Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 15
Inhibition of miR-155 Attenuates CD14+ Monocyte-Mediated Inflammatory Response and Oxidative Stress in Psoriasis Through TLR4/MyD88/NF-κB Signaling Pathway
Authors Li J, Liu Y, Cao Y, Wang J, Zhao X, Jiao J, Li J, Zhang K ![]() , Yin G
, Yin G
Received 30 November 2021
Accepted for publication 27 January 2022
Published 9 February 2022 Volume 2022:15 Pages 193—201
DOI https://doi.org/10.2147/CCID.S350711
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Jeffrey Weinberg
Jiajie Li, Yanmin Liu, Yue Cao, Juanjuan Wang, Xingcheng Zhao, Juanjuan Jiao, Junqin Li, Kaiming Zhang, Guohua Yin
Shanxi Key Laboratory of Stem Cell for Immunological Dermatosis, Taiyuan Central Hospital of Shanxi Medical University, Taiyuan, 030009, Shanxi Province, People’s Republic of China
Correspondence: Guohua Yin, Tel +86 0351-5656079, Email [email protected]
Purpose: Previous studies showed the link of CD14+ monocytes to inflammation and oxidation in psoriasis. In the present study, we investigated the regulatory role of miR-155 in CD14+ monocyte function in psoriasis.
Materials and Methods: CD14+ monocytes were isolated from peripheral blood by magnetic bead separation method and its function was assessed following silence of miR-155 by lentivirus transfection with or without inhibition of TLR4 pathway. CCK8 and EdU were used to assess the proliferation of CD14+ monocytes. Expression levels of SOCS1, TLR4 and MyD88 proteins were determined by Western blotting, while expression levels of IL-6, TNF-α, ROS, MDA and T-AOC were measured by ELISA kit. The expression levels of mRNA for miR-155, NF-κB and its subunit NF-κB-p65 were assessed by q-PCR.
Results: The results showed that compared with normal control CD14+ monocytes, the expression levels of miR-155, NF-κB and NF-κB-p65, TLR4, MyD88 and IL-6, TNF-α were increased, while expression levels of SOCS1 were decreased in CD14+ monocytes from psoriatic patients. Enhanced cell proliferation and oxidation were also observed in CD14+ monocytes from psoriatic patients. Inhibition of miR-155 partially corrected the abnormalities of cell proliferation and expression levels of biomarkers mentioned above in CD14+ monocytes from psoriatic patients. Inhibitions of both TLR4 pathway and miR-155 further corrected abnormalities of proliferation and the above biomarkers in CD14+ monocytes from psoriatic patients.
Conclusion: These results suggest that increased expression levels of miR-155 contribute to CD14+ monocyte-mediated inflammation and oxidation in psoriasis via TLR4 pathway.
Keywords: psoriasis CD14+ monocytes, miR-155, TLR4 pathway, inflammatory, oxidation
Introduction
Psoriasis is a chronic immune inflammatory skin disease characterized by infiltration of inflammatory cells such as monocytes and neutrophils, and epidermal hyperproliferation.1 Monocytes are heterogeneous and have multiple subpopulations. CD14+ monocytes belong to the classic subpopulations and participate in the regulation of diseases.2 The mechanisms of CD14+ monocyte-mediated diseases are complex. Psoriatic patients exhibit an increased number of monocytes in the peripheral blood. Monocytes can be recruited from the blood to the inflamed skin site of psoriasis through CCR6, leading to amplification of the inflammatory response.3 Monocytes can also migrate from the inflamed skin site to the peripheral blood and express higher levels of TLRs.4 In psoriasis, CD14+ monocytes can release high levels of TNF-α, IL-6 and IL-10, the increase of TNF-α and IL-6 is positively correlated with the severity of psoriasis, and the increase of IL-10 plays a positive role in the treatment of psoriasis.5,6
TLRs, which belong to type I transmembrane proteins, are a group of evolutionarily highly conserved pattern recognition receptors that can be activated by a variety of pathology-associated molecules and participate in the occurrence and development of diseases.7 TLR4, a member of the TLR family, is rich in extracellular leucine sequences, which can be activated by bacterial lipopolysaccharide (LPS) to initiate a cascade of protein reactions, inducing the production of pro-inflammatory cytokines, consequently leading to inflammatory and immune responses. LPS binds to the LPS-binding protein (LBP) and is transferred to the CD14 cluster, then recognized and bound by the myeloid differentiation factor-2 (MD-2) on TLR4, forming the LPS/CD14/MD-2/TLR4 complex, which activates intracellular signaling pathways, TLR4/MyD88/NF-κB and TLR4/TRIF/IRF3.8 There are five subtypes of NF-κB in humans, and P65 is an important subunit of NF-κB, which is activated in inflammation.9 In TLR4/MyD88/NF-κB signaling pathway, the downstream of NF-κB is activated through a series of chemical reactions, promoting the release of inflammatory cytokines such as IL-1β, TNF-α and IL-6.10 Over-activation of NF-κB signaling pathway can disturb the oxidative balance, leading to the accumulation of peroxidation products such as reactive oxygen species (ROS), and the decrease of antioxidant T-AOC, resulting in oxidative stress and further aggravation of the inflammatory response.11
miRNA-155 (miR-155), a small RNA, is highly expressed in many diseases mediated by CD14+ monocytes such as the increase of miR-155 in peripheral circulating CD14+ monocytes in patients with multiple sclerosis (MS) and HCV infection, which stimulates the production of inflammatory cytokines and exacerbates the inflammatory response.12,13 It is reported that up-regulated miR-155 in psoriasis can lead to apoptosis of HaCaT cells and impaired function of dermal mesenchymal stem cells in psoriasis, which is an important target for the treatment of psoriasis.14,15 However, the study of miR-155 in psoriatic CD14+ monocytes has not been reported.
miR-155 is closely related to the TLR family. Stimulation of TLR4 and TLR8 ligands, as well as TLR9, can increase the production of miR-155 through different ways, thereby aggravating inflammation and disease,16,17 and the interference of TLR7 signal can inhibit the generation of miR-155.18 Meanwhile, as an early regulator of inflammatory response, miR-155 binds to the 3ʹUTR of cytokine signal suppressor 1 (SOCS1), a negative regulator of TLR signaling, to inhibit SOCS1 translation and regulate disease progression. For example, increased TLR8 gene expression,19 interference with intracellular signaling pathways downstream of TLR-7 and TLR-9, and the synthesis of interferon (IFN) type I synthesis in plasmacytoid dendritic cells.20 The relationship between miR-155 and TLR4 signal is an important research direction, but the related research is very rare in psoriatic CD14+ monocytes. Therefore, in this study, we focused on the regulatory role of miR-155 and TLR4 signaling pathways in psoriatic CD14+ monocytes.
Materials and Methods
Isolation and Culture of CD14+ Monocytes
Blood samples were collected from 10 normal controls and 20 psoriatic patients at the Department of Dermatology of Taiyuan Central Hospital after obtaining informed consent from the participants. Blood sample was mixed with PBS solution at 1:1 ratio and the mixture was then carefully added over the lymphocyte separation solution at 2:1 ratio (mixture: lymphocyte separation solution), followed by centrifugation. The white membrane in the middle was carefully extracted. After determination of the number of cells, 20ul CD14+ magnetic beads (CD14+ magnetic beads, Miltenyi Biotec, Germany) were added to every 107 cells, and placed at 4°C for 10min. CD14+ mononuclear cells were obtained by passing the cell fluid through the magnetic bead separation column. Afterwards, CD14+ mononuclear cells were transferred to six-well plate, and cultured in 1640 complete medium.
Flow Cytometry
The isolated cells were transferred into EP tubes to make 100mL PBS cell suspension, with a cell count of about 105. After addition of 20µl CD14+-FITC antibody (CD14+-FITC antibody, Miltenyi Biotec, Germany) to each tube, cells were incubated at 4°C for 10 minutes in dark. The samples without antibody were used as negative control. The purity of CD14+ monocytes was detected by flow cytometer. Cells with a purity of >85% were used in subsequent experiments.
Treatment of CD14+ Monocytes
The experiment included six groups, ie, normal CD14+ monocytes from normal subjects (N-CD14+), CD14+ monocytes from psoriatic patients (P-CD14+), P-CD14+ treated without miR-155 (P-CD14++miR-155 empty), P-CD14+ treated miR-155 lentivirus (human-miR-155-5p-inhibitor, Shanghai, China) (P-CD14++miR-155 inhibitor), P-CD14+ treated TLR4 inhibitor TAK242 (TAK242, Shanghai, China) (P-CD14++TAK242) and P-CD14+ treated miR-155 lentivirus and TLR4 inhibitor TAK242 (P-CD14++miR-155 inhibitor+TAK242). 293T cells were transfected with lentivirus to obtain empty and inhibited venom, and then transfected with CD14+ monocytes. After successful transfection, the above miR-155 empty group and inhibitor group as well as TAK242 group were obtained. Cells were cultured in 6-well plates with about 106 cells per well at 37 °C with supplement of 10% O2 and 5% CO2.
Cell Proliferation
Cell Count Kit 8 (CCK8, Boster, Wuhan, China) was used to measure cell count according to the instructions. Cells were seeded into 96-well plate at a density of 1×104 with 100µl medium per well. After addition of 10µl CCK8 solution to each well, cells were incubated at 37°C for 2 hours. The optical density was measured at 450 nm with an enzyme reader (n=5), and the total number of cells was calculated. EdU kit (EdU, Ribbio, Guangzhou, China) was used to assess cell proliferation.
Western Blotting
Total protein was extracted from cells by RIPA buffer (Beyotime, Shanghai, China) and measured using BCA kit (Solarbio, Beijing, China). Protein samples were isolated by 10% SDS-PAGE and transferred to PVDF membrane, blocked with 5% skim milk for 30 min. Main antibodies (Anti-TLR4, Rabbit, Novus, Shanghai, China; anti-MyD88, Rabbit, Abcam, Cambridge, UK; anti-SOCS1, Rabbit, CST, Boston, USA; Beta-actin Rabbit, Abcam, Cambridge, UK) were at dilutions of 1:1000. Image density analysis was performed using ImageJ software.
Quantitative RT-PCR (qRT-PCR)
The total RNA was extracted by Trizol method, and the concentration was determined. Reverse transcription was performed using a kit (miScript II RT kit, Germany). Following the instruction of the kit (miScript® SYBY® Green PCR Kit, Germany), the total cDNA was quantitatively detected by real-time fluorescence quantitative PCR, setting the annealing condition to 72 degrees for 1 minute. The relative quantity of target mRNA was determined using the comparative threshold (Ct) method by normalizing target mRNA Ct values. Used primers are as follows: β-actin: Forward: 5’-GCA CCA CAC CTT CTA CAA TGA-3’, Reverse: 5’-GTC ATC TTC TCG CGG TTG GC-3’; NF-κB: Forward: 5’-AGT TGA GGG GAC TTT CCC AGG C-3’, Reverse: 5’-GCC TGG GAA AGT CCC CTC AAC T-3’; NF-κB-p65: Forward: 5’-CGC GGA TCC TGG CTC GTC TGT AGT GCA CG-3’, Reverse: 5’-CCC AAG CTT TAG AAG CCA TCC CGG CAG TC-3’, miR-155-5p: UUA AUG CUA AUU GUG AUA GGG GU.
ELISA Assay
The respective ELISA kits (Xitang, Shanghai, China) were used to measure the content of TNF-α and IL-6, according to the manufacturers’ instruction. Briefly, 100µl of the culture supernatant was added to a 96-well plate, and then various reagents were added successively according to the instructions of the kit. The optical density (OD) value was measured at 450nm. Oxide index test samples were processed according to kit instructions (ROS, MDA, T-AOC, Jiancheng, Nanjing, China), and all detection reagents were added according to the manufacturer’s instruction. Following sequential cell digestion, PBS cleaning and resuspension, ROS was detected under a fluorescence microscope with a wavelength of 488nm. For detection of MDA, 250µl of culture supernatant was loaded into a 96-well plate, and OD values were obtained at 532 nm wavelength with enzyme reader. T-AOC was detected at the wavelength of 405nm.
Statistical Analysis
Each experiment was repeated three times independently. The GraphPad Prism 8 software (San Diego, CA, USA) was used to analyze the data. The data were expressed as mean ± standard error of mean. One-way ANOVA analysis was used to determine the significances. Differences among groups were compared by t-test. All the comparison with P<0.05 considered statistically significant.
Results
The Purity of Isolated CD14+ Monocytes is ≥85% by Flow Cytometry and the Expression Levels of miR-155 are Higher While SOCS1 Protein Levels are Lower in Psoriatic CD14+ Monocytes
We first determined the purity of isolated CD14+ monocytes. The results of flow cytometry showed that isolation of CD14+ monocytes with magnetic beads yielded the purity of CD14+ monocytes ≥85% (Figure 1A), indicating we successfully isolated CD14+ monocytes for subsequent experiments.
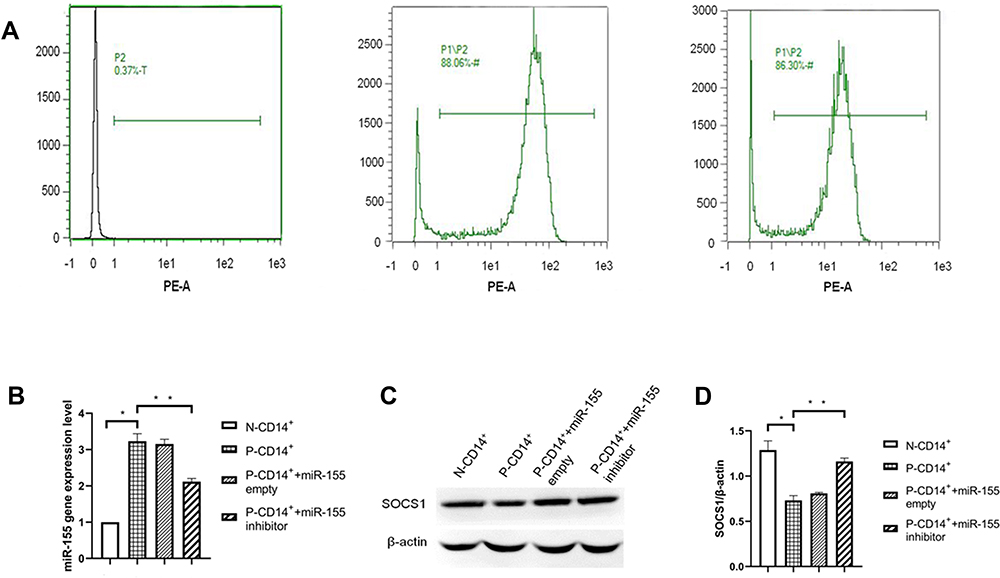 |
Figure 1 Purity of CD14+ monocytes measured by flow cytometer, q-PCR and Western blot for miR-155 and SOCS1 protein. (A) Purity of CD14+ monocytes. (B) Histogram of miR-155 gene expression. (C) Western blot of SOCS1 protein. (D) Histogram of SOCS1 expression. *P<0.05, **p<0.01, n=3. |
We first determined whether miR-155 regulates SOCS1 expression in P-CD14+ monocytes. As expected, expression levels of miR-155 were significantly higher in P-CD14+ monocytes than in N-CD14+ monocytes (Figure 1B. p<0.05). In contrast, expression levels of SOCS1 in P-CD14+ monocytes were lower than that in N-CD14+ monocytes (Figure 1C and D. p<0.05). Inhibition of miR-155 only increased expression levels of SOCS1 (Figure 1C and D. p<0.01), without changing the expression levels of miR-155 in P-CD14+ monocytes (Figure 1B). These results indicate that miR-155 is highly expressed in P-CD14+ monocytes and inhibits SOCS1 expression.
Silencing miR-155 Decreases the Proliferation of Peripheral Blood CD14+ Monocytes in Patients with Psoriasis
Keratinocytes are the main proliferating cells in psoriasis, but the proliferation of inflammatory monocytes also plays an important role in the development of psoriasis. In order to determine whether miR-155 regulates CD14+ monocyte proliferation and the possible signaling pathways, CCK8 and EdU assays were used to assess cell proliferation following inhibition of miR-155 and TLR4. As shown in Figure 2, inhibition of miR-155 and TLR4 respectively decreased proliferation of P-CD14+ monocytes (Figure 2B and C. p<0.01 vs P-CD14+ + miR-155/TAK24 alone). A further decrease in cell proliferation was observed in P-CD14+ monocytes following inhibitions of both miR-155 and TLR4 (Figure 2B and C. p<0.01 vs P-CD14+ + miR-155 or TLR4 inhibitor alone). These results demonstrate that miR-155 and TLR4 synergically enhance CD14+ monocyte proliferation.
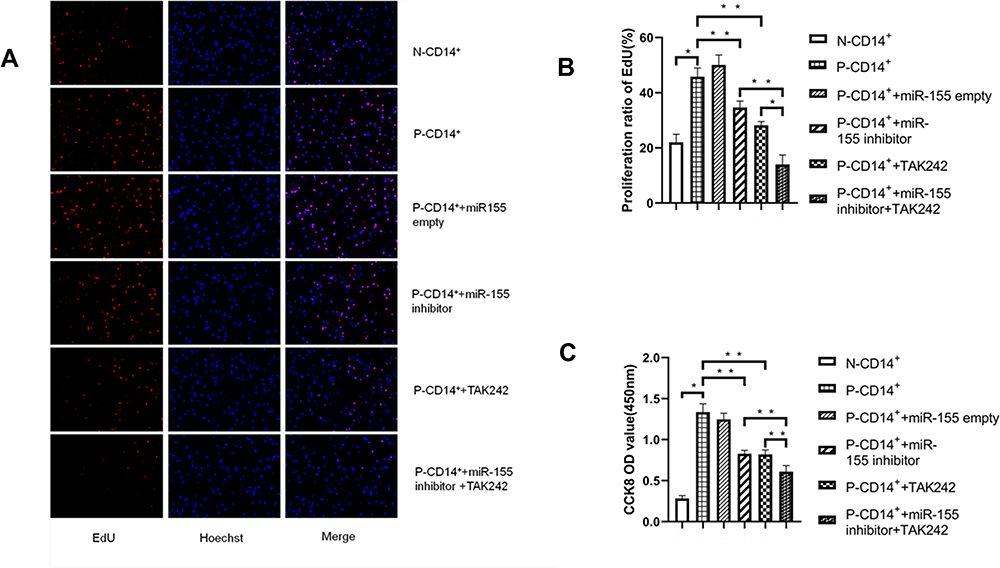 |
Figure 2 miR-155 and TLR4 regulate CD14+ monocyte proliferation. (A) EdU fluorescence staining (×20). (B) P-CD14+ proliferation assessed by EdU assay. (C) P-CD14+ proliferation assessed by CCK8 assay. *P< 0.05, **p<0.01, n=3. |
Inhibition of miR-155 Lowers the Expression Levels of TLR4 and Its Downstream Signaling Molecules
To explore the potential pathway of miR-155 in regulation of CD14+ monocyte function, we measured the expression levels of TLR4 and MyD88 protein, the downstream molecules of miR-155, in N-CD14+ and P-CD14+ monocytes. As seen in Figure 3A and B, expression levels of both TLR4 and MyD88 were significantly higher in P-CD14+ than in N-CD14+ monocytes (p<0.05). Inhibition of miR-155 and TLR4 respectively lowered expression levels of both TLR4 and MyD88 (Figure 3A and B). Again, inhibition of both TLR4 and miR-155 induced a further reduction in TLR4 and MyD88 expression (Figure 3A and B. p<0.05 vs P-CD14+ + miR-155 inhibitor alone).
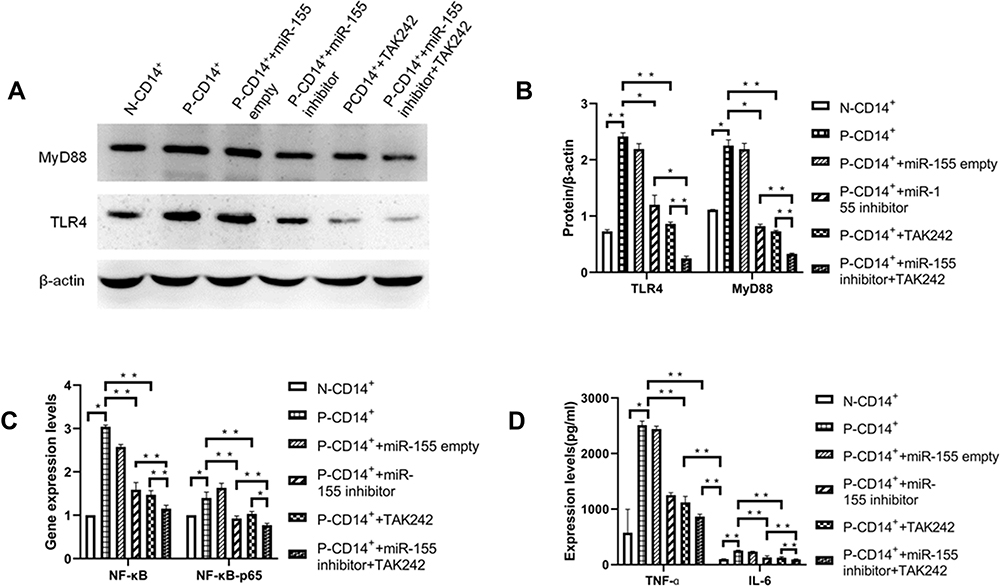 |
Figure 3 Expression of TLR4 and its downstream signaling. (A) Western blot of TLR4 and MyD88 protein. (B) Expressions of TLR4 and MyD88. (C) Expression of NF-κB and NF-κB-p65 mRNA. (D) Expression of TNF-α and IL-6. *P< 0.05, **p<0.01, n=3. |
Because miR-155 can activate its downstream signaling such as TLR4, NF-κB and NF-κB-p65, resulting in release of inflammatory cytokines, including TNF-α and IL-6,21 next we assessed expression levels of these signaling molecules in P-CD14+ monocytes with or without inhibition of miR-155. As shown in Figure 3C and D, expression levels of NF-κB, NF-κB-p65, TNF-α and IL-6 in P-CD14+ monocytes were higher than that in N-CD14+ monocytes, while inhibition of miR-155 lowered expression levels of these biomarkers. Inhibition of both miR-155 and TLR4 induced further reductions in their expression levels. These results suggest that regulation of miR-155 in the inflammation is mediated, in part, by TLR4 signaling pathway.
Inhibition of miR-155 Decreases the Levels of ROS and MDA, and Increases the Level of T-AOC
Oxidative stress and inflammatory response can interact with each other through TLR4 signaling pathway, resulting in the progression of disease.22 Because miR-155 regulates TLR4 expression (Figure 3A and B), we assessed next whether miR-155 also affect oxidative stress in CD14+ monocytes. The results showed that either ROS or MDA levels were markedly elevated in P-CD14+ monocytes vs N-CD14+ monocytes (Figure 4A and B). In contrast, the total antioxidant capacity (T-AOC) was lower in P-CD14+ monocytes vs N-CD14+ monocytes (Figure 4C, p<0.05). Inhibition of miR-155 and TLR4, respectively, partially attenuated the changes in levels of ROS, MDA, and T-AOC. However, simultaneous inhibition of miR-155 and TLR4 induced further reductions in ROS and MDA (Figure 4A and B, p<0.01 vs P-CD14+ + miR-155 inhibitor/TAK242, alone), and an increase in T-AOC (Figure 4C, p<0.01 vs P-CD14+ + miR-155 inhibitor/TAK242, alone). These results demonstrate that miR-155 enhances oxidative stress, in part mediated by TLR4 signaling pathway.
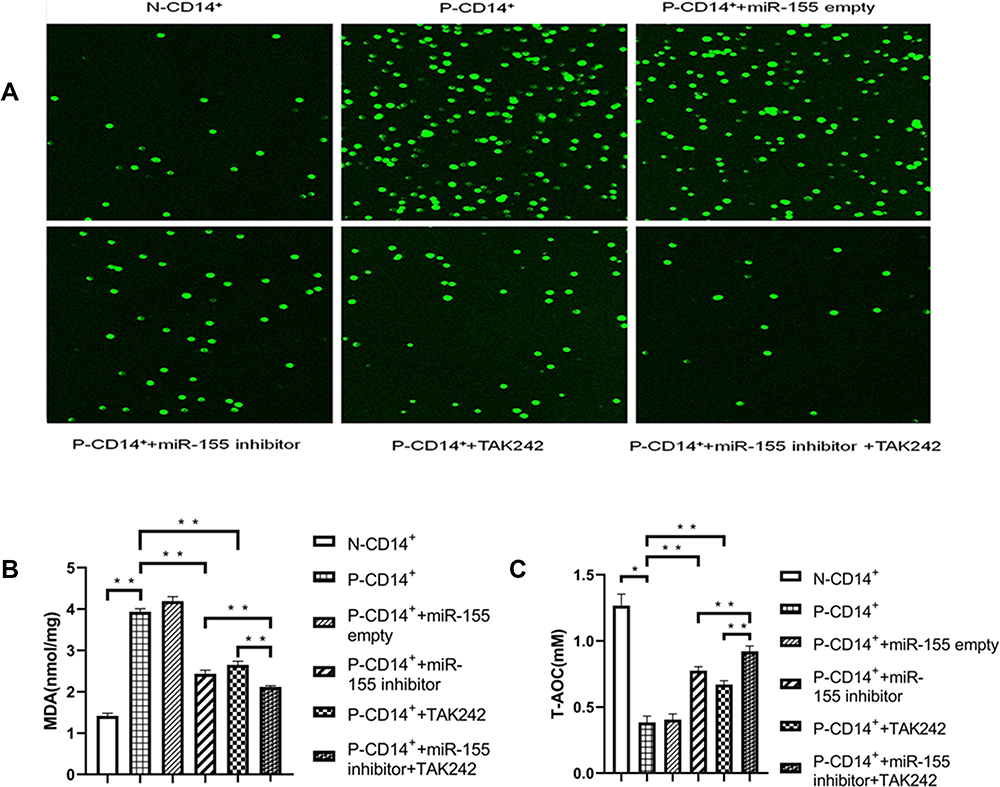 |
Figure 4 Inhibition of miR-155 decreases oxidative stress. (A) Fluorescence of ROS (×10). (B) MDA levels. (C) T-AOC levels. *P< 0.05, **p<0.01, n=3. |
Discussion
Psoriasis is an autoimmune skin disease with complex pathophysiological mechanisms, involving a variety of cell types and molecular mechanisms. Monocytes play a key role in the process of disease through proliferation, differentiation and release of a large number of pro-inflammatory factors.1 However, the underlying mechanisms by which monocytes are involved in the pathogenesis of psoriasis are not very clear. In the present study, we showed a crucial role of miR-155 in regulation of CD14+ monocyte functions, including proliferation, expression levels of TLR4 and its downstream signaling molecules, secretion of proinflammatory cytokine and antioxidant capacity.
Previous studies have shown that the percentage of circulating inflammatory CD14+ monocytes in psoriatic patients is higher than that in healthy controls, and the proliferation activity of CD14+ monocytes is significantly enhanced.23,24 Because inhibition of miR-155 decreased CD14+ monocyte proliferation (Figure 3), the increased CD14+ monocytes are likely attributable to the elevation in miR-155 expression. Consistent with our findings, upregulation of miR-155 in psoriatic keratinocytes leads to excessive proliferation and abnormal differentiation,25 while knockdown of elevated miR-155 in psoriatic tissues significantly reduces cell proliferation.26 Therefore, we believe that the proliferation of psoriatic CD14+ monocytes can be regulated by miR-155. The effect of miR-155 on CD14+ monocyte function is mediated, at least in part, by TLR4 signaling pathway because a) inhibition of miR-155 and TLR4 respectively decreases CD14+ monocyte proliferation and lowers expression levels of TLR4 and its downstream signaling molecules such as NF-κB, TNF-α and IL-6, and b) simultaneous inhibition of miR-155 and TLR4 induces more profound reductions in CD14+ monocyte proliferation and expression levels of NF-κB, TNF-α and IL-6. Similar results have been reported in other studies.27 Our experiments also showed that when TLR4 was inhibited alone without inhibition of miR-155, the changes of the above indicators are similar to those when miR-155 was inhibited alone without inhibition of TLR4, indicating that there are other pathways besides TLR4 for the effect of miR-155 on inflammation and oxidation of psoriatic CD14+ monocytes. We will study more complex mechanisms in future experiments. Moreover, the present study also showed an increased level of TLR4 and MyD88, which both are positively regulated by miR-155, in peripheral blood CD14+ monocytes of psoriatic patients. TLR4 and MyD88 proteins are key proteins in the MyD88-dependent pathway of TLR4 signaling and interaction of TLR4 and MyD88 can further activate TLR4 downstream signaling cascades, such as NF- κB, leading to enhanced cell proliferation, immune response and release of proinflammatory cytokines.28,29 Inhibition of miR-155 lowers expression levels of multiple downstream signaling molecules in TLR4/MyD88/NF-κB pathway, including TNF-α and IL-6, key pro-inflammatory cytokines in psoriasis.30 This is consistent with our results. However, in addition to TLR4, miR-155 also regulates NF-κB-mediated inflammation through DUSP14, SOCS6 and other pathways,31,32 but we did not study it in this experiment. Therefore, we can only say that the pathogenicity of miR-155 in psoriasis is mediated at least in part by the TLR4/MyD88/NF-κB signaling pathway.
Studies have demonstrated a link between oxidative stress and inflammation, and the interaction between the two can accelerate skin damage. Elevated levels of ROS and MDA and decreased levels of T-AOC are important manifestations of oxidative stress. ROS can be increased under the induction of inflammatory cytokines, and the excessive production of ROS in turn further aggravates inflammation.33 Our study showed that oxidative stress and inflammation were activated in psoriasis group, and the levels of inflammatory factors TNF-α, IL-6, ROS and MDA were decreased and T-AOC level was increased in CD14+ monocytes after inhibition of miR-155 and TLR4 pathways, respectively. These results indicate that both miR-155 and TLR4 inhibit inflammation and oxidation. Due to the close relationship between the TLR4 pathway and oxidation and inflammation, we hypothesized that the inhibition of miR-155 on oxidative stress and inflammatory response of CD14+ monocytes in psoriasis may be related to the TLR4 signaling pathway. Other studies have reported results that are consistent with ours.34,35 However, other studies have shown that decreased SOCS1 can directly lead to increased production of ROS through thioredoxin, independent of TLR4.36 In this study, the expression of SOCS1 increased after miR-155 inhibition, and whether this is related to the increase of ROS production remains to be proved in subsequent experiments.
Conclusion
In summary, the present study demonstrates that expression levels of miR-155 were increased in psoriatic CD14+ monocytes. MiR-155 positively regulates proliferation, expression levels of proinflammatory cytokines, and oxidative stress of CD14+ monocytes via TLR4/MyD88/NF-κB signaling pathway. However, whether the effect of miR-155 on CD14+ monocyte function is via other signaling pathways remains to be determined. The results of the present study also suggest that intervention of miR-155 and/or TLR4/MyD88/NF-κB signaling pathway could become a valuable approach in the management of psoriasis.
Acknowledgments
The work was supported by the National Natural Science Foundation of China (No. 81773336).
Ethics Statement
The study was approved by the institutional ethics committee of Taiyuan City Central Hospital (2016005). The study was carried out with the informed consent of the patients themselves and in accordance with the Declaration of Helsinki Principles.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Karabay EA, Demir D, Erman AA. Evaluation of monocyte to high-density lipoprotein ratio, lymphocytes, monocytes, and platelets in psoriasis. An Bras Dermatol. 2020;95(1):40–45. doi:10.1016/j.abd.2019.05.002
2. Hamers AAJ, Dinh HQ, Thomas GD, et al. Human monocyte heterogeneity as revealed by high-dimensional mass cytometry. Arterioscler Thromb Vasc Biol. 2019;39(1):25–36. doi:10.1161/ATVBAHA.118.311022
3. Singh TP, Zhang HH, Borek I, et al. Monocyte-derived inflammatory Langerhans cells and dermal dendritic cells mediate psoriasis-like inflammation. Nat Commun. 2016;7:13581. doi:10.1038/ncomms13581
4. Do JE, Kwon SY, Park S, et al. Effects of vitamin D on expression of Toll-like receptors of monocytes from patients with Behcet’s disease. Rheumatology. 2008;47(6):840–848. doi:10.1093/rheumatology/ken109
5. Takamura N, Yamaguchi Y, Watanabe Y, et al. Downregulated Caveolin-1 expression in circulating monocytes may contribute to the pathogenesis of psoriasis. Sci Rep. 2019;9(1):125. doi:10.1038/s41598-018-36767-5
6. Zhou HB, Huang XF, Cui HJ, et al. miR-155 and its star-form partner miR-155* cooperatively regulate type I interferon production by human plasmacytoid dendritic cells. Blood. 2010;116(26):5885–5894. doi:10.1182/blood-2010-04-280156
7. O’Neill LA, Golenbock D, Bowie AG. The history of Toll-like receptors - redefining innate immunity. Nat Rev Immunol. 2013;13(6):453–460.
8. Kim SJ, Kim HM. Dynamic lipopolysaccharide transfer cascade to TLR4/MD2 complex via LBP and CD14. BMB Rep. 2017;50(2):55–57. doi:10.5483/BMBRep.2017.50.2.011
9. Wang Y, Weng H, Song JF, et al. Activation of the HMGB1-TLR4-NF-κB pathway may occur in patients with atopic eczema. Mol Med Rep. 2017;16(3):2714–2720. doi:10.3892/mmr.2017.6942
10. Akira S, Takeda K. Toll-like receptor signalling. Cold Spring Harb Perspect Biol. 2004;4:499–511.
11. Rommy VB, Laura E, Jaime E. Microglial cell dysregulation in brain aging and neurodegeneration. Front Aging Neurosci. 2015;7(124):124.
12. Moore CS, Rao VTS, Durafourt BA, et al. miR-155 as a multiple sclerosis-relevant regulator of myeloid cell polarization. Ann Neurol. 2013;74(5):709–720. doi:10.1002/ana.23967
13. Zhou Y, Zhang PX, Zheng XY, et al. miR-155 regulates pro- and anti-inflammatory cytokine expression in human monocytes during chronic hepatitis C virus infection. Ann Transl Med. 2021;9(21):1618. doi:10.21037/atm-21-2620
14. Soonthornchai W, Tangtanatakul P, Meephansan J, et al. Down-regulation of miR-155 after treatment with narrow-band UVB and methotrexate associates with apoptosis of keratinocytes in psoriasis. Asian Pac J Allergy Immunol. 2021;39(3):206–213.
15. Hou RX, Liu RF, Zhao XC, et al. Increased miR-155-5p expression in dermal mesenchymal stem cells of psoriatic patients: comparing the microRNA expression profile by microarray. Genet Mol Res. 2016;15(3). doi:10.4238/gmr.15038631
16. Liu F, Fan H, Ren D, et al. TLR9-induced miR-155 and Ets-1 decrease expression of CD1d on B cells in SLE. Eur J Immunol. 2015;45(7):1934–1945. doi:10.1002/eji.201445286
17. Bala S, Tilahun Y, Taha O, et al. Increased microRNA-155 expression in the serum and peripheral monocytes in chronic HCV infection. J Transl Med. 2012;10:151. doi:10.1186/1479-5876-10-151
18. El-Ekiaby N, Hamdi N, Negm M, et al. Repressed induction of interferon-related microRNAs miR-146a and miR-155 in peripheral blood mononuclear cells infected with HCV genotype 4. FEBS Open Bio. 2012;2(1):179–186. doi:10.1016/j.fob.2012.07.005
19. Fadime D, Ilknur K, Öznur A, et al. Investigation of TLR1-9 genes and miR-155 expression in dogs infected with canine distemper. Comp Immunol Microbiol Infect Dis. 2021;79:101711. doi:10.1016/j.cimid.2021.101711
20. Husakova M. MicroRNAs in the key events of systemic lupus erythematosus pathogenesis. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2016;160(3):327–342. doi:10.5507/bp.2016.004
21. Wen Y, Zhang XJ, Dong LP, et al. Acetylbritannilactone modulates microRNA-155-mediated inflammatory response in ischemic cerebral tissues. Mol Med. 2015;21(1):197–209. doi:10.2119/molmed.2014.00199
22. Akhter N, Madhoun A, Arefanian H, et al. Oxidative stress induces expression of the Toll-like receptors (TLRs) 2 and 4 in the human peripheral blood mononuclear cells: implications for metabolic inflammation. Cell Physiol Biochem. 2019;53(1):1–18.
23. Chiu YG, Shao T, Feng C. CD16 (FcRγIII) as a potential marker of osteoclast precursors in psoriatic arthritis. Arthritis Res Ther. 2010;12(1):R14. doi:10.1186/ar2915
24. Yamanaka K, Umezawa Y, Yamagiwa A, et al. Biologic therapy improves psoriasis by decreasing the activity of monocytes and neutrophils. J Dermatol. 2014;41(8):679–685.
25. Yang X, Wang H. miRNAs flowing up and down: the concerto of psoriasis. Front Med. 2021;8:646796.
26. Xu L, Len H, Shi X, et al. MiR-155 promotes cell proliferation and inhibits apoptosis by PTEN signaling pathway in the psoriasis. Biomed Pharmacother. 2017;90:524–530. doi:10.1016/j.biopha.2017.03.105
27. Li C, He H, Zhu M, et al. Molecular characterisation of porcine miR-155 and its regulatory roles in the TLR3/TLR4 pathways. Dev Comp Immunol. 2013;39(1–2):110–116. doi:10.1016/j.dci.2012.01.001
28. Garg A, Aggarwal BB. Nuclear transcription factor-κB as a target for cancer drug development. Leukemia. 2002;16:1053–1068.
29. Taniguchi K, Karin M. NF-κB, inflammation, immunity and cancer: coming of age. Nat Rev Immunol. 2018;18(5):309–324.
30. Kocic H, Damiani G, Stamenkovic B, et al. Dietary compounds as potential modulators of microRNA expression in psoriasis. Ther Adv Chronic Dis. 2019;10:204062231986480. doi:10.1177/2040622319864805
31. Shi Y, Li K, Xu K, et al. MiR-155-5p accelerates cerebral ischemia-reperfusion injury via targeting DUSP14 by regulating NF-κB and MAPKs signaling pathways. Eur Rev Med Pharmacol Sci. 2020;24(3):1408–1419.
32. Ge X, Tang P, Rong Y, et al. Exosomal miR-155 from M1-polarized macrophages promotes EndoMT and impairs mitochondrial function via activating NF-κB signaling pathway in vascular endothelial cells after traumatic spinal cord injury. Redox Biol. 2021;41:101932.
33. Wang Y, Branicky R, Noë A, Hekimi S. Superoxide dismutases: dual roles in controlling ROS damage and regulating ROS signaling. J Cell Biol. 2018;217(6):1915–1928. doi:10.1083/jcb.201708007
34. Yang ZB, Chen WW, Chen HP, et al. MiR-155 aggravated septic liver injury by oxidative stress-mediated ER stress and mitochondrial dysfunction via targeting Nrf-2. Exp Mol Pathol. 2018;105(3):387–394. doi:10.1016/j.yexmp.2018.09.003
35. Fu X, He HD, Li CJ, et al. MicroRNA-155 deficiency attenuates inflammation and oxidative stress in experimental autoimmune prostatitis in a TLR4-dependent manner. Kaohsiung J Med Sci. 2020;36(9):712–720. doi:10.1002/kjm2.12229
36. Jung S, Kim S, Lee C, et al. Mechanism of suppressors of cytokine signaling 1 inhibition of epithelial-mesenchymal transition signaling through ROS regulation in colon cancer cells: suppression of Src leading to thioredoxin up-regulation. Oncotarget. 2016;7(38):62559–62571. doi:10.18632/oncotarget.11537
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.