Back to Journals » International Journal of Women's Health » Volume 17
Mendelian Randomization and Transcriptomics Analysis Reveal Potential Relationships Between Lipids, Immune Cells and PCOS
Authors Xie Y, Zhang Z, Zhao Y, Zhang J, Yin Q
Received 13 April 2025
Accepted for publication 22 July 2025
Published 7 August 2025 Volume 2025:17 Pages 2451—2463
DOI https://doi.org/10.2147/IJWH.S534136
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 6
Editor who approved publication: Dr Vinay Kumar
Yafei Xie, Zelin Zhang, Yuanyuan Zhao, Jiali Zhang, Qiaozhi Yin
School of Clinical Medicine, Chengdu University of Traditional Chinese Medicine, Chengdu, Sichuan, People’s Republic of China
Correspondence: Qiaozhi Yin, Email [email protected]
Background: It has been noted that plasma lipid levels are closely associated with polycystic ovary syndrome (PCOS). Immune cells have also been recognized as critical indicators for evaluating the impact of lipids on disease progression. However, there is still a lack of conclusive causal evidence as to whether immune cells mediate the relationship between lipids and PCOS.
Methods: This study used a two-step two-sample Mendelian randomisation (TSMR) analysis to explore the causal relationship between plasma lipid groups and PCOS, and the mediating role of immune cells in this relationship. Transcriptome analysis was used to further explore the relationship between lipid metabolism, immunity, and PCOS.
Results: The inverse variance weighted (IVW) method and Bayesian weighted Mendelian randomized (BWMR) method identified six plasma lipid groups that exhibited causal effects on PCOS, with no evidence of reverse causality. Eighteen immune cell traits are strongly associated with PCOS. TSMR analysis identified SSC-A on CD4+ as the key factor mediating the causal association of PC (0:18:1_20:4) and PI (18:1_18:2) with PCOS, with mediating effects of − 0.003 (95% CI [− 0.017, 0.012]) and 0.003 (95% CI [− 0.021, 0.026]), respectively. Functional enrichment analysis revealed that PCOS was strongly associated with numerous immune pathways. Immune infiltration evaluation results indicated significant disparities in the distribution of a substantial number of immune cells, especially for all types of CD4+ T cells. Furthermore, Metabolic inference analysis revealed that fatty acid biosynthesis is closely related to PCOS.
Conclusion: Our results provide genetic and transcriptomic evidence substantiating the potential relationship between lipids, immune cells, and PCOS. CD4+ T cells might be key mediators in the causal association between lipids and PCOS. Comprehending this relationship could enhance treatment and prevention strategies for PCOS through the comprehensive management of lipid irregularities. Further experimental studies are required to elucidate the detailed mechanisms involved in this relationship.
Keywords: plasma lipid, immune cell, polycystic ovary syndrome, Mendelian randomization, transcriptomics analysis
Introduction
Lipids, a crucial class of biomolecules ubiquitously present in living organisms, exhibit diverse biological functions and play pivotal roles in energy metabolism, biofilm structure, cell signalling, and other physiological processes.1 Recently, a comprehensive longitudinal lipidomic analysis demonstrated a significant association between lipid profiles and the pathogenesis of acute and chronic inflammation, metabolic diseases, and aging.2
Polycystic ovary syndrome (PCOS) is a prevalent endocrine and metabolic disorder characterized by a spectrum of clinical manifestations including lipid metabolism disorders, dyslipidemia, insulin resistance, hyperandrogenemia, ovulatory dysfunction, oxidative stress, and chronic inflammation.3,4 Epidemiological studies indicate that PCOS affects approximately 5–18% of women of reproductive age globally, and infertility rates may reach 40%-74%.5,6 Beyond its reproductive implications, PCOS also causes diabetes, hyperlipidemia, cardiovascular diseases, and even ovarian and endometrial cancers, significantly impacting patients’ long-term health and wellbeing.7 Recent cross-sectional, cohort, and prospective studies have consistently demonstrated a strong correlation between lipid profiles, lipid metabolism, and PCOS in women of reproductive age.8–11 Observational studies have further elucidated that dysregulated lipid levels exacerbate metabolic risks in PCOS patients,12 contributing to insulin resistance (IR),13 hyperandrogenemia, and impaired glucose metabolism.14 Additionally, altered fatty acid composition in granulosa cells of PCOS patients has been found to lead to mitochondrial dysfunction and glucose metabolism disruption, which in turn impairs the developmental potential of oocytes.15,16
Thus far, the relationship between lipids and PCOS remains unclear, primarily because of limitations in sample sizes for large-scale cohort studies and comprehensive systematic evaluations. Additionally, the potential inverse effect of PCOS on lipid profiles has not been thoroughly investigated. Moreover, the underlying mechanisms by which lipids contribute to PCOS pathogenesis are yet to be fully elucidated. Previous studies have highlighted the critical role of lipids in maintaining immune homeostasis and regulating inflammatory responses.17,18 Furthermore, peripheral white blood cells, immune cells, pro-inflammatory cytokines, and chemokines have been implicated in the development and progression of PCOS.19,20 Based on the aforementioned facts, this study hypothesized that lipids are intricately associated with PCOS, with immune cells potentially serving as key mediating factors in this relationship.
Mendelian randomization (MR) is a method that leverages genetic variance as an instrumental variable to infer causal relationships between a disease and associated risk factors, effectively minimizing the bias arising from confounding factors or reverse causality inherent in observational studies.21 In this study, to explore the mechanisms by which lipids influence PCOS, we employed two-sample MR to reveal the causal effects of lipids on PCOS and two-step MR to identify immune cells mediating the link between lipids and PCOS. In addition, transcriptomic analysis was integrated to comprehensively explore the interplay between lipid metabolism, immune regulation, and PCOS.
Materials and Methods
Study Design
Single nucleotide polymorphisms (SNP) are a common form of variation in the genome that involve the alteration of a single nucleotide sequence. These variations contribute to the genetic diversity among individuals and are often associated with disease susceptibility, drug response, and other phenotypic traits.22 In this study, SNPs were used as validated instrumental variables (IVs) to examine the causal relationships between 179 plasma lipid groups and PCOS. To maximize the accuracy of the results, three important assumptions must always be confirmed. First, the selected IVs are associated with risk factors. Second, IVs is not associated with confounders between exposure and outcome. Third, IVs has no direct effect on outcome, but only affect outcome through exposure. We assessed the causal effects of the 179 plasma lipid groups on PCOS were assessed using the TSMR. Subsequently, immune cells demonstrating a strong correlation with both PCOS and plasma lipid groups were screened as potential mediators by a two-step MR. The first step estimated the lipid causal effect on immune cells. The second step was to calculate the causal effect of the immune cells on PCOS. Finally, we evaluated the mediating role of immune cells in the causal relationship between lipids and PCOS. Subsequently, transcriptomic analysis was performed to further explore the relationship between lipid metabolism, immunity, and PCOS (Figure 1).
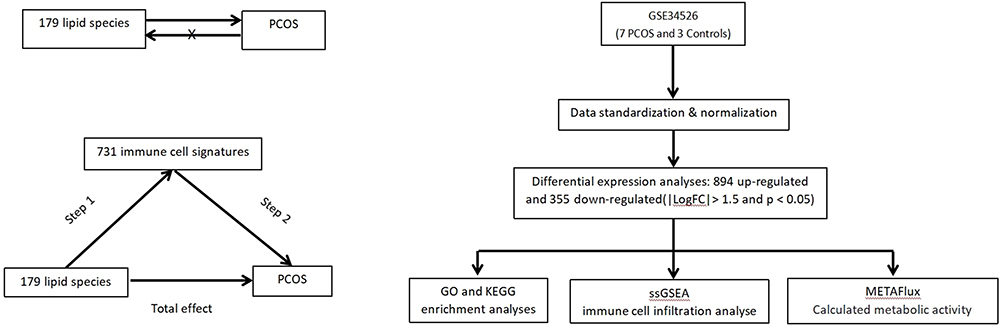 |
Figure 1 Study design. Abbreviations: PCOS, polycystic ovary syndrome; GO, gene ontology; KEGG, kyoto encyclopedia of genes and genomes; ssGSEA, single-sample gene set enrichment analysis. |
Data Sources
Lipid data were obtained by genome-wide association studies (GWAS) of 179 plasma lipid groups from 13 lipid classes in 7174 Finnish individuals.23 Four major lipid categories were covered: glycerophospholipids, glycerolipids, sphingolipids, and sterols. Glycerophospholipids included phosphatidylinositol (PI) (n=10), phosphatidylcholine (PC) (n=46), phosphatidylcholine ether (PCO) (n=27), phosphatidylethanolamine (PE) (n=5), and phosphatidylethanolamine ether (PEO) (n=8). The glycerolipids included triacylglycerol (TAG) (n=38) and diacylglycerol (DAG) (n=6). Sphingolipids included sphingomyelin (SM) (n=11) and ceramides (CER) (n=4). Sterols used included cholesteryl ester (CE) (n=15) and cholesterol (Chol) (n=1).
PCOS data from the FinnGen Consortium version R1024 included a total of 118,870 Europeans (642 cases and 118,228 controls). The diagnostic criteria for PCOS were based on the ICD-9 and ICD-10 criteria (the presence of two of the three criteria: chronic anovulation, hyperandrogenemia, and ultrasonography in the presence of polycystic ovaries).
Immune cell data were derived from 731 immune cell traits in a GWAS of a cohort of 3757 Sardinians. The summary statistics for the 731 immune cell traits disclosed in the GWAS catalog25 included 118 absolute cell counts, 389 mean fluorescence intensities (MFI) of surface antigens, 32 morphological parameters, and 192 relative counts (ratios between cellular levels). The MFI, absolute cell counts, and relative count characteristics included B cells, Conventional Dendritic cells (cDCs), T cell maturation stage, monocytes, myeloid cells, TBNK (T cells, B cells, natural killer cells), and Treg groups, and morphological parameter characteristics included cDC and TBNK groups. All data in this study were derived from research conducted on individuals of European descent to eliminate the potential biases associated with population stratification.
Genetic Instrumental Variable (IV) Selection Criteria
In this study, SNPs as IVs of PCOS were set at P<5e-8. Owing to the low number of eligible SNP loci, the threshold of IVs for plasma lipid groups and immune cell traits was relaxed to P<1e-5 after reading the relevant literature.26 To ensure the independence of each SNP, eligible SNPs were retained according to linkage disequilibrium (LD) conditions (R2=0.001, kb=10000). The outliers in IV were further cleaned using the outlier correction method in the Mendelian randomized multiple effectiveness residuals and outliers (MR-PRESSO) model. Simultaneously, all SNPs with palindromic structures were excluded, thus avoiding possible interference of alleles on the results. Finally, the F-statistic was used to measure the strength of the IV to avoid bias caused by weak IVs.
MR Analysis
In the TSMR analysis, five methods were used in this study: random effects inverse variance weighted (IVW), MR-egger, Weighted median, Simple mode, and Weighted mode. IVW produced reliable causal estimates in the absence of directed pleiotropy and was used as the primary method for estimating the causal relationship between exposure and outcome (P<0.05), suggesting a potential association. In addition, the Bonferroni correction method was applied to circumvent errors that may be introduced by multiple testing, and p-values that exceeded the Bonferroni corrected significance level (0.05/n, where n is the total number of tests) but remained below 0.05 were considered to be signals suggestive of causality. This study adds a Bayesian weighted Mendelian randomized (BWMR) method to ensure the accuracy of the results.
In the mediation MR analyses, we used the IVW approach to calculate the causal effect of plasma lipid groups on immune cells (BetaX), followed by the causal effect of immune cells on PCOS (BetaY). Finally, the coefficient product method was used to derive the mediating effect of the immune cells (BetaXY) and calculate its proportion (BetaXY = BetaX*BetaY).
Sensitivity Analysis
In the sensitivity analyses, the causal effects were analyzed using the heterogeneity test, multiple validity test, and leave-one-out method. Cochran’s Q test was used to calculate the level of heterogeneity, and MR-Egger regression and IVW methods were used to test the heterogeneity of the IVs, with P<0.05, indicating significant heterogeneity. Multiplicity was tested using MR-Egger intercept regression and P < 0.05 was considered significant multiplicity, and the study results were unreliable. The MR-PRESSO test was used to verify the presence of horizontal polyvalence. Sequential elimination of individual SNPs through “leave-one out”sensitivity analysis to sort out the potential impact of individual SNPs.
Transcriptomic Analyses
The PCOS gene expression profile dataset (GSE34526) was obtained from the GEO database. GSE34526 is based on the Affymetrix Human Genome U133 Plus 2.0 Array [HG-U133_Plus_2] platform and consists of 11 ovarian granulosa cell samples, of which seven were from patients with PCOS and three were from normovulatory women.27 Differential gene expression analysis was performed using the R package “limma”,28 with differentially expressed genes (DEGs) identified using the cut-off criteria of |Log2 fold change (LogFC)| > 1.5 and a p-value < 0.05. To further elucidate the biological functions of the DEGs, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses were conducted using the R package “clusterProfiler”, with a significance threshold of p <0.05.29
Single-sample gene set enrichment analysis (ssGSEA), an extension of the GSEA method, was employed to evaluated immune microenvironment by assess the enrichment degree of specific gene sets in each sample.30,31 Using the R package “GSVA”,32 enrichment scores were calculated for 28 tumor-infiltrating lymphocyte (TILs) gene signatures from the TISIDB database (http://cis.hku.hk/TISIDB/download.php) and two immune gene sets summarized by He et al.33 Finally, we employed the METAFlux algorithm to investigate the underlying mechanisms of metabolic abnormalities in PCOS.34 The Wilcoxon rank sum test was used to compare the differences between the two groups, with a p-value < 0.05 considered statistically significant.
Results
The Total Effect of 179 Plasma Lipid Groups on PCOS
Through TSMR combined with BWMR analysis, six plasma lipid groups were identified as potentially associated with PCOS, the F-statistics of all SNPs exceeded the threshold of 10. (The forest maps are shown in Figure 2A). Notably, PC (18:2_20:4) demonstrated a consistent positive causal effect on PCOS across multiple MR methods, including IVW, MR Egger and Weighted median, and the results were all significant (IVW, OR= 1.038, 95% CI[1.003, 1.073], p=0.032) (MR Egger, OR=1.110, 95% CI [1.020, 1.210], P=0.022) (Weighted median, OR= 1.054, 95% CI[1.002, 1.1008], P=0.040). No evidence of reverse causality was detected between the six plasma lipid groups and PCOS (Figure 2B). The sensitivity analyses further supported the robustness of these findings. No heterogeneity or horizontal pleiotropy was detected (Table 1), and no outlier SNPs were identified using MR-PRESSO. Scatter plots confirmed the stability of the causal relationships, and leave-one-out analysis showed that no single SNP disproportionately influenced effect estimates (Figure 2C). In summary, these findings highlight the significance of these six plasma lipid groups in PCOS development.
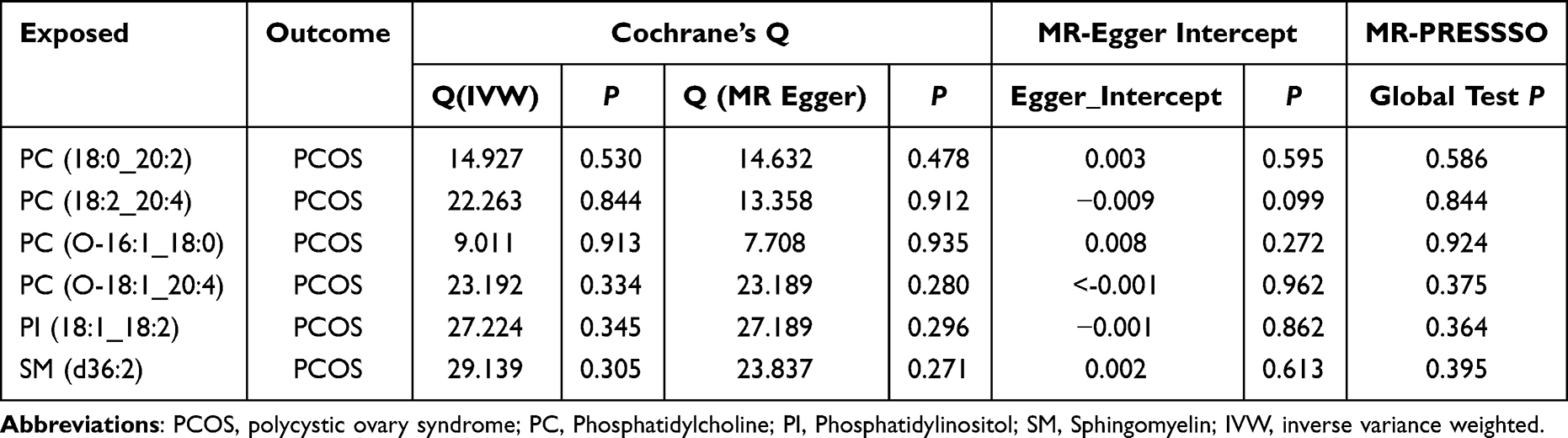 |
Table 1 Sensitivity Analysis Results |
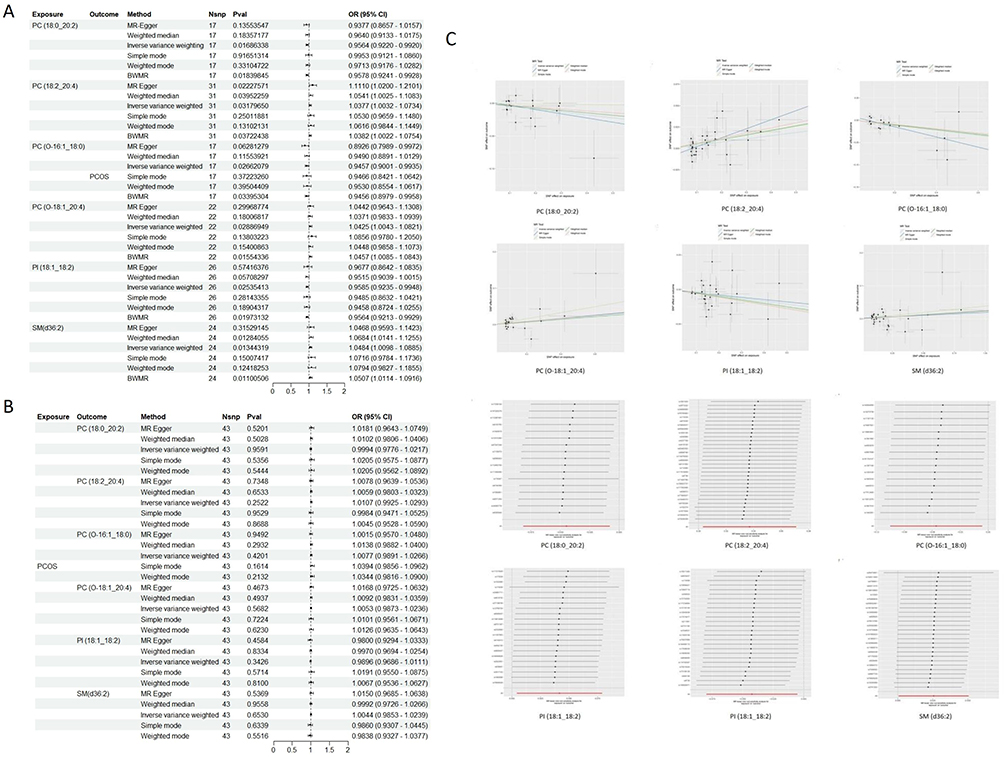 |
Figure 2 MR analyzed six lipids for causal association with PCOS. (A) TSMR and BWMR analysis revealed a causal association between 6 plasma lipid groups and PCOS. (B) Forest plot of the results of the reverse analysis. (C) Scatter plot and “leave-one out” of causal relationship between 6 plasma lipid groups and PCOS. Abbreviations: PCOS, polycystic ovary syndrome; SNP, single nucleotide polymorphism; CI, confidence interval; OR, odds ratio; PC, Phosphatidylcholine; PI, Phosphatidylinositol; SM, Sphingomyelin. |
Results of Two Step MR Analyses
TSMR analysis revealed significant correlations between 18 immune cell traits and PCOS (Figure 3A), including CD80 on CD62L+ myeloid DC (cDC panel), HLA DR on DC (cDC panel), CD33- HLA DR+ AC (Myeloid cell panel), Mo MDSC AC (Myeloid cell panel), CD33 on CD66b++ myeloid cell (Myeloid cell panel) CD33 on Mo MDSC (Myeloid cell panel), CD20- CD38- AC (B cell panel), CD19 on IgD+ CD24+ (B cell panel), CD19 on IgD+ CD38- unsw mem (B cell panel), Secreting Treg %CD4 (Treg panel), Activated & resting Treg AC (Treg panel), CD25hi CD45RA- CD4 not Treg %T cell (Treg panel), CD28+ CD45RA- CD8dim %T cell (Treg panel), CD28- CD25++ CD8br %T cell (Treg panel), CD127- CD8br AC (Treg panel), CD28 on CD28+ CD4+ (Treg panel), CD25 on CD39+ secreting Treg (Treg panel), CD25 on CD39+ CD4+ (Treg panel), CD45RA- CD4+ %CD4+ (Maturation stages of T cell panel), Naive CD4+ %CD4+ (Maturation stages of T cell panel), TD DN (CD4-CD8-) %DN (Maturation stages of T cell panel), CD45 on NK (TBNK panel), FSC-A on HLA DR+ CD8br (TBNK panel), SSC-A on CD4+ (TBNK panel) and HLA DR on B cell (TBNK panel). Clinical studies have demonstrated a close relationship between PCOS and immune dysregulation, particularly involving CD4+ T and NK cells in the peripheral blood of affected women.35 Furthermore, evidence suggests that the MFI of human leukocyte antigen DR (HLA-DR) expressed on dendritic cells (DCs) positively correlates with the ovarian response. Reduced DC abundance in the follicular fluid of PCOS patients disrupts the ovarian immune microenvironment, leading to impaired follicular formation.36 These findings underscore the potential role of immune cells in the pathogenesis of PCOS.
Figure 3 Continued. Figure 3 Two-step MR analysis of forest graphs. (A) Forest plot of causal relationship between immune cells and PCOS. (B) Forest plot of causal relationship between plasma lipid groups and immune cells. Abbreviations: PCOS, polycystic ovary syndrome; SNP, single nucleotide polymorphism; CI, confidence interval; OR, odds ratio; PC, Phosphatidylcholine; PI, Phosphatidylinositol; SM, Sphingomyelin.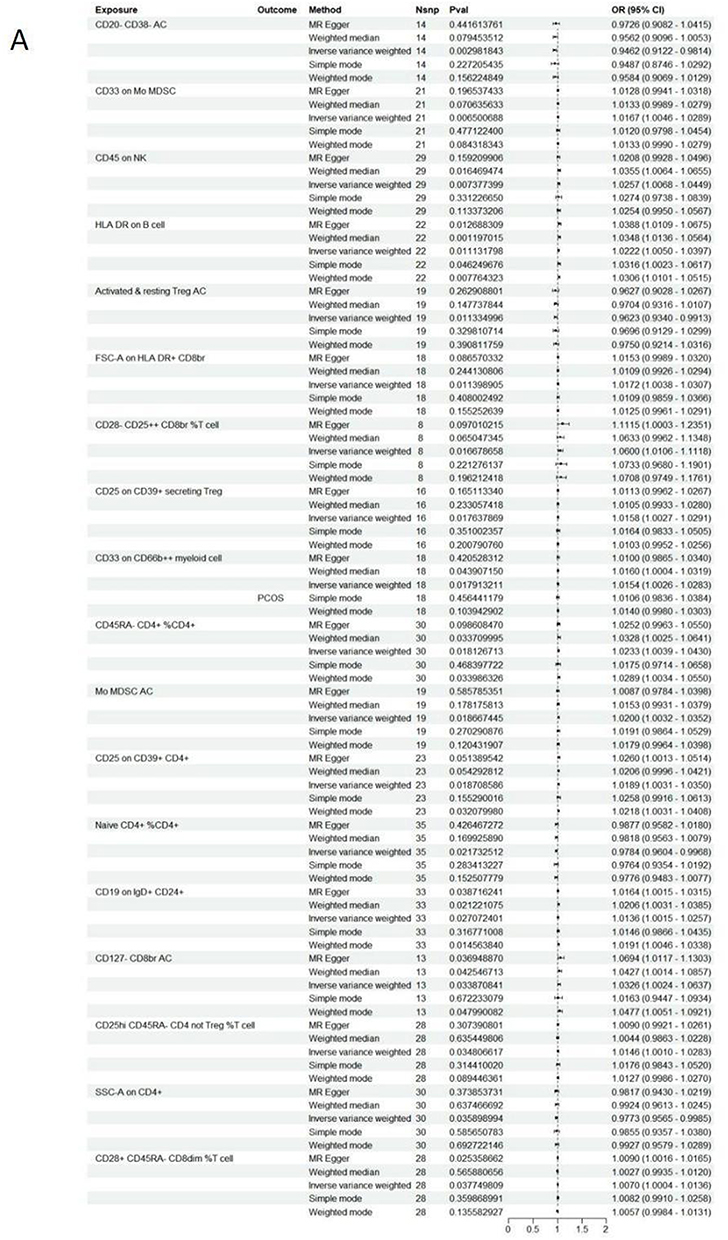
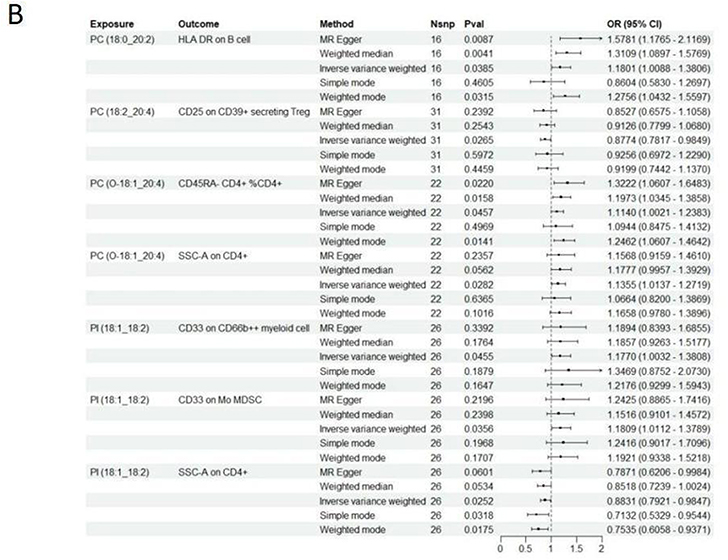
In general, we identified 6 lipids and 18 immune cell traits that are strongly associated with PCOS. However, it is uncertain whether lipids affect PCOS through immune cell mediation. We performed mediation analysis and found that, among all immune cell traits, CD4+ T cells exhibited the largest mediating proportionality and were most closely associated with the causal effects of lipids on PCOS (Figure 3B). Specifically, SSC-A on CD4+ cells emerged as a key mediator linking PC (O-18:1_20:4) to PCOS (mediating effect = 0.003 (95% CI [−0.021, 0.026]). SSC-A on CD4+ also mediated the causal relationship between PI (18:1_18:2) and PCOS, with a mediation effect of −0.003.
DEGs Identification and Functional Enrichment Analysis
We collected GSE34526 gene expression profiles, including seven PCOS patients and three controls. Differential expression analysis identified 1249 DEGs, comprising 894 upregulated and 355 downregulated genes. Visualization of DEGs was achieved using a heatmap (top 30) and volcano plots (Figure 4A). Subsequently, GO and KEGG enrichment analyses were performed on the up-regulated genes (Figure 4B). The GO enrichment results revealed significant enrichment of biological processes (BP) related to the regulation of immune effector processes and leukocyte-mediated immunity. In the cellular component (CC) category, the most significantly enriched terms were secretory granule membrane and outer plasma membrane. For molecular function (MF), Actin binding was the most enriched term for molecular function. Pathway enrichment in the KEGG database primarily involved immune-related pathways, including cytokine and receptor interactions, chemokine signalling pathways, and other immune-related pathways. These findings suggest that the pathology of PCOS is closely linked to immune dysregulation.
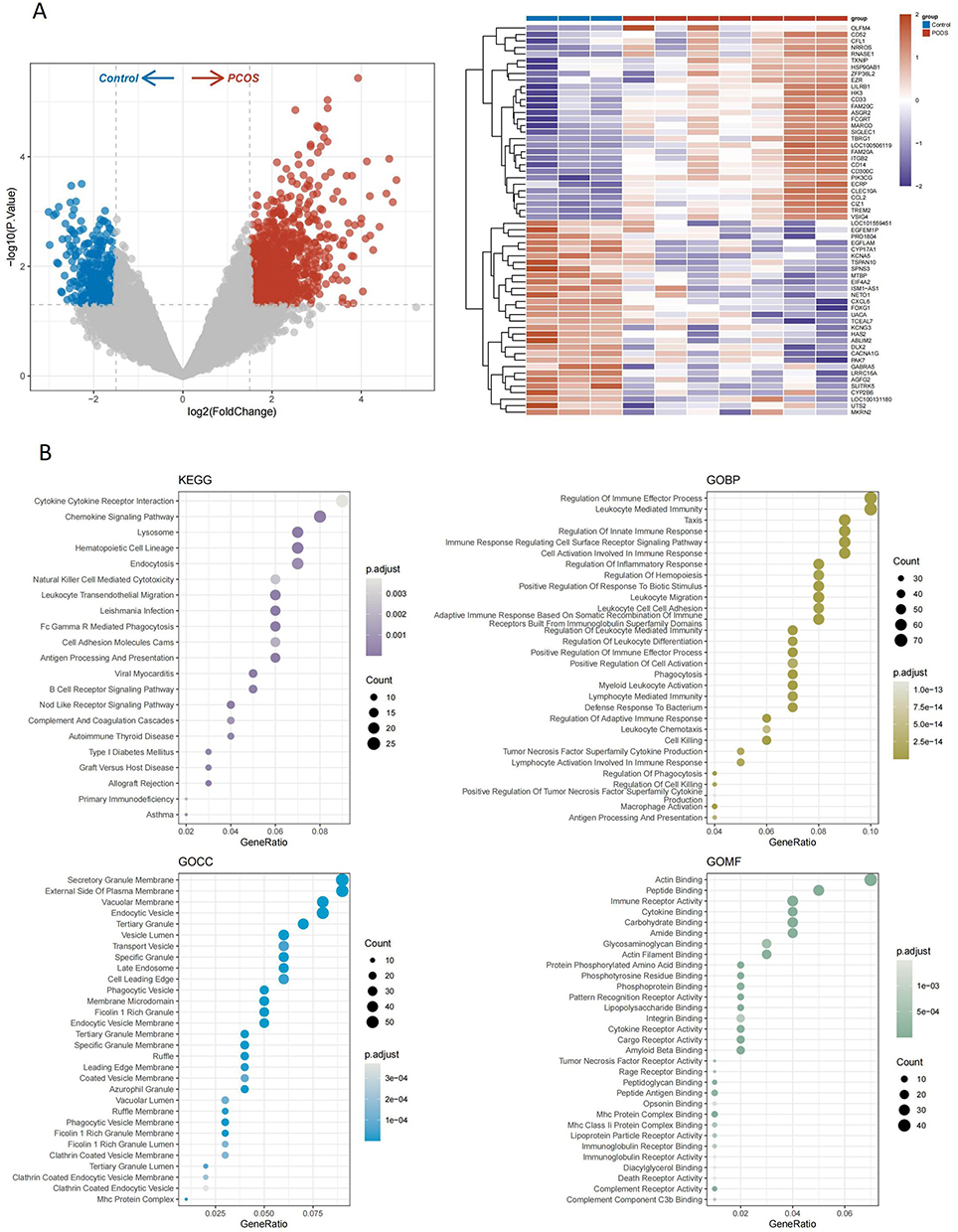 |
Figure 4 Gene Identification and Functional Enrichment Analysis Results. (A) Heat map and volcano map on GSE34526. (B) 10 Bubble plots of GO results and KEGG results for PCOS-expressed genes. Abbreviation: PCOS, polycystic ovary syndrome. |
Immune Microenvironment Analysis and Metabolic Inference
Single-sample gene set enrichment analysis (ssGSEA) was used to quantify immune microenvironment traits by scoring corresponding genesets. By scoring 28 immune cell signatures obtained from the TISIDB database, we found that there were significant differences in the immune infiltration levels of 15 immune cell types between the normal and PCOS groups (Figure 5A). Specifically, patients with PCOS exhibit elevated levels of T-cell infiltration, including central memory CD4 T cells, compared to controls. To ensure the robustness and reproducibility of our findings, we incorporated an additional immune-related gene set curated by He et al. The results of the 16 immune cell type enrichment scores also showed significant differences in the levels of B cells, Macrophages, NK cells, T helper cells, and TIL immune infiltration (Figure 5B). Additionally, 13 immune cell function enrichment scores revealed significant differences in the levels of APC co-stimulation, CCR, checkpoint, HLA, inflammation-promoting, parainflammation, and T-cell co-inhibition were significantly different (Figure 5C). These results suggest that the immune microenvironment is altered in patients with PCOS. Notably, co-stimulatory signals between APCs, such as macrophages, dendritic cells, and T cells (CD80/CD86), have been shown to promote differentiation of Th1 and Th17 cells, release cytokines and chemokines (eg, IL-6, IL-10, IL-18, MIF, and TNF-α) into the bloodstream, and exacerbate insulin resistance and localized inflammation of the ovaries.37 Chronic inflammation is thus considered a key underlying factor contributing to ovarian dysfunction in PCOS.
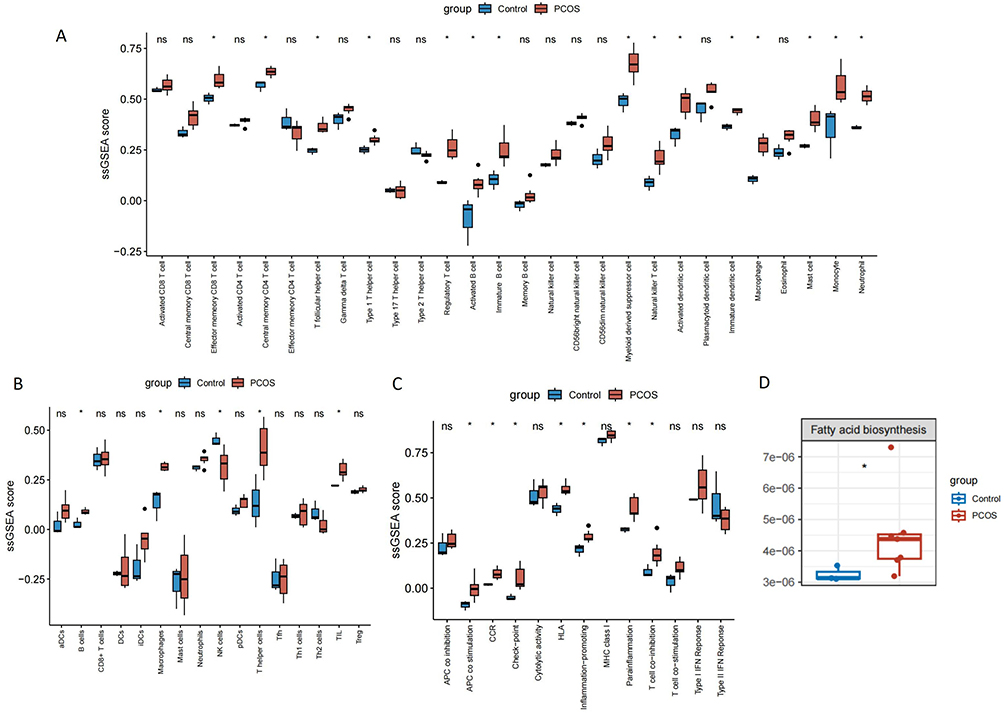 |
Figure 5 PCOS is associated with immune infiltration. (A) Comparison of levels of infiltration of 28 immune cells. (B) Comparison of levels of infiltration of 16 immune cells. (C) Comparison of levels of infiltration of 13 immune cells function (red for PCOS, blue for control, *p < 0.05). (D) The boxplot showed that the biosynthesis of relevant fatty acids was significant between PCOS and control group. Abbreviation: PCOS, polycystic ovary syndrome. |
Finally, to explore whether there were abnormalities in lipid metabolism in PCOS, we used METAflux for metabolic inference. The results showed that the fatty acid biosynthesis activity of PCOS group was significantly higher than that of the normal control group (Figure 5D). Fatty acid biosynthesis is a key process in cellular energy metabolism, wherein fatty acids are synthesized via the fatty acid synthase (FAS) pathway and serve as precursors for the synthesis of phospholipids, such as PI and PC.38
Research has demonstrated that Disruptions in the fatty acid synthesis pathway, particularly in the context of insulin resistance and elevated androgen levels, may lead to an imbalance in the synthesis of PI and PC. This imbalance can interfere with ovarian function and ovulation, thereby promoting the development and progression of PCOS.39 Consequently, targeting the regulation of fatty acid metabolism, PI, and PC may offer novel therapeutic strategies for PCOS management.
Discussion
Ożegowska, and Li et al found that PC and SM were significantly elevated in women with PCOS by serum metabolomics.39,40 In this study, we identified six plasma lipid groups that affect PCOS: PC (18:0_20:2), PC (18:2_20:4), PC (O-16:1_18:0), PC (O-18:1_20:4), PI (18:1_18:2), and SM (d36:2). Our findings are consistent with previous studies. Using a Two-step MR approach, we investigated the role of immune cells in the relationship between these lipids and PCOS. In the first step, 18 immune cells were strongly associated with PCOS. In the second step, MR Analysis showed that SSC-A on CD4+ cells mediated the causal relationship between PC (O:18:1_20:4) and PI (18:1_18:2) and PCOS. The two-step effects of PC were in the same direction and had a clear positive relationship with PCOS.
PC is a critical component of cell membranes, plays a pivotal role in intracellular and intercellular signal transduction, and participates in various physiological processes. During lipid metabolism, PC acts as a carrier for lipid transport, facilitating the movement of lipids throughout the body.41 Arachidonic acid (AA), contained in PC (O:18:1_20:4), is integral to numerous physiological processes, particularly inflammatory responses. AA is metabolized into prostaglandins and leukotrienes through the cyclooxygenase-2 (COX-2) and lipoxygenase (LOX) pathways, which can exacerbate ovarian inflammation and impair normal follicular development, leading to ovulation disorders.42 Additionally, AA and its specific metabolite 5-hydroxyeicosatetraenoic acid (5-HETE) can regulate androgen metabolism and increase androgen secretion, which in turn exacerbates the symptoms of PCOS, such as irregular menstruation and hirsutism.43 IR is a hallmark of PCOS. As an essential component of the cell membrane, PC (O-18:1_20:4) influences the regulation of the insulin signaling pathway. Abnormalities in PC metabolism can disrupt cell membrane fluidity and receptor function, particularly insulin receptors, leading to impaired insulin signaling and exacerbation of IR.43 PCOS is usually accompanied by chronic low-grade inflammation, and infiltration of CD4+ T cells is a potential source of this inflammatory state.35 In this study, transcriptome analysis revealed a significantly elevated infiltration of T cells in patients with PCOS. Cytokines secreted by T cells (IFN-γ, IL-1, etc.) impede normal follicular development and ovulation by exacerbating the inflammatory response in the ovary.44
Fatty acid biosynthesis directly affects the fatty acid composition of PC and PI through the synthesis of fatty acids such as OA, AA, and LA. The fatty acid chains of PC (O:18:1_20:4) and PI (18:1_18:2) are derived from desaturation and lengthening reactions in the fatty acid synthesis pathway. Fatty acid biosynthesis plays a critical role in the development and progression of PCOS, as abnormal fatty acid metabolism is central to the pathogenesis of the disorder by contributing to IR, chronic inflammation, hyperandrogenism, and ovarian dysfunction.45,46 Consequently, regulation of fatty acid synthesis and metabolism may represent an effective therapeutic strategy to alleviate PCOS symptoms.
PCOS is often accompanied by obesity or weight gain, especially with the accumulation of abdominal fat. In this context, CD4+ T cells may promote an immune response in adipose tissue, increase the level of inflammation in the body, and further aggravate the metabolic disorder of PCOS.47 Moreover, the increase and activation of CD4+ T cells in the adipose tissue of obese mice directly affects the insulin signaling pathway, leading to the development of IR.48 CD4+ T cells are involved in the development of PCOS through multiple pathways, especially the inflammatory response, insulin resistance, immune dysregulation, and ovarian function. Regulating the function of immune cells and restoring the balance of the immune system may be a new strategy for treating PCOS.
Conclusions
In this study, we comprehensively investigated the relationship between lipids, immune cells, and PCOS using genetic and transcriptomic analyses. MR analysis revealed that SSC-A on CD4+-mediated PC (O-18:1_20:4) and PI (18:1_18:2) is causally associated with PCOS. Transcriptomic analyses have indicated that fatty acid biosynthesis and CD4+ T cell infiltration exert significant effects on PCOS. However, due to the small sample size of the transcriptomic dataset, its power and generalizability are limited. While this study provides novel insights into PCOS mechanisms, potential diagnostic biomarkers, and therapeutic targets, these findings remain hypothetical at present. It is necessary to clarify the specific mechanisms and interrelationships through cellular experiments, animal model studies, and analyses of clinical samples in the future. In subsequent studies, we will employ integrated lipidomics and immunomics approaches to enable precise stratification of PCOS subtypes.
Data Sharing Statement
The data can be found in public open-access repositories. Data URLs GWAS summary statistics for 179 plasma lipid groups and 731 immune traits are available for download from the GWAS catalogue (study logins: GCST90277238-GCST90277416, GCST90001001-GCST90002000 https://www.ebi.ac.uk/gwas/home). The summary statistics for PCOS are available at https://gwas.mrcieu.ac.uk/datasets/finn-b-E4_POCS/. The GSE34526 dataset was downloaded from https://www.ncbi.nlm.nih.gov/gds/. All codes used in this study are available upon request from the corresponding author.
Ethics Approval and Consent to Participate
The data used in this study were obtained from anonymized public databases and were fully compliant with the exemption criteria specified in Article 32, item 1 and 2 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (issued by the National Health Commission of China, effective February 18, 2023). Consequently, our study does not require additional informed consent or ethical review.
Acknowledgments
The authors of this paper are very grateful to the GWAS, FinnGen, and GEO databases for data support, and the Chengdu Science and Technology Bureau for financial support.
Funding
This study was supported by the Chengdu Science and Technology Bureau(2021-YF05-02042-SN).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Drew D, Boudker O. Ion and lipid orchestration of secondary active transport. Nature. 2024;626(8001):963–974. doi:10.1038/s41586-024-07062-3
2. Hornburg D, Wu S, Moqri M, et al. Dynamic lipidome alterations associated with human health, disease and ageing. Nat Metab. 2023;5(9):1578–1594. doi:10.1038/s42255-023-00880-1
3. Qian Y, Tong Y, Zeng Y, et al. Integrated lipid metabolomics and proteomics analysis reveal the pathogenesis of polycystic ovary syndrome. J Transl Med. 2024;22(1):364. doi:10.1186/s12967-024-05167-x
4. Pereira-Eshraghi CF, Vuguin PP. Polycystic ovary syndrome. Pediatr Rev June. 2024;45(6):363–365. doi:10.1542/pir.2023-006036
5. Joham AE, Norman RJ, Stener-Victorin E, et al. Polycystic ovary syndrome [published correction appears in lancet diabetes endocrinol. 2022;10(11):e11. doi: 10.1016/S2213-8587(22)00281-9]. Lancet Diabetes Endocrinol. 2022;10(9):668–680. doi:10.1016/S2213-8587(22)00163-2
6. Liu X, Zhang J, Wang S. Global, regional, and national burden of infertility attributable to PCOS, 1990-2019. Hum Reprod. 2024;39(1):108–118. doi:10.1093/humrep/dead241
7. Stener-Victorin E, Teede H, Norman RJ, et al. Polycystic ovary syndrome. Nat Rev Dis Primers. 2024;10(1):27. doi:10.1038/s41572-024-00511-3
8. Shafiee MN, Ortori CA, Barrett DA, Mongan NP, Abu J, Atiomo W. Lipidomic biomarkers in polycystic ovary syndrome and endometrial cancer. Int J Mol Sci. 2020;21(13):4753. doi:10.3390/ijms21134753
9. Hussein SRM, Sadiq AM, Johar SA, Nasrawi AJM. Insulin level, lipid profile, and HOMA index in lean and obese patients with polycystic ovary syndrome. J Med Life. 2023;16(8):1258–1263. doi:10.25122/jml-2023-0040
10. Parveen S, Khan S, Khan MM, Gupta B, Ahmad A, Alam R. Association of lipid profile and obesity in patients with polycystic ovary syndrome. Endocr Regul. 2024;58(1):83–90. doi:10.2478/enr-2024-0009
11. Pan X. Metabolic characteristics of obese patients with polycystic ovarian syndrome: a meta-analysis. Gynecol Endocrinol. 2023;39(1):2239934. doi:10.1080/09513590.2023.2239934
12. Mousa A, Huynh K, Ellery SJ, et al. Novel lipidomic signature associated with metabolic risk in women with and without polycystic ovary syndrome. J Clin Endocrinol Metab. 2022;107(5):e1987–e1999. doi:10.1210/clinem/dgab931
13. Kheirollahi A, Teimouri M, Karimi M, et al. Evaluation of lipid ratios and triglyceride-glucose index as risk markers of insulin resistance in Iranian polycystic ovary syndrome women. Lipids Health Dis. 2020;19(1):235. doi:10.1186/s12944-020-01410-8
14. Guo F, Gong Z, Fernando T, Zhang L, Zhu X, Shi Y. The lipid profiles in different characteristics of women with PCOS and the interaction between dyslipidemia and metabolic disorder states: a retrospective study in Chinese population. Front Endocrinol. 2022;13:892125. doi:10.3389/fendo.2022.892125
15. Tian Y, Wang A, Li F, et al. Fatty acid metabolism of granulose cells and follicular fluid and their associations with embryo quality in normal-weight women with PCOS. J Ovarian Res. 2025;18(1):130. doi:10.1186/s13048-025-01711-5
16. Yan H, Wang L, Zhang G, et al. Oxidative stress and energy metabolism abnormalities in polycystic ovary syndrome: from mechanisms to therapeutic strategies. Reprod Biol Endocrinol. 2024;22(1):159. doi:10.1186/s12958-024-01337-0
17. Leuti A, Fazio D, Fava M, Piccoli A, Oddi S, Maccarrone M. Bioactive lipids, inflammation and chronic diseases. Adv Drug Deliv Rev. 2020;159:133–169. doi:10.1016/j.addr.2020.06.028
18. Radzioch D, Giera M, De Sanctis JB. Editorial: quo vadis lipid mediators - lipid mediators implication in inflammation and chronic inflammatory diseases. Front Immunol. 2021;12:699276. doi:10.3389/fimmu.2021.699276
19. Wang J, Yin T, Liu S. Dysregulation of immune response in PCOS organ system. Front Immunol. 2023;14:1169232. doi:10.3389/fimmu.2023.1169232
20. Rudnicka E, Suchta K, Grymowicz M, et al. Chronic low grade inflammation in pathogenesis of PCOS. Int J Mol Sci. 2021;22(7):3789. doi:10.3390/ijms22073789
21. Birney E. Mendelian Randomization. Cold Spring Harb Perspect Med. 2022;12(4):a041302. doi:10.1101/cshperspect.a041302
22. Wu K, Kong F, Zhang J, et al. Recent progress in single-nucleotide polymorphism biosensors. Biosensors. 2023;13(9):864. doi:10.3390/bios13090864
23. Ottensmann L, Tabassum R, Ruotsalainen SE, et al. Genome-wide association analysis of plasma lipidome identifies 495 genetic associations. Nat Commun. 2023;14(1):6934. doi:10.1038/s41467-023-42532-8
24. The FinnGen consortium finngen documentation of R11 release. 2024. Available from: https://finngen.gitbook.io/documentation/.
25. Orrù V, Steri M, Sidore C, et al. Author Correction: complex genetic signatures in immune cells underlie autoimmunity and inform therapy. Nat Genet. 2020;52(11):1266. doi:10.1038/s41588-020-00718-6
26. Chen J, Yu X, Wu X, Chai K, Wang S. Causal relationships between gut microbiota, immune cell, and Non-small cell lung cancer: a two-step, two-sample Mendelian randomization study. J Cancer. 2024;15(7):1890–1897. doi:10.7150/jca.92699
27. Kaur S, Archer KJ, Devi MG, Kriplani A, Strauss JF, Singh R. Differential gene expression in granulosa cells from polycystic ovary syndrome patients with and without insulin resistance: identification of susceptibility gene sets through network analysis. J Clin Endocrinol Metab. 2012;97(10):E2016–21. PMID: 22904171; PMCID: PMC3674289. doi:10.1210/jc.2011-3441
28. Ritchie ME, Phipson B, Wu D, et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015;43(7):e47. doi:10.1093/nar/gkv007
29. Xu S, Hu E, Cai Y, et al. Using clusterProfiler to characterize multiomics data. Nat Protoc. 2024;19(11):3292–3320. doi:10.1038/s41596-024-01020-z
30. Qin Y, Liu Y, Xiang X, et al. Cuproptosis correlates with immunosuppressive tumor microenvironment based on pan-cancer multiomics and single-cell sequencing analysis. Mol Cancer. 2023;22(1):59. doi:10.1186/s12943-023-01752-8
31. Liu S, Zhao X, Meng Q, Li B. Screening of potential biomarkers for polycystic ovary syndrome and identification of expression and immune characteristics. PLoS One. 2023;18(10):e0293447. doi:10.1371/journal.pone.0293447
32. Wang W, Li T, Xie Z, et al. Integrating single-cell and bulk RNA sequencing data unveils antigen presentation and process-related CAFS and establishes a predictive signature in prostate cancer. J Transl Med. 2024;22(1):57. doi:10.1186/s12967-023-04807-y
33. He Y, Jiang Z, Chen C, Wang X. Classification of triple-negative breast cancers based on Immunogenomic profiling. J Exp Clin Cancer Res. 2018;37(1):327. PMID: 30594216; PMCID: PMC6310928. doi:10.1186/s13046-018-1002-1
34. Huang Y, Mohanty V, Dede M, et al. Characterizing cancer metabolism from bulk and single-cell RNA-seq data using METAFlux. Nat Commun. 2023;14(1):4883. doi:10.1038/s41467-023-40457-w
35. He S, Mao X, Lei H, et al. Peripheral blood inflammatory-immune cells as a predictor of infertility in women with polycystic ovary syndrome. J Inflamm Res. 2020;13:441–450. doi:10.2147/JIR.S260770
36. Zhang T, Tian F, Huo R, Tang A, Zeng Y, Duan YG. Detection of dendritic cells and related cytokines in follicular fluid of patients with polycystic ovary syndrome. Am J Reprod Immunol. 2017;78(3):
37. Luan YY, Zhang L, Peng YQ, Li YY, Liu RX, Yin CH. Immune regulation in polycystic ovary syndrome. Clin Chim Acta. 2022;531:265–272. doi:10.1016/j.cca.2022.04.234
38. De Carvalho CCCR, Caramujo MJ. The various roles of fatty acids. Molecules. 2018;23(10):2583. doi:10.3390/molecules23102583
39. Li S, Chu Q, Ma J, et al. Discovery of novel lipid profiles in PCOS: do insulin and androgen oppositely regulate bioactive lipid production? J Clin Endocrinol Metab. 2017;102(3):810–821. doi:10.1210/jc.2016-2692
40. Ożegowska K, Plewa S, Mantaj U, Pawelczyk L, Matysiak J. Serum metabolomics in PCOS women with different body mass index. J Clin Med. 2021;10(13):2811. doi:10.3390/jcm10132811
41. Furse S, de Kroon AI. Phosphatidylcholine’s functions beyond that of a membrane brick. Mol Member Biol. 2015;32(4):117–119. doi:10.3109/09687688.2015.1066894
42. Szczuko M, Kikut J, Komorniak N, Bilicki J, Celewicz Z, Ziętek M. The role of arachidonic and linoleic acid derivatives in pathological pregnancies and the human reproduction process. Int J Mol Sci. 2020;21(24):9628. doi:10.3390/ijms21249628
43. Lin W-L, Chien -M-M, Patchara S, et al. Essential trace element and phosphatidylcholine remodeling: implications for body composition and insulin resistance. J Trace Elements Med Biol. 2024;85:127479. doi:10.1016/j.jtemb.2024.127479
44. Zhou H, Xu J, Hong L, et al. The alterations of circulating mucosal-associated invariant T cells in polycystic ovary syndrome. Front Endocrinol. 2022;13:1038184. doi:10.3389/fendo.2022.1038184
45. Yue X, Zhou Z, Zhang G, et al. Metabolome implies increased fatty acid utilization and histone methylation in the follicles from hyperandrogenic PCOS women. J Nutr Biochem. 2024;125:109548. doi:10.1016/j.jnutbio.2023.109548
46. Elkanawati RY, Sumiwi S, Levita J, et al. Impact of lipids on insulin resistance: insights from human and animal studies. Drug Des Devel Ther. 2024;18:3337–3360. doi:10.2147/DDDT.S468147
47. Shirakawa K, Sano M. Drastic transformation of visceral adipose tissue and peripheral CD4 T cells in obesity. Front Immunol. 2023;13:1044737. doi:10.3389/fimmu.2022.1044737
48. Bensussen A, Torres-Magallanes JA, Roces de Álvarez-Buylla E, et al. Molecular tracking of insulin resistance and inflammation development on visceral adipose tissue. Front Immunol. 2023;14:1014778. doi:10.3389/fimmu.2023.1014778
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.