Back to Journals » Journal of Asthma and Allergy » Volume 18
Mechanism of miR-206 in Regulating KLF4 and Affecting Th17 Cell Differentiation in Neutrophil Asthma Inflammation
Authors Feng L ![]() , Li Y, Zhang L
, Li Y, Zhang L ![]() , Xing Y, Huo R, Cheng E, Tian X
, Xing Y, Huo R, Cheng E, Tian X ![]()
Received 1 May 2025
Accepted for publication 22 October 2025
Published 4 November 2025 Volume 2025:18 Pages 1493—1504
DOI https://doi.org/10.2147/JAA.S537623
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Luis Garcia-Marcos
Liting Feng, Yingna Li, Lulu Zhang, Yanqing Xing, Rujie Huo, Erjing Cheng, Xinrui Tian
Department of Gerontology, The Second Hospital of Shanxi Medical University, Taiyuan, Shanxi, People’s Republic of China
Correspondence: Xinrui Tian, Department of Gerontology, The Second Hospital of Shanxi Medical University, Taiyuan, Shanxi, People’s Republic of China, Tel +8613834575570, Email [email protected]
Purpose: This study explores the role of miR-206 and KLF4 in neutrophilic asthma, focusing on their interaction and the molecular mechanisms by which miR-206 regulates Th17 cell-mediated immune-inflammatory responses through KLF4.
Methods: RT-qPCR and flow cytometry assessed miR-206, KLF4 mRNA, and Th17 cell levels in asthma patients and healthy controls. CD4+ T cells were transfected to overexpress or inhibit miR-206, and KLF4, IL-17, and Th17 cell ratios were analyzed using RT-qPCR, ELISA, and flow cytometry. Correlation analysis evaluated relationships between miR-206, KLF4, and Th17 cells.
Results: Severe asthma patients showed reduced miR-206 and elevated KLF4 mRNA levels, with increased Th17 cells and IL-17. miR-206 downregulated KLF4, negatively correlating with KLF4, Th17 cells, and IL-17. KLF4 positively correlated with Th17 cells and IL-17. miR-206 suppressed Th17 differentiation and IL-17 production by modulating KLF4.
Conclusion: miR-206 is downregulated in acute asthma and targets KLF4, inhibiting Th17 cell differentiation and IL-17 production. These findings highlight miR-206 as a potential therapeutic target for asthma through its anti-inflammatory effects mediated by KLF4 and Th17 regulation.
Keywords: asthma, miR-206, KLF4, Th17, IL-17
Introduction
Bronchial asthma is a chronic inflammatory disorder of the airways characterized by multifaceted cellular involvement, resulting in airway dysfunction, including airway hyperresponsiveness (AHR) and reversible airflow limitation.1,2 Asthma impacts almost 300 million individuals globally, with both prevalence and mortality rates rising year. A recent national cross-sectional survey indicated that the prevalence of asthma among individuals aged 20 years and older in China was 4.2%, equating to approximately 45.7 million persons affected, significantly impacting their quality of life.3 Despite current treatments managing asthma symptoms to some degree, certain patients continue to experience inadequate symptom management and frequent exacerbations due to the heterogeneity of asthma and the unpredictability in patients’ responses to available drugs.4,5 Consequently, a comprehensive investigation into the pathophysiology of asthma and the identification of novel treatment targets hold substantial clinical importance.
Asthma has conventionally been seen as a chronic inflammatory illness of the airways, principally defined by inflammation mediated by T helper 2 (Th2) cells.6 Increasing evidence indicates that Th17 cells, a subpopulation of helper T cells with a distinct development pathway, can secrete cytokines such as IL-17A/F, which are implicated in the immunological response associated with asthma.7 In asthma patients, the expression levels of Th17 cells and their released cytokines IL-17A/F are elevated and significantly linked with asthma severity.8,9 A study involving pediatric patients experiencing acute asthma exacerbation revealed elevated percentages of Th17 cells in peripheral Th cells, increased amounts of IL-17 in sputum, and heightened concentrations of IL-8 and IL-17 in the culture supernatant of PBMCs, all of which were positively linked with sputum neutrophil levels.8 Sorbello noted that asthmatic patients exhibited increased levels of IL-17 in the nasal and bronchial cavities, with IL-17 levels positively correlating with asthma severity and airway neutrophilia.9 These findings suggest that the Th17/IL-17 pathway plays a key role in neutrophilic asthma, which is completely different from the inflammatory mechanism of Th2 type asthma.
There is still a significant clinical unmet need for the Th17 pathway in the current asthma treatment field. Neutrophil asthma patients generally exhibit resistance to glucocorticoid therapy, while interventions targeting a single target, such as IL-17RA, have limited clinical efficacy. This suggests the need to develop multi-target combination therapy strategies. In clinical practice, there is still a dilemma of lacking specific biomarkers, which makes it difficult to accurately identify Th17 related asthma phenotypes and implement precise treatment. More clinical evidence is needed to support the safety and effectiveness of innovative treatment methods, including mesenchymal stem cell therapy and vitamin D regulation. In addition, how to effectively intervene in risk factors closely related to Th17 asthma, such as smoking and obesity, is also an important challenge in current clinical management.10 Targeted regulation of Th17 cell differentiation and function may provide a new therapeutic strategy for the specific subtype of neutrophilic asthma.
MicroRNAs (miRNAs) are a category of endogenous non-coding RNAs, ranging from 20 to 24 nucleotides in length, that are expressed post-transcriptionally in eukaryotes by interacting with mRNAs to regulate gene expression.11 MiR-206, as a member of the small RNA family, has shown important regulatory roles and potential as a biomarker in various inflammatory and non inflammatory diseases. The dysregulation of miR-206 is closely related to various skeletal and muscle diseases. As a soft tissue sarcoma, rhabdomyosarcoma (RMS) can be diagnosed with miR-206 as a biomarker. Clinical trials have shown that miR-206 levels in RMS patients are significantly higher than those in healthy controls, and have been identified as a diagnostic biomarker. MiR-206 also serves as a potential biomarker for disease progression. MiR-206 levels are also elevated in patients with amyotrophic lateral sclerosis (ALS) and Alzheimer’s disease. In addition, miR-206 is also associated with scleroderma, hepatic steatosis, chronic obstructive pulmonary disease, and cardiomyopathy. Therefore, miR-206 is a therapeutic and diagnostic biomarker for many diseases.12 In respiratory inflammatory diseases, it has been proven to target asthma related genes and serve as a biomarker for the disease,13 while also being used to predict individual disease status.14 Its ability to distinguish acute asthma attacks even surpasses traditional clinical scoring systems.15 In experimental studies, the expression of miR-206 was reduced in OVA induced mice, while lncRNA RMRP, as the sponge of miR-206, showed increased expression. By regulating the TGF - β/Smad2 signaling pathway, lncRNA RMRP promotes inflammation and fibrosis, thereby affecting the occurrence of childhood asthma.16 MiR-206 is positioned upstream of the IL-17 A/F gene locus and is capable of regulating IL-17 expression.17 Reports indicate that the expression level of miR-206 is diminished in peripheral blood mononuclear cells (PBMCs) of patients with dermatomyositis.Conversely, IL-17 mRNA level was elevated, and miR-206 expression exhibited a negative correlation with Th17 cells.18 These data indicate that miR-206 may play a role in neutrophilic asthma development by modulating Th17 cell activity. The regulatory function of miR-206 in Th17 cell development and activity in asthma remains unidentified.
Krüppel-like factor 4 (KLF4) is a gene that encodes a transcriptional repressor and is a target of miR-206. Overexpression of miR-206 suppresses the translation of KLF4, while a reduction in miR-206 correlates with an elevation in KLF4 expression.19,20 KLF4 has been shown to activate Th17 cell differentiation and directly regulate IL-17A expression,21,22 with a favorable correlation between Th17 cell expression and KLF4 levels.18 KLF4 can have both pro-inflammatory and anti-inflammatory actions depending on certain environmental conditions, while also regulating pathological and physiological processes.23 In the ovalbumin (OVA)-induced allergic asthma paradigm, the downregulation of KLF4 diminishes the polarization of alternatively activated macrophages (AAM), hence reducing asthma inflammation.24 KLF4 significantly influences Th17 cell development in the context of asthma airway inflammation; nevertheless, it remains unclear whether miR-206 regulates asthma inflammation through the modulation of KLF4.
This study examined the relationship between miR-206 and KLF4 in asthma and its regulation mechanism on Th17 cell development. Through the examination of miR-206 and KLF4 expression levels and their association with Th17 cells in individuals experiencing acute asthma exacerbation and healthy controls, alongside in vitro cellular experiments, we elucidate the molecular mechanism by which miR-206 modulates Th17 cell differentiation via KLF4 targeting. This study enhances the comprehension of asthma pathophysiology and offers a theoretical foundation for developing novel therapeutic options targeting miR-206, holding substantial scientific and clinical importance.
Methods
Acquisition and Analysis of GEO Dataset
This work employed gene expression data from the GEO (Gene Expression Omnibus) database through the following steps: Conduct a search for pertinent GSE series datasets in the GEO database using keywords such as “asthma” or “microRNA” to identify suitable GSE series datasets. Utilize the GEOquery tool to get both raw and processed data for chosen datasets (GSE205876), encompassing expression matrices and sample information. Analyze the data, addressing missing values and outliers to maintain data integrity; compare gene expression in asthma patients with that of healthy controls and identify significantly differentially expressed genes. Heatmaps were created to illustrate the location and expression patterns of differentially expressed genes, offering a comprehensive analytical foundation for investigating the molecular processes of asthma.
Clinical Specimens
This study enrolled 30 patients experiencing acute asthma exacerbation and 30 healthy individuals at the Second Hospital of Shanxi Medical University from 01/03/2022 to 30/09/2023. Our study conducted experiments on both males and females and reported similar results in both genders.The inclusion criteria for asthma patients were: (1) adherence to the diagnostic standards established by the Guidelines for the Prevention and Treatment of Bronchial Asthma; (2) gender-neutral individuals aged 18 to 65 years; (3) patients hospitalized without prior systemic treatment involving glucocorticosteroids or other immunosuppressive agents; (4) exclusion of individuals with concomitant respiratory diseases, allergic conditions, underlying health issues, tumors, immune-related disorders, and significant organ dysfunction. The healthy population was recruited from our health checkup center, where all participants underwent basic examinations, including lung imaging and routine blood tests, during the same period. They were free from respiratory diseases, allergic conditions, immune-related disorders, underlying illnesses, and significant organ dysfunctions. Fundamental characteristics of all individuals, including age, sex, and laboratory results (IL-17, absolute neutrophil count, absolute eosinophil count, IgE) were gathered and documented.Written informed consent was obtained from all participants prior to inclusion in the study.
Acquisition and Analysis of Peripheral Blood Specimens
Five milliliters of peripheral blood were obtained from the subjects using heparin anticoagulation tubes, combined in equal ratios with PBS buffer (Solepol, China), transferred to an equivalent volume of human lymphocyte isolate (Solepol, China), and centrifuged at 2380 rpm for 22 minutes at ambient temperature to isolate the PBMCs in the intermediate white layer. Three milliliters of erythrocyte lysates (Sangong Biotech, China) were subsequently added following a wash with five times the volume of PBS. The PBMCs were lysed for 5–10 minutes at ambient temperature, washed twice with five times the volume of PBS, and preserved at −80°C until analysis following labeling with the subject number and time.
RT-qPCR
PBMCs were isolated from the same patients used for in vitro experiments.Total RNA was extracted from PBMCs using the Total RNA Extraction Kit (Polymeric, China), and cDNA was generated by reverse transcription using the Reverse Transcription cDNA Kit (Novozymes, China) and MicroRNA Stem Loop Reverse Transcription Kit (Novozymes, China), respectively. Subsequently, RT-qPCR was performed using the Fluorescence Real-Time Quantitative PCR Kit (Novozymes, China) to quantify the KLF4 mRNA (with ACTB as internal reference) and miR-206 expression (with U6 as internal reference). All the above operations were performed according to the kit instructions, and three replicate wells were set up for each sample. qPCR reactions were performed according to the following cycling conditions: pre-denaturation at 95°C for 30s, followed by 40 cycles, each cycle including denaturation at 95°C for 10s and extension at 60°C for 30s. The primer sequences were as follows: KLF4 mRNA (ACTB as internal reference) and miR-206 expression (U6 as internal reference). Primer sequences were as follows: KLF4 mRNA upstream (5′-CAAGTCCCGCCGCTCCATTACCAA-3′) and downstream (5′-CCACAGCCGTCCCAGTCACAGTGG-3′); ACTB upstream (5′-CCTGGCACCCAGCACAAT-3′) and downstream (5′- GGGCCGGACTCGTCATAC-3′); miR-206 upstream (5′-CGACGCGATGGAATGTAAGGAAG-3′) and downstream (5′-ATCCAGTGCAGGGTCCGAGG-3′); U6 upstream (5′-CTCGCTTCGGCAGCACA-3′) and downstream (5′ -AACGCTTCACGAATTTGCGT-3′). Relative expression was analyzed by ΔΔCt and 2-ΔΔCt methods.
Flow Cytometry
Two microliters (2 µL/mL) of stimulant were given to each pair of peripheral blood samples, and the supernatant was removed by centrifugation at 2500 rpm for 5 minutes following a 4–5 hour incubation at 37°C with 5% CO2. Following the addition of 20 μL of FITC-labeled anti-CD4 antibody (BD Pharmingen, USA) for labeling, the cells were subsequently fixed and permeabilized using an intracellular fixation and permeabilization buffer set. Twenty microliters of IL-17A-PE (BD Pharmingen, USA) were incubated for 30 minutes in the absence of light for the purpose of sorting Th17 cells. The cells underwent two washes with PBS and were subsequently examined using BD Pharmingen flow cytometry.
CD4+ T Cell Therapy
PBMCs were produced at a concentration of 1 x 107 cells/mL, and CD4+ T cells were isolated by magnetic bead-positive selection. CD4+ T cells were incubated with CD3/CD28 cell activator and IL-2 cytokines for 72 hours. Subsequently, the medium was altered, IL-2 cytokines were introduced, and the mixture was cultured for an additional 48 hours. The incubation conditions were 37°C with 5% CO2. The purity of the isolated CD4+ T lymphocytes was assessed by flow cytometry. CD4+ T cells were transfected with Mimics nc (Sangong Biotech, China), miR-206 agomir (Sangong Biotech, China), Inhibitor nc (Sangong Biotech, China), and miR-206 antagomir (Sangong Biotech, China) with the Lipofectamine RNAiMAX Reagent kit. All transfection experiments were conducted in complete compliance with the LDL. All transfection experiments were conducted in exact accordance with the transfection instructions for Lipofectamine RNAiMAX Reagent. Cells that were successfully transfected were stimulated with 100 μg/mL LPS for 12 hours. RT-qPCR identified alterations in the expression of miR-206 and KLF4 in the cells, as detailed in Section 2.4. Flow cytometry assessed the proportion of Th17 cells across the treatment groups utilizing the identical methodology as described in Section 2.4.
Enzyme-Linked Immunosorbent Assay (ELISA)
IL-17 levels in the cell supernatant were quantified utilizing the IL-17 ELISA Kit (Beinlein, USA) according to the manufacturer’s guidelines. All samples and standards underwent repeated measurements.
Statistical Evaluation
The experimental results were subjected to statistical analysis utilizing SPSS 25.0 and Prism 10.1.2. Results were presented as mean ± standard deviation (mean ± SD), and intergroup comparisons were conducted using the independent samples t-test. Data that were not normally distributed were presented as median (interquartile range) [M(P25, P75)], and group comparisons were conducted using the rank sum test (Mann–Whitney U-test). Comparisons among different groups were evaluated using one-way ANOVA or Welch’s test. Differences were statistically significant when P < 0.05. The relationship between miR-206 expression, KLF4 expression, and Th17 cell count was assessed by Pearson correlation analysis.
Ethical Approval and Declaration
This study has been approved by the Ethics Committee of the Second Hospital of Shanxi Medical University (Approval Number: 2022YX079). All procedures in this study strictly adhered to the ethical principles outlined in the Helsinki Declaration.
Results
Baseline Characterization
Comprehensive details about participant characteristics are shown in Table 1.
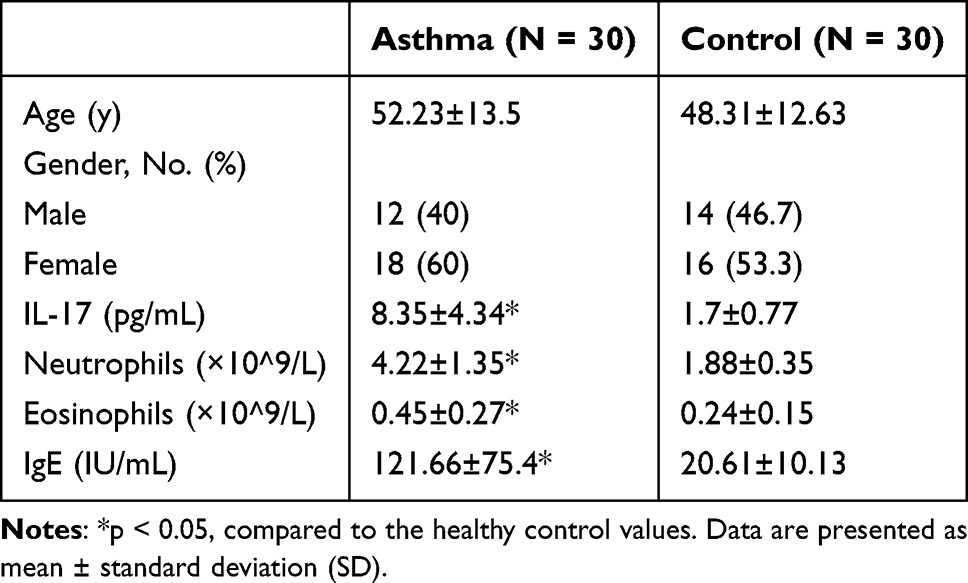 |
Table 1 Demographic Characteristics of Participants |
Decreased miR-206 Expression, Elevated KLF4 Levels, and Increased Th17 Cell Counts in Individuals with Severe Asthma Exacerbation
We obtained miRNA expression data from the GEO database (GSE205876) to analyze the expression levels of miRNAs in individuals undergoing acute asthma exacerbation relative to a healthy population. This data was subsequently utilized to generate heatmaps. The expression level of miR-206 was markedly reduced in asthma patients compared to the healthy population (Figure 1A). The RT-qPCR experiment indicated that miR-206 expression levels were considerably reduced in patients experiencing acute asthma exacerbation compared to the healthy cohort (P<0.05) (Figure 1B). KLF is a target of miR-206, and there exists an autoregulatory feedback loop between miR-206 and KLF4. RT-qPCR was employed to assess KLF4 mRNA expression in individuals with acute asthma exacerbation compared to healthy controls, revealing a significant increase in KLF4 mRNA levels in the asthma cohort (P<0.0001) (Figure 1C). We discovered that KLF4 facilitates the differentiation of Th17 cells and directly modulates the expression of IL-17A, which is crucial in the inflammatory response mechanism associated with asthma. These findings prompted us to postulate that miR-206 may play a role in the developmental phase of asthma by modulating the function of Th17 cells. Consistent with expectations, flow cytometry revealed a large increase in Th17 cells in individuals experiencing acute asthma exacerbation compared to the healthy population (P<0.001) (Figure 1D). In conclusion, the expression level of miR-206 was down-regulated, KLF4 expression was boosted, and the number of Th17 cells was increased in individuals experiencing acute asthma exacerbation. The precise regulatory interaction between miR-206 and KLF4, as well as its impact on Th17 cell development, requires additional research.
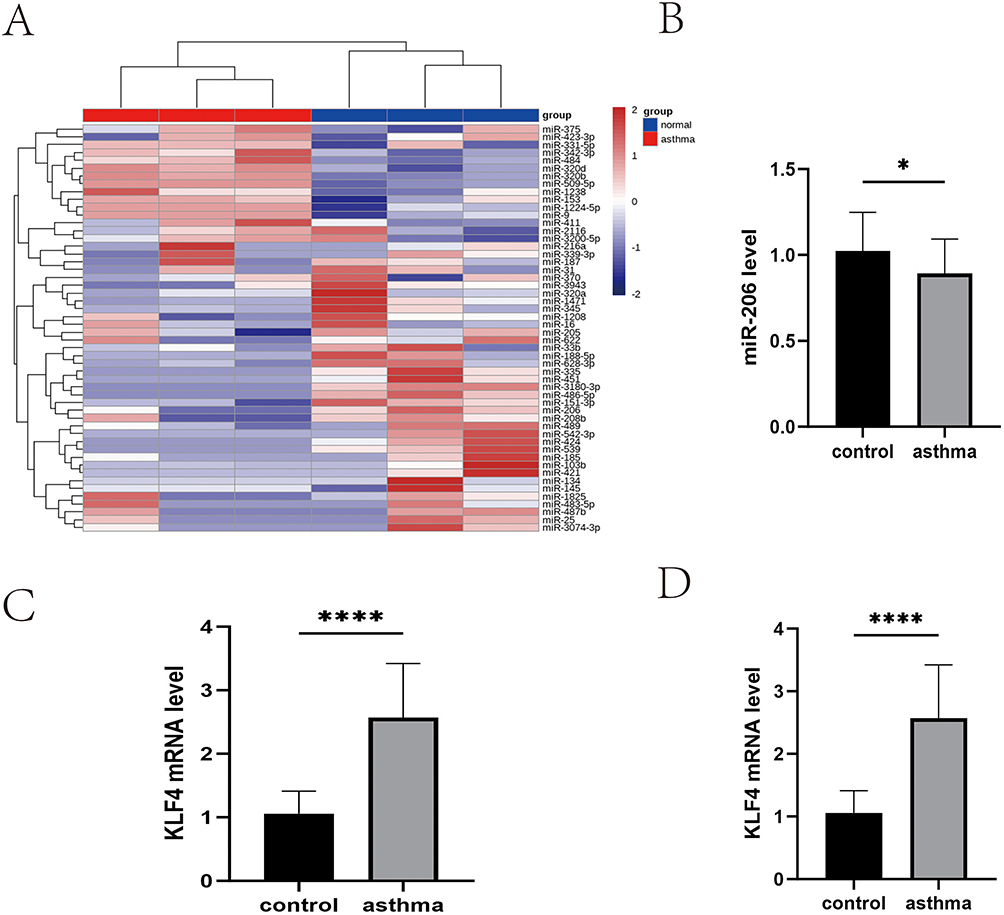 |
Figure 1 Illustrates the expression of miR-206 and KLF4 mRNA, together with the quantity of Th17 cells in patients experiencing acute asthma exacerbations. (A) Heatmap of miRNAs data from the GEO database. (B) Levels of miR-206 in control (healthy) (n=30) and asthma (asthma) (n=30). (C) Levels of KLF4 mRNA in control (healthy) (n=30) and asthma (asthma) (n=30). (D) Levels of Th17 cell number in control (healthy) (n =30) and asthma (asthma) (n=30) Th17 cell counts. *P<0.05, ****P<0.0001. |
The Downregulation of miR-206 and Elevated Expression of KLF4 May Collaboratively Govern Th17 Cell Differentiation
To investigate the correlation between miR-206, KLF4, and Th17 cell count, we performed a systematic analysis. We employed Pearson correlation analysis to elucidate their potential roles in the evolution of asthma and their interactions. The research demonstrated an autoregulatory feedback loop between miR-206 and KLF4, indicating a strong negative connection between the expression levels of miR-206 and KLF4 in individuals experiencing acute asthma exacerbation (r= −0.5232, P<0.01) (Figure 2A). Furthermore, it has been recorded that the expression level of miR-206 was diminished in peripheral blood mononuclear cells (PBMCs) from patients with dermatomyositis. Conversely, the expression level of IL-17 mRNA was elevated, although the expression of miR-206 exhibited a negative correlation with the quantity of Th17 cells. The current investigation revealed that the expression level of miR-206 in individuals experiencing acute asthma exacerbation had a negative correlation with both the quantity of Th17 cells and the concentration of IL-17 (P<0.05) (Figure 2B and C). This study found that the expression level of KLF4 in patients experiencing acute asthma exacerbation positively correlated with the quantity of Th17 cells and the level of IL-17 (P<0.05) (Figure 2D and E). We hypothesized that the Th17 cell-mediated inflammatory response may be modulated by upregulating miR-206 or downregulating KLF4 expression.
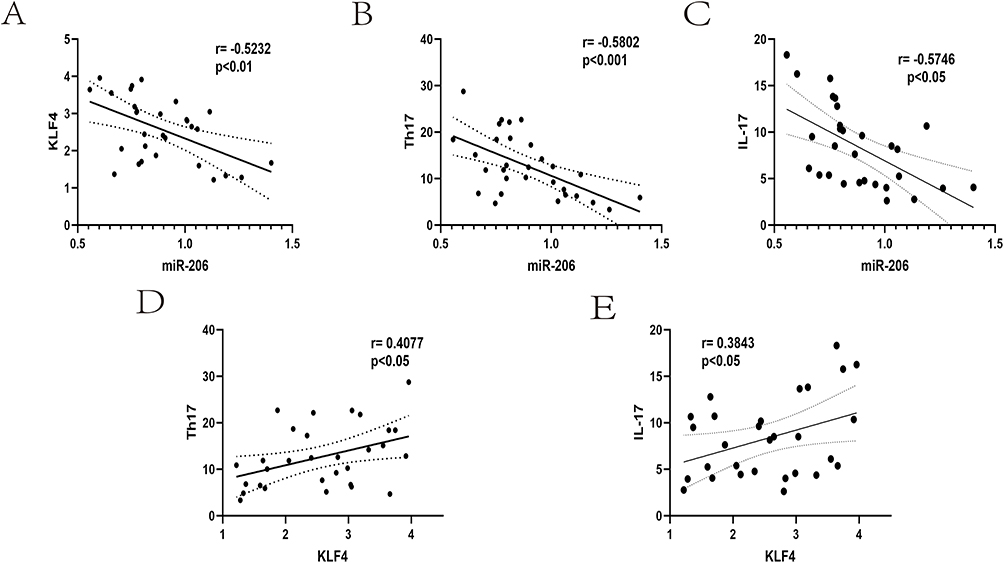 |
Figure 2 Illustrates the correlation among miR-206, KLF4, and Th17 in patients experiencing severe asthma exacerbations. (A) Correlation between miR-206 and KLF4 in patients with acute asthma exacerbation. (B) Correlation between miR-206 expression level and Th17 cell number in patients with acute asthma exacerbation. (C) Correlation between miR-206 expression level and IL-17 in patients with acute asthma exacerbation. (D) Correlation between KLF4 expression level and Th17 cell number in patients with acute asthma exacerbation. (E) Correlation between KLF4 expression level and IL-17 in patients with acute asthma exacerbation. |
miR-206 Diminishes the Prevalence of Th17 Cells
To validate the aforementioned supposition, we extracted CD4+ T cells from the peripheral blood of asthma patients for later experimentation, and a flow cytometry analysis demonstrated that the purity of the sorted CD4+ T cells reached 99.2% (Figure 3A). The expression level of miR-206 was markedly diminished in patients experiencing acute asthma exacerbation.This study revealed a negative correlation between the expression level of miR-206 in patients experiencing acute asthma exacerbation and the quantity of Th17 cells, as well as the concentration of IL-17, leading us to further explore the regulatory influence of miR-206 on Th17 cell proportions. We overexpressed and inhibited miR-206 by transfecting liposomes to investigate the role and mechanism of miR-206 in asthma. In the study, miRNA agomir (miR-206 agonist) and miRNA antagomir (miR-206 inhibitor) were selected as regulatory tools, and these two molecules have been proven to have stable and efficient expression regulation characteristics in previous in vitro and in vivo experimental models.We detected the transfection efficiency using RT-qPCR. The RT qPCR detection results clearly showed that transfection of miR-206 agomir significantly increased miR-206 levels (P<0.0001), while transfection of miR-206 antagomir led to a decrease in miR-206 levels (P<0.001) (Figure 3B). These data fully confirm the successful establishment of an effective miR-206 regulatory model in the experiment. Based on the successful establishment of the miR-206 regulatory model, we further stimulated cells with LPS to simulate the inflammatory state of asthma, providing an experimental basis for subsequent mechanistic research. Flow cytometry revealed the proportion of Th17 cells in the treatment groups: Mimics nc, miR-206 agomir, Inhibitor nc, and miR-206 antagomir. The findings indicated that the proportion of Th17 cells in the miR-206 agomir group was markedly reduced (P<0.05), but the proportion of Th17 cells in the miR-206 antagomir group was dramatically elevated (P<0.01) (Figure 3C and D). This suggested that the overexpression of miR-206 may impede the development of Th17 cells. Conversely, there was no notable alteration in the mimics nc group or the inhibitor nc group when compared to the LPS group, and the intergroup differences were not statistically significant (P>0.05). This work confirmed the significant role of miR-206 in modulating Th17 cell development, namely by down-regulating the number of Th17 cells.
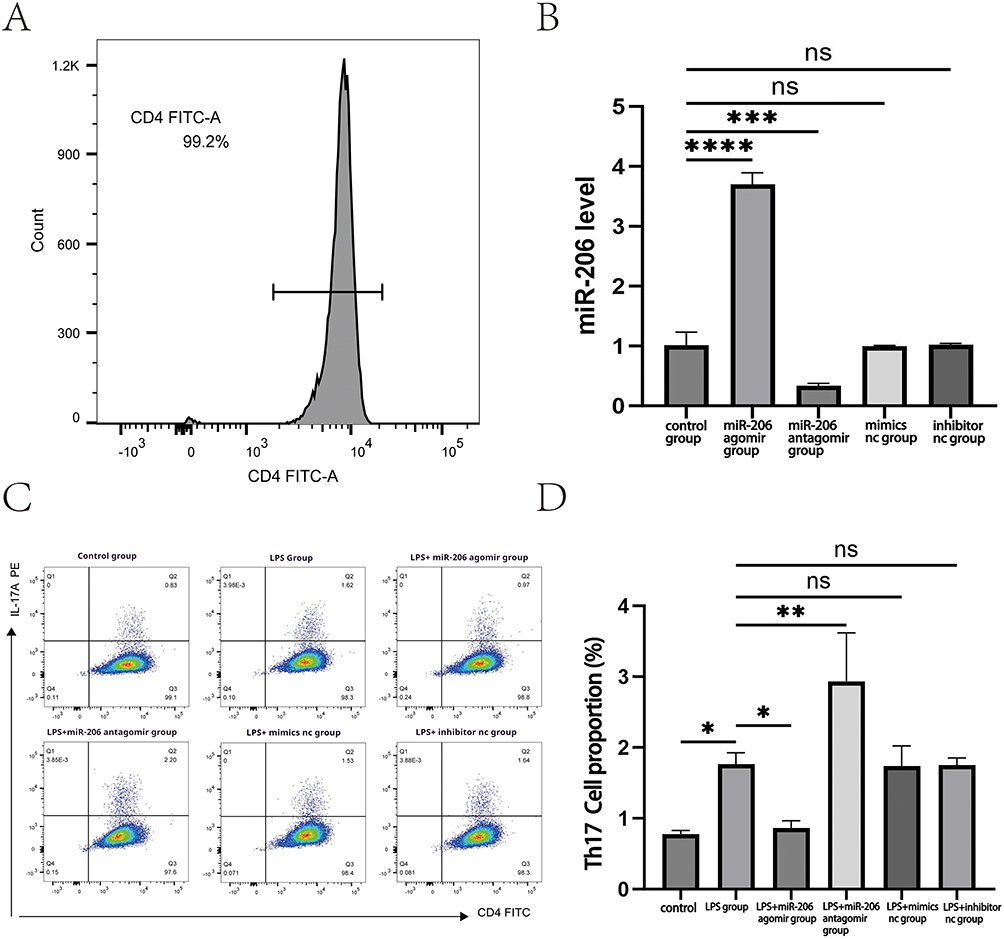 |
Figure 3 Illustrates that miR-206 lowers the proportion of Th17 cells. (A) Concentration of CD4+ T cells. (B) Expression levels of miR-206 in the treatment groups: Mimics nc, miR-206 agomir, Inhibitor nc, and miR-206 antagomir. (C) the percentage of Th17 cells in the treatment groups of Mimics nc, miR-206 agomir, Inhibitor nc, and miR-206 antagomir, together with other treatment groups; (D) analysis of the proportion of Th17 cells in the aforementioned treatment groups. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001,ns P>0.05. |
miR-206 Inhibits KLF4 Expression
As previously stated, miR-206 can inhibit the development of Th17 cells. Nevertheless, its precise mechanism of regulating gene expression requires additional investigation. KLF4 has been identified as a target of miR-206 and is capable of activating Th17 cell differentiation. However, the precise mechanism by which miR-206 modulates asthma inflammation through the targeting of KLF4 requires additional validation. We predicted that miR-206 suppresses Th17 cell development by negatively targeting and modulating KLF4. To validate this theory, we initially assessed the relative expression levels of KLF4 mRNA in the treatment groups of Mimics nc, miR-206 agomir, Inhibitor nc, and miR-206 antagomir utilizing RT-qPCR. The findings indicated a significant reduction in the relative expression level of KLF4 mRNA in the miR-206 agomir group (P<0.05).Conversely, the relative expression level of KLF4 mRNA in the miR-206 antagomir group was markedly elevated (P<0.0001), indicating that miR-206 negatively modulates KLF4 expression (Figure 4). This experiment validated the inhibitory effect of miR-206 on KLF4 expression through RT qPCR analysis, supporting the partial validity of the hypothesis that miR-206 may act by targeting KLF4. However, it should be noted that this study has limitations: it has not directly confirmed the specific function of KLF4 in Th17 cell differentiation, and its direct effect on Th17 cell differentiation has not been verified through functional experiments such as downregulation or upregulation of KLF4 expression.
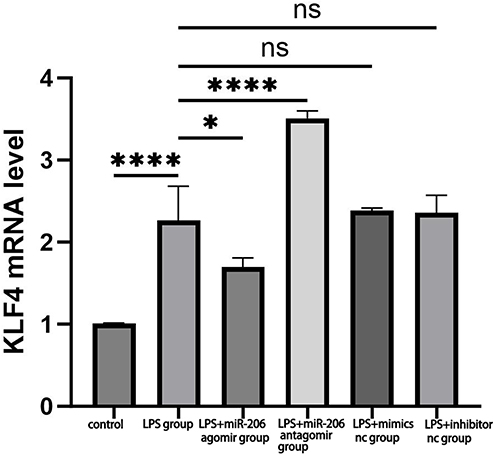 |
Figure 4 Illustrates that miR-206 diminishes the expression of KLF4. Relative expression levels of KLF4 mRNA in the treatment groups: Mimics nc, miR-206 agomir, Inhibitor nc, and miR-206 antagomir. *P < 0.05, ****P < 0.0001,ns P>0.05. |
Therefore, whether the current effect of miR-206 on Th17 cells is indeed mediated by regulating KLF4, and its complete regulatory mechanism in asthma inflammation, still needs further research to clarify, such as conducting KLF4 overexpression or knockout experiments, and combining experimental design with the detection of Th17 cell differentiation indicators to further verify this regulatory pathway.
miR-206 Inhibits the Inflammatory Factor IL-17
miR-206 suppresses the differentiation of Th17 cells, which secrete cytokines like IL-17A/F and contribute to the immune response in asthma by targeting KLF4. miR-206 is situated upstream of the IL-17 gene. We hypothesized that miR-206 may downregulate the expression of the inflammatory factor IL-17 based on this mechanism. To validate this hypothesis, we assessed the expression levels of IL-17 in the treatment groups of Mimics nc, miR-206 agomir, Inhibitor nc, and miR-206 antagomir using ELISA. The experimental findings indicated a significant elevation in the secretion of the inflammatory factor IL-17 following LPS-induced treatment (P<0.01). In the miR-206 agomir group, IL-17 expression was significantly reduced (P<0.05), whereas in the miR-206 antagomir group, IL-17 expression was significantly elevated (P<0.05) (Figure 5). The results further corroborated that miR-206 diminished the LPS-induced production of the inflammatory factor IL-17.
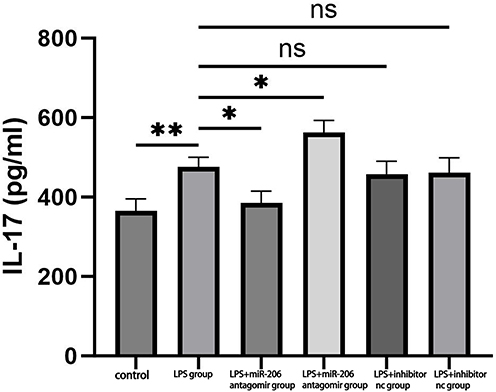 |
Figure 5 Illustrates the expression levels of IL-17 illustrates the expression levels of IL-17 across the treatment groups: Mimics nc, miR-206 agomir, Inhibitor nc, and miR-206 antagomir. *P < 0.05, **P < 0.01,ns P>0.05. |
Discussion
Asthma is a prevalent chronic inflammatory pulmonary disease primarily induced by triggers such as allergens and infections. It is marked by persistent airway inflammation, with recurrent episodes provoking and intensifying airway hyper-responsiveness and remodeling.25 Neutrophilic asthma is an important subtype of asthma with unique pathophysiological features. Unlike classical Th2 asthma, this subtype is mainly associated with non allergic factors such as environmental pollutants, respiratory pathogen infections (such as bacteria or viruses), and autoimmune reactions.10 Epidemiological studies have shown that neutrophilic asthma accounts for a significant proportion of severe asthma patients, and the global disease burden continues to increase. Due to its poor response to conventional glucocorticoid therapy, it has become an important challenge for clinical management and public health.26 Consequently, additional elucidation of asthma’s molecular pathogenesis is required in the pursuit of potential therapeutic targets for the condition.
MiRNAs, as potent gene regulators, play a role in the development of various human diseases and have emerged as potential diagnostic biomarkers.27 miRNAs may become a new therapeutic approach. Numerous studies indicate that miRNAs significantly contribute to asthma development, with miR-206 exhibiting diminished levels in airway remodeling,28 allergic rhinitis, and non-atopic asthma among patients.29 Analysis of the GEO database revealed that the expression level of miR-206 in asthma patients was markedly lower than in the healthy population, offering a novel insight into the molecular underpinnings of asthma. Our study revealed that miR-206 expression levels were significantly diminished in patients experiencing acute asthma exacerbation compared to the healthy population. This outcome not only validates the database findings but also underscores the potential significance of miR-206 in the pathophysiological mechanisms of asthma. The downregulation of miR-206 expression may correlate with heightened airway inflammation in asthma patients, and its reduction may disrupt the normal regulation of the inflammatory response, resulting in excessive release of inflammatory mediators and an escalation of airway hyperresponsiveness.
This study examined the regulatory function of miR-206 in asthma and its possible therapeutic implications. miR-206 is a member of the miR-1 family and is situated on human chromosome 6p12.2, directly upstream of the IL-17 gene.30,31 IL-17-secreting Th17 cells are an important T-cell subpopulation involved in the pathogenesis of asthma airway inflammation, and their aberrant differentiation is closely related to the excessive release of inflammatory factors. kLF4, a transcription factor, is crucial in cell growth, proliferation, differentiation, immunity, and inflammation, and is a target of miR-206. there is an autoregulation between miR-206 and KLF4 feedback loop, where overexpression of miR-206 inhibits KLF4 levels, whereas inhibition of miR-206 promotes KLF4 expression.19,20 KLF4 is instrumental in TCR-mediated T-cell proliferation through the regulation of the cell cycle. It binds to the IL-17A gene promoter to directly enhance its expression, independently of ROR-γt regulation of Th17 cell differentiation, thus directly affecting IL-17 secretion.21,22
Our study additionally verified that the expression of miR-206 was markedly diminished in asthma patients. Conversely, the expression of KLF4 was notably upregulated, accompanied by a substantial increase in the quantity of Th17 cells and IL-17 levels. Through correlation analysis of peripheral blood from asthma patients, we identified that miR-206 exhibited a negative correlation with KLF4, Th17 cell count, and IL-17 levels, whereas KLF4 demonstrated a positive correlation with Th17 cell count and IL-17 levels. We hypothesized that miR-206 may mitigate asthma airway inflammation by negatively regulating KLF4 and Th17 cell differentiation, based on these findings. We further validated this hypothesis through in vitro experiments utilizing miR-206 mimics. The experimental findings indicated that the overexpression of miR-206 markedly reduced the proportion of Th17 cells and inhibited the mRNA levels of KLF4. Inhibition of miR-206, conversely, enhanced KLF4 expression. The findings substantiate that miR-206 exerts a negative regulatory effect on Th17 cell differentiation and function by targeting and suppressing KLF4 expression.
The findings indicate that miR-206 plays a substantial anti-inflammatory role in the pathogenesis of asthma, primarily through the modulation of the KLF4-Th17-IL-17 axis. Under typical circumstances, Th17 cells collaborate with regulatory T cells (Treg) to sustain the dynamic equilibrium of the immune system, with Th17 cells facilitating pro-inflammatory responses and Treg cells providing anti-inflammatory effects. In asthmatics, the equilibrium between Th17 and Treg cells is disturbed, resulting in the suppression of Treg cell anti-inflammatory functions and the hyperactivation of Th17 cells, which contributes to sustained airway inflammation.32 IL-17, a principal effector of Th17 cells, is pivotal in airway inflammation associated with asthma. It intensifies airway inflammatory responses by facilitating neutrophil recruitment and activation. Moreover, IL-17 induces airway hypersecretion of mucus, hyperplasia of goblet cells, differentiation of myofibroblasts, and proliferation of airway smooth muscle, all of which collectively facilitate airway remodeling and exacerbate asthma symptoms.33 miR-206 inhibits the differentiation of Th17 cells and the secretion of IL-17 by targeting and suppressing KLF4 expression, thereby mitigating airway inflammation and pathological changes. Restoring miR-206 expression or augmenting its function may facilitate the reestablishment of Th17/Treg homeostasis and suppress IL-17-mediated inflammatory responses, consequently alleviating asthma symptoms and enhancing patients’ quality of life.
MiR-206 plays a key regulatory role in neutrophilic asthma by interacting with KLF4, Th17 cells, and IL-17. It may inhibit the abnormal differentiation of Th17 cells and excessive secretion of IL-17 by targeting KLF4. Compared with other miRNAs involved in Th17 regulation, miR-206 exhibits a different functional polarity from miR-326. MiR-326 exerts pro-inflammatory effects by inhibiting Ets-1 to promote Th17 differentiation34 while miR-206 inhibits Th17 differentiation by targeting different downstream molecules, exhibiting anti-inflammatory effects. This differentiated regulatory pattern provides a new target selection for asthma treatment. However, the core limitation of current research lies in the lack of in vivo experimental verification. It is not yet clear whether the regulatory effects observed in vitro can maintain the same direction and intensity in a real airway inflammatory environment. Subsequently, animal models need to be constructed, combined with miR-206 overexpression or knockout experiments, to verify its true regulatory effects on airway hyperresponsiveness, inflammatory cell infiltration, and the Th17/IL-17 axis in vivo, and to improve it by combining clinical samples. In terms of clinical translation, there is still a need to overcome challenges such as targeted delivery, stability, safety, and individualized differences, such as how to achieve specific delivery of miRNA to target tissues (such as lungs and immune cells) and reduce off target effects on normal tissues. Exogenous miRNAs are easily cleared in the body’s circulation and need to be chemically modified to enhance their stability, or combined with slow-release carriers to prolong their duration of action and reduce the risk of immune reactions or gene regulation disorders that may arise from long-term use. The expression level and target gene sensitivity of miR-206 in asthma patients may vary, leading to individual differences in the therapeutic effect of miRNA mimetics. Individualized plans should be developed in combination with precision medicine. The potential synergistic effect of MiR-206 regulatory strategy and existing anti-IL-17 biologics can achieve a more comprehensive therapeutic effect. MiR-206 can inhibit Th17 cell differentiation upstream to reduce IL-17 sources, while anti-IL-17 biologics directly neutralize downstream IL-17 molecules. The combination of the two may enhance anti-inflammatory effects through a dual mechanism of “source inhibition+end blockade”, providing a more comprehensive treatment plan for asthma patients. In summary, miR-206 is expected to become a potential target for anti asthma treatment, opening up new avenues for exploring the pathogenesis and developing targeted therapies.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Hammad H, Lambrecht BN. The basic immunology of asthma. Cell. 2021;184(6):1469–1485. doi:10.1016/j.cell.2021.02.016
2. Porsbjerg C, Melén E, Lehtimäki L, Shaw D. Asthma. Lancet. 2023;401(10379):858–873. doi:10.1016/S0140-6736(22)02125-0
3. Huang K, Yang T, Xu J, et al. Prevalence, risk factors, and management of asthma in China: a national cross-sectional study. Lancet. 2019;394(10196):407–418. doi:10.1016/S0140-6736(19)31147-X
4. Charles D, Shanley J, Temple S-N, et al. Real-world efficacy of treatment with benralizumab, dupilumab, mepolizumab and reslizumab for severe asthma: a systematic review and meta-analysis. Clinical and Experimental Allergy: Journal of the British Society for Allergy and Clinical Immunology. 2022;52(5):616–627.
5. Singh D, Garcia G, Maneechotesuwan K, et al. New Versus Old: the Impact of Changing Patterns of Inhaled Corticosteroid Prescribing and Dosing Regimens in Asthma Management. Advances in Therapy. 2022;39(5):1895–1914. doi:10.1007/s12325-022-02092-7
6. Habib N, Pasha MA, Tang DD. Current Understanding of Asthma Pathogenesis and Biomarkers. Cells. 2022;11(17):2764. doi:10.3390/cells11172764
7. Infante-Duarte C, Horton HF, Byrne MC, Kamradt T. Microbial lipopeptides induce the production of IL-17 in Th cells. Journal of Immunology. 2000;165(11):6107–6115. doi:10.4049/jimmunol.165.11.6107
8. Wei Q, Liao J, Jiang M, Liu J, Liang X, Nong G. Relationship between Th17-mediated immunity and airway inflammation in childhood neutrophilic asthma. Allergy, Asthma, and Clinical Immunology: Official Journal of the Canadian Society of Allergy and Clinical Immunology. 2021;17(1):4. doi:10.1186/s13223-020-00504-3
9. Sorbello V, Ciprandi G, Di Stefano A, et al. Nasal IL-17F is related to bronchial IL-17F/neutrophilia and exacerbations in stable atopic severe asthma. Allergy. 2015;70(2):236–240. doi:10.1111/all.12547
10. Zhang X, Xu Z, Wen X, et al. The onset, development and pathogenesis of severe neutrophilic asthma. Immunol Cell Biol. 2022;100(3):144–159. doi:10.1111/imcb.12522
11. Bartel DP. Metazoan MicroRNAs. Cell. 2018;173(1):20–51. doi:10.1016/j.cell.2018.03.006
12. Chua FY, Adams BD. Androgen receptor and miR-206 regulation in prostate cancer. Transcription. 2017;8(5):313–327. doi:10.1080/21541264.2017.1322668
13. Chen R, Piao L-Z, Liu L, Zhang X-F. DNA methylation and gene expression profiles to identify childhood atopic asthma associated genes. BMC Pulmonary Medicine. 2021;21(1):292. doi:10.1186/s12890-021-01655-8
14. Panganiban RP, Wang Y, Howrylak J, et al. Circulating microRNAs as biomarkers in patients with allergic rhinitis and asthma. The Journal of Allergy and Clinical Immunology. 2016;137(5):1423–1432. doi:10.1016/j.jaci.2016.01.029
15. Kho AT, McGeachie MJ, Moore KG, et al. Circulating microRNAs and prediction of asthma exacerbation in childhood asthma. Respiratory Research. 2018;19(1):128. doi:10.1186/s12931-018-0828-6
16. Liu Y, Huo S-G, Xu L, et al. MiR-135b Alleviates Airway Inflammation in Asthmatic Children and Experimental Mice with Asthma via Regulating CXCL12. Immunological Investigations. 2022;51(3):496–510. doi:10.1080/08820139.2020.1841221
17. Haas JD, Nistala K, Petermann F, et al. Expression of miRNAs miR-133b and miR-206 in the Il17a/f locus is co-regulated with IL-17 production in αβ and γδ T cells. PLoS One. 2011;6(5):e20171. doi:10.1371/journal.pone.0020171
18. Tang X, Tian X, Zhang Y, et al. Correlation between the frequency of Th17 cell and the expression of microRNA-206 in patients with dermatomyositis. Clinical & Developmental Immunology. 2013;2013:345347. doi:10.1155/2013/345347
19. Liu N, Wang X, Steer CJ, Song G. MicroRNA-206 promotes the recruitment of CD8+ T cells by driving M1 polarization of Kupffer cells. Gut. 2022;71(8):1642–1655. doi:10.1136/gutjnl-2021-324170
20. Lin -C-C, Liu L-Z, Addison JB, et al. A KLF4-miRNA-206 autoregulatory feedback loop can promote or inhibit protein translation depending upon cell context. Molecular and Cellular Biology. 2011;31(12):2513–2527. doi:10.1128/MCB.01189-10
21. An J, Golech S, Klaewsongkram J, et al. Krüppel-like factor 4 (KLF4) directly regulates proliferation in thymocyte development and IL-17 expression during Th17 differentiation. FASEB Journal: Official Publication of the Federation of American Societies for Experimental Biology. 2011;25(10):3634–3645. doi:10.1096/fj.11-186924
22. Lebson L, Gocke A, Rosenzweig J, et al. Cutting edge: the transcription factor Kruppel-like factor 4 regulates the differentiation of Th17 cells independently of RORγt. Journal of Immunology. 2010;185(12):7161–7164. doi:10.4049/jimmunol.1002750
23. He Z, He J, Xie K. KLF4 transcription factor in tumorigenesis. Cell Death Discovery. 2023;9(1):118. doi:10.1038/s41420-023-01416-y
24. Song Y, Li X, Liu F, Zhu H, Shen Y. Isoalantolactone alleviates ovalbumin-induced asthmatic inflammation by reducing alternatively activated macrophage and STAT6/PPAR-γ/KLF4 signals. Molecular Medicine Reports. 2021;24(4):701. doi:10.3892/mmr.2021.12340
25. Abe Y, Suga Y, Fukushima K, et al. Advances and Challenges of Antibody Therapeutics for Severe Bronchial Asthma. International Journal of Molecular Sciences. 2021;23(1):83. doi:10.3390/ijms23010083
26. Ray A, Kolls JK. Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol. 2017;38(12):942–954. doi:10.1016/j.it.2017.07.003
27. Ho PTB, Clark IM, Le LTT. MicroRNA-Based Diagnosis and Therapy. International Journal of Molecular Sciences. 2022;23(13):7167. doi:10.3390/ijms23137167
28. Pan Y, Liu L, Li S, et al. Activation of AMPK inhibits TGF-β1-induced airway smooth muscle cells proliferation and its potential mechanisms. Scientific Reports. 2018;8(1):3624. doi:10.1038/s41598-018-21812-0
29. Tunçer F, Şahiner ÜM, Ocak M, et al. Comparison of miRNA expression in patients with seasonal and perennial allergic rhinitis and non-atopic asthma. The Turkish Journal of Pediatrics. 2022;64(5):859–868. doi:10.24953/turkjped.2022.410
30. Ma G, Wang Y, Li Y, et al. MiR-206, a Key Modulator of Skeletal Muscle Development and Disease. International Journal of Biological Sciences. 2015;11(3):345–352. doi:10.7150/ijbs.10921
31. Pan J-Y, Sun -C-C, Bi Z-Y, et al. miR-206/133b Cluster: a Weapon against Lung Cancer? Molecular Therapy Nucleic Acids. 2017;8:442–449. doi:10.1016/j.omtn.2017.06.002
32. Thomas R, Qiao S, Yang X. Th17/Treg Imbalance: implications in Lung Inflammatory Diseases. International Journal of Molecular Sciences. 2023;24(5):4865. doi:10.3390/ijms24054865
33. Ramakrishnan RK, Al Heialy S, Hamid Q. Role of IL-17 in asthma pathogenesis and its implications for the clinic. Expert Review of Respiratory Medicine. 2019;13(11):1057–1068. doi:10.1080/17476348.2019.1666002
34. Azimi M, Ghabaee M, Naser Moghadasi A, Izad M. Altered Expression of miR-326 in T Cell-derived Exosomes of Patients with Relapsing-remitting Multiple Sclerosis. Iran J Allergy Asthma Immunol. 2019;18(1):108–113.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.