Back to Journals » International Journal of Nanomedicine » Volume 21
Micro/Nanoparticles in the Nose-Brain Axis: Implications for Pathogenesis and Therapeutic Interventions
Authors Mou YK ![]() , Li Q, Wang Y, Song XY, Wang HR
, Li Q, Wang Y, Song XY, Wang HR ![]() , Liu WC, Cheng YC, Shen H, Ren C
, Liu WC, Cheng YC, Shen H, Ren C ![]() , Song XC
, Song XC ![]()
Received 28 November 2025
Accepted for publication 16 March 2026
Published 3 April 2026 Volume 2026:21 585021
DOI https://doi.org/10.2147/IJN.S585021
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Sachin Mali
Ya-Kui Mou,1– 4,* Quan Li,2,5,* Yao Wang,1– 4,* Xiao-Yu Song,1– 4 Han-Rui Wang,1– 4 Wan-Chen Liu,1– 4 Yuan-Chao Cheng,1– 4 Hui Shen,1– 4 Chao Ren,2,6,7 Xi-Cheng Song1– 4
1Department of Otorhinolaryngology, Head and Neck Surgery, Yantai Yuhuangding Hospital, Qingdao University, Yantai, People’s Republic of China; 2Shandong Provincial Key Laboratory of Neuroimmune Interaction and Regulation, Yantai Yuhuangding Hospital, Yantai, People’s Republic of China; 3Shandong Provincial Clinical Research Center for Otorhinolaryngologic Diseases, Yantai Yuhuangding Hospital, Yantai, People’s Republic of China; 4Yantai Key Laboratory of Otorhinolaryngologic Diseases, Yantai Yuhuangding Hospital, Qingdao University, Yantai, People’s Republic of China; 5Department of Emergency, Yantai Yuhuangding Hospital, Qingdao University, Yantai, People’s Republic of China; 6Department of Neurology, Yantai Yuhuangding Hospital, Qingdao University, Yantai, People’s Republic of China; 7Yantai Municipal Key Medical and Health Laboratory of Yantai Yuhuangding Hospital (Interdisciplinary Brain Science and Geriatric Health Laboratory), Qingdao University, Yantai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Chao Ren, Department of Neurology. Yantai Yuhuangding Hospital, Qingdao University, No. 20, East Road, Zhifu District, Yantai, 264000, People’s Republic of China, Tel +86535 6691999, Fax +86535 6240341, Email [email protected] Xi-Cheng Song, Department of Otolaryngology, Head and Neck Surgery. Yantai Yuhuangding Hospital, Qingdao University, No. 20, East Road, Zhifu District, Yantai, 264000, People’s Republic of China, Tel +86535 6691999, Fax +86535 6240341, Email [email protected]
Abstract: The nose-brain axis (NBA) is a crucial bidirectional deliverypathway between the nasal cavity and the central nervous system (CNS) that influences both neurophysiology and disease progression. In addition to serving as a route for drug delivery, the NBA plays an active role in neurological disorders by mediating inflammatory responses, microbial interactions, and environmental exposure. Emerging evidence suggests that the NBA may be mechanistically relevant to CNS disorders, particularly within neurodegeneration frameworks such as the Braak and dual-hit hypotheses, which emphasize early olfactory or other peripheral involvements. This review explores how micro/nanoparticles interact with the NBA, not only as therapeutic carriers but also as factors contributing to neuroinflammation and neurodegeneration. Pathogenic micro/nanoparticles, including environmental pollutants and industrial nanoparticles, have been implicated in the exacerbation of CNS disorders by triggering oxidative stress and immune activation in both nasal and brain tissues. Conversely, therapeutic micro/nanoparticles such as biomimetic, synthetic, and cell-derived formulations represent promising strategies for modulating neuroinflammation, enhancing neuroprotection, and restoring CNS function through nasal-targeted interventions. However, substantial gaps remain in the existing understanding of the influences of nasal immune responses, microbiota, and barrier integrity on CNS health through the NBA. Addressing these challenges is critical for leveraging micro-/nanoparticles for the prevention and treatment of CNS diseases.
Keywords: nose-brain axis, micro/nanoparticles, neuroinflammation, neurodegenerative diseases
Introduction
The nose-brain axis (NBA) represents a unique and dynamic connection between the nasal cavity and the central nervous system (CNS), providing a pathway for external agents to access the brain and a platform for bidirectional interactions.1 Traditionally, research on the NBA has focused primarily on its ability to bypass the blood–brain barrier (BBB) during drug delivery. By exploiting the olfactory and trigeminal nerve pathways, this route facilitates direct transfer of therapeutic agents from the nasal cavity to the brain, offering a promising non-invasive alternative for treating CNS disorders.2 However, emerging evidence has highlighted the broader biological importance of the NBA as a conduit for organ–organ interactions and systemic health regulation. In addition to drug delivery, this axis mediates bidirectional signaling, linking nasal inflammation, environmental exposure, and infections to CNS pathologies such as neuroinflammation and neurodegeneration.3
Recent studies have highlighted the pathological consequences of nasal insults on the brain, emphasizing that infections, environmental toxins, and chronic inflammation in the nasal cavity can trigger or exacerbate CNS damage. For example, pathogens such as Streptococcus agalactiae and Burkholderia pseudomallei can invade the brain via the olfactory and trigeminal nerves, leading to meningitis and brainstem infections.4,5 Similarly, acute/chronic inflammatory conditions such as allergic rhinitis (AR) and rhinosinusitis may activate pro-inflammatory cytokines and immune mediators that traverse the nasal-CNS interface, potentially contributing to diseases such as depression, Alzheimer’s disease (AD) and Parkinson’s disease (PD).6,7 These findings underscore the importance of the NBA as more than just a passive conduit for drug delivery; it is an active player in CNS health and disease, and is capable of influencing neuroinflammation, neurodegeneration, and overall brain homeostasis.
In addition to its pathological relevance, the NBA has also become the focus of innovation in therapeutic interventions. Advances in nanotechnology have enabled the development of nanoparticles, nanoemulsions, and other formulations designed to optimize drug delivery via this route. These technologies offer distinct advantages, including enhanced drug stability, targeted brain delivery, and reduced systemic side effects.8 These systems have proven particularly effective in managing conditions such as epilepsy, AD, and schizophrenia.9 However, the full potential of this axis remains underexplored, particularly the ability to leverage its bidirectional properties to influence both the nasal cavity and brain. For example, therapeutic interventions targeting inflammatory pathways originating in the nasal cavity can alleviate local pathologies and mitigate the systemic effects on the CNS.
Despite these advancements, substantial gaps remain in the existing understanding of the NBA. The molecular mechanisms underlying the bidirectional delivery between the nasal cavity and CNS are poorly understood. For instance, the role of neurotransmitters such as glutamate in mediating neuroinflammatory responses originating in the nasal cavity warrants further investigation. Moreover, the long-term effects of environmental toxins, pathogens, and chronic nasal inflammation on brain health remain to be fully elucidated.3 These gaps in knowledge not only hinder the development of effective interventions, but also limit the ability to understand and address the bidirectional effects of nasal diseases and CNS function.
This review systematically elaborates on the physiological and anatomical features of the NBA as well as its associated key neural circuits and pathological mechanisms. Building on this foundation, we introduce the dual role concept of micro/nanoparticles in disease and further explore in detail the specific mechanisms through which these particles exert their actions via the NBA in various disorders. Through a comprehensive discussion of NBA functions, this article emphasizes that under both physiological and pathological conditions, the NBA not only serves as a potential route for treating CNS diseases but also acts as a critical hub for bidirectional delivery between the periphery and the CNS. This understanding contributes to the prevention of CNS diseases that may be induced by micro/nanoparticles. In terms of treatment, drug delivery strategies based on the NBA enable precise targeting while reducing systemic side effects, thereby significantly improving therapeutic outcomes. This review aims to deepen our understanding of the NBA and its potential in transforming the treatment of CNS diseases and managing nasal-related disorders with systemic implications. Regulating NBA-related pathways for treating these diseases is expected to become a highly promising clinical direction in the future.
Search Strategy
We conducted a literature search on PubMed using the following keywords in various combinations: nose-brain axis/pathway, neuroinflammation, micro-/nanoparticles, anatomy, physiology, brain, nose, olfactory nerve, trigeminal nerve, immune, microbial. All possible combinations of these keywords were used, and the abstracts of the retrieved records were evaluated. No time restrictions were applied. Additionally, manual searches of the reference lists from articles meeting the inclusion criteria were performed to identify earlier publications. Secondary literature was obtained from the reference lists of key papers, assessed for relevance, and included accordingly.
Anatomy and Physiology of the Nose and Brain
The intricate anatomical and physiological features of the nasal cavity and the brain form the basis for their dynamic bidirectional delivery, enabling therapeutic applications and pathological interactions. The specific anatomy of the NBA can be seen in Figure 1. The nasal cavity serves as a distinct gateway to the CNS via specialized pathways, including the olfactory and trigeminal nerves. These routes allow the direct transport of molecules, cells, and signals between the nasal cavity and the brain, bypassing the restrictive BBB. However, in addition to facilitating therapeutic drug delivery, this unique connectivity also permits the transmission of environmental toxins, pathogens, and inflammatory mediators from the periphery to the CNS. Conversely, CNS dysfunction can also influence nasal health through the neuroimmune and neuroinflammatory pathways, further confirming that the NBA is a critical focus of investigation. This bidirectional interplay underscores the importance of understanding the structural and functional basis of the NBA to advance scientific insights and clinical interventions.
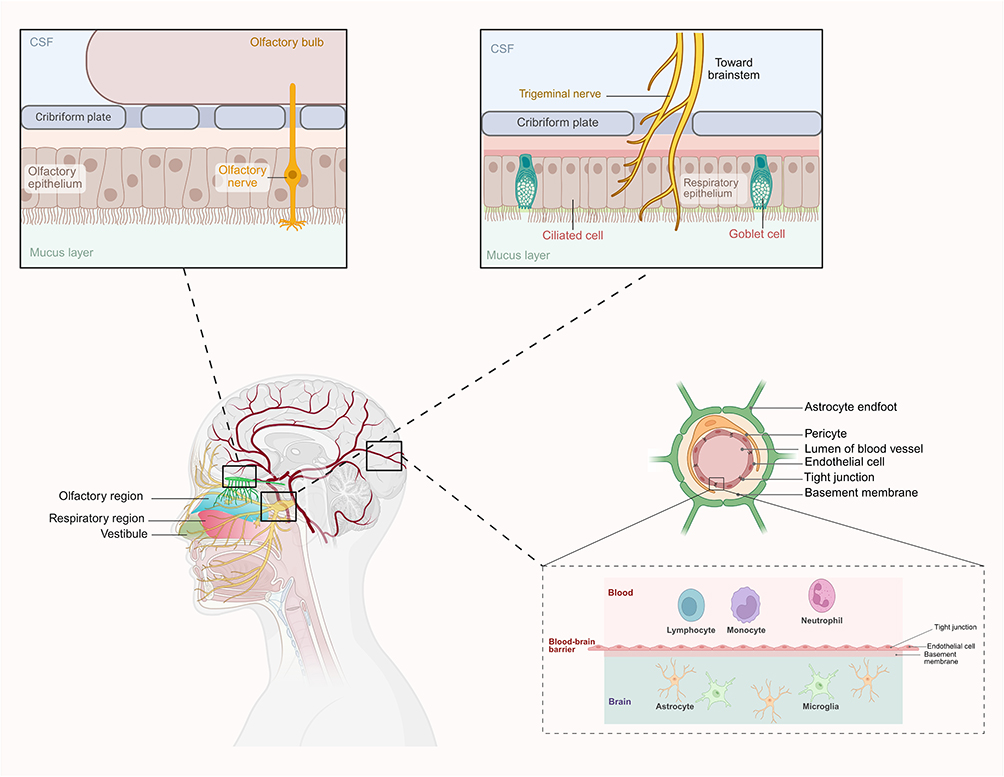 |
Figure 1 The anatomical schematic of the NBA. The nasal cavity functions as a unique portal to the CNS through specialized neural routes, notably the olfactory and trigeminal nerves. These pathways facilitate the direct transfer of molecules, cells, and neural signals from the nasal epithelium to the brain parenchyma, circumventing the selective permeability of the BBB. Abbreviations: NBA, nose-brain axis; CNS, central nervous system; BBB, blood-brain barrier. |
Anatomy and Physiology of the Nasal Cavity
The nasal cavity, which is anatomically divided into the vestibular, respiratory, and olfactory regions, serves as both a functional filter for inhaled air and a direct interface with the CNS. The olfactory region, located in the superior part of the nasal cavity, is lined with the olfactory epithelium containing olfactory sensory neurons (OSNs). These neurons project axons through the cribriform plate to the olfactory bulb, enabling direct neuronal communication with the brain.10 In contrast, the respiratory epithelium is composed of ciliated and goblet cells that produce mucus, which traps particulates and pathogens. Although protective, this mucociliary clearance system poses a barrier to drug delivery. To address this issue, nanoparticle-based delivery systems have been engineered to enhance mucoadhesion and enable deeper penetration into the olfactory epithelium.11 The trigeminal nerve (cranial nerve V) provides sensory innervation to the nasal cavity, and contributes to the perception of irritants and temperature changes. Importantly, its branches extend to the brainstem, offering an alternative route for direct drug transport and a mechanism for transmitting neuroinflammatory signals from the nasal cavity to the CNS.12
Environmental toxins, pathogens, and therapeutic agents exploit these neural pathways. For instance, the olfactory route has been implicated in the entry of pathogens such as Streptococcus pneumoniae, which can migrate along the olfactory nerve to cause meningitis.13,14 Similarly, intranasal delivery of drugs via nanoparticulate systems has shown promise in bypassing the BBB to treat CNS disorders such as AD.15,16
Anatomy and Physiology of the Brain
The brain is the most complex organ in the CNS, with tightly regulated barriers such as the BBB limiting the entry of substances from the systemic circulation. However, the olfactory bulb, located on the ventral surface of the brain, serves as a direct recipient of signals from the nasal cavity. This anatomical connection creates a unique vulnerability to external insults and provides a potential route for targeted drug delivery.17 In addition to processing odorant signals, the olfactory bulb also plays a role in neuroinflammation and neurodegeneration. Pathogens and toxins that migrate along the olfactory nerve can induce microglial activation in the bulb, contributing to the inflammatory cascades implicated in neurodegenerative diseases such as PD.18 In addition to the olfactory pathway, the trigeminal nerve provides an alternative route for interactions between the nasal cavity and brainstem. This nerve is involved in sensory processing and has been implicated in the propagation of neuroinflammatory signals from the periphery to central structures.18
The direct neural connections of the NBA bypass the metabolic and enzymatic barriers imposed by the BBB. This is exemplified by intranasal drug-delivery systems, wherein therapeutic agents delivered via the nasal route achieve rapid CNS concentration. Nanoparticles such as those made from polylactic-co-glycolic acid (PLGA) and chitosan have been designed to optimize delivery through these pathways by leveraging their capacity to cross neuronal and perineural interfaces.19 Understanding the anatomical and physiological interplay between the nasal cavity and brain is critical for advancing therapeutic strategies and mitigating the risks of pathological conditions that exploit this axis.
Mechanisms Linking the NBA
The NBA serves as a pivotal conduit for therapeutic delivery and pathological signaling, leveraging specialized anatomical pathways to facilitate bidirectional delivery between the nasal cavity and CNS. The NBA employs the olfactory and trigeminal nerve pathways as direct ascending conduits. Upon stimulation by allergens, pathogens, or environmental toxins, the nasal mucosa releases pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6). These cytokines disrupt the tight junctions of the nasal epithelium, thereby increasing its permeability. This breakdown allows immune cells and inflammatory mediators, which are normally restricted by the barrier, to accumulate around nerve terminals. Subsequently, these harmful substances and signals can travel directly into the central nervous system along the olfactory and trigeminal nerves via mechanisms such as axonal transport or intercellular transmission, inducing neuronal inflammatory responses and oxidative stress. Concurrently, nasal dysbiosis, characterized by microbial imbalance, exacerbates this inflammatory cascade by disrupting local mucosal immune homeostasis and providing sustained pro-inflammatory stimuli. Furthermore, certain components or metabolites from the dysregulated microbiota may themselves be directly transported upwards along these neural pathways.
Recent evidence has highlighted both extracellular and intracellular mechanisms underlying this transport. Extracellular pathways involve the diffusion of molecules along the olfactory epithelium and mucosal surfaces, enabling their rapid entry into the CNS. Conversely, intracellular transport occurs via axonal migration within sensory neurons, allowing pathogens, nanoparticles, and signaling molecules to bypass the BBB. In addition to these structural mechanisms, emerging evidence has identified the critical roles of neuroimmune signaling and inflammatory mediators in amplifying the effects of nasal insults on brain health. This complex interplay between cellular and molecular mechanisms underscores the importance of the NBA as a central focus for understanding CNS pathology and optimizing therapeutic interventions.
Olfactory Nerve: Direct Transmission Pathway
The olfactory nerve serves as a critical and bidirectional conduit between the nasal cavity and CNS, facilitating the transport of substances and communication of pathological signals. Transport of substances occurs primarily via intracellular and extracellular pathways.20 Intracellular transport involves the internalization of molecules such as nanoparticles and small molecules by OSNs through endocytosis. These substances are then transported along the axonal microtubule network to the olfactory bulb with the assistance of motor proteins such as dynein and kinesin.21 Conversely, extracellular transport bypasses cellular internalization, allowing larger molecules and hydrophilic substances to diffuse through the perineural space or traverse the paracellular junctions between olfactory epithelial cells. These pathways enable both therapeutic agents and potentially harmful molecules, such as environmental toxins, to bypass the BBB and directly access the CNS.22
Importantly, the olfactory nerve is not merely a one-way conduit; evidence indicates a bidirectional interaction between nasal pathology and CNS health. In neurodegenerative diseases such as PD, the olfactory system is often among the earliest affected regions, with olfactory dysfunction preceding motor symptoms. These findings support the hypothesis that pathological proteins such as alpha-synuclein aggregates may propagate from the nasal epithelium to the CNS via the olfactory nerve.1 Conversely, nasal health can be influenced by CNS dysfunction. For example, patients with AR, a chronic nasal inflammatory condition, are at increased risk of depression and anxiety, which is likely mediated by neuroimmune and neuroinflammatory pathways that influence brain health.23
From a therapeutic perspective, the olfactory nerve has been exploited for direct delivery of neuroprotective agents to the CNS. In neurodegenerative diseases, polymeric nanoparticles such as PLGA, which are functionalized with mucoadhesive polymers such as chitosan, have shown promise in delivering drugs to the olfactory bulb. For instance, nanoemulsions have been used to rapidly deliver both hydrophilic and hydrophobic drugs to the affected CNS regions in patients with AD and PD.24 In neuroinflammatory and neuroimmune disorders, nanoparticles loaded with anti-inflammatory agents have been used to reduce CNS inflammation in preclinical models of multiple sclerosis, providing a non-invasive alternative to systemic therapies. Furthermore, diagnostic imaging approaches, such as manganese-enhanced magnetic resonance imaging (MRI), have been used to confirm the rapid transport and distribution of substances delivered intranasally via the olfactory nerve, underscoring the potential of these approaches for monitoring disease progression and evaluating therapeutic outcomes.25
The bidirectional role of the olfactory nerve in CNS health and nasal diseases highlights its unique position as both a therapeutic gateway and pathological conduit. Understanding how this pathway mediates reciprocal interactions, such as the influence of neurodegeneration on olfactory function and that of nasal inflammation on CNS disorders, can offer valuable insights into the complex interplay between the peripheral and central systems. Continued exploration of these mechanisms will improve the ability to diagnose, monitor, and treat disorders that bridge the nasal cavity and CNS.
Trigeminal Nerve: Indirect Transmission
The trigeminal nerve (cranial nerve V) is a critical component of the NBA, which connects the nasal cavity to diverse regions of the CNS. Unlike the olfactory nerve, which provides rapid and direct access to the brain, the trigeminal nerve facilitates sustained and targeted transport to deeper CNS structures, including the brainstem, midbrain, and spinal cord. By bypassing the BBB, this pathway offers substantial advantages for drug delivery and therapeutic interventions, particularly in conditions that require prolonged regional targeting of the CNS.26 Intranasal delivery of substances employs both intracellular and extracellular transport mechanisms along the trigeminal nerve. However, extracellular diffusion through the perineural spaces is typically dominant, albeit slower than that through the olfactory nerve pathways. This mechanism has been validated extensively in preclinical studies. For instance, curcumin-loaded polycaprolactone (PCL) nanoparticles effectively permeate the trigeminal nerve mucosa and accumulate in the brainstem regions within hours of nasal administration.27 Similarly, PLGA nanoparticles have demonstrated efficient transport and bioavailability of neuroprotective agents targeting the brainstem, further validating the therapeutic potential of this route.28
Nanoparticle optimization plays a pivotal role in the development of this drug-delivery pathway. For example, PEGylated PLGA nanoparticles enhance nasal cavity retention and allow sustained drug release, although excessive PEGylation can reduce trigeminal nerve permeability. To address this, functionalized nanoparticles modified with targeting ligands such as rabies virus glycoprotein (RVG) have been developed to improve payload delivery specifically to brainstem regions. This targeting strategy is particularly promising for conditions such as migraines and neurodegenerative diseases.22,29 Other emerging clinical applications have also highlighted the therapeutic potential of the trigeminal pathway. In comparison with intravenous administration, intranasal delivery of eletriptan hydrobromide nanoparticles has been shown to increase the activation of trigeminal neurons, resulting in improved CNS drug levels and substantial therapeutic efficacy for migraine treatment.30 Similarly, solid lipid nanoparticles and lipid-based carriers have shown high bioavailability for neuroprotective agents in preclinical models of PD and AD, further validating the importance of the trigeminal route for long-term therapeutic strategies.28,31
Overall, the trigeminal nerve pathway complements the olfactory route by providing sustained and targeted delivery of larger molecules to critical CNS regions. This dual contribution enhances the potential of NBA as a platform for developing advanced therapeutic interventions, offering a robust solution for conditions requiring precision, prolonged effectiveness, and regional CNS targeting.32,33
Immune and Inflammatory Pathways
The immune and inflammatory pathways of the NBA play central roles in mediating the effects of nasal inflammation on CNS health, while also reflecting the bidirectional relationship between peripheral and central immune responses. Nasal inflammation, driven by allergens, pathogens, or environmental toxins, triggers the release of pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-1β, and IL-6, which disrupt the nasal epithelial barrier and increase its permeability.34,35 This allows immune cells and inflammatory mediators to access the CNS through pathways such as the olfactory and trigeminal nerves. For instance, chronic rhinosinusitis (CRS) is characterized by type 2 inflammation involving cytokines such as IL-4, IL-13, and IL-5, which can impair OSNs but also compromise mucosal integrity, creating an entry point for inflammation to propagate to the CNS.36,37 Conversely, neuroinflammatory conditions such as AD and PD can lead to anosmia and further weaken mucosal defenses, demonstrating the bidirectional impact of nasal and CNS pathologies.38,39
Microglial activation is a hallmark of the CNS inflammation associated with the NBA. Chronic nasal inflammation primes microglia, making them hyperresponsive to subsequent immune challenges. Activated microglia amplify neuroinflammatory responses by releasing pro-inflammatory cytokines, reactive oxygen species (ROS), and neurotoxic mediators, thereby perpetuating a cycle of neuronal damage.40,41 This mechanism is particularly relevant in neurodegenerative diseases such as AD and PD, where sustained microglial activation contributes to the progression of the disease. Additionally, inflammatory mediators, such as matrix metalloproteinases (MMPs), degrade tight junction proteins in both the nasal epithelium and BBB, facilitating the entry of peripheral immune signals into the CNS. For example, type 1 inflammation mediated by interferon-gamma (IFN-γ) induces epithelial-to-mesenchymal transition in the nasal epithelium, exacerbating tissue remodeling and barrier dysfunction.42,43 The bidirectional nature of inflammation complicates the interplay between the periphery and the CNS. Peripheral inflammation, such as CRS or AR, can activate central pathways such as the hypothalamic-pituitary-adrenal (HPA) axis, driving systemic immune responses and neuroinflammatory cascades.44,45 Conversely, CNS inflammation resulting from neurodegenerative conditions can exacerbate nasal inflammation through neuroimmune feedback loops, impairing mucosal barrier function and OSNs.46,47 This cyclical interaction underscores the importance of the NBA in linking nasal health to systemic and CNS pathologies.
While the physiological roles of immune responses and barrier integrity within the NBA have been well emphasized, the specific triggering mechanisms for inflammatory factor release, the precise neural circuits involved in signal transmission, and the molecular pathways and corresponding receptors through which these effects are targeted remain insufficiently explored. Furthermore, the cascade of responses within the CNS triggered by the disruption of nasal mucosal and BBB integrity—such as the activation of specific brain regions and alterations in electrophysiological signals—has yet to be fully elucidated. Further investigation utilizing techniques such as electroencephalographic (EEG) monitoring and optogenetic-based neural tracing is warranted. Validation in animal models would provide more compelling evidence in this regard. Targeting the immune and inflammatory pathways within this axis represents a promising therapeutic avenue for mitigating neuroinflammatory conditions and restoring barrier integrity in both the nasal cavity and CNS.
Microbial Pathways: Interactions Between the Nasal Microbiota and CNS Health
The nasal microbiota, a complex ecosystem of commensal and potentially pathogenic microorganisms, plays a pivotal role in mediating the relationship between the nasal cavity and the CNS through the NBA. Emerging evidence indicates that microbial dysbiosis in the nasal cavity, characterized by imbalances in microbial communities, can disrupt mucosal immunity, trigger inflammatory cascades, and directly or indirectly propagate neuroinflammation. These processes underscore the fact that the nasal microbiota is a crucial mediator in the mechanisms linking the nasal environment to CNS health and pathology.
Microbial dysbiosis facilitates the transfer of inflammatory and immune signals from the nasal cavity to the CNS via multiple mechanisms. Pathogens such as Staphylococcus aureus and S. pneumoniae have been shown to invade the nasal epithelium, releasing toxins and pro-inflammatory molecules that disrupt epithelial integrity and allow immune signaling molecules to access the CNS.48,49 These inflammatory mediators travel through neural routes such as the olfactory and trigeminal nerves, creating direct communication pathways between the nasal cavity and other brain regions. For instance, bacterial endotoxins such as lipopolysaccharides (LPS) can activate Toll-like receptors (TLRs) on neural and epithelial cells, amplifying inflammatory responses that extend to the CNS.17,50 Experimental studies have demonstrated that such neuroinflammatory cascades contribute to neurodegenerative diseases such as AD and PD, further highlighting the role of microbial-mediated pathways in linking nasal health to CNS dysfunction.51 Additionally, recent findings have suggested that nasal Mycoplasma pneumoniae infections can increase systemic and neuroinflammatory responses, contributing to mood disorders such as depression.52 By disrupting immune homeostasis and stimulating the release of pro-inflammatory cytokines, M. pneumoniae may indirectly influence CNS function and exacerbate depressive symptoms, linking nasal infections to neuropsychiatric outcomes.53
In addition to immune modulation, microbial metabolites play an important role in connecting the nasal microbiota to the CNS. Beneficial metabolites such as short-chain fatty acids (SCFAs), which are produced by commensal bacteria, can promote anti-inflammatory effects and enhance BBB integrity, facilitating neuroprotection. Conversely, in states of dysbiosis, the production of neurotoxic byproducts such as hydrogen sulfide and nitric oxide increases, exacerbating oxidative stress and mitochondrial dysfunction in neurons.54,55 Furthermore, certain pathogens, including Chlamydia pneumoniae and Haemophilus influenzae, utilize neural pathways for direct CNS invasion, bypassing the BBB and inducing localized neuroinflammation.56–58 These direct microbial invasions establish clear anatomical and functional connections between the nasal cavity and brain, positioning the microbiota as a key regulator of nose-brain interactions. Systemic interactions between the nasal microbiota and other microbial ecosystems, such as the gut microbiota, further emphasize the holistic nature of NBA. Dysbiosis in the nasal cavity can propagate inflammatory signals through the systemic circulation, amplifying neuroinflammatory responses via shared immune pathways. For example, microbial imbalances in both the gut and nasal cavities have been implicated in the exacerbation of PD, illustrating the interconnectedness of these ecosystems and their collective impact on CNS health.59–61 Together, these findings establish that the nasal microbiota is a critical component of the mechanisms linking the nasal cavity to CNS health through immune, metabolic, and direct microbial pathways.
Insights from the Braak/Dual-Hit Hypothesis
The Braak/dual-hit hypothesis offers a coherent neurodegenerative framework for understanding how peripheral nasal exposures—operating through the nose–brain axis—may initiate or accelerate central proteinopathy.62 It was originally developed to explain how α-synuclein pathology spreads, both in space and over time, in diseases like Parkinson’s. It proposes that an environmental agent, such as a pathogen or toxin, enters the brain through one of two distinct portals: the olfactory epithelium or the gut. Once inside, it moves along neural circuits, propagating from cell to cell in a prion-like fashion.63 Within the NBA, this model reframes the olfactory nerve not merely as a conduit for sensory transduction, but as a vulnerable entry route through which exogenous agents—including ambient micro/nanoparticles, viral pathogens, or inflammatory mediators—can gain access to the central nervous system and trigger pathological protein aggregation.
Experimental and post-mortem evidence lends credence to this naso-central axis of pathogenesis.64 Following intranasal instillation, α-synuclein fibrils have been shown to undergo anterograde and retrograde transport along olfactory sensory neurons, reaching the olfactory bulb within hours and progressively involving limbic structures, the amygdala, and brainstem nuclei over weeks to months. This trajectory closely parallels Braak’s proposed staging scheme for sporadic PD, in which olfactory bulb pathology precedes nigrostriatal involvement. Clinically, the near-ubiquitous presence of olfactory dysfunction in early PD—often manifesting years before motor symptoms—further underscores the nose as a plausible point of entry.65 More recent work using traceable nanoparticles has demonstrated that size, surface charge, and persistence in the nasal mucosa govern not only the efficiency of axonal translocation, but also the subsequent microglial response and induction of α-synuclein misfolding in second-order neurons.
Crucially, the relationship between nasal insult and central neurodegeneration is not unidirectional. The dual-hit hypothesis also accommodates a feedback loop in which established CNS pathology exacerbates peripheral vulnerability. In PD and AD, central α-synuclein or tau deposition has been shown to modulate autonomic and sensory efferents that innervate the nasal mucosa, impairing epithelial barrier integrity and reducing odorant receptor expression.66 This reciprocal crosstalk creates a self-perpetuating cycle: nasal inflammation facilitates CNS entry of pathogenic particles, while ongoing neurodegeneration weakens nasal defenses, rendering the host increasingly susceptible to subsequent environmental hits.
Beyond the olfactory route, the dual-hit framework draws attention to the possibility of synergistic peripheral portals. Micro/nanoparticles deposited in the nasopharynx may be cleared into the gastrointestinal tract, where they can engage the gut–brain axis in parallel. Emerging evidence suggests that nasal and intestinal exposures may act in concert, each capable of seeding pathology and accelerating progression through distinct but converging neural circuits.67 This has implications for experimental design: studies focusing solely on one portal may underestimate the cumulative burden of peripheral proteinopathy induction.
Despite its heuristic value, the Braak/dual-hit hypothesis remains incompletely validated with respect to the NBA, particularly in the context of anthropogenic particulate exposure. Most supporting data derive from rodent models using supraphysiological doses of preformed fibrils or labelled particles; whether chronic, low-dose exposure to real-world environmental particulates is sufficient to initiate authentic α-synuclein pathology in humans remains unresolved. Moreover, the precise biophysical determinants governing axonal uptake, retrograde transport kinetics, and trans-synaptic transfer are only beginning to be delineated.
Micro/Nanoparticles in NBA
According to our literature review, micro/nanoparticles can play a dual role of being pathogenic and therapeutic and the mechanisms can be seen in Figure 2.
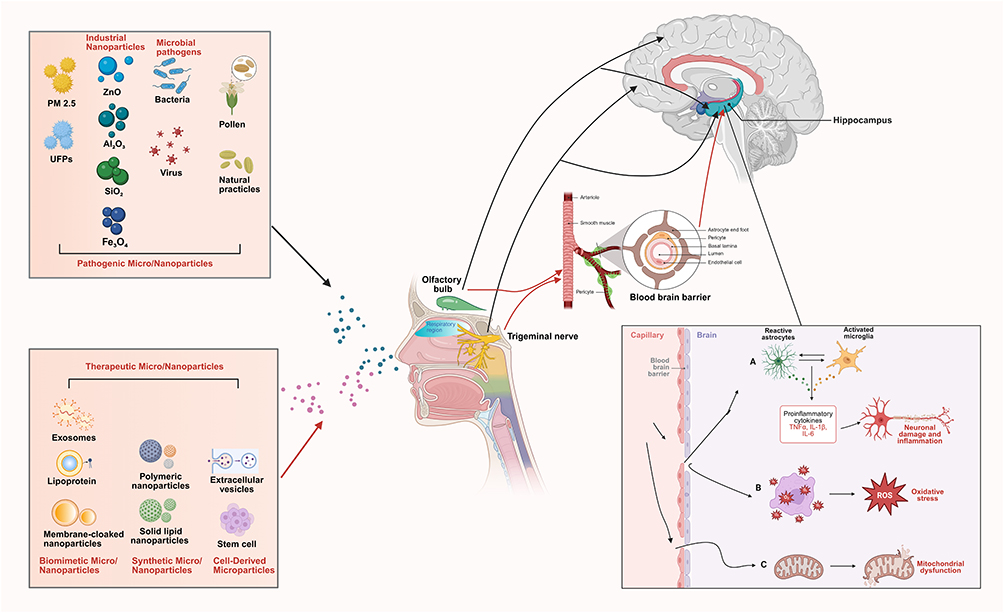 |
Figure 2 Mechanisms related to the pathogenic and therapeutic effects of micro/nanoparticles through NBA. Once the pathogenic and therapeutic micro/nanoparticles enter the nasal cavity, their smaller size allows penetration into the CNS through the nasal mucosa, mainly through the olfactory or trigeminal nerves. Within CNS, (A) these particles activate microglia and astrocytes, triggering the release of pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α, resulting in chronic inflammation, neuronal damage, and the disruption of normal synaptic function. In addition, they can exacerbate neurotoxicity by (B) inducing oxidative stress and (C) disrupting mitochondrial function. (outside the box, the black arrows represent the pathway of pathogenic micro/nanoparticles, while the red arrows represent the pathway of therapeutic micro/nano-particles). Abbreviations: NBA, nose-brain axis; CNS, central nervous system; IL-1β, Interleukin-1β; IL-6, Interleukin-6; TNF-α, tumor necrosis factor-α; UFPs, ultrafine particles; ROS, reactive oxygen species. |
As Pathogenic Agents: harmful micro/nanoparticles from environmental, industrial, microbial, and natural sources can exploit the NBA. Due to their small size and properties, they bypass protective barriers like the nasal epithelium and BBB, entering the CNS. Once inside, they trigger neurotoxicity, neuroinflammation, and oxidative stress, contributing to neurological damage and posing significant public health risks.
As Therapeutic Carriers: in contrast, engineered therapeutic micro/nanoparticles can utilize the same NBA pathway for beneficial purposes. Designed for precise delivery, they transport drugs or genetic material (eg, siRNAs) directly to the brain. This approach effectively treats neuroinflammatory and neurodegenerative conditions while minimizing systemic side effects, turning the NBA into a targeted route for intervention.
The NBA serves as a critical double-edged pathway: it can be infiltrated by pathogenic particles causing neurological disorders, yet it also offers a promising delivery route for precisely engineered therapeutic nanoparticles designed to treat those very disorders. This duality underscores the importance of understanding particle-biomaterial interactions for both toxicology and advanced drug development.
Pathogenic Micro/Nanoparticles
Environmental Pollutants: PM2.5, Ultrafine Particles and Micro-Nanoplastics
Particulate matter (PM), particularly PM2.5 (particulate matter with diameter ≤ 2.5 µm) and ultrafine particles (UFPs; particles with diameter ≤ 100 nm), is one of the most studied contributors to NBA dysfunction. PM2.5 is derived from combustion processes, vehicular emissions, and industrial activities. Its small size enables penetration through the nasal mucosa into the CNS, predominantly via the olfactory and trigeminal nerves.68 Within the CNS, PM2.5 activates microglia and astrocytes, triggering the release of pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α.69,70 This cascade results in chronic inflammation, neuronal damage, and the disruption of normal synaptic functions. In addition, PM2.5-associated heavy metals such as manganese exacerbate neurotoxicity by inducing oxidative stress and disrupting mitochondrial function. These metals promote amyloid-beta deposition and tau hyperphosphorylation, which are hallmarks of AD.71,72 Animal studies have demonstrated a direct relationship between long-term exposure to PM2.5 and cognitive decline, with affected animals exhibiting impairments in spatial memory and learning tasks. UFPs represent an even greater threat because of their nanoscale size and increased surface area, which increase their reactivity and ability to cross biological barriers. Studies have shown that UFPs accumulate in the olfactory bulb and hippocampus, where they induce substantial neuroinflammation and synaptic dysfunction.73,74 Unlike larger PM particles, UFPs directly interact with neuronal structures, thereby amplifying oxidative stress and inflammatory responses.
Micro- and nanoplastics (MNPs, <5 mm and <100 nm) are environmental contaminants found in human tissues including the brain.75 Brain MNP levels are ~12-fold higher than in liver and kidneys (up to 30-fold), with potential effects on neurodevelopment and cognition.76 From 2016 to 2024, brain MNP concentrations rose by ~50%. Compositionally, ~75% are polyethylene (PE), with polypropylene (PP) and others making up the rest.76 Morphologically, brain MNPs are predominantly nanoscale fragments (<200 nm long, <40 nm wide).
Inhaled or systemically circulating MNPs may enter the central nervous system via nasal deposition and olfactory pathways, where they interact with vascular endothelial cells and immune cells.77 Mechanistically, MNPs are proposed to induce oxidative stress, thereby activating the neuroimmune microenvironment.77 This is primarily characterized by the activation of microglia and astrocytes, accompanied by increased release of pro‑inflammatory cytokines such as IL-1β, IL-6, and TNF-α. Such neuroinflammatory responses may synergistically contribute to microvascular obstruction, impaired local cerebral perfusion, synaptic plasticity deficits, and disruptions in neuronal signaling, ultimately exerting detrimental effects on neural network function.78 Elevated brain MNP burdens have been observed in individuals with neurocognitive disorders, including dementia.
The aforementioned evidence not only elucidates the specific mechanisms by which environmental pollutants contribute to the pathogenesis of CNS diseases at the molecular experimental level, but has also been further validated in transgenic animal models. In addition, multi-omics technologies have been employed to identify potential genes involved, supported by large-scale clinical studies. However, human-based experimental evidence has primarily focused on the health effects of PM2.5 derived from traffic and fuel combustion, while PM2.5, micro-nanoplastics from other sources has not been sufficiently examined. Moreover, other air pollutants such as nitrogen oxides and ozone may also act synergistically in disease progression, potentially influencing or confounding the experimental outcomes. Although the present study suggests that PM2.5 and micro-nanoplastics can influence the development of neurodegenerative diseases through neuroinflammatory pathways mediated by the NBA, it did not directly assess the specific pathways involving pro‑inflammatory factors or related genes. Therefore, these findings may not fully capture the overall impact of PM2.5 and micro-nanoplastics on neuropathological health outcomes.
Industrial Nanoparticles
Engineered nanoparticles, which are widely used in consumer products, medical devices, and industrial processes, pose emerging risks to the NBA and CNS. These nanoparticles can penetrate the CNS via the olfactory and trigeminal pathways, bypassing the BBB and subsequently inducing oxidative stress, neuroinflammation, and cellular toxicity. Zinc oxide (ZnO), aluminum oxide (Al2O3), and silica (SiO2) nanoparticles are among the most studied for their neurotoxic effects. For example, ZnO nanoparticles have been shown to accumulate in the olfactory bulb and hippocampus after intranasal exposure, where they trigger mitochondrial dysfunction, generate ROS, and impair dopaminergic neurons. These effects have been linked to motor deficits and an increased risk of PD, highlighting the vulnerability of the NBA to industrial nanoparticles.79,80 Similarly, Al2O3 nanoparticles disrupt hippocampal signaling pathways, induce neuroinflammation, and promote neuronal apoptosis.81 These processes result in synaptic loss, impaired memory formation, and cognitive deficits, underscoring the role of Al2O3 in exacerbating neurodegenerative conditions such as AD. Another nanoparticle of particular concern is airborne magnetite (Fe3O4), a byproduct of urban pollution and industrial activities. Magnetite nanoparticles have been identified in human brain tissue, where they act as catalytic agents for ROS production and exacerbate amyloid-beta aggregation, a hallmark of AD. Their strong association with neurodegenerative pathology emphasizes the systemic implications of chronic nanoparticle exposure.81,82 A particularly concerning nanoparticle is airborne magnetite (Fe3O4), which is associated with urban pollution. Magnetite nanoparticles have been detected in human brain tissue, where they catalyze ROS production and promote amyloid-beta aggregation. Their presence in AD plaques underscores their role in accelerating neurodegenerative processes.83,84 In addition, titanium dioxide (TiO2) nanoparticles, which are commonly used in cosmetics and food products, can enter the CNS following intranasal exposure, where they induce neurobehavioral abnormalities such as anxiety-like behaviors and learning impairments. Studies have highlighted the role of TiO2 nanoparticles in disrupting BBB integrity, increasing neuroinflammation, and altering neurotransmitter levels, particularly in regions critical for cognitive function.85 Furthermore, industrial nanoparticles such as SiO2 and carbon-based particles exacerbate CNS injury by amplifying glial activation and neuroinflammatory responses, which can propagate neuronal damage over time.86
The aforementioned evidence, through integrated analysis combining molecular, cellular, and animal experiments with multi-omics technologies, have comprehensively examined the impact of industrial nanoparticles on CNS diseases and its underlying mechanisms. These findings highlight the bidirectional vulnerability of the NBA to environmental pollutants and industrial nanoparticles, which not only disrupt neuronal homeostasis, but also exacerbate neurodegenerative processes. Given the widespread presence of these nanoparticles in the environment and consumer goods, stringent monitoring, regulation, and further research are essential to mitigate nanoparticle-induced CNS toxicity.87,88 However, the lack of large-scale epidemiological evidence or direct experimental studies conducted in human somewhat diminishes the persuasiveness of these results, warranting further investigation in future research.
Microbial Agents and Pathogens
Microbial pathogens, including bacteria and viruses, exploit the NBA as a critical route for CNS invasion, bypassing the BBB via neural pathways such as the olfactory and trigeminal nerves.89 Bacterial pathogens, such as S. pneumoniae and H. influenzae, adhere to nasal epithelial cells, disrupt mucosal barrier integrity, and gain enhanced access to the CNS through the perineural spaces. These infections often result in severe neurological complications, including meningitis and encephalitis.53,90,91 S. pneumoniae produces pneumolysin, a toxin that induces epithelial cell death, exacerbates neuroinflammation, and triggers neuronal apoptosis, ultimately accelerating brain damage and cognitive deficits.92 Additionally, emerging evidence suggests that nasal infections caused by M. pneumoniae can increase systemic inflammatory responses and contribute to mood disorders, such as depression. And nasal staphylococcus aureus can produce a sex hormone-degrading enzyme that breaks down testosterone and estradiol, thereby reducing the concentrations of dopamine and serotonin in the brains of mouse models and inducing depressive-like behaviors,92 demonstrating the bidirectional impact of microbial dysbiosis on the brain and peripheral immune signaling.
The gut microbiota has been confirmed in numerous significant studies to be associated with the pathogenesis of CNS diseases such as cerebral ischemia and AD, and it can promote the biotransformation of certain drugs.93–95 The gut-brain axis theory provides a new research perspective for revealing the synergistic pathological changes between the brain and gut. According to this theory, there exists a bidirectional regulatory mechanism between the CNS and the gut microbiota. Drug intervention strategies based on the gut-brain axis can advance the treatment of CNS diseases by reducing neuroinflammation and oxidative stress.96 Research by Mishima et al demonstrated that mice with chronic nasal inflammation exhibited significant alterations in the composition of their gut microbiota, with notable sexual dimorphism.97 The gut microbiota regulates the state of nasal inflammation through specific mechanisms, which is then transmitted to higher brain regions via olfactory neural circuits, triggering neuroinflammation in areas such as the piriform cortex, hypothalamus, and amygdala. The gut microbiota may also be involved in the regulation of NBA, warranting further exploration.
Viral pathogens, particularly severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), have garnered substantial attention owing to their capacity to exploit the NBA for entry into the CNS. SARS-CoV-2 binds to angiotensin-converting enzyme 2 (ACE2) receptors expressed on the nasal epithelial cells, facilitating viral replication and subsequent migration to the brain via the olfactory nerve. This form of neuroinvasion has been implicated in anosmia (loss of smell), cognitive impairment, and persistent neurological sequelae such as brain fog and neuroinflammation.98,99 Recent findings have indicated that SARS-CoV-2 induces glial activation and alters neuroimmune homeostasis, linking acute viral infections to long-term neurodegenerative risk factors. Other neurotropic viruses, including herpes simplex virus (HSV) and influenza A virus, similarly utilize the olfactory and trigeminal pathways to invade the CNS, underscoring the susceptibility of the NBA to virus-mediated neuropathology. This growing body of evidence highlights the role of the NBA as a conduit for microbial entry, contributing not only to acute CNS infections, but also to chronic neuroinflammatory conditions and neuropsychiatric sequelae. Understanding the molecular mechanisms underlying microbial neuroinvasion is critical for developing preventive strategies and therapeutics to mitigate CNS pathologies.
Pollen and Other Natural Particles
Natural particles such as pollen are allergens that also contribute to NBA dysfunction. Upon hydration, pollen grains release submicron particles and bioactive molecules such as proteases and lipids. These molecules disrupt the nasal epithelial barrier and activate TLRs, initiating inflammatory responses that can propagate to the CNS. Studies have shown that pollen exposure exacerbates AR and asthma, conditions that are often accompanied by neuroinflammatory responses.100,101 In urban environments, pollen interacts synergistically with air pollutants such as PM2.5, which amplifies immune activation and oxidative stress. These interactions worsen both the respiratory and neurological conditions, linking pollen exposure to a heightened risk of CNS inflammation.102,103 Emerging evidence suggests that pollen-induced neuroinflammation may have long-term effects on cognitive health, although further research is required to confirm these links.
Therapeutic Potential of Micro/Nanoparticles
Biomimetic Micro/Nanoparticles
Biomimetic micro/nanoparticles represent a groundbreaking advancement in the field of drug delivery, particularly for applications targeting the brain via the NBA. These particles are engineered to emulate the structure and function of natural biological systems such as exosomes, cell membranes, and lipoproteins. Their unique ability to mimic the physicochemical and biological properties of natural entities provides several advantages, including enhanced drug compatibility, increased targeting specificity, prolonged circulation time, and minimal immune system recognition. These characteristics make biomimetic nanoparticles highly effective at crossing the BBB and delivering therapeutic agents directly to the CNS.104,105
Recent advances in biomimetic nanoparticle technology have shown significant promise for treating neurodegenerative diseases. A key approach involves using exosome-inspired nanoparticles that mimic the structure and function of natural exosomes—cell-derived vesicles important for intercellular communication. By coating nanoparticles with exosome membranes, researchers have created targeted systems able to deliver therapeutic small interfering RNAs (siRNAs) directly to damaged brain areas.106 For instance, in AD models, such nanoparticles were designed to carry siRNAs against caspase-3 and β-site amyloid precursor protein cleaving enzyme 1 (BACE1).107,108 This approach effectively reduced neuronal death and cleared amyloid plaques, hallmark features of AD’ s pathology. The treatment also improved synaptic integrity and enhanced memory in animal studies, offering a potential non-invasive therapy for AD’s disease. Owing to the success of exosome-based systems, researchers have explored the use of exosome-mimicking nanoparticles modified with specific targeting ligands such as RVG peptides.109 These ligands are known for their high affinity for neuronal cells, making them ideal for enhancing the precision of drug delivery. In Huntington’s disease models, RVG-modified nanoparticles successfully delivered therapeutic RNA molecules to neuronal tissues, resulting in improved motor function and reduced neurodegeneration.110,111
In addition to RNA delivery, the use of bioinspired lipid-coated nanoparticles has emerged as a promising strategy for the treatment of PD. By integrating neural cell membrane components into nanoparticle designs, researchers have developed systems that exhibit superior biocompatibility and retention in the nasal cavity. In addition to facilitating efficient transport to the brainstem, these lipid-coated nanoparticles can also provide neuroprotection by reducing oxidative stress and enhancing mitochondrial function. Preclinical studies have demonstrated that these particles can effectively mitigate the loss of dopaminergic neurons, a critical pathological feature of PD, thereby improving motor coordination and the overall quality of life in animal models.112
Subsequent innovations in biomimetic nanoparticle technology have focused on leveraging hybrid systems that combine natural and synthetic components. For example, cell-membrane-cloaked nanoparticles have been developed to enhance drug delivery and immune evasion. These hybrid systems employ cell membranes from platelets or red blood cells to provide a biologically derived surface coating, which prolongs systemic circulation while enabling active targeting of inflammatory or diseased brain regions.113 These systems have shown promise in preclinical models of multiple sclerosis and traumatic brain injury, in which they have demonstrated the ability to deliver anti-inflammatory and neuroprotective drugs with high precision.114
Another emerging avenue of biomimetic nanoparticle research involves the use of targeted biomimetic vesicles in combination therapies. These vesicles are engineered to co-deliver multiple therapeutic agents such as chemotherapeutics, gene-editing tools, and neuroprotective drugs. For example, biomimetic nanoparticles functionalized with tumor-targeting peptides have been used to deliver a combination of paclitaxel and clustered regularly interspaced short palindromic repeats (CRISPR)-Cas9 constructs in glioblastoma models.115 This approach has been shown to enhance tumor suppression and reduce systemic toxicity, underscoring the potential of biomimetic systems to revolutionize the treatment of aggressive CNS cancers.116
In addition to extensive preclinical studies, significant progress has been made in the clinical translation of exosome-based therapies. Currently, several clinical trials are underway or planned: An exploratory Phase I study of induced pluripotent stem cell-derived exosomes (GD-iEXo-002), administered as nasal drops for refractory focal epilepsy, is ongoing at Peking Union Medical College Hospital in China, sponsored by Guidon Pharmaceutics Ltd; A blinded randomized controlled trial is planned in Moscow, Russia, to investigate intranasal administration of MSC-derived exosomes in extremely low birth weight infants.117
Despite these promising advancements, several challenges regarding clinical translation of biomimetic nanoparticles remain unresolved. Issues such as large-scale manufacturing, stability during storage, and potential immunogenicity must be addressed to ensure their safety and efficacy for human applications.118 Additionally, further studies are required to optimize the pharmacokinetics and biodistribution of these systems, and to evaluate their long-term effects in chronic disease models.119
In conclusion, biomimetic micro/nanoparticles have shown immense potential for transforming the landscape of CNS drug delivery. By emulating the properties of natural biological systems, these nanoparticles offer unprecedented opportunities for targeted, efficient, and non-invasive therapeutic interventions. Future research should focus on refining these technologies and overcoming translational barriers to unlock their full potential in clinical settings.
Synthetic Micro/Nanoparticles
Synthetic micro/nanoparticles have garnered substantial attention as versatile drug-delivery platforms owing to their exceptional ability to be tailored for specific therapeutic applications. Within the context of the NBA, these nanoparticles offer a unique advantage in overcoming the BBB by enabling direct delivery through the olfactory and trigeminal pathways. This targeted approach can improve therapeutic efficacy and minimize systemic side effects, making synthetic nanoparticles highly promising options for treating CNS disorders such as glioblastoma, AD, and PD.
One of the most advanced classes of synthetic nanoparticles is polymeric nanoparticles, which are fabricated from biodegradable materials such as chitosan and PLGA. These polymers are well-known for their biocompatibility, stability, and ability to encapsulate hydrophilic and hydrophobic drugs. Recent studies have explored their application via the nose and brain by designing PLGA-chitosan nanoparticles optimized for nasal retention and mucosal penetration. For instance, these nanoparticles were engineered to co-deliver cetuximab (an epidermal growth factor receptor [EGFR] inhibitor) and metabolic modulators directly to glioblastoma multiforme (GBM) tumor sites via the nasal cavity.114,120 This approach bypassed the systemic circulation and BBB, ensuring precise drug delivery and significantly enhancing anti-tumor efficacy in preclinical models.121 Furthermore, a randomized double-blind trial demonstrated that patients with olfactory dysfunction lasting more than 6 months showed significantly improved olfactory function and a marked reduction in calcium levels in their nasal mucus after receiving chitosan nasal gel.122
Another important class is solid lipid nanoparticles (SLNs), which provide a lipid-based matrix capable of encapsulating lipophilic drugs, protecting them from degradation, and increasing their solubility.123 SLNs have demonstrated remarkable potential for delivering PD therapies through the nose-brain pathway. A notable example is pramipexole-dihydrochloride-loaded SLNs, which were optimized for nasal administration. These nanoparticles showed enhanced uptake through the nasal epithelium and targeted delivery to the substantia nigra of the brain.112 This approach increased dopamine bioavailability, reduced oxidative stress, and mitigated dopaminergic neuronal degeneration in animal models, leading to improved motor function and overall neurological health.124
An emerging innovation in this field is the development of stimuli-responsive nanoparticles designed to release their therapeutic payloads in response to specific conditions, such as ROS, pH changes, or enzymatic activity commonly found in diseased brain regions.125,126 For nose-to-brain applications, ROS-responsive nanoparticles have been engineered to deliver siRNAs targeting the amyloid-beta pathways in AD. When administered intranasally, these nanoparticles utilize the elevated ROS levels in the brain to trigger the release of siRNAs and achieve precise spatiotemporal control.106,127 This strategy significantly reduced amyloid plaque burden and improved cognitive performance in preclinical AD models, underscoring the value of such targeted approaches.105,128
Hybrid nanoparticles combine polymers and lipids to achieve structural stability, improved biocompatibility, and better targeting. They have become promising platforms for nose-to-brain drug delivery by addressing issues like poor mucosal adhesion, short nasal retention, and low BBB permeability.
Sukumar et al developed hybrid nanoparticles with a gelatin methacryloyl (GelMA)-based hydrogel core and a phosphatidylcholine lipid coating for glioblastoma therapy.114 These nanoparticles co-delivered the chemotherapeutic drug temozolomide and the imaging agent indocyanine green via intranasal administration. The GelMA core offered stable, controlled drug release, while the lipid coating enhanced nasal adhesion and retention. This system effectively delivered drugs to brain tumors, improving treatment and enabling real-time imaging.
Raj et al designed lipid–polymer hybrid nanoparticles using a chitosan-grafted polyethylene glycol (PEG) hydrogel and phospholipids to deliver dopamine precursors and antioxidants for Parkinson’s disease.129 Chitosan improved mucoadhesion and sustained release, while PEG increased biocompatibility and reduced immune reactions. Given intranasally, the particles used olfactory and trigeminal pathways to bypass the BBB and reach affected brain regions.112
Bonaccorso et al formulated agarose–lipid hybrid nanoparticles to encapsulate amphotericin B for treating fungal meningitis. The polysaccharide agarose improved drug stability and nasal adhesion, and the lipid layer promoted penetration across the nasal epithelium. Through nose-to-brain pathways, high drug levels were achieved in the meninges, effectively clearing infection while lowering systemic toxicity.130
Ali et al created spanlastic nanoparticles with a deformable lipid bilayer (Span 60 and Tween 80) to deliver piperine for epilepsy management. This flexible system enhanced nasal mucosal penetration, provided prolonged and controlled drug release, and successfully transported piperine to the brain via olfactory and trigeminal routes. As a result, seizure frequency was reduced, neuroprotection was strengthened, and cognitive function was restored in preclinical models.131
Cell-Derived Microparticles
By leveraging the NBA to bypass the BBB and deliver therapeutic molecules directly to the brain, cell-derived microparticles, particularly extracellular vesicles (EVs) such as exosomes, offer promising platforms for CNS therapies. EVs derived from mesenchymal stem cells (MSCs) have shown potential in modulating neuroinflammation and clearing amyloid plaques in AD.132 In addition, neuron-derived EVs (NDEVs) have demonstrated superior neuronal targeting, facilitating synaptic recovery in traumatic brain injury.133,134 In PD, dopaminergic neuron-derived exosomes effectively restored motor function by delivering dopamine and antioxidants to the substantia nigra,134,135 and macrophage-derived EVs, which target tumors with integrin-functionalized vesicles to reduce growth and invasiveness, have been explored for glioblastoma treatment.136 Astrocyte-derived exosomes have also shown promise in the treatment of fungal meningitis, achieving high CNS drug concentrations and reducing inflammation.137,138 Although EVs display remarkable versatility and adaptability in CNS applications, challenges such as scalability, standardized production, and disease-specific targeting optimization must be addressed to ensure successful clinical translation.
In addition to EVs, the intranasal delivery of stem cells represents a transformative and non-invasive approach for CNS therapies, effectively bypassing the BBB via the olfactory and trigeminal pathways. This method capitalizes on the inherent ability of stem cells to home onto injury sites, modulate inflammation, and promote neuroregeneration, indicating their promising therapeutic potential across diverse CNS disorders. For instance, the intranasal delivery of stem cells has shown efficacy in conditions ranging from neurodegenerative diseases such as PD139 and AD140 to glioblastoma141 and ischemic stroke.142 Studies have consistently demonstrated that targeted migration of stem cells to affected brain regions, facilitated by nasal transport mechanisms, results in reduced neuroinflammation, enhanced neuronal repair, and improved functional recovery.143,144 Clinical and preclinical studies have also highlighted the versatility of this approach. Jiang et al demonstrated the preliminary clinical benefits of intranasal neural stem cell transplantation in patients with PD, with significant improvements in motor function observed at six months post-treatment. Jeon et al demonstrated efficient glioblastoma targeting via paracrine signaling and immune modulation. Furthermore, Wei et al demonstrated that intranasally delivered hypoxia-preconditioned MSCs enhanced repair in ischemic stroke models by localizing to ischemic regions and reducing the infarct volume. An open-label, single-center Phase I/II trial evaluating the safety and efficacy of allogenic adipose MSC-derived exosomes in Alzheimer’s disease patients is being conducted at Ruijin Hospital, Shanghai Jiao Tong University, sponsored by Cellular Biomedicine Group Ltd.145 Despite these promising advancements, challenges persist in optimizing the delivery efficiency, scalability, and therapeutic consistency of cell-derived microparticles such as exosomes and stem cells. Key hurdles include standardizing production, increasing targeting precision, and addressing the variability in efficacy across CNS pathologies. Future efforts should focus on refining the engineering strategies and exploring the synergies between exosomes and stem cell technologies to increase their therapeutic potential and clinical applicability.
Inspirations from the NBA for Peripheral-Central Communications
Insights from the NBA to Body-Brain Communication
Body-brain communication has emerged as a transformative framework for understanding the dynamic interactions between peripheral organs and the CNS. Since the discovery of the gut-brain axis, which highlights the role of microbial metabolites in influencing brain function and behavior, studies have revealed several other communication axes, including the heart-brain, lung-brain, liver-brain, kidney-brain, spleen-brain, and muscle-brain axes.146 Each of these pathways provides unique insights into the impact of peripheral organ systems on CNS homeostasis, disease progression, and neuroimmune modulation. For instance, the gut-brain axis, one of the most well-studied body-brain communication systems, highlights the role of gut microbiota-derived metabolites such as SCFAs in regulating neuroinflammation and cognitive function.147 Similarly, the lung-brain axis emphasizes the effect of respiratory pathogens and environmental pollutants on neuroinflammation and neurodegeneration, particularly through systemic immune activation.148 Moreover, the heart-brain axis highlights the intricate relationships among cardiovascular health, cerebral blood flow, and cognitive decline, with increasing evidence linking heart dysfunction to neurodegenerative diseases.149
In addition to the visceral organ axes, sensory organs are gaining recognition for their role in body-brain communication. For example, the eye-brain axis can mediate neurodegenerative and inflammatory processes, and retinal changes are associated with CNS diseases such as AD and multiple sclerosis.150 Similarly, studies on the throat-brain and larynx-brain axes have demonstrated how inflammatory or microbial disruptions in the upper respiratory tract can influence brain function via shared neural pathways.151 As discussed in this review, the NBA represents a distinct and critical communication route because of its direct anatomical connection to the CNS. While other body-brain axes predominantly rely on systemic circulation to transmit signals, the NBA bypasses the BBB through the olfactory and trigeminal pathways. This direct link enables the rapid transport of pathogens, cytokines, and therapeutic agents, establishing the NBA as both a pathogenic and therapeutic pathway. Importantly, the NBA also shares some characteristics with other axes, particularly in its reliance on microbial communities and immune signaling mechanisms to mediate peripheral-central communication. Exploration of body-brain axes, including the gut-brain, heart-brain, and lung-brain systems, has opened new avenues for understanding systemic diseases and their impact on brain health. However, the role of the nasal microbiota, as well as the environmental and inflammatory factors within the NBA, deserves further investigation to determine the full potential of the NBA in modulating neuroimmune pathways and influencing CNS pathology. Future studies focusing on the shared mechanisms across the body-brain axes, such as immune activation, microbial dysbiosis, and neuroinflammation, may provide integrated therapeutic strategies for systemic and CNS disorders.
Advantages of the NBA Over Other Body-Brain Axes
Owing to its anatomical and physiological properties, the NBA is a uniquely efficient pathway for CNS interventions. Unlike the gut-brain and lung-brain axes, which rely on systemic circulation and intermediary processes for signal transduction, the NBA bypasses the BBB via the olfactory and trigeminal nerves. This direct access enables rapid and localized delivery of therapeutic agents such as nanoparticles and stem cells to specific brain regions, minimizing systemic dilution and delays.152 For example, nanoparticles loaded with anti-inflammatory drugs can effectively target inflamed hippocampal regions through the NBA and reduce neuroinflammation without causing systemic toxicity. Furthermore, the NBA can enhance therapeutic precision by minimizing off-target effects that are common in systemic delivery. For instance, dopamine-loaded nanoparticles and siRNA therapies for neurodegenerative diseases such as PD and AD have demonstrated significant improvements in targeting the substantia nigra and amyloid plaques, respectively, while reducing peripheral toxicity.153 Additionally, the non-invasive nature of nasal delivery avoids the risks associated with invasive methods, making it more accessible for patients with chronic conditions and vulnerable populations.
In addition to CNS-targeted therapies, the NBA holds substantial potential for systemic modulation. In comparison with the gut-brain axis, the NBA enables faster signal transmission, where microbial metabolites such as SCFAs rely on slower systemic pathways.154 The NBA can also regulate other body-brain communication systems, such as gut-brain signaling, by modulating vagal nerve activity.155 Olfactory stimulation has been shown to improve autonomic balance and reduce inflammation linked to lung-brain interactions during respiratory infections. Recent advancements in nanotechnology and stem cell-derived exosomes have further enhanced the bioavailability and targeting capabilities of the NBA. These strategies have demonstrated potential as neuroregenerative therapies for ischemic stroke and AD, highlighting the versatility of this axis in addressing a broad range of CNS and systemic disorders.156 This integration of cutting-edge therapeutic strategies positions the NBA as a cornerstone for future systemic and CNS-targeted therapeutic advancements.
Therapeutic Opportunities and Safety Imperatives: “Treating Nasal Diseases Through the Brain” and “Treating Brain Diseases Through the Nose”
The bidirectional delivery facilitated by the NBA offers a foundation for innovative therapeutic strategies, allowing for the integrated treatment of both CNS and nasal disorders. This dual potential opens up transformative opportunities to address diseases through targeted and cross-acting interventions. For “treating brain diseases through the nose”, the intranasal delivery of therapeutic agents has demonstrated significant success in bypassing the BBB to directly target the affected CNS regions. For example, dopamine-loaded nanoparticles can improve motor deficits in PD models by efficiently delivering therapeutic cargo to dopaminergic neurons in the substantia nigra.157 Similarly, exosome-based carriers loaded with siRNAs and neuroprotective agents can reduce amyloid plaque deposition, mitigate neuroinflammation, and promote neuronal repair in AD.158 In addition to neurodegeneration, the intranasal delivery of anti-inflammatory agents and growth factors has shown promise for ischemic stroke recovery by facilitating neuroregeneration and reducing brain tissue damage.159
Conversely, for “treating nasal diseases through the brain”, neuromodulation of CNS circuits offers novel approaches for managing nasal pathologies. For instance, CRS and AR are strongly influenced by neurogenic inflammation mediated through the central neural pathways. Targeting brain areas associated with sensory processing and autonomic control can alleviate nasal inflammation and improve symptom management.160 Studies have also highlighted the potential of brain-derived signaling to influence nasal mucosal immunity, restore epithelial barrier function, and reduce susceptibility to recurrent infections. For example, central interventions that modulate the HPA axis are effective in reducing nasal inflammation triggered by systemic stress responses.161 Furthermore, interventions targeting the nasal microbiome provide exciting avenues for bidirectional therapy. The nasal cavity harbors diverse microbial communities that influence local and CNS health. Dysbiosis of the nasal microbiota has been linked to neurodegenerative and inflammatory conditions such as CRS, cognitive decline, and depression. Regulation of the nasal microbiota through probiotics, antimicrobial agents, or immune-modulating therapies has shown potential for improving nasal mucosal immunity, while indirectly modulating CNS inflammation and cognitive function.146,162
The therapeutic promise of the nose–brain axis is inseparable from a clear-eyed commitment to safety—and to the principles of Safe-and-Sustainable-by-Design.163 The very features that enable non-invasive access to the CNS also open the door to distinct risks. Approaches that aim to treat nasal disease by modulating central circuits need to be assessed not just for efficacy, but for neural selectivity: imprecise targeting could just as easily disrupt olfactory signaling or spill into circuits involved in mood and cognition. On the other side of the equation, intranasal nanoparticles designed to reach the brain bring their own set of concerns—mucosal barrier disruption, accumulation in the olfactory bulb, chronic microglial activation driven by prolonged particle persistence, and unclear clearance trajectories.10 Delivery efficiency alone is no longer sufficient as a benchmark. Translational development must now contend with biodistribution, degradability, residence time, and the distinction between local and remote toxicity. An SSbD-oriented mindset—one that favors biodegradable materials, surface chemistries that evolve after barrier crossing, and dosing regimens that respect the regenerative capacity of the nasal epithelium—needs to move from aspirational to foundational.164 The real question going forward is not whether NBA-based therapeutics can work, but whether they can do so without introducing new, long-term neurobiological liabilities.
Summary and Prospects
The NBA is not only a transformative pathway for drug delivery but also a bidirectional communication system between the nasal cavity and the CNS, facilitating intricate organ–organ interactions. Unlike traditional therapeutic strategies that focus solely on the delivery to the BBB, strategies focused on the NBA are based on the direct influence of peripheral signals, ranging from cytokines to environmental particulates, on CNS health and function. For example, inflammatory cytokines and microRNAs triggered by environmental pollutants, such as PM2.5, propagate neuroinflammation and cognitive impairment through the olfactory and trigeminal pathways. Conversely, therapeutic interventions targeting brain disorders, such as neurodegenerative diseases or neuroinflammation, can utilize the NBA to modulate nasal or peripheral immune responses, exemplifying the potential for “treating brain diseases through the nose” and “treating nasal diseases through the brain”. This reciprocal influence underscores the broader importance of the NBA as an interactive system rather than merely a unidirectional drug-delivery pathway. We have made a summary of potential micro/nanoparticles that play a role in disease pathogenesis and therapy through the NBA which can be seen in Table 1.72,109,110,165–193
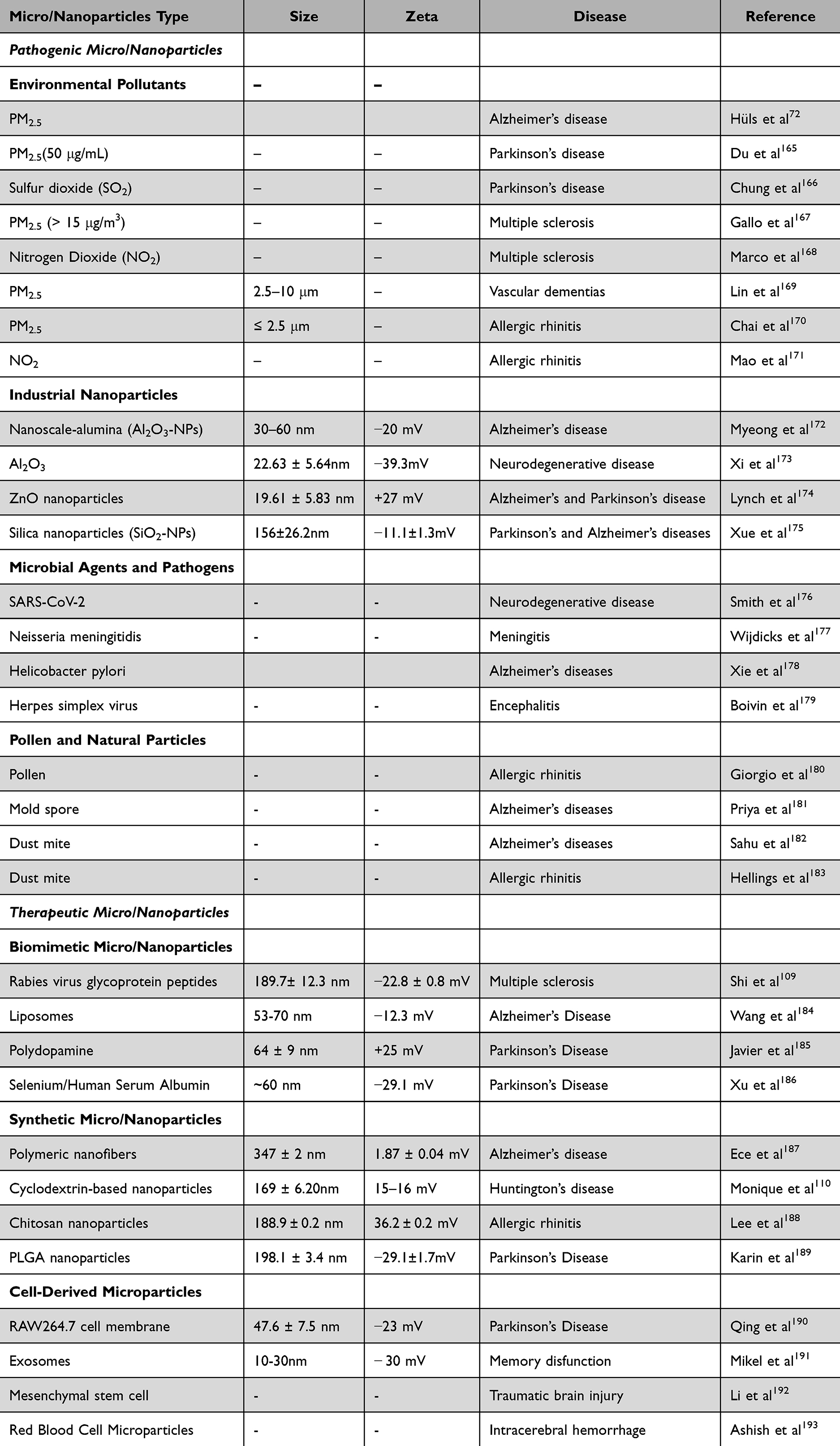 |
Table 1 Potential Micro/Nanoparticles Acting Through the Nose-Brain Axis in Disease Pathogenesis and Treatment |
The implications of the NBA extend beyond CNS-targeted therapies, serving as a model for understanding and leveraging other body-brain communication systems. While axes such as the gut-brain or lung-brain pathways depend on systemic intermediaries, the NBA offers a direct, localized mechanism to study peripheral-to-central signaling and vice versa. By exploring the interplay between nasal inflammation and CNS disorders or brain-directed interventions that mitigate nasal diseases, studies on the NBA can inspire new approaches to address systemic inflammation, neuroimmune crosstalk, and organ-specific pathologies. Moreover, these findings can provide a platform for the integration of environmental toxicology, neuroscience, and precision medicine, thereby driving innovations in diagnostic biomarkers, neuroprotective strategies, and systemic disease management.
Current research on the NBA remains largely at the preclinical stage. Future efforts should prioritize multicenter clinical studies to further evaluate its specific therapeutic efficacy. Building on this, machine learning techniques—such as causal inference models and graph neural networks—can be employed to integrate multimodal data (eg, proteomic, metabolomic, and microbiome profiles). This approach aims to reverse-engineer the core pathway networks connecting peripheral biomarkers with central imaging features, thereby uncovering novel therapeutic targets. Additionally, predictive models based on baseline clinical, biochemical, and imaging characteristics can be constructed to simulate treatment outcomes and optimize patient selection prior to therapy, thus maximizing the efficiency of medical resources. By systematically decoding the language of the nasal-brain axis and leveraging intelligent technologies to translate this knowledge into personalized and preventive diagnostic and therapeutic strategies, our ultimate goal extends beyond restoring a single sensory function. This integrated perspective will position the NBA not only as a gateway for CNS therapies, but also as a foundational framework for advancing the understanding of peripheral-to-central communications and inter-organ interactions across the body.
Data Sharing Statement
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Key Research and Development Program of Shandong Province (Competitive Innovation Platform Project: 2024CXPT091; Major Science and Technology Innovation Project: 2022CXGC020506); the National Natural Science Foundation of China (82571293, 82371119, 82271146); the Taishan Scholar Project (tstp20250512, tsqn202312392); the Natural Science Foundation of Shandong Province (ZR2024MH034); the Ministry of Education Industry-University-Research Innovation Fund for Chinese Universities: New Technology and Medical Integration Innovation Special Project (2025XJS011); Science and Technology Innovation development plan of Yantai (2023ZDCX033); the youth talent training program from Qingdao medical college of Qingdao University (No. RZ2300002690).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Wang Y, Song XY, Wei SZ, et al. Brain response in allergic rhinitis: profile and proposal. J Neurosci Res. 2023;101(4):480–26. doi:10.1002/jnr.25159
2. Bahadur S, Pardhi DM, Rautio J, Rosenholm JM, Pathak K. Intranasal nanoemulsions for direct nose-to-brain delivery of actives for CNS disorders. Pharmaceutics. 2020;12(12):1230. doi:10.3390/pharmaceutics12121230
3. Reid WK. Neurodegenerative diseases due to neurotoxins passing through the nose-to-brain pathway. Med Res Arch. 2023;11(6). doi:10.18103/mra.v11i6.3898
4. Chacko A, Delbaz A, Choudhury IN, et al. Streptococcus agalactiae infects glial cells and invades the central nervous system via the olfactory and trigeminal nerves. Front Cell Infect Microbiol. 2022;12:793416. doi:10.3389/fcimb.2022.793416
5. St John JA, Walkden H, Nazareth L, et al. Burkholderia pseudomallei rapidly infects the brain stem and spinal cord via the trigeminal nerve after intranasal inoculation. Infect Immun. 2016;84(9):2681–2688. doi:10.1128/IAI.00361-16
6. Delbaz A, Chen M, Jen FE, et al. Neisseria meningitidis induces pathology-associated cellular and molecular changes in trigeminal schwann cells. Infect Immun. 2020;88(4). doi:10.1128/IAI.00955-19
7. Mou YK, Wang HR, Zhang WB, Zhang Y, Ren C, Song XC. Allergic rhinitis and depression: profile and proposal. Front Psychiatry. 2021;12:820497. doi:10.3389/fpsyt.2021.820497
8. Nguyen TT, Maeng HJ. Pharmacokinetics and pharmacodynamics of intranasal solid lipid nanoparticles and nanostructured lipid carriers for nose-to-brain delivery. Pharmaceutics. 2022;14(3):572. doi:10.3390/pharmaceutics14030572
9. Khatri DK, Preeti K, Tonape S, et al. Nanotechnological advances for nose to brain delivery of therapeutics to improve the parkinson therapy. Curr Neuropharmacol. 2023;21(3):493–516. doi:10.2174/1570159X20666220507022701
10. Gandhi S, Shastri DH, Shah J, Nair AB, Jacob S. Nasal delivery to the brain: harnessing nanoparticles for effective drug transport. Pharmaceutics. 2024;16(4):481. doi:10.3390/pharmaceutics16040481
11. Lochhead JJ, Thorne RG. Intranasal delivery of biologics to the central nervous system. Adv Drug Deliv Rev. 2012;64(7):614–628. doi:10.1016/j.addr.2011.11.002
12. Khan AR, Liu M, Khan MW, Zhai G. Progress in brain targeting drug delivery system by nasal route. J Control Release. 2017;268:364–389. doi:10.1016/j.jconrel.2017.09.001
13. Cho Y, Seo EU, Hwang KS, Kim H, Choi J, Kim HN. Evaluation of size-dependent uptake, transport and cytotoxicity of polystyrene microplastic in a blood-brain barrier (BBB) model. Nano Converg. 2024;11(1):40. doi:10.1186/s40580-024-00448-z
14. Lochhead JJ, Davis TP. Perivascular and perineural pathways involved in brain delivery and distribution of drugs after intranasal administration. Pharmaceutics. 2019;11(11):598. doi:10.3390/pharmaceutics11110598
15. Awad R, Avital A, Sosnik A. Polymeric nanocarriers for nose-to-brain drug delivery in neurodegenerative diseases and neurodevelopmental disorders. Acta Pharm Sin B. 2023;13(5):1866–1886. doi:10.1016/j.apsb.2022.07.003
16. Ruan S, Li J, Ruan H, et al. Microneedle-mediated nose-to-brain drug delivery for improved Alzheimer’s disease treatment. J Control Release. 2024;366:712–731. doi:10.1016/j.jconrel.2024.01.013
17. Goel H, Kalra V, Verma SK, Dubey SK, Tiwary AK. Convolutions in the rendition of nose to brain therapeutics from bench to bedside: feats & fallacies. J Control Release. 2022;341:782–811. doi:10.1016/j.jconrel.2021.12.009
18. Mwema A, Muccioli GG, Des Rieux A. Innovative drug delivery strategies to the CNS for the treatment of multiple sclerosis. J Control Release. 2023;364:435–457. doi:10.1016/j.jconrel.2023.10.052
19. Agrawal M, Saraf S, Saraf S, et al. Recent strategies and advances in the fabrication of nano lipid carriers and their application towards brain targeting. J Control Release. 2020;321:372–415. doi:10.1016/j.jconrel.2020.02.020
20. Wong CYJ, Baldelli A, Hoyos CM, Tietz O, Ong HX, Traini D. Insulin delivery to the brain via the nasal route: unraveling the potential for alzheimer’s disease therapy. Drug Deliv Transl Res. 2024;14(7):1776–1793. doi:10.1007/s13346-024-01558-1
21. Goli VVN, Tatineni S, Hani U, et al. Pharmacokinetics and pharmacodynamics of a nanostructured lipid carrier co-encapsulating artemether and miRNA for mitigating cerebral malaria. Pharmaceuticals. 2024;17(4):466. doi:10.3390/ph17040466
22. Costa CP, Moreira JN, Sousa Lobo JM, Silva AC. Intranasal delivery of nanostructured lipid carriers, solid lipid nanoparticles and nanoemulsions: a current overview of in vivo studies. Acta Pharm Sin B. 2021;11(4):925–940. doi:10.1016/j.apsb.2021.02.012
23. Shen X, Cui Z, Wei Y, et al. Exploring the potential to enhance drug distribution in the brain subregion via intranasal delivery of nanoemulsion in combination with borneol as a guider. Asian J Pharm Sci. 2023;18(6):100778.
24. Jiang L, Wang AD, Li B, Cui TH, Lu YF. Electrons dynamics control by shaping femtosecond laser pulses in micro/nanofabrication: modeling, method, measurement and application. Light Sci Appl. 2018;7:17134.
25. Romashchenko A, Petrovski D, Trotsky SY, et al. High resolution quantitative tracing and modulation of nanoparticles’ nose-to-brain transmission. J Phys. 2020;1461.
26. Adams F, Zimmermann CM, Baldassi D, et al. Pulmonary siRNA delivery with sophisticated amphiphilic poly(Spermine Acrylamides) for the treatment of lung fibrosis. Small. 2024;20(22):e2308775. doi:10.1002/smll.202308775
27. Lu B, Wei L, Shi G, Du J. Nanotherapeutics for alleviating anesthesia-associated complications. Adv Sci. 2024;11(15):e2308241. doi:10.1002/advs.202308241
28. Zha S, Wong KL, All AH. Intranasal delivery of functionalized polymeric nanomaterials to the brain. Adv Healthc Mater. 2022;11(11):e2102610. doi:10.1002/adhm.202102610
29. Zheng Y, Cui L, Lu H, et al. Nose to brain: exploring the progress of intranasal delivery of solid lipid nanoparticles and nanostructured lipid carriers. Int J Nanomed. 2024;19:12343–12368. doi:10.2147/IJN.S497480
30. Esim O, Savaser A, Ozkan CK, et al. Nose to brain delivery of eletriptan hydrobromide nanoparticles: preparation, in vitro/in vivo evaluation and effect on trigeminal activation. J Drug Delivery Sci Technol. 2020;59:101919. doi:10.1016/j.jddst.2020.101919
31. Khodabandelou S, Nazem Z, Komaki A, et al. Development of silibinin-loaded nanostructured lipid carriers for Alzheimer’s disease induced by amyloid beta in Wistar rats. J Mater Chem B. 2024;12(44):11426–11443. doi:10.1039/D4TB00775A
32. Wu D, Chen Q, Chen X, Han F, Chen Z, Wang Y. The blood-brain barrier: structure, regulation, and drug delivery. Signal Transduct Target Ther. 2023;8(1):217.
33. Shukla R, Kumar A, Flora SJS. Chapter 5 - Nanotechnological advances in direct nose-to-brain drug delivery for neurodegenerative disorders and other neuroailments. In: Pardeshi CV, Souto EB, editors. Direct Nose-to-Brain Drug Delivery. Academic Press; 2021:73–91.
34. Yuan T, Zheng R, Liu J, et al. Role of yes-associated protein in interleukin-13 induced nasal remodeling of chronic rhinosinusitis with nasal polyps. Allergy. 2021;76(2):600–604. doi:10.1111/all.14699
35. Kotas ME, Patel NN, Cope EK, et al. IL-13-associated epithelial remodeling correlates with clinical severity in nasal polyposis. J Allergy Clin Immunol. 2023;151(5):1277–1285. doi:10.1016/j.jaci.2022.12.826
36. Carsuzaa F, Bequignon E, Bainaud M, et al. Oncostatin M counteracts the fibrotic effects of TGF-beta1 and IL-4 on nasal-polyp-derived fibroblasts: a control of fibrosis in chronic rhinosinusitis with nasal polyps? Int J Mol Sci. 2022;23(11):6308. doi:10.3390/ijms23116308
37. Bachert C, Hicks A, Gane S, et al. The interleukin-4/interleukin-13 pathway in type 2 inflammation in chronic rhinosinusitis with nasal polyps. Front Immunol. 2024;15:1356298.
38. Jiang L, Tang H, Lin T, et al. Epithelium-derived kallistatin promotes CD4(+) T-cell chemotaxis to T(H)2-type inflammation in chronic rhinosinusitis. J Allergy Clin Immunol. 2024;154(1):120–130.
39. Hara Y, Jha MK, Mattoo H, et al. Interleukin 4 directly activates olfactory neurons and induces loss of smell in mice. J Allergy clin Immunol. 2023;151(2):AB128. doi:10.1016/j.jaci.2022.12.403
40. Wangler LM, Godbout JP. Microglia moonlighting after traumatic brain injury: aging and interferons influence chronic microglia reactivity. Trends Neurosci. 2023;46(11):926–940.
41. Tabor JB, Penner LC, Galarneau JM, et al. Plasma biomarkers of traumatic brain injury in adolescents with sport-related concussion. JAMA Network Open. 2024;7(9):e2431959.
42. Liu P, Qin D, Deng Z, et al. TET2 deficiency exacerbates nasal polypogenesis by inducing epithelial-to-mesenchymal transition. Allergy. 2022;77(11):3452–3455. doi:10.1111/all.15446
43. Yan B, Lan F, Li J, Wang C, Zhang L. The mucosal concept in chronic rhinosinusitis: focus on the epithelial barrier. J Allergy Clin Immunol. 2024;153(5):1206–1214. doi:10.1016/j.jaci.2024.01.015
44. Rubio T, Viana R, Moreno-Estelles M, Campos-Rodriguez A, Sanz P. TNF and IL6/Jak2 signaling pathways are the main contributors of the glia-derived neuroinflammation present in Lafora disease, a fatal form of progressive myoclonus epilepsy. Neurobiol Dis. 2023;176:105964.
45. Gao C, Jiang J, Tan Y, Chen S. Microglia in neurodegenerative diseases: mechanism and potential therapeutic targets. Signal Transduct Target Ther. 2023;8(1):359. doi:10.1038/s41392-023-01588-0
46. Walker KA, Le Page LM, Terrando N, Duggan MR, Heneka MT, Bettcher BM. The role of peripheral inflammatory insults in alzheimer’s disease: a review and research roadmap. Mol Neurodegener. 2023;18(1):37.
47. Le Thuc O, Garcia-Caceres C. Obesity-induced inflammation: connecting the periphery to the brain. Nat Metab. 2024;6(7):1237–1252.
48. Zhao Y, Chen J, Hao Y, et al. Predicting the recurrence of chronic rhinosinusitis with nasal polyps using nasal microbiota. Allergy. 2022;77(2):540–549.
49. Gioacchini FM, Ferlito S, Ralli M, et al. Nasal microbiota and neuroinflammation: relationship between nasal flora and multiple sclerosis onset/progression. Life. 2022;12(12). doi:10.3390/life12122043
50. Bang S, Shin YH, Ma X, et al. A cardiolipin from Muribaculum intestinale induces antigen-specific cytokine responses. J Am Chem Soc. 2023;145(43):23422–23426. doi:10.1021/jacs.3c09734
51. Ma C, Hao X, Gao L, et al. Extracellular vesicles released from macrophages infected with Mycoplasma pneumoniae stimulate proinflammatory response via the TLR2-NF-kappaB/JNK signaling pathway. Int J Mol Sci. 2023;24(10).
52. Jiang Z, Li S, Zhu C, Zhou R, Leung PHM. Mycoplasma pneumoniae infections: pathogenesis and vaccine development. Pathogens. 2021;10(2). doi:10.3390/pathogens10020119
53. Yu XC, Yang JJ, Jin BH, et al. A strategy for bypassing the blood-brain barrier: facial intradermal brain-targeted delivery via the trigeminal nerve. J Control Release. 2017;258:22–33. doi:10.1016/j.jconrel.2017.05.001
54. Chidambaram SB, Essa MM, Rathipriya AG, et al. Gut dysbiosis, defective autophagy and altered immune responses in neurodegenerative diseases: tales of a vicious cycle. Pharmacol Ther. 2022;231:107988. doi:10.1016/j.pharmthera.2021.107988
55. Li B, Wang D, Zhang C, et al. Role of respiratory system microbiota in development of lung cancer and clinical application. Imeta. 2024;3(5):e232. doi:10.1002/imt2.232
56. Choudhury IN, Chacko A, Delbaz A, et al. Antimicrobial responses of peripheral and central nervous system glia against Staphylococcus aureus. Sci Rep. 2021;11(1):10722. doi:10.1038/s41598-021-90252-0
57. Zhou X, Kumar P, Bhuyan DJ, Jensen SO, Roberts TL, Munch GW. Neuroinflammation in alzheimer’s disease: a potential role of nose-picking in pathogen entry via the olfactory system? Biomolecules. 2023;13(11):1568. doi:10.3390/biom13111568
58. Yu Y, Tang X, Zhou L, et al. Cascade-targeting polymeric particles eliminate intracellular C. neoformans in fungal infection therapy. J Control Release. 2024;373:399–409. doi:10.1016/j.jconrel.2024.07.047
59. Varesi A, Campagnoli LIM, Fahmideh F, et al. The interplay between gut microbiota and parkinson’s disease: implications on diagnosis and treatment. Int J Mol Sci. 2022;23(20). doi:10.3390/ijms232012289
60. Wang C, Zong S, Cui X, et al. The effects of microglia-associated neuroinflammation on alzheimer’s disease. Front Immunol. 2023;14:1117172. doi:10.3389/fimmu.2023.1117172
61. Nayak V, Patra S, Rout S, et al. Regulation of neuroinflammation in alzheimer’s disease via nanoparticle-loaded phytocompounds with anti-inflammatory and autophagy-inducing properties. Phytomedicine. 2024;122:155150. doi:10.1016/j.phymed.2023.155150
62. Hawkes CH, Del Tredici K, Braak H. Parkinson’s disease: a dual-hit hypothesis. Neuropathol Appl Neurobiol. 2007;33(6):599–614. doi:10.1111/j.1365-2990.2007.00874.x
63. Borghammer P, Just MK, Horsager J, et al. A postmortem study suggests a revision of the dual-hit hypothesis of Parkinson’s disease. NPJ Parkinsons Dis. 2022;8(1):166. doi:10.1038/s41531-022-00436-2
64. Graham UM, Pinto JM, Weuve J, et al. Nose-to-brain translocation of inhaled ultrafine elongated particles: facts and mysteries. Front Toxicol. 2025;7:1655149. doi:10.3389/ftox.2025.1655149
65. Fullard ME, Morley JF, Duda JE. Olfactory dysfunction as an early biomarker in parkinson’s disease. Neurosci Bull. 2017;33(5):515–525. doi:10.1007/s12264-017-0170-x
66. Dan X, Wechter N, Gray S, Mohanty JG, Croteau DL, Bohr VA. Olfactory dysfunction in aging and neurodegenerative diseases. Ageing Res Rev. 2021;70:101416. doi:10.1016/j.arr.2021.101416
67. Mathias K, Petronilho F, Danielski LG. Nose-to-brain axis: mechanistic links between nasal microbiome dysbiosis, neuroinflammation, and brain disorders. Neuroscience. 2026;598:19–28. doi:10.1016/j.neuroscience.2026.01.039
68. Shou Y, Huang Y, Zhu X, Liu C, Hu Y, Wang H. A review of the possible associations between ambient PM2.5 exposures and the development of Alzheimer’s disease. Ecotoxicol Environ Saf. 2019;174:344–352. doi:10.1016/j.ecoenv.2019.02.086
69. Cristaldi A, Fiore M, Oliveri Conti G, et al. Possible association between PM(2.5) and neurodegenerative diseases: a systematic review. Environ Res. 2022;208:112581. doi:10.1016/j.envres.2021.112581
70. Liu C, Meng L, Gao Y, et al. PM2.5 triggers tau aggregation in a mouse model of tauopathy. JCI Insight. 2024;9(14). doi:10.1172/jci.insight.176703
71. Lee SH, Lin CY, Chen TF, et al. Distinct brain lipid signatures in response to low-level PM(2.5) exposure in a 3xTg-alzheimer’s disease mouse inhalation model. Sci Total Environ. 2022;838(Pt 4):156456. doi:10.1016/j.scitotenv.2022.156456
72. Li Z, Liang D, Ebelt S, et al. Differential DNA methylation in the brain as potential mediator of the association between traffic-related PM(2.5) and neuropathology markers of alzheimer’s disease. Alzheimers Dement. 2024;20(4):2538–2551. doi:10.1002/alz.13650
73. Calderon-Garciduenas L, Ayala A. Air pollution, ultrafine particles, and your brain: are combustion nanoparticle emissions and engineered nanoparticles causing preventable fatal neurodegenerative diseases and common neuropsychiatric outcomes? Environ Sci Technol. 2022;56(11):6847–6856. doi:10.1021/acs.est.1c04706
74. Liu F, Liu C, Liu Y, Wang J, Wang Y, Yan B. Neurotoxicity of the air-borne particles: from molecular events to human diseases. J Hazard Mater. 2023;457:131827. doi:10.1016/j.jhazmat.2023.131827
75. Amato-Lourenco LF, Dantas KC, Junior GR, et al. Microplastics in the olfactory bulb of the human brain. JAMA Network Open. 2024;7(9):e2440018. doi:10.1001/jamanetworkopen.2024.40018
76. Nihart AJ, Garcia MA, El Hayek E, et al. Bioaccumulation of microplastics in decedent human brains. Nat Med. 2025;31(4):1114–1119. doi:10.1038/s41591-024-03453-1
77. Huang H, Hou J, Li M, Wei F, Liao Y, Xi B. Microplastics in the bloodstream can induce cerebral thrombosis by causing cell obstruction and lead to neurobehavioral abnormalities. Sci Adv. 2025;11(4):eadr8243. doi:10.1126/sciadv.adr8243
78. Prust M, Meijer J, Westerink RHS. The plastic brain: neurotoxicity of micro- and nanoplastics. Part Fibre Toxicol. 2020;17(1):24. doi:10.1186/s12989-020-00358-y
79. Yan Y, Huang W, Lu X, et al. Zinc oxide nanoparticles induces cell death and consequently leading to incomplete neural tube closure through oxidative stress during embryogenesis. Cell Biol Toxicol. 2024;40(1):51. doi:10.1007/s10565-024-09894-1
80. Yan Y, Wang G, Luo X, et al. Endoplasmic reticulum stress-related calcium imbalance plays an important role on Zinc oxide nanoparticles-induced failure of neural tube closure during embryogenesis. Environ Int. 2021;152:106495. doi:10.1016/j.envint.2021.106495
81. Sun W, Lu Z, Chen X, et al. Aluminum oxide nanoparticles impair working memory and neuronal activity through the GSK3beta/BDNF signaling pathway of prefrontal cortex in rats. ACS Chem Neurosci. 2022;13(23):3352–3361. doi:10.1021/acschemneuro.2c00383
82. Sun W, Li J, Li X, et al. Aluminium oxide nanoparticles compromise spatial memory performance and proBDNF-mediated neuronal function in the hippocampus of rats. Part Fibre Toxicol. 2022;19(1):34. doi:10.1186/s12989-022-00477-8
83. Park SJ, Lee J, Lee S, et al. Exposure of ultrafine particulate matter causes glutathione redox imbalance in the hippocampus: a neurometabolic susceptibility to alzheimer’s pathology. Sci Total Environ. 2020;718:137267. doi:10.1016/j.scitotenv.2020.137267
84. Mussalo L, Lampinen R, Avesani S, et al. Traffic-related ultrafine particles impair mitochondrial functions in human olfactory mucosa cells - implications for alzheimer’s disease. Redox Biol. 2024;75:103272. doi:10.1016/j.redox.2024.103272
85. Hou S, Li C, Wang Y, et al. Silica nanoparticles cause activation of NLRP3 inflammasome in-vitro model-using microglia. Int J Nanomed. 2022;17:5247–5264. doi:10.2147/IJN.S372485
86. Wei W, Sun H, Yang B, Song E, Song Y. Coronal ApoE protein combines with LRP1 to inactivate GSK3β that mitigates silica nanoparticle-induced brain lesion. ACS Chem Neurosci. 2024;15(4):808–815. doi:10.1021/acschemneuro.3c00728
87. Heilgeist S, Sekine R, Sahin O, Stewart RA. Finding nano: challenges involved in monitoring the presence and fate of engineered titanium dioxide nanoparticles in aquatic environments. Water. 2021;13(5):734. doi:10.3390/w13050734
88. Gatou M-A, Syrrakou A, Lagopati N, Pavlatou EA. Photocatalytic TiO2-based nanostructures as a promising material for diverse environmental applications: a review. Reactions. 2024;5(1):135–194.
89. Butowt R, von Bartheld CS. The route of SARS-CoV-2 to brain infection: have we been barking up the wrong tree? Mol Neurodegener. 2022;17(1):20. doi:10.1186/s13024-022-00529-9
90. Du W, Li H, Tian B, et al. Development of nose-to-brain delivery of ketoconazole by nanostructured lipid carriers against cryptococcal meningoencephalitis in mice. Colloids Surf B Biointerfaces. 2019;183:110446. doi:10.1016/j.colsurfb.2019.110446
91. Gay F. Bacterial transportable toxins of the nasopharyngeal microbiota in multiple sclerosis. Nose-to-Brain Direct Rev Neurol. 2019;175(10):644–649.
92. Xiang G, Wang Y, Ni K, et al. Nasal Staphylococcus aureus carriage promotes depressive behaviour in mice via sex hormone degradation. Nat Microbiol. 2025;10(10):2425–2440. doi:10.1038/s41564-025-02120-6
93. Zhang S, Wu Z, Zhang S, et al. The intricate microbial-gut-brain axis in alzheimer’s disease: a review of microbiota-targeted strategies. Food Funct. 2025;16(21):8320–8344. doi:10.1039/D5FO03139G
94. Zhuang C, Chen Q, Dou X, et al. Pathogenic mechanisms of cerebral ischemia and potential gut-brain axis-oriented therapeutic strategies. Phytomedicine. 2025;148:157264. doi:10.1016/j.phymed.2025.157264
95. Hu E, Li Z, Li T, et al. A novel microbial and hepatic biotransformation-integrated network pharmacology strategy explores the therapeutic mechanisms of bioactive herbal products in neurological diseases: the effects of astragaloside IV on intracerebral hemorrhage as an example. Chin Med. 2023;18(1):40. doi:10.1186/s13020-023-00745-5
96. Hui Z, Lai-Fa W, Xue-Qin W, Ling D, Bin-Sheng H, Li JM. Mechanisms and therapeutic potential of chinonin in nervous system diseases. J Asian Nat Prod Res. 2024;26(12):1405–1420. doi:10.1080/10286020.2024.2371040
97. Mishima Y, Osaki T, Shimada A, Kamiya S, Hasegawa-Ishii S. Sex-dependent differences in the gut microbiota following chronic nasal inflammation in adult mice. Sci Rep. 2021;11(1):4640. doi:10.1038/s41598-021-83896-5
98. Takahara Y, Sumitomo T, Kono M, et al. Pneumolysin contributes to dysfunction of nasal epithelial barrier for promotion of pneumococcal dissemination into brain tissue. mSphere. 2024;9(10):e0065524. doi:10.1128/msphere.00655-24
99. Borisova T, Komisarenko S. Air pollution particulate matter as a potential carrier of SARS-CoV-2 to the nervous system and/or neurological symptom enhancer: arguments in favor. Environ Sci Pollut Res Int. 2021;28(30):40371–40377. doi:10.1007/s11356-020-11183-3
100. Shahbaz MA, Kuivanen S, Mussalo L, et al. Exposure to urban particulate matter alters responses of olfactory mucosal cells to SARS-CoV-2 infection. Environ Res. 2024;249:118451. doi:10.1016/j.envres.2024.118451
101. Schiavoni G, D’Amato G, Afferni C. The dangerous liaison between pollens and pollution in respiratory allergy. Ann Allergy Asthma Immunol. 2017;118(3):269–275. doi:10.1016/j.anai.2016.12.019
102. Visez N, Hamze M, Vandenbossche K, et al. Uptake of ozone by allergenic pollen grains. Environ Pollut. 2023;331(Pt 1):121793. doi:10.1016/j.envpol.2023.121793
103. Lindqvist M, Leth-Moller KB, Linneberg A, et al. Natural course of pollen-induced allergic rhinitis from childhood to adulthood: a 20-year follow up. Allergy. 2024;79(4):884–893. doi:10.1111/all.15927
104. Papapostolou G, Kiotseridis H, Romberg K, et al. Cognitive dysfunction and quality of life during pollen season in children with seasonal allergic rhinitis. Pediatr Allergy Immunol. 2021;32(1):67–76. doi:10.1111/pai.13328
105. Li J, Peng H, Zhang W, et al. Enhanced Nose-to-brain delivery of combined small interfering rnas using lesion-recognizing nanoparticles for the synergistic therapy of alzheimer’s disease. ACS Appl Mater Interfaces. 2023;15(46):53177–53188. doi:10.1021/acsami.3c08756
106. Yang X, Yang W, Xia X, et al. Intranasal delivery of BACE1 siRNA and rapamycin by dual targets modified nanoparticles for alzheimer’s disease therapy. Small. 2022;18(30):e2203182. doi:10.1002/smll.202203182
107. Zhou Y, Zhu F, Liu Y, et al. Blood-brain barrier-penetrating siRNA nanomedicine for alzheimer’s disease therapy. Sci Adv. 2020;6(41). doi:10.1126/sciadv.abc7031
108. Wang P, Zheng X, Guo Q, et al. Systemic delivery of BACE1 siRNA through neuron-targeted nanocomplexes for treatment of alzheimer’s disease. J Control Release. 2018;279:220–233. doi:10.1016/j.jconrel.2018.04.034
109. Yin N, Zhao Y, Liu C, et al. Engineered nanoerythrocytes alleviate central nervous system inflammation by regulating the polarization of inflammatory microglia. Adv Mater. 2022;34(27):e2201322. doi:10.1002/adma.202201322
110. Mendonça MCP, Cronin MF, Cryan JF, O’Driscoll CM. Modified cyclodextrin-based nanoparticles mediated delivery of siRNA for huntingtin gene silencing across an in vitro BBB model. Eur J Pharm Biopharm. 2021;169:309–318. doi:10.1016/j.ejpb.2021.11.003
111. Zhang L, Wu T, Shan Y, et al. Therapeutic reversal of Huntington’s disease by in vivo self-assembled siRNAs. Brain. 2021;144(11):3421–3435. doi:10.1093/brain/awab354
112. Raj R, Wairkar S, Sridhar V, Gaud R. Pramipexole dihydrochloride loaded chitosan nanoparticles for nose to brain delivery: development, characterization and in vivo anti-Parkinson activity. Int J Biol Macromol. 2018;109:27–35. doi:10.1016/j.ijbiomac.2017.12.056
113. Chang Y, Cai X, Syahirah R, et al. CAR-neutrophil mediated delivery of tumor-microenvironment responsive nanodrugs for glioblastoma chemo-immunotherapy. Nat Commun. 2023;14(1):2266. doi:10.1038/s41467-023-37872-4
114. Sukumar UK, Bose RJC, Malhotra M, et al. Intranasal delivery of targeted polyfunctional gold-iron oxide nanoparticles loaded with therapeutic microRNAs for combined theranostic multimodality imaging and presensitization of glioblastoma to temozolomide. Biomaterials. 2019;218:119342. doi:10.1016/j.biomaterials.2019.119342
115. Tu K, Deng H, Kong L, et al. Reshaping tumor immune microenvironment through acidity-responsive nanoparticles featured with CRISPR/Cas9-mediated programmed death-ligand 1 attenuation and chemotherapeutics-induced immunogenic cell death. ACS Appl Mater Interfaces. 2020;12(14):16018–16030. doi:10.1021/acsami.9b23084
116. Boyuklieva R, Pilicheva B. Micro- and nanosized carriers for nose-to-brain drug delivery in neurodegenerative disorders. Biomedicines. 2022;10(7):1706. doi:10.3390/biomedicines10071706
117. Gotoh S, Kawabori M, Fujimura M. Intranasal administration of stem cell-derived exosomes for central nervous system diseases. Neural Regen Res. 2024;19(6):1249–1255. doi:10.4103/1673-5374.385875
118. Mistry A, Stolnik S, Illum L. Nose-to-brain delivery: investigation of the transport of nanoparticles with different surface characteristics and sizes in excised porcine olfactory epithelium. Mol Pharm. 2015;12(8):2755–2766. doi:10.1021/acs.molpharmaceut.5b00088
119. Elkomy MH, Abo El-Ela FI, Zaki RM, et al. Intranasal nanotransferosomal gel for quercetin brain targeting: II. antidepressant effect in an experimental animal model. Pharmaceutics. 2023;15(8):2095. doi:10.3390/pharmaceutics15082095
120. Ferreira NN, Granja S, Boni FI, et al. Modulating chitosan-PLGA nanoparticle properties to design a co-delivery platform for glioblastoma therapy intended for nose-to-brain route. Drug Deliv Transl Res. 2020;10(6):1729–1747. doi:10.1007/s13346-020-00824-2
121. Ferreira NN, de Oliveira Junior E, Granja S, et al. Nose-to-brain co-delivery of drugs for glioblastoma treatment using nanostructured system. Int J Pharm. 2021;603:120714. doi:10.1016/j.ijpharm.2021.120714
122. Algarni MA, Abdelazim MH, Alsulami FT, et al. Therapeutic potential of chitosan nasal gel in addressing olfactory dysfunction: a clinical trial and mechanistic study. Am J Rhinol Allergy. 2025;39(3):237–244. doi:10.1177/19458924251322058
123. Becker Peres L, Becker Peres L, de Araujo PHH, Sayer C. Solid lipid nanoparticles for encapsulation of hydrophilic drugs by an organic solvent free double emulsion technique. Colloids Surf B Biointerfaces. 2016;140:317–323. doi:10.1016/j.colsurfb.2015.12.033
124. Trivedi R, Minglani VV, El-Gazzar AM, et al. Optimization of pramipexole-loaded in situ thermosensitive intranasal gel for parkinson’s disease. Pharmaceuticals. 2024;17(2):172. doi:10.3390/ph17020172
125. Cao Z, Zuo X, Liu X, Xu G, Yong KT. Recent progress in stimuli-responsive polymeric micelles for targeted delivery of functional nanoparticles. Adv Colloid Interface Sci. 2024;330:103206. doi:10.1016/j.cis.2024.103206
126. Zhou H, Chen DS, Hu CJ, Hong X, Shi J, Xiao Y. Stimuli-responsive nanotechnology for RNA delivery. Adv Sci. 2023;10(36):e2303597. doi:10.1002/advs.202303597
127. Wang C, Song X, Li P, et al. Multifunctional nanocarrier for synergistic treatment of alzheimer’s disease by inhibiting beta-amyloid aggregation and scavenging reactive oxygen species. ACS Appl Mater Interfaces. 2024;16(21):27127–27138. doi:10.1021/acsami.4c02825
128. Lee D, Shen AM, Shah M, Garbuzenko OB, Minko T. In vivo evaluation of nose-to-brain delivery of liposomal donepezil, memantine, and BACE-1 siRNA for alzheimer’s disease therapy. Int J Mol Sci. 2024;25(19):10357.
129. Bhattamisra SK, Shak AT, Xi LW, et al. Nose to brain delivery of rotigotine loaded chitosan nanoparticles in human SH-SY5Y neuroblastoma cells and animal model of parkinson’s disease. Int J Pharm. 2020;579:119148. doi:10.1016/j.ijpharm.2020.119148
130. Bonaccorso A, Musumeci T, Serapide MF, Pellitteri R, Uchegbu IF, Puglisi G. Nose to brain delivery in rats: effect of surface charge of rhodamine B labeled nanocarriers on brain subregion localization. Colloids Surf B Biointerfaces. 2017;154:297–306. doi:10.1016/j.colsurfb.2017.03.035
131. Gupta I, Adin SN, Rashid MA, Alhamhoom Y, Aqil M, Mujeeb M. Spanlastics as a potential approach for enhancing the nose-to-brain delivery of piperine: in vitro prospect and in vivo therapeutic efficacy for the management of epilepsy. Pharmaceutics. 2023;15(2):641. doi:10.3390/pharmaceutics15020641
132. Samaridou E, Walgrave H, Salta E, et al. Nose-to-brain delivery of enveloped RNA - cell permeating peptide nanocomplexes for the treatment of neurodegenerative diseases. Biomaterials. 2020;230:119657. doi:10.1016/j.biomaterials.2019.119657
133. Zhao C, Deng Y, He Y, Huang X, Wang C, Li W. Decreased level of exosomal miR-5121 released from microglia suppresses neurite outgrowth and synapse recovery of neurons following traumatic brain injury. Neurotherapeutics. 2021;18(2):1273–1294. doi:10.1007/s13311-020-00999-z
134. Kodali M, Madhu LN, Reger RL, et al. A single intranasal dose of human mesenchymal stem cell-derived extracellular vesicles after traumatic brain injury eases neurogenesis decline, synapse loss, and BDNF-ERK-CREB signaling. Front Mol Neurosci. 2023;16:1185883. doi:10.3389/fnmol.2023.1185883
135. Ye J, Sun X, Jiang Q, et al. Umbilical cord blood-derived exosomes attenuate dopaminergic neuron damage of Parkinson’s disease mouse model. J Nanobiotechnology. 2024;22(1):567. doi:10.1186/s12951-024-02773-1
136. Tian T, Liang R, Erel-Akbaba G, et al. Immune checkpoint inhibition in GBM primed with radiation by engineered extracellular vesicles. ACS Nano. 2022;16(2):1940–1953. doi:10.1021/acsnano.1c05505
137. Qian Y, Li X, Li G, et al. Astrocyte-derived exosomal miR-148a-3p suppresses neuroinflammation and restores neurological function in traumatic brain injury by regulating the microglial phenotype. eNeuro. 2024;11(2):ENEURO.0336–23.2024. doi:10.1523/ENEURO.0336-23.2024
138. Mavroudis I, Balmus IM, Ciobica A, Nicoara MN, Luca AC, Palade DO. The role of microglial exosomes and miR-124-3p in neuroinflammation and neuronal repair after traumatic brain injury. Life. 2023;13(9):1924. doi:10.3390/life13091924
139. Jiang S, Wang H, Yang C, et al. Phase 1 study of safety and preliminary efficacy of intranasal transplantation of human neural stem cells (ANGE-S003) in parkinson’s disease. J Neurol Neurosurg Psychiatry. 2024;95(12):1102–1111. doi:10.1136/jnnp-2023-332921
140. Santamaria G, Brandi E, Vitola P, et al. Intranasal delivery of mesenchymal stem cell secretome repairs the brain of Alzheimer’s mice. Cell Death Differ. 2021;28(1):203–218. doi:10.1038/s41418-020-0592-2
141. Jeon S, Park SH, Kim E, Kim JY, Kim SW, Choi H. A magnetically powered stem cell-based microrobot for minimally invasive stem cell delivery via the intranasal pathway in a mouse brain. Adv Healthc Mater. 2021;10(19):e2100801. doi:10.1002/adhm.202100801
142. Borlongan CV, Lee JY, D’Egidio F, de Kalbermatten M, Garitaonandia I, Guzman R. Nose-to-brain delivery of stem cells in stroke: the role of extracellular vesicles. Stem Cells Transl Med. 2024;13(11):1043–1052. doi:10.1093/stcltm/szae072
143. Gutova M, Cheng JP, Adhikarla V, et al. Intranasally administered l-myc-immortalized human neural stem cells migrate to primary and distal sites of damage after cortical impact and enhance spatial learning. Stem Cells Int. 2021;2021:5549381. doi:10.1155/2021/5549381
144. Jiang Y, Zhang P, Zhang X, Lv L, Zhou Y. Advances in mesenchymal stem cell transplantation for the treatment of osteoporosis. Cell Prolif. 2021;54(1):e12956. doi:10.1111/cpr.12956
145. Xie X, Song Q, Dai C, et al. Clinical safety and efficacy of allogenic human adipose mesenchymal stromal cells-derived exosomes in patients with mild to moderate alzheimer’s disease: a phase I/II clinical trial. Gen Psychiatr. 2023;36(5):e101143. doi:10.1136/gpsych-2023-101143
146. Mou YK, Song XY, Wang HR, et al. Understanding the nose-brain axis and its role in related diseases: a conceptual review. Neurobiol Dis. 2024;202:106690. doi:10.1016/j.nbd.2024.106690
147. Hashimoto K, Wei Y, Yang C. Special issue on “A focus on brain-body communication in understanding the neurobiology of diseases”. Neurobiol Dis. 2024;201:106666. doi:10.1016/j.nbd.2024.106666
148. Hosang L, Canals RC, van der Flier FJ, et al. The lung microbiome regulates brain autoimmunity. Nature. 2022;603(7899):138–144. doi:10.1038/s41586-022-04427-4
149. Jammal Salameh L, Bitzenhofer SH, Hanganu-Opatz IL, Dutschmann M, Egger V. Blood pressure pulsations modulate central neuronal activity via mechanosensitive ion channels. Science. 2024;383(6682):eadk8511. doi:10.1126/science.adk8511
150. Yin X, Zhang S, Lee JH, et al. Compartmentalized ocular lymphatic system mediates eye-brain immunity. Nature. 2024;628(8006):204–211. doi:10.1038/s41586-024-07130-8
151. Bin NR, Prescott SL, Horio N, Wang Y, Chiu IM, Liberles SD. An airway-to-brain sensory pathway mediates influenza-induced sickness. Nature. 2023;615(7953):660–667. doi:10.1038/s41586-023-05796-0
152. Xu K, Duan S, Wang W, et al. Nose-to-brain delivery of nanotherapeutics: transport mechanisms and applications. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2024;16(2):e1956. doi:10.1002/wnan.1956
153. Nigam K, Kaur A, Tyagi A, et al. In vitro & in vivo evaluations of PLGA nanoparticle based combinatorial drug therapy for baclofen and lamotrigine for neuropathic pain management. J Microencapsul. 2022;39(2):95–109. doi:10.1080/02652048.2022.2041751
154. Misra SK, Pathak K. Nose-to-brain targeting via nanoemulsion: significance and evidence. Colloids Interfaces. 2023;7(1):23. doi:10.3390/colloids7010023
155. Taha E, Shetta A, Nour SA, Naguib MJ, Mamdouh W. Versatile nanoparticulate systems as a prosperous platform for targeted nose-brain drug delivery. Mol Pharm. 2024;21(3):999–1014. doi:10.1021/acs.molpharmaceut.3c00588
156. Kamei N, Suwabe S, Arime K, et al. Investigation of the transport pathways associated with enhanced brain delivery of peptide drugs by intranasal coadministration with penetratin. Pharmaceutics. 2021;13(11):1745. doi:10.3390/pharmaceutics13111745
157. Monge-Fuentes V, Biolchi Mayer A, Lima MR, et al. Dopamine-loaded nanoparticle systems circumvent the blood-brain barrier restoring motor function in mouse model for Parkinson’s Disease. Sci Rep. 2021;11(1):15185. doi:10.1038/s41598-021-94175-8
158. Jahangard Y, Monfared H, Moradi A, Zare M, Mirnajafi-Zadeh J, Mowla SJ. Therapeutic effects of transplanted exosomes containing miR-29b to a rat model of alzheimer’s disease. Front Neurosci. 2020;14:564. doi:10.3389/fnins.2020.00564
159. Zhou X, Deng X, Liu M, et al. Intranasal delivery of BDNF-loaded small extracellular vesicles for cerebral ischemia therapy. J Control Release. 2023;357:1–19. doi:10.1016/j.jconrel.2023.03.033
160. Shah P, Dubey P, Vyas B, et al. Lamotrigine loaded PLGA nanoparticles intended for direct nose to brain delivery in epilepsy: pharmacokinetic, pharmacodynamic and scintigraphy study. Artif Cells Nanomed Biotechnol. 2021;49(1):511–522. doi:10.1080/21691401.2021.1939709
161. Koo J, Lim C, Oh KT. Recent advances in intranasal administration for brain-targeting delivery: a comprehensive review of lipid-based nanoparticles and stimuli-responsive gel formulations. Int J Nanomed. 2024;19:1767–1807. doi:10.2147/IJN.S439181
162. Shin S, Jo H, Agura T, et al. Use of surface-modified porous silicon nanoparticles to deliver temozolomide with enhanced pharmacokinetic and therapeutic efficacy for intracranial glioblastoma in mice. J Mater Chem B. 2024;12(37):9335–9344. doi:10.1039/D4TB00631C
163. Abbate E, Ragas AMJ, Caldeira C, et al. Operationalization of the safe and sustainable by design framework for chemicals and materials: challenges and proposed actions. Integr Environ Assess Manag. 2025;21(2):245–262. doi:10.1093/inteam/vjae031
164. Groenewold M, Bleeker EAJ, Noorlander CW, et al. Governance of advanced materials: shaping a safe and sustainable future. NanoImpact. 2024;35:100513. doi:10.1016/j.impact.2024.100513
165. Wang Y, Li C, Zhang X, et al. Exposure to PM2.5 aggravates parkinson’s disease via inhibition of autophagy and mitophagy pathway. Toxicology. 2021;456:152770. doi:10.1016/j.tox.2021.152770
166. Jo S, Kim YJ, Park KW, et al. Association of NO2 and other air pollution exposures with the risk of parkinson disease. JAMA Neurol. 2021;78(7):800–808. doi:10.1001/jamaneurol.2021.1335
167. Tateo F, Grassivaro F, Ermani M, Puthenparampil M, Gallo P. PM2.5 levels strongly associate with multiple sclerosis prevalence in the Province of Padua, Veneto Region, North-East Italy. Mult Scler. 2019;25(13):1719–1727. doi:10.1177/1352458518803273
168. Ponzano M, Schiavetti I, Bergamaschi R, et al. The impact of PM2.5, PM10 and NO2 on Covid-19 severity in a sample of patients with multiple sclerosis: a case-control study. Mult Scler Relat Disord. 2022;68:104243. doi:10.1016/j.msard.2022.104243
169. Zhang Z, Chen L, Wang X, et al. Associations of air pollution and genetic risk with incident dementia: a prospective cohort study. Am J Epidemiol. 2023;192(2):182–194. doi:10.1093/aje/kwac188
170. Piao CH, Fan Y, Nguyen TV, Song CH, Kim HT, Chai OH. PM2.5 exposure regulates Th1/Th2/Th17 cytokine production through NF-kappaB signaling in combined allergic rhinitis and asthma syndrome. Int Immunopharmacol. 2023;119:110254. doi:10.1016/j.intimp.2023.110254
171. Luo P, Ying J, Li J, et al. Air pollution and allergic rhinitis: findings from a prospective cohort study. Environ Sci Technol. 2023;57(42):15835–15845. doi:10.1021/acs.est.3c04527
172. Shah SA, Yoon GH, Ahmad A, Ullah F, Ul Amin F, Kim MO. Nanoscale-alumina induces oxidative stress and accelerates amyloid beta (Abeta) production in ICR female mice. Nanoscale. 2015;7(37):15225–15237. doi:10.1039/C5NR03598H
173. Liu H, Zhang W, Fang Y, et al. Neurotoxicity of aluminum oxide nanoparticles and their mechanistic role in dopaminergic neuron injury involving p53-related pathways. J Hazard Mater. 2020;392:122312. doi:10.1016/j.jhazmat.2020.122312
174. Guo Z, Zhang P, Luo Y, et al. Intranasal exposure to ZnO nanoparticles induces alterations in cholinergic neurotransmission in rat brain. Nano Today. 2020;35:100977. doi:10.1016/j.nantod.2020.100977
175. Wu J, Wang C, Sun J, Xue Y. Neurotoxicity of silica nanoparticles: brain localization and dopaminergic neurons damage pathways. ACS Nano. 2011;5(6):4476–4489. doi:10.1021/nn103530b
176. Rahmati M, Yon DK, Lee SW, et al. New-onset neurodegenerative diseases as long-term sequelae of SARS-CoV-2 infection: a systematic review and meta-analysis. J Med Virol. 2023;95(7):e28909. doi:10.1002/jmv.28909
177. van de Beek D, Brouwer M, Hasbun R, Koedel U, Whitney CG, Wijdicks E. Community-acquired bacterial meningitis. Nat Rev Dis Primers. 2016;2:16074. doi:10.1038/nrdp.2016.74
178. Xie J, Cools L, Van Imschoot G, et al. Helicobacter pylori-derived outer membrane vesicles contribute to Alzheimer’s disease pathogenesis via C3-C3aR signalling. J Extracell Vesicles. 2023;12(2):e12306. doi:10.1002/jev2.12306
179. Piret J, Boivin G. Immunomodulatory strategies in herpes simplex virus encephalitis. Clin Microbiol Rev. 2020;33(2). doi:10.1128/CMR.00105-19
180. Ciprandi G, De Amici M, Tosca M, Fuchs D. Tryptophan metabolism in allergic rhinitis: the effect of pollen allergen exposure. Hum Immunol. 2010;71(9):911–915. doi:10.1016/j.humimm.2010.05.017
181. Phuna ZX, Madhavan P. A closer look at the mycobiome in Alzheimer’s disease: fungal species, pathogenesis and transmission. Eur J Neurosci. 2022;55(5):1291–1321. doi:10.1111/ejn.15599
182. Sahu B, Nookala S, Floden AM, et al. House dust mite-induced asthma exacerbates Alzheimer’s disease changes in the brain of the App(NL-G-F) mouse model of disease. Brain Behav Immun. 2024;121:365–383. doi:10.1016/j.bbi.2024.07.038
183. Callebaut I, Steelant B, Backaert W, et al. Brain activation after nasal histamine provocation in house dust mite allergic rhinitis patients. Allergy. 2021;76(6):1879–1882. doi:10.1111/all.14677
184. Feng Q, Zhang X, Zhao X, et al. Intranasal delivery of pure nanodrug loaded liposomes for alzheimer’s disease treatment by efficiently regulating microglial polarization. Small. 2024;20(50):e2405781. doi:10.1002/smll.202405781
185. Garcia-Pardo J, Novio F, Nador F, et al. Bioinspired theranostic coordination polymer nanoparticles for intranasal dopamine replacement in parkinson’s disease. ACS Nano. 2021;15(5):8592–8609. doi:10.1021/acsnano.1c00453
186. Xu K, Huang P, Wu Y, et al. Engineered selenium/human serum albumin nanoparticles for efficient targeted treatment of parkinson’s disease via oral gavage. ACS Nano. 2023;17(20):19961–19980. doi:10.1021/acsnano.3c05011
187. Guler E, Yekeler HB, Parviz G, et al. Vitamin B(12)-loaded chitosan-based nanoparticle-embedded polymeric nanofibers for sublingual and transdermal applications: two alternative application routes for vitamin B(12). Int J Biol Macromol. 2024;258(Pt 2):128635. doi:10.1016/j.ijbiomac.2023.128635
188. Mu D, Zhou L, Shi L, et al. Quercetin-crosslinked chitosan nanoparticles: a potential treatment for allergic rhinitis. Sci Rep. 2024;14(1):4021. doi:10.1038/s41598-024-54501-2
189. Danz K, Fleddermann J, Koch M, et al. Evaluation of the transport and binding of dopamine-loaded PLGA nanoparticles for the treatment of parkinson’s disease using in vitro model systems. Pharmaceutics. 2024;16(5):571. doi:10.3390/pharmaceutics16050571
190. Zheng Q, Liu H, Zhang H, et al. Ameliorating mitochondrial dysfunction of neurons by biomimetic targeting nanoparticles mediated mitochondrial biogenesis to boost the therapy of parkinson’s disease. Adv Sci. 2023;10(22):e2300758. doi:10.1002/advs.202300758
191. Etxebeste-Mitxeltorena M, Niza E, Fajardo CM, et al. Neuroprotective properties of exosomes and chitosan nanoparticles of Tomafran, a bioengineered tomato enriched in crocins. Nat Prod Bioprospect. 2024;14(1):9. doi:10.1007/s13659-023-00425-9
192. Zhang L, Lin Y, Bai W, Sun L, Tian M. Human umbilical cord mesenchymal stem cell-derived exosome suppresses programmed cell death in traumatic brain injury via PINK1/Parkin-mediated mitophagy. CNS Neurosci Ther. 2023;29(8):2236–2258. doi:10.1111/cns.14159
193. Rehni AK, Cho S, Quero HN, et al. Red blood cell microparticles limit hematoma growth in intracerebral hemorrhage. Stroke. 2022;53(10):3182–3191. doi:10.1161/STROKEAHA.122.039641
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.