Back to Journals » Nutrition and Dietary Supplements » Volume 12
Insights into Novel Infant Milk Formula Bioactives
Authors Skolnick J, Chou C ![]() , Miklavcic J
, Miklavcic J ![]()
Received 11 September 2019
Accepted for publication 15 January 2020
Published 18 February 2020 Volume 2020:12 Pages 11—19
DOI https://doi.org/10.2147/NDS.S192099
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Chandrika Piyathilake
Jordan Skolnick,1 Claire Chou,1 John Miklavcic1,2
1Schmid College of Science and Technology, Chapman University, Orange, CA 92866, USA; 2School of Pharmacy, Chapman University, Irvine, CA 92618, USA
Correspondence: John Miklavcic
Schmid College of Science and Technology, Chapman University Tel +1 714-516-5520
Fax +1 714-532-6048
Email [email protected]
Abstract: Human milk provides optimal infant nutrition for early life development. However, there are situations in which human milk feeding is not the most viable option for infant nutrition. Some factors include maternal illness, lactation insufficiency, storage complications, and time constraints. As a result, modern infant formula has become a substitute or alternative when breastfeeding is not possible or is inadequate for infants. In the optimal design of formula, many factors are considered in creating a product that is safe and provides a nutritional profile either equivalent to human milk or in optimal alignment with infant needs. Human milk composition serves as a reference for infant formula formulation. The purpose of this literature review is to provide a synopsis of recent advances in infant formula research in the past 5 years (2014– 2018). Specifically, the breadth of bioactive components of human milk that are not yet available in infant formulas and dose optimization requirements in full-term infants are reviewed. The nutritional value of infant formulas has been improved by adding functional bioactive ingredients such as choline, strains of probiotics, and prebiotic oligosaccharides. There are considerable variations in the composition of available infant formulas between manufacturers and across different countries. Formulas still generally lack important contents such as antibodies and exosomes that are present in human milk. Some of the adverse health outcomes associated with formula feeding as compared with human milk feeding are attributed to the different composition of the foods. This literature review summarizes the most recent research aimed at advancing infant formula composition to narrow the difference in health outcomes between human milk and formula-fed infants.
Keywords: nutrition, neonate, pediatrics, infant formula, ganglioside, miRNA
Introduction
The American Academy of Pediatrics (AAP) and the World Health Organization (WHO) recommend that infants are exclusively breastfed for 6 months after birth for optimal growth and development and that infants should continue to breastfeed up to 2 years along with complementary foods.1 In the case infants are not breastfed and banked or donor milk may not be available, infant formula is the most adequate food alternative.2,3 The health benefits of human milk consumption are greater than those of infant formula due to the nutritional and bioactive components in human milk that are absent in infant formula or not present in optimal concentration relative to the infant stage of growth and development.4 In this context, bioactives are compounds in diet that elicit physiologic effects in the consumer. Important bioactives for newborn health may include whole bacterial or mammalian cells, nanovesicles derived from cells, membrane fragments, or macromolecules. Despite the shortcomings of infant formula, it serves as the most viable substitution for human milk. Therefore, it benefits researchers, practitioners, and industry to develop a nutritionally adequate substitute for human milk which can ensure similar acute and chronic health outcomes compared to human milk feeding.
In consideration of adding novel bioactive agents to infant formulas, human milk is used as a model. In general, commercially available infant formula products are nutritionally interchangeable and there is minimal evidence to support that one infant formula is better than another.5 The composition of infant formula has great importance as it can have a significant impact on long-term health.6 As a result, when knowledge on optimizing infant nutrition progresses, industry is continuously adapting the composition of infant formula to align growth and development outcomes in formula-consuming infants with those seen in infants that consume human milk. These changes can include alternative sources of bioactive ingredients, consideration of dose optimization, or addition of novel bioactives.
Literature reviews to date have primarily focused on a specific bioactive in extensive detail or on particular health outcomes related to infant milk formula composition. The purpose of this literature review is to provide a comprehensive synopsis of recent advances in infant formula research pertaining to the inclusion, or dose optimization, of novel bioactives. Accordingly, a range of clinical health outcomes are reviewed in relation to the plethora of bioactives present, or being considered for inclusion, in infant formulas.
Methods
JSTOR, Embase, MedlinePlus, and Pubmed bibliographic databases of published scientific research were accessed for this literature review. The following search terms were used: infant nutrition, neonatal nutrition, baby formula, infant formula, human milk, human breast milk, breastfeeding, and [(human milk OR formula) AND (protein content OR lactalbumin OR lactoferrin OR probiotics OR prebiotics OR oligosaccharide OR essential fat OR polyunsaturated fat OR omega-3 OR omega-6 OR choline OR lutein OR ganglioside OR milk fat globule membrane OR microRNA]. Peer-reviewed scientific research selected for inclusion in this work was primarily human clinical intervention studies and were published between 2014 and 2018. Other literature published before 2014 was included in this work to provide scope, context, and background to the bioactive agent topic subsections.
Novel Bioactives
Protein Content
Bioactive proteins in human milk provide immune protection, assist in digestive processes, contribute towards gut development, and act as carriers for some nutrients.7 Human milk and infant formula may differ in protein composition and concentration. Human milk provides total protein needs and is rich in essential amino acids.8,9 Whereas commercially available infant formulas are typically made from bovine milk which has a much higher total protein concentration.10 Some infant formula products have soy as the primary protein source in order to address allergy to cow’s milk. This is less common, as in the United States about 69% of infants consume cow’s milk formula and only 12% consume soy formula.11
In order for infants to avoid drastically exceeding protein requirements, the quantity and quality of protein in infant formula should match closely to human milk. In infant formula and human milk, higher protein content increases the risk of infants being overweight and obese later in life.6,12 Rapid early growth during the first 2 years of life is associated with the likelihood of being overweight and obese during childhood.13 Recent formula products developed contain lower protein concentrations with improved protein quality.7 Recent clinical evaluations of infant formulas with lowered protein content show that low-protein content formula is safe and promotes adequate growth.14 Currently, formulas meet protein requirements of 1.8–3.0 g/100 kcal but there is no consensus on the optimal amount of protein in infant formulas.15 In fact, the European Food Safety Authority (EFSA) had proposed to lower the maximum protein content of infant formula to reduce progressive weight gain in formula-fed infants.16
Infant formula products generally contain adequate protein concentrations that provide sufficient quantities of all essential amino acids to meet the needs of developing infants.17 However, protein concentrations in human milk change over time as protein content is higher at the initial stages of lactation and decrease during the later stages of lactation. In earlier stages of lactation, human milk contains 1.4–1.6 g protein/100 mL of protein. The concentration of protein lowers to 0.8–1.0 g/100 mL after 3 months of lactation and to 0.7–0.8 g/100 mL after 6 months.1 These changes in human milk protein concentration over the course of lactation necessitate consideration for optimal age-specific products to provide adequate nutrition to infants at growth and development milestones before being weaned.
α-Lactalbumin
α-Lactalbumin is a whey protein found in human milk at a concentration of 28% of total protein and in bovine milk at 3% of total protein.18 α-Lactalbumin provides essential amino acids and the bovine form is added to infant formulas.10 Since the introduction of α-lactalbumin to infant formula, the proportion of α-lactalbumin has increased in product over time since its structural elucidation reveals it has a high proportion of tryptophan, lysine, and cysteine; and is available and easily obtained from an enriched whey protein fraction.19
α-Lactalbumin is bioavailable to the infant. It is digested and absorbed and the oligopeptides and amino acids have been shown to support critical immune functions such as phagocytosis by macrophages and growth of Bifidobacteria.20,21 In addition, the α-lactalbumin peptides are antimicrobial and have shown activity in the colon against colonization of Escherichia coli, K. pneumoniae, Staphylococcus aureus, Staphylococcus epidermis, Streptococci, and Candida albicans.22 Supplementation of α-lactalbumin in infant formula containing a protein concentration of 1 g protein/100 mL showed normal growth of infants up to 4 months of age.23 This study demonstrates that the addition of α-lactalbumin to infant formula supports a mechanism by which total protein concentration of formula can be lowered, the protein needs of the infant can still be met, and that growth outcomes are still supported. Importantly, it has been shown that α-lactalbumin does not affect iron absorption from formula in infants. Since iron status can vary greatly depending on weight and time of gestation in infants, research supports the addition of a standard dose of α-lactalbumin to formula with respect to iron requirement.24,25
Lactoferrin
Lactoferrin is a multifunctional whey protein found in human milk that has antibacterial and antiviral properties; it is involved in immune function, increased iron uptake, and inhibition of carcinogenesis.26 The concentration is variable in colostrum, but lactoferrin comprises 15–20% of mature human milk protein content.10,27 The addition of lactoferrin to formula from a bovine source is costly and can exhaust raw product resources from cow milk containing only a very low quantity of the protein. Additionally, it is difficult to preserve during infant formula production due to denaturing at high temperatures.27,28
Commercially added lactoferrin has similar bioactive properties as human lactoferrin. When lactoferrin was added to infant formula, it was found that the incidence of respiratory illnesses decreased while hematocrit increased.27,29 However, enriched lactoferrin did not improve iron absorption since bovine lactoferrin does not bind to human lactoferrin receptors.30 Lactoferrin is enriched in a small number of commercially available infant formulas and is recognized as a safe ingredient by the Food and Drug Administration.27,31 Even still, lactoferrin is highly regulated to ensure it is of high quality before its addition in infant formula.32 Furthermore, there are still gaps in knowledge regarding an optimal dosage of enriched lactoferrin in infant formula products.31
Probiotics
Probiotics are being added to infant formula and have been shown to have multiple beneficial effects (Table 1). Probiotics are microorganisms that can be present in human milk. The most common strains include Staphylococci, Streptococci, lactic acid bacteria, and Bifidobacteria.33 When consumed, probiotics may influence the gut microbiota composition of the infant which is colonized by over 1014 microorganisms.33 The intestinal microbiota aids in digestion, gut homeostasis, immune function, and in maintaining stool firmness.33 When probiotics are not consumed in human milk or formula, the rates of negative health outcomes such as necrotizing enterocolitis, infection, and underweight infants increase.33 During infant development, the gut profile of microbiota is not static and can be influenced by environmental factors like diet.33 The method of delivery, length of gestation, and antibiotic use can also change the microbial composition of the gut.33 Thus, there is interest in determining which strains could be consumed to counteract deleterious environmental influences on gut microbiome composition.
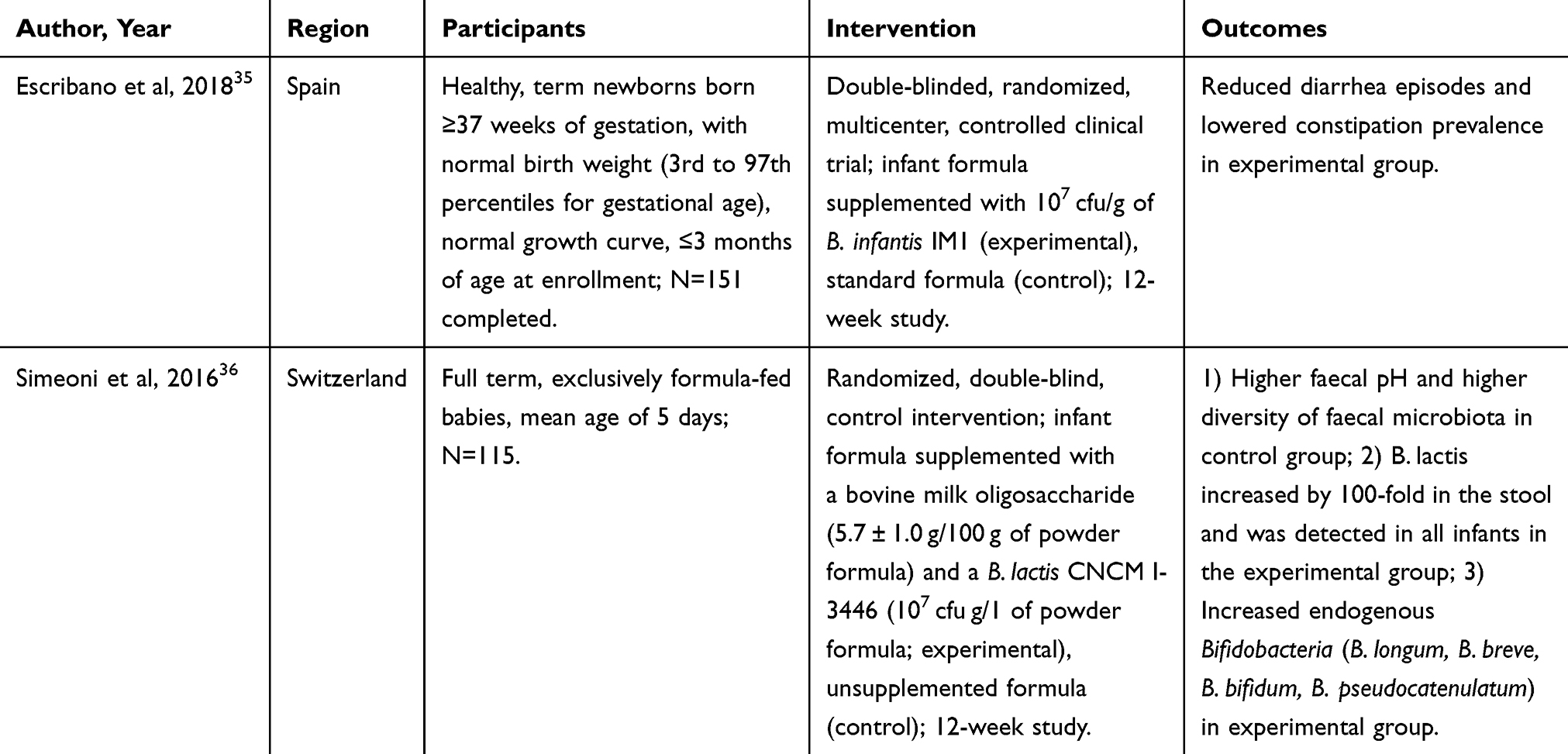 |
Table 1 Human Clinical Research on Health Outcomes Related to Inclusion of Probiotics in Infant Milk Formulae |
Some infant formula manufacturers have started incorporating probiotics into products. Efficacy varies greatly between strains of bacteria used; however, the use has been shown to be safe even if there is no observable effect on health outcomes.33–36 A study on infants less than 3 months old at enrollment who received Bifidobacterium infantis IM1 in a dose of 107 colony-forming units (cfu) per gram for 12 weeks had less incidence of diarrhea, reduction in Proteobacteria in stool samples, and higher stool frequency when compared to the control arm.35 Additionally, synbiotics, the use of both probiotics and prebiotic, have shown additive beneficial effects. Research on supplementing Bifidobacterium lactis and bovine milk-derived oligosaccharides in formula for infants averaging 5 days old showed a reduction in the number of infants with gastrointestinal infections and diarrhea.36 The evidence for inclusion of probiotics in infant formula is convincing, and future work should focus on which specific or combination of strains should be included, the dosage, and whether live cultures or lysed fragments are optimal for enhanced infant health outcomes.
Prebiotics
Prebiotics are non-digestible oligosaccharides that stimulate the growth and function of bacteria in the gastrointestinal tract.33,37 Prebiotics are found in high concentrations in human milk but are found only in trace amounts in cow’s milk. Thus, it is imperative for product manufacturers to fortify infant formulas with prebiotics above and beyond the amount intrinsic in bovine milk in order to elicit positive physiologic effects. Products are most often supplemented with galacto-oligosaccharide, fructo-oligosaccharide or polydextrose.37 These bioactive prebiotics have been shown to increase the growth of beneficial microorganisms like Bifidobacteria in the gut and resilience of the gut microbiome to metabolic stressors.37 The discrepancies in infection rates, stool pH, and stool consistency between infants consuming human milk and those fed formula are lessened when formula is supplemented with prebiotics.37 Apart from allergy, possible side effects identified include bloating, abdominal discomfort, and flatulence in doses of over 20 g/day in infants up to 28 days old.33
Human Milk Oligosaccharides
Human milk oligosaccharides (HMO) are complex, non-digestible carbohydrates that are found in human milk and are not present in bovine milk.38 HMOs have many potential benefits for developing infants, and the biological roles and functions of HMOs have been studied more extensively only in recent years.38 More than 200 HMOs have been discovered in human milk, where 2-fucosyllactose is the predominant oligosaccharide as it comprises 20–30% of HMOs.39 The functions, either additive or synergistic, of all HMOs have not yet been elucidated. In addition, the mechanism of action of most HMOs is unknown, but some may function as prebiotics and stimulate the growth of Bifidobacteria which colonize the infant gut and prevent pathogen adhesion to mucosal surfaces to reduce risks of infections.40 It has been shown that HMOs also support gastrointestinal and immune functions and that there are lower risks of viral infections in infants consuming HMOs from human milk.41,42
When evidence for the addition of HMOs to formula was scant, many producers added this bioactive to formulas simply because HMOs are found in human milk.43 Recently, research on HMO addition to infant formulas revealed evidence showing that prebiotic-enriched formula-fed infants display a profile of gastrointestinal microbiota that is similar to breastfed infants.43 HMO-supplemented infants do not have significant differences in length, weight, and head circumference in comparison to breastfed infants and non-prebiotic supplemented infants.44 Prebiotic-supplemented infants have increased frequencies of stool in comparison to human milk-fed infants and stool pH levels equal to human milk-fed infants.45
In a recent clinical study, it was found that infant formula that is enriched with 2ʹfucosyllactose and lacto-N-neotetraose is safe and supports adequate infant growth and development. In addition, it was found that there is lower incidence of bronchitis and medication use including antipyretics and antibiotics among infants that were fed HMO-supplemented formula.46 The addition of 2ʹ-fucosyllactose and lacto-N-neotetraose has contributed towards closing the compositional gap between human milk and infant formula. It is anticipated that more HMOs will be added to infant formula as it is safe for infants and promotes beneficial health outcomes in early life.47
Polyunsaturated Fatty Acids
Polyunsaturated fatty acids (PUFAs) have been added to infant formula products to function as bioactive ingredients (Table 2) that aid in eye and brain development. PUFAs have many biological effects on membrane function and composition, production of inflammatory mediators, and immune responses.4,48 A meta-analysis published in 2012 showed that PUFA supplementation in infant formulas did not have significant effects on improving early infant cognition.49 However, intervention studies have shown that PUFA supplementation has positive developmental outcomes that resemble trajectories seen in infants fed human milk with respect to cognitive function, visual acuity, and immune response.50,51
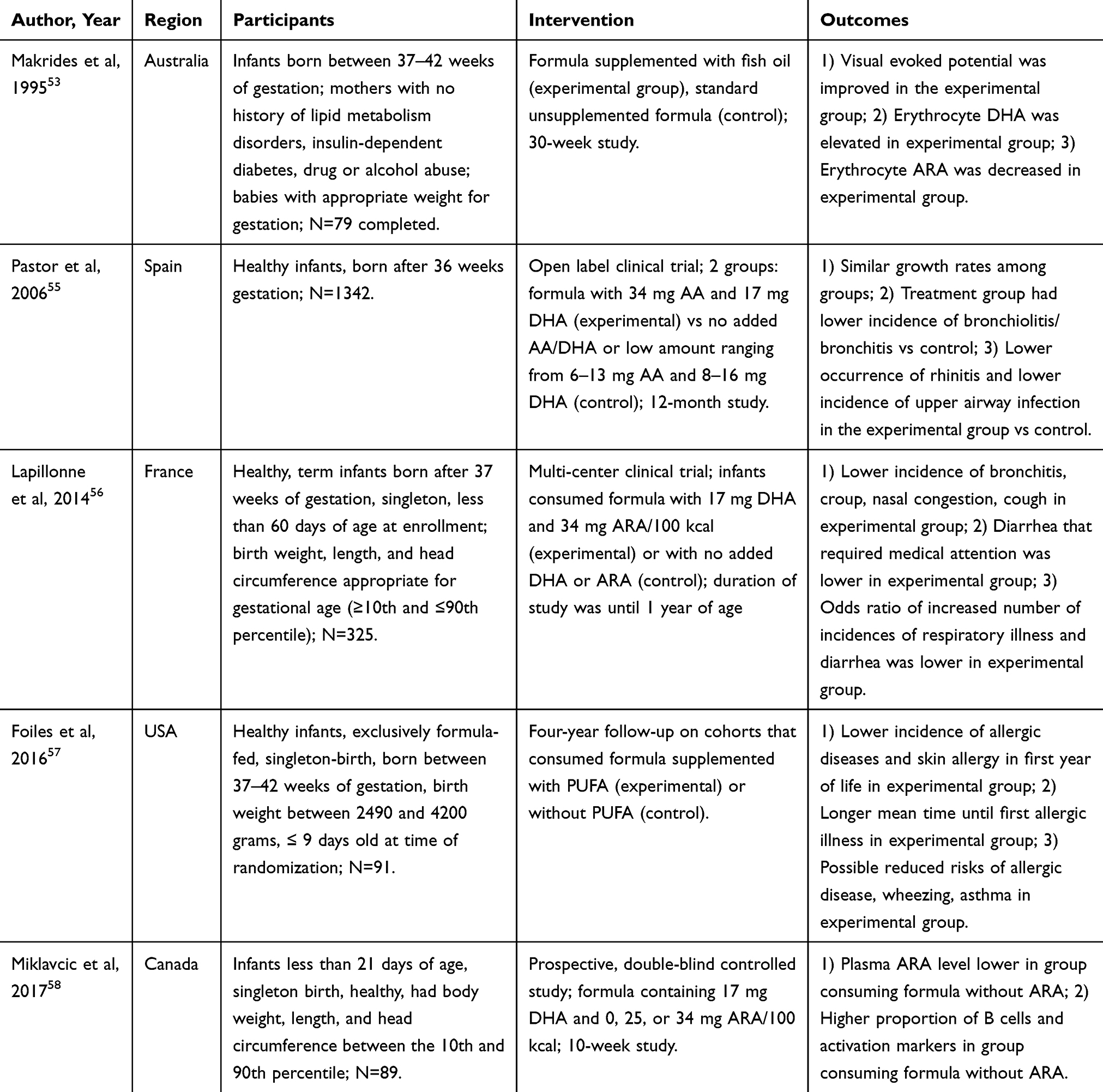 |
Table 2 Human Clinical Research on Health Outcomes Related to Inclusion of Polyunsaturated Fatty Acids in Infant Milk Formulae |
Docosahexaenoic acid (DHA) and arachidonic acid (AA) are PUFAs that are found in human milk and have important roles in the development of plasma membrane constituents.5 DHA and AA are added to formula instead of the respective n-6 and n-3 precursors since the elongation and desaturation processes are slow and inefficient resulting in a very low conversion rate. Infants that are fed formulas supplemented with PUFAs have higher levels of DHA and AA in plasma or red blood cells compared with infants that are breastfed or fed formulas that are not supplemented.53,54 Infants that are fed formulas that are fortified with DHA and AA have lower incidences of upper respiratory infections and allergies, bronchitis, nasal congestion, cough, diarrhea that requires medical attention, atopic dermatitis, eczema, and contact dermatitis.55–57
Although the concentration of n-6 and n-3 PUFAs in human milk varies highly depending on maternal diet,52 there is insufficient data that supports the addition of DHA to formula without AA as well.50 Infant formulas that include AA and DHA have beneficial effects on the immune system of developing infants. The absence of AA in infant formula affects populations of circulating T cells and macrophages and also causes an increase in the presence of B cells expressing CD20+ and CD54+ activation markers compared to infant formula with AA.58 The proportion of AA and DHA in supplemented infant formula varies. Further research is needed to determine the exact dose and ratio of PUFAs for optimal growth and development of humoral and cell-mediated immunity.50
Choline
Choline is an essential bioactive micronutrient that plays a role in cell signaling, structure of plasma membrane, muscle function, and lipid transport. Choline is found in human milk in a water-soluble form and is essential in fetal cognitive development. The presence of choline alters brain and spinal cord structure and function and can lower risks of neural tube defects.59
The European Society for Pediatric Gastroenterology, Hepatology and Nutrition recommends inclusion of no more than 50 mg/100 kcal of choline in infant formula products. Currently, there are no known cases of adverse effects of having high choline intakes. Inadequate choline levels can inhibit vitamin B12 and folic acid metabolism. Current research on choline supplementation in infants has been limited to preterm infants and more studies need to be performed to examine its properties in infant formula.
Lutein
Lutein is a carotenoid found in human milk and the concentration varies depending on maternal diet, with intake of fruits and vegetables increasing content.60 It functions as a structural component of the eye, an antioxidant, and as a filter for blue light. Lutein plays a biological and physiological role in the visual development and function in infants;61 and infants are reliant on breastfeeding to receive adequate quantities.62 Differences in serum lutein content between breastfed and formula-fed infants are seen as early as 1 month of age. Exclusively breastfed neonates had 6 times the average serum lutein level in comparison to infants consuming formula. Even in infants consuming formula fortified with lutein to a concentration similar to human milk, serum levels were drastically lower than in infants consuming human milk suggesting a difference in bioavailability.63
Recent studies demonstrated that lutein-supplemented infant formula is safe and supports the visual and cognitive development of infants.60–62,64 Presently, there is no research that lutein supplementation influences brain accretion during infancy. A recent study found that the concentration of lutein is higher in several regions of the brain during infancy than in adulthood which may indicate that lutein may play an important role in early neural development.60 Currently, there are a number of infant formulas that contain lutein. However, further research on lutein must be conducted to ensure efficacy for optimal growth and development outcomes in infants.61
Ganglioside
Gangliosides are sialylated glycosphingolipids commonly found in the lipid membranes of cells. They play a large role in neurological development, intracellular trafficking, immune cell signaling, gut integrity, and preventing infections.65,66 Gangliosides are present in human milk, with disialoganglioside GD3 and monosialodihexosylganglioside (GM3) being the primary molecular species.67 Total concentration of ganglioside is highest in early stages of lactation and is predominantly comprised of GD3. However, ganglioside GD3 content decreases over period of lactation and GM3 becomes more prominent.67 In infant milk formulas the content of gangliosides is provided from the bovine milk base, which has lower ganglioside content than human milk.67 Supplementation of infants with GD3 has shown beneficial effects on neurologic development. Infants aged 2-8 weeks showed increased Hand and Eye Coordination IQ, Performance IQ, and General IQ on the Griffiths scales when formula was supplemented with 2–3 mg of ganglioside/100 g of infant formula.68 There is a case to be made for the addition of gangliosides to formula; however, very few studies have been performed in human infants despite the observed safety, tolerability, and efficacy of this bioactive.
MicroRNA
MicroRNA (miRNA) are bioactive components found in human milk. They are small non-coding RNA molecules that regulate the expression of genes involved in the cell cycle, proliferation, differentiation, apoptosis, and the immune response. It was recently discovered that human milk contains various miRNAs that are bioavailable, circulate systemically, and are thought to be involved in infant development.4,69 In addition to human milk, bovine milk contains miRNA in an exosome vehicle for which bioavailability to human tissue has been demonstrated utilizing an in vitro CaCo-2 cell model, a human colorectal tumor cell line.70 There is emerging research that miRNAs that are present in human milk can enter phenotypically normal and tumor cells and affect biological functions.69 Some miRNAs found in bovine milk have similar nucleotide sequences to those from human milk but analogous effects have not been directly demonstrated.70 Currently, human and bovine milk miRNA are not detected in many infant formulas.69 This may be due to degradation of miRNA caused by homogenization and heating of milk.70 Future research in this area could consider whether bovine milk miRNAs are resistant to processing steps in the production of infant formulas, and whether human milk miRNA elicits physiologic effects different from those of bovine miRNA.
Conclusion
This literature review summarizes research on the recent advances in research related to the addition of novel bioactive compounds to infant formula (2014–2018). These advances consist of changing the source from which certain ingredients or bioactives are derived, altering the dose of bioactives, such as including specific protein in formula products, and adding novel bioactive components like HMO and miRNA. Health outcomes related to consumption of infant formula do not emulate outcomes in infants that consume human milk exclusively. However, adding bioactive ingredients such as gangliosides, PUFA, and probiotics to infant formulas achieves growth and developmental outcomes in infants that are more in alignment with outcomes observed in infants exclusively consuming human milk. The addition of novel functional bioactive agents that also promote health outcomes beyond growth, namely immunity, cognition, and temperament, is of paramount interest for future work. Currently, no uniform composition of bioactive compounds exists among commercially available infant formulas. The optimization of infant formula composition may be realized with more research in longitudinal cohort studies following acute and long-term health outcomes of children to ensure the efficaciousness of novel bioactives.
Acknowledgment
This research is funded by Chapman University and by the Chapman University Center for Undergraduate Excellence Scholarly/Creative Grant.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Martin CR, Ling P-R, Blackburn GL. Review of infant feeding: key features of breast milk and infant formula. Nutrients. 2016;8(5):279. doi:10.3390/nu8050279
2. WHO. Infant and young children feeding. 2003.
3. Pediatrics AAo. Safe and Healthy Beginnings: a Resource Toolkit for Hospitals and Physicians’ Offices. 2009.
4. Mosca F, Gianni ML. Human milk: composition and health benefits. Pediatr Med Chir. 2017;39(2):155. doi:10.4081/pmc.2017.155
5. O’Connor NR. Infant formula. Am Fam Physician. 2009;79(7):565–570.
6. Luque V, Closa-Monasterolo R, Escribano J, Ferre N. Early programming by protein intake: the effect of protein on adiposity development and the growth and functionality of vital organs. Nutr Metab Insights. 2015;8(Suppl 1):49–56. doi:10.4137/NMI.S29525
7. Haschke F, Haiden N, Thakkar SK. Nutritive and bioactive proteins in breastmilk. Ann Nutr Metab. 2016;69(Suppl 2):17–26. doi:10.1159/000452820
8. Lonnerdal B, Forsum E, Gebre-Medhin M, Hambraeus L. Breast milk composition in Ethiopian and Swedish mothers. II. Lactose, nitrogen, and protein contents. Am J Clin Nutr. 1976;29(10):1134–1141. doi:10.1093/ajcn/29.10.1134
9. Feng P, Gao M, Burgher A, Zhou TH, Pramuk K. A nine-country study of the protein content and amino acid composition of mature human milk. Food Nutr Res. 2016;60:31042. doi:10.3402/fnr.v60.31042
10. Lonnerdal B. Infant formula and infant nutrition: bioactive proteins of human milk and implications for composition of infant formulas. Am J Clin Nutr. 2014;99(3):712S–717S. doi:10.3945/ajcn.113.071993
11. Rossen LM, Simon AE, Herrick KA. Types of infant formulas consumed in the United States. Clin Pediatr (Phila). 2016;55(3):278–285. doi:10.1177/0009922815591881
12. Heinig MJ, Nommsen LA, Peerson JM, Lonnerdal B, Dewey KG. Intake and growth of breast-fed and formula-fed infants in relation to the timing of introduction of complementary foods: the DARLING study. Davis area research on lactation, infant nutrition and growth. Acta Paediatr. 1993;82(12):999–1006. doi:10.1111/j.1651-2227.1993.tb12798.x
13. Weng SF, Redsell SA, Nathan D, Swift JA, Yang M, Glazebrook C. Estimating overweight risk in childhood from predictors during infancy. Pediatrics. 2013;132(2):e414–e421. doi:10.1542/peds.2012-3858
14. Liotto N, Orsi A, Menis C, et al. Clinical evaluation of two different protein content formulas fed to full-term healthy infants: a randomized controlled trial. BMC Pediatr. 2018;18(1):59. doi:10.1186/s12887-018-1046-6
15. Patro-Golab B, Zalewski BM, Kouwenhoven SM, et al. Protein concentration in milk formula, growth, and later risk of obesity: a systematic review. J Nutr. 2016;146:551–564. doi:10.3945/jn.115.223651
16. ESFA. Scientific opinion on the essential composition of infant and follow-on formulae. ESFA J. 2014;12(7):3760.
17. Trabulsi J, Capeding R, Lebumfacil J, et al. Effect of an alpha-lactalbumin-enriched infant formula with lower protein on growth. Eur J Clin Nutr. 2011;65(2):167–174. doi:10.1038/ejcn.2010.236
18. Heine WE, Klein PD, Reeds PJ. The importance of alpha-lactalbumin in infant nutrition. J Nutr. 1991;121(3):277–283. doi:10.1093/jn/121.3.277
19. Davis AM, Harris BJ, Lien EL, Pramuk K, Trabulsi J. Alpha-lactalbumin-rich infant formula fed to healthy term infants in a multicenter study: plasma essential amino acids and gastrointestinal tolerance. Eur J Clin Nutr. 2008;62(11):1294–1301. doi:10.1038/sj.ejcn.1602848
20. Jaziri M, Migliore-Samour D, Casabianca-Pignede MR, Keddad K, Morgat JL, Jolles P. Specific binding sites on human phagocytic blood cells for Gly-Leu-Phe and Val-Glu-Pro-Ile-Pro-Tyr, immunostimulating peptides from human milk proteins. Biochim Biophys Acta. 1992;1160(3):251–261. doi:10.1016/0167-4838(92)90085-R
21. Kee HJ, Hong YH, Kim ER, et al. Effect of enzymatically hydrolyzed alpha-LA fractions with pepsin on growth promoting of Bifidobacterium longum ATCC 15707. Korean J Dairy Sci. 1998;(1998(20):61–68.
22. Pellegrini A, Thomas U, Bramaz N, Hunziker P, von Fellenberg R. Isolation and identification of three bactericidal domains in the bovine alpha-lactalbumin molecule. Biochim Biophys Acta. 1999;1426(3):439–448. doi:10.1016/S0304-4165(98)00165-2
23. Oropeza-Ceja LG, Rosado JL, Ronquillo D, et al. Lower protein intake supports normal growth of full-term infants fed formula: a randomized controlled trial. Nutrients. 2018;10:7. doi:10.3390/nu10070886
24. Szymlek-Gay EA, Lonnerdal B, Abrams SA, Kvistgaard AS, Domellof M, Hernell O. alpha-Lactalbumin and casein-glycomacropeptide do not affect iron absorption from formula in healthy term infants. J Nutr. 2012;142(7):1226–1231. doi:10.3945/jn.111.153890
25. Lonnerdal B. Development of iron homeostasis in infants and young children. Am J Clin Nutr. 2017;106(Suppl6):1575S–1580S. doi:10.3945/ajcn.117.155820
26. Lonnerdal B. Nutritional roles of lactoferrin. Curr Opin Clin Nutr Metab Care. 2009;12(3):293–297. doi:10.1097/MCO.0b013e328328d13e
27. O’Callaghan DM, O’Mahony JA, Ramanujam KS, Burgher AM. Dehydrated dairy products/infant formulae. In: Fuquay JW, editor. Encyclopedia of Dairy Sciences (Second Edition). Academic Press; 2011:135–145.
28. Wang B, Timilsena YP, Blanch E, Adhikari B. Lactoferrin: structure, function, denaturation and digestion. Crit Rev Food Sci Nutr. 2019;59(4):580–596. doi:10.1080/10408398.2017.1381583
29. Lonnerdal B, Jiang R, Du X. Bovine lactoferrin can be taken up by the human intestinal lactoferrin receptor and exert bioactivities. J Pediatr Gastroenterol Nutr. 2011;53(6):606–614. doi:10.1097/MPG.0b013e318230a419
30. Aly E, Ros G, Frontela C. Structure and functions of lactoferrin as ingredient in infant formulas. J Food Res. 2013;2(4):25–36. doi:10.5539/jfr.v2n4p25
31. Manzoni P. Clinical Benefits of Lactoferrin for Infants and Children. J Pediatr. 2016;173(Suppl):S43–S52. doi:10.1016/j.jpeds.2016.02.075
32. Wakabayashi H, Yamauchi K, Abe F. Quality control of commercial bovine lactoferrin. Biometals. 2018;31(3):313–319. doi:10.1007/s10534-018-0098-2
33. Bertelsen RJ, Jensen ET, Ringel-Kulka T. Use of probiotics and prebiotics in infant feeding. Best Pract Res Clin Gastroenterol. 2016;30(1):39–48. doi:10.1016/j.bpg.2016.01.001
34. Maldonado-Lobon JA, Gil-Campos M, Maldonado J, et al. Long-term safety of early consumption of Lactobacillus fermentum CECT5716: a 3-year follow-up of a randomized controlled trial. Pharmacol Res. 2015;95–96:12–19. doi:10.1016/j.phrs.2015.01.006
35. Escribano J, Ferre N, Gispert-Llaurado M, et al. Bifidobacterium longum subsp infantis CECT7210-supplemented formula reduces diarrhea in healthy infants: a randomized controlled trial. Pediatr Res. 2018;83(6):1120–1128. doi:10.1038/pr.2018.34
36. Simeoni U, Berger B, Junick J, et al. Gut microbiota analysis reveals a marked shift to bifidobacteria by a starter infant formula containing a synbiotic of bovine milk-derived oligosaccharides and Bifidobacterium animalis subsp. lactis CNCM I-3446. Environ Microbiol. 2016;18(7):2185–2195. doi:10.1111/1462-2920.13144
37. Vandenplas Y, De Greef E, Veereman G. Prebiotics in infant formula. Gut Microbes. 2014;5(6):681–687. doi:10.4161/19490976.2014.972237
38. Bode L, Jantscher-Krenn E. Structure-function relationships of human milk oligosaccharides. Adv Nutr. 2012;3(3):383S–391S. doi:10.3945/an.111.001404
39. Bode L. Human milk oligosaccharides: every baby needs a sugar mama. Glycobiology. 2012;22(9):1147–1162. doi:10.1093/glycob/cws074
40. Penders J, Thijs C, Vink C, et al. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics. 2006;118(2):511–521. doi:10.1542/peds.2005-2824
41. Lewis ZT, Totten SM, Smilowitz JT, et al. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome. 2015;3:13. doi:10.1186/s40168-015-0071-z
42. Bode L. The functional biology of human milk oligosaccharides. Early Hum Dev. 2015;91(11):619–622. doi:10.1016/j.earlhumdev.2015.09.001
43. Vandenplas Y, Zakharova I, Dmitrieva Y. Oligosaccharides in infant formula: more evidence to validate the role of prebiotics. Br J Nutr. 2015;113(9):1339–1344. doi:10.1017/S0007114515000823
44. Bozensky J, Hill M, Zelenka R, Skyba T. Prebiotics do not influence the severity of atopic dermatitis in infants: a randomised controlled trial. PLoS One. 2015;10(11):e0142897. doi:10.1371/journal.pone.0142897
45. Wopereis H, Sim K, Shaw A, Warner JO, Knol J, Kroll JS. Intestinal microbiota in infants at high risk for allergy: effects of prebiotics and role in eczema development. J Allergy Clin Immunol. 2018;141(4):1334–1342 e1335. doi:10.1016/j.jaci.2017.05.054
46. Puccio G, Alliet P, Cajozzo C, et al. Effects of infant formula with human milk oligosaccharides on growth and morbidity: a randomized multicenter trial. J Pediatr Gastroenterol Nutr. 2017;64(4):624–631. doi:10.1097/MPG.0000000000001520
47. Donovan SM, Comstock SS. Human milk oligosaccharides influence neonatal mucosal and systemic immunity. Ann Nutr Metab. 2016;69(Suppl 2):42–51. doi:10.1159/000452818
48. Miklavcic JJ, Badger TM, Bowlin AK, et al. Human breast-milk feeding enhances the humoral and cell-mediated immune response in neonatal piglets. J Nutr. 2018;148(11):1860–1870. doi:10.1093/jn/nxy170
49. Qawasmi A, Landeros-Weisenberger A, Leckman JF, Bloch MH. Meta-analysis of long-chain polyunsaturated fatty acid supplementation of formula and infant cognition. Pediatrics. 2012;129(6):1141–1149. doi:10.1542/peds.2011-2127
50. Lien EL, Richard C, Hoffman DR. DHA and ARA addition to infant formula: current status and future research directions. Prostaglandins Leukot Essent Fatty Acids. 2018;128:26–40. doi:10.1016/j.plefa.2017.09.005
51. Richard C, Lewis ED, Field CJ. Evidence for the essentiality of arachidonic and docosahexaenoic acid in the postnatal maternal and infant diet for the development of the infant’s immune system early in life. Appl Physiol Nutr Metab. 2016;41(5):461–475. doi:10.1139/apnm-2015-0660
52. Carlson SE. Docosahexaenoic acid and arachidonic acid nutrition in early development. Adv Pediatr. 2016;63(1):453–471. doi:10.1016/j.yapd.2016.04.011
53. Makrides M, Neumann M, Simmer K, Pater J, Gibson R. Are long-chain polyunsaturated fatty acids essential nutrients in infancy? Lancet. 1995;345(8963):1463–1468. doi:10.1016/S0140-6736(95)91035-2
54. Innis SM, Lupton BA, Nelson CM. Biochemical and functional approaches to study of fatty acid requirements for very premature infants. Nutrition. 1994;10(1):72–76.
55. Pastor N, Soler B, Mitmesser SH, Ferguson P, Lifschitz C. Infants fed docosahexaenoic acid- and arachidonic acid-supplemented formula have decreased incidence of bronchiolitis/bronchitis the first year of life. Clin Pediatr (Phila). 2006;45(9):850–855. doi:10.1177/1073858406289801
56. Lapillonne A, Pastor N, Zhuang W, Scalabrin DM. Infants fed formula with added long chain polyunsaturated fatty acids have reduced incidence of respiratory illnesses and diarrhea during the first year of life. BMC Pediatr. 2014;14:168. doi:10.1186/1471-2431-14-168
57. Foiles AM, Kerling EH, Wick JA, Scalabrin DM, Colombo J, Carlson SE. Formula with long-chain polyunsaturated fatty acids reduces incidence of allergy in early childhood. Pediatr Allergy Immunol. 2016;27(2):156–161. doi:10.1111/pai.12515
58. Miklavcic JJ, Larsen BM, Mazurak VC, et al. Reduction of arachidonate is associated with increase in b-cell activation marker in infants: a randomized trial. J Pediatr Gastroenterol Nutr. 2017;64(3):446–453. doi:10.1097/MPG.0000000000001283
59. Zeisel SH. Choline: critical role during fetal development and dietary requirements in adults. Annu Rev Nutr. 2006;26:229–250. doi:10.1146/annurev.nutr.26.061505.111156
60. Zielinska MA, Wesolowska A, Pawlus B, Hamulka J. Health effects of carotenoids during pregnancy and lactation. Nutrients. 2017;9(8):838. doi:10.3390/nu9080838
61. Kon IY, Gmoshinskaya MV, Safronova AI, Alarcon P, Growth VY. Tolerance assessment of a lutein-fortified infant formula. Pediatr Gastroenterol Hepatol Nutr. 2014;17(2):104–111. doi:10.5223/pghn.2014.17.2.104
62. Capeding R, Gepanayao CP, Calimon N, et al. Lutein-fortified infant formula fed to healthy term infants: evaluation of growth effects and safety. Nutr J. 2010;9:22. doi:10.1186/1475-2891-9-22
63. Bettler J, Zimmer JP, Neuringer M, DeRusso PA. Serum lutein concentrations in healthy term infants fed human milk or infant formula with lutein. Eur J Nutr. 2010;49(1):45–51. doi:10.1007/s00394-009-0047-5
64. Mackey AD, Albrecht D, Oliver J, Williams T, Long AC, Price P. Tplasma carotenoid concentrations of infants are increased by feeding a milk-based infant formula supplemented with carotenoids. J Sci Food Agric. 2013;93(8):1945–1952. doi:10.1002/jsfa.2013.93.issue-8
65. Ryan JM, Rice GE, Mitchell MD. The role of gangliosides in brain development and the potential benefits of perinatal supplementation. Nutr Res. 2013;33(11):877–887. doi:10.1016/j.nutres.2013.07.021
66. Miklavcic JJ, Schnabl KL, Mazurak VC, Thomson AB, Clandinin MT. Dietary ganglioside reduces proinflammatory signaling in the intestine. J Nutr Metab. 2012;2012:280286. doi:10.1155/2012/280286
67. Palmano K, Rowan A, Guillermo R, Guan J, McJarrow P. The role of gangliosides in neurodevelopment. Nutrients. 2015;7(5):3891–3913. doi:10.3390/nu7053891
68. Zheng L, Fleith M, Giuffrida F, O’Neill BV, Schneider N. Dietary polar lipids and cognitive development: a narrative review. Adv Nutr. 2019;10:1163–1176. doi:10.1093/advances/nmz051
69. Golan-Gerstl R, Elbaum Shiff Y, Moshayoff V, Schecter D, Leshkowitz D, Reif S. Characterization and biological function of milk-derived miRNAs. Mol Nutr Food Res. 2017;61:10. doi:10.1002/mnfr.v61.10
70. Zempleni J, Aguilar-Lozano A, Sadri M, et al. Biological activities of extracellular vesicles and their cargos from bovine and human milk in humans and implications for infants. J Nutr. 2017;147(1):3–10. doi:10.3945/jn.116.238949
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.