Back to Journals » International Journal of Nanomedicine » Volume 20
Mesenchymal Stem Cell-Derived Extracellular Vesicles in Alzheimer’s Disease: A Novel Cell-Free Therapeutic Strategy and Diagnostic Biomarker
Authors Wang X ![]() , Yang F, Chen P, Yang M, Deng Y, Zhan Z
, Yang F, Chen P, Yang M, Deng Y, Zhan Z
Received 31 July 2025
Accepted for publication 23 November 2025
Published 3 December 2025 Volume 2025:20 Pages 14375—14391
DOI https://doi.org/10.2147/IJN.S556625
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Xing Zhang
Xiaoling Wang,1,2 Fulan Yang,3 Puwen Chen,1 Mei Yang,1 Yuxin Deng,4 Zhao Zhan5
1The First School of Clinical Medicine, Gannan Medical University, Ganzhou, 341000, People’s Republic of China; 2College of Medical Technology, Gannan Medical University, Ganzhou, 341000, People’s Republic of China; 3Department of Breast Surgery, Ganzhou People’s Hospital, Ganzhou, 341000, People’s Republic of China; 4Center for Clinical Laboratories, The Affiliated Hospital of Guizhou Medical University, Guiyang, 550001, People’s Republic of China; 5Department of Clinical Laboratory, The First Affiliated Hospital of Yangtze University, Jingzhou, 434000, People’s Republic of China
Correspondence: Yuxin Deng, Email [email protected] Zhao Zhan, Email [email protected]
Abstract: With the ongoing trend of population aging worldwide, the incidence of Alzheimer’s disease (AD) is steadily increasing. In the absence of effective therapeutic options for atypical forms of AD, reducing its prevalence and improving treatment outcomes have become pressing priorities. Mesenchymal stem cell-derived extracellular vesicles (MSC-EVs) have attracted growing attention as a new cell-free therapeutic approach for AD due to their high stability, low immunogenicity, and minimal tumorigenic risk. This review provides a comprehensive overview of the pathological mechanisms underlying AD, highlights the diagnostic potential of MSC-EVs, and elaborates on their therapeutic advantages and mechanisms of action. Furthermore, it addresses the key challenges and considerations associated with the clinical translation of MSC-EVs.
Keywords: extracellular vesicles, Alzheimer’s disease, mesenchymal stem cell, β-amyloid protein
Introduction
Alzheimer’s disease (AD) is the most common cause of dementia in older adults, accounting for 60% to 80% of all dementia cases.1 In 2024, an estimated 6.9 million individuals aged 65 and older in the United States were living with AD. Worse still, the prevalence of AD continues to rise, and this number is expected to reach 13.8 million by 2060.2 AD places a substantial burden on patients and their families, both economically and physically.
The pathogenesis of AD is primarily associated with the abnormal accumulation of β-amyloid (Aβ) plaques3 and the aggregation of hyperphosphorylated tau protein in neurofibrillary tangles (NFTs).4 The symptom of AD varies depending on the specific brain regions affected. The most common manifestation is an insidious onset of progressively worsening dementia, typically characterized by episodic memory impairment, which accounts for the majority of cases. In contrast, some patients present with atypical clinical phenotypes, often with an earlier age of onset, and predominantly exhibit deficits in visuospatial processing, language, or motor function. These atypical cases are frequently diagnosed at later stages, leading to missed opportunities for timely intervention.5,6 At present, the treatment of AD relies primarily on pharmacological interventions, and the most commonly prescribed agents are acetylcholinesterase inhibitors. However, current therapies are only capable of temporarily slowing disease progression, and their efficacy is limited by the timing of intervention.7 As a result, caring for AD patients imposes significant economic and caregiving burdens. Therefore, there is an urgent need to develop novel therapeutic strategies that can more effectively modify disease progression and improve clinical outcomes.
In recent years, mesenchymal stem cells (MSCs) have attracted significant interest as potential cell-based therapeutic agents due to their robust regenerative capacity. MSCs have the ability to home to sites of tissue injury and facilitate neural repair and angiogenesis through the secretion of neurotrophic factors and angiogenic modulators.8 MSCs have been widely investigated and applied in various clinical contexts, exhibiting immunomodulatory,9 antifibrotic,10 and regenerative properties11 in diseases such as atopic dermatitis, organ transplantation, and diabetic nephropathy.12–14 Increasing evidence, however, suggests that the therapeutic benefits of MSCs are largely mediated by their paracrine release of extracellular vesicles (EVs).15,16
EVs are small lipid bilayer vesicles ranging from 30 to 2000 nanometers in diameter, secreted by a variety of cell types, including stem cells, immune cells, and tumor cells.17–19 EVs were first isolated from sheep reticulocytes by Pan20 and Johnstone,21 who coined the term “exosomes”. However, due to current technical limitations in separating EV subtypes originating from distinct biogenetic pathways, most isolation methods are unable to distinguish between them reliably. In response, the International Society for Extracellular Vesicles (ISEV) issued updated guidelines in 2023, recommending against the routine use of biogenesis-based terms such as exosomes and microvesicles. Instead, ISEV advocates for the use of the umbrella term “EVs”, along with operational extensions where appropriate. ISEV further encourages the classification of EV subtypes based on physical characteristics (eg, size, density) or cellular origin. According to commonly accepted size-based criteria, EVs with a diameter smaller than 200 nm are referred to as small extracellular vesicles (sEVs), while those larger than 200 nm are known as large extracellular vesicles (LEVs). Vesicle mimetics produced under experimental conditions involving physical cell disruption (eg, extrusion) are termed artificial cell-derived vesicles (ACDVs). EV mimetics synthesized de novo from molecular components or engineered as hybrid constructs are referred to as synthetic vesicles (SVs).22 In this review, the terms sEVs and LEVs are used according to the original studies’ reported vesicle sizes, while ACDVs and SVs are used based on the method of synthesis. The term EVs is used in contexts where definitions are being discussed or where the original source did not specify vesicle size or origin. EVs are known to play important roles in various diseases by mediating intercellular communication, delivering proteins and nucleic acids, and modulating immune and inflammatory responses.23 To date, most EV studies have focused on vesicles of small sizes.
Techniques for EV Isolation
Currently, numerous techniques exist for EV isolation. The initial guidelines issued by ISEV in 2014 emphasized that no consensus “gold standard” for EV isolation and purification has been established.24 Commonly used methods include differential ultracentrifugation (dUC), density gradient centrifugation, immunoaffinity capture, precipitation, and size-exclusion chromatography (SEC). Subsequent guideline updates issued by ISEV continue to assert that no single method is universally optimal; each has distinct advantages and limitations, and researchers should select the most appropriate approach based on their downstream applications and scientific objectives.22,25 (i) dUC is the most widely used method for the isolation and concentration of EVs across diverse research settings. This technique operates by subjecting EV-containing fluids to one or more rounds of low-speed centrifugation to remove cells and tissue debris, followed by high-speed centrifugation with precisely adjusted speed and duration based on sedimentation coefficients to recover EVs of varying diameters. This approach is cost-effective but time-consuming, and the EV populations isolated at different centrifugation speeds often exhibit overlapping sizes and content.26 (ii) Density gradient centrifugation is a more stringent ultracentrifugation method that employs a gradient formed from media and aqueous buffers with progressively decreasing densities from bottom to top. The EV-containing sample is loaded either at the bottom or the top of the gradient and then subjected to ultracentrifugation to separate distinct EV subpopulations. This technique yields EVs with high specificity and purity; however, it is labor-intensive and typically associated with low recovery efficiency.27 (iii) SEC entails applying EV-containing samples to chromatographic columns packed with matrices of defined pore size, where larger particles are excluded and elute first, while smaller particles penetrate the matrix and elute later. SEC is rapid and highly specific. However, commercial columns are costly, and samples may contain protein contaminants.28 (iv) Immunoaffinity capture isolates EVs based on surface markers or charge, providing rapid and highly selective isolation but at high reagent cost and with potential for non-specific binding. (v) Precipitation methods reduce EV solubility using polymers such as polyethylene glycol, inducing vesicle precipitation followed by low-speed centrifugation. However, the latest ISEV guidelines strongly discourage these kits due to chemical contamination and poor sample purity.29 Table 1 summarizes the principles, advantages, and limitations of these methods to guide researchers in selecting the most suitable EV isolation technique for their studies.
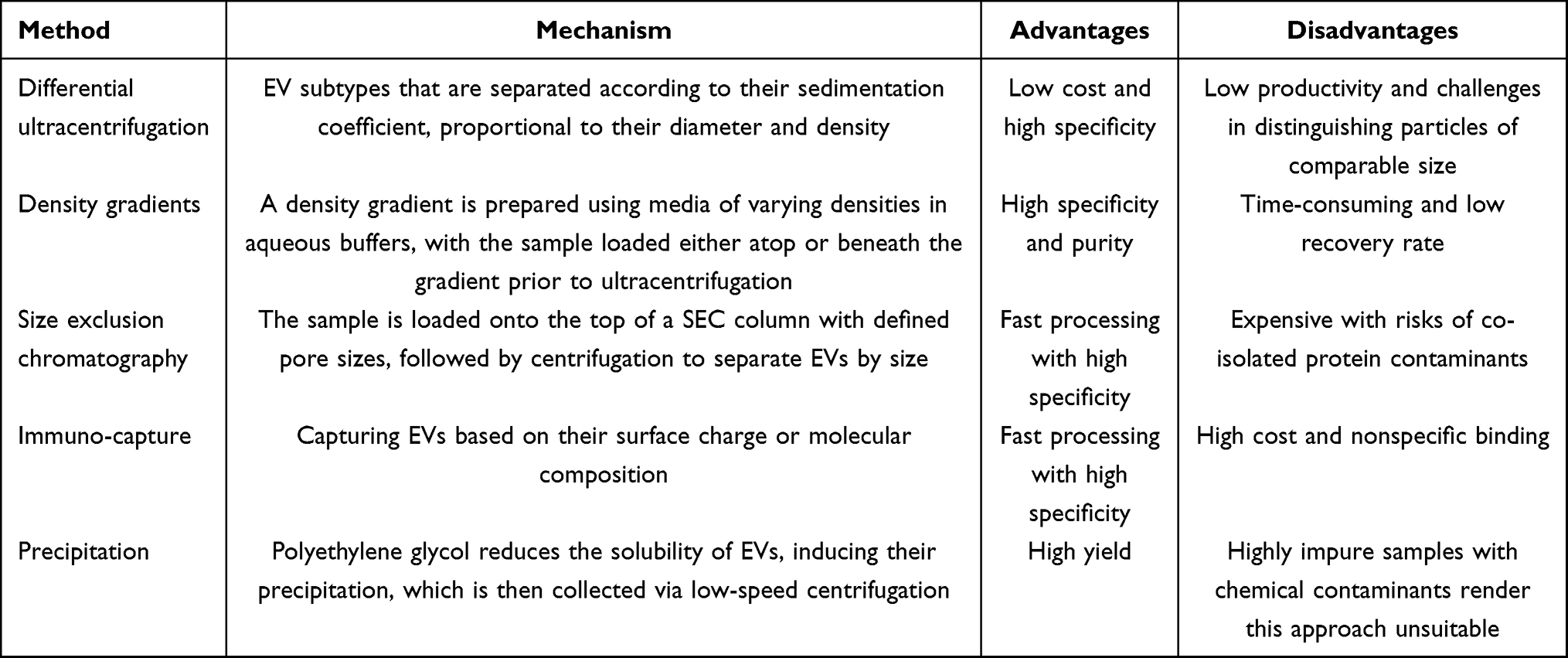 |
Table 1 Isolation Methods for Extracellular Vesicles |
Pathogenic Mechanisms of AD
Despite being recognized for decades and exhibiting a steadily increasing prevalence, the pathogenic mechanisms of AD remain to be fully revealed. The leading hypotheses center around the amyloid cascade hypothesis and the formation of NFTs resulting from hyperphosphorylated tau protein. Additional contributing factors include synaptic dysfunction, neuroinflammation, mitochondrial impairment, and cholinergic deficits.30,31 In this section, we provide a detailed description of these mechanisms and summarize them in Figure 1, aiming to enhance our understanding of AD pathogenesis and to inform novel strategies for its diagnosis and treatment.
 |
Figure 1 Pathogenic mechanisms of AD. The pathogenesis of Alzheimer’s disease mainly includes synaptic dysfunction, abnormal deposition of Aβ, hyperphosphorylation of Tau, neuroinflammation, and mitochondrial dysfunction. (This figure was created with Biorender.com.). |
Amyloid Cascade Hypothesis
Aβ is a peptide composed of 39 to 43 amino acids, predominantly generated from the amyloid precursor protein (APP) through sequential proteolytic cleavage by α-secretase (ADAM10), β-secretase (BACE1), and γ-secretase.32 Aβ is a critical pathological protein in AD, whose abnormal aggregation forms extracellular amyloid plaques deposited across various brain regions. These plaques exacerbate neurotoxicity by activating microglia to release pro-inflammatory cytokines. Aβ exhibits neurotoxicity, particularly toward neurons, inducing lipid dysregulation and disruption of calcium homeostasis, ultimately leading to neuronal injury and apoptosis.7,33,34 APP undergoes two distinct processing pathways, with Aβ production arising exclusively from the abnormal pathway: (a) The normal metabolic pathway is the non-amyloidogenic pathway, where APP is sequentially cleaved by α-secretase and γ-secretase, releasing soluble APPα (sAPPα), P3 peptides, and the APP intracellular domain (AICD) without generating Aβ; (b) The abnormal metabolic pathway is the amyloidogenic pathway, in which APP is sequentially cleaved by β-secretase (BACE1) and γ-secretase, producing soluble APPβ (sAPPβ), AICD, and Aβ peptides (typically Aβ40 and Aβ42, and the latter is more prone to aggregation).35 The dysregulation of APP processing via the amyloidogenic pathway is a key driver in the pathogenesis of AD. In summary, Aβ derived from APP has several pathological effects: (i) oligomeric Aβ aggregates are highly neurotoxic, causing severe synaptic dysfunction;36 (ii) Aβ activates microglia to release pro-inflammatory cytokines, such as interleukin-1β (IL-1β) and tumor necrosis factor-alpha (TNF-α), thus exacerbating neuronal injury;37 and (iii) Aβ may indirectly promote tau hyperphosphorylation and lead to NFTs.38 Currently, therapeutics targeting Aβ are being developed, which brings hope for innovative treatments of AD, with several candidates progressing through clinical trials.
Tau Hyperphosphorylation and NFTs
Tau is a microtubule-associated protein primarily localized in neurons, where it functions in stabilizing microtubules, preserving cytoskeletal integrity, and facilitating axonal transport.39,40 In the pathogenesis of AD, the hyperphosphorylation of Tau and the subsequent formation of NFTs represent one of the key pathological hallmarks. Under normal physiological conditions, the microtubule-binding domains of Tau interact with tubulin to promote microtubule polymerization, thereby maintaining the stability of the axonal transport system. Importantly, the activity of Tau protein is modulated by phosphorylation, and excessive phosphorylation disrupts its normal physiological function.41 In the pathogenesis of AD, multiple aberrant kinases, such as glycogen synthase kinase-3β (GSK-3β) and cyclin-dependent kinase 5 (CDK5), contribute to Tau hyperphosphorylation, thereby resulting in the loss of its microtubule-stabilizing function.42 Hyperphosphorylated Tau tends to self-aggregate, forming NFTs that accumulate in the cytoplasm, ultimately contributing to neuronal damage and cell death.43 Furthermore, some studies suggest that Aβ plaques may act as upstream triggers of Tau pathology by promoting Tau phosphorylation (resulting in p-Tau) and facilitating its propagation across brain regions.44
Synaptic Dysfunction and Neurotransmitter Imbalance
The cholinergic system plays a vital role in cognitive function. Acetylcholine, a key neurotransmitter in the human brain, is widely distributed across the cerebral cortex, basal ganglia, and forebrain. It facilitates synaptic transmission by enabling communication between neurons. In AD, degeneration of the nucleus basalis results in the loss of cholinergic signaling, particularly affecting the cholinergic neurons and axonal projections to the cerebral cortex. Aβ oligomers can bind to postsynaptic N-methyl-D-aspartate (NMDA) receptors, triggering calcium influx and oxidative stress, which collectively impair synaptic plasticity.45 Furthermore, hyperphosphorylated Tau accumulates within neurons to form NFTs, disrupting the axonal transport of presynaptic vesicles and ultimately impairing neurotransmitter release.46
Neuroinflammation
Neuroinflammation plays a critical role in the pathogenesis of AD. Aβ deposits activate microglia and astrocytes, which subsequently trigger inflammatory responses via activation of Toll-like receptors (TLR2, TLR4, and TLR6) and inflammasomes belonging to the intracellular nucleotide-binding oligomerization domain (NOD)-like receptor (NLR) family.47,48 Microglia recruited in association with Aβ are termed disease-associated microglia (DAM), exhibiting a dual role in AD progression.49 Under physiological conditions, microglia contribute to homeostasis by phagocytosing Aβ and clearing cellular debris.50 However, chronically activated DAM releases pro-inflammatory cytokines, such as TNF-α, IL-1β, and interferon-gamma (IFN-γ), which can enhance APP processing, thus leading to increased production of Aβ. Importantly, in response to these pro-inflammatory cytokines, microglia upregulate inducible nitric oxide synthase (iNOS), resulting in elevated nitric oxide (NO) synthesis. This heightened NO production facilitates peptide aggregation, impairs synaptic plasticity, and contributes to synaptic dysfunction.51 Furthermore, studies have shown that the triggering receptor expressed on myeloid cells 2 (TREM2), which is expressed on myeloid-lineage cells, is essential for microglia to fully develop DAM functions. Loss of the TREM2 compromises microglial clearance of Aβ and has been shown to increase the risk of AD in humans.52
Mitochondrial Dysfunction
Mitochondrial dysfunction is not only involved in the pathogenesis of AD but also accelerates its progression. Research has demonstrated that glucose metabolism is significantly reduced in the brains of AD patients, resulting in decreased mitochondrial ATP synthesis. This deficit leads to an insufficient energy supply for neurons and impairs synaptic function.53,54 Moreover, mitochondria serve as the primary source of reactive oxygen species (ROS). Dysfunction of the mitochondrial electron transport chain (ECT) in AD leads to excessive ROS accumulation, which damages lipids, proteins, and DNA, thereby exacerbating neuronal injury.55 In addition, mitochondria are crucial for intracellular calcium homeostasis, and impaired mitochondrial calcium buffering capacity in AD patients causes calcium overload, ultimately activating apoptotic signaling pathways.56
The Role of EVs in AD
The Potential of EVs as Biomarkers for AD
EVs are present in various biological fluids, a property that makes them ideal candidates for liquid biopsy in disease diagnostics. Currently, the diagnosis of AD primarily relies on amyloid positron emission tomography (PET) scan and the cerebrospinal fluid (CSF) concentrations of Aβ and tau proteins.5 EVs isolated from plasma, serum, urine, saliva, and CSF have demonstrated diagnostic potential for AD. Among these, plasma- and serum-derived EVs offer the advantages of minimal invasiveness, high sensitivity, and specificity, making them particularly suitable as diagnostic biomarkers.57 Moreover, saliva and urine are more easily accessible and contain fewer high-abundance interfering molecules compared to plasma and serum, rendering them more applicable to clinical settings, especially for large-scale preclinical screening or routine monitoring of disease progression.58 EVs have also been isolated from CSF and exhibit high disease specificity. As carriers of diverse genetic and protein cargoes, including microRNAs and proteins, EVs represent a rich source of potential biomarkers. Table 2 summarizes EV-associated miRNAs and proteins that have shown potential as diagnostic biomarkers for AD in recent studies.
 |
Table 2 miRNAs and Proteins in EVs as Biomarkers for AD |
The Potential of EV-Derived miRNAs as Biomarkers for AD
MicroRNAs (miRNAs) are short (19–25 nucleotides), single-stranded, non-coding RNAs. They are not only emerging as therapeutic targets but also hold promise as diagnostic biomarkers.69 Due to their encapsulation within EVs, miRNAs are protected from enzymatic degradation, rendering them significantly more stable This property has increasingly attracted attention to EVs as a source of diagnostic biomarkers.70 In AD, studies investigating EV-associated miRNAs have primarily been cross-sectional and focused on samples from plasma, serum, and CSF. Serpente et al identified four differentially expressed miRNAs in neuron-derived small EVs (NDEVs) from plasma—miR-23a-3p, miR-223-3p, and miR-190a-5p were significantly upregulated, while miR-100-3p was downregulated—indicating their potential as AD biomarkers.59 Similarly, Kumar et al reported that miR-106b-5p was significantly overexpressed in plasma-derived sEVs from AD patients and demonstrated high diagnostic accuracy in distinguishing between AD and mild cognitive impairment (MCI).60 In Visconte et al, miR-106a-5p, miR-16-5p, miR-223-3p, miR-25-3p, miR-30b-5p, miR-92a-3p, and miR-451a were found to be significantly upregulated in plasma-derived sEVs from individuals with AD. Notably, in prodromal AD, miR-16-5p, miR-25-3p, miR-92a-3p, and miR-451a exhibited characteristic upregulation, indicating their potential as promising biomarkers for early-stage diagnosis.61 In addition, Reho et al identified five downregulated miRNAs, including miR-6780b-5p, miR-27b-3p, miR-15b-5p, miR-3940-5p, and miR-29c-3p, in serum-derived NDEVs, which could serve as candidate biomarkers for preclinical AD detection.62 Interestingly, Ryu et al investigated the association between salivary EVs and AD and reported a significantly upregulated miR-485-3p in EVs isolated from the saliva of AD patients, which positively correlated with Aβ deposition. This method is non-invasive and cost-effective, making it highly suitable for early detection of AD and for routine monitoring of treatment response.63 Moreover, EVs from other bodily fluids have also demonstrated diagnostic value. For instance, Kumar et al found that miR-455-3p was significantly upregulated in CSF-derived sEVs from AD patients compared to healthy controls. The upregulated expression of miR-455-3p in the AD brain highlights its strong potential as a clinical diagnostic biomarker.64
The Potential of EV Proteins as Biomarkers for AD
Besides miRNAs, proteins carried by EVs also exhibit significant potential as biomarkers. Muraoka et al found that tau phosphorylated at serine 396 (pS396 tau) and Amyloid-β (1–42) peptide (Aβ1-42) were significantly upregulated in sEVs isolated from the brain tissue of AD patients compared to controls. Additionally, annexin A5 (ANXA5) was significantly increased. Together with pS396 tau and Aβ1-42, ANXA5 may serve as promising biomarker candidates in biofluid samples from AD patients.65 Jia et al confirmed a strong correlation between proteins in plasma-derived sEVs and those in CSF, demonstrating that growth-associated protein 43 (GAP43), synaptosomal-associated protein 25 (SNAP25), and synaptotagmin-1 in plasma sEVs exhibited diagnostic accuracy comparable to CSF biomarkers for distinguishing AD or amnestic mild cognitive impairment (aMCI) patients from controls, with slightly improved discrimination between AD and MCI cases.66 Moreover, multiple candidate proteins for AD diagnosis have been identified in blood-derived EVs. Upregulated proteins include Ig-like domain-containing protein (A0A0G2JRQ6), complement C1q subcomponent subunit C (C1QC), complement component C9, platelet glycoprotein Ib beta chain (GP1BB), and Ras suppressor protein 1 (RSU1), while ADA10 (disintegrin and metalloproteinase domain 10) is among the downregulated proteins.67 Elevated C1q in CSF-derived EVs may be linked to early cognitive impairment and could contribute to immune regulation in the pathogenesis of AD.68
EVs as Drug Delivery Systems
EVs are considered promising drug delivery vehicles for various diseases. They have been investigated in multiple contexts, eg, using genetic engineering to encapsulate the photosensitizer Chlorin e6 and the immunoadjuvant R848 into sEVs to modulate the tumor microenvironment in prostate cancer.71 In clinical applications, many drugs reach only a small fraction of the target lesion to exert therapeutic effects and may cause increased toxicity and adverse side effects.72 As natural delivery vehicles, EVs exhibit excellent biocompatibility and the ability for intercellular communication. They are capable of delivering diverse therapeutic agents, including DNA, RNA, and proteins, and can cross the blood-brain barrier (BBB), thus making them particularly well-suited for AD. Zhu et al demonstrated that sEVs dual-functionalized with Angiopep-2 and trans-activator of transcription peptides can penetrate the BBB and directly reach glioma lesions within tumors.73 Based on these properties, EVs are capable of efficiently crossing BBB, thereby enhancing intracerebral drug concentrations. Compared to conventional therapeutic delivery strategies for AD, EV-based delivery can mitigate complications such as intracranial infection and drug-induced toxicity. Moreover, the lipid bilayer of EVs enhances target affinity and improves delivery efficiency. Most critically, EVs provide protection for therapeutic cargo from enzymatic degradation.74,75 Corynoxine-B (Cory-B), a natural autophagy inducer, has been shown to reduce Aβ and APP by promoting autophagic activity in neuronal cells. However, due to its poor BBB permeability, Cory-B exhibits limited bioavailability. To address this limitation, Iyaswamy et al utilized engineered neuron-derived sEVs as carriers. These sEVs demonstrated a robust ability to cross the BBB, facilitating targeted delivery of Cory-B into the brain of AD model mice. Consequently, the treatment significantly improved cognitive function by specifically acting on neurons overexpressing APP and Aβ.76
EVs Participate in the Clearance of AD Pathogenic Proteins
EVs are involved in the clearance of pathogenic proteins. Aβ and tau are classical pathogenic proteins in AD and are considered key contributors to cognitive decline.77 Research has shown that sEVs derived from neural stem cells (NSCs) can promote the non-amyloidogenic processing of APP by enhancing the activity of α-secretase while simultaneously suppressing that of β- and γ-secretase, thereby significantly reducing Aβ production. In addition, NSC-derived sEVs have been found to attenuate neuroinflammation.78 Peng et al used electroporation to load miR-206-3p into EVs derived from mesenchymal stem cells (MSCs) in rats. These engineered MSC-EVs were delivered intranasally (a non-invasive administration route) to AD-modeled mice, leading to a reduction in Aβ deposition, enhanced neurogenesis, and preservation of synaptic plasticity.79 Notably, while EVs can aid in the clearance of pathogenic proteins, they may also contribute to the propagation of such proteins within the brain.80
Beyond their primary functions, EVs can also serve as imaging agents to monitor therapeutic progress. By labeling EVs with techniques like electroporation or direct incubation, researchers can track their distribution in vivo using imaging modalities such as magnetic resonance and nuclear imaging. Thereby, EVs function as effective, non-invasive tracking tools that are crucial for optimizing treatments and diagnostics.81,82 This is particularly revealing for central nervous system disorders, where this method illustrates how EVs operate as drug carriers—including their uptake, clearance, and specific organ targeting.83
Therapeutic Potential of MSCs
MSCs are multipotent stem cells with self-renewal capacity and multilineage differentiation, and they are widely used in clinical trials.84 MSCs can be isolated from various tissues, including umbilical cord, bone marrow, adipose tissue, and muscle.85 They exhibit numerous therapeutic properties, such as anti-inflammatory effects, immunomodulation, tissue regeneration, and antifibrotic activity.86,87 MSCs exert their functions through autocrine, paracrine, and endocrine signaling by secreting a broad range of chemokines, cytokines, growth factors, anti-inflammatory mediators, and metabolites, thereby contributing to angiogenesis, immune regulation, and cell recruitment.88,89 To date, MSCs have demonstrated important therapeutic efficacy across various diseases. Transplantation of MSCs holds significant potential due to their immunomodulatory capacity and ability to promote tissue repair. Neuroinflammation frequently occurs during the pathological progression of AD, and MSCs can modulate the activity of astrocytes and microglia either directly or indirectly. They regulate the balance between pro- and anti-inflammatory cytokines via multiple signaling pathways involving nuclear transcription factors (eg, NFs), thus producing significant therapeutic effects in AD.90 For instance, after intrathecal injection of autologous adipose-derived MSCs, most patients exhibit significant improvement in their American Spinal Injury Association Impairment Scale (AIS) score without experiencing severe adverse events.91 Beyond neurological applications, MSCs have also been employed in the treatment of endocrine and metabolic disorders, cardiac repair, and autoimmune diseases.89,92–94 However, challenges such as immune rejection, uncontrolled proliferation after transplantation, and limited differentiation capacity remain in stem cell-based therapies. These limitations have shifted attention toward MSC-derived EVs (MSC-EVs), which are more stable and exhibit similar therapeutic functions to MSCs.95
Therapeutic Advantages of MSC-EVs Over MSC-Based Cell Therapy
EVs secreted by MSCs represent a promising alternative to conventional MSC-based cell therapy. MSC-EVs retain the therapeutic functions of their parental MSCs and serve as critical mediators of intercellular communication.96 These vesicles are enriched with bioactive proteins, miRNAs, and functional lipids that reflect the physiological properties of the source cells.97,98 Compared with cell-based therapies, MSC-EVs offer several distinct advantages, which are summarized in Table 3:
 |
Table 3 Comparison of MSCs-EVs and MSCs Cell Therapy in Disease Treatment |
- Safety. Although MSCs have low immunogenicity, there is still a risk of immune rejection following transplantation. Additionally, MSC transplantation may lead to abnormal differentiation, potentially resulting in tumorigenesis or the formation of ectopic tissues.99,100 It is also noteworthy that intravenous administration of MSCs may cause pulmonary embolism.101,102 Compared with MSC-based cell therapy, MSC-EVs are cell-free, exhibit lower immunogenicity, and avoid the risks of unwanted differentiation associated with the transplantation of live cells.103,104
- Targeting capability and diverse routes of administration. MSC-EVs are substantially smaller than MSCs, allowing for more efficient trafficking to target tissues. Most notably, MSC-EVs are capable of crossing the BBB via transcytosis and exerting direct effects within the central nervous system, offering therapeutic potential for neurodegenerative disorders and diseases involving neuroinflammation.105 Clinically, MSC-EVs allow for precise dose control and can be administered through various routes, including intravenous injection, local delivery, or aerosol inhalation.106–108 In contrast, MSC-based cell therapies require careful consideration of delivery methods, eg, local injection may cause tissue injury, while intravenous administration risks pulmonary entrapment.109 However, MSCs-EV-based therapies are limited by the half-life of vesicular cargo and their intrinsic targeting efficiency, and they cannot fully replicate the functional properties of MSCs, such as differentiation and direct cellular replacement of damaged tissues.110
- Cost and ethical considerations. MSC-EVs offer several advantages, including ease of storage and transportation, high stability, scalability for large-scale production, lower manufacturing costs, and the absence of ethical controversies.111 In contrast, MSC-based therapies require rigorous cell culture conditions, and donor heterogeneity may influence therapeutic efficacy.112,113 Moreover, the cultivation of MSCs is expensive, and sourcing from embryonic stem cells raises ethical concerns.114 Nevertheless, therapies based on MSC-EVs still face challenges, such as complex isolation and purification processes and difficulties in achieving standardization.115,116
Therapeutic Applications of MSC-EVs in AD
MSC-EVs exhibit a strong capacity to traverse the blood-brain barrier (BBB), which makes them promising for AD.117 A growing body of research has demonstrated that MSC-EVs can facilitate Aβ clearance, modulate tau protein hyperphosphorylation, regulate neuroinflammatory responses, and ameliorate synaptic and cognitive dysfunction. These findings support the potential of MSC-EVs as a therapeutic strategy for AD (see Figure 2).118
 |
Figure 2 Therapeutic applications of MSC-EVs in AD. MSC-EVs can facilitate Aβ clearance, modulate tau protein hyperphosphorylation, regulate neuroinflammatory responses, and ameliorate synaptic and cognitive dysfunction. (This figure was created with Biorender.com.). |
MSC-EVs Facilitate Aβ Degradation
In the healthy brain, the production and degradation of Aβ are balanced. However, abnormal metabolism disrupts this balance. When the clearance capacity of lysosomes or glial cells is overwhelmed, pathogenic proteins are released into the extracellular space and propagate across various brain regions. The mechanisms underlying the production of Aβ, involving APP and BACE1, have been detailed in the previous section that covers the pathogenesis of AD. Key enzymes responsible for Aβ degradation include insulin-degrading enzyme (IDE) and neprilysin (NEP).119 IDE is a protease that binds to and degrades substrates in tissue extracts and also degrades Aβ, representing a critical link between diabetes and AD.120 Dysregulated expression of these enzymes leads to pathological accumulation of Aβ. NEP, also known as neutral endopeptidase, is a membrane-associated zinc metallopeptidase. Genetic deficiency of NEP results in impaired degradation of both exogenous and endogenous Aβ, causing abnormal accumulation of the toxic Aβ42 isoform in the brain.121 Numerous studies have confirmed that promoting Aβ clearance is a viable therapeutic strategy for AD. The pathogenesis of AD involves the PI3K/Akt/mTOR signaling pathway, autophagy dysfunction, and neuroinflammation. Ebrahim et al reported upregulated expression of BACE1, presenilin-1 (PS-1), and APP in the brains of AD-modeled rats, correlating closely with increased production of Aβ alongside a reduction in IDE expression. Treatment with MSCs-sEVs combined with two autophagy inhibitors produced significantly greater reductions in BACE1, PS-1, and APP expression, bringing their levels closer to those observed in control animals compared to autophagy inhibition alone. Histopathological examination revealed that amyloid plaques were significantly smaller in rats receiving the combined treatment.122 In a recent investigation, BACE1-targeting siRNA and berberine (BBR), a potent anti-inflammatory alkaloid with notable neuroprotective effects, were loaded into MSCs-sEVs via ultrasound-assisted encapsulation and co-incubated with neuronal cells. Sun et al demonstrated that this approach significantly decreased BACE1 expression and Aβ accumulation compared with untreated AD models. Furthermore, IL-6 and TNF-α were substantially suppressed, resulting in attenuated neuroinflammation. Importantly, the engineered MSCs-sEVs were delivered intranasally, a non-invasive and effective route enabling vesicles to bypass the blood-brain barrier and reach brain tissue directly. This approach not only reduces Aβ production at the source but also shifts microglial activation from a pro-inflammatory to an anti-inflammatory state, thus ultimately ameliorating cognitive deficits in AD-modeled mice.123
MSC-EVs Modulate Tau Protein Hyperphosphorylation
Tau protein hyperphosphorylation, which leads to the formation of NFTs, represents another key pathological hallmark of AD. Specifically, miR-132-3p is one of the most abundant miRNAs in brain tissue. In the hippocampus, decreased miR-132-3p has been associated with reduced synaptogenesis, increased production of Aβ, and enhanced Tau hyperphosphorylation. Ma et al demonstrated that miR-132-3p within MSCs-sEVs can downregulate RAS p21 protein activator 1 (RASA1) while upregulating Ras expression. This cascade leads to increased phosphorylation of protein kinase B (AKT) and GSK-3β, a critical kinase regulating both Aβ production and Tau phosphorylation. Accordingly, treatment with miR-132-3p-enriched MSCs-sEVs significantly attenuated Tau hyperphosphorylation in neurons subjected to oxygen-glucose deprivation (OGD).124
Anti-Neuroinflammatory Effects of MSC-EVs
Numerous studies have demonstrated that MSC-EVs modulate neuroinflammation by balancing pro-inflammatory and anti-inflammatory cytokines. In the brain tissues, a critical event in neuroinflammation is the activation and polarization of microglia towards the pro-inflammatory M1 phenotype.125 Losurdo et al administered MSCs-sEVs intranasally to microglia and neurons in AD-modelled mice and observed a significant reduction in the expression of CD68, a marker of activated microglia, in treated microglia. This suggests that MSC-EVs inhibit microglial polarization towards the M1 pro-inflammatory phenotype.126 Similarly, curcumin-pretreated MSCs-sEVs have been shown to upregulate CD206, an M2 anti-inflammatory microglial marker, while downregulating CD86, an M1 pro-inflammatory marker.74
In addition, miR-146a is an anti-inflammatory miRNA that suppresses NF-κB signaling by targeting interleukin-1 receptor-associated kinase 1 (IRAK1) and tumor necrosis factor receptor-associated factor 6 (TRAF6). Nakano et al demonstrated that sEVs derived from bone marrow mesenchymal stem cells (BM-MSCs) could deliver miR-146a to astrocytes, effectively downregulating NF-κB activity and exerting anti-inflammatory effects.127
Neuroprotective Effects of MSC-EVs
The length and density of neuronal synapses are closely linked to the progression of AD. In recent studies, Fang et al demonstrated that the administration of MSC-EVs enriched with miR-124 into a rat model of spinal cord injury promoted autophagy and mitigated oxidative stress, thereby exerting neuroprotective effects.128 Earlier research by Zhou et al showed that MSC-EVs facilitated motor function recovery in aged rhesus monkeys following primary motor cortex (M1) injury by promoting homeostatic ramified microglia, reducing injury-associated neuronal hyperexcitability, and enhancing synaptic plasticity within the peri-infarct cortex. Activated hypertrophic microglia secrete C1q, and the increased expression of C1q-positive hypertrophic microglia in the peri-lesional M1 area correlated with MSC-EVs treatment and improved functional outcomes.129
Clinical Trials of MSC-EVs in AD
As summarized in this review, the therapeutic potential of MSC-EVs has been extensively investigated in various preclinical studies using animal models and cell cultures. However, clinical studies exploring MSC-EVs for the treatment of AD remain limited.
To date, three clinical trials have evaluated MSC-EVs therapy for COVID-19. Among these, two studies employed nebulized MSC-EVs treatment (ChiCTR2000030261, IRCT20200217046526N2),130,131 demonstrating that MSC-EVs can effectively attenuate inflammatory responses with minimal adverse events. Another trial indicated that intravenous infusion of MSC-EVs may reverse cytokine storm in critically ill patients.132 Beyond COVID-19, MSC-EVs are also being investigated in Phase I/II clinical trials for conditions such as anal fistula, knee osteoarthritis, and Sjögren’s syndrome.133–135 Xie et al from Shanghai Jiao Tong University conducted a clinical evaluation of allogeneic human adipose-derived MSC-sEVs for safety and efficacy in patients with mild to moderate AD (NCT04388982). Nine patients received intranasal administration of MSCs-sEVs at different doses diluted in physiological saline (2×108, 4×108, and 8×108 particles/mL) twice weekly for 12 weeks. The treatment was found to be safe and well-tolerated. For future studies, doses of at least 4×108 particles are recommended for randomized Phase II and III clinical trials.136 Despite limitations, including small sample size and non-rigorous inclusion/exclusion criteria, this study provides encouraging preliminary evidence supporting the clinical potential of MSC-EVs for AD treatment.
Advantages and Challenges of MSC-EVs in AD Treatment
Extensive research indicates that MSC-EVs serve as innate delivery vehicles capable of transporting diverse bioactive cargos, including nucleic acids (miRNAs, mRNAs), proteins, and lipids, across BBB. This enables targeted delivery of therapeutic molecules to the brain, facilitating modulation of neuroinflammation and enhancement of neuronal survival. Compared to MSC therapy, MSC-EVs lack proliferative capacity, thereby reducing the risk of tumorigenesis.137,138 Additionally, MSC-EVs demonstrate lower immunogenicity and diminished potential for transplant rejection.103,139 Crucially, MSC-EVs offer versatile administration routes such as intranasal nebulization, intravenous injection, and localized delivery.106–108 Moreover, MSC-EVs have a higher potential for bioengineering modifications to enhance drug payload delivery and targeting specificity, thus enabling personalized treatment strategies.140 In contrast, MSC-based therapies must carefully consider the transplantation route to optimize efficacy and safety. From a translational perspective, MSC-EVs can be produced on a large scale, exhibit good stability, and pose no ethical concerns.
As a promising therapeutic tool, MSC-EVs still face significant challenges, mainly in the following aspects: (1) Isolation, purification, and storage of MSC-EVs require ongoing optimization and standardization to improve comparability and reproducibility. Heterogeneity arises from EVs derived from MSCs of different sources. Current isolation methods, which rely primarily on density, size, and surface charge, may result in low yield or contamination.22 Clinical application depends on time-efficient, cost-effective, and convenient methods, so existing isolation techniques need further improvement. (2) MSC cell therapies possess differentiation capabilities for tissue repair, which MSC-EVs currently lack.110 (3) Some harmful factors secreted by MSCs via paracrine signaling may also be present in MSC-EVs, and whether these factors negatively affect disease progression remains unclear. (4) Data on the optimal administration routes, dosing regimens, and potential side effects of MSC-EVs in different diseases remain insufficient.141 MSC-EVs provide a breakthrough for treating AD. However, before broader clinical application, more experimental work is required to overcome technical bottlenecks and accelerate clinical translation.
Conclusion
Considerable efforts have been devoted to treating AD. However, therapeutic outcomes remain unsatisfactory. As an emerging cell-free therapeutic agent, MSC-EVs contribute to AD treatment by facilitating the clearance of pathogenic proteins, modulating neuroinflammation, and enhancing cognitive and motor functions. Compared to traditional cell therapies, MSC-EVs provide distinct advantages and are regarded as a promising novel strategy for treating AD. In parallel, EVs serve as highly ideal liquid biopsy tools for AD diagnostics. The ability to track labeled EVs non-invasively using imaging techniques further solidifies their role as powerful theranostic agents, bridging diagnostic assessment with therapeutic monitoring.
Abbreviations
AD, Alzheimer’s disease; MSC-EVs, mesenchymal stem cell-derived extracellular vesicles; Aβ, β-amyloid; NFTs, neurofibrillary tangles; MSCs, mesenchymal stem cells; EVs, extracellular vesicles; ISEV, International Society for Extracellular Vesicles; sEVs, small extracellular vesicles; LEVs, large extracellular vesicles; ACDVs, artificial cell-derived vesicles; SVs: synthetic vesicles; dUC, differential ultracentrifugation; SEC, size-exclusion chromatography; ADAM10, α-secretase; BACE1, β-secretase; APP, amyloid precursor protein; sAPPα, soluble APPα; AICD, APP intracellular domain; sAPPβ, soluble APPβ; IL-1β, interleukin-1β; TNF-α, tumor necrosis factor-alpha; GSK-3β, glycogen synthase kinase-3β; CDK5, cyclin-dependent kinase 5; NMDA, N-methyl-D-aspartate; NOD, intracellular nucleotide-binding oligomerization domain; NLR, NOD-like receptor; DAM, disease-associated microglia; IFN-γ, interferon-gamma; Inos, inducible nitric oxide synthase; NO, nitric oxide; TREM2, triggering receptor expressed on myeloid cells 2; ROS, reactive oxygen species; ECT, electron transport chain; PET, positron emission tomography; CSF, cerebrospinal fluid; miRNAs, microRNAs; NDEVs, neuron-derived small EVs; MCI, mild cognitive impairment; ANXA5, annexin A5; GAP43, growth-associated protein 43; SNAP25, synaptosomal-associated protein 25; aMCI, amnestic mild cognitive impairment; C1QC, complement C1q subcomponent subunit C; GP1BB, platelet glycoprotein Ib beta chain; RSU1, Ras suppressor protein 1; BBB, blood-brain barrier; Cory-B, Corynoxine-B; NSCs, neural stem cells; AIS, American Spinal Injury Association Impairment Scale; IDE, insulin-degrading enzyme; NEP, neprilysin; PS-1, presenilin-1; BBR, berberine; RASA1, RAS p21 protein activator 1; AKT, protein kinase B; OGD, oxygen-glucose deprivation; IRAK1, interleukin-1 receptor-associated kinase 1; TRAF6, tumor necrosis factor receptor-associated factor 6; BM-MSCs, bone marrow mesenchymal stem cells; M1, primary motor cortex.
Acknowledgments
We would like to express our profound gratitude to all participants for their invaluable contributions to this research. Additionally, we appreciate the assistance of BioRender.com in creating the figures for this study. We would like to thank KetengEdit (www.ketengedit.com) for its linguistic assistance during the preparation of this manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Municipal level scientific research plan project of Ganzhou Municipal Health Commission (2022-2-62), Science and Technology Plan of Jiangxi Provincial Administration of Traditional Chinese Medicine (2022B495).
Disclosure
The authors state no conflict of interest.
References
1. R AA. Pathogenesis of Alzheimer’s disease. Int J Mol Sci. 2022;24(1):107. doi:10.3390/ijms24010107
2. Alzheimer’s association. 2024 Alzheimer’s disease facts and figures. Alzheimers Dement. 2024;20(5):3708–3821. doi:10.1002/alz.13809
3. Gou X, Fu Y, Li J, Xiang J, Yang M, Z Y. Impact of nanoplastics on Alzheimer ‘s disease: enhanced amyloid-β peptide aggregation and augmented neurotoxicity. J Hazard Mater. 2024;465:133518. doi:10.1016/j.jhazmat.2024.133518
4. Horie K, Barthélemy NR, Sato C, B RJ. CSF tau microtubule binding region identifies tau tangle and clinical stages of Alzheimer’s disease. Brain. 2021;144(2):515–527. doi:10.1093/brain/awaa373
5. Dubois B, von Arnim CAF, Burnie N, Bozeat S, C J. Biomarkers in Alzheimer’s disease: role in early and differential diagnosis and recognition of atypical variants. Alzheimer’s Res Ther. 2023;15(1):175. doi:10.1186/s13195-023-01314-6
6. Graff-Radford J, Yong KXX, Apostolova LG, et al. New insights into atypical Alzheimer’s disease in the era of biomarkers. Lancet Neurol. 2021;20(3):222–234. doi:10.1016/s1474-4422(20)30440-3
7. Twarowski B, H M. Inflammatory processes in Alzheimer’s disease-pathomechanism, diagnosis and treatment: a review. Int J Mol Sci. 2023;24(7):6518. doi:10.3390/ijms24076518
8. Andrzejewska A, Dabrowska S, Lukomska B, J M. Mesenchymal Stem Cells for Neurological Disorders. Adv Sci. 2021;8(7):2002944. doi:10.1002/advs.202002944
9. Massaro F, Corrillon F, Stamatopoulos B, et al. Age-related changes in human bone marrow mesenchymal stromal cells: morphology, gene expression profile, immunomodulatory activity and miRNA expression. Front Immunol. 2023;14:1267550. doi:10.3389/fimmu.2023.1267550
10. Brizio M, Mancini M, Lora M, et al. Cytokine priming enhances the antifibrotic effects of human adipose derived mesenchymal stromal cells conditioned medium. Stem Cell Res Ther. 2024;15(1):329. doi:10.1186/s13287-024-03916-9
11. Ferro F, Spelat R, Shaw G, et al. Regenerative and anti-inflammatory potential of regularly fed, starved cells and extracellular vesicles in vivo. Cells. 2022;11(17):2969. doi:10.3390/cells11172696
12. Liu H, Wang J, Yue G, Xu J. Placenta-derived mesenchymal stem cells protect against diabetic kidney disease by upregulating autophagy-mediated SIRT1/FOXO1 pathway. Renal Failure. 2024;46(1):2303396. doi:10.1080/0886022x.2024.2303396
13. Dreyer GJ, Drabbels JJ, de Fijter JW, van Kooten C, Reinders ME, Heidt S. Cell-free DNA measurement of three genomes after allogeneic MSC therapy in kidney transplant recipients indicates early cell death of infused MSC. Front Immunol. 2023;14:1240347. doi:10.3389/fimmu.2023.1240347
14. Lee H, Lee TW, Chandrasekharan A, et al. Injectable self-crosslinkable thiolated hyaluronic acid for stem cell therapy of atopic dermatitis. ACS Biomater Sci Eng. 2022;8(4):1613–1622. doi:10.1021/acsbiomaterials.1c01374
15. Chen J, Liu Y, Zhang J, et al. External application of human umbilical cord-derived mesenchymal stem cells in hyaluronic acid gel repairs foot wounds of types i and ii diabetic rats through paracrine action mode. Stem Cells Transl Med. 2023;12(10):689–706. doi:10.1093/stcltm/szad050
16. Yang H, Zhou Y, Ying B, Dong X, Qian Q, Gao S. Effects of human umbilical cord mesenchymal stem cell-derived exosomes in the rat osteoarthritis models. Stem Cells Transl Med. 2024;13(8):803–811. doi:10.1093/stcltm/szae031
17. Wang L, Wang W, Hu D, et al. Tumor-derived extracellular vesicles regulate macrophage polarization: role and therapeutic perspectives. Front Immunol. 2024;15:1346587. doi:10.3389/fimmu.2024.1346587
18. Wu R, Fan X, Wang Y, et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles in Liver Immunity and Therapy. Front Immunol. 2022;13:833878. doi:10.3389/fimmu.2022.833878
19. Cheng J, Zhang Y, Ma L, et al. Macrophage-derived extracellular vesicles-coated palladium nanoformulations modulate inflammatory and immune homeostasis for targeting therapy of ulcerative colitis. Adv Sci. 2023;10(33):e2304002. doi:10.1002/advs.202304002
20. Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor. Cell. 1983;33(3):967–978. doi:10.1016/0092-8674(83)90040-5
21. Harding CV, Heuser JE, Stahl PD. Exosomes: looking back three decades and into the future. J Cell Biol. 2013;200(4):367–371. doi:10.1083/jcb.201212113
22. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
23. Mas-Bargues C, Alique M. Extracellular vesicles as “Very Important Particles” (VIPs) in aging. Int J Mol Sci. 2023;24(4):4250. doi:10.3390/ijms24044250
24. Lötvall J, Hill AF, Hochberg F, et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: a position statement from the international society for extracellular vesicles. J Extracell Vesicles. 2014;3:26913. doi:10.3402/jev.v3.26913
25. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the international society for extracellular vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi:10.1080/20013078.2018.1535750
26. Visan KS, Lobb RJ, Ham S, et al. Comparative analysis of tangential flow filtration and ultracentrifugation, both combined with subsequent size exclusion chromatography, for the isolation of small extracellular vesicles. J Extracell Vesicles. 2022;11(9):e12266. doi:10.1002/jev2.12266
27. Karimi N, Dalirfardouei R, Dias T, Lötvall J, Lässer C. Tetraspanins distinguish separate extracellular vesicle subpopulations in human serum and plasma - Contributions of platelet extracellular vesicles in plasma samples. J Extracell Vesicles. 2022;11(5):e12213. doi:10.1002/jev2.12213
28. Guo J, Wu C, Lin X, et al. Establishment of a simplified dichotomic size-exclusion chromatography for isolating extracellular vesicles toward clinical applications. J Extracell Vesicles. 2021;10(11):e12145. doi:10.1002/jev2.12145
29. Zhang M, Lu Y, Wang L, Mao Y, Hu X, Chen Z. Current status of research on small extracellular vesicles for the diagnosis and treatment of urological tumors. Cancers. 2022;15(1):100. doi:10.3390/cancers15010100
30. Monteiro AR, Barbosa DJ, Remião F, Silva R. Alzheimer’s disease: insights and new prospects in disease pathophysiology, biomarkers and disease-modifying drugs. Biochem Pharmacol. 2023;211:115522. doi:10.1016/j.bcp.2023.115522
31. Thakur S, Dhapola R, Sarma P, Medhi B, Reddy DH. Neuroinflammation in Alzheimer’s disease: current progress in molecular signaling and therapeutics. Inflammation. 2023;46(1):1–17. doi:10.1007/s10753-022-01721-1
32. Matuszyk MM, Garwood CJ, Ferraiuolo L, Simpson JE, Staniforth RA, Wharton SB. Biological and methodological complexities of beta-amyloid peptide: implications for Alzheimer’s disease research. J Neurochem. 2022;160(4):434–453. doi:10.1111/jnc.15538
33. Khan S, Barve KH, Kumar MS. Recent advancements in pathogenesis, diagnostics and treatment of Alzheimer’s disease. Curr Neuropharmacol. 2020;18(11):1106–1125. doi:10.2174/1570159x18666200528142429
34. Princen K, Van Dooren T, van Gorsel M, et al. Pharmacological modulation of septins restores calcium homeostasis and is neuroprotective in models of Alzheimer’s disease. Science. 2024;384(6699):eadd6260. doi:10.1126/science.add6260
35. Yang Y, Zhang J, Yang X, et al. Dysregulated APP expression and α-secretase processing of APP is involved in manganese-induced cognitive impairment. Ecotoxicol Environ Saf. 2021;220:112365. doi:10.1016/j.ecoenv.2021.112365
36. Wang X, Jia J. Magnolol improves Alzheimer’s disease-like pathologies and cognitive decline by promoting autophagy through activation of the AMPK/mTOR/ULK1 pathway. Biomed Pharmacother. 2023;161:114473. doi:10.1016/j.biopha.2023.114473
37. Vilella A, Bodria M, Papotti B, et al. PCSK9 ablation attenuates Aβ pathology, neuroinflammation and cognitive dysfunctions in 5XFAD mice. Brain Behav Immun. 2024;115:517–534. doi:10.1016/j.bbi.2023.11.008
38. Gratuze M, Jiang H, Wang C, Xiong M, Bao X, Holtzman DM. APOE antibody inhibits Aβ-associated tau seeding and spreading in a mouse model. Ann Neurol. 2022;91(6):847–852. doi:10.1002/ana.26351
39. McMillan PJ, Benbow SJ, Uhrich R, et al. Tau-RNA complexes inhibit microtubule polymerization and drive disease-relevant conformation change. Brain. 2023;146(8):3206–3220. doi:10.1093/brain/awad032
40. Robles-Gómez ÁA, Ordaz B, Lorea-Hernández JJ, Peña-Ortega F. Deleterious and protective effects of epothilone-D alone and in the context of amyloid β- and tau-induced alterations. Front Mol Neurosci. 2023;16:1198299. doi:10.3389/fnmol.2023.1198299
41. Zhang D, Zhang W, Ming C, et al. P-tau217 correlates with neurodegeneration in Alzheimer’s disease, and targeting p-tau217 with immunotherapy ameliorates murine tauopathy. Neuron. 2024;112(10):1676–1693.e12. doi:10.1016/j.neuron.2024.02.017
42. Baltissen D, Bold CS, Rehra L, et al. APPsα rescues CDK5 and GSK3β dysregulation and restores normal spine density in Tau transgenic mice. Front Cell Neurosci. 2023;17:1106176. doi:10.3389/fncel.2023.1106176
43. Zhong S, Ye J, Deng Y, et al. Quercetagitrin inhibits tau accumulation and reverses neuroinflammation and cognitive deficits in P301S-tau transgenic mice. Molecules. 2023;28(9):3964. doi:10.3390/molecules28093964
44. Bassil F, Brown HJ, Pattabhiraman S, et al. Amyloid-beta (Aβ) plaques promote seeding and spreading of alpha-synuclein and tau in a mouse model of Lewy body disorders with Aβ pathology. Neuron. 2020;105(2):260–275.e6. doi:10.1016/j.neuron.2019.10.010
45. Islam MS, Lai CC, Wang LH, Lin HH. Inhibition of NMDA receptor activation in the rostral ventrolateral medulla by amyloid-β Peptide in rats. Biomolecules. 2023;13(12):1736. doi:10.3390/biom13121736
46. Sawant N, Kshirsagar S, Reddy PH, Reddy AP. Protective effects of SSRI, Citalopram in mutant APP and mutant Tau expressed dorsal raphe neurons in Alzheimer’s disease. Biochim Biophys Acta Mol Basis Dis. 2024;1870(2):166942. doi:10.1016/j.bbadis.2023.166942
47. Sangineto M, Ciarnelli M, Cassano T, et al. Metabolic reprogramming in inflammatory microglia indicates a potential way of targeting inflammation in Alzheimer’s disease. Redox Biol. 2023;66:102846. doi:10.1016/j.redox.2023.102846
48. Moonen S, Koper MJ, Van Schoor E, et al. Pyroptosis in Alzheimer’s disease: cell type-specific activation in microglia, astrocytes and neurons. Acta Neuropathol. 2023;145(2):175–195. doi:10.1007/s00401-022-02528-y
49. Kimura K, Subramanian A, Yin Z, et al. Immune checkpoint TIM-3 regulates microglia and Alzheimer’s disease. Nature. 2025;641(8063):718–731. doi:10.1038/s41586-025-08852-z
50. Wang S, Sudan R, Peng V, et al. TREM2 drives microglia response to amyloid-β via SYK-dependent and -independent pathways. Cell. 2022;185(22):4153–4169.e19. doi:10.1016/j.cell.2022.09.033
51. Shen Y, Liu F, Zhang M. Therapeutic potential of plant-derived natural compounds in Alzheimer’s disease: targeting microglia-mediated neuroinflammation. Biomed Pharmacother. 2024;178:117235. doi:10.1016/j.biopha.2024.117235
52. Wang X, Xie Y, Fan X, Wu X, Wang D, Zhu L. Intermittent hypoxia training enhances Aβ endocytosis by plaque associated microglia via VPS35-dependent TREM2 recycling in murine Alzheimer’s disease. Alzheimer’s Res Ther. 2024;16(1):121. doi:10.1186/s13195-024-01489-6
53. Leng L, Yuan Z, Pan R, et al. Microglial hexokinase 2 deficiency increases ATP generation through lipid metabolism leading to β-amyloid clearance. Nat Metab. 2022;4(10):1287–1305. doi:10.1038/s42255-022-00643-4
54. Sun Y, Zhang H, Zhang X, et al. Promotion of astrocyte-neuron glutamate-glutamine shuttle by SCFA contributes to the alleviation of Alzheimer’s disease. Redox Biol. 2023;62:102690. doi:10.1016/j.redox.2023.102690
55. Zheng J, Xie Y, Ren L, et al. GLP-1 improves the supportive ability of astrocytes to neurons by promoting aerobic glycolysis in Alzheimer’s disease. Mol Metabol. 2021;47:101180. doi:10.1016/j.molmet.2021.101180
56. Joshi M, Joshi S, Khambete M, Degani M. Role of calcium dysregulation in Alzheimer’s disease and its therapeutic implications. Chem Biol Drug Des. 2023;101(2):453–468. doi:10.1111/cbdd.14175
57. Guo Z, Tian C, Shi Y, et al. Blood-based CNS regionally and neuronally enriched extracellular vesicles carrying pTau217 for Alzheimer’s disease diagnosis and differential diagnosis. Acta Neuropathologica Commun. 2024;12(1):38. doi:10.1186/s40478-024-01727-w
58. Cai Y, Chen T, Cai Y, et al. Surface protein profiling and subtyping of extracellular vesicles in body fluids reveals non-CSF biomarkers of Alzheimer’s disease. J Extracell Vesicles. 2024;13(4):e12432. doi:10.1002/jev2.12432
59. Serpente M, Fenoglio C, D’Anca M, et al. MiRNA profiling in plasma neural-derived small extracellular vesicles from patients with Alzheimer’s disease. Cells. 2020;9(6):1443. doi:10.3390/cells9061443
60. Kumar A, Su Y, Sharma M, et al. MicroRNA expression in extracellular vesicles as a novel blood-based biomarker for Alzheimer’s disease. Alzheimers Dement. 2023;19(11):4952–4966. doi:10.1002/alz.13055
61. Visconte C, Fenoglio C, Serpente M, et al. Altered extracellular vesicle miRNA profile in prodromal alzheimer’s disease. Int J Mol Sci. 2023;24(19):14749. doi:10.3390/ijms241914749
62. Reho P, Kalia V, Jackson GL, et al. Preclinical Alzheimer’s disease shows alterations in circulating neuronal-derived extracellular vesicle microRNAs in a multiethnic cohort. Alzheimers Dement. 2025;21(3):e70050. doi:10.1002/alz.70050
63. Ryu IS, Kim DH, Ro JY, et al. The microRNA-485-3p concentration in salivary exosome-enriched extracellular vesicles is related to amyloid β deposition in the brain of patients with Alzheimer’s disease. Clin Biochem. 2023;118:110603. doi:10.1016/j.clinbiochem.2023.110603
64. Kumar S, Reddy PH. Elevated levels of MicroRNA-455-3p in the cerebrospinal fluid of Alzheimer’s patients: a potential biomarker for Alzheimer’s disease. Biochim Biophys Acta Mol Basis Dis. 2021;1867(4):166052. doi:10.1016/j.bbadis.2020.166052
65. Muraoka S, DeLeo AM, Sethi MK, et al. Proteomic and biological profiling of extracellular vesicles from Alzheimer’s disease human brain tissues. Alzheimers Dement. 2020;16(6):896–907. doi:10.1002/alz.12089
66. Jia L, Zhu M, Kong C, et al. Blood neuro-exosomal synaptic proteins predict Alzheimer’s disease at the asymptomatic stage. Alzheimers Dement. 2021;17(1):49–60. doi:10.1002/alz.12166
67. Cai H, Pang Y, Wang Q, et al. Proteomic profiling of circulating plasma exosomes reveals novel biomarkers of Alzheimer’s disease. Alzheimer’s Res Ther. 2022;14(1):181. doi:10.1186/s13195-022-01133-1
68. Chatterjee M, Özdemir S, Kunadt M, et al. C1q is increased in cerebrospinal fluid-derived extracellular vesicles in Alzheimer’s disease: a multi-cohort proteomics and immuno-assay validation study. Alzheimers Dement. 2023;19(11):4828–4840. doi:10.1002/alz.13066
69. Nazri HM, Greaves E, Quenby S, Dragovic R, Tapmeier TT, Becker CM. The role of small extracellular vesicle-miRNAs in endometriosis. Hum Reprod. 2023;38(12):2296–2311. doi:10.1093/humrep/dead216
70. Matsuzaka Y, Yashiro R. Therapeutic strategy of mesenchymal-stem-cell-derived extracellular vesicles as regenerative medicine. Int J Mol Sci. 2022;23(12):6480. doi:10.3390/ijms23126480
71. Wang D, Wan Z, Yang Q, et al. Sonodynamical reversion of immunosuppressive microenvironment in prostate cancer via engineered exosomes. Drug Delivery. 2022;29(1):702–713. doi:10.1080/10717544.2022.2044937
72. Wang X, Hao X, Zhang Y, et al. Bioinspired adaptive microdrugs enhance the chemotherapy of malignant glioma: beyond their nanodrugs. Adv Mater. 2024;36(32):e2405165. doi:10.1002/adma.202405165
73. Zhu Z, Zhai Y, Hao Y, et al. Specific anti-glioma targeted-delivery strategy of engineered small extracellular vesicles dual-functionalised by Angiopep-2 and TAT peptides. J Extracell Vesicles. 2022;11(8):e12255. doi:10.1002/jev2.12255
74. Bashirrohelleh MA, Bavarsad K, Khodadadi A, Shohan M, Asadirad A. Curcumin-enhanced stem cell exosomes: a novel approach to modulating neuroinflammation and improving cognitive function in a rat model of Alzheimer’s disease. Eur J Pharmacol. 2025;999:177695. doi:10.1016/j.ejphar.2025.177695
75. Wang X, Li D, Li G, et al. Enhanced therapeutic potential of hybrid exosomes loaded with paclitaxel for cancer therapy. Int J Mol Sci. 2024;25(7):3645. doi:10.3390/ijms25073645
76. Iyaswamy A, Thakur A, Guan XJ, et al. Fe65-engineered neuronal exosomes encapsulating corynoxine-B ameliorate cognition and pathology of Alzheimer’s disease. Signal Transduct Target Ther. 2023;8(1):404. doi:10.1038/s41392-023-01657-4
77. Liu X, Chen C, Jiang Y, et al. Brain-derived extracellular vesicles promote bone-fat imbalance in Alzheimer’s disease. Int J Bio Sci. 2023;19(8):2409–2427. doi:10.7150/ijbs.79461
78. Khan MI, Jeong ES, Khan MZ, Shin JH, Kim JD. Stem cells-derived exosomes alleviate neurodegeneration and Alzheimer’s pathogenesis by ameliorating neuroinflamation, and regulating the associated molecular pathways. Sci Rep. 2023;13(1):15731. doi:10.1038/s41598-023-42485-4
79. Peng D, Liu T, Lu H, et al. Intranasal delivery of engineered extracellular vesicles loaded with miR-206-3p antagomir ameliorates Alzheimer’s disease phenotypes. Theranostics. 2024;14(19):7623–7644. doi:10.7150/thno.103596
80. Leroux E, Perbet R, Caillierez R, et al. Extracellular vesicles: major actors of heterogeneity in tau spreading among human tauopathies. Mol Ther. 2022;30(2):782–797. doi:10.1016/j.ymthe.2021.09.020
81. Aafreen S, Feng J, Wang W, Liu G. Theranostic extracellular vesicles: a concise review of current imaging technologies and labeling strategies. Extracell Vesicles Circ Nucl Acids. 2023;4(1):107–132. doi:10.20517/evcna.2023.01
82. Chuo ST, Chien JC, Lai CP. Imaging extracellular vesicles: current and emerging methods. J Biomed Sci. 2018;25(1):91. doi:10.1186/s12929-018-0494-5
83. Gao Y, Chu C, Jablonska A, Bulte JWM, Walczak P, Janowski M. Imaging as a tool to accelerate the translation of extracellular vesicle-based therapies for central nervous system diseases. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2021;13(3):e1688. doi:10.1002/wnan.1688
84. Chen W, Peng J, Tang X, Ouyang S. MSC-derived exosome ameliorates pulmonary fibrosis by modulating NOD 1/NLRP3-mediated epithelial-mesenchymal transition and inflammation. Heliyon. 2025;11(2):e41436. doi:10.1016/j.heliyon.2024.e41436
85. Nguyen VVT, Welsh JA, Tertel T, et al. Inter-laboratory multiplex bead-based surface protein profiling of MSC-derived EV preparations identifies MSC-EV surface marker signatures. J Extracell Vesicles. 2024;13(6):e12463. doi:10.1002/jev2.12463
86. Hou L, Zhu Z, Jiang F, et al. Human umbilical cord mesenchymal stem cell-derived extracellular vesicles alleviated silica induced lung inflammation and fibrosis in mice via circPWWP2A/miR-223-3p/NLRP3 axis. Ecotoxicol Environ Saf. 2023;251:114537. doi:10.1016/j.ecoenv.2023.114537
87. Yang Y, Chen Y, Liu Y, et al. Mesenchymal stem cells and pulmonary fibrosis: a bibliometric and visualization analysis of literature published between 2002 and 2021. Front Pharmacol. 2023;14:1136761. doi:10.3389/fphar.2023.1136761
88. Hass R. Role of MSC in the tumor microenvironment. Cancers. 2020;12(8):2107. doi:10.3390/cancers12082107
89. Vilar A, Hodgson-Garms M, Kusuma GD, et al. Substrate mechanical properties bias MSC paracrine activity and therapeutic potential. Acta Biomater. 2023;168:144–158. doi:10.1016/j.actbio.2023.06.041
90. Zhang L, Dong ZF, Zhang JY. Immunomodulatory role of mesenchymal stem cells in Alzheimer’s disease. Life Sci. 2020;246:117405. doi:10.1016/j.lfs.2020.117405
91. Bydon M, Qu W, Moinuddin FM, et al. Intrathecal delivery of adipose-derived mesenchymal stem cells in traumatic spinal cord injury: Phase I trial. Nat Commun. 2024;15(1):2201. doi:10.1038/s41467-024-46259-y
92. Zaripova LN, Midgley A, Christmas SE, et al. Mesenchymal stem cells in the pathogenesis and therapy of autoimmune and autoinflammatory diseases. Int J Mol Sci. 2023;24(22):16040. doi:10.3390/ijms242216040
93. Dias IX, Cordeiro A, Guimarães JAM, KR S. Potential and limitations of induced pluripotent stem cells-derived mesenchymal stem cells in musculoskeletal disorders treatment. Biomolecules. 2023;13(9):1342. doi:10.3390/biom13091342
94. Sid-Otmane C, Perrault LP, Ly HQ. Mesenchymal stem cell mediates cardiac repair through autocrine, paracrine and endocrine axes. J Transl Med. 2020;18(1):336. doi:10.1186/s12967-020-02504-8
95. Trohatou O, Roubelakis MG. Mesenchymal Stem/Stromal Cells in Regenerative Medicine: past, Present, and Future. Cell Reprogramming. 2017;19(4):217–224. doi:10.1089/cell.2016.0062
96. Potter DR, Miyazawa BY, Gibb SL, et al. Mesenchymal stem cell-derived extracellular vesicles attenuate pulmonary vascular permeability and lung injury induced by hemorrhagic shock and trauma. J Trauma Acute Care Surg. 2018;84(2):245–256. doi:10.1097/ta.0000000000001744
97. Pu Y, Li C, Qi X, et al. Extracellular Vesicles from NMN Preconditioned Mesenchymal Stem Cells Ameliorated Myocardial Infarction via miR-210-3p Promoted Angiogenesis. Stem Cell Rev Rep. 2023;19(4):1051–1066. doi:10.1007/s12015-022-10499-6
98. Rezaie J, Nejati V, Mahmoodi M, Ahmadi M. Mesenchymal stem cells derived extracellular vesicles: a promising nanomedicine for drug delivery system. Biochem Pharmacol. 2022;203:115167. doi:10.1016/j.bcp.2022.115167
99. Zhou Y, Wen LL, Li YF, et al. Exosomes derived from bone marrow mesenchymal stem cells protect the injured spinal cord by inhibiting pericyte pyroptosis. Neural Regen Res. 2022;17(1):194–202. doi:10.4103/1673-5374.314323
100. Hassaan NA, Mansour HA. Exosomal therapy is a luxury area for regenerative medicine. Tissue Cell. 2024;91:102570. doi:10.1016/j.tice.2024.102570
101. Preda MB, Lupan AM, Neculachi CA, et al. Evidence of mesenchymal stromal cell adaptation to local microenvironment following subcutaneous transplantation. J Cell Mol Med. 2020;24(18):10889–10897. doi:10.1111/jcmm.15717
102. Hoang VT, Le DS, Hoang DM, et al. Impact of tissue factor expression and administration routes on thrombosis development induced by mesenchymal stem/stromal cell infusions: re-evaluating the dogma. Stem Cell Res Ther. 2024;15(1):56. doi:10.1186/s13287-023-03582-3
103. Bulut Ö, Gürsel İ. Mesenchymal stem cell derived extracellular vesicles: promising immunomodulators against autoimmune, autoinflammatory disorders and SARS-CoV-2 infection. Turk J Biol. 2020;44(3):273–282. doi:10.3906/biy-2002-79
104. Tsai HI, Wu Y, Liu X, et al. Engineered small extracellular vesicles as a FGL1/PD-L1 Dual-targeting delivery system for alleviating immune rejection. Adv Sci. 2022;9(3):e2102634. doi:10.1002/advs.202102634
105. Phelps J, Hart DA, Mitha AP, Duncan NA, Sen A. Extracellular vesicles generated by mesenchymal stem cells in stirred suspension bioreactors promote angiogenesis in human-brain-derived endothelial cells. Int J Mol Sci. 2024;25(10):5219. doi:10.3390/ijms25105219
106. Zhao R, Wang L, Wang T, Xian P, Wang H, Long Q. Inhalation of MSC-EVs is a noninvasive strategy for ameliorating acute lung injury. J Control Release. 2022;345:214–230. doi:10.1016/j.jconrel.2022.03.025
107. Jin T, Liu X, Li G, Sun S, Xie L. Intravenous injection of BMSCs modulate tsRNA expression and ameliorate lung remodeling in COPD mice. Stem Cell Res Ther. 2024;15(1):450. doi:10.1186/s13287-024-04066-8
108. Zhang B, Lai RC, Sim WK, Tan TT, Lim SK. An assessment of administration route on MSC-sEV therapeutic efficacy. Biomolecules. 2024;14(6):622. doi:10.3390/biom14060622
109. de Witte SFH, Luk F, Sierra Parraga JM, et al. Immunomodulation by therapeutic Mesenchymal Stromal Cells (MSC) is triggered through phagocytosis of MSC by monocytic cells. Stem Cells. 2018;36(4):602–615. doi:10.1002/stem.2779
110. Xie Z, Yu W, Zheng G, et al. TNF-α-mediated m(6)A modification of ELMO1 triggers directional migration of mesenchymal stem cell in ankylosing spondylitis. Nat Commun. 2021;12(1):5373. doi:10.1038/s41467-021-25710-4
111. Powsner EH, Kronstadt SM, Nikolov K, Aranda A, Jay SM. Mesenchymal stem cell extracellular vesicle vascularization bioactivity and production yield are responsive to cell culture substrate stiffness. Bioeng Transl Med. 2025;10(3):e10743. doi:10.1002/btm2.10743
112. Kolliopoulos V, Tiffany A, Polanek M, Harley BAC. Donor sex and passage conditions influence msc osteogenic response in mineralized collagen scaffolds. Adv Healthcare Mater. 2024;13(27):e2400039. doi:10.1002/adhm.202400039
113. Baudequin T, Naudot M, Dupont S, et al. Donor variability alters differentiation and mechanical cohesion of tissue-engineered constructs with human endothelial/MSC co-culture. Int J Artif Organs. 2021;44(11):868–879. doi:10.1177/03913988211051758
114. Yuan S, Li G, Zhang J, Chen X, Su J, Zhou F. Mesenchymal stromal cells-derived extracellular vesicles as potential treatments for osteoarthritis. Pharmaceutics. 2023;15(7):1814. doi:10.3390/pharmaceutics15071814
115. Wang T, Gong Y, Lin H, et al. Heat shock strengthens the protective potential of MSCs in liver injury by promoting EV release through upregulated autophagosome formation. J Extracell Vesicles. 2025;14(5):e70084. doi:10.1002/jev2.70084
116. Daga KR, Larey AM, Morfin MG, et al. Microglia morphological response to mesenchymal stromal cell extracellular vesicles demonstrates EV therapeutic potential for modulating neuroinflammation. J Biol Eng. 2024;18(1):58. doi:10.1186/s13036-024-00449-w
117. Zhang J, Buller BA, Zhang ZG, et al. Exosomes derived from bone marrow mesenchymal stromal cells promote remyelination and reduce neuroinflammation in the demyelinating central nervous system. Exp Neurol. 2022;347:113895. doi:10.1016/j.expneurol.2021.113895
118. Ye Y, Gao M, Shi W, et al. The immunomodulatory effects of mesenchymal stem cell-derived extracellular vesicles in Alzheimer’s disease. Front Immunol. 2023;14:1325530. doi:10.3389/fimmu.2023.1325530
119. Gholampour M, Basu MK, Swerdlow RH, Zhuo X, Haeri M. Cell-specific protein expression in Alzheimer’s disease prefrontal cortex. Alzheimers Dement. 2025;21(6):e70339. doi:10.1002/alz.70339
120. Corraliza-Gomez M, Bermejo T, Lilue J, et al. Insulin-degrading enzyme (IDE) as a modulator of microglial phenotypes in the context of Alzheimer’s disease and brain aging. J Neuroinflammation. 2023;20(1):233. doi:10.1186/s12974-023-02914-7
121. Qian C, Yang C, Lu M, et al. Activating AhR alleviates cognitive deficits of Alzheimer’s disease model mice by upregulating endogenous Aβ catabolic enzyme Neprilysin. Theranostics. 2021;11(18):8797–8812. doi:10.7150/thno.61601
122. Ebrahim N, Al Saihati HA, Alali Z, et al. Exploring the molecular mechanisms of MSC-derived exosomes in Alzheimer’s disease: autophagy, insulin and the PI3K/Akt/mTOR signaling pathway. Biomed Pharmacother. 2024;176:116836. doi:10.1016/j.biopha.2024.116836
123. Sun C, Sha S, Shan Y, et al. Intranasal delivery of BACE1 siRNA and berberine via engineered stem cell exosomes for the treatment of Alzheimer’s disease. Int J Nanomedicine. 2025;20:5873–5891. doi:10.2147/ijn.S506793
124. Ma X, Wang Y, Shi Y, et al. Exosomal miR-132-3p from mesenchymal stromal cells improves synaptic dysfunction and cognitive decline in vascular dementia. Stem Cell Res Ther. 2022;13(1):315. doi:10.1186/s13287-022-02995-w
125. Wang W, Li Y, Ma F, et al. Microglial repopulation reverses cognitive and synaptic deficits in an Alzheimer’s disease model by restoring BDNF signaling. Brain Behav Immun. 2023;113:275–288. doi:10.1016/j.bbi.2023.07.011
126. Losurdo M, Pedrazzoli M, D’Agostino C, et al. Intranasal delivery of mesenchymal stem cell-derived extracellular vesicles exerts immunomodulatory and neuroprotective effects in a 3xTg model of Alzheimer’s disease. Stem Cells Transl Med. 2020;9(9):1068–1084. doi:10.1002/sctm.19-0327
127. Nakano M, Kubota K, Kobayashi E, et al. Bone marrow-derived mesenchymal stem cells improve cognitive impairment in an Alzheimer’s disease model by increasing the expression of microRNA-146a in hippocampus. Sci Rep. 2020;10(1):10772. doi:10.1038/s41598-020-67460-1
128. Fang C, Qian J, Tu BZ, Xia X, Jia CY, Shen CL. MiR-124 delivered by extracellular vesicles from mesenchymal stem cell exerts neuroprotective effects by stabilizing the p62-Keap1-Nrf2 pathway after spinal cord injury in rats. Mol Neurobiol. 2025;62(7):8328–8340. doi:10.1007/s12035-025-04755-2
129. Zhou Y, Bhatt H, Mojica CA, et al. Mesenchymal-derived extracellular vesicles enhance microglia-mediated synapse remodeling after cortical injury in aging Rhesus monkeys. J Neuroinflammation. 2023;20(1):201. doi:10.1186/s12974-023-02880-0
130. Zarrabi M, Shahrbaf MA, Nouri M, et al. Allogenic mesenchymal stromal cells and their extracellular vesicles in COVID-19 induced ARDS: a randomized controlled trial. Stem Cell Res Ther. 2023;14(1):169. doi:10.1186/s13287-023-03402-8
131. Chu M, Wang H, Bian L, et al. Nebulization therapy with umbilical cord mesenchymal stem cell-derived exosomes for COVID-19 pneumonia. Stem Cell Rev Rep. 2022;18(6):2152–2163. doi:10.1007/s12015-022-10398-w
132. Sengupta V, Sengupta S, Lazo A, Woods P, Nolan A, Bremer N. Exosomes derived from bone marrow mesenchymal stem cells as treatment for severe COVID-19. Stem Cells Dev. 2020;29(12):747–754. doi:10.1089/scd.2020.0080
133. Figueroa-Valdés AI, Luz-Crawford P, Herrera-Luna Y, et al. Clinical-grade extracellular vesicles derived from umbilical cord mesenchymal stromal cells: preclinical development and first-in-human intra-articular validation as therapeutics for knee osteoarthritis. J Nanobiotechnol. 2025;23(1):13. doi:10.1186/s12951-024-03088-x
134. Pak H, Hadizadeh A, Heirani-Tabasi A, et al. Safety and efficacy of injection of human placenta mesenchymal stem cells derived exosomes for treatment of complex perianal fistula in non-Crohn’s cases: clinical trial Phase I. J Gastroenterol Hepatol. 2023;38(4):539–547. doi:10.1111/jgh.16110
135. Habibi A, Khosravi A, Soleimani M, Nejabat M, Dara M, Azarpira N. Efficacy of topical mesenchymal stem cell exosome in Sjögren’s syndrome-related dry eye: a randomized clinical trial. BMC Ophthalmol. 2025;25(1):299. doi:10.1186/s12886-025-04078-9
136. Xie X, Song Q, Dai C, et al. Clinical safety and efficacy of allogenic human adipose mesenchymal stromal cells-derived exosomes in patients with mild to moderate Alzheimer’s disease: a phase I/II clinical trial. Gen Psychiatry. 2023;36(5):e101143. doi:10.1136/gpsych-2023-101143
137. Han T, Song P, Wu Z, et al. MSC secreted extracellular vesicles carrying TGF-beta upregulate Smad 6 expression and promote the regrowth of neurons in spinal cord injured rats. Stem Cell Rev Rep. 2022;18(3):1078–1096. doi:10.1007/s12015-021-10219-6
138. Yari H, Mikhailova MV, Mardasi M, et al. Emerging role of mesenchymal stromal cells (MSCs)-derived exosome in neurodegeneration-associated conditions: a groundbreaking cell-free approach. Stem Cell Res Ther. 2022;13(1):423. doi:10.1186/s13287-022-03122-5
139. Liu WZ, Ma ZJ, Li JR, Kang XW. Mesenchymal stem cell-derived exosomes: therapeutic opportunities and challenges for spinal cord injury. Stem Cell Res Ther. 2021;12(1):102. doi:10.1186/s13287-021-02153-8
140. Li Y, Wang Y, Zhang Y, et al. Engineered mesenchymal stem cell-derived extracellular vesicles: kill tumors and protect organs. Theranostics. 2024;14(16):6202–6217. doi:10.7150/thno.99618
141. Fan Y, Chen Z, Zhang M. Role of exosomes in the pathogenesis, diagnosis, and treatment of central nervous system diseases. J Transl Med. 2022;20(1):291. doi:10.1186/s12967-022-03493-6
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.