Back to Journals » Drug Design, Development and Therapy » Volume 14
Integrated Network Pharmacology Analysis and Experimental Validation to Reveal the Mechanism of Anti-Insulin Resistance Effects of Moringa oleifera Seeds
Authors Huang Q, Liu R, Liu J, Huang Q ![]() , Liu S
, Liu S ![]() , Jiang Y
, Jiang Y
Received 10 June 2020
Accepted for publication 27 August 2020
Published 2 October 2020 Volume 2020:14 Pages 4069—4084
DOI https://doi.org/10.2147/DDDT.S265198
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Jianbo Sun
Qiong Huang,1– 3,* Rong Liu,1– 3,* Jing Liu,1– 3 Qi Huang,1– 3 Shao Liu,1– 3 Yueping Jiang1– 3
1Department of Pharmacy, Xiangya Hospital, Central South University, Changsha 410008, People’s Republic of China; 2Institute of Hospital Pharmacy, Central South University, Changsha 410008, People’s Republic of China; 3Institute for Rational and Safe Medication Practices, National Clinical Research Center for Geriatric Disorders, Xiangya Hospital, Central South University, Changsha 410008, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shao Liu; Yueping Jiang
Department of Pharmacy, Xiangya Hospital, Central South University, 87 Xiangya Road, Changsha, Hunan 4100080, People’s Republic of China
Email [email protected]; [email protected]
Background and Purpose: Insulin resistance (IR) is one of the factors that results in metabolic syndrome, type 2 diabetes mellitus and different aspects of cardiovascular diseases. Moringa oleifera seeds (MOS), traditionally used as an antidiabetic food and traditional medicine in tropical Asia and Africa, have exhibited potential effects in improving IR. To systematically explore the pharmacological mechanism of the anti-IR effects of MOS, we adopted a network pharmacology approach at the molecular level.
Methods: By incorporating compound screening and target prediction, a feasible compound-target-pathway network pharmacology model was established to systematically predict the potential active components and mechanisms of the anti-IR effects of MOS. Biological methods were then used to verify the results of the network pharmacology analysis.
Results: Our comprehensive systematic approach successfully identified 32 bioactive compounds in MOS and 44 potential targets of these compounds related to IR, as well as 37 potential pathways related to IR. Moreover, the network pharmacology analysis revealed that glycosidic isothiocyanates and glycosidic benzylamines were the major active components that improved IR by acting on key targets, such as SRC, PTPN1, and CASP3, which were involved in inflammatory responses and insulin-related pathways. Further biological research demonstrated that the anti-IR effects of MOS were mediated by increasing glucose uptake and modulating the expression of SRC and PTPN1.
Conclusion: Our study successfully predicts the active ingredients and potential targets of MOS for improving IR and helps to illustrate mechanism of action at a systemic level. This study not only provides new insights into the chemical basis and pharmacology of MOS but also demonstrates a feasible method for discovering potential drugs from traditional medicines.
Keywords: network pharmacology, Moringa oleifera seeds, IR, active components, action mechanism
Introduction
Insulin resistance (IR) is caused by the resistance of cells to the effects of insulin. Due to this phenomenon, the biological effects of insulin on target tissues, such as liver, muscle, and adipose tissue, are reduced or lost. As a result, glucose uptake is reduced and glucose output from the liver is increased, leading to a variety of pathological and clinical manifestations.1 Prolonged IR is known to cause health disorders, including obesity, type 2 diabetes mellitus, non-alcoholic fatty liver, and cardiovascular diseases.2–5
The mechanism of IR is very complicated and involves various pathways and targets, including increased phosphorylation of insulin receptor substrate (IRS) protein via serine/threonine kinases, such as mitogen-activated protein kinase (MAPK) 8, I-kappa-B-kinase beta (IKKB), and protein kinase C; increased IRS-1 proteasome degradation via the mTOR signaling pathway; decreased activation of signaling molecules including phosphoinositide 3-kinase (PI3K) and serine/threonine-protein kinase (AKT); and increased activity of phosphatases including tyrosine-protein phosphatases (PTPs), phosphatidylinositol 3,4,5-trisphosphate 3-phosphatase and dual-specificity protein phosphatase (PTEN), and protein phosphatase 2A (PP2A).6–10 IR is the main pathophysiological symptom of type 2 diabetes mellitus. IR is mainly ascribed to dysfunction of glucose transporter 4 (GLUT4), which transports glucose from the cytoplasm to membrane. GLUT4 dysfunction is related to insulin signaling pathway abnormalities in adipose and skeletal muscle cells.11 Obesity can affect type 2 diabetes mellitus due to deposition of excess free fatty acids and triglycerides in the liver and muscles. In the liver, excessive oxidation of free fatty acids eventually results in the secretion of large amounts of reactive oxygen species (ROS), which activate c-Jun N-terminal kinase (JNK). After activation, JNK phosphorylates threonine and serine residues in IRS-1 and inhibits tyrosine phosphorylation of insulin-stimulated IRS-1, thereby preventing the coupling of IRS-1 to the regulatory subunit p85 of PI3K.12 In obesity, a large number of inflammatory cytokines, such as resistin and leptin, are also secreted by adipose tissues and activate the production of cytokine signaling inhibitors (suppressor of cytokine signaling, SOCSs) in hepatocytes and muscle cells. Excessive SOCSs bind to insulin receptor-IRS-1/2, thus blocking the insulin-induced tyrosine phosphorylation of IRS-1/2, and ultimately inhibiting PI3K/Akt activation, leading to IR.13,14
Moringa oleifera (MO) is a perennial angiosperm plant belonging to the Moringa genus and the Moringaceae family. MO is also known as the horseradish tree or the drumstick tree in tropical Asia and Africa, and is commonly used for medicinal and nutritional purposes.15 MO is used as a traditional medicine in tropical Africa, America, and Asia for therapeutic applications in several disorders including arterial hypertension and diabetes.16,17 The leaves, fruits, flowers, and immature pods of this tree are used as a highly nutritious vegetable in tropical regions of Africa, America, and Asia.18,19
The seeds, leaves, and fruits of MO contain significant amounts of important phytochemicals such as polysaccharides, phenolics, flavonoids, isothiocyanates, benzylamines, vitamins, sterols, and amino acids.18,20–22 To date, phenolics, flavonoids, sterols, isothiocyanates, and benzylamines are the most commonly reported components of M. oleifera seeds (MOS).23–27 Among these components, glucosinolates, isothiocyanate, and thiocarbamates are characteristic and exert multiple potent bioactivities.21,23–28
MOS extracts have been shown to possess pharmacological properties and commercial utility, such as metal antidote, anti-oxidant, anti-asthmatic, anti-arthritic, anti-bacterial, and antitumor, nematicidal, hypoglycemic and hepatoprotective activities as well as alleviating scopolamine-induced learning and memory impairment in mice.16,21,29–31 The extracts of M. oleifera leaf and M. oleifera bark have been shown to alleviate IR.32–37 However, the potential of MOS to improve IR remains to be fully elucidated.
In contrast to the classical principle of “one drug corresponds to one target, one target corresponds to one disease”, compound medicines or traditional medicines, which usually consist of multiple medicines or complicated active components act on multiple targets in a synergistic manner. Clarification of the mechanisms of action of multi-component and multi-targeted medicines is a challenge. With the rapid development of bioinformatics, network pharmacology has become a novel approach to effective and systematic investigations of the mechanism of action, and discovery potential bioactive ingredients in traditional medicines.38 Network pharmacology is a promising bioinformatics method used to investigate the mechanism underlying the interactions between drugs and their protein targets, and is especially applicable to natural botanical medicines with multiple component. Thus, network pharmacology combines systems biology, omics, and computational biology to elucidate the mechanism of drug action with a holistic perspective at the systemic level.39,40
In this study, we first evaluated the anti-IR effect of MOS extracts. Second, to elucidate the potent active components and investigate the mechanism by which the anti-IR effects of MOS are mediated, we adopted a network pharmacology approach involving drug-likeness evaluation, oral bioavailability prediction, component target prediction, IR-related to target collection, and network construction combined with network analysis. Third, we used molecular docking technology to verify the results of our network analysis. Finally, the key network analysis results were further verified by biological research in a HepG2 cell model. As a result, we identified glycosidic isothiocyanates and glycosidic benzylamines as potential major active components by acting on three core targets consisting of proto-oncogene tyrosine-protein kinase (SRC), tyrosine-protein phosphatase non-receptor type 1 (PTPN1), and caspase-3 (CASP3). The three core targets were significantly related to the regulation of inflammatory response, IR, and insulin pathway. To our knowledge, this study is the first to use a network pharmacological approach to study the anti-IR effects and mechanism of action of MOS, providing a powerful and promising platform for the development and application of MOS to IR-related disease therapy.
Materials and Methods
Chemical Constituents Database Building
All chemical constituents of M. oleifera seeds were collected from online published literature in the following electronic literature databases: PubMed (http://www.ncbi.nlm.nih.gov), SciFinder (http://scifinder.cas.org), and the China National Knowledge Infrastructure (CNKI) (http://cnki.net/). A total of 46 chemical constituents were collected.
Active Compound Screening
As an orally administered medicine, we predicted the oral bioavailability (OB), drug-likeness and Caco-2 permeability of MOS. OB in vivo (%F), the fraction of the orally administered dose that reaches the systemic circulation unchanged, is a good indicator of the efficiency of oral administration for drug delivery into the systemic circulation, and one of the most commonly used pharmacokinetic parameters in drug screening cascades.41,42 According to Lipinski’s rule of five, drug-likeness (DL) describes the pharmacokinetic and pharmaceutical properties of compounds, including molecular weight, hydrogen bond donors, hydrogen bond acceptors, octanol-water partition coefficient log P (AlogP) and rotatable bonds,43 and is an important indicator of the chemical suitability of the compound. Compounds that meet both the OB and DL criteria are more likely to be accepted as candidate compounds. Orally administered medicine is required to permeate the intestine for absorption. Caco-2 permeability is a valuable predictor of in vivo intestinal permeability. Therefore, in this study, the active compounds were screened by OB ≥30%, DL ≥0.18 and Caco-2 ≥–0.4.43,44 Isothiocyanates and benzylamines in MO have been reported to have significant pharmacological activity on the improvement of IR.45,46 Therefore, all isothiocyanates and benzylamines were selected for further investigation regardless of whether these compounds met the requirements.
Protein Target Database Construction
Active Compound-Target Prediction
Predicting interactions between active compounds and target proteins can help decipher the underlying biological mechanisms and is a critical phase in the drug discovery process. In general, an active compound may interact with multiple target proteins. The structures of all of the candidate active compounds were transformed into SDF or Canonical SMILES formats. To obtain as many targets as possible, several databases were employed to this study, including the Swiss Target Prediction (Swiss, http://www.swiss target prediction.ch/),47 Similarity Ensemble Approach (SEA, http://sea.bkslab.org/), PharmMapper Server (PharmMapper, http://lilab.ecust.edu.cn/pharmmapper/submit_file.php)48 and STITCH (http://stitch.embl.de/) databases, and all of the targets were limited to Homo sapiens. The obtained targets were then imported into UniProt (https://www.uniprot.org/) to obtain the standard gene name and predict their functions.
Protein Target Database Construction
GeneCards is a human gene database (http://www.genecards.org/) that provides comprehensive, user-friendly information on all annotated and predicted human genes, and automatically integrates gene-centric data from approximately 150 web sources, including genomic, transcriptomic, proteomic, genetic, clinical and functional information.49 IR-associated targets were screened from GeneCards using “insulin resistance” as the key search term and a cut-off score ≥10. By comparing IR-associated targets with active compound targets, the common targets were screened to build the protein target database.
Network Construction, Gene Ontology (GO) Analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analysis
Protein-protein interactions (PPI) are critically important in many cellular processes, including signal transduction, regulation of gene expression, and cell migration.43 The targets obtained were imported into STRING (https://string-db.org/) to study protein-protein interactions.50 To improve the reliability of the data, the protein-protein interactions were further filtered with a minimum combined score of 0.7 (high confidence),51 and the targets without interactions were removed. The remaining PPIs were used for network construction and analysis. The DAVID (Functional Annotation Result Summary, https://david.ncifcrf.gov/summary.jsp) webserver was used for GO enrichment analysis and KEGG pathway analysis. GO enrichment and KEGG pathways with P-values <0.01 were selected, while IR-resistance irrelevant pathways were not.43 Compound-target-pathway networks were constructed using Cytoscape 3.7.0 (http://www.cytoscape.org/). In graphical representations of the networks, compounds, targets and pathways were described by nodes, and the interactions were encoded by edges.
Target Verification by Molecular Docking Using the Molecular Operating Environment Tool
The Molecular Operating Environment (MOE, version 2018) tool is used to predict the binding affinities of related compounds from a known active series and the binding mode of a known active ligand in addition to the identification of new ligands by virtual screening. The candidate compounds were drawn by ChemDraw in the SDF format. Crystallographic structures of proteins were downloaded from the Protein Data Bank (https://www.rcsb.org/). The candidate compounds and the corresponding targets were uploaded to the software of MOE to obtain docking scores (S values). It is generally believed that the lower the energy when the conformation of ligand binding to the receptor is stable, the greater the possibility that an interaction will occur; binding energy (docking score, S) ≤− 5.0 kJ/mol was used as the screening criterion.52
Experimental Validation
Chemicals and Reagents
MOSs were purchased from Kunming district (Yunnan Province, China) in 2019. The materials were authenticated by the authors, and a voucher specimen (ID 2,019,001) was deposited at the authors’ laboratory. HepG2 cells were purchased from the Bioresource Collection and Research Center (Shanghai, China) and cultured in Dulbecco’s modified Eagle’s medium (DMEM) (Thermo Fisher Scientific, New York, USA) with 10% heat-inactivated fetal bovine serum (FBS), and 1% penicillin/streptomycin. Insulin (Sigma, 91077C) and CCK-8 (Dojindo, CK04) were also used.
Sample Preparation
MOS (10 kg) was extracted with H2O (10 L, 3χ1 h). The aqueous extracts were evaporated under reduced pressure and lyophilized to yield a dark brown dried aqueous extracts (2650 g). The MOS residue was further extracted with 90% ethanol (10L, 3χ1 h). The ethanol extracts were then evaporated under reduced pressure, and lyophilized to yield a dark brown dried ethanol extract (413 g).
Cell Viability
HepG2 cells (1×104 cells per well) were seeded in a 96-well flat-bottomed culture plates and incubated at 37°C overnight in a humidified environment under 5% CO2 and 95% air to allow cell adherence. Cells were then treated with MOS extracts. After removal of the medium, 100 χL CCK8 was added to each well and the plate was incubated for a further 2 h. The absorbance was measured at 570 nm with a microplate reader (BioTek Instruments).
Glucose Consumption Assay in the HepG2-Insulin Resistance Model (HepG2-IR)
HepG2 cells (1×104 cells per well) were treated with insulin (10–6 mol/L) for 24 h with or without MOS extracts (1.0 ng/mL–10 μg/mL). Glucose concentration was quantified using glucose oxidase-peroxidase (GOD-POD) kits (Nanjing Jiancheng Biology Engineering Institute, F006-1-1). The absorbance of samples was measured at 505 nm and glucose uptake was calculated based on the difference between the glucose content of the final cell supernatant and the original medium. Each sample was analyzed in triplicate.
Western Blot Analysis
HepG2 cells (1×104 cells per well) were seeded in a 96-well flat-bottomed culture plate and incubated at 37°C overnight in a humidified environment under 5% CO2. The proteins were extracted from HepG2 cells. The samples were then separated by 10% SDS-PAGE and transferred to 0.45 µm polyvinylidene difluoride (PVDF) membranes. Total proteins were analyzed by immunoblotting using anti-SRC (Boster Biological Technology Co. Ltd. PB9059; 1:1000), anti-CASP3 (Boster Biological Technology Co. Ltd. PB9188; 1:1000), anti-PTPN1 (Boster Biological Technology Co. Ltd. A00613-1; 1:1000), and anti-β-actin (Affinity T0022; 1:3000) as primary detection antibodies. Finally, the membranes were visualized using an ImageQuantLAS imaging system (General Electric Company, Boston, USA). This experiment was repeated in triplicate. For quantitative analysis, the density of bands was analyzed using Image J software (National Institute of Mental Health, Bethesda, MD, USA).
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 7.0 software. All data were expressed as mean ± standard deviation (SD) of triplicate experiments. One-way analysis of variance (ANOVA) was used to analyze the data from multiple groups. A P-value <0.05 was considered to indicate statistical significance.
Results
Database Construction
Compounds Database Construction and Target Prediction
A total of 46 chemical constituents of MOS were identified in multiple databases, and 32 active compounds were screened (Figure 1). It is worth noting that isothiocyanates and benzylamines have relatively low OB values, but are known to produce effective anti-IR activity or improve glucose tolerance.45,46 Hence, eight isothiocyanates and 15 benzylamines were also selected for inclusion in the next step of our analysis, despite not fulfilling the criteria for OB and DL. Indeed, the present data showed that almost all candidate compounds showed low oral bioavailability (OB ≤30%) and poor drug-like characteristics (DL ≤0.18). However, isothiocyanates and benzylamines were dominant active components in MOS. In addition to the potential pharmacological effects of compounds, another nine compounds (OB ≥30%, DL ≥0.18 and Caco-2 ≥–0.4), including phenylpropanoids and flavonoids, were also regarded as candidate compounds. Screening of MOS yielded a total of 32 candidate compounds.
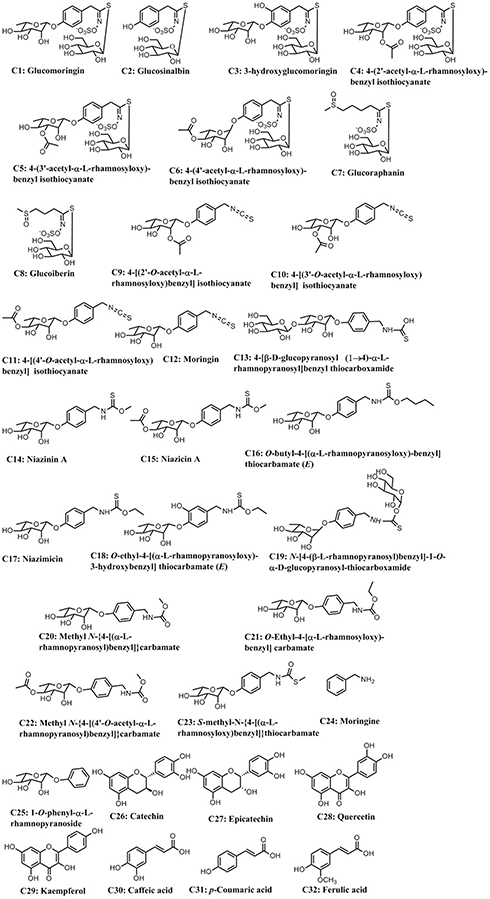 |
Figure 1 Structures of 32 potentially active compounds. |
By searching the Swiss Target Prediction, Similarity Ensemble Approach, and PharmMapper Server databases, a total of 71 targets of candidate compounds were identified. These targets were tyrosine-protein phosphatase-associated targets, insulin-like growth factor-binding proteins, mitogen-activated protein kinase-associated targets, and other anti-tumor- or anti-inflammatory-associated targets.
Protein Target Database Construction
By searching GeneCards, 555 IR-associated targets (score ≥10) were identified. By comparison of 555 IR-associated targets with 71 compound targets, and considering protein-protein interactions (PPI score >0.7), a total of 44 common targets were screened and used to build the protein target database (Table 1). The PPIs of 44 IR-associated targets are shown in Figure 2. The thickness of the line between the two targets indicates the strength of the supported data, with thicker lines indicating stronger interactions.43 These targets included mainly anti-inflammatory-associated targets, such as MAPKs, nitric oxide synthases (NOSs), anti-diabetic-associated targets, such as PTPN1, PTPN11, IGF1R, PIK3R1, and other inflammatory- or IR-associated targets.
 |
Table 1 Forty-Four Insulin Resistance-Related Targets of M. oleifera Seeds Information |
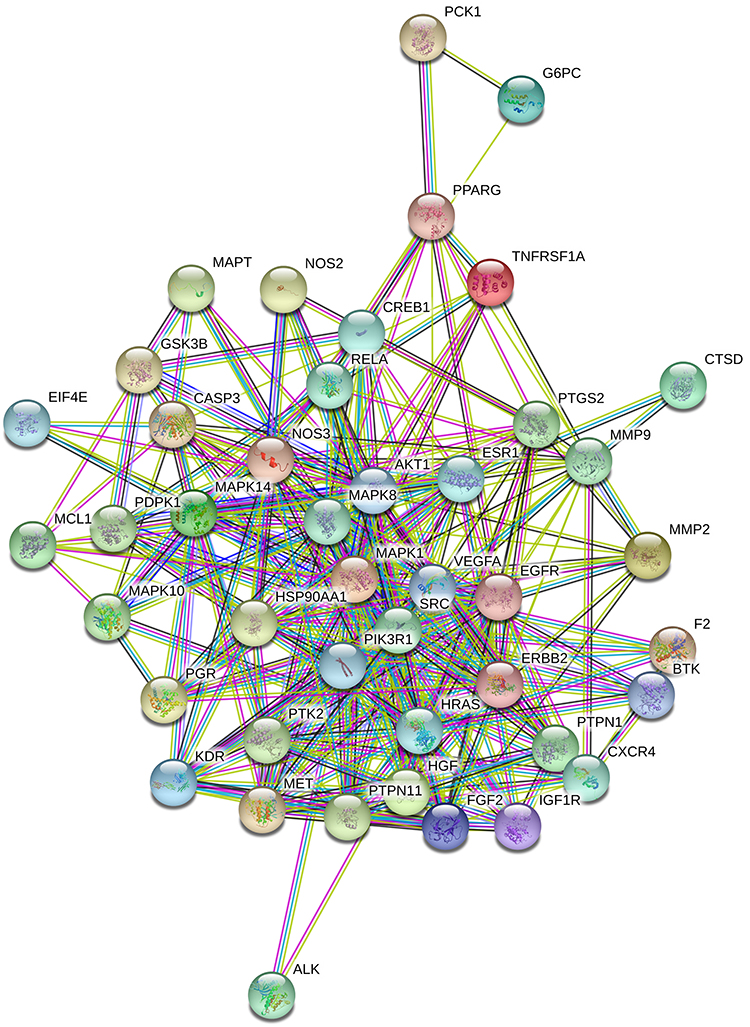 |
Figure 2 Protein-protein interactions (P > 0.7) of 44 insulin-resistance-related targets of MOS. |
Bioinformatics Analysis
Gene Ontology and KEGG Pathway Enrichment Analysis
GO enrichment analysis was then performed to further investigate the molecular function, biological process, and cellular component of the component’s targets using DAVID bioinformatics resources (version 6.8). The top 15 GO enrichment terms are shown in Figure 3 and the pathway enrichment categories (P <0.01) are shown in Figure 4. According to the GO enrichment analysis, the functions of target proteins were classified into two categories: inflammatory response and insulin resistance. In detail, the top three terms in the GO biological processes were as follows: the PI3K/Akt signaling pathway, IR and focal adhesion via insulin-like growth factor II binding, and insulin-like growth factor I binding and transmembrane receptor protein tyrosine kinase activation in the cytosol, extracellular exosome and membrane raft. Insulin-like growth factor binding indicates that insulin-like growth factors bind insulin-like growth factor systems to exert their biological effects. Insulin-like growth factors are involved in the regulation of metabolic pathways53 and insulin sensitivity.54 Insulin-like growth factor 1 receptor (IGF1R), a transmembrane receptor, belongs to the receptor tyrosine kinase family,55 which has been shown to increase the sensitivity of the liver and muscle to insulin and to improve the ability of insulin to mediate physiological functions that protect against type 2 diabetes.56 Insulin stimulates translocation of the IGF2R to the cell surface, promoting insulin-like growth factor 2 (IGF2) clearance. IGF2 may increase glucose uptake into human adipocytes.57 Tyrosine phosphorylation of key proteins in the insulin signaling pathway is critical for insulin action.57,58
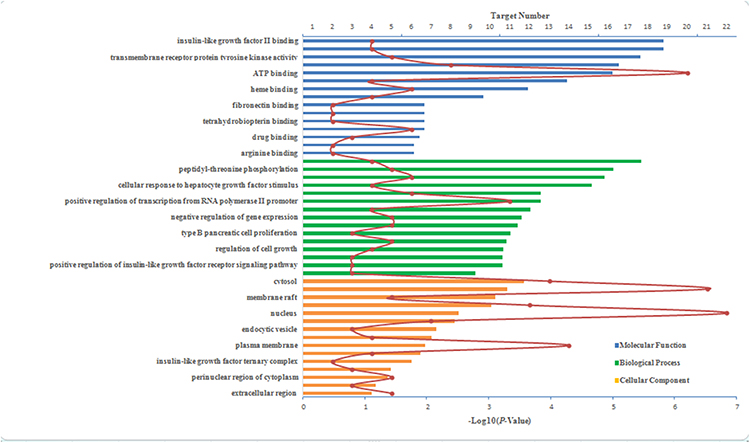 |
Figure 3 Top 15 GO enrichment terms. |
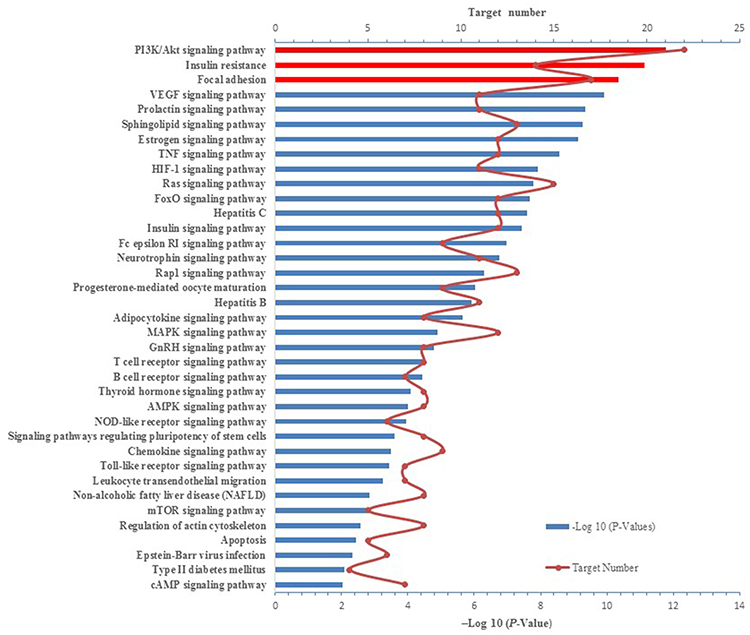 |
Figure 4 KEGG pathway enrichment analysis of 44 insulin-resistance-related targets. |
According to the KEGG enrichment analysis and after deleting irrelevant broad-spectrum pathways,43 a total of 37 potential pathways were identified with a cut-off P-value of 0.01. The top three enriched pathways with the lowest P-values. Therefore, considered to be the most important pathways (Figure 4), were PI3K/Akt signaling pathway, IR, and focal adhesion. The PI3K/Akt pathway is an important insulin signaling pathway that is closely related to type 2 diabetes caused by IR. This pathway plays an important role in the regulatory effects of insulin on blood sugar, the activation of glucose transporters, and the initiation of glucose uptake into cell storage or decomposition.59 Insulin-like growth factor I ameliorates type 2 diabetes by binding to insulin-like growth factor I receptor located on the cell membrane, resulting in activation of its downstream PI3K/Akt pathway.56 Dysregulation of the PI3K/AKT pathway in various tissues leads to conditions such as obesity and type 2 diabetes by increasing IR. In turn, IR exacerbates the PI3K/AKT pathway, forming a vicious cycle.60 Focal adhesion, otherwise known as cell-matrix adhesion, plays essential roles in important biological processes, including cell motility, proliferation and differentiation, as well as regulation of gene expression and cell survival. Furthermore, focal adhesion was identified as an important pathway in IR.61 Focal adhesion kinase (FAK), a non-receptor tyrosine kinase, has recently been implicated in the regulation of IR by regulating glucose uptake in skeletal muscle cells.61,62
The two main therapeutic effects of MOS are mediated by its ability to regulate inflammatory responses and improve IR. Through the synergistic effects on these two systems, MOS can alleviate IR. Thus, the theory of MOS treatment is based on its ability to modulate the release of inflammatory factors that interfere with insulin function.
Network Analysis
A global view of the compound-target-pathway network, which contained 117 nodes and 561 edges (Figure 5), was constructed to further understand the molecular mechanism by which MOS functions to improve IR. This network suggested the existence of complex correlations among different compounds and targets, and provided a clear understanding of the complex network relationships in terms of the “one ingredient-multiple targets” and “one target-multiple ingredients” models.
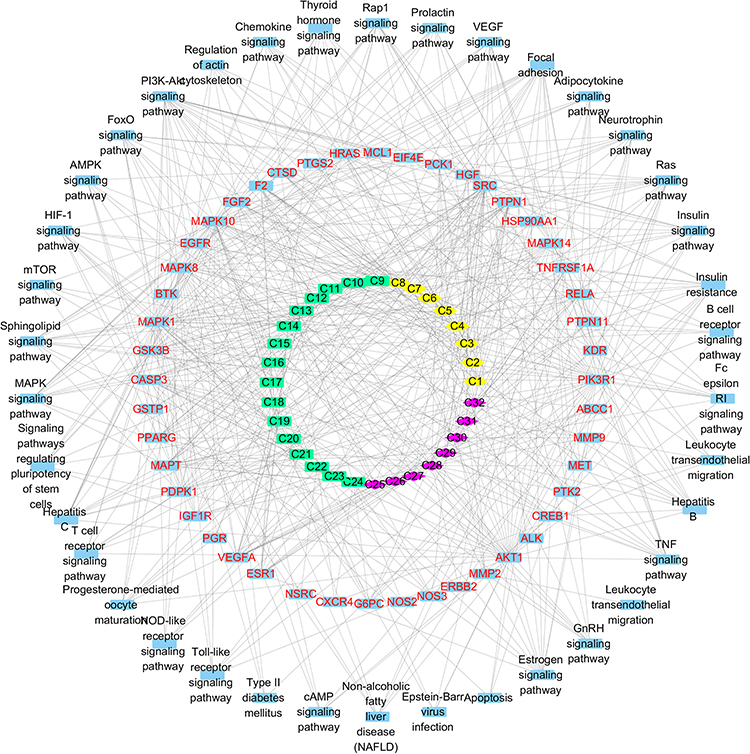 |
Figure 5 The compound-target-pathway network of MOS. Different species of active compounds are depicted by different shapes and colors. The compounds are classified into three groups: isothiocyanates (yellow hexagons), benzylamine (green squares), and phenolic compounds (purple diamonds). |
The top three target nodes linked to the greatest number of compound nodes were SRC, PTPN1, and microtubule-associated protein tau (MAPT) (Figure 6A). These targets are involved in multiple pathways. For example, focal adhesion and the vascular endothelial growth factor (VEGF) signaling pathway, IR and the insulin signaling pathway, the MAPK signaling pathway, and regulation of the actin cytoskeleton. The top three compound nodes linked to the greatest number of target nodes were quercetin (compound 28), caffeic acid (compound 30), and ferulic acid (compound 32). However, isothiocyanates and benzylamines are known to be active compounds that improve IR.45,46 As isothiocyanates and benzylamines might function synergistically to improve IR, we identified SRC, PTPN1, and CASP3 as the top three targets linked to the greatest number of isothiocyanates and benzylamines (Figure 6B). These targets are involved in multiple important pathways, such as focal adhesion, IR, the insulin signaling pathway, and the MAPK signaling pathway. These results indicate that MOS improves IR in a multi-target, multi-pathway, and synergistic manner.
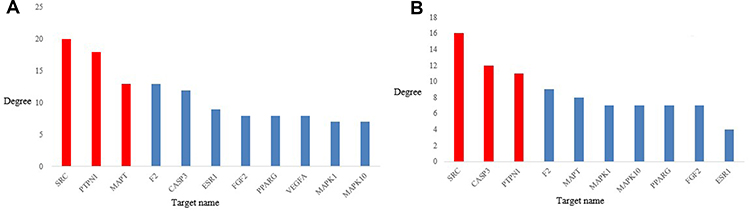 |
Figure 6 (A) Top 10 target nodes linked to the greatest number of compound nodes. (B) Top 10 target nodes linked to more isothiocyanates and benzylamines. (Red bars indicate the top 3 target nodes). |
Target Validation by Molecular Docking Analysis
Although SRC, PTPN1 and MAPT were linked to the greatest number of compound nodes, SRC, PTPN1, and CASP3 were the top three targets linked to the greatest number of isothiocyanates and benzylamines. SRC, PTPN1, and CASP3 were selected as targets for docking, because isothiocyanates and benzylamines were the dominant active constituents that improve IR. Docking scores (S values) between compounds and targets were calculated by MOE. Three pairs of compound-target docking relationships were built and the results are shown in Figure 7 and Table 2. The S values of the vast majority of compounds were below −5.0. However, the S values of compound 24 were greater than −5.0 for the three targets. The S values of compounds 30, 31 and 32 were also greater than −5.0 for the target SRC. Among them, all targets of diglycosidic isothiocyanates (compounds 3–6) showed the most effective binding, and all targets of glycosidic benzylamines showed good binding. These results provided important evidence in support of the discovery if possible effective compounds shown in Figure 7. Taken together, isothiocyanates and benzylamines are the major and effective constituents that mediate the anti-IR effects of MOS.
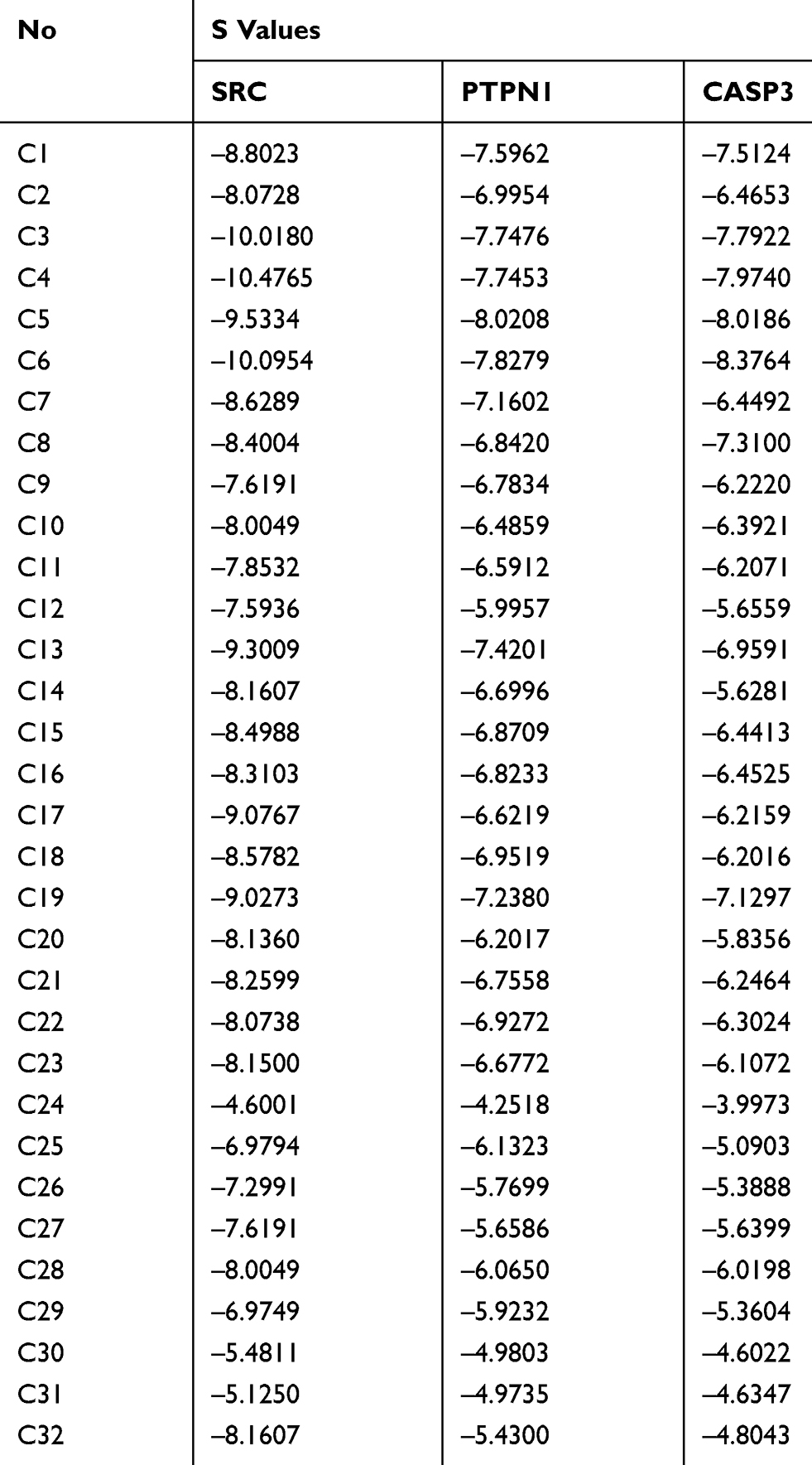 |
Table 2 Detailed Docking Values (S) of 32 Potential Active Compounds for SRC, PTPN1, and CASP3 |
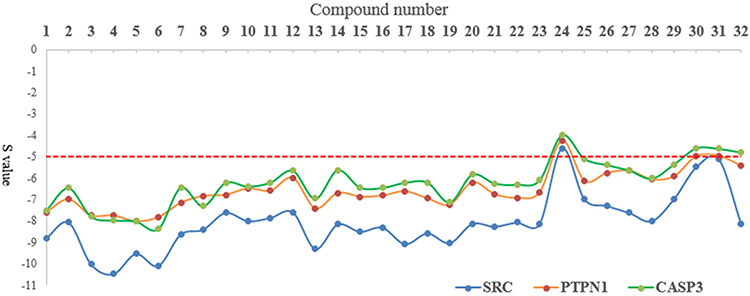 |
Figure 7 Line chart of molecular docking results (S values) of 32 potentially active compounds related to SRC, PTPN1, and CASP3. |
Experimental Validation
Effects of MOS Extracts on Glucose Consumption in Insulin-Resistant HepG2 Cells
The cytotoxic effects of MOS extracts were investigated in insulin-resistant HepG2 cells. There were no significant differences in the cytotoxic effects of MOS on cells treated with extracts in the range of 1.0 ng/mL to 10 μg/mL. Treatment with aqueous and ethanol extracts of MOS both significantly increased glucose consumption by HepG2-IR cells in a concentration-dependent manner. As shown in Figure 8, the glucose consumption of HepG2-IR cells treated with MOS extracts at 10 ng/mL, 100 ng/mL, 1 μg/mL or 10 μg/mL was approximately 2.0-fold higher, respectively, than that in the control culture without MOS extracts. Previous publications have reported that benzylamine-rich and isothiocyanate-rich MO leaf or bark extracts effectively improved glucose tolerance, and lowered blood insulin levels.32–36,45,46 These results further highlighted the roles of benzylamines and isothiocyanates as the major effective anti-IR-mediating components of MOS.
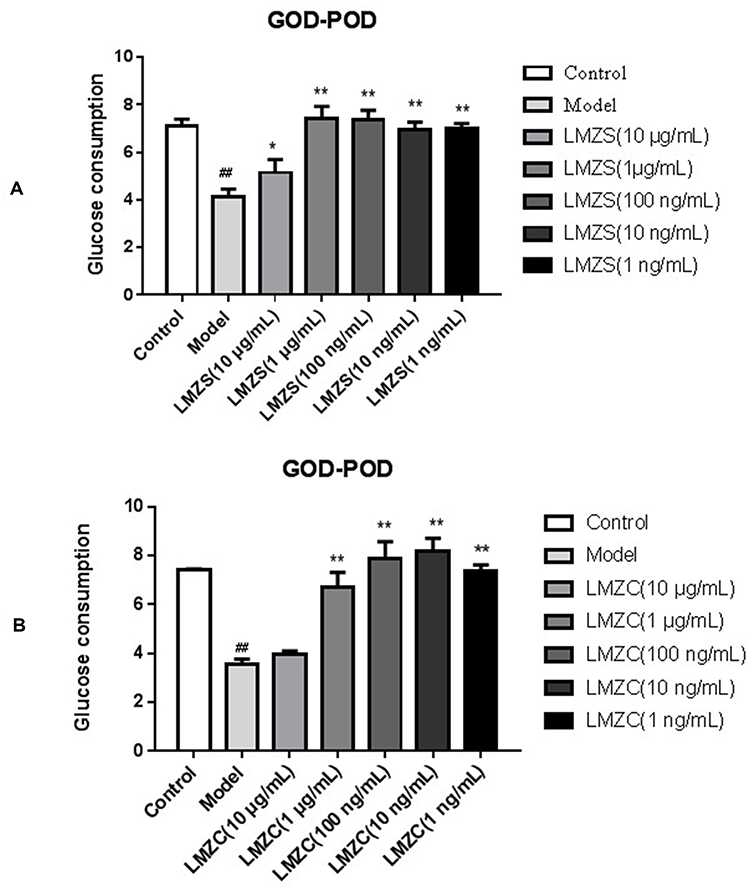 |
Figure 8 Glucose consumptions of MOS extracts in the HepG2-IR model (A) MOS aqueous extract; (B) MOS ethanol extract; Model vs Control, ##P < 0.01; MOS aqueous extract or MOS ethanol extract vs Model, *P < 0.05, **P < 0.01). |
Effects of MOS Extracts on SRC, PTPN1 and CASP3 Protein Expression
The major effective compounds of MOS, such as isothiocyanates and benzylamines, are not commercially available and difficult to purify in the laboratory. Therefore, we used aqueous and ethanol extracts of MOS to verify the effects of these components on SRC, PTPN1, and CASP3 targets. SRC, a non-receptor tyrosine kinase and a member of the SRC family of protein tyrosine kinases, regulates a wide range of cellular functions, and hyperactivity of SRC is involved in impaired glucose metabolism.63 The HepG2 cells in the model groups were treated with 1 μm insulin. SRC expression in the model groups was significantly higher than the blank control groups (Figures 9 and 10). This demonstrated the successful generation of the model, and confirmed that insulin upregulates SRC expression in HepG2 cells compared with that in the model group.64 In the concentration range of 100 ng/mL–10 μg/mL, the ethanol extract of MOS significantly suppressed SRC expression in HepG2 cells (Figure 9). Similarly, in the concentration range of 10 ng/mL–10 μg/mL, the aqueous extract of MOS also significantly suppressed SRC expression in HepG2 cells (Figure 10). At 10 ng/mL, the aqueous extract of MOS significantly effects suppressed SRC expression, whereas the ethanol extract of MOS had no effect. In the concentration range of 10 ng/mL–10 μg/mL, the aqueous and ethanol extracts of MOS both significantly downregulated the PTPN1 expression compared with that in the model group (Figures 9 and 10). Moreover, the aqueous extract of MOS exerted more effective suppression of PTPN1 than the ethanol extract of MOS.
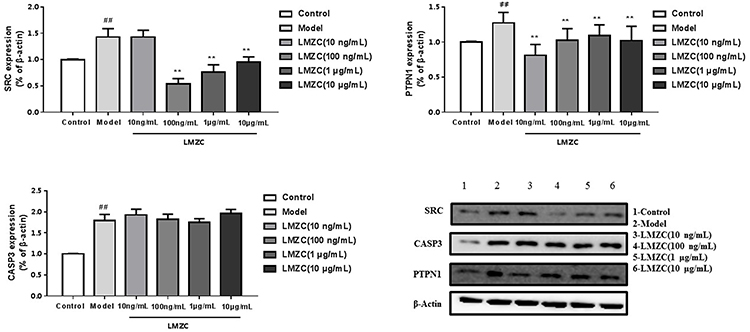 |
Figure 9 Effect of MOS ethanol extract on the expression of SRC, PTPN1, and CASP3 (Model vs Control, ##P < 0.01; MOS ethanol extract vs Model, **P < 0.01). |
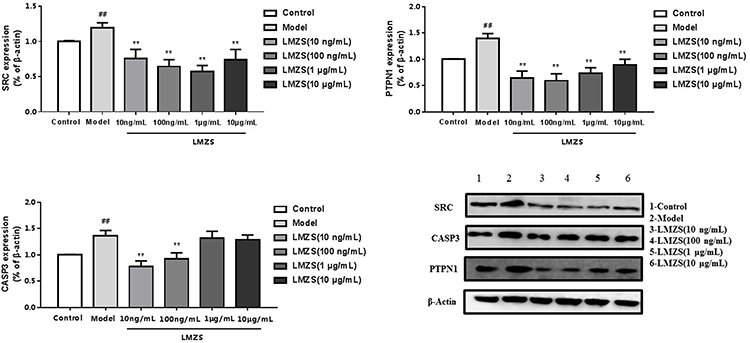 |
Figure 10 Effect of MOS aqueous extract on the expression of SRC, PTPN1, and CASP3 (Model vs Control, ##P < 0.01; MOS aqueous extract vs Model, **P < 0.01). |
CASP3 is involved in the activation cascade of caspases responsible for apoptosis execution. CASP3 induces islet beta cell apoptosis. In this study, CASP3 expression in model group was significantly higher than that in the blank control group. However, the ethanol extract of MOS treated group did downregulate CASP3 expression. In the concentration range of 10 ng/mL or 100 ng/mL, treatment with the aqueous extract of MOS significantly downregulated CASP3 expression, but it did not significantly suppress CASP3 expression following treatment (1 μg/mL or 10 μg/mL). It can be speculated that this results is due to the toxic effects of MOS extracts on CASP3 expression in HepG2 cells. In the molecular docking analysis (Figure 7), the S values of most compounds and CASP3 were greater than those of the other targets. This demonstrated that compounds in MOS improve IR mainly by acting on SRC and PTPN1B rather than CASP3.
Discussion
MOS is known to exert anti-diabetic effects possibly by improving IR, although the active components and their mechanisms of action are unclear. In this study, we used the network pharmacology approach to predict the potentially active components and their mechanisms of action. Our network pharmacology analysis revealed that glycosidic isothiocyanates and glycosidic benzylamines were the major active components responsible for improving IR. Biological studies showed that the aqueous and ethanol extracts of MOS both exhibited anti-IR effects, although the effects of the aqueous extract of MOS were stronger than those of the ethanol extract. Our biological studies implicated the macropolar compounds as the active compounds, which was consistent with the results of our network pharmacology analysis. Although most glycosidic isothiocyanates and glycosidic benzylamines did not meet the screening requirements, these compounds were still included in the network construction because of their reported anti-IR effects. Hence, structural modification of glycosidic isothiocyanates and glycosidic benzylamines is highlighted as a viable approach to the development of novel anti-IR agents.
The results of preliminary KEGG pathway analysis included cancer related pathways, such as proteoglycans in cancer, pathways in cancer, and prostate cancer, because many targets such as SRC, CASP3, and IGF1R were involved in either multiple cancer pathways or insulin resistance-related pathways. It can be speculated that the mechanisms underlying the pathogenesis of IR and tumorigenesis show some similarities through the involvement of common pathways such as the PI3K/PTEN/Akt signaling pathway.65 Although IR is involved in the pathogenesis of cancers,66 the aim of this study was to clarify the pathogenesis of IR rather than cancer. Therefore, to establish an effective network pharmacology model, we deleted irrelevant broad-spectrum pathways, such as those involved in cancer. Subsequently, the predominant inflammatory response- and insulin-resistance-related pathways were found to include the PI3K/Akt signaling pathway, IR, and focal adhesion. Growing evidence has indicated the role of the chronic low-grade inflammatory response in the pathogenesis of IR.67
The molecular docking analysis results (Figure 7) showed that the pattern of binding ability of most of the potent active compounds and three targets were as follows: SRC ≥PTPN1 ≥CASP3. Combined with the results of Western blot analysis, our molecular docking analysis results clearly demonstrated that the anti-IR effects of MOS were exerted mainly via SRC and PTPN1 rather than CASP3. KEGG pathway analysis indicated that SRC was involved in processes such as focal adhesion, VEGF signaling, and prolactin signaling and PTPN1 was involved in IR and the insulin signaling pathway. Although PTPN1 was involved in only two pathways, as one of the top three targets, it was identified as a key target linked to a large number of compounds.
Although network pharmacology can provide us with a vast amount of information in mechanistic studies and prediction of the potent active compounds in a complicated multiple component system, the results of network pharmacology should be analyzed with a degree of caution. Furthermore, biological verification is an essential step to confirm that the predicted key targets are the actual key targets and to rule out the existence of irrelevant or non-objective pathways. Numerous natural products, such as flavonoids, phenols, and phenyl propanoids, show the ability to regulate glucose metabolism and improve IR activity.11,68 Although glycosidic isothiocyanates and glycosidic benzylamines have been reported to play a major role in anti-IR activity, other compounds such as 26, 27, 28, 29, 30, 31 and 32 may possess potential activity mediated via various pathways. Moreover, these compounds may play a synergistic role in the anti-IR activity of MOS.
To date, there is no consensus on the criteria for a network pharmacology method. The chemical constituents databases, active compound screening rules, protein target databases, PPI screening score, and molecular docking software vary widely between different studies. Thus, the results of the same study might be a little bit different. Therefore, comprehensive and effective criteria for the design of future network pharmacology studies are required to ensure the reliability and consistency of results.
Conclusion
In this study, we adopted a systematic network pharmacology approach to investigation of the anti-IR effects of MOS. Furthermore, network pharmacology analysis combined with validation of our experimental results in a HepG2-IR cell model indicate that the anti-IR effects of MOS are manifested by mechanisms that regulate inflammatory responses, insulin resistance and the insulin signaling pathway. Moreover, glycosidic isothiocyanates and glycosidic benzylamines are implicated as the major active components that act on SRC and PTPN1 as important targets. Our results provide the basis of further studies to fully elucidate the anti-IR effects of MOS.
Acknowledgment
The authors greatly appreciate the technical support about MOE from Prof. Dongsheng Cao (Central South University, Changsha, China) during the conduct of this study.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper and report no conflicts of interest for this work.
References
1. Taylor R. Insulin resistance and type 2 diabetes. Diabetes. 2012;61(4):778–779. doi:10.2337/db12-0073
2. Reaven G. Insulin resistance, hypertension, and coronary heart disease. J Clin Hypertens. 2003;5(4):269–274.
3. Gupta A, Gupta V. Metabolic syndrome: what are the risks for humans? Biosci Trends. 2010;5:204–212.
4. Neuschwander-Tetri BA. Non-alcoholic fatty liver disease. BMC Med. 2017;15(1):45.
5. Cederberg A, Enerbäck S. Insulin resistance and type 2 diabetes–an adipocentric view. Curr Mol Med. 2003;3(2):107–125.
6. Martins AR, Nachbar RT, Gorjao R, et al. Mechanisms underlying skeletal muscle insulin resistance induced by fatty acids: importance of the mitochondrial function. Lipids Health Dis. 2012;11:30.
7. Bouzakri K, Roques M, Gual P, et al. Reduced activation of phosphatidylinositol-3 kinase and increased serine phosphorylation of insulin receptor substrate-1 in primary culture of skeletal muscle cells from patients with type 2 diabetes. Diabetes. 2003;52(6):1319–1325.
8. Choi K, Kim Y. Molecular mechanism of insulin resistance in obesity and type 2 diabetes. Korean J Intern Med. 2010;25(2):119–129.
9. Plomgaard P, Bouzakri K, Krogh-Madsen R, Mittendorfer B, Zierath JR, Pedersen BK. Tumor necrosis factor-alpha induces skeletal muscle insulin resistance in healthy human subjects via inhibition of Akt substrate phosphorylation. Diabetes. 2005;54(10):2939–2945.
10. Steinberg GR, Michell BJ, van Denderen BJ, et al. Tumor necrosis factor alpha-induced skeletal muscle insulin resistance involves suppression of AMP-kinase signaling. Cell Metab. 2006;4(6):465–474. doi:10.1016/j.cmet.2006.11.005
11. Saadeldeen FSA, Niu Y, Wang HL, et al. Natural products: regulating glucose metabolism and improving insulin resistance. Food Sci Hum Wellness. 2020. doi:10.1016/j.fshw.2020.04.005
12. Ando H, Takamura T, Matsuzawanagata N, et al. The hepatic circadian clock is preserved in a lipid-induced mouse model of non-alcoholic steatohepatitis. Biochem Biophys Res Commun. 2009;380(3):684–688. doi:10.1016/j.bbrc.2009.01.150
13. Lebrun P, Cognard E, Bellon-Paul R, et al. Constitutive expression of suppressor of cytokine signalling-3 in skeletal muscle leads to reduced mobility and overweight in mice. Diabetologia. 2009;52(10):2201–2212.
14. Yaspelkis BB, Kvasha IA, Figueroa TY. High-fat feeding increases insulin receptor and IRS-1 coimmunoprecipitation with SOCS-3, IKKalpha/beta phosphorylation and decreases PI-3 kinase activity in muscle. Am J Physiol Regul Integr Comp Physiol. 2009;296(6):1709–1715. doi:10.1152/ajpregu.00117.2009
15. Minaiyan M, Asghari G, Taheri D, Saeidi M, Nasr-Esfahani S. Anti-inflammatory effect of Moringa oleifera Lam. seeds on acetic acid-induced acute colitis in rats. Avicenna J Phytomed. 2014;4(2):127–136.
16. Randriamboavonjy JI, Rio M, Pacaud P, Loirand G, Tesse A. Moringa oleifera seeds attenuate vascular oxidative and nitrosative stresses in spontaneously hypertensive rats. Oxid Med Cell Longev. 2017;4129459.
17. Lunyera J, Wang D, Maro V, et al. Traditional medicine practices among community members with diabetes mellitus in northern tanzania: an ethnomedical survey. BMC Complement Altern Med. 2016;16(1):282.
18. Muhammad HI, Asmawi MZ, Khan NAK. A review on promising phytochemical, nutritional and glycemic control studies on Moringa oleifera Lam. in tropical and sub-tropical regions. Asian Pac J Trop Biomed. 2016;6(10):896–902.
19. Kumar Saini RK, Sivanesan I, Keum YS. Phytochemicals of Moringa oleifera: a review of their nutritional, therapeutic and industrial significance. 3 Biotech. 2016;6:203.
20. Leone A, Spada A, Battezzati A, et al. Genetic, ethnopharmacology, phytochemistry and pharmacology of Moringa oleifera leaves: an overview. Int J Mol Sci. 2015;16:12791–12835.
21. Wang F, Zhong HH, Chen WK, et al. Potential hypoglycaemic activity phenolic glycosides from Moringa oleifera seeds. Nat Prod Res. 2017;31(16):1869–1874.
22. Amaglo NK, Bennett RN, Curto RBL, et al. Profiling selected phytochemicals and nutrients in different tissues of the multipurpose tree Moringa oleifera L., grown in Ghana. Food Chem. 2010;122:1047–1054.
23. Giacoppo S, Soundara Rajan TS, Nicola GRD, et al. The isothiocyanate isolated from Moringa oleifera shows potent anti-inflammatory activity in the treatment of murine subacute parkinson’s disease. Rejuvenation Res. 2017;20(1):50–63.
24. Bennett RN, Mellon FA, Foidl N, et al. Profiling glucosinolates and phenolics in vegetative and reproductive tissues of the multi-purpose trees Moringa oleifera L. (Horseradish Tree) and Moringa stenopetala L. J Agric Food Chem. 2003;51:3546–3553.
25. Costa RA, Sousa OVD, Hofer E, Mafezoli J, Barbosa FG, Vieira RHSDF. Thiocarbamates from Moringa oleifera seeds bioactive against virulent and multidrug-resistant vibrio species. Biomed Res Int. 2017;7963747.
26. Guevara AP, Vargas C, Sakurai H, et al. An antitumor promoter from Moringa oleifera Lam. Mutation Res. 1999;440:181–188.
27. Singh RSG, Negi PS, Radha C. Phenolic composition, antioxidant and antimicrobial activities of free and bound phenolic extracts of Moringa oleifera seed flour. J Funct Foods. 2013;5:1883–1891.
28. Cheenpracha S, Park EJ, Yoshida WY, et al. Potential anti-inflammatory phenolic glycosides from the medicinal plant Moringa oleifera fruits. Bioorg Med Chem. 2010;18:6598–6602.
29. Oluduro OA, Aderiye BI, Connolly JD, Akintayo ET, Famurewa O. Characterization and antimicrobial Activity of 4-(β-D-glucopyranosyl-1→4-α-L-rhamnopyranosyloxy)-benzyl thiocarboxamide; a novel bioactive compound from Moringa oleifera seed extract. Folia Microbiol. 2010;55(5):422–426.
30. Abdull Razis AF, Ibrahim MD, Kntayya SB. Health benefits of Moringa oleifera. Asian Pac J Cancer Prev. 2014;15:8571–8576.
31. Medeirosa MLSD, Mourab MCD, Napoleãob TH, et al. Nematicidal activity of a water soluble lectin from seeds of Moringa oleifera. Int J Biol Macromol. 2018;108:782–789.
32. Amelia D, Santoso B, Purwanto B, et al. Effects of Moringa oleifera on insulin levels and folliculogenesis in polycystic ovary syndrome model with insulin resistance. Immunol Endocr Metab Agents Med Chem. 2018;18:22–31.
33. Metwally FM, Rashad HM, Ahmed HH, Mahmoud AA, Raouf ERA, Abdalla AM. Molecular mechanisms of the anti-obesity potential effect of Moringa oleifera in the experimental model. Asian Pac J Trop Biomed. 2017;7(3):214–221.
34. Lopez M, Rios-Silva M, Huerta M, et al. Effects of Moringa oleifera leaf powder on metabolic syndrome induced in male wistar rats: a preliminary study. J Int Med Res. 2018;46(8):3327–3336.
35. Sosa-Gutiérrez JA, Valdéz-Solana MA, Forbes-Hernández TY, et al. Effects of Moringa oleifera leaves extract on high glucose-induced metabolic changes in HepG2 cells. Biology. 2018;7(3):37.
36. Tuorkey MJ. Effects of M. oleifera aqueous leaf extract in alloxan induced diabetic mice. Interv Med Appl Sci. 2016;8(3):109–117.
37. Sholapur HN, Patil BM. Effect of Moringa oleifera bark extracts on dexamethasone-induced insulin resistance in rats. Drug Res. 2013;63:527–531.
38. Hao DC, Xiao PG. Network pharmacology: a rosetta stone for traditional chinese medicine. Drug Dev Res. 2014;75(5):299–312.
39. Hopkins AL. Network pharmacology. Nat Biotechnol. 2007;25(10):1119–1126.
40. Hopkins AL. Network pharmacology: the next paradigm in drug discovery. Nat Chem Biol. 2008;4(11):682–690.
41. Tao W, Xu X, Wang X, et al. Network pharmacology-based prediction of the active ingredients and potential targets of chinese herbal Radix Curcumae formula for application to cardiovascular disease. J Ethnopharmacol. 2013;145:1–10.
42. Tang F, Tang Q, Tian Y, Fan Q, Huang Y. Network pharmacology-based prediction of the active ingredients and potential targets of Mahuang Fuzi Xixin decoction for application to allergic rhinitis. J Ethnopharmacol. 2015;176:402–412.
43. Shi XQ, Yue SJ, Tang YP, et al. A network pharmacology approach to investigate the blood enriching mechanism of Danggui buxue Decoction. J Ethnopharmacol. 2019;235:227–242.
44. Zhu J, Yi X, Zhang Y, Pan Z, Zhong L, Huang P. Systems pharmacology-based approach to comparatively study the independent and synergistic mechanisms of Danhong injection and Naoxintong capsule in ischemic stroke treatment. Evid Based Complement Alternat Med. 2019;1056708.
45. Iffiú-Soltész Z, Wanecq E, Lomba A, et al. Chronic benzylamine administration in the drinking water improves glucose tolerance, reduces body weight gain and circulating cholesterol in high-fat diet-fed mice. Pharmacol Res. 2010;61(4):355–363.
46. Waterman C, Rojas-Silva P, Tumer TB, et al. Isothiocyanate-rich Moringa oleifera extract reduces weight gain, insulin resistance and hepatic gluconeogenesis in mice. Mol Nutr Food Res. 2015;59(6):1013–1024.
47. Daina A, Michielin O, Zoete V. SwissTargetPrediction: updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019;47(W1):W357–W364.
48. Wang X, Shen Y, Wang S, et al. PharmMapper 2017 update: a web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017;45:W356W360.
49. Shawky E. Prediction of potential cancer-related molecular targets of north african plants constituents using network pharmacology-based analysis. J Ethnopharmacol. 2019;238:111826.
50. Zhai J, Song Z, Wang Y, et al. Zhixiong capsule (ZXC), a traditional chinese patent medicine, prevents atherosclerotic plaque formation in rabbit carotid artery and the related mechanism investigation based on network pharmacology and biological research. Phytomedicine. 2019;59:152776.
51. Chen Y, Dong J, Liu J, et al. Network pharmacology-based investigation of protective mechanism of Aster tataricus on lipopolysaccharide-induced acute lung injury. Int J Mol Sci. 2019;20:543.
52. Zong Y, Ding M, Jia K, Ma S, Ju W. Exploring the active compounds of Da-Yuan-Yin in treatment of novel coronavirus (2019-nCoV) pneumonia based on network pharmacology and molecular docking method. Chin Trad Herbal Drugs. 2020;51(4):836–844.
53. Inzaghi E, Baldini Ferroli B, Fintini D, Grossi A, Nobili V, Cianfarani S. Insulin-like growth factors and metabolic syndrome in obese children. Horm Res Paediatr. 2017;87(6):400–404.
54. Kujawska-Luczak M, Szulinska M, Skrypnik D, et al. The influence of orlistat, metformin and diet on serum levels of insulin-like growth factor-1 in obeses women with and without insulin resistance. J Physiol Pharmacol. 2018;69(5):737–745.
55. Dong L, Hou X, Liu F, et al. Regulation of insulin resistance by targeting the insulin‐like growth factor 1 receptor with microRNA-122-5p in hepatic cells. Cell Biol Int. 2019;43(5):553–564.
56. Halperin-Sheinfeld M, Gertler A, Okun E, Sredni B, Cohen HY. The tellurium compound, AS101, increases SIRT1 level and activity and prevents type 2 diabetes. Aging. 2012;4(6):436–447.
57. Lin X, Tang S, Gui W, et al. Circulating miR-143-3p inhibition protects against insulin resistance in metabolic syndrome via targeting of the insulin-like growth factor 2 receptor. Transl Res. 2019;205:33–43.
58. Ahmad F, Goldstein BJ. Increased abundance of specific skeletal muscle protein-tyrosine phosphatases in a genetic model of insulin-resistant obesity and diabetes mellitus. Metabolism. 1995;44(9):1175–1184.
59. Czech MP. Insulin action and resistance in obesity and type 2 diabetes. Nat Med. 2017;23(7):804–814.
60. Huang X, Liu G, Guo J, Su Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int J Biol Sci. 2018;14(11):1483–1496.
61. Bisht B, Goel HL, Dey CS. Focal adhesion kinase regulates insulin resistance in skeletal muscle. Diabetologia. 2007;50(5):1058–1069.
62. Bisht B, Dey CS. Focal adhesion kinase contributes to insulin-induced actin reorganization into a mesh harboring Glucose transporter-4 in insulin resistant skeletal muscle cells. BMC Cell Biol. 2008;9:48.
63. Sato H, Nagashima K, Ogura M, et al. Src regulates insulin secretion and glucose metabolism by influencing subcellular localization of glucokinase in pancreatic β-cells. J Diabetes Investig. 2016;7(2):171–178.
64. Liu Y, Chen XL, Wang L, Martins-Green M. Insulin antagonizes thrombin-induced microvessel leakage. J Vasc Res. 2017;54(3):143–155.
65. Li A, Qiu M, Zhou H, Wang T, Guo W. PTEN, insulin resistance and cancer. Curr Pharm Des. 2017;23(25):3667–3676.
66. Djiogue S, Nwabo Kamdje AH, Vecchio L, et al. Insulin resistance and cancer: the role of insulin and IGFs. Endocr Relat Cancer. 2013;20(1):R1R17.
67. Matulewicz N, Karczewska-Kupczewska M. Insulin resistance and chronic inflammation. Postepy Hig Med Dosw. 2016;70:1245–1258.
68. Cui LL, Wang JM, Wang MK, et al. Chemical composition and glucose uptake effect on 3T3-L1 adipocytes of Ligustrum lucidum Ait. Flowers. Food Sci Hum Wellness. 2020;9:124–129.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.