Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 19
Low Humidity Alters Epidermal Gene Expression in vivo
Authors Wen S, Wang X, Ye L ![]() , Gong C, Liu D, Man MQ
, Gong C, Liu D, Man MQ ![]()
Received 27 November 2025
Accepted for publication 29 January 2026
Published 4 February 2026 Volume 2026:19 577853
DOI https://doi.org/10.2147/CCID.S577853
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Anne-Claire Fougerousse
Si Wen,1 Xiaohua Wang,2 Li Ye,1 Canyi Gong,3 Dan Liu,3 Mao-Qiang Man3,4
1Evaluation Center for Skin Care Product Efficacy, Dermatology Hospital of Southern Medical University, Guangzhou, Guangdong, 510091, People’s Republic of China; 2Dermatology, Dermatology Hospital of Southern Medical University, Guangzhou, Guangdong, 510091, People’s Republic of China; 3Formulation Center, Dermatology Hospital of Southern Medical University, Guangzhou, Guangdong, 510091, People’s Republic of China; 4Department of Dermatology, The First Hospital of Hebei Medical University, Shijiazhuang, Hebei, 050023, People’s Republic of China
Correspondence: Mao-Qiang Man, Formulation center, Dermatology Hospital of Southern Medical University, 2 Lujing Road, Guangzhou, 510091, People’s Republic of China, Email [email protected]
Introduction: Environmental factors, including humidity, affect cutaneous functions. The influences of low humidity on epidermal gene expression and whether an emollient can attenuate its effects are unknown.
Methods: We employed RNAseq technique to analyze epidermal gene expression in mice exposed to low humidity (15%) with or without topical treatment with an emollient (Dr. Yu Skin Barrier Repair Cream®) for 6 months. Mice maintained in normal humidity (55± 5%) served as normal controls.
Results: Exposure of mice to low humidity down-regulated 159 genes and upregulated 105 genes in the epidermis. Topical applications of the emollient upregulated 195 genes and downregulated 280 genes in mice exposed to low humidity. Low humidity primarily downregulated genes associated with interaction between viral proteins and cytokines, and cytokine-cytokine interaction pathways, while upregulating genes associated with IL-17 and TNF signaling pathways. Topical emollient upregulated signaling pathways associated with fatty acid elongation, peroxisome proliferator-activated receptors (PPAR) and fatty acid metabolism in mice exposed to low humidity, while down-regulated genes were associated with inflammation.
Conclusion: These results demonstrate that low humidity alters epidermal gene expression, and topical emollient regulates epidermal gene expression in mice exposed to low humidity, providing a rationale for utility of emollients in low-humidity environment and the management of dry skin.
Keywords: environment, humidity, epidermis, emollient, RNA sequencing, cytokines
Introduction
A number of environmental factors can influence epidermal functions. Alteration in epidermal function can be reflected in the changes in epidermal biophysical properties. Previous studies showed that sun-exposure can induce dose- and gender-dependent changes in stratum corneum hydration levels and transepidermal water loss rates (TEWL).1,2 Air pollution is another common factor negatively impacting epidermal functions. Air pollution can trigger and accelerate skin aging and some skin disorders such as atopic dermatitis and psoriasis.3–5 Likewise, exposure of skin to particulate matter for 24 hours increases the production of proinflammatory cytokines such as IL-1α and IL-8 in vitro, likely via induction of reactive oxygen species, which activate inflammasome.6–8 Moreover, several studies have demonstrated that particulate matter regulates multiple epidermal functions, including increases in epidermal permeability, decreases in expression levels of differentiation marker-related proteins, as well as reductions in keratinocyte proliferation in vivo and in vitro.9,10 Importantly, the levels of urocanic acid (UCA) and cis/trans-UCA, the products of filaggrin degradation, in the stratum corneum reversely correlate with levels of air PM2.5.10 Furthermore, environmental temperature also affects epidermal function. Previous studies showed that epidermal permeability barrier recovery is accelerated at temperature ranged 36°C to 40°C by activation of transient receptor potential receptor 4, while temperature at either 34°C or 42°C delayed barrier recovery.11 Immersion of hand to either cold (11.13°C) or hot water (41.29 oC) for 10 minutes significantly increases both TEWL and skin surface pH. Exposure of skin to cold water also increases stratum corneum hydration levels.12 Although exposure of intact skin to 4°C does not alter basal TEWL,12 exposure of barrier-disrupted skin to ice for 3.5 hours markedly increased TEWL.13 Correspondingly, working in low temperature environment (fish processing) increases TEWL along with reduction in stratum corneum hydration levels.14,15 Collectively, this line of evidence indicates the regulatory role of environmental factors in epidermal functions.
In addition, humidity also affects epidermal function. When relative humidity is under 50%, the water content in the stratum corneum will be less than 10%, causing decreases in skin softness and pliability.16 Dry skin, an indicator of reduced stratum corneum hydration, is a common phenomenon in dry environment. Denda et al showed that exposure of mice to low humidity (relative humidity<10%) for 2 weeks increases stratum corneum lipid content and decreases TEWL,17 while exposure of mice to low humidity (relative humidity<10%), not high humidity (relative humidity>80%), for 48 hours increases epidermal DNA synthesis, which can be attenuated by topical application of either glycerol or petrolatum.18 Forty-eight hours prior to and after barrier disruption, keeping mice in low humidity environment enhances epidermal hyperplasia, accompanied by increases in density of mast cells in the dermis.18 Moreover, transferring mice from high humidity (relative humidity>80%), not normal humidity (relative humidity 40–70%), to low humidity environment for 2 days increases TEWL by 6- to 7-fold, while lowering stratum corneum hydration levels.19 In humans, working 12 hours daily in the environment at relative humidity of 1.5% for 2 weeks significantly lowers both TEWL and stratum corneum hydration levels.20 Furthermore, studies showed a higher prevalence of pruritus and atopic dermatitis in subjects working in low humidity than in normal humidity environment.21,22 However, the impacts of low humidity on epidermal gene expression have not been elucidated yet. Additionally, because of the adverse impact of low humidity on skin, emollients are often used to improve skin condition, including lowering cytokine levels in the epidermis.23 However, the influences of low humidity on epidermal gene expression and whether an emollient can attenuate its effects are unknown. In the present study, we performed RNA sequencing to elucidate gene expression profile in mice exposed to low humidity with or without emollient treatments.
Materials and Methods
Animals
The body’s functions are optimal in young individuals, where a remarkable changes in gene expression may not be easily observed upon exposure to low humidity. In humans, the epidermal function such as stratum corneum hydration levels declines at age of ≈50 years,24 which is about 12-month-old in mice.25 Therefore, 12-month-old mice were used in this study. Female C57BL/6J mice were purchased from and housed at Guangdong Medical Laboratory Animal Center, China. One group of mice (n=6) was housed in a humidity chamber at temperature of 22 ± 1°C and relative humidity of 55 ± 5%, while another group of mice (n=11) was kept in a humidity chamber at temperature of 22°C and relative humidity of 15% for 6 months. Mice exposed to low humidity were divided into two groups, ie, a. both flanks were treated topically with emollient (n=6) twice daily, and b. untreated controls (n=5). Since Dr. Yu Skin Barrier Repair Cream®, containing Olea europaea (olive) fruit oil, Carthamus tinctorius (safflower) seed oil, Butyrospermum parkii (shea butter), Persea gratissima (avocado) oil, and Oryza sativa (rice) bran oil (Jahwa United Co. Shanghai, China) has shown good efficacy and safety in improving stratum corneum hydration,26,27 we chose it in our study. The cream was applied to both flanks with an index finger. Throughout the experiment, the hair was shaved as needed with an electric clipper. After 6 months, mice were euthanized with overdose isoflurane, followed by cervical dislocation. Afterward, skin samples were collected from both flanks of the mice, followed by heat separation of the epidermis from the dermis, as described previously.28 Briefly, the subcutaneous tissue was removed with a surgical scalpel, followed by placing the skin on a Petri dish with dermal side facing down. The Petri dish was then transferred to a water bath and incubated at 60°C for 60 seconds. Afterward, the epidermis was collected by scraping the epidermis with a surgical scalpel. This study was approved by the ethics committee of Guangdong Medical Laboratory Animal Center (D202410-2). All animal procedures followed the guidelines for the ethical review of laboratory animal welfare People’s Republic of China National Standard GB/T 35892‐2018.
Total RNA Extraction
The epidermis was homogenized with IKA T10 homogenizer in cold. According to the manufacturer’s instruction, total RNA was extracted from the epidermis using Trizol (Thermo Fisher Scientific, USA, Lot:252612) Plus RNeasy mini kits (Qiagen, Germany; Lot:166012799). RNA concentration and purity were assessed using a Nano Drop and Agilent 2100 bioanalyzer (Thermo Fisher Scientific, MA, USA).
mRNA Library Construction and Sequencing
About 1 µg of total RNA was incubated with oligo(dT) magnetic beads to bind polyadenylated mRNA. After washing, the purified mRNA was eluted from the beads. The enriched mRNA was fragmented at 94°C for 15 minutes in the presence of divalent cations to generate fragments of approximately 200–300 nucleotides. First-strand cDNA synthesis was then performed using random hexamer primers and ProtoScript II Reverse Transcriptase. A second-strand cDNA was synthesized using a master mix containing dUTP in place of dTTP, followed by incubation with A-Tailing Mix and RNA Index Adapters to end repair. Afterwards, PCR was used to amplify cDNA fragments obtained from previous step, and PCR products were purified with Ampure XP Beads, and dissolved in EB solution. Agilent Technologies 2100 bioanalyzer was used to assess the quality and size distribution of the prepared libraries. The double stranded PCR products from previous step were denatured by heat and circularized by the splint oligo sequence. The single strand circle DNA was formatted as the final library. The final library was amplified with phi29 to make DNA nanoball containing more than 300 copies per molecule. DNA nanoballs were loaded into the patterned nanoarray and single-end 50 base reads were read through on BGIseq500 platform (BGI-Shenzhen, China).
Analysis of RNA-Seq Data
SOAPnuke (v1.5.2) was used to filter the sequencing data, including a. removal of reads with sequencing adapter; b. removal of reads with low-quality base ratio (base quality less than or equal to 5) >20%; c. removal of reads with unknown base (“N” base) ratio >5%.29 Afterwards, clean reads were stored in FASTQ format. The clean reads were mapped onto the current Mus musculus genome (GCF_000001635.26_GRCm38.p6) using HISAT2 (v2.0.4).30 Bowtie2 (v2.2.5) was used to align the clean reads to the reference coding gene set.31 A software package, RSEM (v1.2.12), was used to calculate the expression levels of genes.32
Identification of Differentially Expressed Genes (DEGs)
Differentially expressed genes between the groups were determined using the Bioconductor package DESeq2 (v1.4.5).33 Genes with Q value ≤0.05 were defined as differentially expressed genes. | log2(Fold change) |≥1 in comparison was set as the threshold for significantly differentially expressed genes. The software ClustVis 2.0 (https://biit.cs.ut.ee/clustvis/) was used to create graphs of heatmap and principal component analysis (PCA) plots. Read counts were normalized using the median-of-ratios method in DESeq2. Since the samples were from the same batch of experiments, the data were unlikely to be affected by batch effects. No batch effect correction was performed.
Functional Enrichment Analysis of DEGs
In order to reveal the functional significances of DEGs, we performed GO (http://www.geneontology.org/) and KEGG (https://www.kegg.jp/) enrichment analysis of annotated differentially expressed genes using Phyper (https://en.wikipedia.org/wiki/Hypergeometric_distribution) based on Hypergeometric test. Based on physiological roles and functions, pathways and terms were grouped into different categories. The enrichment of up- and down-regulated genes was separately analyzed. The significant levels of pathways and terms were corrected by a false discovery rate (FDR) adjusted P value (Q value) with a threshold of Q value ≤0.05 by Bonferroni.34 Only those GO terms and KEGG pathways with Q value ≤0.05 were defined as significant ones.
Results
Epidermal Gene Expression in Mice Exposed to Low Humidity with or without Emollient Treatment
We first assessed whether exposure of mice to low humidity can alter epidermal gene expression. Indeed, exposure of mice to low humidity for 6 months altered a whole panel of epidermal gene expression compared to mice living in normal humidity environment (Figure 1). To overview the variance of gene expression among the groups, we performed principal component (PC) analysis. As seen in Figure 2, PC1 and PC2 explained 44.5% and 11.4% of the total variance, respectively, totaling 55.9% of variance. No significant batch effects were observed in the principal component analysis.
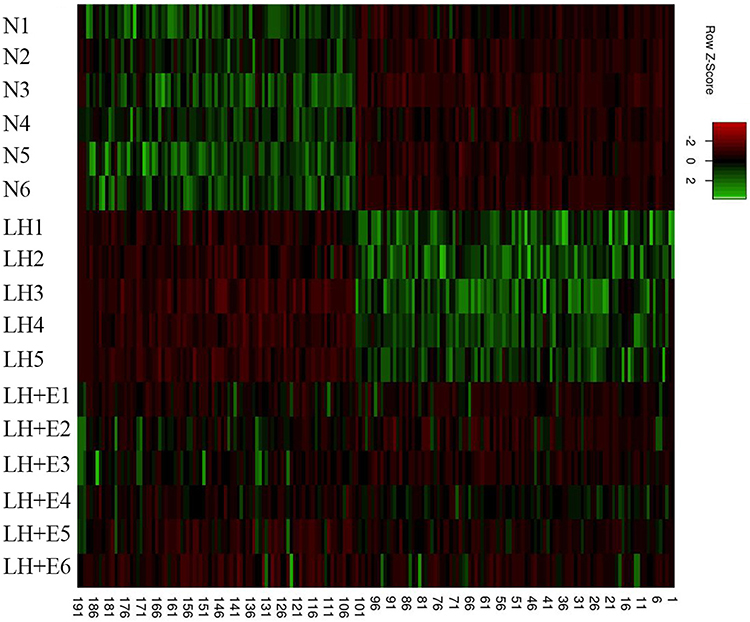 |
Figure 1 Heatmap of gene expression. N1-N6 represent date of samples from mice without any treatment and were housed in normal humidity environment; LH1-LH5 show data of samples from mice exposed to low humidity; LH+E1 – LH+E6 are data of samples from mice exposed to low humidity and treated with emollient. |
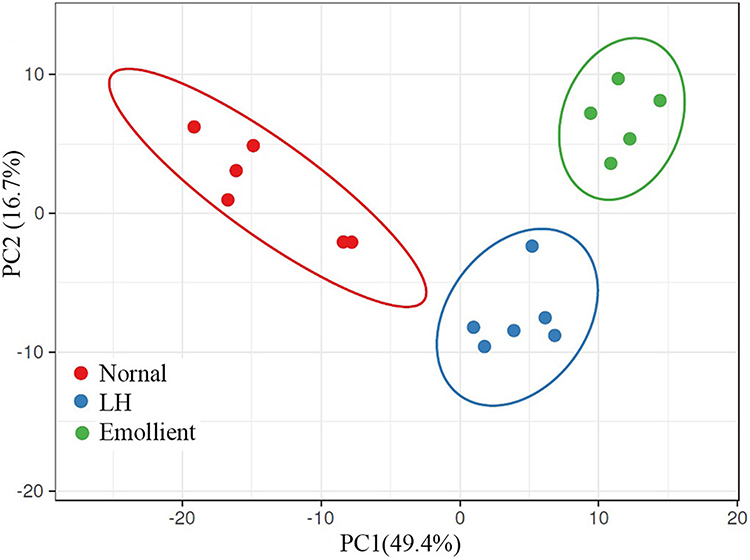 |
Figure 2 The results of the principal component analysis. (N) Mice without any treatment were housed in normal humidity environment; LH: Mice were exposed to low humidity; Emollient: Mice were exposed to low humidity and treated with emollient. |
Then we identified differentially expressed genes (DEG) (|log2 fold change| ≥1.0, Q value<0.05) between the groups. Overall, there were 264 DEGs, including 105 upregulated and 159 downregulated genes, in mice exposed to low humidity vs those living in normal humidity environment, and 195 upregulated and 280 downregulated genes in mice exposed to low humidity with vs without treatment with emollient (Figure 3). Interestingly, treatments with emollient could partially reverse the changes in expression levels of some genes altered by low humidity (Figure S1). For example, low humidity markedly upregulatthe expressionion of serum amyloid A2 (saa2) (log2 4.8-fold increase vs normal, Q=0.001381), which is linked to inflammation. Treatments with emollient downregulated saa2 expression (log2 5-fold decrease vs low humidity alone, Q<0.001). Similarly, low humidity upregulated expression levels of Gm5414 (log2 3.2-fold increase vs normal). Topical emollient downregulated this gene (log2 2.3-fold decrease vs low humidity alone). In contrast, low humidity downregulated expression levels of serpinb1c (log2 1.9-fold decrease vs normal), while topical emollient treatment upregulated serpinb1c expression (log2 2.8-fold increase vs low humidity alone). Consistent with previous studies,35 the present study demonstrated that low humidity increased expression levels of inflammatory cytokines (such as CXCL1, CXCL13, IL-1β, IL-6, IL-24). Topical emollient markedly downregulated the expression of these cytokines in mice exposed to low humidity. Another worth noting finding was the increase in epidermal very-long-chain fatty acid elongase 3 (Elovl3) in emollient treated mice (log2=2 fold vs low humidity alone, Q<0.0001). Elovl3 is involved in the synthesis of long chain fatty acid, a key component for epidermal permeability barrier.36 Moreover, emollient upregulated expression of acyl-CoA:diacylglycerol acyltransferase gene 1 (DGAT1) (log2=1.4 fold vs low humidity alone, Q<0.0001). Taken together, these results demonstrate that topical emollient can partially reverse some epidermal gene expression that are altered by low humidity.
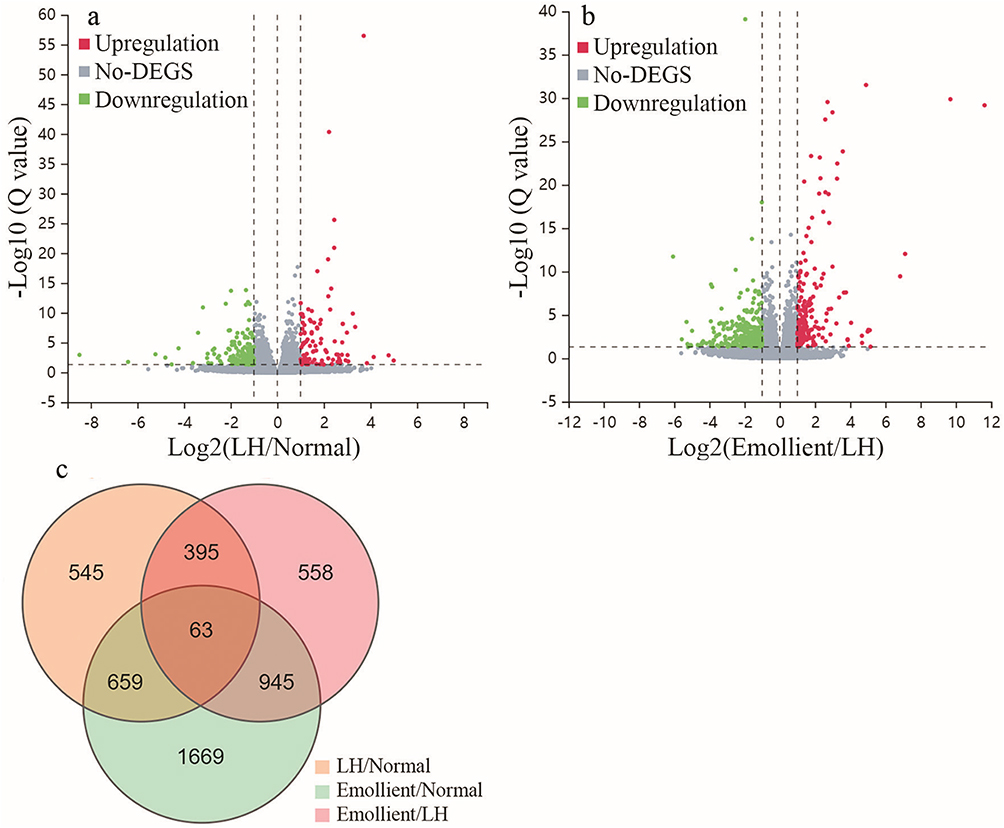 |
Figure 3 Differentially Expressed Genes in the Epidermis of Mice Exposed to Low Humidity. Significantly and differentially expressed genes (DEGs) are defines as those genes with log2 ≥1-fold changes and Q≤0.05. (a) Volcano plot of DEGs in mice exposed to low humidity (LH) vs normal controls; (b) Volcano plot of DEGs in mice exposed to LH-treated with vs without treatment with the cream; (c) Venn diagram of DEGs in each group. N=6 for normal control and emollient-treated group; N=5 for low humidity group. Abbreviation: LH, low humidity. |
Functional and Pathway Enrichment Analyses of DEGs
To reveal the functional significance of DEGs in the epidermis of mice exposed to low humidity, we performed gene ontology (GO) analysis. As seen in Figure 4a, low humidity prominently downregulated genes enriched in regulation of transcription and melanin metabolism in comparison to normal controls. Treatments with emollient upregulated genes enriched in lipid and sterol biosynthesis in mice exposed to low humidity (Figure 4a). Among the significantly downregulated genes, the numbers of genes enriched in regulation of transcription and immune response process were 25 and 11, respectively (Figure 4a). Topical emollient increased expression of 32 genes associated with lipid metabolism. Low humidity upregulated genes mainly enriched in GO terms of keratinization (15 genes, rich ratio=0.2459) and keratinocyte differentiation (9 genes, rich ratio=0.12) (Figure 4b). Whereas, topical emollient downregulated genes enriched in GO terms of keratinization (18 genes, rich ratio=0.295) and keratinocyte differentiation (15 genes, rich ratio=0.2) in mice exposed to low humidity. Moreover, topical emollient also downregulated genes enriched in inflammation and immune response, including inflammatory response (28 genes, rich ratio=0.074) and immune system process (27 genes, rich ratio=0.0515) in the epidermis of mice exposed to low humidity.
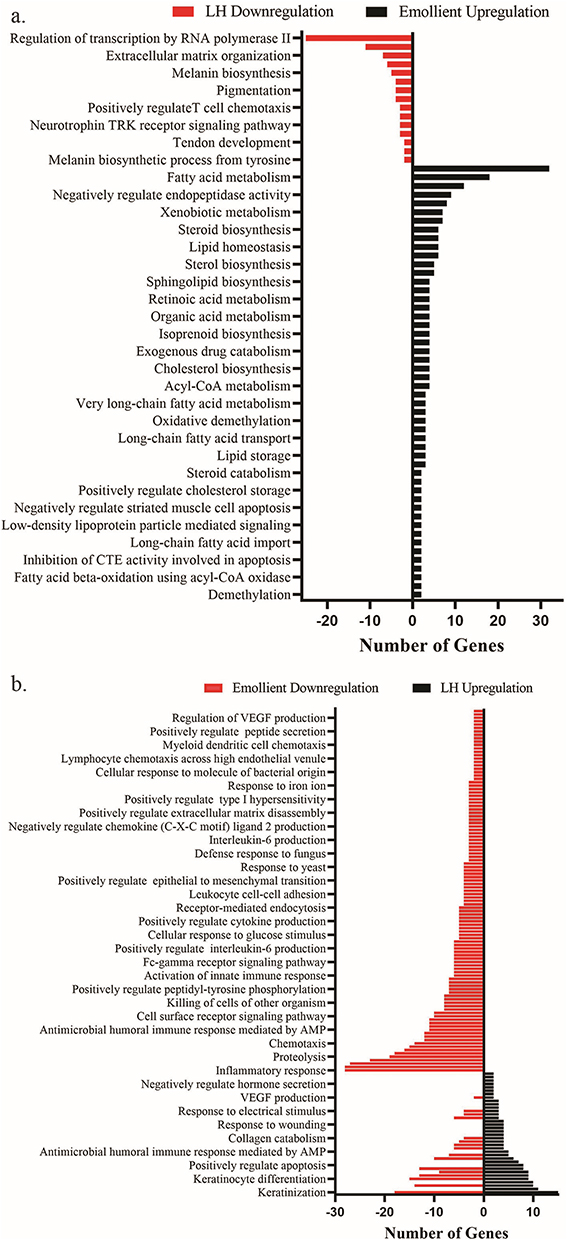 |
Figure 4 GO Analyses of Low Humidity-Induced Changes in Epidermal Gene Expression. (a) Gene ontology (GO) analysis of genes downregulated by low humidity compared to normal controls and genes upregulated by emollient; (b) Genes upregulated by low humidity compared to normal controls and genes downregulated by emollient. Q value <0.05 for all. Abbreviations: RTK, Receptor tyrosine kinase; CTE, Cysteins-type endopeptidase; AMP, Antimicrobial peptide; VEGF, Vascular endothelial growth factor. |
We analyzed next the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways of DEGs. In comparison to normal controls, low humidity downregulated genes associated with signaling pathways of viral protein interaction with cytokine-cytokine receptor (7 genes, rich ratio=0.074468), cytokine-cytokine receptor interaction (11 genes, rich ratio=0.037801), and herpes simplex virus 1 infection (19 genes, rich ratio=0.042222) (Figure 5a). Treatments with emollient significantly upregulated genes enriched in the signaling pathways associated with retinol metabolism (9 genes, rich ratio=0.092784), peroxisome proliferator-activated receptor (PPAR) (6 genes, rich ratio=0.067416) and fatty acid metabolism (15 genes, rich ratio=0.086207) in the epidermis of mice exposed to low humidity. Additionally, low humidity-upregulated genes were mainly enriched in the signaling pathways linked to IL-17 (11 genes, rich ratio=0.120879), TNF (8 genes, rich ratio=0.070796) and cytokine-cytokine receptor (6 genes, rich ratio=0.020619) signaling pathways (Figure 5b). Interestingly, treatment with emollient downregulated genes enriched in the signaling pathways linked to inflammatory response, which were upregulated by low humidity. For example, emollient downregulated 17 genes associated with IL-17 signaling pathway in the mouse epidermis exposed to low humidity (Figure 5b). These results indicate that low humidity regulates epidermal gene expression, which can be partially reversed by topical emollient, in mice, suggesting the importance of emollient in maintaining epidermal functions in low humidity environment.
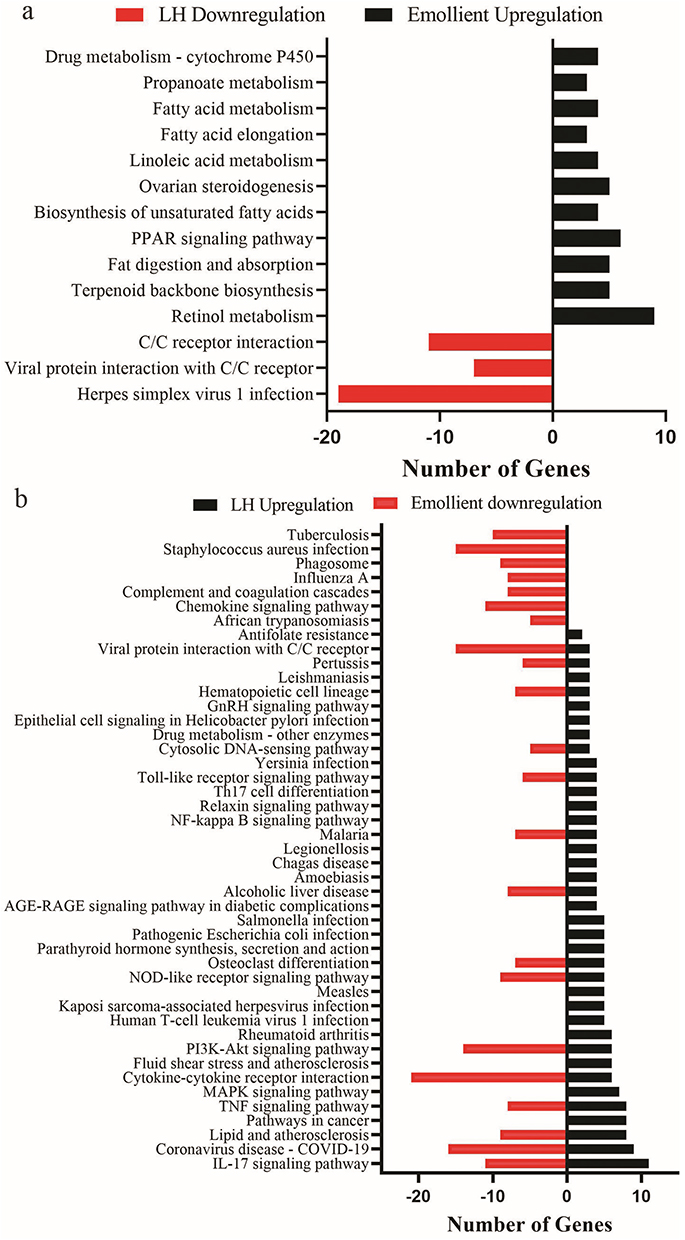 |
Figure 5 KEGG Anayses of Changes in Epidermal Gene Expression. (a) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways that are significantly downregulated by low humidity and upregulated by emollient in mice exposed to low humidity; (b) KEGG pathways that are significantly upregulated by low humidity and downregulated by emollient in mice exposure to low humidity. Q value <0.05 for all. Abbreviation: C/C, Cytokine and cytokine. |
Discussion
Although emollients have been widely used in the management of various cutaneous conditions, influences of emollients on epidermal gene expression are poorly understood. We show here that topical applications of an emollient regulated a number of gene expression in mice exposed to low humidity. In agreement with previous findings that topical applications of an emollient improve cutaneous inflammation, stratum hydration and epidermal permeability barrier in several skin conditions such as psoriasis, atopic dermatitis, chronic eczema, and pruritus hiemalis,26,27 we show here that topical application of the emollient markedly upregulated PPAR pathway in mice exposed to low humidity. Other worth noting KEGG pathways upregulated by topical emollient were terpenoid backbone biosynthesis and fatty acid elongation with rich ratios of 0.21 (Q<0.001) and 0.1 (Q<0.05), respectively. PPAR, terpenoid backbone biosynthesis and fatty acid elongation are involved in epidermal lipid synthesis, a requirement for epidermal permeability barrier function. In addition, emollient upregulated expression of Dgat1, a key enzyme in triglyceride synthesis,37 while triglyceride can be hydrolyzed to glycerol. The latter enhances the stratum corneum hydration and epidermal permeability barrier function.38,39 The underlying mechanisms by which topical emollient regulate epidermal gene expression are not clear. It is possible that the linoleic acid enriched ingredients such as avocado oil in the cream contribute to the upregulation of PPAR signaling pathway. Previous studies demonstrated that activation of PPAR increases epidermal lipid synthesis, while inhibiting cutaneous inflammation.40,41 Hence, topical applications of this emollient upregulated PPAR signaling, leading to downregulation of expression of genes related to inflammation, upregulation of epidermal lipid production and differentiation in mice exposed to low humidity. Of course, emollient-induced elevations in stratum corneum hydration can also contribute to downregulation of inflammation-related signaling pathways. However, whether other moisturizers have effects comparable to the one used in the present study remains unknown. Nonetheless, the present study clearly shows that low humidity alters epidermal gene expression, which can be partially reversed by topical emollient in mice. Since the activation of PPAR can enhance the epidermal permeability barrier and stratum corneum hydration, while inhibiting inflammation, emollients containing PPAR activators, such as fatty acids, may be a valuable addition to the management of certain skin conditions, including seasonal dry skin.
Previous studies showed that exposure of mice to low humidity increases inflammatory infiltrates and mast cell degranulation.18 Similarly, epidermal cytokine levels in humans are increased in dry environment.22 However, the transcriptional basis for these changes is not clear. We demonstrate here that low humidity increased expression levels of a number of genes enriched in the signaling pathways associated with inflammation in mice. The most noticeable signaling pathways upregulated by low humidity were IL-17 and TNF, providing a transcriptional basis for cutaneous inflammation in dry environment. Other signaling pathways, including cytokine-cytokine interaction, IL-17 cell differentiation and cytosolic DNA-sensing pathways, were also markedly upregulated by low humidity. Among these upregulated pathways, IL-17 pathway is particularly clinically relevant to psoriasis, which often relapses in dry seasons such as winter and spring.42,43 Although low temperature can increase the risk for psoriasis,43,44 temperature unlikely accounts for the increased expression of inflammation-related genes in mice exposed to low humidity because the temperature was the same among those mice. Instead, low humidity can be the major contributor to the changes in epidermal gene expression, including inflammatory response-related genes, again providing a transcriptional basis for onset and/exacerbation of psoriasis in low humidity seasons. Thus, improvements in epidermal functions, including stratum corneum hydration levels, can mitigate inflammatory dermatoses such as psoriasis. This assumption is supported by the evidence that a. more systemic and biologic treatments are deployed to treat psoriasis in spring, a low humidity season;45 b. topical emollient downregulated expression levels of a number of immune-related genes as shown here and inflammatory infiltrates in mice exposed to low humidity18 and c. topical emollient prevents the development of psoriasis.46–48 Moreover, low humidity decreases stratum corneum hydration levels,49 which negatively correlates with serum levels of pro-inflammatory cytokines in humans.50 Chronic inflammation is linked to the pathogenesis of some systemic disorders, including obesity, cardiovascular disease, type 2 diabetes and neural disorders.51 Accordingly, topical emollient lowers serum levels of proinflammatory cytokines in aged humans and mice, and mRNA levels for proinflammatory cytokines in the mouse skin.52,53 Hence, improvement in stratum corneum hydration can downregulate expression of inflammation-related genes in the epidermis, possibly leading to mitigation of some inflammation-associated systemic disorders. The results of the present study also suggest that the underlying mechanisms by which topical emollients ameliorate pruritus hiemalis may involve the downregulation of inflammatory pathways.
In summary, low humidity alters epidermal gene expression in mice, including significant upregulation of serum amyloid A2, Gm5414, and proinflammatory cytokines while down-regulating serpinb1c gene. Topical applications of an emollient can partially reverse low humidity-induced changes in expression levels of some epidermal genes. Moreover, topical emollient significantly upregulates Elovl3 and DGAT1 expression, likely via activation of PPAR. The findings of the present study suggest that utilizing ingredients that target PPAR in emollients can be a valuable approach in managing skin conditions characterized by dry skin and inflammation. However, whether other emollients can also regulate epidermal gene expression remains to be elucidated. In addition, further studies are needed to determine the functional significance of low humidity-induced changes in epidermal gene expression with/without emollient.
Data Sharing Statement
Data are deposited in Science Data Bank, entitled “Low humidity regulates gene expression” (https://www.scidb.cn/en/datalist), with free access.
Ethics Approval
This work was reviewed and approved by the ethics committee of Guangdong Medical Laboratory Animal Center (D202410-2).
Disclosure
All authors declare no conflicts of interest in this work.
References
1. Liu Z, Fluhr JW, Song SP, et al. Sun-induced changes in stratum corneum function are gender and dose dependent in a Chinese population. Skin Pharmacol Physiol. 2010;23(6):313–12. doi:10.1159/000314138
2. Liu Z, Song S, Luo W, Elias PM, Man MQ. Sun-induced changes of stratum corneum hydration vary with age and gender in a normal Chinese population. Skin Res Technol. 2012;18(1):22–28. doi:10.1111/j.1600-0846.2011.00536.x
3. Ahn K. The role of air pollutants in atopic dermatitis. J Allergy Clin Immunol. 2014;134(5):993–999. discussion 1000. doi:10.1016/j.jaci.2014.09.023
4. Burke KE. Mechanisms of aging and development-A new understanding of environmental damage to the skin and prevention with topical antioxidants. Mech Ageing Dev. 2018;172:123–130. doi:10.1016/j.mad.2017.12.003
5. Kamiya K, Kishimoto M, Sugai J, Komine M, Ohtsuki M. Risk factors for the development of psoriasis. Int J Mol Sci. 2019;20(18):4347. doi:10.3390/ijms20184347
6. Ferrara F, Prieux R, Woodby B, Valacchi G. Inflammasome activation in pollution-induced skin conditions. Plast Reconstr Surg. 2021;147(1S–2):15S–24S. doi:10.1097/PRS.0000000000007617
7. Jin SP, Li Z, Choi EK, et al. Urban particulate matter in air pollution penetrates into the barrier-disrupted skin and produces ROS-dependent cutaneous inflammatory response in vivo. J Dermatol Sci. 2018;S0923–1811(18)30202–0. doi:10.1016/j.jdermsci.2018.04.015
8. Verdin A, Cazier F, Fitoussi R, et al. An in vitro model to evaluate the impact of environmental fine particles (PM0.3-2.5) on skin damage. Toxicol Lett. 2019;305:94–102. doi:10.1016/j.toxlet.2019.01.016
9. Dijkhoff IM, Drasler B, Karakocak BB, et al. Impact of airborne particulate matter on skin: a systematic review from epidemiology to in vitro studies. Part Fibre Toxicol. 2020;17(1):35. doi:10.1186/s12989-020-00366-y
10. Kim BE, Kim J, Goleva E, et al. Particulate matter causes skin barrier dysfunction. JCI Insight. 2021;6(5):e145185. doi:10.1172/jci.insight.145185
11. Denda M, Sokabe T, Fukumi-Tominaga T, Tominaga M. Effects of skin surface temperature on epidermal permeability barrier homeostasis. J Invest Dermatol. 2007;127(3):654–659. doi:10.1038/sj.jid.5700590
12. Herrero-Fernandez M, Montero-Vilchez T, Diaz-Calvillo P, Romera-Vilchez M, Buendia-Eisman A, Arias-Santiago S. Impact of water exposure and temperature changes on skin barrier function. J Clin Med. 2022;11(2):298. doi:10.3390/jcm11020298
13. Halkier-Sørensen L, Menon GK, Elias PM, Thestrup-Pedersen K, Feingold KR. Cutaneous barrier function after cold exposure in hairless mice: a model to demonstrate how cold interferes with barrier homeostasis among workers in the fish-processing industry. Br J Dermatol. 1995;132(3):391–401. doi:10.1111/j.1365-2133.1995.tb08672.x
14. Halkier-Sørensen L, Thestrup-Pedersen K. Skin physiological changes in employees in the fish processing industry immediately following work. A field study. Contact Dermatitis. 1991;25(1):19–24. doi:10.1111/j.1600-0536.1991.tb01767.x
15. Halkier-Sørensen L, Thestrup-Pedersen K. The relationship between skin surface temperature, transepidermal water loss and electrical capacitance among workers in the fish processing industry: comparison with other occupations. A field study. Contact Dermatitis. 1991;24(5):345–355. doi:10.1111/j.1600-0536.1991.tb01749.x
16. Rycroft RJ. Low humidity and microtrauma. Am J Ind Med. 1985;8(4–5):371–373. doi:10.1002/ajim.4700080417
17. Denda M, Sato J, Masuda Y, et al. Exposure to a dry environment enhances epidermal permeability barrier function. J Invest Dermatol. 1998;111(5):858–863. doi:10.1046/j.1523-1747.1998.00333.x
18. Denda M, Sato J, Tsuchiya T, Elias PM, Feingold KR. Low humidity stimulates epidermal DNA synthesis and amplifies the hyperproliferative response to barrier disruption: implication for seasonal exacerbations of inflammatory dermatoses. J Invest Dermatol. 1998;111(5):873–878. doi:10.1046/j.1523-1747.1998.00364.x
19. Sato J, Denda M, Chang S, Elias PM, Feingold KR. Abrupt decreases in environmental humidity induce abnormalities in permeability barrier homeostasis. J Invest Dermatol. 2002;119(4):900–904. doi:10.1046/j.1523-1747.2002.00589.x
20. Chou TC, Lin KH, Wang SM, et al. Transepidermal water loss and skin capacitance alterations among workers in an ultra-low humidity environment. Arch Dermatol Res. 2005;296(10):489–495. doi:10.1007/s00403-005-0541-4
21. Chou TC, Lin KH, Sheu HM, et al. Alterations in health examination items and skin symptoms from exposure to ultra-low humidity. Int Arch Occup Environ Health. 2007;80(4):290–297. doi:10.1007/s00420-006-0133-4
22. Sato M, Fukayo S, Yano E. Adverse environmental health effects of ultra-low relative humidity indoor air. J Occup Health. 2003;45(2):133–136. doi:10.1539/joh.45.133
23. Kikuchi K, Kobayashi H, Hirao T, Ito A, Takahashi H, Tagami H. Improvement of mild inflammatory changes of the facial skin induced by winter environment with daily applications of a moisturizing cream. A half-side test of biophysical skin parameters, cytokine expression pattern and the formation of cornified envelope. Dermatology. 2003;207(3):269–275. doi:10.1159/000073089
24. Man MQ, Xin SJ, Song SP, et al. Variation of skin surface pH, sebum content and stratum corneum hydration with age and gender in a large Chinese population. Skin Pharmacol Physiol. 2009;22(4):190–199. doi:10.1159/000231524
25. Cottam NC, Ofori K, Stoll KT, et al. From circuits to lifespan: translating mouse and human timelines with neuroimaging-based tractography. J Neurosci. 2025;45(12):e1429242025. doi:10.1523/JNEUROSCI.1429-24.2025
26. Liu M, Li X, Chen XY, Xue F, Zheng J. Topical application of a linoleic acid-ceramide containing moisturizer exhibit therapeutic and preventive benefits for psoriasis vulgaris: a randomized controlled trial. Dermatol Ther. 2015;28(6):373–382. doi:10.1111/dth.12259
27. Yang Q, Liu M, Li X, Zheng J. The benefit of a ceramide-linoleic acid-containing moisturizer as an adjunctive therapy for a set of xerotic dermatoses. Dermatol Ther. 2019;32(4):e13017. doi:10.1111/dth.13017
28. Mao-Qiang M, Elias PM, Feingold KR. Fatty acids are required for epidermal permeability barrier function. J Clin Invest. 1993;92(2):791–798. doi:10.1172/JCI116652
29. Li R, Li Y, Kristiansen K, Wang J. SOAP: short oligonucleotide alignment program. Bioinformatics. 2008;24(5):713–714. doi:10.1093/bioinformatics/btn025
30. Kim D, Langmead B, Salzberg SL. HISAT: a fast spliced aligner with low memory requirements. Nat Methods. 2015;12(4):357–360. doi:10.1038/nmeth.3317
31. Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012;9(4):357–359. doi:10.1038/nmeth.1923
32. Li B, Dewey CN. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinf. 2011;12(1):323. doi:10.1186/1471-2105-12-323
33. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15(12):550. doi:10.1186/s13059-014-0550-8
34. Abdi H. The bonferonni and sˇidák corrections for multiple comparisons. Encycl Meas Stat. 2007;1:1–9.
35. Ashida Y, Denda M. Dry environment increases mast cell number and histamine content in dermis in hairless mice. Br J Dermatol. 2003;149(2):240–247. doi:10.1046/j.1365-2133.2003.05408.x
36. Westerberg R, Tvrdik P, Undén AB, et al. Role for ELOVL3 and fatty acid chain length in development of hair and skin function. J Biol Chem. 2004;279(7):5621–5629. doi:10.1074/jbc.M310529200
37. Yen CL, Stone SJ, Koliwad S, Harris C, Farese RV. Thematic review series: glycerolipids. DGAT enzymes and triacylglycerol biosynthesis. J Lipid Res. 2008;49(11):2283–2301. doi:10.1194/jlr.R800018-JLR200
38. Choi EH, Man MQ, Wang F, et al. Is endogenous glycerol a determinant of stratum corneum hydration in humans? J Invest Dermatol. 2005;125(2):288–293. doi:10.1111/j.0022-202X.2005.23799.x
39. Fluhr JW, Gloor M, Lehmann L, Lazzerini S, Distante F, Berardesca E. Glycerol accelerates recovery of barrier function in vivo. Acta Derm Venereol. 1999;79(6):418–421. doi:10.1080/000155599750009825
40. Schmuth M, Jiang YJ, Dubrac S, Elias PM, Feingold KR. Thematic review series: skin lipids. Peroxisome proliferator-activated receptors and liver X receptors in epidermal biology. J Lipid Res. 2008;49(3):499–509. doi:10.1194/jlr.R800001-JLR200
41. Sheu MY, Fowler AJ, Kao J, et al. Topical peroxisome proliferator activated receptor-alpha activators reduce inflammation in irritant and allergic contact dermatitis models. J Invest Dermatol. 2002;118(1):94–101. doi:10.1046/j.0022-202x.2001.01626.x
42. Monin L, Gaffen SL. Interleukin 17 family cytokines: signaling mechanisms, biological activities, and therapeutic implications. Cold Spring Harb Perspect Biol. 2018;10(4):a028522. doi:10.1101/cshperspect.a028522
43. Pan R, Zhang J. Epidemiology and treatment of psoriasis: a Chinese perspective. Psoriasis: Targets and Ther. 2014;37–47.
44. Sun H, Wang X, Zhang X, Wang L, Zou Y, Hu H. The association between apparent temperature and psoriasis outpatient visits: a time-series study in Hefei, China. Environ Res Commun. 2023;5(3):035003. doi:10.1088/2515-7620/acc013
45. Liang H, Kirk B, Polinski JM, Yue X, Kilpatrick RD, Gelfand JM. Impact of season and other factors on initiation, discontinuation, and switching of systemic drug therapy in patients with psoriasis: a retrospective study. JID Innov. 2022;3(2):100171. doi:10.1016/j.xjidi.2022.100171
46. Li X, Yang Q, Zheng J, et al. Efficacy and safety of a topical moisturizer containing linoleic acid and ceramide for mild-to-moderate psoriasis vulgaris: a multicenter randomized controlled trial. Dermatol Ther. 2020;33(6):e14263. doi:10.1111/dth.14263
47. Man MQ, Ye L, Hu L, Jeong S, Elias PM, Lv C. Improvements in epidermal function prevent relapse of psoriasis: a self-controlled study. Clin Exp Dermatol. 2019;44(6):654–657. doi:10.1111/ced.13888
48. Seité S, Khemis A, Rougier A, Ortonne JP. Emollient for maintenance therapy after topical corticotherapy in mild psoriasis. Exp Dermatol. 2009;18(12):1076–1078. doi:10.1111/j.1600-0625.2009.00903.x
49. Tagami H, Kobayashi H, Zhen XS, Kikuchi K. Environmental effects on the functions of the stratum corneum. J Investig Dermatol Symp Proc. 2001;6(1):87–94. doi:10.1046/j.0022-202x.2001.00016.x
50. Yang B, Lv C, Ye L, et al. Stratum corneum hydration inversely correlates with certain serum cytokine levels in the elderly, possibly contributing to inflammaging. Immun Ageing. 2023;20(1):7. doi:10.1186/s12979-023-00331-1
51. Man MQ, Elias PM. Could inflammaging and its sequelae be prevented or mitigated? Clin Interv Aging. 2019;14:2301–2304. doi:10.2147/CIA.S235595
52. Hu L, Mauro TM, Dang E, et al. Epidermal dysfunction leads to an age-associated increase in levels of serum inflammatory cytokines. J Invest Dermatol. 2017;137(6):1277–1285. doi:10.1016/j.jid.2017.01.007
53. Ye L, Mauro TM, Dang E, et al. Topical applications of an emollient reduce circulating pro-inflammatory cytokine levels in chronically aged humans: a pilot clinical study. J Eur Acad Dermatol Venereol. 2019;33(11):2197–2201. doi:10.1111/jdv.15540
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.