Back to Journals » Cancer Management and Research » Volume 11
Integrative gene expression profiling reveals that dysregulated triple microRNAs confer paclitaxel resistance in non-small cell lung cancer via co-targeting MAPT
Authors Cai Y, Jia R, Xiong H, Ren Q, Zuo W, Lin T, Lin R, Lei Y, Wang P, Dong H, Zhao H, Zhu L, Fu Y, Zeng Z, Zhang W, Wang S ![]()
Received 12 May 2019
Accepted for publication 22 July 2019
Published 5 August 2019 Volume 2019:11 Pages 7391—7404
DOI https://doi.org/10.2147/CMAR.S215427
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
Yuanming Cai, 1,* Ruxue Jia, 2, 3,* Haozhe Xiong, 4 Qun Ren, 2, 3 Weimin Zuo, 2, 3 Tingting Lin, 2, 3 Rong Lin, 2 Yan Lei, 2 Ping Wang, 2 Huiyue Dong, 2 Hu Zhao, 2, 3 Ling Zhu, 3 Yunfeng Fu, 3 Zhiyong Zeng, 5 Wei Zhang, 6 Shuiliang Wang 2, 3
1Undergraduate Grade 2015, The 5th Clinical Medical School of Xinjiang Medical University, Urumchi, People’s Republic of China; 2Department of Urology, The 900th Hospital of the Joint Logistics Team, Fujian Medical University, Fuzhou, Fujian, People’s Republic of China; 3Fujian Key Laboratory of Transplant Biology, Affiliated Dongfang Hospital, Xiamen University School of Medicine, Fuzhou, Fujian, People’s Republic of China; 4Undergraduate Grade 2017, Department of Bioengineering, College of Life Science, Fujian Normal University, Fuzhou, Fujian, People’s Republic of China; 5Department of Thoracic Surgery, The 900th Hospital of the Joint Logistics Team, Fujian Medical University, Fuzhou, Fujian, People’s Republic of China; 6Department of Emergency, The 900th Hospital of the Joint Logistics Team, Fujian Medical University, Fuzhou, Fujian, People’s Republic of China
*These authors contributed equally to this work
Background: Paclitaxel has shown significant anti-tumor activity against non-small cell lung cancer (NSCLC); however, resistance to paclitaxel frequently occurs and represents a significant clinical problem and its underlying molecular mechanism remains elusive.
Methods: Long-term treatment of culture cell with paclitaxel was carried out to mimic the development of acquired drug resistance in NSCLC. Cell proliferation and clonogenic assay and apoptosis evaluation were carried out to determine the efficacy of paclitaxel on NSCLC cells. Western blot analyses were performed to determine the expression and activation of proteins. Apoptosis enzyme-linked immunosorbent assay was used to quantify cytoplasmic histone-associated DNA fragments. Microarray analyses were applied to explore both mRNA and miRNA expression profiles in NSCLC cells followed by integrative analysis. qRT-PCR was carried out to verify the differentially expressed mRNAs and miRNAs.
Results: The expression of 652 genes was shown to be changed at least 2-fold in paclitaxel-resistant NSCLC (H460_TaxR) cells with 511 upregulated and 141 downregulated as compared with that in parental H460 cells. The differentially expressed genes were functionally enriched in regulating the cell proliferation, cell death, and response to endogenous stimulus, and clustered in pathways such as cancer and signaling by the G protein-coupled receptor (GPCR). Moreover, 43 miRNAs were shown to be differentially expressed in H460_TaxR cells with 15 upregulated and 28 downregulated as compared with parental H460 cells. A total of 289 pairs of miRNA-potential target gene were revealed in H460_TaxR cells by bioinformatics analysis. Furthermore, integrative analysis of miRNAs and gene expression profiles revealed that dysregulated miR-362-3p, miR-766-3p, and miR-6507-3p might confer paclitaxel resistance in NSCLC via targeting MAPT simultaneously.
Conclusion: Our findings suggested that specific manipulation of MAPT-targeting miRNAs may be a novel strategy to overcome paclitaxel resistance in patients with NSCLC especially large-cell lung carcinoma.
Keywords: integrative analysis, non-small cell lung cancer, paclitaxel-resistance, gene expression profile, miRNAs
Corrigendum for this paper has been published
Introduction
Lung cancer remains the most commonly diagnosed cancer and the leading cause of cancer-related deaths, with 2.1 million new cases and 1.8 million deaths in 2018 worldwide.1,2 Of all pathological types, non-small cell lung cancer (NSCLC) accounts for approximate 85% of all lung cancers and more than 65% of the patients with NSCLC present with locally advanced or metastatic disease.3,4
The major treatment options for patients with NSCLC are determined on the basis of histologic features and staging according to the eighth edition of the TNM (T, N, and M represent tumor, lymph node, and metastasis, respectively) classification for lung cancer.5 Generally, the current treatment for lung cancer patients who have been diagnosed at an early stage is surgical resection followed by chemotherapy; however, majority of the patients will eventually experience disease progression and require further treatment.6,7 Paclitaxel, either as single agent or combined with other therapeutics, has shown significant anti-NSCLC activity.8–10 In advanced stages of NSCLC, the efficacy of paclitaxel in combination with a platinum compound has also been confirmed by a meta-analysis of 16 randomized trials.11 However, either intrinsic or acquired resistance to paclitaxel frequently occurs and represents a significant clinical problem.12 We had previously shown that microRNA-mediated epigenetic targeting of survivin significantly enhances the antitumor activity of paclitaxel against NSCLC.13 In the current report, we investigated both the gene and miRNAs expression profiles in paclitaxel-resistant large-cell lung carcinoma cells, a pathological type of relative small population of NSCLC. An integrative analysis of miRNAs and gene expression profiles was also carried out.
Materials and methods
Reagents and antibodies
Paclitaxel was obtained from the 900th Hospital of the Joint Logistics Team pharmacy (the Former Fuzhou General Hospital). The CellTiter96 AQ cell proliferation kit (Cat.#G3582) was a product of Promega (Madison, WI, USA). Oligonucleotides were synthesized in Sangon (Shanghai, China). Antibodies against PARP (Cat.#9542) and Caspase-3 (Cat.#9665) were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA); antibody against β-actin (Cat.#A5441) was the product of Sigma (St. Louis, MO, USA).
Cells and cell culture
Human large-cell lung carcinoma cell line H460 was obtained from American Type Culture Collection (Manassas, VA, USA) and maintained in RPMI1640 medium supplemented with 10% fetal bovine serum (FBS). All cells were cultured in a 37ºC humidified atmosphere containing 95% air and 5% CO2 and were split twice a week.
To mimic the development of acquired paclitaxel resistance in vitro, parental H460 cells were maintained in culture medium with low dose of paclitaxel started from 1 nmol/L initially. Cells were then split twice a week and maintained in culture medium with gradually increased dose of paclitaxel every 2 weeks. After 6 months, paclitaxel-resistance NSCLC cells developed from H460 were maintained in culture medium with 40 nmol/L of paclitaxel for normal culture.
Cell viability assay
The CellTiter96 AQ cell proliferation kit (Cat.#G3582, Promega) was used to determine cell viability as previously described.13 Briefly, cells were plated onto 96-well plates for 24 hrs, and then grown in either RPMI1640 medium with 0.5% FBS as control or the same medium containing different concentrations of paclitaxel, and then incubated for another 72 hrs. During this period, the medium was refreshed daily maintaining the same treatment. After reading all wells at 490 nm with a microplate reader, the percentages of surviving cells from each group relative to controls, defined as 100% survival, were determined by reduction of 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt (MTS).
Clonogenic assay
The parental and paclitaxel-resistant human SCLC cancer cells H460 were plated onto 6-well plates and incubated at 37°C with 5% CO2. After 24 hrs, the culture medium was replaced daily with 2 mL fresh medium containing 0.5% FBS or the same medium containing indicated concentrations of paclitaxel for long-term incubation. After 2 weeks, cells were stained with 0.5% crystal violet (dissolved in 25% methanol) and clone number was quantified with QuantiOne software of Fluor-STM Multimager (Bio-Rad Laboratories, Inc., Hercules, CA) at the end of the experiments.
Western blotting analysis and quantification of apoptosis
Protein expression and activation were determined by Western blotting analysis as previously described.14 In brief, equal amounts of cell lysates in a buffer (containing 50 mmol/L Tris (pH 7.4), 50 mmol/L NaCl, 0.5% NP40, 50 mmol/L NaF, 1 mmol/L Na3VO4, 1 mmol/L phenylmethylsulfonyl fluoride, 25 μg/mL leupeptin, and 25 μg/mL aprotinin) were boiled in sodium dodecyl sulfate (SDS) sample buffer (0.0625 mol/L Tris (pH 6.8), 2% SDS, 10% Glycerol, 5% 2-mercaptoethanol, 0.002% Bromophenol-B), resolved by SDS-polyacrylamide gel electrophoresis and Western blotted with specific antibodies directed against PARP (1:1000), Caspase-3 (1:1000), or β-actin (1:10000), as described in the figure legends. For quantification of apoptosis, an apoptosis enzyme-linked immunosorbent assay kit (Cat.#11774425001, Roche Diagnostics Corp., Indianapolis, IN, USA) was used to quantitatively measure cytoplasmic histone-associated DNA fragments (mononucleosomes and oligonucleosomes) as previously reported.14
Microarray analysis of both mRNA and miRNA followed by integrative analysis
Total RNAs were prepared from parental and paclitaxel-resistant H460 cells (H460_Parental and H460_TaxR, respectively) with miRNeasy Mini Kit (QIAGEN, GmBH, Germany). For mRNA and miRNA profiling, Agilent G3 Human (8*60 K) Chip and Agilent Human miRNA Chip (8*60 K) V19.0 (Agilent technologies, Santa Clara, CA, USA) were used, respectively. Microarray analysis was performed in triplicate in Shanghai Biochip Co., Ltd (Shanghai, China). Either mRNA or miRNA with two-fold change and a P<0.05 between parental and paclitaxel-resistant H460 cells was taken as significantly differentially expressed. The mRNA and miRNA expression profiles were then subjected to integrative analysis in Shanghai Center for Bioinformation Technology (Shanghai, China).
Analysis of differentially expressed mRNAs and miRNAs with real-time quantitative reverse transcriptase PCR (qRT-PCR)
Total RNA was prepared using TRIZOL reagent (Invitrogen, Carlsbad, CA, USA). First-strand cDNA was generated using High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Carlsbad, CA, USA) following the manufacturer’s instructions. To quantify the mRNA level of MAPT (forward primer: 5ʹ-AAGATCGGCTCCACTGAGAA-3; reverse primer: 5ʹ-ATGAGCCACACTTGGAGGTC-3ʹ), qRT-PCR was performed using the FastStart Universal SYBR Green Master Mixes (Roche) by a 7900HT Fast Real-Time PCR system (Applied Biosystems) as we had described previously.15 The expression of β-actin was used as an internal control (forward primer: 5ʹ-AGAGCTACGAGCTGCCTGAC-3; reverse primer: 5ʹ-AGCACTGTGTTGGCGTACAG-3ʹ). The expression of mature miR-362-3p, miR-766-3p, miR-6507-3p, and RNU6B was measured by qRT-PCR using TaqMan Assays (Applied Biosystems) as described previously.15 The relative mRNA and miRNA levels were calculated using the comparative Ct method (2−ΔΔCt).
Statistical analysis
All results were confirmed by at least three independent experiments. Data are presented as mean±SD. Student’s t-test or one-way ANOVA was carried out to analyze the differences between two groups or multiple groups. Values of P<0.05 were considered significant.
Results
Establishment of paclitaxel-resistant NSCLC cells
To establish a stable NSCLC cell subline with significant resistance to paclitaxel, parental H460 cells were subjected to growth in culture medium with gradually increased dose of paclitaxel persistently to mimic the development of acquired drug resistance in vitro. Six months later, NSCLC cell subline with significant resistance to paclitaxel was developed from parental H460 without obvious morphology change (data not shown). The subline was nominated as H460_TaxR. As shown in Figure 1A, the IC50 values of paclitaxel were 4.824 nmol/L and 41.209 nmol/L for parental and paclitaxel-resistant H460 cells, respectively. The IC50 value for paclitaxel-resistant H460 cells increased 8.542-fold as compared with that of parental H460 cells. The significant resistance to anti-proliferative/anti-survival effects of paclitaxel in H460_TaxR cells was further confirmed by clonogenic assay (Figure 1B and C). Moreover, results from both Western blotting analysis and quantification of apoptosis showed that while treatment with as low as a dose of 1.5 nmol/L of paclitaxel induced significant apoptosis in parental H460 cells; however, for H460_TaxR cells, even treatment with 12 nmol/L of paclitaxel could only induce a few apoptosis (Figure 1D and E). Collectively, our results demonstrated that an NSCLC cell subline H460_TaxR with significant resistance to paclitaxel was established successfully.
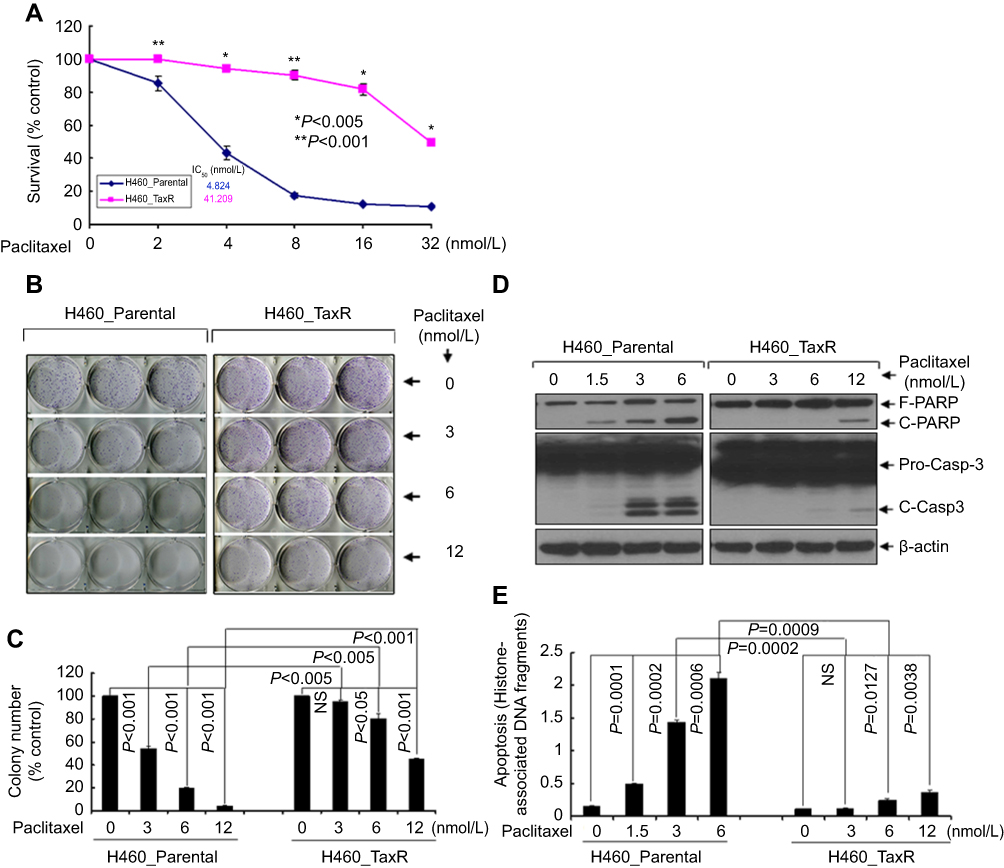 |
Figure 1 Identification of paclitaxel-resistant NSCLC cells. Notes: (A) Human NSCLC cells (H460_Parental and H460_TaxR) treated with indicated concentrations of paclitaxel for 72 h were subjected to cell viability assay. (B, C) H460_Parental and H460_TaxR cells were grown in triplicates in the absence or presence of indicated concentrations of paclitaxel for 2–3 weeks. The pictures and numbers of the cell colonies were obtained by the QuantiOne software of Fluor-STM Multimager. (D, E) H460_Parental and H460_TaxR cells were treated with indicated concentrations of paclitaxel for 24 hrs. Cells were collected and subjected to Western blot analyses of PARP, Casp-3 or β-actin (D), or apoptotic-ELISA (E). Abbreviations: F-PARP, full length of poly(ADP-ribose) polymerase; C-PARP, cleaved PARP; Pro-Casp-3, Caspase-3; C-Casp-3, cleaved caspase-3; ELISA, enzyme-linked immunosorbent assay. |
Gene expression profile in paclitaxel-resistant NSCLC cells
Next, to gain a more comprehensive knowledge of the underlying molecular mechanism of paclitaxel-resistance in NSCLC, microarray analysis was carried out to investigate the gene expression profile in H460_TaxR cells with significant resistance to paclitaxel. The expression of 652 genes was found to be changed at least 2-fold in H460_TaxR cells with 511 upregulated and 141 downregulated as compared with parental H460 cells (Figure 2, P<0.05). Of these genes, the top 10 genes that were most significantly up- or downregulated in H460_TaxR cells are shown in Table 1. Gene Ontology (GO, http://www.geneontology.org) function enrichment analysis based on our own microarray data showed that the differentially expressed genes in H460_TaxR cells mainly involved in regulating the cell proliferation, cell death, and response to endogenous stimulus. (Table 2). Meanwhile, Gene Set Enrichment Analysis (GSEA) revealed that those differentially expressed genes were clustered in pathways such as cancer,16 signaling by GPCR, cytokine and cytokine receptor interaction, and so on (Table 3).
 |
Figure 2 mRNA expression profiles in parental and paclitaxel-resistant NSCLC cells. Note: The heatmap from unsupervised hierarchical clustering showed mRNAs with high expression in red and mRNAs with low expression in green. |
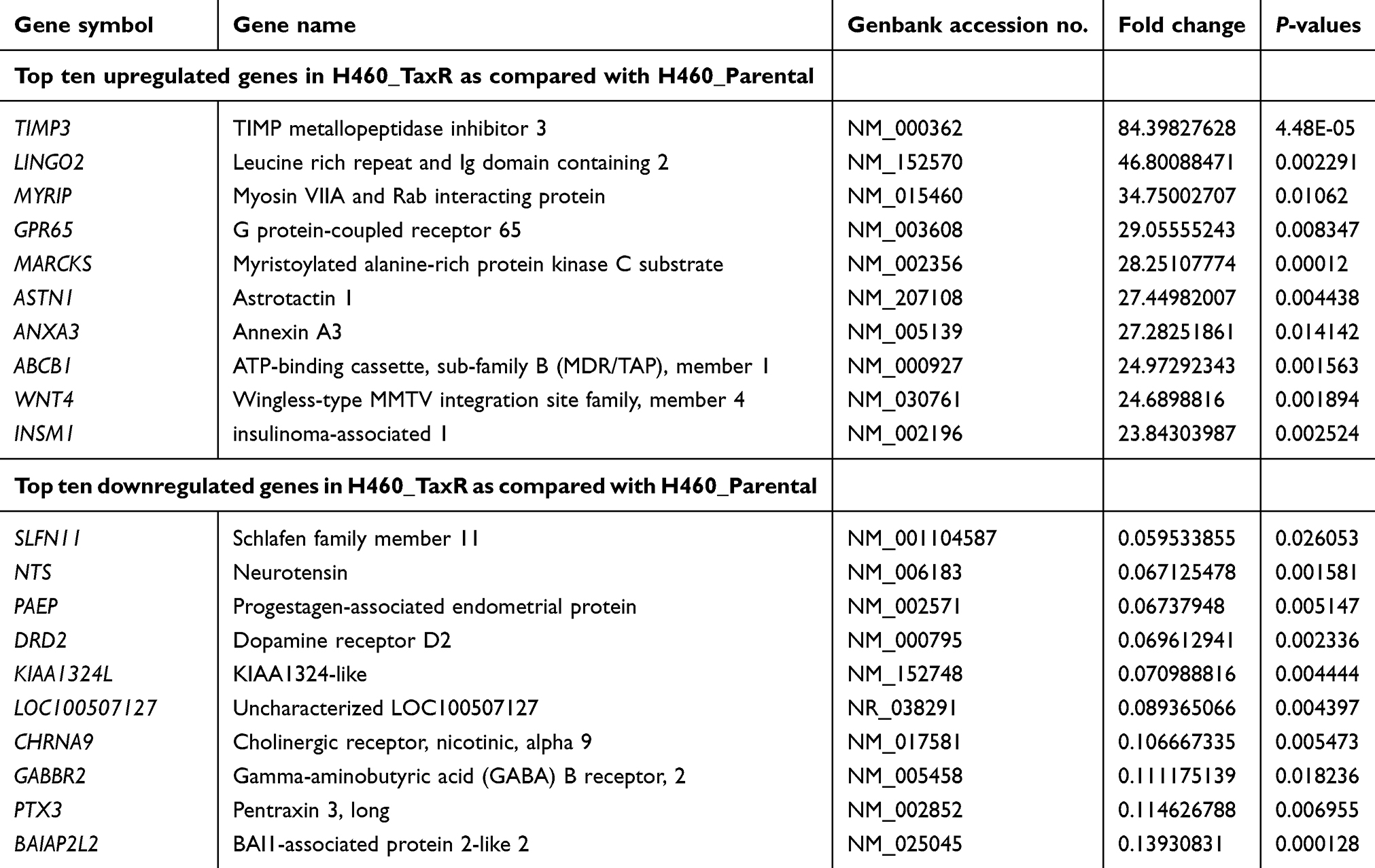 |
Table 1 Representative differentially expressed genes in paclitaxel-resistant NSCLC cells |
 |
Table 2 Function enrichment analysis of differentially expressed genes in paclitaxel-resistant NSCLC cells |
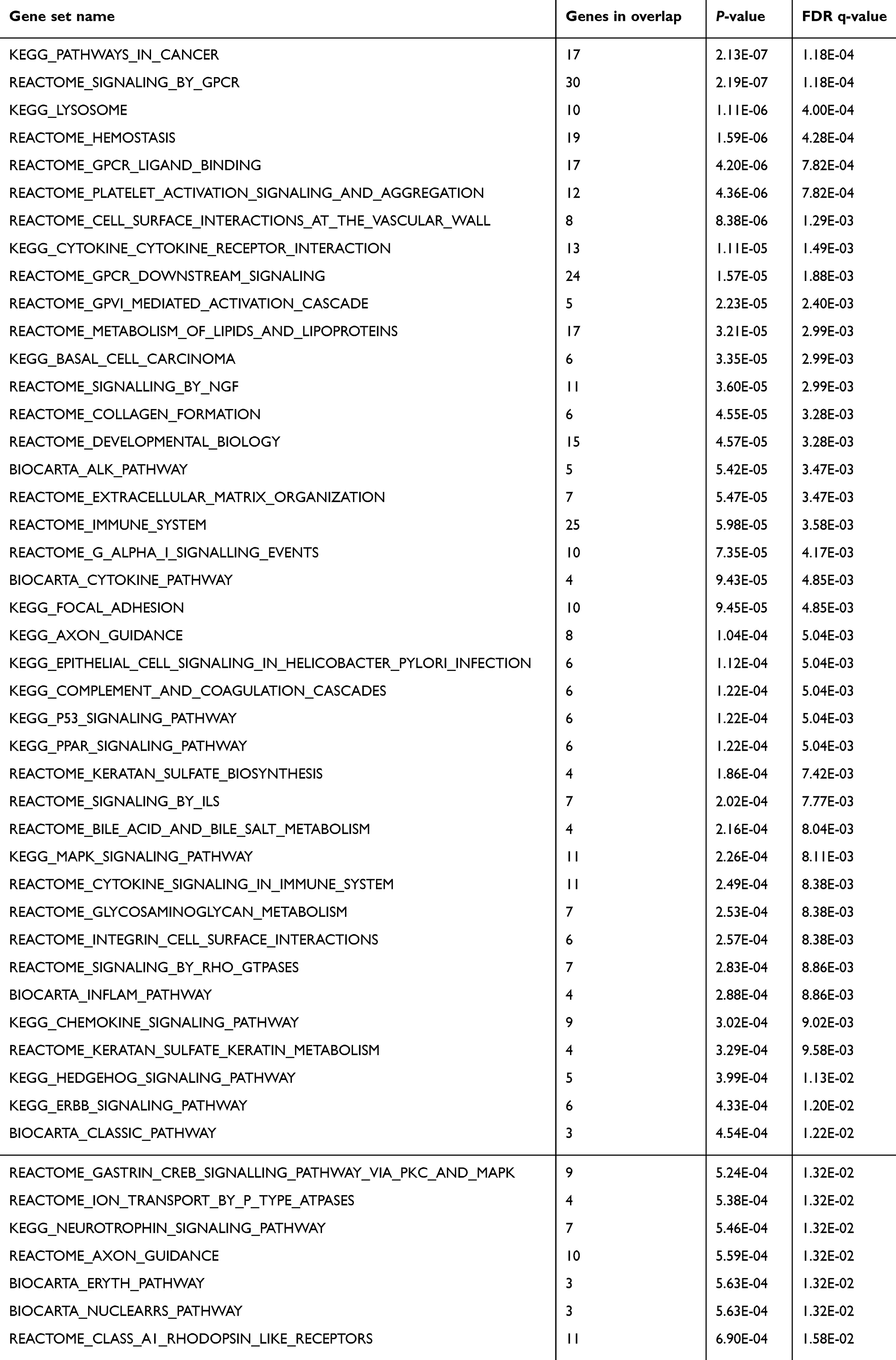 |
Table 3 Clustered pathways analysis of differentially expressed genes in paclitaxel-resistant NSCLC cells |
MiRNAs expression profile in paclitaxel-resistant NSCLC cells
MiRNA has emerged as an important regulator in chemoresistance.17 Thus, microarray analysis was also carried out to investigate the miRNAs expression profile in H460_TaxR cells with significant resistance to paclitaxel in our study. As shown in Figure 3 and Table 4, the expression of 43 miRNAs was found to be changed at least 2-fold in H460_TaxR cells with 15 upregulated and 28 downregulated as compared with parental H460 cells (P<0.05). A total of 289 pairs of miRNA-potential target gene were revealed in paclitaxel-resistant NSCLC cells by bioinformatics analysis.
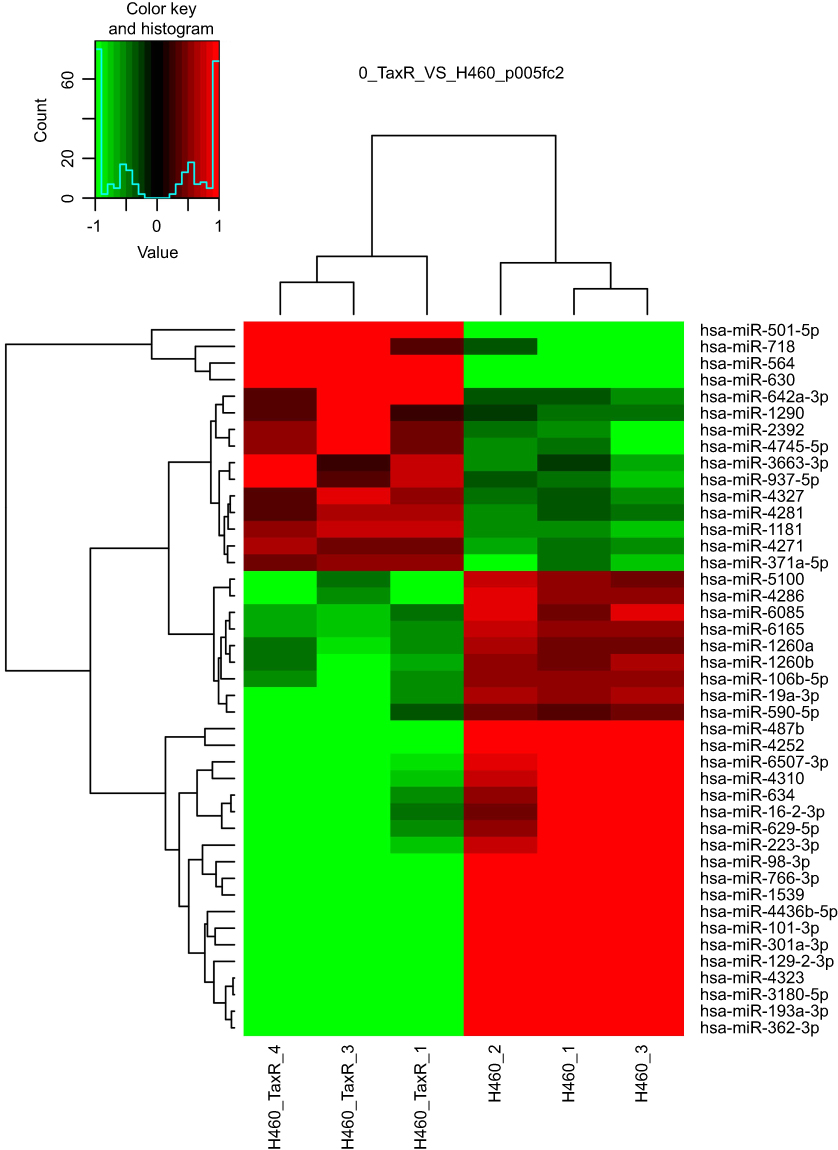 |
Figure 3 miRNA expression profiles in parental and paclitaxel-resistant NSCLC cells. Note: The heatmap from unsupervised hierarchical clustering showed miRNAs with high expression in red and miRNAs with low expression in green. |
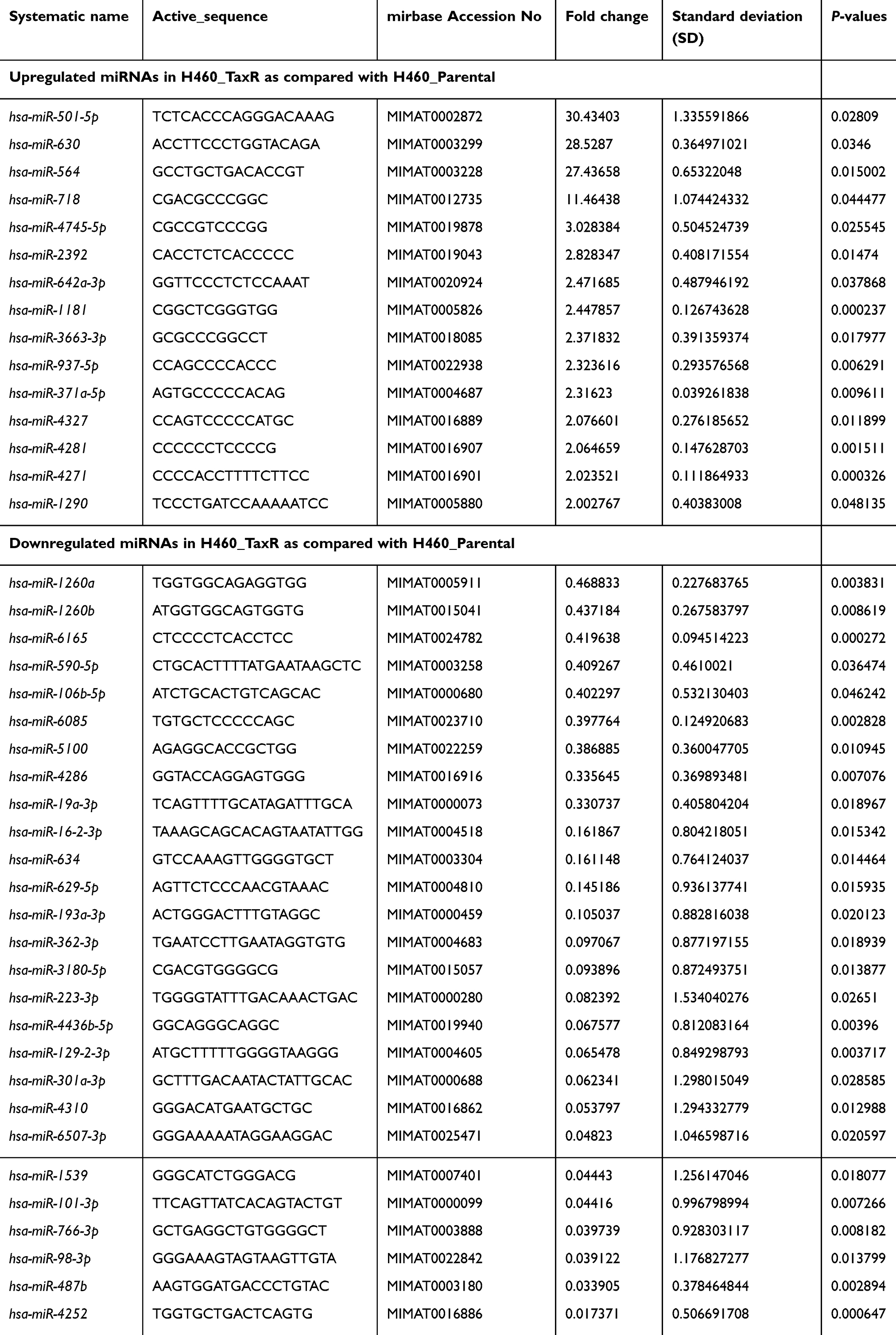 |
Table 4 Differentially expressed miRNAs in paclitaxel-resistant NSCLC cells |
Integrative analysis of miRNAs and gene expression profiles revealed that dysregulated MAPT-targeting microRNAs confer paclitaxel resistance in NSCLC
Cumulative studies had indicated that microtubule-associated protein tau (MAPT) play an important role in mediating the sensitivity of paclitaxel in several types of tumor including breast cancer.18,19 Since our current gene expression profiling also showed that the MAPT mRNA was significantly upregulated in H460_TaxR cells as compared with parental H460 (Fold-change=6.31, P=3.46E-03; Figure 4A), we then focused our integrative analysis on candidate miRNAs targeting MAPT mRNA. Of all 28 downregulated miRNAs in H460_TaxR cells, 3 of them including miR-362-3p, miR-766-3p, and miR-6507-3p whose expression was verified by qRT-PCR were predicted to target MAPT mRNA via searching in miRDB (http://www.mirdb.org) with at least one binding site in its 3ʹ-UTR (Figure 4B and C). Meanwhile, miR-362-3p and miR-766-3p were also predicted to target MAPT mRNA by TargetScan (http://www.targetscan.org). Furthermore, a MAPT-subnetwork was constructed via searching in STRING database (http://string-db.org) in combination with analysis of potential targets of miR-362-3p, miR-766-3p, and miR-6507-3p. As shown in Figure 4D, 13 of 14 potential targets of miR-362-3p, miR-766-3p, and miR-6507-3p were upregulated in H460_TaxR cells (Table 5) and could form a MAPT-subnetwork with protein–protein interactions. Taken together, our integrative analysis of miRNAs and gene expression profiles revealed that dysregulated a couple of microRNAs might confer paclitaxel resistance in NSCLC especially large-cell lung carcinoma via targeting MAPT simultaneously.
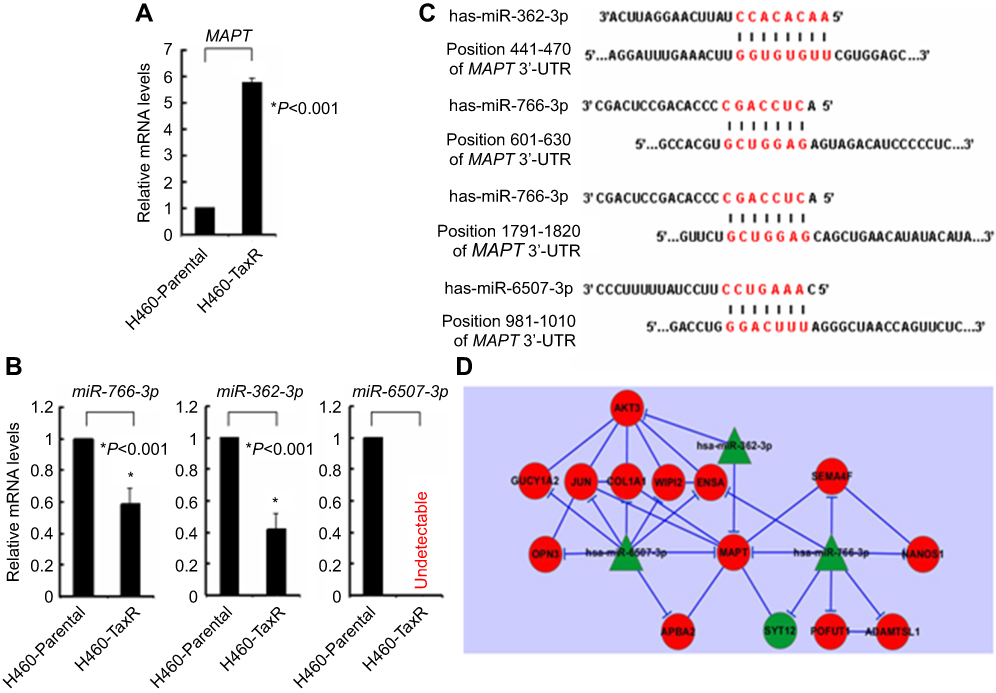 |
Figure 4 MAPT-subnetwork in paclitaxel-resistant NSCLC cells. Notes: (A) qRT-PCR analysis of mRNA level of MAPT in H460_Parental and H460_TaxR. (B) qRT-PCR analysis of miRNAs levels of miR-766-3p, miR-362-3p, and miR-6507-3p in H460_Parental and H460_TaxR. (C) Diagram of targeting sequences of miR-766-3p, miR-362-3p, and miR-6507-3p in 3ʹ-UTR of MAPT mRNA. (D) Diagram of constructed MAPT-subnetwork in paclitaxel-resistant non-small cell lung cancer cells. Green, downregulated; Red, upregulated. Abbreviations: MAPT, microtubule-associated protein tau; qRT-PCR, quantitative reverse transcriptase PCR; 3ʹ-UTR, 3ʹ-untranslated region. |
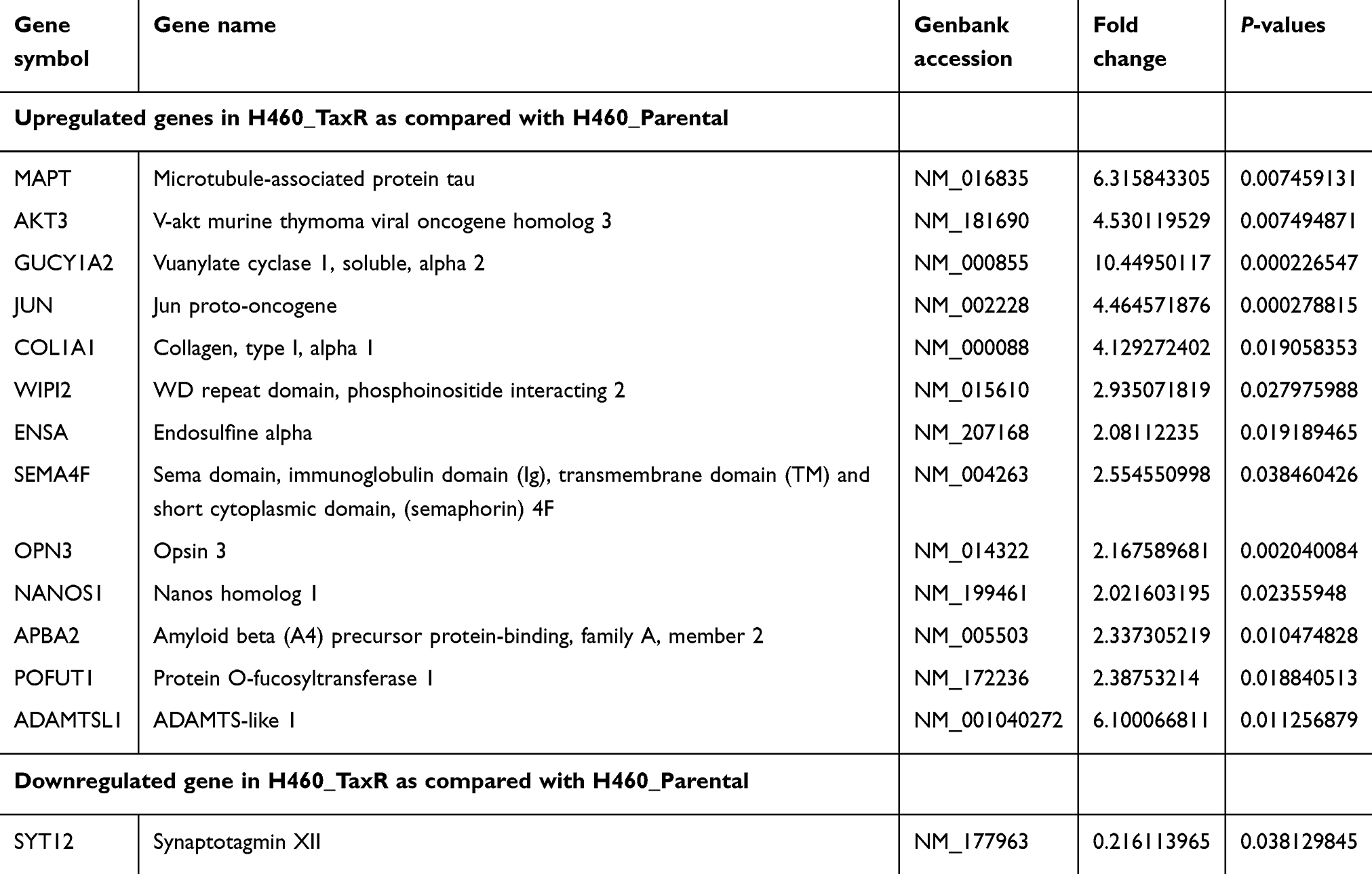 |
Table 5 Differentially expressed genes in MAPT-subnetwork in paclitaxel-resistant NSCLC cells |
Discussion
Paclitaxel, initially derived from the bark of the Pacific yew tree, has long been used to treat NSCLC either as single-agent or combined with other therapeutics.8,9,19 Treatment with paclitaxel disables spindle division and causes cell cycle arrest in phase G1/G2 of mitosis as well as induces apoptosis.20 However, resistance to paclitaxel remains one of the main causes of treatment failure in NSCLC.12 It is of particular significance to elucidate the underlying mechanism of paclitaxel resistance and identify new strategy to abrogate it.
Mechanisms of the resistance to paclitaxel are complex as indicated by cumulative studies, which include alterations in microtubule dynamics, altered expression of β-tubulin isotypes, and also deregulated signaling pathways.21–24 In our own serial studies, we have demonstrated that activation of PI-3K/Akt signaling plays a critical role in ErbB2/ErbB3-mediated therapeutic resistance to paclitaxel mainly via upregulation of survivin.14,25 In addition, we have also found that microRNA-mediated epigenetic targeting of survivin significantly enhances the antitumor activity of paclitaxel against NSCLC.12 Unexpectedly, as we did not observe the change of survivin in our established paclitaxel-resistant NSCLC cells (data not shown), an integrative gene expression profiling was applied to explore the possible novel underlying molecular mechanism of paclitaxel-resistance in NSCLC in our current study. We showed that a total of 43 miRNAs and 652 protein-encoding genes, functionally enriched in regulating the fundamental biological processes including cell proliferation and clustered in pathways such as cancer, are differentially expressed in paclitaxel-resistant NSCLC cells, which suggest that the mechanism of resistance to paclitaxel is more complicated than we had ever recognized.
Upon binding to a pocket in beta-tubulin, paclitaxel inhibits the microtubule depolymerization process and interferes the progression of normal cell cycle.19 MAPT, being a microtubule-associated protein which promotes the assembly of tubulin into microtubules to stabilize microtubule structure,26 has been shown to compete with paclitaxel via sharing the same site.27 As anticipated, a large body of studies had indicated that MAPT plays an important role in mediating the sensitivity of paclitaxel in several types of tumor.18,19,28–30 Consistently, we also found that the mRNA expression of MAPT was significantly upregulated in paclitaxel-resistant NSCLC cells. However, the underlying mechanism of deregulated expression of MAPT remains largely unknown currently. A study by Wu and colleagues showed that miR-34c-5p determines the chemosensitivity of gastric cancer to paclitaxel via regulating MAPT.31 Recently, miR-186 was also showed to regulate paclitaxel sensitivity of NSCLC via targeting MAPT.32 Interestingly, although both aforementioned two miRNAs remain unchanged in our established paclitaxel-resistant NSCLC cells, our integrative analysis of miRNAs and gene expression profiles revealed that deregulated miR-362-3p, miR-766-3p, and miR-6507-3p and their 13 potential targets could form a MAPT-subnetwork with protein–protein interactions and confer paclitaxel resistance in NSCLC (Figure 4D). Currently, since the molecular mechanism underlying deregulated miR-362-3p, miR-766-3p, and miR-6507-3p remains largely unclear, it would be of great interest to unveil it. Besides, whether there exists a causal relationship between specific silence of triple microRNAs (miR-362-3p, miR-766-3p, and miR-6507-3p) and upregulation of MAPT awaits further investigation. Taken together, our findings shed new light on the mechanism of paclitaxel resistance in NSCLC especially large-cell lung carcinoma and suggested that specific manipulation of MAPT-targeting miRNAs may be a novel strategy to overcome paclitaxel resistance in patients with NSCLC in the future.
Abbreviations
NSCLC, non-small cell lung cancer; MAPT, microtubule-associated protein tau; qRT-PCR, quantitative reverse transcriptase PCR; 3ʹ-UTR, 3ʹ-untranslated region; PARP, poly (ADP-ribose) polymerase; C-PARP, cleaved PARP; C-Casp-3, cleaved Caspase-3; ELISA, enzyme-linked immunosorbent assay.
Data sharing statement
Supplementary data is available on request.
Acknowledgments
This work was funded in part by the National Natural Science Foundation of China (No. 81772848), Joint Funds for the Innovation of Science and Technology from Fujian Province (No. 2017Y9127), the Natural Science Foundation of Fujian Province of China (Nos. 2017J01223, 2018J01349), and the Research Project of Military Logistics (No. CNJ13J011).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Ferlay J, Colombet M, Soerjomataram I, et al. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int J Cancer. 2019;144(8):1941–1953. doi:10.1002/ijc.31937
3. Devesa SS, Bray F, Vizcaino AP, Parkin DM. International lung cancer trends by histologic type: male: female differences diminishing and adenocarcinoma rates rising. Int J Cancer. 2005;117(2):294–299. doi:10.1002/(ISSN)1097-0215
4. Morgensztern D, Ng SH, Gao F, Govindan R. Trends in stage distribution for patients with non-small cell lung cancer: a national cancer database survey. J Thorac Oncol. 2010;5(1):29–33. doi:10.1097/JTO.0b013e3181c5920c
5. Reck M, Heigener DF, Mok T, Soria JC, Rabe KF. Management of non-small-cell lung cancer: recent developments. Lancet. 2013;382(9893):709–719. doi:10.1016/S0140-6736(13)61502-0
6. Reck M, Rabe KF. Precision diagnosis and treatment for advanced non-small-cell lung cancer. N Engl J Med. 2017;377(9):849–861. doi:10.1056/NEJMra1703413
7. Lu T, Yang X, Huang Y, et al. Trends in the incidence, treatment, and survival of patients with lung cancer in the last four decades. Cancer Manag Res. 2019;11:943–953. doi:10.2147/CMAR.S187317
8. Socinski MA. Single-agent paclitaxel in the treatment of advanced non-small cell lung cancer. Oncologist. 1999;4(5):408–416.
9. Belani CP. Paclitaxel and docetaxel combinations in non-small cell lung cancer. Chest. 2000;117(4Suppl 1):144S–141S. doi:10.1378/chest.117.4_suppl_1.144s
10. Sandler A, Gray R, Perry MC, et al. Paclitaxel-carboplatin alone or with bevacizumab for non-small-cell lung cancer. N Engl J Med. 2006;355(24):2542–2550. doi:10.1056/NEJMoa061884
11. NSCLC Meta-Analyses Collaborative Group. Chemotherapy in addition to supportive care improves survival in advanced non-small-cell lung cancer: a systematic review and meta-analysis of individual patient data from 16 randomized controlled trials. J Clin Oncol. 2008;26(28):4617–4625. doi:10.1200/JCO.2008.17.7162.
12. Orr GA, Verdier-Pinard P, McDaid H, Horwitz SB. Mechanisms of taxol resistance related to microtubules. Oncogene. 2003;22(47):7280–7295. doi:10.1038/sj.onc.1206934
13. Wang S, Zhu L, Zuo W, et al. MicroRNA-mediated epigenetic targeting of Survivin significantly enhances the antitumor activity of paclitaxel against non-small cell lung cancer. Oncotarget. 2016;7(25):37693–37713. doi:10.18632/oncotarget.9264
14. Wang S, Huang X, Lee CK, Liu B. Elevated expression of erbB3 confers paclitaxel resistance in erbB2-overexpressing breast cancer cells via upregulation of Survivin. Oncogene. 2010;29(29):4225–4236. doi:10.1038/onc.2010.180
15. Wang S, Huang J, Lyu H, et al. Functional cooperation of miR-125a, miR-125b, and miR-205 in entinostat-induced downregulation of erbB2/erbB3 and apoptosis in breast cancer cells. Cell Death Dis. 2013;4:e556. doi:10.1038/cddis.2013.79
16. Subramanian A, Tamayo P, Mootha VK, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci USA. 2005;102(43):15545–15550. doi:10.1073/pnas.0506580102
17. Garofalo M, Croce CM. MicroRNAs as therapeutic targets in chemoresistance. Drug Resist Updat. 2013;16(3–5):47–59. doi:10.1016/j.drup.2013.05.001
18. Xie S, Ogden A, Aneja R, Zhou J. Microtubule-binding proteins as promising biomarkers of paclitaxel sensitivity in cancer chemotherapy. Med Res Rev. 2016;36(2):300–312. doi:10.1002/med.21378
19. Shi X, Sun X. Regulation of paclitaxel activity by microtubule-associated proteins in cancer chemotherapy. Cancer Chemother Pharmacol. 2017;80(5):909–917. doi:10.1007/s00280-017-3398-2
20. Amos LA, Löwe J. How Taxol stabilises microtubule structure. Chem Biol. 1999;6(3):65–69. doi:10.1016/S1074-5521(99)89002-4
21. Gonçalves A, Braguer D, Kamath K, et al. Resistance to Taxol in lung cancer cells associated with increased microtubule dynamics. Proc Natl Acad Sci USA. 2001;98(20):11737–11742. doi:10.1073/pnas.191388598
22. Kavallaris M, Kuo DY, Burkhart CA, et al. Taxol-resistant epithelial ovarian tumors are associated with altered expression of specific beta-tubulin isotypes. J Clin Invest. 1997;100(5):1282–1293. doi:10.1172/JCI119642
23. Atjanasuppat K, Lirdprapamongkol K, Jantaree P, Svasti J. Non-adherent culture induces paclitaxel resistance in H460 lung cancer cells via ERK-mediated up-regulation of βIVa-tubulin. Biochem Biophys Res Commun. 2015;466(3):493–498. doi:10.1016/j.bbrc.2015.09.057
24. Yu D, Jing T, Liu B, et al. Overexpression of ErbB2 blocks Taxol-induced apoptosis by upregulation of p21Cip1, which inhibits p34Cdc2 kinase. Mol Cell. 1998;2(5):581–591.
25. Chen J, Ren Q, Cai Y, et al. Mesenchymal stem cells drive paclitaxel resistance in ErbB2/ErbB3-coexpressing breast cancer cells via paracrine of neuregulin 1. Biochem Biophys Res Commun. 2018;501(1):212–219. doi:10.1016/j.bbrc.2018.04.218
26. Drechsel DN, Hyman AA, Cobb MH, Kirschner MW. Modulation of the dynamic instability of tubulin assembly by the microtubule-associated protein tau. Mol Biol Cell. 1992;3(10):1141–1154. doi:10.1091/mbc.3.10.1141
27. Kar S, Fan J, Smith MJ, Goedert M, Amos LA. Repeat motifs of tau bind to the insides of microtubules in the absence of taxol. EMBO J. 2003;22(1):70–77. doi:10.1093/emboj/cdg001
28. Mimori K, Sadanaga N, Yoshikawa Y, et al. Reduced tau expression in gastric cancer can identify candidates for successful paclitaxel treatment. Br J Cancer. 2006;94(12):1894–1897. doi:10.1038/sj.bjc.6603182
29. Smoter M, Bodnar L, Duchnowska R, Stec R, Grala B, Szczylik C. The role of Tau protein in resistance to paclitaxel. Cancer Chemother Pharmacol. 2001;68(3):553–557. doi:10.1007/s00280-011-1696-7
30. Wagner P, Wang B, Clark E, Lee H, Rouzier R, Pusztai L. Microtubule Associated Protein (MAP)-Tau: a novel mediator of paclitaxel sensitivity in vitro and in vivo. Cell Cycle. 2005;4(9):1149–1152. doi:10.4161/cc.4.9.2038
31. Wu H, Huang M, Lu M, et al. Regulation of microtubule- associated protein tau (MAPT) by miR-34c-5p determines the chemosensitivity of gastric cancer to paclitaxel. Cancer Chemother Pharmacol. 2013;71(5):1159–1171. doi:10.1007/s00280-013-2108-y
32. Ye J, Zhang Z, Sun L, Fang Y, Xu X, Zhou G. miR-186 regulates chemo-sensitivity to paclitaxel via targeting MAPT in non-small cell lung cancer (NSCLC). Mol Biosyst. 2016;12(11):3417–3424. doi:10.1039/c6mb00576d
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.