Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 7
MicroRNA binding site polymorphisms as biomarkers in cancer management and research
Authors Cipollini M, Landi S, Gemignani F
Received 31 January 2014
Accepted for publication 2 May 2014
Published 23 July 2014 Volume 2014:7 Pages 173—191
DOI https://doi.org/10.2147/PGPM.S61693
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Monica Cipollini, Stefano Landi*, Federica Gemignani*
Department of Biology, University of Pisa, Pisa, Italy
*These authors contributed equally to this work
Abstract: MicroRNAs (miRNAs) are important regulators of eukaryotic gene expression. They have been implicated in a broad range of biological processes, and miRNA-related genetic alterations probably underlie several human diseases. Single nucleotide polymorphisms of transcripts may modulate the posttranscriptional regulation of gene expression by miRNAs and explain interindividual variability in cancer risk and in chemotherapy response. On the basis of recent association studies published in the literature, the present review mainly summarizes the potential role of miRNAs as molecular biomarkers for disease susceptibility, diagnosis, prognosis, and drug-response prediction in tumors. Many clues suggest a role for polymorphisms within the 3' untranslated regions of KRAS rs61764370, SET8 rs16917496, and MDM4 rs4245739 as SNPs in miRNA binding sites highly promising in the biology of human cancer. However, more studies are needed to better characterize the composite spectrum of genetic determinants for future use of markers in risk prediction and clinical management of diseases, heading toward personalized medicine.
Keywords: miRSNP, 3'-UTR target binding site, cancer risk, biomarkers
Introduction
MicroRNAs (miRNAs) are short, noncoding RNAs of 22–27 nucleotides that regulate gene expression through binding to cognate sequences, preferentially 3′ untranslated regions (UTR) regions, of mRNAs. The degree of complementarity around nucleotides 2–7 of a miRNA, the “seed” region, is the most important known determinant of recognition of an mRNA by a targeting miRNA.1 Based on the importance of seed pairing, multiple bioinformatics algorithms have been developed to predict miRNA-binding sites in mRNA sequences.2–6 Alterations of miRNA::mRNA interactions, although restricted to a few nucleotides, can have profound effects on the control of gene expression, as showed by Clop et al.7 These authors report, for the first time in mammals, that the G to A transition within the 3′-UTR of GDF8 creates a target site for mir-1 and mir-206, causing a translational inhibition of the myostatin gene and, hence, contributing to the muscular hypertrophy of Texel sheep.7 Following this and other evidence, it was hypothesized that a miRNA::mRNA pairing could be affected by a number of factors, including miRNA expression levels, the presence of SNPs within miRNA genes, and the presence of SNPs located within miRNA-binding target sites, now defined as miRSNPs and typically located at the 3′-UTR of mRNAs. In the same year Clop et al published their report, Chen et al found that negative selection in humans is stronger on computationally predicted conserved miRNA binding sites than on other conserved sequence motifs in 3′-UTRs, providing independent support for the target prediction model and explicitly demonstrating the contribution of miRNAs to Darwinian fitness.8 In 2007, Mishra et al demonstrated that variant DHFR 829C>T (rs34764978), falling near a miR-24 binding site, causes the overexpression of human dihydrofolate reductase, contributing to methotrexate resistance.9 A year later, for the first time, a putative miRSNP was also associated with the risk for colorectal cancer among Caucasians.10 Thus, in recent years, there has been increasing interest in the role of posttranscriptional regulation of gene expression by miRNAs11 and in the influence of miRSNPs on cancer risk12,13 and clinical outcomes.14–16 Actually, a growing number of studies have suggested that miRSNPs constitute a promising novel class of polymorphic variations worth investigation, with the potential of opening new areas of research in cancer biology and clinical oncology.17 Moreover, it has been suggested that miRSNPs could be employed as useful biomarkers in the study of disease progression, patient prognosis,18 and treatment efficacy of cancer.19,20
This article focuses on the studies of miRSNPs related to cancer. The review includes case–case studies in which the associations between miRSNPs and clinical outcome were evaluated. Moreover, case-control association studies were also included, with the aim of suggesting a possible role for miRSNPs in increased susceptibility to cancer. A concise resume of the results is reported in Table 1.
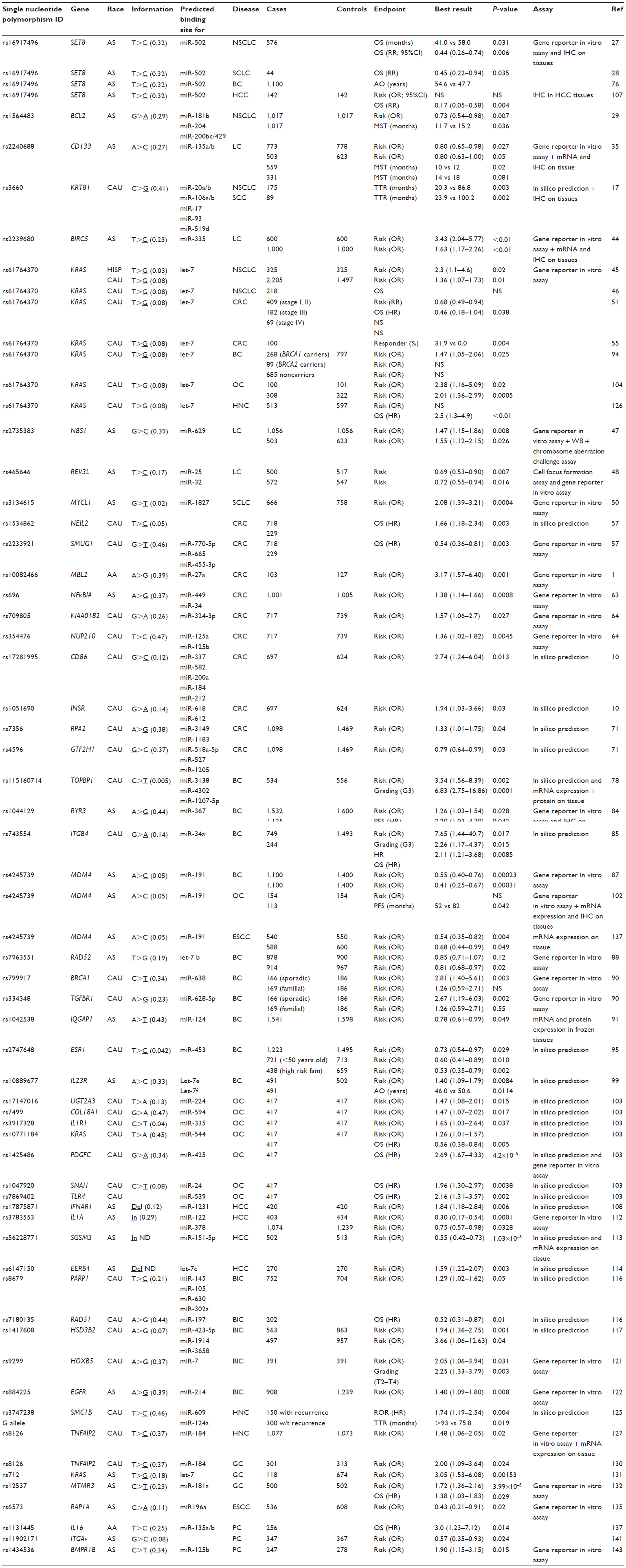 | Table 1 List of micro-RNA binding sites evaluated in epidemiologic studies |
Lung cancer
miRSNPs as biomarkers of prognosis in lung cancer
Lung cancer (LC) continues to be the leading cause of cancer-related deaths worldwide because of its high incidence, malignant behavior, and lack of major advancements in treatment strategy. Non-small-cell lung cancer (NSCLC) accounts for about 80% of all cases, with less than 15% of patients surviving beyond 5 years.21 Thus, specific prognostic biomarkers to be added to the standard tumor, node, and metastasis staging system may improve the medical care of patients with NSCLC.22–25 Because SET8 is found overexpressed in various types of tumor, including LC,26 Takawa et al evaluated whether SET8 rs16917496 T>C, a miRSNP falling within miR-502 binding site, could be associated with the overall survival (OS).26 Indeed, in a Chinese population, the CC genotype was associated with longer OS and reduced risk for death for NSCLC.27 The results are consistent with the in vitro observations that the variant C allele may decrease the expression of SET8 through enhancing the binding capacity of miR-502.27
The same SNP was further analyzed in 44 cases and 44 controls, and the C allele was confirmed to be independently associated with longer OS in NSCLC patients.28 Another study provided the evidence that the rs1564483 A allele, located within the 3′-UTR of BCL2, was associated with a significantly lower risk for LC in male Chinese patients and with a favorable OS in advanced NSCLC patients.29 This effect was more obvious in smoking patients, in stage IIIA patients, and in patients without surgery undergone to chemotherapy or radiotherapy. The authors speculated that the rs1564483 G-to-A substitution might change the stem-loop structure of 3′-UTR or introduce a miRNA binding site, which may affect BCL2 mRNA stability or its expression levels.29 Several miRNAs, including miR-181b,30 miR-200bc/429,31 and miR-204,32 have been reported to bind to the BCL2 3′-UTR in correspondence with the miRSNP, thereby modulating BCL2 mRNA levels. When the expression of BCL2 is decreased, the balance between pro- and antiapoptotic pathways could be shifted in favor of a proapoptotic activity, thus contributing to protecting lung cells from genotoxicants and carcinogenesis.33,34
Another miRSNP, the rs2240688, within the 3′-UTR of CD133 was significantly associated with a decreased risk for LC in Asian patients: compared with individuals with the AA genotype, those patients with CA or CC genotypes showed a 20% decreased risk in an exploratory sample set, confirmed in an independent sample set of validation.35 In addition, the C allele conferred a significantly favorable prognosis, with the median OS of CC + CA patients significantly longer than that found in patients carrying the AA genotype. Functional assays revealed that the A>C transversion creates a new binding site for miR-135a/b and causes a decreased expression of the CD133 mRNA.35 CD133 has been identified as a pleiotropic marker of cancer cells stemness in various human tumors. Both clinical analyses and laboratory studies have shown that CD133 plays a critical role in tumorigenesis and tumor progression.36,37 Increased CD133 expression was observed in several human cancer tissues38,39 and was reported to be associated with poor prognosis. In another study on NSCLC, significant differences in time to recurrence were found when Caucasian patients were analyzed for KRT81 rs3660 (20.3 months for the CC genotype versus 86.8 months for the CG + GG genotypes), especially among patients at stage I.17 The SNP is located within predicted binding sites of several miRNAs, including miR-17, miR-93, miR-20b, miR-519d, miR-520g, miR-520h, miR-519c-3p, miR-519b-3p, miR-519a, and miR-765, some of which were shown to be deregulated in NSCLC.40 KRT81 encodes for a protein known as Hb-1, a type of hair keratin that is physiologically expressed in hair shafts. Keratins are proteins expressed in all types of epithelial cells,41 with different expression patterns among different carcinomas,42 and they are extensively used as diagnostic markers.
In addition, survivin was studied for its miRSNPs. Survivin is overexpressed in many types of human cancer, including LC, and is considered a promising therapeutic target.43 Interestingly, the expression of its encoding gene, BIRC5, was found to be correlated with a SNP (rs2239680) within the 3′-UTR in normal lung tissues.44 This putative miRSNP was evaluated in two independent sets of samples from a Chinese Han population, and the C allele was associated with a significantly increased risk for LC and advanced staging. Furthermore, a reporter gene assay showed that rs2239680 T>C change caused an altered regulation of BIRC5 mRNA expression through the effect on miR-335::mRNA pairing.
miRSNPs as biomarkers of susceptibility to LC
A study showing the importance of miRSNPs within the 3′-UTR of the protooncogene KRAS was presented in 2008.45 In this region, there are at least 10 different target sites for one of the first discovered miRNAs (let-7), and one of them, rs61764370, within the let-7 complementary site 6 (LCS6) was extensively studied in relation to several types of cancer, including LC. This miRSNP was evaluated for its association with the risk for NSCLC in two independent sample sets: one consisting of 325 cases and 325 controls from New Mexico45 and other replicating the findings on 2,205 cases and 1,497 controls of Caucasian origin.45 The LCS6 variant allele was significantly associated with increased risk for NSCLC among moderate smokers. However, rs61764370 was not found to be associated with OS in LC patients, suggesting a poor clinical utility in NSCLC.46
The CC genotype of rs2735383, a functional miRSNP within the 3′-UTR of the NBS1 gene, was associated with a significantly increased risk for LC when compared with GG or GC genotypes in a study consisting of 1,559 cases and 1,679 controls, all Han Chinese.47 The CC genotype caused a decrease of the mRNA expression through the alteration of the miR-629 binding site. In turn, a deficient expression of NBS1 may induce deficiencies in the DNA repair and increased mutagen sensitivity, providing a possible explanation for its relationship with the risk for LC. In another Chinese study (1,072 patients and 1,064 cancer-free controls), the variant C of rs465646 within the 3′-UTR of REV3L was associated with decreased risk for LC.48 REV3Lp constitutes the catalytic subunit of DNA polymerase zeta, the major participant in trans-lesion DNA synthesis, one error-prone DNA repair system.49 Consistent with this role, T allele showed a stronger binding affinity for miR-25 and miR-32, resulting in significantly weaker reporter expression levels, as confirmed by additional experiments.48 Another case-control association study showed that rs3134615 T allele within the 3′-UTR of MYCL1 was associated with a significantly increased risk for small cell lung cancer (SCLC).50 This miRSNP is located within the binding site for miR-1827. The G>T change may inhibit the interaction of miR-1827 with MYCL1 mRNA, resulting in higher expression of MYCL1. Because MYCL1 is a member of the MYC oncogene family, which plays a critical role in carcinogenesis, individuals carrying the rs3134615 T allele are expected to have elevated risk for the development of SCLC. miR-1827 may play a role in lung carcinogenesis by functioning as a tumor suppressor, and further studies of this miRNA in cancer are warranted.
Colorectal cancer
miRSNPs as biomarkers of prognosis in colorectal cancer
Colorectal cancer (CRC) is the second most common cancer and the fourth-leading cause of cancer death worldwide.51 The tumor node, and metastasis staging system is currently the main tool to provide prognostic information, being highly predictive for prognosis at the extremes, although less predictive for intermediate stages.52,53 According to current guidelines, adjuvant chemotherapy is not administered to early-stage patients (T1-3-N0-M0), as 5-year OS is more than 70%. Nevertheless, 20%–30% of them will die of CRC within 5 years. Unfortunately, molecular markers enabling us to detect these aggressive forms are lacking. Several authors undertook studies to evaluate whether miRSNPs could represent prognosis markers to be used to this end.
The KRAS rs61764370 (within LCS6) was evaluated in the early CRC stages in the prospective Netherlands Cohort Study.51 The T allele was associated with a higher CRC risk and a shorter OS compared with the G allele. In patients with advanced disease, no clear associations were observed.
Response to therapy
rs61764370 also was associated with the response to cetuximab, a monoclonal antibody directed toward the epidermal growth factor receptor (EGFR).54 In a recent study on Caucasians, it was shown that among patients with wild-type KRAS, 31.9% of those with the rs61764370 TT genotype presented a complete or a partial response, whereas none of those with the TG + GG genotypes responded.55 This finding adds knowledge of the role of KRAS in the cure of the disease. However, these findings were not in agreement with the results obtained in a previous work.56 In summary, KRAS genotypes deserve further validation as prognostic biomarkers and consideration in therapy decision-making, especially for early-stage patients.
Concerning the most commonly used therapies based on 5-fluorouracil (5-FU), miRSNPs in BER genes were evaluated, and interesting results were found for miRSNPs rs1534862 within NEIL2 and rs223392 within SMUG1.57 Both miRSNPs were found to be associated with OS, with the stronger association for TT homozygotes of rs223392 after stratification for 5-FU-based chemotherapy. This is in agreement with the fact that SMUG1 and NEIL2 are among the main DNA glycosylases involved in the response to damages induced by 5-FU.58–61 A functional in vitro assay showed that the SMUG1 T allele, compared with the G allele, caused a reduced expression of a reporter gene. Thus, it was suggested that SMUG1 excision activity, modulated by miRSNP rs223392, could affect the toxicity caused by 5-FU.58
miRSNPs as biomarkers of susceptibility in colorectal cancer
Chronic intestinal inflammation has been identified as a risk factor for CRC.62 Therefore, it is possible that functionally important genetic variants of inflammatory mediators, such as mannose-binding lectin 2 (MBL2), are also associated with susceptibility to CRC. Four MBL2-specific allele variants in linkage disequilibrium located in the 3′-UTR region of the gene were associated with a higher risk for CRC in African-Americans.1 In particular, C allele of rs10082466 was associated with increased risk, and it was predicted to create a novel binding site for miR-27a and miR-27b. The increased binding affinity predicted for the C allele of rs10082466 was reflected by a significant decrease in normalized luciferase activity compared with the negative control. More important, the C allele of rs10082466 was associated with lower plasma MBL levels and activity in cases and controls, as would be expected for a regulatory interaction involving a germ-line polymorphism. miR-27a binds more efficiently to the C allele, which is consistent with the observed lower plasma MBL levels and activity.
Nuclear factor kB (NFkB) plays a key role in the regulation of apoptosis. The function of NFkB is inhibited by binding to the NFkB inhibitor, and the disruption of the balance of NFkB and the NFkB inhibitor is related to the development of many diseases, including tumors. Therefore, it was hypothesized that SNPs within the 3′-UTR of NFkBIA were associated with CRC susceptibility.63 Both A>G polymorphisms (rs696) were associated with an increased risk for CRC among Chinese patients. For rs696, the GG genotype was associated with a statistically significantly increased risk compared with AA + GA. Moreover, the authors found that the change from A to G in the 3′-UTR of NFkBIA decreased luciferase activities, as assessed by an in vitro reporter assay. These experiments suggested that NFkBIA 2758 A>G variants may affect mRNA stability, likely generating a novel seed site for miR-449a. More experiments showed that miR-449a reduced the relative luciferase activities via the NFkBIA 3′-UTR target site created by the A allele. The results indicate that A allele strengthens the binding of miR-449a with 3′-UTR of NFkBIA, which in turn inhibits the expression of NFkBIA. This polymorphism could be a genetic marker for susceptibility to CRC.
In another case-control association study on Caucasians, the AA homozygotes for rs709805 (within the predicted gene KIAA0182) showed an increased CRC risk compared with in the GG + GA group, and the CC homozygotes for rs354476 (NUP210) had an increased risk compared with the TT + TC group.64 In vitro assays carried out to test the differences between the common and variant 3′-UTRs of NUP210 and KIAA0182 showed that only the T allele of rs354476 was associated with a reduced expression of the reporter gene. NUP210 encodes the nuclear pore glycoprotein 210 involved in the structural organization of the nuclear pore complex.65 During mitosis, Ser1880 of glycoprotein 210 is phosphorylated by the cyclin B-p34cdc235.66 An increased expression of NUP210 was found also in other types of cancer, such as cervical cancer.67
Positive associations between risk for CRC and two miRSNPs (CD86 rs17281995 and INSR rs1051690) were also found in another study on Caucasians.10 Five different miRNAs (miR-337, miR-582, miR-200a, miR-184, and miR-212) bind to a target site that contains the same polymorphism within CD86. CD86, with CD80, is a costimulatory ligand expressed on the surface of the antigen-presenting cells (dendritic cells, macrophages, and B cells) in the immune system.68 Two different miRNAs bind to the same polymorphism within INSR: miR-612 and miR-618. INSR encodes for insulin receptor, and after the binding of insulin to the extracellular portion, a second messenger system diverges into two separate pathways that regulate distinct biological effects: the phosphoinositide-3-kinase pathway and the mitogen-activated protein kinase pathway.69
In addition, SNPs residing within the 3′-UTRs of genes involved in pathways such as DNA repair, DNA signaling, or apoptosis may indirectly contribute to affecting the individual risk of developing CRC.45,70 The role of miRSNPs in genes specifically involved in the nucleotide excision repair (NER) pathway was investigated. rs7356 within RPA2 and rs4596 within GTF2H1 were associated with rectal cancer risk in Caucasian patients.71 rs4596 is located within a target region of the predicted miR-518a-5p and miR-527 that showed the highest energy-binding level to 3′-UTR with the G allele. Interestingly, findings from association studies showed that G-allele carriers were at decreased cancer risk. The GTF2H1 (general transcription factor IIH, polypeptide 1, 62 kDa) encodes for a component of the core-TFIIH basal transcription factor involved in NER and, when in complex with Cdk-activating kinase (CAK), also is involved in RNA transcription. Variant A allele of rs7356 in RPA2 (replication protein A 32 kDa subunit) was associated with increased risks. The G allele of rs7356 is more prone to bind miR-3149 and miR-1183, eventually resulting in a stronger negative regulation on target gene expression.
Breast cancer
miRSNPs as biomarkers of prognosis in breast cancer
Breast cancer (BC) is the most frequently diagnosed cancer and one of the leading causes of cancer death among women worldwide.72,73 Germ-line mutations in BRCA1 and BRCA2 account for only 5% of all BC cases in the general population.74 Other low-penetrance genetic variants, especially in as-yet-unknown combinations, are expected to explain most BC incidence.75 Investigators have hypothesized that the 3′-UTRs of miRNA target genes may harbor part of these variants. The genotype CC of SET8 rs16917496, a BC candidate gene, was associated with earlier age of onset when compared with TT in Asian patients.76 Another candidate miRSNP, rs115160714, within TOPBP1, encoding for topoisomerase IIb binding protein 1 (TopBP1), was evaluated in relation to BC by Forma et al in Caucasian patients.77,78 Heterozygotes (CT) and homozygotes (TT) had significantly increased risk for BC compared with common homozygotes (CC). Moreover, patients with a tumor classified as high grade (G3) or T2-T4N1M0 were carriers of the variant allele (T) more often than expected. In agreement with these findings, TOPBP1 mRNA and protein expression were found to be increased in individuals with the CT or TT genotype. Three candidate miRNAs, miR-3138, miR-4302, and miR-1207-5p, were predicted to bind to the 3′-UTR of TopBP1. Thus, the study raised the hypothesis that a genetic variation of TopBP1 may be involved in the etiology of BC. The biological bases for explaining its role in BC could rely on the fact that TopBP1 shares structural functional similarities with BRCA1 and is involved in cell survival, DNA replication, DNA damage repair, and cell cycle checkpoints.79,80
Because calcium and vitamin D intake were associated with mechanisms of carcinogenesis of the mammary gland,81,82 and in addition, breast calcifications are an important risk factor for BC,83 Zhang et al84 evaluated the role of miRSNPs within the 3′-UTR of RYR3, a CICR (calcium-induced calcium release) protein playing a crucial role in cellular Ca2+ homeostasis. After the analysis of 1,532 breast cancer cases and 1,600 healthy Chinese women, rs1044129 was found to be associated with BC risk, calcification, and progression-free survival. These findings were also supported by in vitro assays showing that miR-367 binds more tightly to the A allele of rs1044129 than to the G allele and represses RYR3 expression more strongly.
Concerning the role of integrins for BC, a Swedish study evaluated whether miRSNPs within the 3′-UTR of ITGA3, ITGA6, ITGAv, ITGB3, ITGB4, and ITGB5 genes could be associated with BC clinical outcome and risk.85 Detailed clinical data of 749 Swedish incident patients with follow-up within 15 years were evaluated and compared with data from 1,493 matched controls. The strongest association was observed between the rare A allele of the SNP rs743554 within ITGB4 and the risk for estrogen receptor-negative carcinomas. The same allele also was associated with worse OS compared with the common allele. None of the remaining putative miRSNPs were significantly associated with BC risk. In silico analysis predicted that A allele may cause a loss of the binding site for the miR-34a. The association between the ITGB4 and hormone-receptor status may be explained by the fact that integrin-mediated signal transduction pathways regulate estrogen receptor α (ER-α) in mouse mammary epithelial cells.86
miRSNPs as biomarkers of susceptibility in breast cancer
Several studies investigated the role of miRSNPs as possible risk factors for BC. In 1,100 BC cases and 1,400 controls from two regions of China, the AC and CC genotypes of MDM4 rs4245739 were significantly associated with decreased BC risk compared with the AA genotype.87 In another Chinese study, the variant C allele rs7963551 within the 3′-UTR of RAD52 was associated with a reduced BC risk.88 Luciferase activity assay showed a higher expression level for C allele compared with A allele, which might be a result of a reduced inhibition from a weakened binding capacity of miRNA to the 3′-UTR of RAD52 harboring C allele. These findings suggested that rs7963551, one of the miRSNPs located with a let-7 binding site, may alter expression of RAD52, contributing to the development of BC. RAD52 could play a crucial role for BC, considering its role in the homologous recombination repair in cooperation with BRCA1 and BRCA2.89 For these latter genes, Nicoloso et al90 reported that T allele rs799917 within the 3′-UTR of BRCA1 is associated with susceptibility to BC in Caucasians and that this risk is particularly increased for the sporadic form. The authors showed that miR-638 interacts more strongly with C allele of SNP rs799917 than with the T allele; this difference was also confirmed at the protein level.90
Concerning the pathway of the mitogen-activated protein kinase, several pieces of evidence suggested that deficiencies of IQGAP1 (IQ motif-containing GTPase-activating protein 1) could be related to cancer development. Thus, the genotypes of IQGAP1 were evaluated in a Chinese association study on 1,541 BC patients and 1,598 controls.91 The TT genotype of the putative miRSNP rs1042538 was associated with a significantly lower risk for BC compared with the AA genotype. The results were consistent with the finding that the expression levels of IQGAP1 protein were significantly higher in the TT genotype. Lim et al92 verified experimentally that the A-to-T variant disrupts a miRNA target site for miR-124, and thus the A allele causes miR-124 to bind more tightly with IQGAP1 mRNA, leading to the down-regulation of the encoded protein. Functional studies established that IQGAP1 interacts with and regulates the actin-Cdc42/Rac1-mitogen-activated protein kinase pathway, contributing to its role in cell migration and invasion.93 In the same pathway, the association between miRSNPs KRAS rs61764370 and BC risk also was evaluated. A German study was carried out on 268 BRCA1-positive families, 89 BRCA2-positive families, 685 BRCA1/BRCA2-negative families, and 797 geographically matched controls.94 The allele frequency of the KRAS variant was found to be increased only among patients with BC positive for BRCA1 mutations (compared with controls). However, when a larger sample set was analyzed by including other family members in addition to the index cases, the association could not be replicated.
In another German study of 1,223 BC families and 1,495 unrelated controls, a significant association was revealed for the T allele within the 3′-UTR of ESR rs2747648, and in particular, among premenopausal women.95 According to in silico analyses, T allele attenuates the binding of miR-453, leading to higher ESR1 protein levels. ESR1 is a member of the nuclear receptor family, a group of hormone-inducible transcription factors that activates gene expression by recruiting multiple coactivators. Clinical studies have shown that depletion of ESR1 significantly reduces BC risk, providing further support for the observed protective effect of the C allele in premenopausal women.96,97
Other studies on BC susceptibility focused on the role of immunity and inflammation. Interleukin 23 (IL23) and its receptor (IL23R) guide T cells toward a T-helper type 17 (Th17) phenotype characterized by IL-17A production.98 A microenvironment constituted by tumor-infiltrating cells releasing high levels of IL-17 is a poor prognostic factor for BC. Thus, the effect of the miRSNP IL23R rs10889677 was evaluated in Chinese Han women, and the A allele was associated with an increased risk for BC.99 Zwiers et al reported that A allele could determine the loss of binding capacity for the miRNAs let-7e and let-7f.100
Ovarian cancer
miRSNPs as biomarkers of prognosis in ovarian cancer
Epithelial ovarian cancer (EOC) is the fifth most common cancer in women.101 At the time of diagnosis, more than 80% of patients present late-stage malignancies with a survival rate less than 30% at 5 years.102 A recent study on Chinese patients hypothesized that MDM4 could affect chemosensitivity and progression of EOC.102 The authors found that the common AA genotype rs4245739 is more frequent in patients with high-grade carcinomas and that when the analysis was stratified considering cases not expressing the estrogen receptor, patients with the AA genotype have an increased risk for recurrence and tumor-related death. Further analyses showed that the miRSNP creates a putative target site for miR-191 (a miRNA highly expressed in normal and tumor tissues) and that this acquisition causes downregulation of MDM4 expression, thereby significantly delaying ovarian carcinoma progression and tumor-related death.
In another study on 417 Caucasian cases and controls, the authors investigated 238 SNPs from eight miRNA processing genes and 134 genes for EOC predisposition and association with clinical outcome and treatment response.103 Four miRSNPs (UGT2A3 rs17147016, COL18A1 rs7499, IL1R1 rs3917328, and KRAS rs10771184, alias rs12245) were associated with increased risks. The variant allele of KRAS was also associated with longer OS and favorable treatment outcome. Moreover, the variant alleles within PDGFC rs1425486 and TLR4 rs7869402 were associated with poor response to therapies. PDGFC is a member of the platelet-derived growth factor family, which encodes a mitogenic factor for cells of mesenchymal origin, and with an in vitro reporter assay, the authors demonstrate that rs1425486 alleles could differentially affect miR-425 targeting in ovarian cancer cells, suggesting that PDGFC is a putative target for miR-425.
miRSNPs as biomarkers of susceptibility to ovarian cancer
The Connecticut Ovarian Cancer Case-Control study, consisting of 320 patients and 328 controls, mostly from Northern Italy, showed a significant increased risk of developing EOC for carriers of KRAS G allele rs61764370.104 In EOC, KRAS overexpression was shown to disrupt the EGFR-signaling pathway, a pathway found frequently deregulated in the disease. This could provide a rationale for explaining the association. However, no increased risks were observed in a larger study of 8,669 cases and 10,012 controls, mostly from Northern Europe and Northern America.101 Although false-positive results of the Connecticut study could not be ruled out because of the small sample size, positive associations could be also ascribed to regional differences. It is quite impressive that when the analysis was stratified considering patients affected by a familial form of EOC and negative for mutations within BRCA1 or BRCA2 (ie, 31 diagnosed for hereditary breast and ovarian cancer syndrome), the frequency of the carriers of the variant allele was much higher (61%) compared with what was expected (14.5%).104 Although the study was somewhat limited by the small number of uninformative patients, these findings support the hypothesis that the G allele could be a genetic marker of increased risk for hereditary breast and ovarian cancer.
Hepatocellular carcinoma
mirSNPs as biomarkers of prognosis to hepatocellular carcinoma
Hepatocellular carcinoma (HCC) is the fifth most common malignancy worldwide and the third leading cause of cancer death.105 Because of its high fatality, its incidence and mortality rates are almost equal. The incidence of HCC is rising steeply in Asia and Africa, where hepatitis B and C viruses (HBV and HBC) are more prevalent.106 The SNP in the miR-502 binding site of the SET8 3′-UTR was examined for its predictive power relative to HCC outcomes.107 The SET8 CC genotype was associated with longer postoperative OS in Asian patients with HCC and with reduced SET8 protein levels, according to the immunostaining of 51 HCC tissue samples. C allele located within the seed region was perfectly matched with G allele in miR-502. These data suggest that an altered expression of SET8, at least in part ascribed to miR-502, affects HCC outcome. Thus, an analysis of genetic polymorphisms within miRNA binding sites may help identify patient subgroups with poor prognosis.
miRSNPs as biomarkers of susceptibility to hepatocellular carcinoma
HBV and HCV are the main etiological risk factors for HCC worldwide.106 Thus, several studies were undertaken to ascertain the role of miRSNPs involved in modulating the inflammation and/or in the immune response to these infections. A study of 420 Chinese patients and unrelated controls showed that the 4-bp In/Del miRSNP rs17875871 within the 3′-UTR of IFNAR1 was associated with the risk for HCC and that the association was more pronounced in a subgroup of patients positive for B-hepatitis.108 In silico predictions suggested that rs17875871 is located in the seed region of a miR-1231 predicted target sequence. IFNAR1 encodes for a membrane protein needed to compose the receptors for interferon alpha and beta. The binding with the ligand activates STAT1 and STAT2 cascades.109 Several pieces of evidence showed the involvement of IFNAR1 in HBV and HCV replication and chronic infection.110 Genetic polymorphisms within IFNAR1 were also found to be associated with clinical presentation and outcomes after HBV infection.111 Thus, IFNAR1 may affect the inflammatory process related to HBV and HCV infections, thereby contributing to HCC susceptibility.
In another study, the miRSNP rs3783553 within IL1A 3′-UTR was evaluated in association with HCC in two independent Asian populations. The variant allele consisting of a 4-bp (TTCA) insertion was associated with decreased risks.112 In addition, it was shown in vitro and in vivo that this allele disrupts the binding sites for miR-122 and miR-378, thereby increasing the expression of IL-1a. These findings suggest that functional polymorphism rs3783553 could contribute to HCC susceptibility. Considering that IL-1a affects not only various phases of the malignant process, such as carcinogenesis, tumor growth, and invasiveness, but also patterns of interactions between malignant cells and the host’s immune system, these results indicate that IL-1a may be a promising target for immunotherapy, early diagnosis, and intervention of HCC.
Concerning pathways of intracellular signaling, a Chinese study carried out on rs56228771 found an association between a 4-bp In/Del miRSNP within the 3′-UTR of SGSM3 and a significantly decreased risk for HCC.113 Tissue samples with In/In genotype had the highest levels of SGSM3, about 1.52-fold and 2.93-fold higher than that with In/Del and Del/Del genotype, respectively. Bioinformatics predictions showed that the insertion allele disrupts a binding site for miRNA-151-5p and, thus, causes SGSM3 to upregulate. Moreover, an association between HCC susceptibility and a 12-bp In/Del polymorphism rs6147150 (within the 3′-UTR of ERBB4) was found: carriers of the Del allele had a 1.59-fold increased risk for HCC. Bioinformatics analysis suggests that rs6147150 lies within a predicted binding site for let-7c, and it could be hypothesized that let-7c tightly binds ERBB4 transcripts containing the 12-bp deletion allele, negatively regulating ERBB4 expression.114 These findings suggested that common genetic polymorphisms in ERBB4 may affect HCC risk, at least in part via let7c-mediated regulation, which may be involved in the pathogenesis of HCC.
Bladder cancer
miRSNPs as biomarkers of prognosis or response to therapy in bladder cancer
Bladder cancer (BlC) is the fourth most common cancer among men in the United States and accounts for 3% of the cancer deaths.72 The fact that BlC incidence is three to four times higher in men suggests that risk may be modified by hormone levels.115 Among BlC patients of Caucasian origins undergoing radiotherapy, carriers of the RAD51 rs7180135 minor allele showed longer OS.116 miR-197 was predicted to bind more tightly to the G allele, likely resulting in a reduction of RAD51 expression. RAD51 is involved in DSB homologous recombination repair; thus, in carcinoma, alteration of RAD51 expression could potentiate radiosensitization. Low RAD51 mRNA expression has previously been associated with lower local recurrence and improved survival after adjuvant chemotherapy and radiotherapy treatments in BC.116 The results suggest that the RAD51 rs7180135 genotype, through the alteration of miR-197 binding site, may affect radiosensitivity and radiotherapy outcome in BlC. If successfully validated, this might be used clinically as a predictive marker of radiotherapy outcome. In the same study, rs8679 in 3′-UTRs PARP1 was associated with risk for BlC. The variant was predicted to decrease the strength of binding with miR-145, possibly increasing PARP1 expression.
miRSNPs as biomarkers of susceptibility to bladder cancer
In agreement with the hypothesis that a key role in modulating this risk could be played by hormone level, the A allele of miRSNP rs1417608 within the 3′-UTR of the hormone regulation gene 3-beta-hydroxysteroid dehydrogenase type 2 (HSD3B2) was associated with nearly a 2-fold increased risk for BlC in patients of Caucasian origins.117 HSD3B2 encodes a NAD+, dependent microsomal enzyme that catalyzes biosynthesis of dihydrotestosterone and dihydroprogesterone. Interestingly, this association was confirmed in another study (the Texas BlC Study), in which the SNP rs1341015, in linkage disequilibrium with rs1417608, was evaluated. Moreover, two other SNPs in strong linkage disequilibrium with rs1417608 (rs1819698 and rs1538989) were associated with significantly increased risks for prostate cancer in people of European descent.118 The variant allele of rs1819698 was also computationally predicted to disrupt a miRNA binding site for miR-3658. In summary, all these observations support the notion that hormone synthesis deregulation, through HSD3B2, may be important in the etiology of BlC.
In recent years, the HOX gene family has also been associated with human diseases, especially cancers. For instance, HOXB5 has been reported to be related to human diseases, including acute myeloid leukemia,119 EOC,120 and urological carcinomas. In BlC, the frequency of the G allele of HOXB5 rs9299, a miRSNP falling within miRNA-7 binding site, was higher among Caucasian patients with BlC compared with healthy controls, and it was found to be correlated with the risk for high grade and high stage.121 The expression of the HOXB5 mRNA with the G allele was significantly higher than the mRNA with the A allele in both cancer tissues and cell lines. In summary, the results suggest this miRSNP may affect HOXB5 expression, which in turn may affect bladder tumorigenesis.
In another study, the association between EGFR miRSNP rs884225 T>C and BlC risk was examined among Chinese patients.122 Results showed that the CC genotype was associated with a significantly increased risk compared with TT + TC genotypes. In addition, luciferase reporter gene assay confirmed that T-to-C substitution could increase the EGFR expression. On the basis of the bioinformatics analysis, rs884225 polymorphism lies within a predicted binding site for miR-214; however, in vitro experiments could not confirm such a prediction, and thus it is unclear whether rs884225 is a functional polymorphism or is a proxy for other variations nearby.122 In any case, much evidence shows that overexpression of EGFR plays an important role in regulating carcinogenesis by mediating cell mortality, apoptosis, tumor invasion, and metastasis. Thus, EGFR differential regulation at the 3′-UTR level might constitute a susceptibility factor that warrants being explored further.
Head and neck cancer
miRSNPs as biomarkers of prognosis in head and neck cancer
Squamous cell carcinoma of the head and neck (HNSCC), which includes cancers of the oral cavity, pharynx, and larynx, is one of the six most common cancers worldwide, accounting for 35% of all cancers in the United States.123 Most early-stage patients can be cured with surgery, radiotherapy, and chemotherapy. However, second primary tumors (SPTs) and local-regional recurrence negatively affect their long-term prognosis. It has been reported that 15%–25% of HNSCC patients will develop SPT/recurrence during the first 5 years after initial diagnosis.124 Thus, the development of clinical biomarkers predicting SPT/recurrence could be very important for the surveillance and targeted chemoprevention of high-risk patients. Genetic variations in miRNA-binding sites are reported to be associated with the risk for HNSCC and with SPT/recurrence in Caucasian patients with early stages.125 In particular, the rare homozygous genotype of the miRSNP rs3747238 within SMC1B was associated with an increased SPT/recurrence risk and reduced OS. The variant allele is predicted to create de novo binding sites for miR-609 and miR-124a, resulting in lower SMC1B expression. SMC1B is involved in chromosome structure maintenance during meiosis and mitosis, and its reduced expression could be related to potentially increased genome instability and greater cancer progression risk. For the same type of cancer, the genotype KRAS rs61764370, the miRSNP affecting a let-7 miRNA-binding site, was evaluated. Although no significant associations with the risk were described, the variant G allele was associated with a significantly reduced OS compared with a common allele.126 This observation suggested that the miRSNP could be associated with tumor progression, rather than initiation. Moreover, in the presence of the variant allele, KRAS expression was increased. Amplified KRAS promotes the growth in HNSCC, and its immune-histochemical positivity for K-ras protein was associated with late stages and increased tumor size.
miRSNPs as biomarkers of susceptibility in head and neck cancer
In HNSCC, the C allele of rs8126 within TNFAIP2 was associated with increased risks among Caucasians.127 This allele was also associated with higher mRNA expression levels of TNFAIP2 compared with the T allele in blood lymphocytes of 64 cases. rs8126 C allele was predicted to disrupt a miR-184 binding site, providing a possible explanation for the altered TNFAIP2 expression level.127TNFAIP2 mRNA is detectable in many human tissues and most hematopoietic cell lines.128 Although the function of the encoded protein is unknown, it was involved in apoptosis, and the gene was found induced by the tumor necrosis factor α in human endothelial cells.129 Moreover, TNFAIP2 was found to be highly expressed in nasopharyngeal carcinoma tumor cells when compared with adjacent normal tissues, and the increased expression of TNFAIP2 was significantly associated with shorter OS in nasopharyngeal carcinoma patients without distant metastasis.127
Other tumors
Gastric and esophageal cancer
Gastric cancer (GC) is one of the most common malignancies worldwide, accounting for 8% of total cancer cases and 10% of total cancer deaths, although both its incidence and mortality have been declining in the latest decade.72 Not many studies have investigated the role of miRSNPs in GC. However, interestingly, the genotype CC of rs8126 within TNFAIP2, already associated with increased risk of HNSCC, was also associated with increased risks for GC among Caucasians.130 Studies carried out on other miRSNPs revealed that the TT genotype of KRAS rs712, another miRSNP falling within a let-7 binding site within the 3′-UTR, was associated with an increased risk among Asian patients.131 Moreover, increased risks for GC were also found, among Asian patients, for the variant allele of rs12537, located within the 3′-UTR of MTMR3.132 This allele was also associated with poor OS, and GC tissues from carriers of the T allele showed lower MTMR3 mRNA expression levels than CC homozygotes. Luciferase assay revealed that miR-181a directly targeted MTMR3, and its suppressive effect was enhanced when the C allele was substituted by its T-variant. MTMR3, myotubularin-related protein 3, is ubiquitously expressed and has been demonstrated to regulate autophagy.133 However, little is known about its role on cancer.
Esophageal cancer (ESCC) is one of the most aggressive cancers, and its incidence worldwide has significantly increased in recent years.72 After complete surgical removal of the primary tumor, the 5-year survival rate is 50%–80% for stage I, 10%–40% for stage II, and 10%–15% for stage III disease. Patients with distant metastases (stage IV) who are treated with palliative chemotherapy have a median survival of less than 1 year.134 In an association study of Chinese patients, authors found that the CC genotype rs6573 within the 3′-UTR of RAP1A was associated with an increased risk compared with CA or AA genotypes.135 In addition, the C allele was more frequently represented among patients with stage III or IV disease. The change A>C in the binding site for miR-196a, was associated with a high constitutive expression of RAP1A. RAP1A is involved in a wide range of biological processes, including cell proliferation, differentiation, and cell motility and an abnormal RAP1A activation contributes to the tumorigenic processes. Expression of RAP1 at high levels can morphologically transform Swiss 3T3 fibroblasts and form tumors when injected into nude mice.136
rs4245739, a SNP within MDM4, was another miRSNP associated with the risk of developing ESCC.137 C allele had a significantly decreased risk compared with A allele in a Chinese population. In the genotype-phenotype correlation analysis of 29 human ESCC and paired esophagus tissue samples, AC + CC genotypes were associated with a statistically significant decrease of MDM4 mRNA expression. The change C to A was predicted to disrupt the interaction between miR-191 and mRNA, thereby increasing MDM4 expression in cancer cells.
Prostate cancer
Prostate cancer (PC) remains the second leading cause of cancer-related death in men in the United States.138 Androgen deprivation therapy is the most commonly used first-line treatment for advanced PC. Despite a generalized positive response, within 2–3 years, the disease progresses to a castration-resistant status in 20% of patients, and the life expectancy becomes approximately 16–18 months. A variety of prediction parameters, such as tumor stage, Gleason score, and prostate-specific antigen kinetics, have been used in clinical practice to define the presentation of PC and adapt the treatment strategy.139 However, their prognostic capabilities are still limited and might be improved by the incorporation of other markers. In the following work, the authors explored the role of miRSNPs in ALOX15, RAF1, IL-16, and IL-18 (genes of biologic relevance to PC) and were evaluated in a longitudinal screening of high-risk people.140 Analysis showed a statistically borderline association between IL-16 rs1131445 TT genotype and earlier age at diagnosis only among African-Americans, not Caucasians. Another study, carried out evaluating miRSNPs in five integrin genes, failed to show any association with PC prognosis; however, GC carriers of rs11902171 within 3′-UTR of ITGAv were associated with decreased risk among Chinese patients.141 Integrins have been implicated in the genesis and development of many tumors and act as indispensable partners of oncogenes in the transformation of normal cells.142 The transversion G to C in rs11902171 may alter targets for the mir-382, mir-30a-3p, and mir-30e-3p. Finally, the T allele of rs1434536 within the 3′-UTR of BMPR1B was found associated with the risk for localized PC in Asian.143 The association was stronger among patients older than 70 years, suggesting that rs1434536 was a more appropriate predictor for PC in older people. The C allele gave a reduced luciferase activity relative to the T allele, likely because the C-to-T substitution causes a reduced binding of miR-125b to BMPR1B mRNA. This gene encodes a member of the bone morphogenetic protein receptor family of transmembrane serine/threonine kinases, and its regulation could affect PC cells homing and growth at distant metastatic sites.144
Leukemia
Chronic lymphocytic leukemia is a B-cell malignancy and one of the most common non-Hodgkin lymphomas. About 69,740 cases of non-Hodgkin lymphoma are expected to be diagnosed in the United States in 2013.145 In the present study on 745 chronic lymphocytic leukemia Caucasian cases and 1,521 controls, the strongest association with chronic lymphocytic leukemia risk was observed with a common SNP located within the 3′-UTR of IRF8 (rs1044873, log additive odds ratio =0.7; P=1.81 × 10−6).146 rs1044873 is located in the 3′-UTR of the IRF8 gene and therefore is potentially located within a target region for miRNA. However, bioinformatics analysis did not support this. According to PolymiRTS, rs1044873 is not within any validated miRNA target. In addition, in acute myeloid leukemia, a polymorphic nucleotide T deletion is present in the 3′-UTR of NPM1, was associated with adverse outcomes, and could independently predict shortened survival in patients with de novo acute myeloid leukemia.147 In particular, patients carrying a homozygous delT genotype had higher relapse rates (59% versus 31%; P=0.051) and significantly shortened OS (median, 9 months versus 12 months; P=0.016) and relapse-free survival (median, 5 months versus 12 months; P=0.007) than patients carrying a non homozygous genotype. The nucleotide T deletion created an illegitimate binding NPM1 for miR-337-5p, which was widely expressed in different acute myeloid leukemia subtypes and inhibited NPM1 expression.
Discussion
MiRSNPs represent a promising class of genetic variations worth being deeply investigated as markers of individual susceptibility to complex diseases, to prognosis, and in clinically relevant decision-making. Although it should be acknowledged that most of the illustrated studies have not been replicated in independent laboratories yet, it also should be noticed that some of the findings reported here were consistent among different tumor types and had some experimental evidence based on in vitro assays. In particular, several variants within the 3′-UTR of the protooncogene KRAS were of interest. It has been shown that there are ten different LCSs within the 3′-UTR of KRAS mRNA to induce KRAS downregulation. A miRSNP within the sixth site (also termed KRAS-LCS6) has been identified and demonstrated to affect KRAS expression. The rs61764370 determines the change of the ancestral T allele to a G allele, disrupts the let-7 miRNA binding site, and causes an increased KRAS expression.45 This natural change might represent a mechanism of KRAS activation. This variant is relatively uncommon; in fact, is almost absent in Native Americans and East Asians, is very rare in Africans, and has a minor allele frequency of about 7% in the European populations.148
This variant allele is associated with increased risk for NSCLC,46 BC, and EOC. Other groups had also reported that the KRAS variants were associated with decreased OS in HNSCC and increased OS in NSCLC and, in addition, seem to modulate therapeutic responses in CRC patients.51,54 It is also worth noting that rs712 and rs10771184, miRSNPs falling within other LCSs of KRAS mRNA, were associated with increased risks for GC and EOC and OS. Thus, overall, there are many clues to the role of polymorphisms within KRAS 3′-UTR in the biology of human cancer. A miRSNP highly promising for its role in human tumors is rs16917496. This miRSNP, within the 3′-UTR of SET8, is associated with longer OS in patients with NSCLC27 and HCC107 and with the age at diagnosis of BC.76 The rational linking of the regulation of SET8 to cancer could reside in the multiple functions of the encoded protein that is involved in the advancement of cell cycle through the S-phase149 and in the transcriptional regulation,150 genome stability,151 apoptosis, and cell-cycle arrest.152 Most of these actions can be explained by SET8 having a well-defined function in the p53 pathway by mono-methylating p53 at lysine 382 and suppressing the p53-mediated transcription activation of target genes.153
Finally, it is worth mentioning here miRSNP rs4245739 within MDM4. The A allele was associated with the risk for BC87 and ESCC,132 as well as an unfavorable prognosis in EOC.103 Mdm4 was originally discovered as a p53-interacting protein through screening of a mouse embryo cDNA expression library.154 The p53-binding domain of Mdm4 interacts with the transactivation domain of p53 to represses its transcriptional activity. High levels of Mdm4 are also found in a variety of human cancers: HNSCC, retinoblastoma, melanoma, and BC. Mdm4 upregulation in malignancies is mostly ascribed to MDM4 gene amplification.155 However, the C-to-A substitution also could constitute an alternative mechanism of activation. In fact, it could disrupt the interaction between miR-191 and mRNA, thereby increasing MDM4 expression. In summary, a new emerging field of study supports the notion that gene regulation through miRNAs could explain part of the phenotypic variability observed in humans. More studies are needed to better characterize the composite spectrum of genetic determinants for a futuristic use of markers in risk prediction and clinical management of diseases, moving toward personalized medicine.
Disclosure
The authors report no conflicts of interest in this work.
References
Zanetti KA, Haznadar M, Welsh JA, et al. 3′-UTR and functional secretor haplotypes in mannose-binding lectin 2 are associated with increased colon cancer risk in African Americans. Cancer Res. 2012;72(6):1467–1477. | |
John B, Enright AJ, Aravin A, Tuschl T, Sander C, Marks DS. Human MicroRNA targets. PLoS Biol. 2004;2(11):e363. | |
Krek A, Grün D, Poy MN, et al. Combinatorial microRNA target predictions. Nat Genet. 2005;37(5):495–500. | |
Lewis BP, Shih IH, Jones-Rhoades MW, Bartel DP, Burge CB. Prediction of mammalian microRNA targets. Cell. 2003;115(7):787–798. | |
Rusinov V, Baev V, Minkov IN, Tabler M. MicroInspector: a web tool for detection of miRNA binding sites in an RNA sequence. Nucleic Acids Res. 2005;33(Web Server issue):W696–W700. | |
Kiriakidou M, Nelson PT, Kouranov A, et al. A combined computational-experimental approach predicts human microRNA targets. Genes Dev. 2004;18(10):1165–1178. | |
Clop A, Marcq F, Takeda H, et al. A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nat Genet. 2006;38(7):813–818. | |
Chen K, Rajewsky N. Natural selection on human microRNA binding sites inferred from SNP data. Nat Genet. 2006;38(12):1452–146. | |
Mishra PJ, Humeniuk R, Mishra PJ, Longo-Sorbello GS, Banerjee D, Bertino JR. A miR-24 microRNA binding-site polymorphism in dihydrofolate reductase gene leads to methotrexate resistance. Proc Natl Acad Sci U S A. 2007;104(33):13513–13518. | |
Landi D, Gemignani F, Naccarati A, et al. Polymorphisms within micro-RNA-binding sites and risk of sporadic colorectal cancer. Carcinogenesis. 2008;29(3):579–584. | |
Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat Rev Genet. 2008;9(2):102–114. | |
Chen K, Song F, Calin GA, Wei Q, Hao X, Zhang W. Polymorphisms in microRNA targets: a gold mine for molecular epidemiology. Carcinogenesis. 2008;29(7):1306–1311. | |
Saunders MA, Liang H, Li WH. Human polymorphism at microRNAs and microRNA target sites. Proc Natl Acad Sci U S A. 2007;104(9):3300–3305. | |
Hu Z, Chen J, Tian T, et al. Genetic variants of miRNA sequences and non-small cell lung cancer survival. J Clin Invest. 2008;118(7):2600–2608. | |
Christensen BC, Moyer BJ, Avissar M, et al. A let-7 microRNA-binding site polymorphism in the KRAS 3′ UTR is associated with reduced survival in oral cancers. Carcinogenesis. 2009;30(6):1003–1007. | |
Christensen BC, Avissar-Whiting M, Ouellet LG, et al. Mature microRNA sequence polymorphism in MIR196A2 is associated with risk and prognosis of head and neck cancer. Clin Cancer Res. 2010;16(14):3713–3720. | |
Campayo M, Navarro A, Viñolas N, et al. A dual role for KRT81: a miR-SNP associated with recurrence in non-small-cell lung cancer and a novel marker of squamous cell lung carcinoma. PLoS One. 2011;6(7):e22509. | |
Salzman DW, Weidhaas JB. SNPing cancer in the bud: microRNA and microRNA-target site polymorphisms as diagnostic and prognostic biomarkers in cancer. Pharmacol Ther. 2013;137(1):55–63. | |
Mishra PJ, Mishra PJ, Banerjee D, Bertino JR. MiRSNPs or MiR-polymorphisms, new players in microRNA mediated regulation of the cell: Introducing microRNA pharmacogenomics. Cell Cycle. 1, 2008;7(7):853–858. | |
Mishra PJ, Bertino JR. MicroRNA polymorphisms: the future of pharmacogenomics, molecular epidemiology and individualized medicine. Pharmacogenomics. 2009;10(3):399–416. | |
Xu J, Yin Z, Gao W, et al. Genetic variation in a microRNA-502 minding site in SET8 gene confers clinical outcome of non-small cell lung cancer in a Chinese population. PLoS One. 2013;8(10):e77024. | |
Ludwig JA, Weinstein JN. Biomarkers in cancer staging, prognosis and treatment selection. Nat Rev Cancer. 2005;5(11):845–856. | |
Wu X, Zhao H, Amos CI, et al. p53 Genotypes and Haplotypes Associated with Lung Cancer Susceptibility and Ethnicity. J Natl Cancer Inst. 2002;94(9):681–690. | |
Gurubhagavatula S, Liu G, Park S, et al. XPD and XRCC1 genetic polymorphisms are prognostic factors in advanced non-small-cell lung cancer patients treated with platinum chemotherapy. J Clin Oncol. 2004;22(13):2594–2601. | |
Sweeney C, Nazar-Stewart V, Stapleton PL, Eaton DL, Vaughan TL. Glutathione S-transferase M1, T1, and P1 polymorphisms and survival among lung cancer patients. Cancer Epidemiol Biomarkers Prev. 2003;12(6):527–533. | |
Takawa M, Cho HS, Hayami S, et al. Histone lysine methyltransferase SETD8 promotes carcinogenesis by deregulating PCNA expression. Cancer Res. 2012;72(13):3217–3227. | |
Xu J, Yin Z, Gao W, et al. Genetic variation in a microRNA-502 minding site in SET8 gene confers clinical outcome of non-small cell lung cancer in a Chinese population. PLoS One. 2013;8(10):e77024. | |
Ding C, Li R, Peng J, Li S, Guo Z. A polymorphism at the miR-502 binding site in the 3′ untranslated region of the SET8 gene is associated with the outcome of small-cell lung cancer. Exp Ther Med. 2012;3(4):689–692. | |
Xu P, Liu L, Wang J, et al. Genetic variation in BCL2 3′-UTR was associated with lung cancer risk and prognosis in male Chinese population. PLoS One. 2013;8(8):e72197. | |
Zhu W, Shan X, Wang T, Shu Y, Liu P. miR-181b modulates multidrug resistance by targeting BCL2 in human cancer cell lines. Int J Cancer. 2010;127(11):2520–2529. | |
Zhu W, Xu H, Zhu D, et al. miR-200bc/429 cluster modulates multidrug resistance of human cancer cell lines by targeting BCL2 and XIAP. Cancer Chemother Pharmacol. 2012;69(3):723–731. | |
Sacconi A, Biagioni F, Canu V, et al. miR-204 targets Bcl-2 expression and enhances responsiveness of gastric cancer. Cell Death Dis. 2012;3:e423. | |
Pakunlu RI, Wang Y, Tsao W, Pozharov V, Cook TJ, Minko T. Enhancement of the efficacy of chemotherapy for lung cancer by simultaneous suppression of multidrug resistance and antiapoptotic cellular defense: novel multicomponent delivery system. Cancer Res. 2004;64(17):6214–6224. | |
Garbuzenko OB, Saad M, Pozharov VP, Reuhl KR, Mainelis G, Minko T. Inhibition of lung tumor growth by complex pulmonary delivery of drugs with oligonucleotides as suppressors of cellular resistance. Proc Natl Acad Sci U S A. 2010;107(23):10737–10742. | |
Cheng M, Yang L, Yang R, et al. A microRNA-135a/b binding polymorphism in CD133 confers decreased risk and favorable prognosis of lung cancer in Chinese by reducing CD133 expression. Carcinogenesis. 2013;34(10):2292–2299. | |
Elsaba TM, Martinez-Pomares L, Robins AR, et al. The stem cell marker CD133 associates with enhanced colony formation and cell motility in colorectal cancer. PLoS One. 2010;5(5):e10714. | |
Vaughan AE, Halbert CL, Wootton SK, Miller AD. Lung cancer in mice induced by the jaagsiekte sheep retrovirus envelope protein is not maintained by rare cancer stem cells, but tumorigenicity does correlate with Wnt pathway activation. Mol Cancer Res. 2012;10(1):86–95. | |
Cortes-Dericks L, Galetta D, Spaggiari L, Schmid RA, Karoubi G. High expression of octamer-binding transcription factor 4A, prominin-1 and aldehyde dehydrogenase strongly indicates involvement in the initiation of lung adenocarcinoma resulting in shorter disease-free intervals. Eur J Cardiothorac Surg. 2012;41(6):e173–e181. | |
Yang ZL, Zheng Q, Yan J, Pan Y, Wang ZG. Upregulated CD133 expression in tumorigenesis of colon cancer cells. World J Gastroenterol. 2011;17(7):932–937. | |
Patnaik SK, Kannisto E, Knudsen S, Yendamuri S. Evaluation of microRNA expression profiles that may predict recurrence of localized stage I non-small cell lung cancer after surgical resection. Cancer Res. 2010;70(1):36–45. | |
Moll R, Franke WW, Schiller DL, Geiger B, Krepler R. The catalog of human cytokeratins: patterns of expression in normal epithelia, tumors and cultured cells. Cell. 1982;31(1):11–24. | |
Moll R, Divo M, Langbein L. The human keratins: biology and pathology. Histochem Cell Biol. 2008;129(6):705–733. | |
Warnecke-Eberz U, Baldus SE, Bollschweiler E, Hoelscher AH, Metzger R. Up-regulation of survivin mRNA might be a marker for non-invasive detection of non-small cell lung cancer rather than for prognosis. Anticancer Res. 2008;28(3A):1525–1529. | |
Zu Y, Ban J, Xia Z, et al. Genetic variation in a miR-335 binding site in BIRC5 alters susceptibility to lung cancer in Chinese Han populations. Biochem Biophys Res Commun. 2013;430(2):529–534. | |
Chin LJ, Ratner E, Leng S, et al. A SNP in a let-7 microRNA complementary site in the KRAS 3′ untranslated region increases non-small cell lung cancer risk. Cancer Res. 2008;68(20):8535–8540. | |
Nelson HH, Christensen BC, Plaza SL, Wiencke JK, Marsit CJ, Kelsey KT. KRAS mutation, KRAS-LCS6 polymorphism, and non-small cell lung cancer. Lung Cancer. 2010;69(1):51–53. | |
Yang L, Li Y, Cheng M, et al. A functional polymorphism at microRNA-629-binding site in the 3′-untranslated region of NBS1 gene confers an increased risk of lung cancer in Southern and Eastern Chinese population. Carcinogenesis. 2012;33(2):338–347. | |
Zhang S, Chen H, Zhao X, et al. REV3L 3′UTR 460 T>C polymorphism in microRNA target sites contributes to lung cancer susceptibility. Oncogene. 2013;32(2):242–250. | |
Knobel PA, Kotov IN, Felley-Bosco E, Stahel RA, Marti TM. Inhibition of REV3 expression induces persistent DNA damage and growth arrest in cancer cells. Neoplasia. 2011;13(10):961–970. | |
Xiong F, Wu C, Chang J, et al. Genetic variation in an miRNA-1827 binding site in MYCL1 alters susceptibility to small-cell lung cancer. Cancer Res. 2011;71(15):5175–5181. | |
Smits KM, Paranjape T, Nallur S, et al. A let-7 microRNA SNP in the KRAS 3′UTR is prognostic in early-stage colorectal cancer. Clin Cancer Res. 2011;17(24):7723–7731. | |
Klump B, Nehls O, Okech T, et al. Molecular lesions in colorectal cancer: impact on prognosis? Original data and review of the literature. Int J Colorectal Dis. 2004;19(1):23–42. | |
Wallner M, Herbst A, Behrens A, et al. Methylation of serum DNA is an independent prognostic marker in colorectal cancer. Clin Cancer Res. 15, 2006;12(24):7347–7352. | |
Lièvre A, Bachet JB, Boige V, et al. KRAS mutations as an independent prognostic factor in patients with advanced colorectal cancer treated with cetuximab. J Clin Oncol. 2008;26(3):374–379. | |
Sebio A, Paré L, Páez D, et al. The LCS6 polymorphism in the binding site of let-7 microRNA to the KRAS 3′-untranslated region: its role in the efficacy of anti-EGFR-based therapy in metastatic colorectal cancer patients. Pharmacogenet Genomics. 2013;23(3):142–147. | |
Kjersem JB, Ikdahl T, Guren T, et al. Let-7 miRNA-binding site polymorphism in the KRAS 3′UTR; colorectal cancer screening population prevalence and influence on clinical outcome in patients with metastatic colorectal cancer treated with 5-fluorouracil and oxaliplatin ± cetuximab. BMC Cancer. 2012;20(12):534–542. | |
Pardini B, Rosa F, Barone E, et al. Variation within 3′-UTRs of base excision repair genes and response to therapy in colorectal cancer patients: A potential modulation of microRNAs binding. Clin Cancer Res. 2013;19(21):6044–6056. | |
Wyatt MD, Wilson DM 3rd. Participation of DNA repair in the response to 5-fluorouracil. Cell Mol Life Sci. 2009;66(5):788–799. | |
Ingraham HA, Tseng BY, Goulian M. Mechanism for exclusion of 5-fluorouracil from DNA. Cancer Res. 1980;40(4):998–1001. | |
Wallace SS, Murphy DL, Sweasy JB. Base excision repair and cancer. Cancer Lett. 2012;327(1–2):73–89. | |
Kavli B, Sundheim O, Akbari M, et al. hUNG2 is the major repair enzyme for removal of uracil from U:A matches, U:G mismatches, and U in single-stranded DNA, with hSMUG1 as a broad specificity backup. J Biol Chem. 2002;277(42):39926–39936. | |
Terzic J, Grivennikov S, Karin E, Karin M. Inflammation and colon cancer. Gastroenterology. 2010;138(6):2101–2114. | |
Song S, Chen D, Lu J, et al. NFκB1 and NFκBIA polymorphisms are associated with increased risk for sporadic colorectal cancer in a southern Chinese population. PLoS One. 2011;6(6):e21726. | |
Landi D, Gemignani F, Pardini B, et al. Identification of candidate genes carrying polymorphisms associated with the risk of colorectal cancer by analyzing the colorectal mutome and microRNAome. Cancer. 2012;118(19):4670–4680. | |
Greber UF, Senior A, Gerace L. A major glycoprotein of the nuclear pore complex is a membrane-spanning polypeptide with a large lumenal domain and a small cytoplasmic tail. EMBO J. 1990;9(5):1495–1502. | |
Favreau C, Worman HJ, Wozniak RW, Frappier T, Courvalin JC. Cell cycle-dependent phosphorylation of nucleoporins and nuclear pore membrane protein Gp210. Biochemistry. 1996;35(24):8035–8044. | |
Rajkumar T, Sabitha K, Vijayalakshmi N, et al. Identification and validation of genes involved in cervical tumourigenesis. BMC Cancer. 2011;11:80. | |
Orabona C, Grohmann U, Belladonna ML, et al. CD28 induces immunostimulatory signals in dendritic cells via CD80 and CD86. Nat Immunol. 2004;5(11):1134–1142. | |
Taniguchi CM, Emanuelli B, Kahn CR. Critical nodes in signalling pathways: insights into insulin action. Nat Rev Mol Cell Biol. 2006;7(2):85–96. | |
Ryan BM, Robles AI, Harris CC. Genetic variation in microRNA networks: the implications for cancer research. Nat Rev Cancer. 2010;10(6):389–402. | |
Naccarati A, Pardini B, Stefano L, et al. Polymorphisms in miRNA-binding sites of nucleotide excision repair genes and colorectal cancer risk. Carcinogenesis. 2012;33(7):1346–1351. | |
Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61(2):69–90. | |
Chen YC, Hunter DJ. Molecular epidemiology of cancer. CA Cancer J Clin. 2005;55(1):45–54. | |
McPherson K, Steel CM, Dixon JM. ABC of breast diseases. Breast cancer-epidemiology, risk factors, and genetics. BMJ. 2000;321(7261):624–628. | |
Pharoah PD, Tyrer J, Dunning AM, Easton DF, Ponder BA; SEARCH Investigators. Association between common variation in 120 candidate genes and breast cancer risk. PLoS Genet. 2007;3(3):e42. | |
Zhang BL, Song FJ, Zheng H, Zhang LN, Zhao YR, Chen KX. [SNP rs16917496 within SET8 3′UTR is associated with the age of onset of breast cancer]. Zhonghua Zhong Liu Za Zhi. 2012;34(11):835–837. Chinese. | |
Forma E, Brys M, Krajewska W. TopBP1 in DNA damage response. In: Kruman I, editor. DNA Repair/Book 4. Rijeka, Croatia: INTECH Open Access Publisher; 2011;281–304. | |
Forma E, Brzeziańska E, Krześlak A, et al. Association between the c.*229C>T polymorphism of the topoisomerase IIβ binding protein 1 (TopBP1) gene and breast cancer. Mol Biol Rep. 2013;40(5):3493–3502. | |
Xu YJ, Leffak M. ATRIP from TopBP1 to ATR – in vitro activation of a DNA damage checkpoint. Proc Natl Acad Sci U S A. 2010;107(31):13561–13562. | |
Glover JN. Insights into the molecular basis of human hereditary breast cancer from studies of the BRCA1 BRCT domain. Fam Cancer. 2005;(1):89–93. | |
Xue L, Lipkin M, Newmark H, Wang J. Influence of dietary calcium and vitamin D on diet-induced epithelial cell hyperproliferation in mice. J Natl Cancer Inst. 1999;91(2):176–181. | |
Goodwin PJ, Ennis M, Pritchard KI, Koo J, Hood N. Prognostic effects of 25-hydroxyvitamin D levels in early breast cancer. J Clin Oncol. 2009;27(23):3757–3763. | |
Tse GM, Tan PH, Cheung HS, Chu WC, Lam WW. Intermediate to highly suspicious calcification in breast lesions: a radio-pathologic correlation. Breast Cancer Res Treat. 2008;110(1):1–7. | |
Zhang L, Liu Y, Song F, et al. Functional SNP in the microRNA-367 binding site in the 3′UTR of the calcium channel ryanodine receptor gene 3 (RYR3) affects breast cancer risk and calcification. Proc Natl Acad Sci U S A. 2011;108(33):13653–13658. | |
Brendle A, Lei H, Brandt A, et al. Polymorphisms in predicted microRNA-binding sites in integrin genes and breast cancer: ITGB4 as prognostic marker. Carcinogenesis. 2008;29(7):1394–1399. | |
Novaro V, Roskelley CD, Bissell MJ. Collagen-IV and laminin-1 regulate estrogen receptor alpha expression and function in mouse mammary epithelial cells. J Cell Sci. 2003;116(Pt 14):2975–2986. | |
Liu J, Tang X, Li M, et al. Functional MDM4 rs4245739 genetic variant, alone and in combination with P53 Arg72Pro polymorphism, contributes to breast cancer susceptibility. Breast Cancer Res Treat. 2013;140(1):151–157. | |
Jiang Y, Qin Z, Hu Z, et al. Genetic variation in a hsa-let-7 binding site in RAD52 is associated with breast cancer susceptibility. Carcinogenesis. 2013;34(3):689–693. | |
Lao JP, Oh SD, Shinohara M, Shinohara A, Hunter N. Rad52 promotes postinvasion steps of meiotic double-strand-break repair. Mol Cell. 2008;29(4):517–524. | |
Nicoloso MS, Sun H, Spizzo R, et al. Single-nucleotide polymorphisms inside microRNA target sites influence tumor susceptibility. Cancer Res. 2010;70(7):2789–2798. | |
Zheng H, Song F, Zhang L, et al. Genetic variants at the miR-124 binding site on the cytoskeleton-organizing IQGAP1 gene confer differential predisposition to breast cancer. Int J Oncol. 2011;38(4):1153–1161. | |
Lim LP, Lau NC, Garrett-Engele P, et al. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature. 2005;433(7027):769–773. | |
Noritake J, Watanabe T, Sato K, Wang S, Kaibuchi K. IQGAP1: a key regulator of adhesion and migration. J Cell Sci. 2005;118(Pt 10):2085–2092. | |
Hollestelle A, Pelletier C, Hooning M, et al. Prevalence of the variant allele rs61764370 T>G in the 3′UTR of KRAS among Dutch BRCA1, BRCA2 and non-BRCA1/BRCA2 breast cancer families. Breast Cancer Res Treat. 2011;128(1):79–84. | |
Tchatchou S, Jung A, Hemminki K, et al. A variant affecting a putative miRNA target site in estrogen receptor (ESR) 1 is associated with breast cancer risk in premenopausal women. Carcinogenesis. 2009;30(1):59–64. | |
Fishman J, Osborne MP, Telang NT. The role of estrogen in mammary carcinogenesis. Ann N Y Acad Sci. 1995;768:91–100. | |
Martin G, Davio C, Rivera E, et al. Hormone dependence of mammary tumors induced in rats by intraperitoneal NMU injection. Cancer Invest. 1997;15(1):8–17. | |
Chen WC, Lai YH, Chen HY, Guo HR, Su IJ, Chen HH. Interleukin-17-producing cell infiltration in the breast cancer tumour microenvironment is a poor prognostic factor. Histopathology. 2013;63(2):225–233. | |
Wang L, Liu W, Jiang W, et al. A miRNA binding site single-nucleotide polymorphism in the 3′-UTR region of the IL23R gene is associated with breast cancer. PLoS One. 2012;7(12):e49823. | |
Zwiers A, Kraal L, van de Pouw Kraan TC, Wurdinger T, Bouma G, Kraal G. Cutting edge: a variant of the IL-23R gene associated with inflammatory bowel disease induces loss of microRNA regulation and enhanced protein production. J Immunol. 2012;188(4):1573–1577. | |
Pharoah PD, Palmieri RT, Ramus SJ, et al; Ovarian Cancer Association Consortium. The role of KRAS rs61764370 in invasive epithelial ovarian cancer: implications for clinical testing. Clin Cancer Res. 2011;17(11):3742–3750. | |
Wynendaele J, Böhnke A, Leucci E, et al. An illegitimate microRNA target site within the 3′ UTR of MDM4 affects ovarian cancer progression and chemosensitivity. Cancer Res. 2010;70(23):9641–9649. | |
Liang D, Meyer L, Chang DW, et al. Genetic variants in MicroRNA biosynthesis pathways and binding sites modify ovarian cancer risk, survival, and treatment response. Cancer Res. 2010;70(23):9765–9776. | |
Ratner E, Lu L, Boeke M, et al. A KRAS-variant in ovarian cancer acts as a genetic marker of cancer risk. Cancer Res. 2010;70(16):6509–6515. | |
Gomaa AI, Khan SA, Toledano MB, Waked I, Taylor-Robinson SD. Hepatocellular carcinoma: epidemiology, risk factors and pathogenesis. World J Gastroenterol. 2008;14(27):4300–4308. | |
Caldwell S, Park SH. The epidemiology of hepatocellular cancer: from the perspectives of public health problem to tumor biology. J Gastroenterol. 2009;44 Suppl 19:96–101. | |
Guo Z, Wu C, Wang X, Wang C, Zhang R, Shan B. A polymorphism at the miR-502 binding site in the 3′-untranslated region of the histone methyltransferase SET8 is associated with hepatocellular carcinoma outcome. Int J Cancer. 2012;131(6):1318–1322. | |
Zhou C, Yu Q, Chen L, Wang J, Zheng S, Zhang J. A miR-1231 binding site polymorphism in the 3′UTR of IFNAR1 is associated with hepatocellular carcinoma susceptibility. Gene. 2012;507(1):95–98. | |
de Weerd NA, Samarajiwa SA, Hertzog PJ. Type I interferon receptors: biochemistry and biological functions. J Biol Chem. 2007;282(28):20053–20057. | |
Zhang Q, Gong R, Qu J, et al. Activation of the Ras/Raf/MEK pathway facilitates hepatitis C virus replication via attenuation of the interferon-JAK-STAT pathway. J Virol. 2012;86(3):1544–1554. | |
Song le H, Xuan NT, Toan NL, et al. Association of two variants of the interferon-alpha receptor-1 gene with the presentation of hepatitis B virus infection. Eur Cytokine Netw. 2008;19(4):204–210. | |
Gao Y, He Y, Ding J, et al. An insertion/deletion polymorphism at miRNA-122-binding site in the interleukin-1alpha 3′ untranslated region confers risk for hepatocellular carcinoma. Carcinogenesis. 2009;30(12):2064–2069. | |
Wang C, Zhao H, Zhao X, et al. Association between an insertion/deletion polymorphism within 3′UTR of SGSM3 and risk of hepatocellular carcinoma. Tumour Biol. 2014;35(1):295–301. | |
Yu Q, Zhou CX, Chen NS, Zheng SD, Shen LM, Zhang JK. A polymorphism within ErbB4 is associated with risk for hepatocellular carcinoma in Chinese population. World J Gastroenterol. 2012;18(4):383–387. | |
Miyamoto H, Yang Z, Chen YT, et al. Promotion of bladder cancer development and progression by androgen receptor signals. J Natl Cancer Inst. 4, 2007;99(7):558–568. | |
Teo MT, Landi D, Taylor CF, et al. The role of microRNA-binding site polymorphisms in DNA repair genes as risk factors for bladder cancer and breast cancer and their impact on radiotherapy outcomes. Carcinogenesis. 2012;33(3):581–586. | |
Andrew AS, Hu T, Gu J, et al. HSD3B and gene-gene interactions in a pathway-based analysis of genetic susceptibility to bladder cancer. PLoS One. 2012;7(12):e51301. | |
Beuten J, Gelfond JA, Franke JL, Leach RJ, et al. Single and multigenic analysis of the association between variants in 12 steroid hormone metabolism genes and risk of prostate cancer. Cancer Epidemiol Biomarkers Prev. 2009;18(6):1869–1880. | |
Giampaolo A, Sterpetti P, Bulgarini D, et al. Key functional role and lineage-specific expression of selected HOXB genes in purified hematopoietic progenitor differentiation. Blood. 1994;84(11):3637–3647. | |
Morgan R, Plowright L, Harrington KJ, Michael A, Pandha HS. Targeting HOX and PBX transcription factors in ovarian cancer. BMC Cancer. 2010;10:89. | |
Luo J, Cai Q, Wang W, et al. A microRNA-7 binding site polymorphism in HOXB5 leads to differential gene expression in bladder cancer. PLoS One. 2012;7(6):e40127. | |
Chu H, Wang M, Jin H, et al. EGFR 3′UTR 774T>C polymorphism contributes to bladder cancer risk. Mutagenesis. 2013;28(1):49–55. | |
Wu X, Gu J, Dong Q, et al. Joint effect of mutagen sensitivity and insulin-like growth factors in predicting the risk of developing secondary primary tumors and tumor recurrence in patients with head and neck cancer. Clin Cancer Res. 2006;12(23):7194–7201. | |
Khuri FR, Kim ES, Lee JJ, et al. The impact of smoking status, disease stage, and index tumor site on second primary tumor incidence and tumor recurrence in the head and neck retinoid chemoprevention trial. Cancer Epidemiol Biomarkers Prev. 2001;10(8):823–829. | |
Zhang X, Yang H, Lee JJ, et al. MicroRNA-related genetic variations as predictors for risk of second primary tumor and/or recurrence in patients with early-stage head and neck cancer. Carcinogenesis. 2010;31(12):2118–2123. | |
Christensen BC, Moyer BJ, Avissar M, et al. A let-7 microRNA-binding site polymorphism in the KRAS 3′ UTR is associated with reduced survival in oral cancers. Carcinogenesis. 2009;30(6):1003–1007. | |
Liu Z, Wei S, Ma H, et al. A functional variant at the miR-184 binding site in TNFAIP2 and risk of squamous cell carcinoma of the head and neck. Carcinogenesis. 2011;32(11):1668–1674. | |
Park DJ, Vuong PT, de Vos S, Douer D, Koeffler HP. Comparative analysis of genes regulated by PML/RAR alpha and PLZF/RAR alpha in response to retinoic acid using oligonucleotide arrays. Blood. 2003;102(10):3727–3736. | |
Sarma V, Wolf FW, Marks RM, Shows TB, Dixit VM. Cloning of a novel tumor necrosis factor-alpha-inducible primary response gene that is differentially expressed in development and capillary tube-like formation in vitro. J Immunol. 1992;148(10):3302–3312. | |
Xu Y, Ma H, Yu H, et al. The miR-184 binding-site rs8126 T>C polymorphism in TNFAIP2 is associated with risk of gastric cancer. PLoS One. 2013;8(5):e64973. | |
Li ZH, Pan XM, Han BW, et al. A let-7 binding site polymorphism rs712 in the KRAS 3′ UTR is associated with an increased risk of gastric cancer. Tumour Biol. 2013;34(5):3159–3163. | |
Lin Y, Nie Y, Zhao J, et al. Genetic polymorphism at miR-181a binding site contributes to gastric cancer susceptibility. Carcinogenesis. 2012;33(12):2377–2383. | |
Walker DM, Urbé S, Dove SK, Tenza D, Raposo G, Clague MJ. Characterization of MTMR3 an inositol lipid 3-phosphatase with novel substrate specificity. Curr Biol. 2001;11(20):1600–1605. | |
Stahl M, Mariette C, Haustermans K, Cervantes A, Arnold D; ESMO Guidelines Working Group. Oesophageal cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann Oncol. 2013;24(6):vi51–vi56. | |
Wang K, Li J, Guo H, et al. MiR-196a binding-site SNP regulates RAP1A expression contributing to esophageal squamous cell carcinoma risk and metastasis. Carcinogenesis. 2012;33(11):2147–2154. | |
Altschuler DL, Ribeiro-Neto F. Mitogenic and oncogenic properties of the small G protein Rap1b. Proc Natl Acad Sci U S A. 1998;95(13):7475–7479. | |
Zhou L, Zhang X, Li Z, et al. Association of a genetic variation in a miR-191 binding site in MDM4 with risk of esophageal squamous cell carcinoma. PLoS One. 2013;8(5):e64331. | |
American Cancer Society. Cancer Facts and Figures 2013. Atlanta, GA: American Cancer Society; 2013. Available from: http://www.cancer.org/acs/groups/content/@epidemiologysurveilance/documents/document/acspc-036845.pdf. Accessed July 4, 2014. | |
Gomella LG, Liu XS, Trabulsi EJ, et al. Screening for prostate cancer: the current evidence and guidelines controversy. Can J Urol. 2011;18(5):5875–5883. | |
Hughes L, Ruth K, Rebbeck TR, Giri VN. Genetic variation in IL-16 miRNA target site and time to prostate cancer diagnosis in African-American men. Prostate Cancer Prostatic Dis. 2013;16(4):308–314. | |
Liu J, Huang J, He Y, Liu J, Liao B, Liao G. Genetic variants in the integrin gene predicted microRNA-binding sites were associated with the risk of prostate cancer. Mol Carcinog. 2014;53(4):280–285. | |
Goel HL, Li J, Kogan S, Languino LR. Integrins in prostate cancer progression. Endocr Relat Cancer. 2008;15(3):657–664. | |
Feng N, Xu B, Tao J, et al. A miR-125b binding site polymorphism in bone morphogenetic protein membrane receptor type IB gene and prostate cancer risk in China. Mol Biol Rep. 2012;39(1):369–373. | |
Pham LK, Liang M, Adisetiyo HA, et al. Contextual effect of repression of bone morphogenetic protein activity in prostate cancer. Endocr Relat Cancer. 2013;20(6):861–874. | |
Surveillance, Epidemiology, and End Results Program (SEER) Stat Fact Sheets: Non-Hodgkin Lymphoma [webpage on the Internet]. Bethesda, MD: National Cancer Institute. Available from: http://seer.cancer.gov/statfacts/html/nhl.html. Accessed July 4, 2014. | |
Slager SL, Achenbach SJ, Asmann YW, et al. Mapping of the IRF8 gene identifies a 3′UTR variant associated with risk of chronic lymphocytic leukemia but not other common non-Hodgkin lymphoma subtypes. Cancer Epidemiol Biomarkers Prev. 2013;22(3):461–466. | |
Cheng CK, Kwan TK, Cheung CY, et al. A polymorphism in the 3′-untranslated region of the NPM1 gene causes illegitimate regulation by microRNA-337-5p and correlates with adverse outcome in acute myeloid leukemia. Haematologica. 2013;98(6):913–917. | |
Caiola E, Rulli E, Fruscio R, Buda A, Broggini M, Marabese M. KRas-LCS6 polymorphism does not impact on outcomes in ovarian cancer. Am J Cancer Res. 2012;2(3):298–308. | |
Tardat M, Brustel J, Kirsh O, et al. The histone H4 Lys 20 methyltransferase PR-Set7 regulates replication origins in mammalian cells. Nat Cell Biol. 2010;12(11):1086–1093. | |
Li Z, Nie F, Wang S, Li L. Histone H4 Lys 20 monomethylation by histone methylase SET8 mediates Wnt target gene activation. Proc Natl Acad Sci U S A. 2011;108(8):3116–3123. | |
Oda H, Okamoto I, Murphy N, et al, Reinberg D. Monomethylation of histone H4-lysine 20 is involved in chromosome structure and stability and is essential for mouse development. Mol Cell Biol. 2009;29(8):2278–2295. | |
Centore RC, Havens CG, Manning AL, et al. CRL4(Cdt2)-mediated destruction of the histone methyltransferase Set8 prevents premature chromatin compaction in S phase. Mol Cell. 2010;40(1):22–33. | |
Shi X, Kachirskaia I, Yamaguchi H, et al. Modulation of p53 function by SET8-mediated methylation at lysine 382. Mol Cell. 2007;27(4):636–646. | |
Markey MP. Regulation of MDM4. Front Biosci (Landmark Ed). 2011;16:1144–1156. | |
Li Q, Lozano G. Molecular pathways: targeting Mdm2 and Mdm4 in cancer therapy. Clin Cancer Res. 2013;19(1):34–41. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.