Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 8
Lixisenatide accelerates restoration of normoglycemia and improves human beta-cell function and survival in diabetic immunodeficient NOD–scid IL-2rgnull RIP-DTR mice engrafted with human islets
Authors Yang C, Loehn M, Jurczyk A, Przewozniak N, Leehy L, Herrera P, Shultz L, Greiner D, Harlan D, Bortell R
Received 25 April 2015
Accepted for publication 3 June 2015
Published 20 August 2015 Volume 2015:8 Pages 387—398
DOI https://doi.org/10.2147/DMSO.S87253
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Ming-Hui Zou
Chaoxing Yang,1 Matthias Loehn,2 Agata Jurczyk,1 Natalia Przewozniak,1 Linda Leehy,1 Pedro L Herrera,3 Leonard D Shultz,4 Dale L Greiner,1 David M Harlan,5 Rita Bortell1
1Program in Molecular Medicine, Diabetes Center of Excellence, University of Massachusetts Medical School, Worcester, MA, USA; 2Sanofi-Aventis, Diabetes Division, Frankfurt, Germany; 3University of Geneva, Geneva, Switzerland; 4The Jackson Laboratory, Bar Harbor, ME, USA; 5Department of Medicine, Diabetes Center of Excellence, University of Massachusetts Medical School, Worcester, MA, USA
Objective: Glucagon-like peptide-1 induces glucose-dependent insulin secretion and, in rodents, increases proliferation and survival of pancreatic beta cells. To investigate the effects on human beta cells, we used immunodeficient mice transplanted with human islets. The goal was to determine whether lixisenatide, a glucagon-like peptide-1 receptor agonist, improves human islet function and survival in vivo.
Methods: Five independent transplant studies were conducted with human islets from five individual donors. Diabetic human islet-engrafted immunodeficient mice were treated with lixisenatide (50, 150, and 500 µg/kg) or vehicle. Islet function was determined by blood glucose, plasma human insulin/C-peptide, and glucose tolerance tests. Grafts were analyzed for total beta- and alpha-cell number, percent proliferation, and levels of apoptosis.
Results: Diabetic mice transplanted with marginal human islet mass and treated with lixisenatide were restored to euglycemia more rapidly than vehicle-treated mice. Glucose tolerance tests, human plasma insulin, and glucose-stimulation indices of lixisenatide-treated mice were significantly improved compared to vehicle-treated mice. The percentages of proliferating or apoptotic beta cells at graft recovery were not different between lixisenatide-treated and vehicle-treated mice. Nevertheless, in one experiment we found a significant twofold to threefold increase in human beta-cell numbers in lixisenatide-treated compared to vehicle-treated mice.
Conclusion: Diabetic human islet-engrafted immunodeficient mice treated with lixisenatide show improved restoration of normoglycemia, human plasma insulin, and glucose tolerance compared to vehicle-treated mice engrafted with the same donor islets. Because the proliferative capacity of human beta cells is limited, improved beta-cell survival coupled with enhanced beta-cell function following lixisenatide treatment may provide the greatest benefit for diabetic patients with reduced functional islet mass.
Keywords: GLP-1 receptor agonist, lixisenatide, human islet transplant, beta cells, glucose tolerance tests, plasma insulin
Introduction
Glucagon-like peptide-1 (GLP-1) and GLP-1 receptor agonists have been reported to improve beta-cell function and viability.1–3 At the beta-cell level, GLP-1 and its receptor agonists were found to induce beta-cell proliferation and decrease beta-cell apoptosis in rodents and in vitro.3–6 In vivo, GLP-1 receptor agonists preserve beta-cell mass in multiple animal models of diabetes,3,7–9 although an effect on beta-cell mass/number has not yet been demonstrated in humans. In clinical trials with type 2 diabetes (T2D) patients, GLP-1 receptor agonists lowered both fasting and postprandial glucose concentrations;10,11 GLP-1 receptor agonists also potentiate glucose-dependent insulin secretion, and thus have a low propensity to cause hypoglycemia.12 However, in patients with long-standing type 1 diabetes, C-peptide secretion was not increased, although insulin sensitivity improved.13,14
Lixisenatide is a recently developed GLP-1 receptor agonist with potent binding affinity and extended biological activity.15–18 In patients with T2D, once-daily administration of lixisenatide improved glycemic control by decreasing both postprandial and fasting glucose levels; moreover, these effects were both immediate and sustained.19–22 In addition to increased insulin secretion in response to meal-related or glucose stimulation, a reduction in endogenous glucose production and slowing of gastric emptying also contribute to the reduction in postprandial blood glucose levels observed with lixisenatide and other GLP-1 receptor agonists.22–25 In vitro studies with an INS-1 rat pancreatic beta-cell line indicates that GLP-1 and its receptor agonists, including lixisenatide, protect against lipid- and cytokine-induced apoptosis.26 Even so, it is not possible to directly assess beta-cell apoptosis in patients due to the inaccessibility of pancreatic islets, and clinical studies are difficult to control due to the variability between patients in regards to age, sex, diet, and other lifestyle factors.
A useful method to interrogate human islet function in vivo is to transplant human islets into diabetic immunodeficient nonobese diabetic–severe combined immunodeficiency (NOD–scid) IL-2 receptor common gamma chain (IL-2rgnull) (NSG) mice.27 Many diabetes-inducing chemicals, such as streptozotocin, may cause damage to other organs. Also, the chemical-induced destruction of endogenous beta cells is not always complete, rendering problematic the interpretation of long-term transplantation studies with exogenous islets. To circumvent these issues, we utilized a strain of transgenic NSG mice that uses the rat insulin promoter (RIP) to drive human diphtheria toxin receptor (DTR) expression in the animal’s beta cells. When treated with low doses of diphtheria toxin, the NSG RIP-DTR mouse model allows complete and specific ablation of mouse pancreatic beta cells and thereby avoids broadly toxic agents such as streptozotocin. The goal of this study was to investigate the efficacy of lixisenatide to promote human beta-cell function, proliferation, and survival using diabetic NSG RIP-DTR mice engrafted with marginal amounts of human pancreatic islets.
Methods
Mice and diabetes induction
NOD.Cg-PrkdcscidIl2rgtm1Wjl Tg(Ins2-HBEGF)6832 Ugfm/Sz mice, referred to as NSG RIP-DTR mice, were developed at the Jackson Laboratory, Bar Harbor, ME, USA, by backcrossing the RIP-DTR transgene from a B6;CBA-RIP-DTR stock kindly provided by Pedro Herrera. The original B6;CBA Tg(Ins2-HBEGF)6832 Ugfm/Sz mice were made by injecting the construct into B6;CBA eggs. The transgene was backcrossed using a marker-assisted speed congenic method to the NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ (abbreviated as NOD–scid IL-2rgnull or NSG) strain background. These NSG RIP-DTR mice express the human DTR driven by a RIP. The RIP-DTR transgene was then fixed to homozygosity. All mice were housed in a specific pathogen-free facility and maintained in accordance with the Institutional Animal Care and Use Committee of the University of Massachusetts Medical School.28
To induce diabetes, male NSG RIP-DTR mice (8–12 weeks old) were injected intraperitoneally (ip) with 20 ng diphtheria toxin (List Biological Laboratories, Campbell, CA, USA) diluted in sterile phosphate-buffered saline (PBS). Blood glucose was monitored with an Accu-chek Aviva Plus glucometer (Hoffman-La Roche Ltd, Basel, Switzerland) to confirm diabetes (blood glucose >300 mg/dL on 2 consecutive days). Diabetic mice that were not transplanted with human islets within 1 week were given insulin implants (LinShin Canada Inc., Scarborough, ON, Canada) to prevent metabolic decompensation until human islets were available.
Pharmacokinetic analyses
An initial pharmacokinetic study with unengrafted, euglycemic NSG mice was performed to determine the plasma levels of lixisenatide over a 24-hour period following treatment. All mice were injected subcutaneously (sc) with vehicle alone or with 50 μg/kg, 150 μg/kg, or 500 μg/kg lixisenatide (provided by Sanofi-Aventis, Frankfurt, Germany). Blood was collected in potassium ethylenediaminetetraacetic acid (K-EDTA) tubes at 0, 5, 15, 30, 60, and 120 minutes and 4, 8, and 24 hours from three mice/group at each time point; plasma was stored at −80°C. High-performance liquid chromatography analysis of lixisenatide levels in blood plasma was performed by Sanofi-Aventis. Based on these time-course data (Figure S1), the mice in the transplant studies were treated twice daily with the same concentrations of lixisenatide as in the pharmacokinetic study.
Human islet transplantation and lixisenatide treatment
Human islets were obtained from the Integrated Islet Distribution Program under protocols approved by the Institutional Review Board of the University of Massachusetts Medical School. Islets were transplanted into the subrenal capsular space as previously described;29 insulin implants were removed upon transplant. Five independent transplant studies were performed, each with human islets from a single donor. One day post-transplant, the mice were randomized into four groups, with five to seven mice in each group. The mice were injected sc twice daily with lixisenatide (50, 150, or 500 μg/kg/injection) or vehicle until graft removal at ~4 weeks post-transplant.
Glucose tolerance test, plasma insulin/C-peptide, and glucose stimulation index
For the glucose tolerance test (GTT), mice were fasted for 5–6 hours, and blood glucose was measured following ip injection of glucose (2.0 g/kg body weight). To measure plasma levels of human insulin and C-peptide, heparinized blood from transplanted mice was collected with protease inhibitor (aprotinin; Sigma-Aldrich Co, St Louis, MO, USA). Non-fasting blood samples were collected just prior to drug/vehicle treatment. On alternate weeks, the mice were fasted for 5–6 hours prior to glucose injection (2 g/kg, ip); in the Donor 1 study, arginine (1 g/kg, ip) was given in addition to glucose. Blood was collected at 0 (fasted) and 15 minutes (stimulated) post-injection; the glucose stimulation index was determined as the ratio of plasma insulin at 15 and 0 minutes. All plasma was stored at −80°C until analyzed by human-specific enzyme-linked immunosorbent assay (ELISA) (ALPCO Diagnostics, Salem, NH, USA).
Bromodeoxyuridine treatment, immunofluorescence staining, and TUNEL assay
Human islet-engrafted mice were provided drinking water containing 0.8 mg/mL of bromodeoxyuridine (BrdU) ad libitum for 7 days prior to nephrectomy of the graft-bearing kidney. Euglycemic mice at the time of nephrectomy were followed for reversion to hyperglycemia for confirmation of human islet graft function. Islet graft-bearing kidneys were fixed in 10% neutral-buffered formalin. Paraffin-embedded sections were stained with guinea pig anti-insulin (Dako, Carpinteria, CA, USA), mouse anti-glucagon (Abcam, Cambridge, England), and rat anti-BrdU (Accurate Chemical, Westbury, NJ, USA); secondary Alexa Fluor antibodies (Alexa Fluor 647, 594, 488) were from Life Technologies (Carlsbad, CA, USA), and 4′,6-diamidino-2-phenylindole (DAPI) was from Sigma-Aldrich Co. Insulin+, insulin+BrdU+, glucagon+, and glucagon+BrdU+ cells were visualized by fluorescence microscopy (Nikon Eclipse Ti series; Nikon Corporation, Tokyo, Japan). Terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) assay was performed as per manufacturer’s instructions (Hoffman-La Roche Ltd). All counts were performed with Nikon NIS Elements software.
Total beta- and alpha-cell counts in human islet grafts
To determine total beta- and alpha-cell numbers in the islet grafts, 5 μm serial sections were cut through the entire graft and immunostained for insulin and glucagon. Beginning at the outer edge of the graft, images of the entire section were taken and stitched with a Nikon Eclipse Ti series microscope with motorized x–y stage. Subsequent serial sections were counted at 20 μm intervals to avoid duplicate counting of the same cells; all counts were performed with Nikon NIS Elements software.
Statistical analyses
Time-course data were analyzed by two-way analysis of variance (ANOVA) with Tukey’s or Holm–Sidak’s multiple comparisons test; insulin levels, cell counts, proliferation, and TUNEL data were analyzed by one-way ANOVA with Bonferroni’s or Tukey’s multiple comparisons test when comparing the four groups. Percent diabetes survival (Kaplan–Meier) was analyzed by Mantel–Cox log rank test. All statistical analyses were performed with GraphPad Prism (San Diego, CA, USA); P-values <0.05 were considered significant.
Results
Lixisenatide treatment accelerates restoration of normoglycemia in diabetic mice engrafted with human islets
Diabetic NSG RIP-DTR mice were engrafted with human islets from a single donor, randomized into four groups, and treated with vehicle (control) and lixisenatide at 50 μg/kg (low dose), 150 μg/kg (medium dose), and 500 μg/kg (high dose). A total of five independent studies were done with human islets from five donors; none of the donors were diagnosed as diabetic. The demographic characteristics of the islet donors and the numbers of islet equivalents (IEQs) transplanted into each mouse for each study are shown in Table 1.
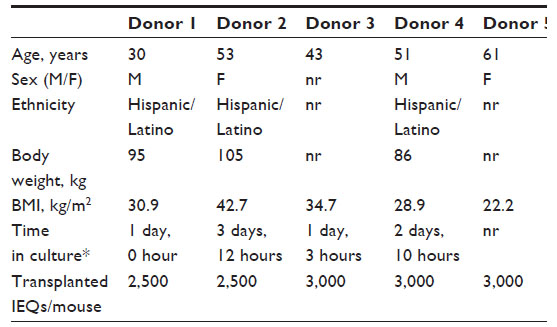 | Table 1 Demographic characteristics of human islet donors |
Blood glucose and body weight measurements of the mice were taken prior to islet transplant and then twice weekly following transplantation (Figure 1A and B). In four of the five transplant studies, mice treated with lixisenatide exhibited significant improvement in blood glucose control more rapidly than mice receiving vehicle alone (Figure 1A). In the Donor 2 study, only the high-dose lixisenatide mice showed significant improvement in blood glucose (at 9 and 13 days post-transplant). The mice in all other groups remained hyperglycemic; consequently, all mice engrafted with Donor 2 islets were euthanized without further analysis.
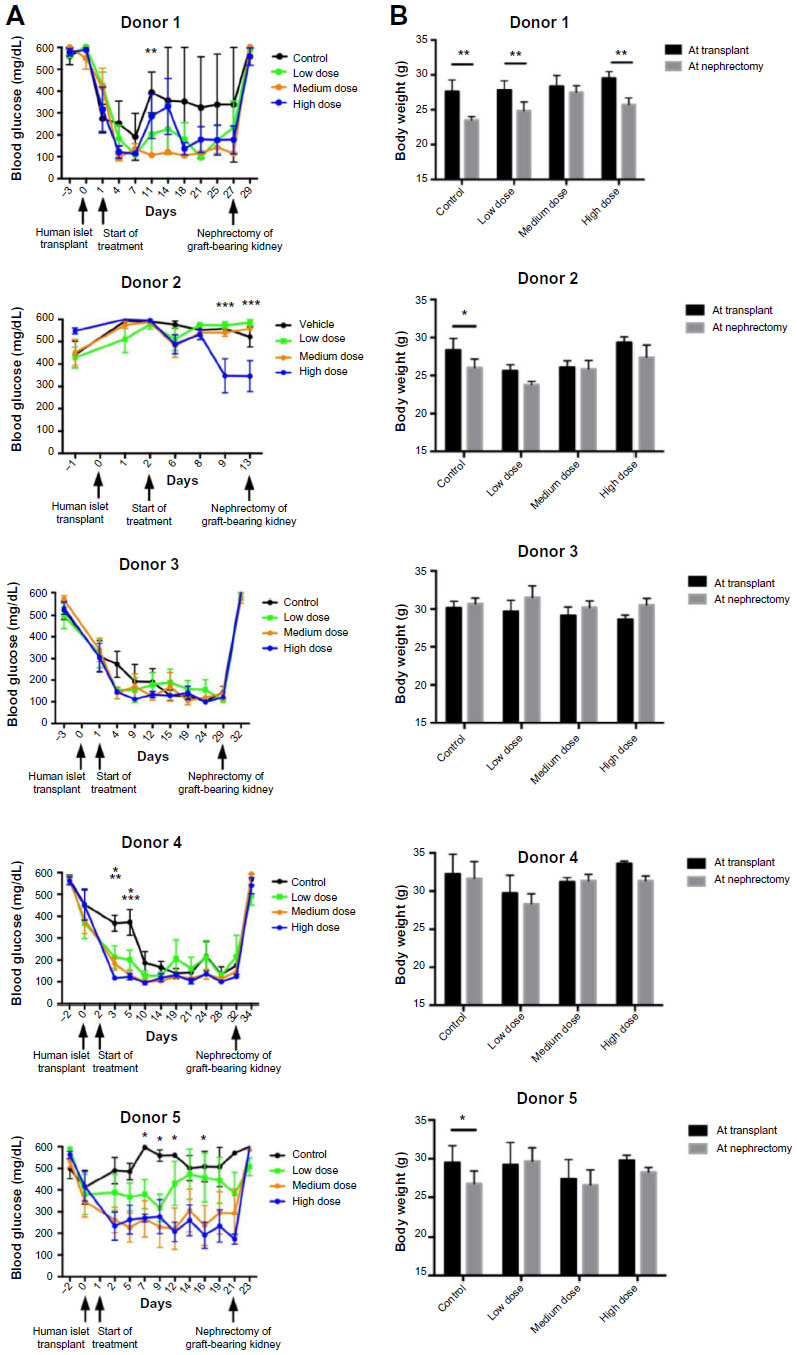 | Figure 1 Blood glucose and body weights of control and lixisenatide-treated mice engrafted with human islets. |
A combined survival curve analysis for transplant studies with islets from Donors 1, 3, 4, and 5 showed that lixisenatide treatment improved recovery from diabetes with high statistical power (Figure 2). The median time for the mice to become diabetes free was 12 days for the vehicle control group, and 5, 3, and 4 days for low-, medium-, and high-dose lixisenatide-treated groups, respectively. On surgical excision of the graft-bearing kidney, all mice became acutely hyperglycemic (blood glucose >500 mg/dL, Figure 1A), thus verifying that the human islet graft was responsible for maintaining blood glucose levels.
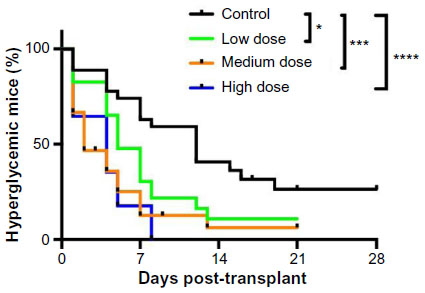 | Figure 2 Lixisenatide treatment improves recovery from hyperglycemia in human islet-engrafted mice. |
Lixisenatide treatment is associated with improved body weight maintenance
Mouse body weights were measured during each transplant study. In the Donor 1 study, all mice lost significant weight between the time of transplant and nephrectomy, except the medium-dose group (Figure 1B), which also showed the best blood glucose control. In transplant studies with islets from Donors 2 and 5, mice in the control group lost a small, but significant, amount of weight. This weight loss was not unexpected because these control mice were hyperglycemic throughout the trial. However, it is interesting to note that none of the lixisenatide-treated mice in the Donor 2 and 5 studies showed significant weight loss, even though some of the mice in the lower dose groups were also hyperglycemic. In transplant studies with islets from Donors 3 and 4, in which normoglycemia was eventually restored in all mice, there was no significant change in body weight of the mice in either the control or lixisenatide-treated groups.
Glucose-stimulated human insulin secretion is increased in islet-engrafted mice treated with lixisenatide
At 2 weeks post-transplant, the fasting levels of human insulin showed no significant differences between control and any of the lixisenatide-treatment groups (Figure 3A). However, glucose-stimulated levels of human insulin were significantly increased with medium-dose lixisenatide treatment in Donors 1 and 4 transplant studies (Figure 3B). Of note, the stimulation index (ratio of stimulated to fasting human insulin) was significantly increased compared to controls in the medium-dose group for Donor 1, and both medium- and high-dose groups for Donors 3 and 4 transplants (Figure 3C). Mice in the Donor 2 and 5 transplant studies were excluded because the controls (as well as some of the treated mice) were hyperglycemic.
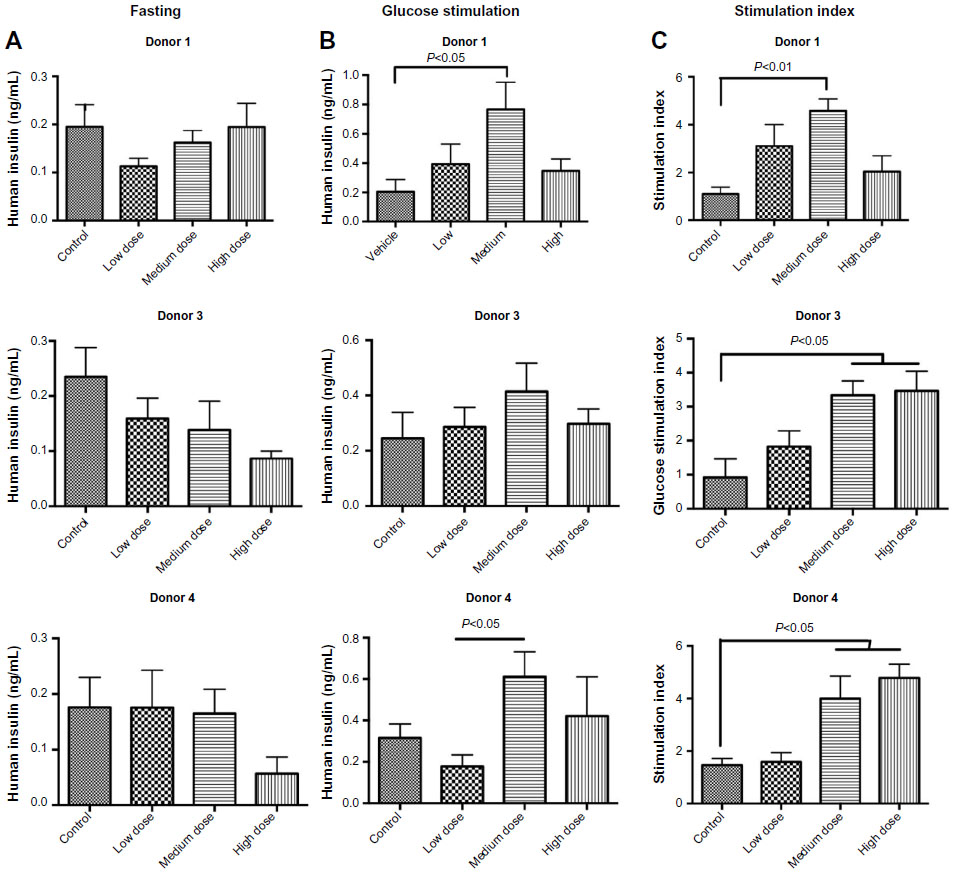 | Figure 3 Fasting and glucose-stimulated human insulin levels and stimulation indices in control and lixisenatide-treated mice engrafted with human islets. |
Lixisenatide treatment improves GTTs in human islet-engrafted mice
Consistent with the increased human insulin levels in lixisenatide-treated mice in response to glucose, lixisenatide treatment also significantly improved responses in GTTs in both the Donor 3 and 4 studies at 4 weeks post-transplant (Figure 4). Although mice in the Donor 4 study received uninterrupted drug treatment until the GTT, mice in the Donor 3 study continued to exhibit significantly improved glucose tolerance, even though lixisenatide treatment had been discontinued for 4 days prior to the GTT. In the Donor 3 transplant study, the mice treated with lixisenatide showed significant differences from controls in the medium-dose (P<0.01) and high-dose (P<0.05) groups, whereas in the Donor 4 transplant study, the low-dose lixisenatide group showed significant differences from the control group (P<0.05).
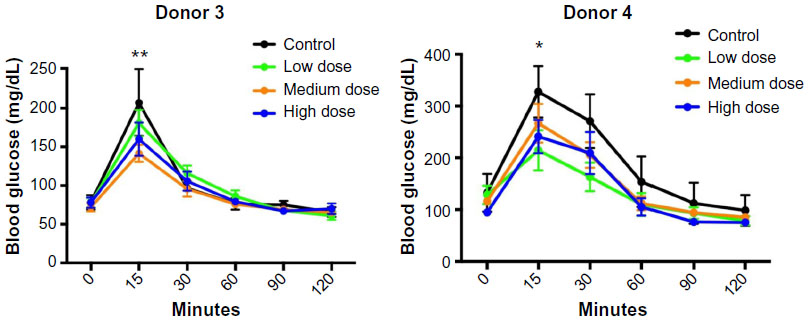 | Figure 4 GTTs in control and lixisenatide-treated mice engrafted with human islets. |
Apoptosis and proliferation analyses of human islet grafts recovered from control and lixisenatide-treated mice
Treatment with GLP-1 receptor agonists has been reported to protect rodent pancreatic beta cells from apoptosis.1 To determine whether lixisenatide treatment modulates human beta-cell apoptosis, the islet grafts were recovered on days 29, 32, and 21 (for Donor 3, 4, and 5 studies, respectively) and examined by TUNEL staining (Figure 5). We found no significant differences in beta-cell apoptosis between the control and lixisenatide-treated groups at the time points examined (at the end of each islet transplant study). In addition, although there was a variability between donor islets, the average percent beta-cell apoptosis observed at the end of each study was very low (<1%).
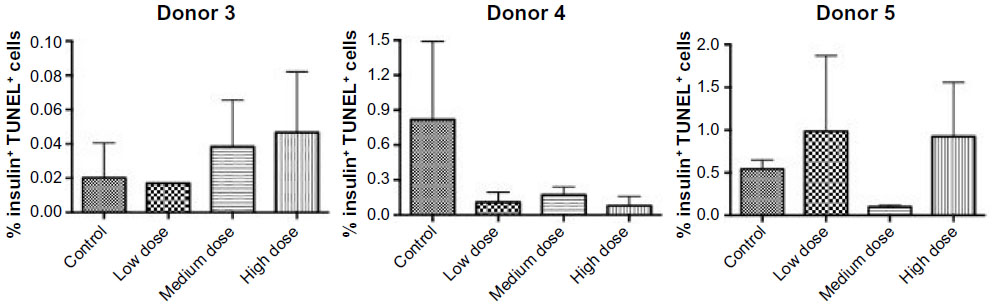 | Figure 5 Percent TUNEL-positive human beta cells in islet grafts recovered from control and lixisenatide-treated mice. |
To determine whether lixisenatide treatment induced human beta (or alpha)-cell proliferation, the mice were supplied with BrdU in their drinking water for 1 week prior to recovery of the graft-bearing islets. The percentages of BrdU+ beta cells (Figure 6A) and BrdU+ alpha cells (Figure 6B) for each human islet graft are shown for control and lixisenatide-treatment groups from Donor 3, 4, and 5 transplant studies combined. Consistent with previous reports,30–32 most islet grafts had very low levels of proliferating human beta cells, but neither beta- nor alpha-cell proliferation was significantly affected by lixisenatide treatment (as measured by BrdU incorporation during the last 7 days of each islet transplant study).
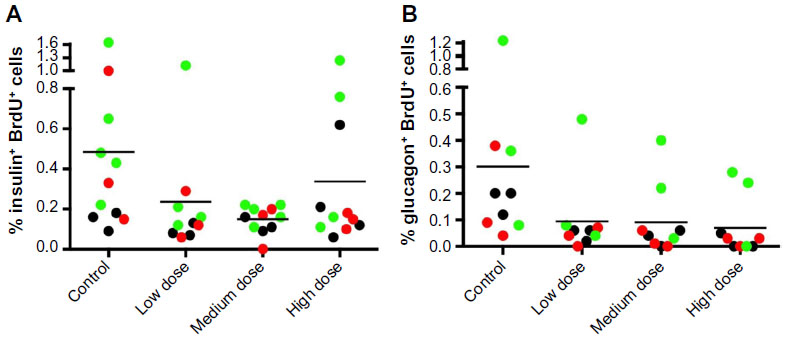 | Figure 6 Percent human beta- and alpha-cell proliferation in each islet graft recovered from control and lixisenatide-treated mice. |
Quantitation of total beta and alpha cells in human islet grafts recovered from control and lixisenatide-treated mice
Our proliferation and apoptosis analyses reflect lixisenatide effects on the human islet grafts only as a “snapshot” at late stages of engraftment. Therefore, to better interrogate the effect of lixisenatide on human beta-cell survival throughout the ~30-day treatment period, we counted total beta- and alpha-cell numbers within the recovered islet grafts. We observed considerable donor-to-donor variability with regard to the numbers of beta (Figure 7A) and alpha (Figure 7B) cells in the recovered islet grafts, even though the same numbers of IEQs were transplanted in each of the three studies examined.
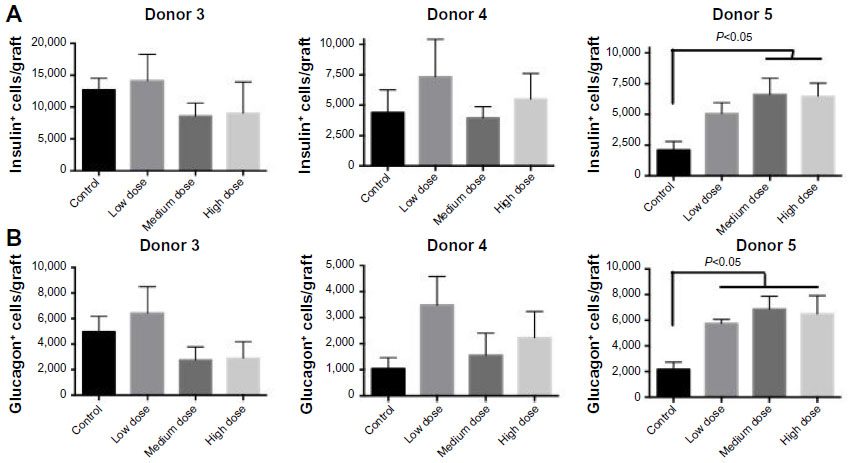 | Figure 7 Total beta- and alpha-cell counts in human islet grafts recovered from control and lixisenatide-treated mice. |
In the Donor 5 transplant study, medium- and high-dose lixisenatide-treated mice displayed significantly greater beta-cell numbers within the graft compared to control mice. In this study, an average of ~2,000 beta cells was detected in the islet grafts of control mice compared to an average of ~12,500 and ~4,500 beta cells in the control groups of Donor 3 and 4 studies, respectively. Interestingly, Donor 5 control mice remained hyperglycemic throughout the study, whereas normoglycemia was restored in control group mice in the Donor 3 and 4 studies. The beta (and alpha)-cell counts in the low-dose lixisenatide-treated group from the Donor 5 study were approximately twofold higher than the control group, even though the blood glucose of these mice remained elevated (~400 mg/dL, Figure 1A). The blood glucose values in the medium- and high-dose groups were significantly lower than the control group, and these mice had approximately threefold higher numbers of beta (and alpha) cells. In support of this, the islet grafts from three vehicle control mice appear noticeably smaller than from lixisenatide-treated mice (Figure 8), consistent with the significantly lower beta- and alpha-cell counts in the control group.
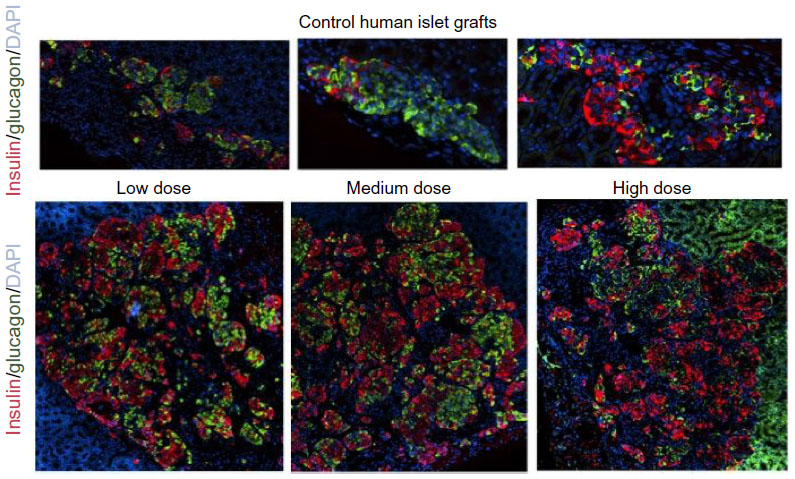 | Figure 8 Photomicrographs of human islet grafts from control and lixisenatide-treated mice. |
Discussion
In this study, we demonstrated that lixisenatide treatment significantly improved human beta-cell function and survival in diabetic NSG RIP-DTR mice engrafted with human islets. The human islets were derived from both male and female islet donors, aged 30–61 years, with body mass index (BMI) from 22.2 to 42.7 kg/m2; none was diagnosed as having diabetes. Five independent islet transplant studies were conducted, and mice were treated with three different doses of lixisenatide (50, 150, and 500 μg/kg) and vehicle control. A significantly accelerated recovery from diabetes was observed in lixisenatide-treated mice compared to controls, with the median time to a diabetes-free condition of 5, 3, and 4 days for low-, medium-, and high-dose lixisenatide groups, respectively, compared to controls at 12 days. The numbers of human islets transplanted were 2,500 IEQs in Donor 1 and 2 studies, and 3,000 IEQs in studies with Donors 3, 4, and 5. Although human islets from nondiabetic donors can comprise as few as 28% or as many as 75% beta cells,33 in our experience, ~3,000 human IEQs is typically sufficient to restore normoglycemia in the NSG RIP-DTR model.
Plasma levels of human insulin and C-peptide in fed or fasted islet-engrafted mice were variable or not significantly different between the control and lixisenatide-treatment groups. In contrast, mice treated with medium- and high-dose lixisenatide (compared to control) had significantly increased glucose-stimulated plasma levels of human insulin and glucose-stimulation indices for each of the three donors that were analyzed. This is consistent with human studies in T2D patients and nondiabetic subjects, in whom lixisenatide stimulated insulin secretion when blood glucose levels were increased, but not at normoglycemia.34 In support of this, GTTs were significantly improved in mice treated with lixisenatide compared to control mice engrafted with the same donor islets. Similarly, in a study with T2D patients insufficiently controlled on metformin, lixisenatide treatment provided a significantly greater reduction in postprandial plasma glucose than in placebo control.35
To measure the proliferation of human beta and alpha cells in the islet grafts, control and lixisenatide-treated mice were given BrdU in their drinking water 7 days before removal of the graft-bearing kidney. No significant differences were seen between lixisenatide treatment and control groups, although individual islet grafts derived from Donors 4 and 5 tended to have more BrdU+ cells in control mice compared to lixisenatide-treated animals engrafted with the same islets. Of note, control mice in the Donor 4 and 5 transplant studies had higher blood glucose levels than lixisenatide-treated mice, and we and others have reported that even mild hyperglycemia induces human beta-cell proliferation in human islet-engrafted mice.31,32 Nonetheless, beta cells from humans have a very low proliferation rate compared to rodents, and hyperglycemia induction results in only a ~0.5% proliferative rate in human beta cells.30–32
GLP-1 and its receptor agonists have been reported to inhibit beta-cell apoptosis in short-term culture of insulinoma cells and freshly isolated human and rodent islets.8,26,36 At recovery of the human islet grafts ~4 weeks post-transplant, we observed no significant difference between lixisenatide-treated and control groups in beta-cell apoptosis (as measured by TUNEL staining). However, it is possible that lixisenatide may have had an anti-apoptotic effect on the islet grafts at earlier stages in the post-transplant period. Indeed, it has been estimated that up to 70% of islet mass may be lost in the early post-transplant period, even in immunodeficient or syngeneic transplant models.37,38 Thus, it is likely that lixisenatide may have modulated beta-cell survival at earlier post-transplant time periods. In support of this, GLP-1 receptor agonist treatment of diabetic mice engrafted with syngeneic islets reversed the loss of both the number and mass of islets grafts at 1 and 3 days post-transplant.39 With longer GLP-1 receptor agonist treatment (2–3 weeks), both Ins2Akita (Akita) and Leprdb mice had increased islet mass and elevated pancreatic insulin content compared with controls.7,8 Similarly, we found that lixisenatide treatment dramatically improved total beta- and alpha-cell numbers in human islet grafts from Donor 5.
In the Donor 5 transplant study, the human islets failed to lower blood glucose levels in the control group, and mice in the low-dose lixisenatide group also remained hyperglycemic. Remarkably, the insulin+ (beta) and glucagon+ (alpha) cell counts in the islet grafts of mice treated with low-dose lixisenatide were twofold greater than in the control group, and mice treated with medium- and high-dose lixisenatide had threefold greater numbers of alpha and beta cells. Because the low rate of human beta-cell proliferation cannot account for these differences in beta-cell numbers, these data are consistent regarding the role of lixisenatide to preserve human beta-cell survival.
It is possible that lixisenatide’s effect to maintain beta-cell viability may be secondary to its ability to increase insulin secretion and lower blood glucose, thus preventing glucotoxicity. However, in Akita mice, the protective effect of GLP-1 receptor agonist on pancreatic islet mass was found to be independent of lowered blood glucose levels.7 Similarly, in our Donor 5 study, the low-dose lixisenatide group showed a twofold increase in beta-cell number compared to the control group, even though mice in this group remained hyperglycemic throughout the study. These data suggest that lixisenatide may have additional islet-protective effects on human beta cells, such as that previously reported in rodent diabetes models, in which GLP-1 receptor agonists acted to reduce beta-cell endoplasmic reticulum stress.3,7–9 Taken together, our data are consistent with a role for lixisenatide to preserve human beta-cell function and survival in vivo, in particular when numbers of functional beta cells are limiting, as would be found in individuals with T2D.
Acknowledgment
Sanofi-Aventis funded this research.
Author contributions
CY, ML, AJ, LDS, DLG, DMH, and RB contributed to the conception and design of the study and interpretation of the data. CY, AJ, NP, and LL performed experiments and analyzed the data. All authors contributed toward data analysis, drafting and critically revising the paper and agree to be accountable for all aspects of the work. None of this work has been published or submitted elsewhere.
Disclosure
Dr Loehn is an employee of Sanofi-Aventis. The authors report no other conflicts of interest in this work.
References
Drucker DJ. Glucagon-like peptides: regulators of cell proliferation, differentiation, and apoptosis. Mol Endocrinol. 2003;17(2):161–171. | |
Garber AJ. Incretin effects on beta-cell function, replication, and mass: the human perspective. Diabetes Care. 2011;34(Suppl 2):S258–S263. | |
Kwon DY, Kim YS, Ahn IS, et al. Exendin-4 potentiates insulinotropic action partly via increasing beta-cell proliferation and neogenesis and decreasing apoptosis in association with the attenuation of endoplasmic reticulum stress in islets of diabetic rats. J Pharmacol Sci. 2009;111(4):361–371. | |
Brubaker PL, Drucker DJ. Minireview: glucagon-like peptides regulate cell proliferation and apoptosis in the pancreas, gut, and central nervous system. Endocrinology. 2004;145(6):2653–2659. | |
Egan JM, Bulotta A, Hui H, Perfetti R. GLP-1 receptor agonists are growth and differentiation factors for pancreatic islet beta cells. Diabetes Metab Res Rev. 2003;19(2):115–123. | |
Perfetti R, Merkel P. Glucagon-like peptide-1: a major regulator of pancreatic beta-cell function. Eur J Endocrinol. 2000;143(6):717–725. | |
Yamane S, Hamamoto Y, Harashima S, et al. GLP-1 receptor agonist attenuates endoplasmic reticulum stress-mediated beta-cell damage in Akita mice. J Diabetes Investig. 2011;2(2):104–110. | |
Yusta B, Baggio LL, Estall JL, et al. GLP-1 receptor activation improves beta cell function and survival following induction of endoplasmic reticulum stress. Cell Metab. 2006;4(5):391–406. | |
Tsunekawa S, Yamamoto N, Tsukamoto K, et al. Protection of pancreatic beta-cells by exendin-4 may involve the reduction of endoplasmic reticulum stress; in vivo and in vitro studies. J Endocrinol. 2007;193(1):65–74. | |
Fehse F, Trautmann M, Holst JJ, et al. Exenatide augments first- and second-phase insulin secretion in response to intravenous glucose in subjects with type 2 diabetes. J Clin Endocrinol Metab. 2005;90(11):5991–5997. | |
Vilsboll T, Zdravkovic M, Le-Thi T, et al. Liraglutide, a long-acting human glucagon-like peptide-1 analog, given as monotherapy significantly improves glycemic control and lowers body weight without risk of hypoglycemia in patients with type 2 diabetes. Diabetes Care. 2007; 30(6):1608–1610. | |
Nauck MA, Heimesaat MM, Behle K, et al. Effects of glucagon-like peptide 1 on counterregulatory hormone responses, cognitive functions, and insulin secretion during hyperinsulinemic, stepped hypoglycemic clamp experiments in healthy volunteers. J Clin Endocrinol Metab. 2002;87(3):1239–1246. | |
Rother KI, Spain LM, Wesley RA, et al. Effects of exenatide alone and in combination with daclizumab on beta-cell function in long-standing type 1 diabetes. Diabetes Care. 2009;32(12):2251–2257. | |
Sarkar G, Alattar M, Brown RJ, et al. Exenatide treatment for 6 months improves insulin sensitivity in adults with type 1 diabetes. Diabetes Care. 2014;37(3):666–670. | |
Elkinson S, Keating GM. Lixisenatide: first global approval. Drugs. 2013;73(4):383–391. | |
Petersen AB, Knop FK, Christensen M. Lixisenatide for the treatment of type 2 diabetes. Drugs Today (Barc). 2013;49(9):537–553. | |
Christensen M, Knop FK, Holst JJ, Vilsboll T. Lixisenatide, a novel GLP-1 receptor agonist for the treatment of type 2 diabetes mellitus. IDrugs. 2009;12(8):503–513. | |
Werner U, Haschke G, Herling AW, Kramer W. Pharmacological profile of lixisenatide: a new GLP-1 receptor agonist for the treatment of type 2 diabetes. Regul Pept. 2010;164(2–3):58–64. | |
Fonseca VA, Alvarado-Ruiz R, Raccah D, Boka G, Miossec P, Gerich JE. Efficacy and safety of the once-daily GLP-1 receptor agonist lixisenatide in monotherapy: a randomized, double-blind, placebo-controlled trial in patients with type 2 diabetes (GetGoal-Mono). Diabetes Care. 2012;35(6):1225–1231. | |
Horowitz M, Rayner CK, Jones KL. Mechanisms and clinical efficacy of lixisenatide for the management of type 2 diabetes. Adv Ther. 2013; 30(2):81–101. | |
Petersen AB, Christensen M. Clinical potential of lixisenatide once daily treatment for type 2 diabetes mellitus. Diabetes Metab Syndr Obes. 2013;6:217–231. | |
Ahren B, Gautier JF, Berria R, Stager W, Aronson R, Bailey CJ. Pronounced reduction of postprandial glucagon by lixisenatide: a meta-analysis of randomized clinical trials. Diabetes Obes Metab. 2014; 16(9):861–868. | |
Meier JJ. GLP-1 receptor agonists for individualized treatment of type 2 diabetes mellitus. Nat Rev Endocrinol. 2012;8(12):728–742. | |
Cervera A, Wajcberg E, Sriwijitkamol A, et al. Mechanism of action of exenatide to reduce postprandial hyperglycemia in type 2 diabetes. Am J Physiol Endocrinol Metab. 2008;294(5):E846–E852. | |
Linnebjerg H, Park S, Kothare PA, et al. Effect of exenatide on gastric emptying and relationship to postprandial glycemia in type 2 diabetes. Regul Pept. 2008;151(1–3):123–129. | |
Tews D, Werner U, Eckel J. Enhanced protection against cytokine- and fatty acid-induced apoptosis in pancreatic beta cells by combined treatment with glucagon-like peptide-1 receptor agonists and insulin analogues. Horm Metab Res. 2008;40(3):172–180. | |
Shultz LD, Ishikawa F, Greiner DL. Humanized mice in translational biomedical research. Nat Rev Immunol. 2007;7(2):118–130. | |
Shultz LD, Lyons BL, Burzenski LM, et al. Human lymphoid and myeloid cell development in NOD/LtSz-scid IL2R gamma null mice engrafted with mobilized human hemopoietic stem cells. J Immunol. 2005;174(10):6477–6489. | |
Jurczyk A, Diiorio P, Brostowin D, et al. Improved function and proliferation of adult human beta cells engrafted in diabetic immunodeficient NOD-scid IL2rgamma(null) mice treated with alogliptin. Diabetes Metab Syndr Obes. 2013;6:493–499. | |
Perl S, Kushner JA, Buchholz BA, et al. Significant human beta-cell turnover is limited to the first three decades of life as determined by in vivo thymidine analog incorporation and radiocarbon dating. J Clin Endocrinol Metab. 2010;95(10):E234–E239. | |
Levitt HE, Cyphert TJ, Pascoe JL, et al. Glucose stimulates human beta cell replication in vivo in islets transplanted into NOD-severe combined immunodeficiency (SCID) mice. Diabetologia. 2011;54(3):572–582. | |
Diiorio P, Jurczyk A, Yang C, et al. Hyperglycemia-induced proliferation of adult human beta cells engrafted into spontaneously diabetic immunodeficient NOD-Rag1null IL2rgammanull Ins2Akita mice. Pancreas. 2011;40(7):1147–1149. | |
Brissova M, Fowler MJ, Nicholson WE, et al. Assessment of human pancreatic islet architecture and composition by laser scanning confocal microscopy. J Histochem Cytochem. 2005;53(9):1087–1097. | |
Becker RH, Stechl J, Msihid J, Kapitza C. Lixisenatide resensitizes the insulin-secretory response to intravenous glucose challenge in people with type 2 diabetes – a study in both people with type 2 diabetes and healthy subjects. Diabetes Obes Metab. 2014;16(9):793–800. | |
Ratner RE, Rosenstock J, Boka G, Investigators DRIS. Dose-dependent effects of the once-daily GLP-1 receptor agonist lixisenatide in patients with type 2 diabetes inadequately controlled with metformin: a randomized, double-blind, placebo-controlled trial. Diabet Med. 2010; 27(9):1024–1032. | |
Farilla L, Bulotta A, Hirshberg B, et al. Glucagon-like peptide 1 inhibits cell apoptosis and improves glucose responsiveness of freshly isolated human islets. Endocrinology. 2003;144(12):5149–5158. | |
Biarnes M, Montolio M, Nacher V, Raurell M, Soler J, Montanya E. Beta-cell death and mass in syngeneically transplanted islets exposed to short- and long-term hyperglycemia. Diabetes. 2002;51(1):66–72. | |
Davalli AM, Ogawa Y, Ricordi C, Scharp DW, Bonner-Weir S, Weir GC. A selective decrease in the beta cell mass of human islets transplanted into diabetic nude mice. Transplantation. 1995;59(6):817–820. | |
Toyoda K, Okitsu T, Yamane S, et al. GLP-1 receptor signaling protects pancreatic beta cells in intraportal islet transplant by inhibiting apoptosis. Biochem Biophys Res Commun. 2008;367(4):793–798. |
Supplementary material
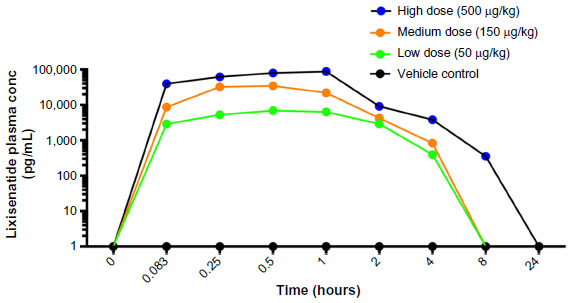 | Figure S1 Pharmacokinetic analysis of control and lixisenatide treatments. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.