleus, it has long been assumed that they lack deoxyribonucleic acid and ribonucleic acid. On the other hand, erythrocytes contain a diverse range of ribonucleic acids, of which micro-ribonucleic acids are key component. Changes in this micro-ribonucleic acid protect cells from death and adenine triphosphate depletion, and they are linked to specific storage lesions. As a result, changes in micro-ribonucleic acid in stored erythrocytes may be used as a marker to assess the quality and safety of stored erythrocytes. Therefore, this review ams to review the role of microRNA in stored packed red blood cells as quality indicator. Google Scholar, PubMed, Scopus, and Z-libraries are used for searching articles and books. The article included in this paper was written in the English language and had the full article. During long storage of RBCs, miR-16-2-3p, miR-1260a, miR-1260b, miR-4443, miR-4695-3p, miR-5100, let-7b, miR-16, miRNA-1246, MiR-31-5p, miR-203a, miR-654-3p, miR-769-3p, miR-4454, miR-451a and miR-125b- 5p are up regulated. However, miR-96, miR-150, miR-196a, miR-197, miR-381 and miR-1245a are down regulated after long storage of RBCs. The changes of this microRNAs are linked to red blood cell lesions. Therefore, micro-ribonucleic acids are the potential quality indicator in stored packed red blood cells in the blood bank. Particularly, micro-ribonucleic acid-96 is the most suitable biomarker for monitoring red blood cell quality in stored packed red blood units.
Keywords: blood bank, microRNA, packed red blood cell
Introduction
Blood transfusions save hundreds of lives around the world per day. The most commonly transfused blood component worldwide is concentrated red blood cells (RBC).1 Blood transfusions are one of the most common clinical therapeutic procedures for acute or chronic blood loss, with approximately 85 million RBC units transfused each year around the world.2 Blood and blood component storage allows for better control of blood supply safety, adequacy, and management.3 Red blood cells can now be kept in a solution of saline adenine–glucose–mannitol (SAGM) as a combination anticoagulant and energy source for up to 6 weeks at a temperature of +4°C for an adequate and available blood supply, however storage may compromise the efficacy and quality of blood components.4
During storage, blood cells continue their metabolic activity, releasing byproducts into the medium and undergoing biologic and immunologic alterations that may have an impact on RBC function and survival.5,6 Despite recent improvements in blood storage conditions, a wide range of detrimental molecular, biochemical, and physical modifications known as “storage lesions” have been observed during preservation.1,5 Metabolic effects, shape changes, membrane loss, rheological alterations, losses of membrane carbohydrates, oxidative damage to lipids and proteins, changes in oxygen affinity, and changes in oxygen delivery are all part of the storage lesion that red blood cells go through. Additionally, there are concerns associated with shorter RBC lifespan, increased RBC adherence to endothelium, potassium buildup, and the loss of active proteins, lipids, and microvesicles.7
Storage damages include changes in cell flexibility and a decrease in glucose, diphosphoglycerate, and adenine triphosphate (ATP) levels, as well as acidosis caused by lactic acid accumulation. Deactivation of the sodium/potassium ATPase pump results in a drop in sodium and a rise in potassium levels in plasma.4 Other morphological and molecular alterations, as well as oxidative damage to lipids, proteins, and carbohydrates, contributed to the sphero-echinocyte shape. Spherical cells have a lower surface area-to-volume ratio than discoid cells, which leads to higher osmotic fragility and decreased cell deformability and survival.8
The two key elements that influence the process of causing a red blood cell storage lesion are the length of storage period and donor variability. Even when the same amount of time and ex vivo storage conditions are used, donor variation may affect the quality of the red blood cell units obtained from various blood donors. Donor genetic and environmental characteristics are directly related to donor variation.9,10 Red blood cell storage capacity may be impacted by the inherent traits of blood donors (RBCs). Hemolysis, redox, and metabolic characteristics of stored RBCs as well as their post-transfusion recovery have been demonstrated to be influenced by environmental (alcohol use,11 smoking12 and genetic (sex,13 ethnicity,14 hemoglobin (Hb) mutation.15 The increase in red blood storage lesion was caused by factors including blood donors’ aging, smoking, BMI, and decreased level of physical activity.16 Additionally, having a male sex, being Asian or African American, having sickle cell disease, thalassemia, or lacking glucose-6-phosphate dehydrogenase may alter RBC rheology or antioxidant capability and compromise RBC recovery during storage and after transfusion.15,17,18
RBCs’ homeostasis is affected by storage on three levels: energy metabolism, redox metabolism, and cell membrane, leading to a clearly characterized phenotype of morphologic, structural, and functional alterations. This phenotype is characterized by loss of normal shape due to the buildup of free hemoglobin, heme, lactate, oxidized glutathione, malondialdehyde, potassium, inflammatory lipids, and extracellular vesicles, including microvesicles. Additionally, preserved packed red blood cells showed decreased PH, glutathione, phospholipids, cholesterol, and 2,3-diphosphoglycerate levels.7,8 Additionally, improper ATP-centered metabolism and oxidative stress contribute to RBC storage lesions (RBCSL). High energy phosphate compound metabolites are used as energy sources to preserve membrane integrity and, hence, RBC survival. This process causes the RBCs to lose energy and valuable deformability, physiological surface area, topography, and shape.19
In general, these alterations have an impact on the quality of RBCs that have been preserved for a long time. Even when transfused within the 42 days, stored RBC has a worse quality including the higher fraction of RBC eliminated after transfusion, reduced oxygen delivery capability, reduced deformability and adverse clinical consequences.20–25 Also, these changes may have been linked to a negative impact on transfusion.8,26 Transfusions of old RBCs have been linked to greater rates of morbidity and mortality in specific patient groups, particularly in newborns and infants after massive transfusions. Transfusions of RBC with elevated potassium levels have been linked to cardiac hyperkalemia and neonatal arrhythmia.4,8 Prolonged RBC preservation has been linked to an increased risk of problems and poor clinical results. Patients with heart disease who had “older” RBC transfusions may develop thrombotic problems.27 Prolonged RBC preservation in trauma patients may raise the risk of deep vein thrombosis and bacterial infections.28,29 The alterations in RBCs and bioactive chemicals accumulated during storage are thought to be the cause of these problems.30 Hemoglobin or hematocrit, percent hemolysis, PH, lactate, electrolyte panels, and 2,3-diphosphate glycerides are among the RBC quality indicators tested by conventional techniques. The novel method for checking the quality of packed RBCs in blood banks is micro-ribonucleic acid (miRNA).31
MicroRNAs are short, non-coding RNA molecules that control how genes are expressed in cells. They are 22 nucleotide lengths in their mature state with great stability and long half-life. The miRNA molecule binds to a messenger RNA (mRNA) with complementarity in the 3’-untranslated region, causing translation to be disrupted and gene expression to be prevented.32 In humans, there are more than 2500 mature miRNAs, resulting in a greater family of microRNAs.33 Those miRNAs are essential for several biological processes, including cell proliferation, tissue development, cell growth, apoptosis,34 signal transmission, immunomodulation, and hematopoiesis.35 MiRNAs are thought to regulate more than 60% of human protein-coding genes.36
For many years, microRNAs have been successfully used as diagnostic markers for a variety of diseases.37 It is possible that changes in miRNA expression levels cause disease and cellular abnormalities, or that diseases cause dysregulated miRNA expression.38 MicroRNAs are involved in a wide range of biological processes, and it has been suggested that when they are dysfunctional, human illnesses can result.36,39,40 They play a function in the inflammatory response, solid cancer and leukemia, and neurological illnesses and are involved in the pathogenesis of a variety of diseases.41 It can be used as valuable clinical biomarkers for diagnosis and prognosis of specific diseases, as well as monitoring therapy responses, due to their unique qualities of disease specificity, high stability, and accessibility.42
Researchers have studied the physiological and biochemical alterations that occur in stored RBCs. RBC must use its accessible posttranscriptional machinery in the cytoplasm to be alive and functional, particularly during in vitro storage, and to control all essential physiologic functions because it is enucleated.43 Despite the lack of ribosomal and large-sized RNAs in mature RBCs, miRNAs are numerous and diverse in mature erythrocytes.44 MicroRNAs are non-coding tiny single RNA molecules that play a key role in a variety of biological processes in RBCs such as cell differentiation, proliferation, and apoptosis. As a result, miRNA alterations in stored RBCs could be used as biomarkers for storage lesions. Due to the linkage between miRNA and its target mRNAs in terms of gene regulation, the underlying processes of miRNA as a quality indicator in stored packed RBCs may offer insight into related RBCSL.45 Researchers have so far paid little attention to the comprehensive study of RBCSL in terms of the exact nature of the occurrence and the molecular components involved.46 Therefore, this review aims to review the role of miRNA as a quality indicator for stored packed RBCs in blood banks. For preparing this review, Google Scholar, PubMed, Scopus and Z-libraries were used for searching articles and books. The keywords such as, Blood bank, microRNA and red blood cells. The article included in this paper was written in the English language and had the full article. Articles that are outdated are excluded from the review. This means the article included only published over 2000 G.C. The reference was entered by endnote Version 20 in Vancouver style.
MicroRNA Expression in Stored Packed RBC as Quality Tool
Compared to other blood cells, mature RBCs have an abundance of miRNAs, which are the negative regulators of cellular mRNAs.45 During storage, the mRNA and miRNA present in stored RBCs undergo morphological and molecular alterations. The miRNA and mRNA expression profiles are altered during storage in RBC. It is reasonable to postulate that miRNAs provide translational regulation of existing mRNAs in these cells. These decisional steps within the stored RBCs will ultimately determine the fate of the RBCs, that is survival during storage by resisting the onset of physiological changes or ultimately undergoing cell death.38
The effect of storage on the erythrocyte transcriptome has not been thoroughly investigated. Because adult erythrocytes lack a nucleus, it has long been assumed that they lack deoxy nucleotide acid (DNA) and RNA. On the other hand, erythrocytes contain a diverse range of RNAs, of which miRNAs are a key component, and these miRNAs persist beyond terminal differentiation.47,48 Changes in these miRNAs protect cells from death and ATP depletion, and they are linked to specific storage lesions.49 As a result, changes in miRNA in stored erythrocytes may be used as a marker to assess the quality and safety of stored erythrocytes.50 MiRNAs should be considered when looking for novel and stable biomarkers in stored packed RBCs for a number of reasons, including the fact that they are very stable in RBCs, some miRNAs can only be expressed in certain cells, and miRNA levels are simple to measure using common laboratory methods, even more advanced and powerful ones.51 The following facts also pertain to miRNAs and cellular apoptosis as they relate to the quality of RBCs that have been stored. The first is that mature RBCs contain a large amount of miRNAs. White blood cell miRNAs have little effect on the second RBC miRNA study because RBC miRNAs create a significant abundance of white blood cells.44 The third is that miRNAs are widely known for controlling how genes and mRNAs expressed within cells, particularly those involved in differentiation and apoptosis.45
The RBC storage lesion is frequently accompanied by considerable transcriptome changes. The miRNAs must be expressed at a high level and target to have a biological effect, and they may even serve a unique role in anucleated cells’ physiological activities. As a result, miRNAs are suitable prospective markers for investigating storage lesions in RBC storage because of their abundance and their stable structure.52 As cells differentiate into reticulocytes, the nuclei are expelled, but translation activities and cytoplasmic RNAs (including mRNAs and miRNAs) are still observable.53
Understanding how miRNAs work in RBCs as they are being stored may shed light on the mechanisms underlying storage defects.54 Through mRNA degradation or translation inhibition, microRNAs regulate cellular processes like apoptosis. This regulatory role of the miRNA is more pertinent to understanding the events connected to RBC stored damage during storage. These cells must rely on the available cytoplasmic posttranscriptional regulatory systems for survival because they are enucleated and have lost their transcriptional regulatory mechanisms.55 As a result, these miRNAs are anticipated to play an important role in erythroid cell posttranscriptional regulation. The expression of numerous miRNAs can substantially change during storage, according to RBC profiling studies, and apoptosis-associated miRNAs have been discovered.38,45
The miRNA of RBCs changes dramatically during storage. For example, the up regulation of miRNA-16-2-3p, miRNA-1260a, miRNA-1260b, miRNA-4443, miRNA-4695-3p, miRNA-5100, lethal (let)-7b, miRNA-16, miRNA-1246, miRNA-31-5p, miRNA-203a, miRNA-654-3p, miRNA-769-3p, miRNA-4454, miRNA-451a and miRNA-125b-5p occurred during long-term preservation of erythrocytes.45,56–59 Although miRNA-96, miRNA-150, miRNA-196a, miRNA-197, miRNA-381, and miRNA-1245a are down regulated after 20 days of storage.45,50,58 The significance of miRNAs in RBC storage lesions has been confirmed by microRNA dysregulation, and this finding raises the possibility that miRNAs in RBCs serve as possible quality markers in stored RBCs.45
Each miRNA in stored RBCs has many functions. Also, the mechanism of how the miRNA is used as the quality tool for stored packed RBCs is based on their function. Based on the research using the target scan tool revealed that let-7b, miRNA-16, miRNA-96, miRNA-196a, miRNA-150, and miRNA-197 each can target on 819, 733, 787, 211, 142, and 140 genes of mRNA, respectively. At least one gene is related to the RBC membrane and apoptosis and targeted by each miRNA.60 MicroRNA-96 appears to target at least 17 conserved mRNAs (genes) associated with RBC membrane and apoptosis.45 Potential targets for miR-96 included septin family genes, cathepsin B, docking protein 4, CAPNS1, caspase 2, and apoptosis-associated tyrosine kinase. The role of miRNA-96 could be the inhibition of apoptosis in mature RBCs. An increase in miRNA-96 expression level in RBCs may inhibit apoptosis. Thereby, this inhibition of apoptosis may result in the accumulation of lipid peroxidation and decrease in antioxidant resources within the RBCs, which can affect the quality of RBC that would be transfused to a blood recipient. Becuase the accumulation of lipid peroxidation is one key components of RBC storage lesion.60 MicroRNA-196a was found to protect stored RBCs from cell death and ATP loss. The variations in miRNA during erythrocyte storage are linked to certain storage lesions. As a result, miRNA has the potential to be used as a biomarker to assess the quality and safety of RBCs.55 MicroRNA-96 is most likely a good candidate for testing as a potential biomarker for evaluating the quality of RBCs in stored blood units.60
The most prevalent miRNAs that have been found to be intracellularly linked to aragonite 2 in 24-hour-stored RBCs are miRNA-451a, miRNA-16, let-7, and miRNA-486-5p.61 In most cells and tissues, miRNA-16 is one of the most prevalently cloned miRNAs. It was discovered to be stable and abundant in RBC storage.62 It is a significant miRNA that participates in a variety of biological activities., includes encouraging cell apoptosis and suppresses cell growth.63,64 Lethal-7 was one of the miRNAs which are crucial for cell maturation and development,65 one of the useful markers of RBC storage is lethal-7b, and the storage of RBCs for up to 40 days causes its levels to rise. It demonstrates that this miRNA is involved in preserving the RBC membrane’s integrity.45
Researchers have determined that miRNA-451a plays a crucial role in the homeostasis of RBCs. Furthermore, it has been observed that miRNA-451a positively controls terminal erythroid development, inhibits the expression of 14-3-3zeta, and protects RBCs from oxidative stress.66,67 The significant regulatory roles for miRNA-381 and miRNA-1245a are inducing apoptosis.68,69 After 1 week of storage, the expression levels of miRNA-381 and miRNA-1245a in RBC were reduced, suggesting that these two miRNAs can resist the negative effects of storage damage. MicroRNA-381 and miRNA-1245a may therefore be employed as indicators for RBC storage products.50 According to reports, miRNA-150-3p is crucial in preventing the apoptotic process. Additionally, it has been demonstrated to play a role in differentiating erythrocytes.70 MicroRNA-769-3p overexpression may be crucial for RBC survival and promotes apoptosis.71 The function of miRNA-1246 is to promote apoptosis while suppressing cellular survival.72 The function of miRNA-4695-3p is to prevent RBCs from apoptosis.73 The cell growth is inhibited by miRNA-4443.74
For the miRNA molecule with multiple functions, miRNA-125b is small and non-coding. It is one of the earliest miRNAs to be discovered. It is essential to many functions, including the control of RBCs’ differentiation, and apoptosis.75 MicroRNA-769-3p overexpression dramatically reduced cell growth and increased apoptosis.76 The miRNA-203a-3p has many impacts, including reducing cell growth and promoting apoptosis.77 MiRNA-654-3p overexpression could induce apoptosis by targeting the mRNA for the ras protein activator like 2 genes.78 The overexpression of miRNA-5100 can induce apoptosis.79 The fact that these miRNAs have a direct link to the mRNAs that are expected to be their apoptosis-related targets will also demonstrate that there was a connection relevant to the storage lesions in RBCs. Given their relationship to mRNA, it would make sense to think that these miRNAs will serve as appropriate indications for storage lesions. Therefore, this microRNA profile may indicate certain types of storage lesions in RBCs Table 1.38
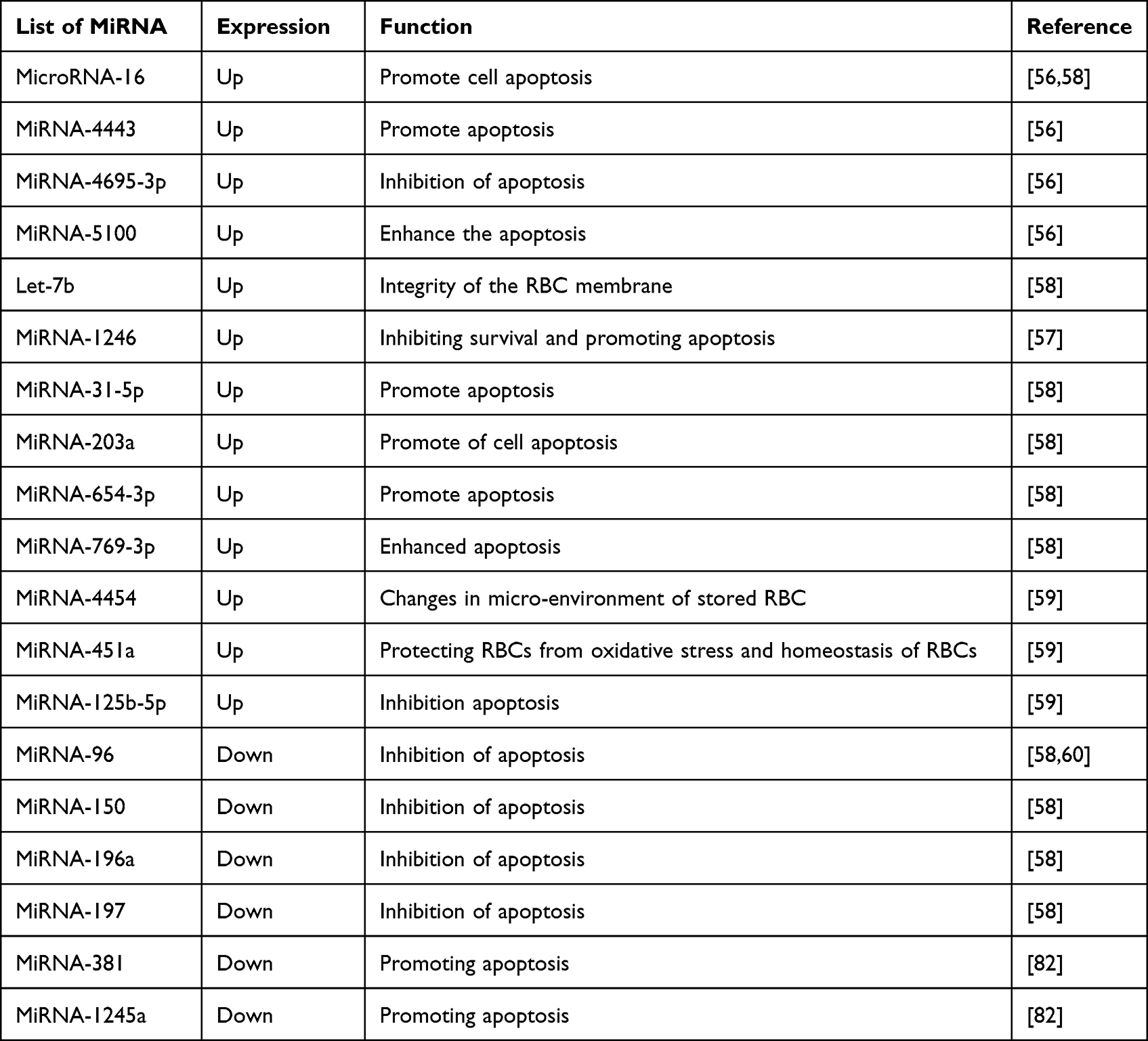 |
Table 1 Summary of the Expression of miRNA in Stored Packed RBCs |
MiRNAs are a novel method for confirming the quality of packed RBCs in blood banks. But, they have some drawbacks. These are the lack of reproducibility across technologies for biomarker discovery in RBCs, some miRNAs are released from blood cells during the storage period and could lead to false results,80 the extraction and analysis of miRNAs from the RBCs requires long times and needs experienced personnel, needs sophisticated laboratories and expensive for detections of miRNA in stored packed RBC.81 Finally, all miRNA findings need additional validation processes and an exact standardization of all pre-analytical and analytical procedures before being applied to clinical practice in order to account for any potential technical biases.
Conclusion
Red blood cells have a comprehensive storage lesion that involves metabolic consequences, shape alterations, membrane loss, rheological changes, losses of membrane carbohydrates, and oxidative damage to lipids. Additionally, the proteins’ modifications to oxygen affinity and delivery, the RBCs’ enhanced endothelial adherence and shorter lifespan, as well as the incidental dangers of potassium buildup and the loss of active proteins, lipids, and microvesicles. The miRNA-16-2-3p, miRNA-1260a, miRNA-1260b, miRNA-4443, miRNA-4695-3p, miRNA-5100, let-7b, miRNA-16, miRNA-1246, miRNA-31-5p, miRNA-203a, miRNA-654-3p, miRNA-769-3p, miRNA-4454, miRNA-451a and miRNA-125b-5p increased during long storage of erythrocytes. However, miRNA-96, miRNA-150, miRNA-196a, miRNA-197, miRNA-381 and miRNA-1245a are down regulated after long storage of RBCs. MicroRNA alterations in stored RBCs can be used as biomarkers for storage lesions. Particularly, miRNA-96 is likely the best candidate to be tested as a potential biomarker for assessing the quality of RBCs in blood units that have been kept. Therefore, recommended for blood bank, the microRNAs are tested as a potential quality biomarker for packed red blood cell. Also, the integration of both biochemical and microRNA detection test is used as quality indicator of packed RBCs. Recommended for researchers, there needs a further study about the role of miRNA as quality indicator for stored packed RBCs and the mechanism how the miRNA used as a quality indicator for stored RBCs. There is a limited study in the role of miRNA as quality tool for stored RBCs, and the mechanism is also not well addressed.
Abbreviations
ATP, adenosine tri phosphate; DNA, deoxyribonucleic acid; Let, lethal; MiRNAs, microribonucleic acid; MRNA, messenger ribonucleic acid; NGS, next-generation sequencing; PCR, polymerase chain reaction; RBC, red blood cell; RBCSL, red blood cell storage lesion; RT, reverse transcriptase; RT-qPCR, reverse transcription quantitative polymerase chain reaction; SAGM, saline–adenine–glucose–mannitol.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Zimrin B, Hess R. Current issues relating to the transfusion of stored red blood cells. Int J Blood Transfus. 2009;96(2):93–103.
2. Carson J, Guyatt G. AABB releases clinical practice guidelines on red blood cell transfusion and storage. J Am Med Assoc. 2021;3(1):28–32.
3. Antonelou H, Seghatchian J. Insights into red blood cell storage lesion: toward a new appreciation. Transfus Apher Sci. 2016;55(3):292–301. doi:10.1016/j.transci.2016.10.019
4. Hess R. Measures of stored red blood cell quality. Int Soc Blood Transfus. 2014;107(1):1–9.
5. Obrador R, Musulin S, Hansen B. Red blood cell storage lesion. J Vet Emerg Crit Care. 2015;25(2):187–199. doi:10.1111/vec.12252
6. Cummings A, Abelson L, Rozanski A, Sharp R. The effect of storage on ammonia, cytokine, and chemokine concentrations in feline whole blood. J Vet Emerg Crit Care. 2016;26(5):639–645. doi:10.1111/vec.12510
7. Hess R. Red cell changes during storage. Transfus Apher Sci. 2010;43(1):51–59. doi:10.1016/j.transci.2010.05.009
8. D’Alessandro A, Kriebardis G, Rinalducci S, et al. An update on red blood cell storage lesions, as gleaned through biochemistry and omics technologies. Transfusion. 2015;55(1):205–219. doi:10.1111/trf.12804
9. Antonelou MH, Seghatchian J. Insights into red blood cell storage lesion: toward a new appreciation. Transfus Apher Sci. 2016;55(3):292–301.
10. Tzounakas VL, Kriebardis AG, Papassideri IS, Antonelou MH. Donor-variation effect on red blood cell storage lesion: a close relationship emerges. Proteomics Clin Appl. 2016;10(8):791–804. doi:10.1002/prca.201500128
11. D’Alessandro A, Fu X, Reisz JA, et al. Ethyl glucuronide, a marker of alcohol consumption, correlates with metabolic markers of oxidant stress but not with hemolysis in stored red blood cells from healthy blood donors. Transfusion. 2020;60(6):1183–1196. doi:10.1111/trf.15811
12. Stefanoni D, Fu X, Reisz JA, et al. Nicotine exposure increases markers of oxidant stress in stored red blood cells from healthy donor volunteers. Transfusion. 2020;60(6):1160–1174. doi:10.1111/trf.15812
13. Tzounakas VL, Anastasiadi AT, Drossos PV, et al. Sex-related aspects of the red blood cell storage lesion. Blood Transfus. 2021;19(3):224–236. doi:10.2450/2020.0141-20
14. D’Alessandro A, Fu X, Kanias T, et al. Donor sex, age and ethnicity impact stored red blood cell antioxidant metabolism through mechanisms in part explained by glucose 6-phosphate dehydrogenase levels and activity. Haematologica. 2021;106(5):1290–1302. doi:10.3324/haematol.2020.246603
15. Tzounakas VL, Anastasiadi AT, Stefanoni D, et al. Beta thalassemia minor is a beneficial determinant of red blood cell storage lesion. Haematologica. 2022;107(1):112–125. doi:10.3324/haematol.2020.273946
16. Kanias T, Lanteri MC, Page GP, et al. Ethnicity, sex, and age are determinants of red blood cell storage and stress hemolysis: results of the REDS-III RBC-Omics study. Blood Adv. 2017;1(15):1132–1141. doi:10.1182/bloodadvances.2017004820
17. Raciti PM, Francis RO, Spitalnik PF, Schwartz J, Jhang JS. Acquired hemoglobin variants and exposure to glucose-6-phosphate dehydrogenase deficient red blood cell units during exchange transfusion for sickle cell disease in a patient requiring antigen-matched blood. J Clin Apher. 2013;28(4):325–329. doi:10.1002/jca.21255
18. Francis RO, Jhang J, Hendrickson JE, Zimring JC, Hod EA, Spitalnik SL. Frequency of glucose-6-phosphate dehydrogenase-deficient red blood cell units in a metropolitan transfusion service. Transfusion. 2013;53(3):606–611. doi:10.1111/j.1537-2995.2012.03765.x
19. Lang E, Pozdeev I, Xu C, et al. Storage of erythrocytes induces suicidal erythrocyte death. Cell Physiol Biochem. 2016;39(2):668–676. doi:10.1159/000445657
20. Qu L, Triulzi J. Clinical effects of red blood cell storage. Cancer Control. 2015;22(1):26–37. doi:10.1177/107327481502200105
21. Koch G, Figueroa I, Li L, Sabik F, Mihaljevic T, Blackstone H. Red blood cell storage: how long is too long? Ann Thorac Surg. 2013;96(5):1894–1899. doi:10.1016/j.athoracsur.2013.05.116
22. Refaai A, Blumberg N. The transfusion dilemma--weighing the known and newly proposed risks of blood transfusions against the uncertain benefits. Best Pract Res Clin Anaesthesiol. 2013;27(1):17–35. doi:10.1016/j.bpa.2012.12.006
23. Flegel A, Natanson C, Klein G. Does prolonged storage of red blood cells cause harm? Br J Haematol. 2014;165(1):3–16. doi:10.1111/bjh.12747
24. Jy W, Ricci M, Shariatmadar S, Gomez-Marin O, Horstman H, Ahn S. Microparticles in stored red blood cells as potential mediators of transfusion complications. Transfusion. 2011;51(4):886–893. doi:10.1111/j.1537-2995.2011.03099.x
25. Hod A, Brittenham M, Billote B, et al. Transfusion of human volunteers with older, stored red blood cells produces extravascular hemolysis and circulating non-transferrin-bound iron. Blood. 2011;118(25):6675–6682. doi:10.1182/blood-2011-08-371849
26. Doctor A, Spinella P. Effect of processing and storage on red blood cell function in vivo. Semin Perinatol. 2012;36(4):248–259. doi:10.1053/j.semperi.2012.04.005
27. Czubak-Prowizor K, Rywaniak J, Zbikowska M. Red blood cell supernatant increases activation and agonist-induced reactivity of blood platelets. Thromb Res. 2020;196(3):543–549. doi:10.1016/j.thromres.2020.10.023
28. Spinella C, Carroll L, Staff I, et al. Duration of red blood cell storage is associated with increased incidence of deep vein thrombosis and in hospital mortality in patients with traumatic injuries. Crit Care. 2009;13(5):R151. doi:10.1186/cc8050
29. Juffermans P, Vlaar P, Prins J, Goslings C, Binnekade M. The age of red blood cells is associated with bacterial infections in critically ill trauma patients. Blood Transfus. 2012;10(3):290–295. doi:10.2450/2012.0068-11
30. Sparrow L. Red blood cell storage and transfusion-related immunomodulation. Blood Transfus. 2010;8(3):26–30.
31. D’Alessandro A, Dzieciatkowska M, Nemkov T, Hansen KC. Red blood cell proteomics update: is there more to discover? Blood Transfus. 2017;15(2):182–187. doi:10.2450/2017.0293-16
32. Dave V, Ngo T, Pernestig A, et al. MicroRNA amplification and detection technologies: opportunities and challenges for point of care diagnostics. Lab Invest. 2019;99(4):452–469. doi:10.1038/s41374-018-0143-3
33. Kozomara A, Griffiths S. miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014;42(1):68–73. doi:10.1093/nar/gkt1181
34. Esquela A, Slack F. Oncomirs - microRNAs with a role in cancer. Nat Rev Cancer. 2006;6(4):259–269. doi:10.1038/nrc1840
35. Undi B, Kandi R, Gutti K. MicroRNAs as haematopoiesis regulators. Adv Hematol. 2013;20(13):695–754.
36. Friedman C, Farh K, Burge B, Bartel P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009;19(1):92–105. doi:10.1101/gr.082701.108
37. Lu M, Zhang Q, Deng M, et al. An analysis of human microRNA and disease associations. PLoS One. 2008;3(10):34. doi:10.1371/journal.pone.0003420
38. Ryan P, Atreya C. Blood cell microRNAs: what are they and what future do they hold? Transfus Med Rev. 2011;25(3):247–251. doi:10.1016/j.tmrv.2011.01.005
39. Chen K, Rajewsky N. The evolution of gene regulation by transcription factors and microRNAs. Nat Rev Genet. 2007;8(2):93–103. doi:10.1038/nrg1990
40. Kim J, Inoue K, Ishii J, et al. A microRNA feedback circuit in midbrain dopamine neurons. Science. 2007;317(58):1220–1224. doi:10.1126/science.1140481
41. Vishnoi A, Rani S. MiRNA biogenesis and regulation of diseases: an overview. Methods Mol Biol. 2017;1509(3):1–10.
42. Hayes J, Peruzzi P, Lawler S. MicroRNAs in cancer: biomarkers, functions and therapy. Trends Mol Med. 2014;20(8):460–469. doi:10.1016/j.molmed.2014.06.005
43. Bratosin D, Estaquier J, Petit F, et al. Programmed cell death in mature erythrocytes: a model for investigating death effector pathways operating in the absence of mitochondria. Cell Death Differ. 2001;8(12):1143–1156. doi:10.1038/sj.cdd.4400946
44. Chen Y, Wang Y, Telen J, Chi T. The genomic analysis of erythrocyte microRNA expression in sickle cell diseases. PLoS One. 2008;3(6):23–60.
45. Kannan M, Atreya C. Differential profiling of human red blood cells during storage for 52 selected microRNAs. Transfusion. 2010;50(7):1581–1588. doi:10.1111/j.1537-2995.2010.02585.x
46. Flatt F, Bawazir M, Bruce J. The involvement of cation leaks in the storage lesion of red blood cells. Front Physiol. 2014;5:214. doi:10.3389/fphys.2014.00214
47. Yang H, Doss F, Walzer A, et al. Angiogenin-mediated tRNA cleavage as a novel feature of stored red blood cells. Br J Haematol. 2019;185(4):760–764. doi:10.1111/bjh.15605
48. Azzouzi I, Moest H, Wollscheid B, Schmugge M, Eekels M, Speer O. Deep sequencing and proteomic analysis of the microRNA-induced silencing complex in human red blood cells. Exp Hematol. 2015;43(5):382–392. doi:10.1016/j.exphem.2015.01.007
49. Chen P, Hong J, Chi J. Discovery, genomic analysis, and functional role of the erythrocyte RNAs. Curr Pathobiol Rep. 2017;5(1):43–48. doi:10.1007/s40139-017-0124-z
50. Jian F, Peng Y, Bian M. Expression and bioinformatics analysis of key miRNAs in stored red blood cells. Transfus Med Hemother. 2022;21(3):1–8.
51. Andreasen D, Fog U, Biggs W, et al. Improved microRNA quantification in total RNA from clinical samples. Blood Transfus. 2010;50(4):6–9.
52. Zhang Y, Huang G, Yuan Z, et al. Profiling and bioinformatics analysis revealing differential circular RNA expression about storage lesion regulatory in stored red blood cells. Transfus Med Hemother. 2022;49(2):76–87. doi:10.1159/000519626
53. Sangokoya C, LaMonte G, Chi T. Isolation and characterization of microRNAs of human mature erythrocytes. Methods Mol Biol. 2010;667(3):193–203.
54. Kannan M, Kulkarni S, Atreya C. Apoptotic microRNA profiling of packed red blood cells during storage. Blood. 2009;114(22):3145–3149. doi:10.1182/blood.V114.22.3145.3145
55. Sarachana T, Kulkarni S, Atreya D. Evaluation of small noncoding RNAs in ex vivo stored human mature red blood cells: changes in noncoding RNA levels correlate with storage lesion events. Transfusion. 2015;55(11):2672–2683. doi:10.1111/trf.13235
56. Haberberger A, Kirchner B, Riedmaier I, et al. Changes in the microRNA expression profile during blood storage. BMJ Open Sport Exer Med. 2018;4(1):354–360. doi:10.1136/bmjsem-2018-000354
57. Kong Y, Tian X, He R, et al. The accumulation of exosome-associated microRNA-1246 and microRNA-150-3p in human red blood cell suspensions. J Transl Med. 2021;19(1):225–228. doi:10.1186/s12967-021-02887-2
58. Chen X, Xie X, Xing Y, Yang X, Yuan Z, Wei Y. MicroRNA dysregulation associated with red blood cell storage. Transfus Med Hemother. 2018;45(6):397–402. doi:10.1159/000489321
59. Huang H, Zhu J, Fan L, et al. MicroRNA profiling of exosomes derived from red blood cell units: implications in transfusion-related immunomodulation. Biomed Res Int. 2019;19(2):459–465.
60. Zare S, Mousavi K, Maghsudlu M, Shahabi M. miRNA96 expression level within red blood cells is probably associated with RSL indicators during the storage of red blood cell units. Transfus Apher Sci. 2021;60(3):103–122. doi:10.1016/j.transci.2021.103122
61. Vu L, Ragupathy V, Kulkarni S, Atreya C. Analysis of Argonaute 2-microRNA complexes in ex vivo stored red blood cells. Transfusion. 2017;57(12):2995–3000. doi:10.1111/trf.14325
62. Chen CZ, Li L, Lodish F, Bartel P. MicroRNAs modulate hematopoietic lineage differentiation. Science. 2004;303(5654):83–86. doi:10.1126/science.1091903
63. Linsley P, Schelter J, Burchard J, et al. Transcripts targeted by the microRNA-16 family cooperatively regulate cell cycle progression. Mol Cell Biol. 2007;27(6):2240–2252. doi:10.1128/MCB.02005-06
64. Yan X, Liang H, Deng T, et al. The identification of novel targets of miR-16 and characterization of their biological functions in cancer cells. Mol Cancer. 2013;12(5):92–95. doi:10.1186/1476-4598-12-92
65. Wulczyn G, Smirnova L, Rybak A, et al. Post-transcriptional regulation of the let-7 microRNA during neural cell specification. FASEB j. 2007;21(2):415–426. doi:10.1096/fj.06-6130com
66. Patrick M, Zhang C, Tao Y, et al. Defective erythroid differentiation in miR-451 mutant mice mediated by 14-3-3zeta. Genes Dev. 2010;24(15):1614–1619. doi:10.1101/gad.1942810
67. Rasmussen D, Simmini S, Abreu-Goodger C, et al. The miR-144/451 locus is required for erythroid homeostasis. J Exp Med. 2010;207(7):1351–1358. doi:10.1084/jem.20100458
68. Bolandghamat Pour Z, Nourbakhsh M, Mousavizadeh K, et al. Up-regulation of miR-381 inhibits NAD+ salvage pathway and promotes apoptosis in breast cancer cells. Excli J. 2019;18(6):683–696. doi:10.17179/excli2019-1431
69. Liao W, Zhang Y. MicroRNA-381 facilitates autophagy and apoptosis in prostate cancer cells via inhibiting the RELN-mediated PI3K/AKT/mTOR signaling pathway. Life Sci. 2020;254(4):117–122. doi:10.1016/j.lfs.2020.117672
70. Wan Y, Cui R, Gu J, et al. Identification of four oxidative stress-responsive microRNAs, miR-34a-5p, miR-1915-3p, miR-638, and miR-150-3p, in hepatocellular carcinoma. Oxid Med Cell Longev. 2017;2017(1):189–198. doi:10.1155/2017/5189138
71. Luo E, Chang Y, Sher Y, et al. MicroRNA-769-3p down-regulates NDRG1 and enhances apoptosis in MCF-7 cells during reoxygenation. Sci Rep. 2014;4(2):590–594. doi:10.1038/srep05908
72. Bhagirath D, Yang T, Bucay N, et al. microRNA-1246 is an exosomal biomarker for aggressive prostate cancer. Cancer Res. 2018;78(7):1833–1844. doi:10.1158/0008-5472.CAN-17-2069
73. Yang Y, Hou Y, Li J, Zhang F, Du Q. Characterization of antiapoptotic microRNAs in primary Sjögren’s syndrome. Cell Biochem Funct. 2020;38(8):1111–1118. doi:10.1002/cbf.3569
74. Meerson A, Yehuda H. Leptin and insulin up-regulate miR-4443 to suppress NCOA1 and TRAF4, and decrease the invasiveness of human colon cancer cells. BMC Cancer. 2016;16(1):882–885. doi:10.1186/s12885-016-2938-1
75. Le M, Xie H, Zhou B, et al. MicroRNA-125b promotes neuronal differentiation in human cells by repressing multiple targets. Mol Cell Biol. 2009;29(19):5290–5305. doi:10.1128/MCB.01694-08
76. Luo E, Chang Y, Sher Y, et al. MicroRNA-769-3p down-regulates NDRG1 and enhances apoptosis in MCF-7 cells during reoxygenation. Sci Rep. 2014;4(1):5908–5911.
77. Liu H, Zhang Y, Zhu B, et al. MiR-203a-3p regulates the biological behaviors of ovarian cancer cells through mediating the Akt/GSK-3β/Snail signaling pathway by targeting ATM. J Ovarian Res. 2019;12(1):60–65. doi:10.1186/s13048-019-0532-2
78. Xiong J, Xing S, Dong Z, et al. miR‑654‑3p suppresses cell viability and promotes apoptosis by targeting RASAL2 in non‑small‑cell lung cancer. Mol Med Rep. 2021;23(2):78–82. doi:10.3892/mmr.2020.11721
79. Zhang H, Li H, Wang G, et al. MKL1/miR-5100/CAAP1 loop regulates autophagy and apoptosis in gastric cancer cells. Neoplasia. 2020;22(5):220–230. doi:10.1016/j.neo.2020.03.001
80. de Gonzalo-Calvo D, Pérez-Boza J, Curado J, Devaux Y. Challenges of microRNA-based biomarkers in clinical application for cardiovascular diseases. Clin Transl Med. 2022;12(2):585–587.
81. Vickers K, Palmisano B, Shoucri B, Shamburek R, Remaley A. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat Cell Biol. 2011;13(4):423–433. doi:10.1038/ncb2210
82. Jian F, Peng Y, Bian M. Expression and bioinformatics analysis of key miRNAs in stored red blood cells. Transfus Med Hemother. 2022;30(3):1–8.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.