Back to Journals » International Journal of Women's Health » Volume 18
Indirect Roles of Immunophenotypes in the Association Between Glycerophospholipid Metabolism and Endometrial Cancer Risk: A Mendelian Randomization Study
Authors Shen Y ![]() , Yan Y, Lin P, Wang Y, Li Y, Li H
, Yan Y, Lin P, Wang Y, Li Y, Li H ![]() , Li Y, Dai Y
, Li Y, Dai Y
Received 12 November 2025
Accepted for publication 30 March 2026
Published 15 April 2026 Volume 2026:18 580946
DOI https://doi.org/10.2147/IJWH.S580946
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Vinay Kumar
Yichen Shen,1,* Yijing Yan,2,* Peizhe Lin,3,* Yulin Wang,1 Yafang Li,1 Huhu Li,4 Yanqi Li,1 Yongna Dai1
1School of Traditional Chinese Medicine, Tianjin University of Traditional Chinese Medicine, Tianjin, People’s Republic of China; 2Department of Traditional Chinese Medicine, National Cancer Center/National Clinical Research Center for Cancer/Cancer Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing, People’s Republic of China; 3Guang’anmen Hospital, Chinese Academy of Chinese Medical Sciences, Beijing, People’s Republic of China; 4School of Integrative Medicine, Tianjin University of Traditional Chinese Medicine, Tianjin, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yongna Dai, Email [email protected] Yanqi Li, Email [email protected]
Background: The incidence of endometrial cancer (EC) has continued to rise in recent years. Increasing evidence suggests that lipid metabolic alterations in serum metabolites are associated with cancer progression.
Methods: This research applied a two-step Mendelian Randomization (MR), two-sample MR methodology to explore the potential causal associations between glycerophospholipid metabolism and EC risk, as well as possible indirect roles of immunophenotypes, drawing data from the Genome-Wide Association Study Catalog and EC Association Consortium databases. The Inverse Variance Weighted, MR-Egger regression, Weighted Median Estimator, Simple Mode, and Weighted Mode approaches formed a comprehensive array of MR methodologies predominantly employed to assess causality. Several sensitivity analyses were implemented to verify the reliability of MR results.
Results: Genetically predicted levels of three metabolites were associated with EC risk, with evidence suggesting partially indirect pathways involving four immunophenotypes. 1-(1-enyl-palmitoyl)-GPC (p-16:0) was found to diminish EC risk, potentially through indirect pathways involving, specifically CD33dim HLA-DR+ CD11b- AC, and CD25 on memory B cells, explaining 3.99% and 5.68% to the total effect, respectively. Additionally, 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1) decreased EC risk, with partial mediation by B cell % CD3- lymphocyte, accounting for a 3.25% share of the indirect effect. Conversely, Eicosenedioate (C20:1-DC) increased EC risk, with its effect being mediated by CD38 on PB/PC cells, exerting a indirect effect of up to 10.09%.
Conclusion: This investigation provides genetic evidence supporting potential causal associations between glycerophospholipid metabolism and EC, highlighting possible indirect pathways of immunophenotypes. The derived insights offer novel pathways for the diagnosis and therapeutic intervention of EC.
Keywords: endometrial cancer, glycerophospholipid metabolism, immunophenotypes, mendelian randomization, indirect effects
Introduction
Endometrial cancer (EC), recognized as one of the most common gynecological malignancies in developed nations, has shown a continuous increase in both incidence and mortality across Europe, the United States.1 More than 400,000 new cases are diagnosed worldwide each year,2 and projections indicate that the incidence rate of this cancer may escalate by 40–50% over the next decade.3 Presently, the screening and diagnostic protocols for EC primarily rely on hysteroscopy or biopsy of endometrial specimens. However, these invasive procedures are still constrained by high costs, inherent risks, and discomfort,4 making it more hinder early-stage detection. Therefore, identifying reliable biomarkers from non-invasive biological samples, particularly blood-based indicators, may facilitate early detection and improve risk stratification strategies.
Metabolomics holds potential for the non-invasive diagnosis and screening of EC by providing comprehensive molecular information that may help elucidate mechanisms underlying cancer development and pathological features relevant to early diagnosis and targeted therapy.5 In clinical practice, serum metabolite profiling offers practical advantages due to its minimal invasiveness, repeatability, and suitability for large-scale population screening. Importantly, circulating metabolites not only reflect systemic metabolic status but also serve as functional readouts of tumor-associated biochemical alterations, thereby bridging clinical biomarker discovery and mechanistic investigations of cancer biology.6 Serum metabolomics, typically performed using untargeted metabolomic approach, exhibits high sensitivity and throughput, and has been increasingly explored as a significant resource for the discovery of biomarker discovery for cancer surveillance and screening.7 Glycerophospholipids are major structural components of cellular membranes and play essential roles in membrane fluidity, signal transduction, and inflammatory regulation. Dysregulation of glycerophospholipid metabolism may influence tumor cell proliferation, oxidative stress responses, and remodeling of the immune microenvironment, highlighting the potential clinical value of detecting serum metabolites for identifying individuals at elevated risk.8,9 Nevertheless, despite growing interest in serum metabolomics for EC research, several challenges remain, including ambiguous potential causal associations and specific pathways between glycerophospholipid metabolism and EC, insufficient validation across large patient cohorts, and the inability to adequately control for confounding factors.
By leveraging randomly allocated genetic variants as instrumental variables, Mendelian randomization (MR) enables the assessment of potential causal associations between exposures and specific outcomes while reducing bias from confounding and reverse causality.10 MR has been increasingly applied to investigate metabolic traits and cancer susceptibility, providing more robust evidence for causal inference compared with conventional observational studies. Previous MR studies have identified causal associations between circulating metabolites and multiple malignancies, including colorectal cancer, breast cancer, and prostate cancer, highlighting the value of genetically informed approaches in uncovering metabolic risk factors and potential therapeutic targets.11–13
Moreover, lipid metabolism is closely intertwined with immune regulation. Glycerophospholipid metabolites participate in membrane receptor organization, antigen presentation processes, and immune cell signaling pathways, thereby potentially influencing immune cell activation, differentiation, and effector functions. Increasing evidence suggests that metabolic reprogramming of immune cells plays a crucial role in tumor immune surveillance and immune escape.6 Therefore, investigating the interplay between glycerophospholipid metabolism and immunophenotypes may provide additional insights into immune-related pathways involved in EC development.
Within the scope of this investigation, single nucleotide polymorphisms (SNPs) were utilized as instrumental variables (IVs). We performed a two-sample MR analysis to evaluate potential causal associations between glycerophospholipid metabolites and EC risk. Furthermore, a two-step MR framework was implemented to examine whether immunophenotypes may partially explain the associations between glycerophospholipid metabolism and EC. This study aims to provide additional insights into the biological processes potentially linking lipid metabolism with EC and to identify candidate biomarkers for risk assessment and targeted prevention.
Materials and Methods
Study Design
By utilizing two-sample MR and two-step MR approaches, a comprehensive analysis was conducted with 1400 serum metabolites serving as exposure variables to investigate potential causal associations between these metabolites and EC outcomes. The direct effect of serum metabolites on EC was quantified as the overall effect (β) employing the TSMR technique. The discrete causal influences of serum metabolites on immune cells (β1) and immune cells on EC (β2) were individually examined through the two-step MR framework, with the indirect associations’ magnitudes ascertained by multiplying the respective causal estimates derived from the two-step MR. The indirect associations’ proportion attributable to immune cells within the serum metabolites-EC continuum was delineated by apportioning the fraction of the indirect associations relative to the overall effect. The indirect associations’ magnitudes were ascertained using the formula: β1×β2. The indirect association’s percentage was derived using the formula: (β1×β2)/β. This analytical framework enables decomposition of total effects into direct and indirect components, facilitating exploration of potential immune-related pathways linking metabolites with EC risk. The derived beta coefficients were transformed into odds ratios (ORs), and 95% confidence intervals (CIs) were ascertained for this research. CIs for indirect associations were estimated using the Delta method to account for variance propagation in the product of coefficients (Figure 1).
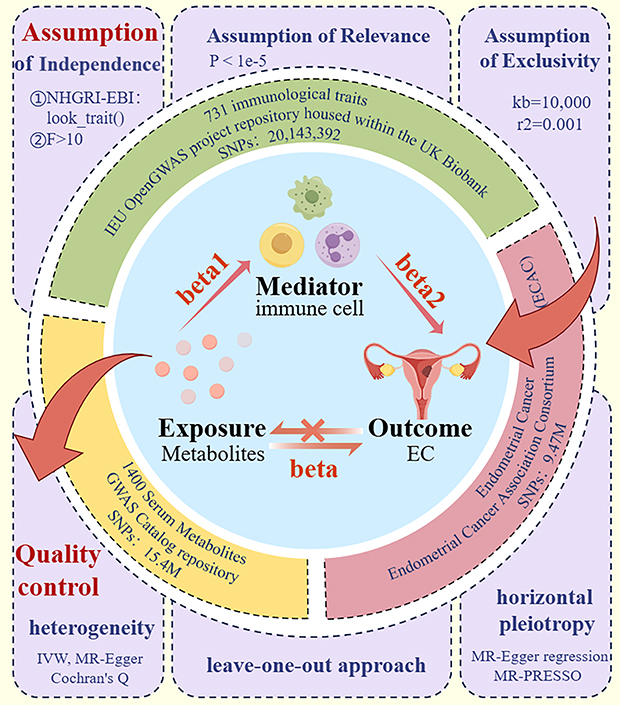 |
Figure 1 General flow chart of the study. TSMR and two-step MR framework were applied to explore potential associations between 1400 serum metabolites and EC. The total effect was estimated using TSMR, while the indirect effect was calculated as β1×β2, where β1 represents the effect of metabolites on immune cells and β2 represents the effect of immune cells on EC. Data came from GWAS Catalog, UK Biobank with 731 immune traits, and ECAC with over 12,000 EC cases. SNPs satisfying the three core MR assumptions were selected as IVs, and instrument strength was assessed using the F-statistic and R². The IVW method was used as the primary analysis. Effect estimates are presented as ORs with 95% CIs. Sensitivity analyses, including heterogeneity tests, were conducted to ensure the robustness of the results. |
Data Sources
Descriptive statistical data pertaining to human serum metabolites were extracted from the Genome-Wide Association Study (GWAS) Catalog repository. A total of 1400 serum metabolites were registered within the database under the accession numbers GCST90199621 to GCS90201020.
Comprehensive statistical analyses pertaining to immunological characteristics were derived from the Integrative Epidemiology Unit Open GWAS project repository housed within the United Kingdom Biobank. A total of 731 immunological traits were ascertained, each assigned a unique GWAS catalog identifier ranging from GCST90001391 to GCST90002121.
Aggregate statistical data pertaining to EC were sourced from the EC Association Consortium (ECAC). The dataset encompassed 108,979 control subjects and 12,906 individuals with EC, drawn from various European countries, culminating in the identification of approximately 9.47 million SNPs.
Theoretically, All GWAS summary statistics were obtained from independent consortia with no known participant overlap. To further reduce potential bias caused by sample overlap, linkage disequilibrium score regression (LDSC) was performed to evaluate genetic correlations between exposure and outcome datasets.
All original GWAS studies had received ethical approval from their respective institutional review boards, and all participants provided informed consent. The present study used only de-identified publicly available summary-level data and therefore required no additional ethical approval.
Screening of IVs
Within the framework of MR analysis, to optimize the precision of the analysis and mitigate potential errors, these IVs must rigorously adhere to three fundamental presuppositions: (1) Assumption of Relevance: The IV exhibits a robust correlation with serum metabolites, serving as a proxy for the exposure factor. (2) Assumption of Independence: The IV is posited to be orthogonal to the confounding variables that may influence the outcome, thereby ensuring its exogeneity. (3) Assumption of Exclusivity: The IV is hypothesized to exert its influence on the outcome solely through its interaction with serum metabolites, modulating immune cell characteristics.
SNPs with high correlation with serum metabolites, immune profile, and EC at the genome-wide level were selected as IVs by association analysis, and the screening threshold condition was P < 1e−5 to meet the first MR hypothesis. To ensure independence between IVs, linkage disequilibrium (LD) clumping was performed using an r² threshold < 0.001 within a 10,000 kb window. Potential duplicates or palindromes were eliminated using the harmonise_data() function with the action=2 parameter. In addition, the Steiger directionality test was conducted to verify that the genetic variants explained more variance in the exposure than in the outcome, ensuring correct causal direction. Concurrently, Instrument strength was evaluated by calculating the F-statistic for each SNP using the formula F=β2/SE2. SNPs with F<10 were considered weak instruments and were excluded to reduce weak instrument bias. Ultimately, The NHGRI-EBI Catalog, a repository of curated genome-wide association database, was leveraged to ascertain the linkage between SNPs and phenotypic manifestations, thereby facilitating the mitigation of confounding variables. This process was executed utilizing the look_trait() function within the R package FastTraitR. The coefficient of determination (R-squared, R2) was computed utilizing the formula: R2=2×Minor Allele Frequency (MAF)×(1-MAF)×β2, where β represents the allelic effect size. The F-statistic was derived from the formula: 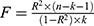 , with k signifying the count of IVs, and n denoting the sample size.
, with k signifying the count of IVs, and n denoting the sample size.
MR Analysis
The implementation was facilitated by the R package Two-Sample MR (version 4.3.2). A comprehensive array of MR methodologies, encompassing the Inverse Variance Weighted (IVW), MR-Egger regression, Weighted Median Estimator (WME), Simple Mode, and Weighted Mode approaches, were predominantly utilized to evaluate the causality. The IVW approach was designated as the principal reference method for evaluating causal effects, due to its superior statistical power relative to the alternative four MR techniques. The threshold for statistical significance was set at P<0.05, indicating a notable link between the exposure and outcome variables. Additionally, an inverse MR analysis was implemented to address the potential confounding influence of reverse causality. Ultimately, the intermediary function of immune cells within the associative framework between serum metabolites and EC was investigated using a two-step MR approach.
Quality Control
To ascertain the robustness of the study’s findings, sensitivity analyses were conducted, encompassing heterogeneity, horizontal gene multiplicity test, and the leave-one-out approach. Using Cochran’s Q statistic, the IVW method and MR-Egger’s regression analysis were employed in evaluating heterogeneity, as facilitated by the MR_heterogeneity function. A P-value exceeding 0.05 suggested that the observed discrepancies were not statistically significant, thereby negating the effect of heterogeneity on the study’s outcomes. The presence of horizontal multiplicity was verified by the MR-Egger intercept and MR-PRESSO statistical assessment, which indicated the presence of horizontal multiplicity when the MR-Egger intercept was statistically significant. The effect of individual SNPs on causality was determined by eliminating each SNP, in turn, using leave-one-out analysis. Sensitivity analyses were separately conducted for each step of the two-step MR indirect pathway to ensure robustness of indirect effect estimates.
Results
TSMR Analysis
Causal Effects of Serum Metabolites and EC
TSMR analysis of serum metabolites and EC was performed with IVW as the principal referent. A P-value threshold of P<0.05 was set as the benchmark for determining whether the potential causal effect was statistically significant. The analysis revealed that 72 serum metabolites were significantly associated with EC, comprising 45 risk factors and 27 protective factors (Figure 2).
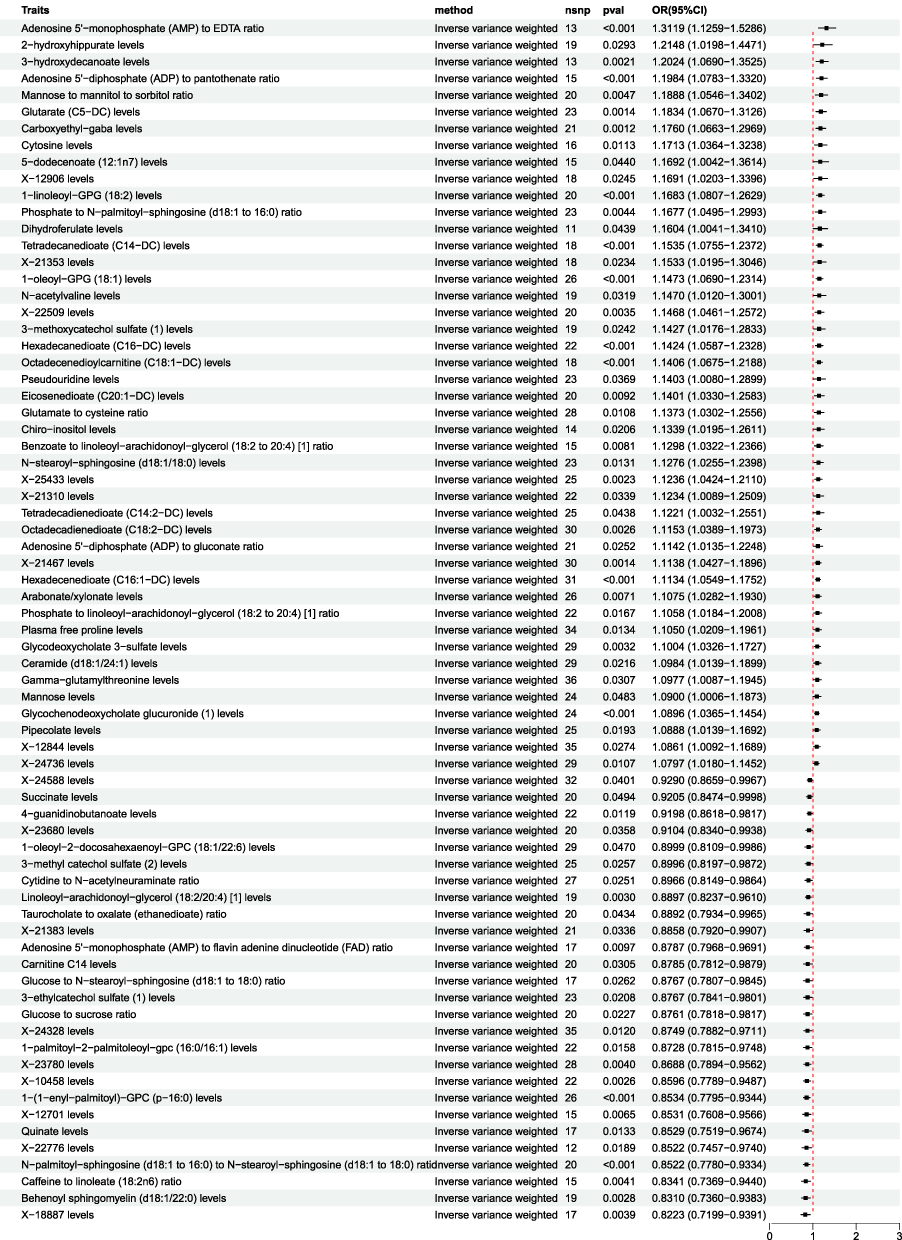 |
Figure 2 Forest plot showing the MR analysis of serum metabolites and EC. The IVW method revealed that 72 serum metabolites were significantly associated with endometrial cancer (P<0.05), including 45 risk factors (OR>1) and 27 protective factors (OR<1). |
Selected Serum Metabolites Associated with EC
Based on the analysis results, five glycerophospholipid metabolites that exhibit significant and strong correlations with EC and hold important clinical significance were selected for further analysis, specifically 1-(1-enyl-palmitoyl)-GPC (p-16:0) levels, 1-oleoyl-GPG (18:1), 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1), 1-oleoyl-2-docosahexaenoyl-GPC (18:1/22:6), and Eicosenedioate (C20:1-DC) (Figure 3).
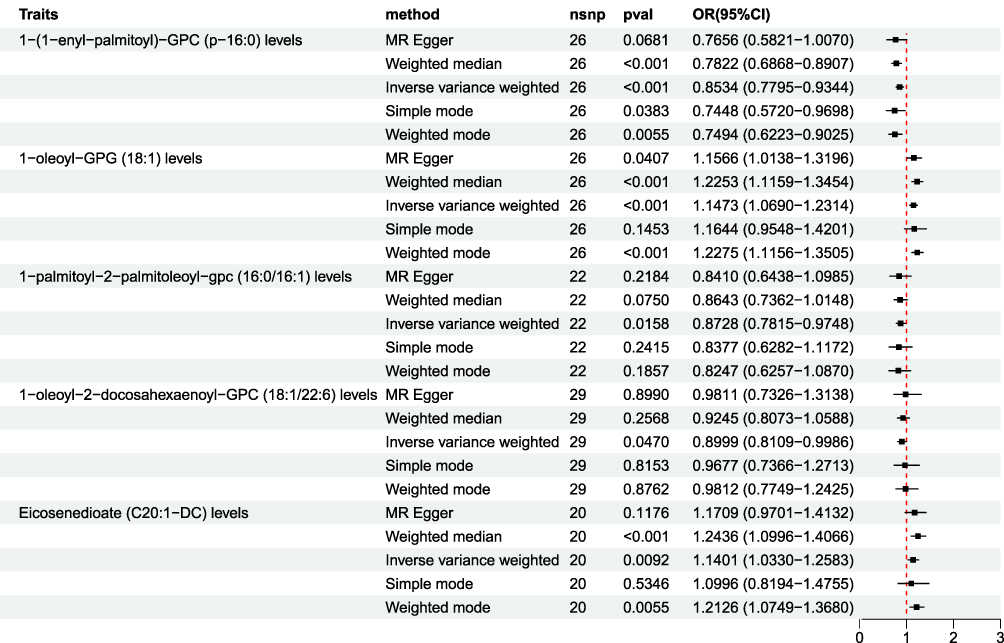 |
Figure 3 Forest plot displaying five methods of MR analysis (IVW, MR-Egger, weighted median, simple mode, and weighted mode) for five glycerophospholipid metabolites associated with EC. The analysis of IVW results indicated that levels of 1-oleoyl-GPG (18:1) and Eicosenedioate (C20:1-DC) were positively correlated with EC, exhibiting ORs of 1.1473 and 1.1401 (p<0.05). 1-(1-enyl-palmitoyl)-GPC (p-16:0), 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1), and 1-oleoyl-2-docosahexaenoyl-GPC (18:1/22:6) exhibited OR values below 1, showing negative correlations with EC (p<0.05). |
Causal Effects of EC and Serum Metabolites
MR causality analyses, utilizing EC as the independent variable and glycerophospholipid metabolites as the dependent variables, identified a total of 123 SNPs. Across all five applied algorithms, it was observed that the five glycerophospholipid metabolites implicated in EC exhibited P-values exceeding 0.05 in the inverse MR analyses (Table 1). This suggests that genetic predisposition to EC does not significantly influence the levels of these glycerophospholipid metabolites.
 |
Table 1 Results of Inverse MR Analysis of EC and Serum Metabolites |
Sensitivity Analysis
The results were analyzed for sensitivity, encompassing heterogeneity testing, horizontal pleiotropy assessment, and leave-one-out validation, which were conducted to scrutinize the robustness of the findings. Specifically, Cochran’s Q test for the IVW method applied to the aforementioned five glycerophospholipid metabolites produced non-significant findings (P>0.05), indicating that potential heterogeneity-induced bias did not confound the study’s outcomes. The MR-Egger intercepts for these metabolites also revealed non-significance (P>0.05), signifying the absence of genetic pleiotropy bias. MR-PRESSO test did not have horizontal gene pleiotropy. The leave-one-out approach demonstrated that excluding any SNP did not notably change the effect size estimates, which supports the consistency of the causal inferences (Figure 4A–E). As delineated in, The sensitivity analysis further substantiated the study’s findings, with no evidence indicating that any SNPs compromised the overarching causality (Table 2 and Figure 5A–E; Figure 6A–E; Figure 7A–E).
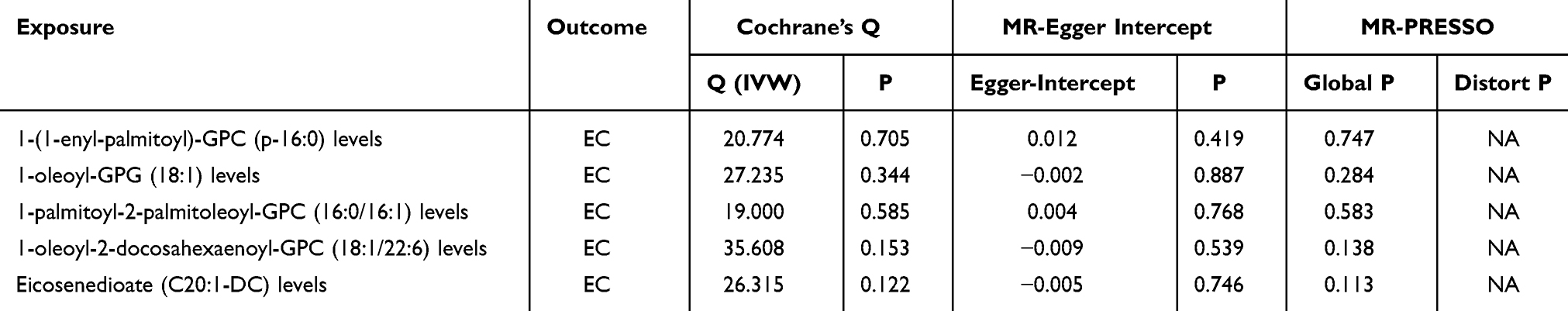 |
Table 2 Results of Sensitivity Analysis of Glycerophospholipid Metabolites |
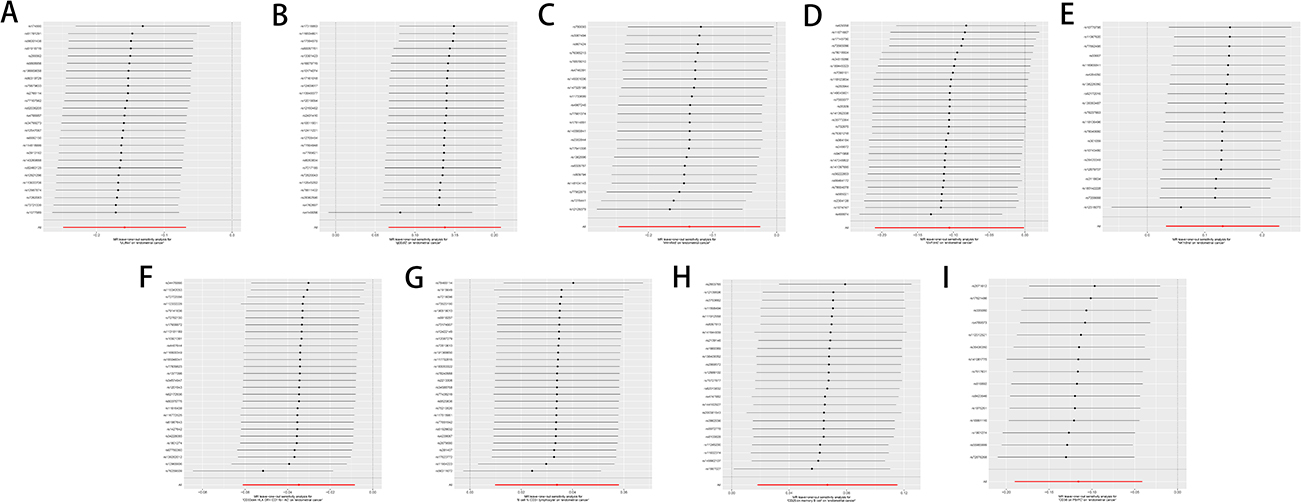 |
Figure 4 Leave-one-out method analysis of glycerophospholipid metabolites and immunophenotypes with EC. (A) Glycerophospholipid metabolites 1-(1-enyl-palmitoyl)-GPC (p-16:0) levels; (B) Glycerophospholipid metabolites 1-oleoyl-GPG (18:1) levels; (C) Glycerophospholipid metabolites 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1) levels; (D) Glycerophospholipid metabolites 1-oleoyl-2-docosahexaenoyl-GPC (18:1/22:6) levels; (E) Glycerophospholipid metabolites Eicosenedioate (C20:1-DC) levels; (F) Immunophenotype CD33dim HLA-DR+ CD11b− AC; (G) Immunophenotype B cell % CD3− lymphocyte; (H) Immunophenotype CD25 on memory B cell; (I) Immunophenotype CD38 on PB/PC. |
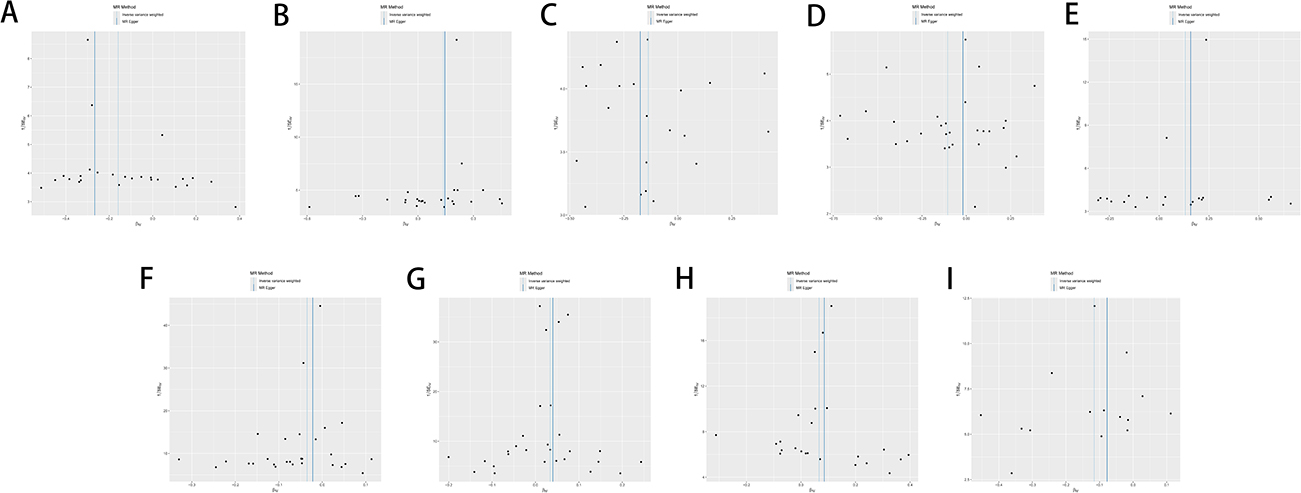 |
Figure 5 Funnel plot sensitivity analysis of glycerophospholipid metabolites and immunophenotypes with EC. (A) Glycerophospholipid metabolites 1-(1-enyl-palmitoyl)-GPC (p-16:0) levels; (B) Glycerophospholipid metabolites 1-oleoyl-GPG (18:1) levels; (C) Glycerophospholipid metabolites 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1) levels; (D) Glycerophospholipid metabolites 1-oleoyl-2-docosahexaenoyl-GPC (18:1/22:6) levels; (E) Glycerophospholipid metabolites Eicosenedioate (C20:1-DC) levels; (F) Immunophenotype CD33dim HLA-DR+ CD11b− AC; (G) Immunophenotype B cell % CD3− lymphocyte; (H) Immunophenotype CD25 on memory B cell; (I) Immunophenotype CD38 on PB/PC. |
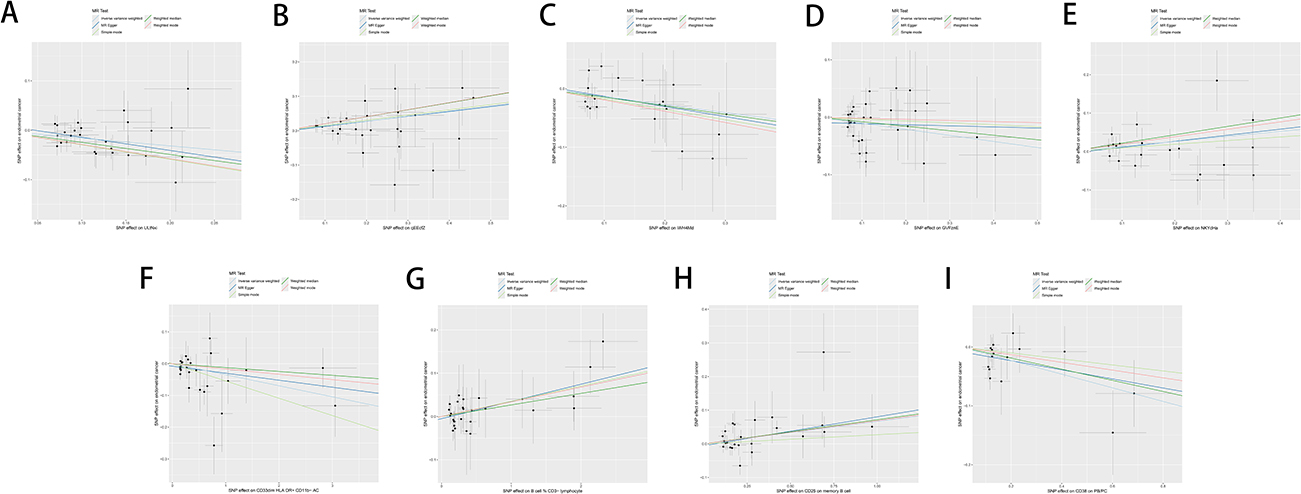 |
Figure 6 Scatter plot sensitivity analysis of glycerophospholipid metabolites and immunophenotypes with EC. (A) Glycerophospholipid metabolites 1-(1-enyl-palmitoyl)-GPC (p-16:0) levels; (B) Glycerophospholipid metabolites 1-oleoyl-GPG (18:1) levels; (C) Glycerophospholipid metabolites 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1) levels; (D) Glycerophospholipid metabolites 1-oleoyl-2-docosahexaenoyl-GPC (18:1/22:6) levels; (E) Glycerophospholipid metabolites Eicosenedioate (C20:1-DC) levels; (F) Immunophenotype CD33dim HLA-DR+ CD11b− AC; (G) Immunophenotype B cell % CD3− lymphocyte; (H) Immunophenotype CD25 on memory B cell; (I) Immunophenotype CD38 on PB/PC. |
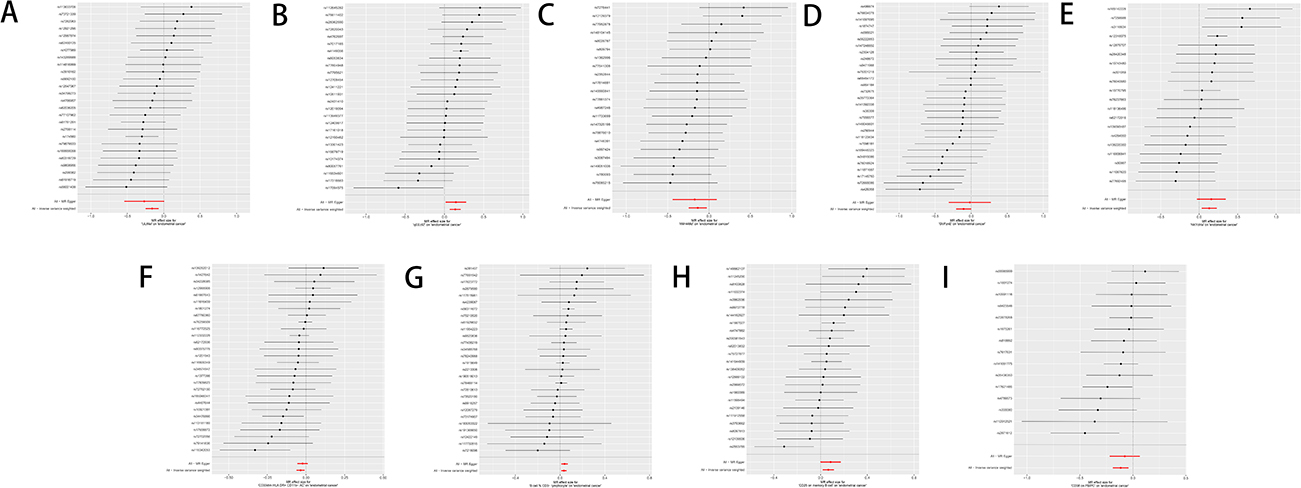 |
Figure 7 Forest plot sensitivity analysis of glycerophospholipid metabolites and immunophenotypes with EC. (A) Glycerophospholipid metabolites 1-(1-enyl-palmitoyl)-GPC (p-16:0) levels; (B) Glycerophospholipid metabolites 1-oleoyl-GPG (18:1) levels; (C) Glycerophospholipid metabolites 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1) levels; (D) Glycerophospholipid metabolites 1-oleoyl-2-docosahexaenoyl-GPC (18:1/22:6) levels; (E) Glycerophospholipid metabolites Eicosenedioate (C20:1-DC) levels; (F) Immunophenotype CD33dim HLA-DR+ CD11b− AC; (G) Immunophenotype B cell % CD3− lymphocyte; (H) Immunophenotype CD25 on memory B cell; (I) Immunophenotype CD38 on PB/PC. |
Two-Step MR Analysis
Indirect Effects of Immune Cells on Glycerophospholipid Metabolites-EC Risk
An analysis of indirect effects was conducted to assess the potential intermediary role of immune cells within the associative framework between glycerophospholipid metabolites and EC. TSMR, encompassing 731 immune cells in relation to EC, identified 16 immune cells with a potential association with EC. Subsequently, these 16 immune cells, along with the glycerophospholipid metabolites 1-oleoyl-GPG (18:1), Eicosenedioate (C20:1-DC), 1-(1-enyl-palmitoyl)-GPC (p-16:0), 1-oleoyl-2-docosahexaenoyl-GPC (18:1/22:6), and 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1), underwent further TSMR analysis. The findings revealed that 8 immune cells were potentially associated with the aforementioned 5 glycerophospholipid metabolites, with 4 immune cells exhibiting OR values aligned with the directionality of the initial TSMR results. Notably, a P-value threshold of below 0.05 was observed, denoting statistical significance and warranting progression to subsequent analytical phases (Figure 8).
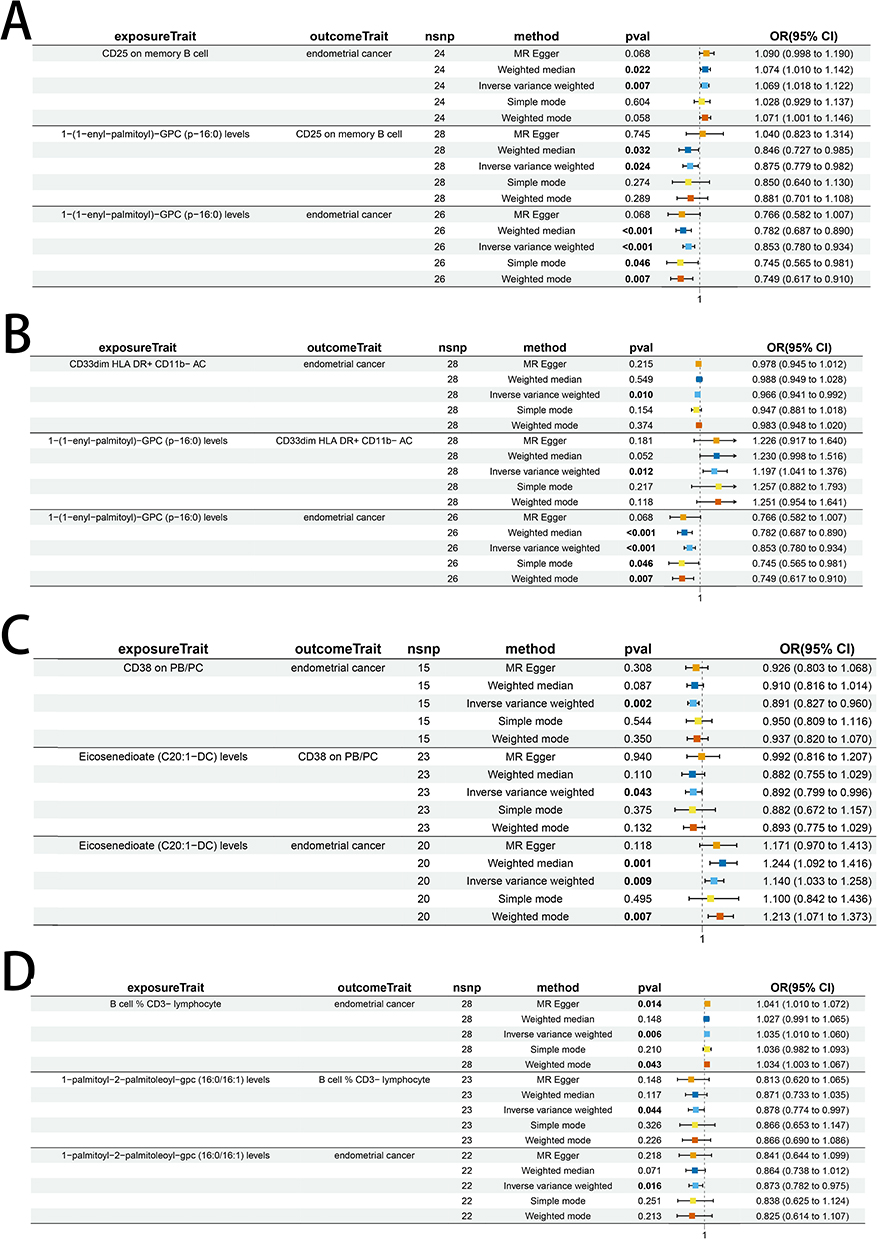 |
Figure 8 Results of two-step MR analysis of glycerophospholipid metabolite-immune cell-EC. (A) Metabolite 1-(1-enyl-palmitoyl)-GPC (p-16:0) levels, mediated by immune phenotype CD25 on memory B cells; (B) Metabolite 1-(1-enyl-palmitoyl)-GPC (p-16:0) levels, mediated by immune phenotype CD33dim HLA DR+ CD11b-AC; (C) Metabolite Eicosenedioate (C20:1-DC) levels, mediated by immune phenotype CD38 ON PB/PC; (D) Metabolite 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1) levels, mediated by immune phenotype: B cell % CD3- lymphocyte. The effect estimates include β1 (metabolites → immune cells) and β2 (immune cells → EC), along with corresponding ORs and 95% CIs. |
Two-step MR findings indicated a bivariate correlation between the glycerophospholipid metabolite 1-(1-enyl-palmitoyl)-GPC (p-16:0) and specific immune cell populations. Specifically, a positive correlation was observed with CD33dim HLA DR+ CD11b- AC, and a negative correlation with CD25 on memory B cells. The above suggests that the glycerophospholipid metabolite may modulate CD25 on memory B cells and CD33dim HLA DR+ CD11b- AC expression, potentially influencing the risk of EC. Additionally, the serum metabolites Eicosenedioate (C20:1-DC) and 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1) were found to suppress the activity of immune cell CD38 on PB/PC and the percentage of B cell % CD3- lymphocyte, respectively. The downregulation of CD38 on PB/PC may increase the risk of EC, while a reduction in the percentage of B cell % CD3- lymphocyte may correlate with a decreased risk of EC.
The indirect associations exerted by the aforementioned quartet of immune cells on the risk association between glycerophospholipid metabolites and EC were ascertained through a series of MR analyses. Specifically, this process involved the computation of beta coefficients (Table 3). The study’s findings revealed that the intermediary roles of immune cells, specifically CD33dim HLA DR+ CD11b- AC, CD25 on memory B cells, and B cell % CD3- lymphocyte, in the association between glycerophospholipid metabolites and EC, constituted contributions of 3.99%, 5.68%, and 3.25%, respectively. These percentages represent modest influences within the broader potential pathway. In contrast, the indirect effect of CD38 on PB/PC cells in the relationship between the serum metabolite Eicosenedioate (C20:1-DC) and EC was more pronounced, accounting for 10.09% of the total effect, indicative of a notably substantial role for this immune cell subset in mediating the risk association between Eicosenedioate (C20:1-DC) and EC.
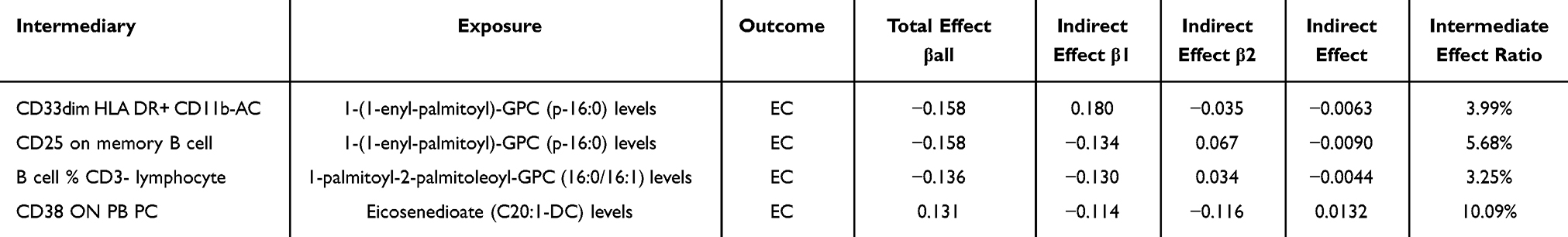 |
Table 3 Calculation of Glycerophospholipid Metabolite-Immune Cell-EC Mediated Effects |
Sensitivity Analysis
Sensitivity analyses, encompassing assessment of heterogeneity, evaluation of horizontal pleiotropy, and a leave-one-out approach, were executed to appraise the robustness of the study’s findings. Specifically, Cochran’s Q test for IVW method applied to the aforementioned quartet of immune cells resulted in non-significant findings (P>0.05), indicating that potential heterogeneity bias did not confound the study’s outcomes. The MR-Egger intercepts for these immune cells also revealed non-significance (P>0.05), signifying the absence of genetic pleiotropy bias. MR-PRESSO test did not have horizontal gene pleiotropy. Furthermore, the leave-one-out analysis demonstrated that no particular SNP significantly affected the estimates of effect size, thereby attesting to the stability of the causal inferences (Table 4 and Figure 4F–I; Figure 5F–I, Figure 6F–I; Figure 7F–I).
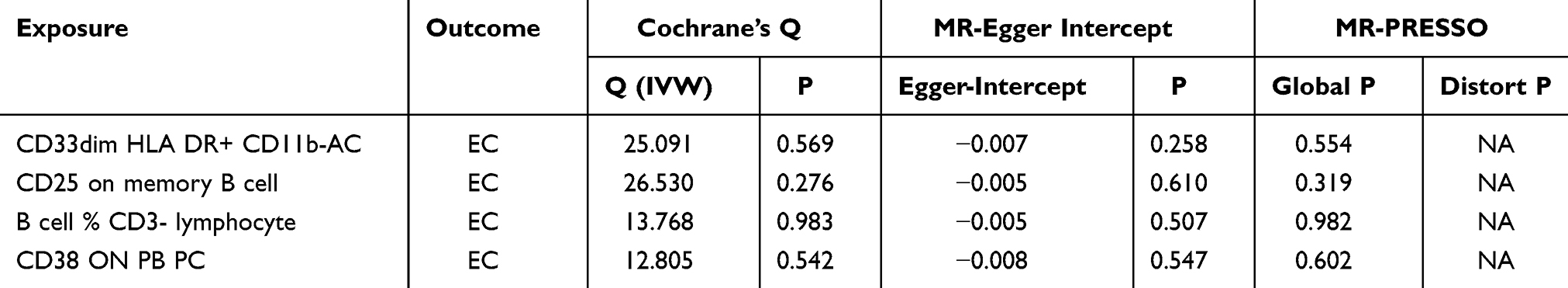 |
Table 4 Results of Immune Cell Sensitivity Analysis |
Discussion
Utilizing TSMR and two-step MR analyses, the potential association between glycerophospholipid metabolites and EC, along with the indirect role of immune cell traits, was scrutinized. The findings revealed significant correlations with elevated glycerophospholipid metabolite levels of 1-oleoyl-GPG (18:1) and Eicosenedioate (C20:1-DC), which were associated with increased EC risk. Additionally, 1-(1-enyl-palmitoyl)-GPC (p-16:0), 1-oleoyl-2-docosahexaenoyl-GPC (18:1/22:6), and 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1) exhibited protective effects against EC. The analysis of indirect effects indicated that four immune cell types exerted significant effects on three glycerophospholipid metabolites. Specifically, the reduction in EC risk associated with 1-(1-enyl-palmitoyl)-GPC (p-16:0) was partially mediated by immune cells CD33dim HLA DR+ CD11b- AC and CD25 on memory B cells, accounting for 3.99% and 5.68% of the effect, respectively. The effect of 1-palmitoyl-2-palmitoleoyl-GPC (16:0/16:1) in reducing the risk of EC was partly mediated by a 3.25% increase in the percentage of immune cells B cell % CD3- lymphocyte. The impact of Eicosenedioate (C20:1-DC) on the elevated risk of EC was partially mediated by CD38 on PB/PC cells, contributing to 10.09% of the effect. These insights may elucidate novel mechanisms underlying the etiology and progression of EC and identify potential therapeutic targets for intervention.
Glycerophosphocholine (GPC) metabolites are central intermediates in choline phospholipid metabolism, serving both as building blocks for membrane phospholipids and as intracellular signaling molecules. Their levels reflect the balance between membrane synthesis and degradation, which is particularly important in rapidly proliferating tumor cells14,15 GPC is produced from phosphatidylcholine via phospholipase A2 and lysophospholipase activity, linking membrane remodeling to bioactive lipid pools. It modulates mitochondrial function, Ca2+ signaling, NAD+/NADH balance, and ROS levels, all of which are essential for tumor cell survival and immune cell activation. Additionally, GPC influences downstream pathways including mTOR signaling and the OXPHOS/glycolysis balance, establishing a mechanistic basis for context-dependent effects.15 In cancer, GPC exhibits tumor-specific dual effects. In breast and cancers, elevated GPC promotes membrane biosynthesis and tumor cell proliferation. In hepatocellular carcinoma, excess GPC induces lipid accumulation in the tumor microenvironment, supporting immunosuppressive conditions.16,17
This study focuses on EC and provides evidence that GPC metabolites may have a protective role against EC. Our findings indicate that GPC partially reduces EC risk through specific immune cells, including CD33dim HLA-DR+ CD11b- cells, CD25 on memory B cells, and B cell % CD3- lymphocytes. Mechanistically, GPC metabolism modulates tumor-associated immune cells via metabolic reprogramming. In MDSCs, GPC enhances fatty acid oxidation and lipid droplet accumulation, promoting a less suppressive phenotype that limits their inhibitory effect on cytotoxic T cells and thereby reduces tumor-promoting activity.18,19 In B cells, including CD25 on memory B cells and B cell % CD3- lymphocytes, GPC maintains mitochondrial function and phospholipid synthesis to enhance activation and antibody production, supporting anti-tumor immunity.20,21 Our MR analyses provide genetically-informed evidence that predicted alterations in GPC metabolites are associated with reduced EC susceptibility. These findings are consistent with the mechanistic framework.20 GPC enhances the function of immune cells that mediate anti-tumor effects, thereby lowering EC risk. Overall, these results suggest that GPC metabolic alterations can modulate key immune phenotypes to exert tumor-suppressive effects in EC, highlighting its potential role in EC prevention.
Eicosenedioate, a serum metabolite classified within the arachidonic acid family and a member of the ω-6 polyunsaturated fatty acids, plays a pivotal role in promoting EC development by modulating the inflammatory microenvironment and molecular signaling pathways. Eicosenedioate is metabolized through one of three pathways: cyclooxygenase-2 (COX-2), 5-lipoxygenase (5-LOX), and P450 epoxygenase. The direct link between Eicosenedioate and EC remains to be clarified. Previous research has predominantly centered on the COX-2 and 5-LOX pathways and their associated products.22 Within the COX-2 pathway, prostaglandin F2α (PGF2α) regulates the inflammatory microenvironment in endometrial adenocarcinoma by adjusting chemokine (C-X-C motif) ligand 1 (CXCL1) expression and directing neutrophil chemotaxis through FP receptor signaling. Moreover, PGF2α stimulates angiogenesis in endometrial adenocarcinoma tissue through epidermal growth factor receptor (EGFR) trans-activation and vascular endothelial growth factor (VEGF) mRNA upregulation, thereby accelerating EC progression.23 Prostaglandin E2 (PGE2), via E-prostanoid receptor 4 (EP4) receptor activation, enhances small ubiquitin-like modifier-1 (SUMO-1) activity in endometrial carcinoma cells, driving EC proliferation and invasion.24 Notably, it has been shown that the serum levels of total soluble CD38 molecules were significantly elevated in EC and were associated with low survival rates.25,26 CD38 expression showed a high correlation with COX-2, and inhibiting COX-2 markedly reduced the differentiation of human B cells into CD38+ IgM and CD38+ IgG- secreting cells.27 This evidence further confirms the potential association between elevated COX-2 expression and adverse prognosis in EC. In our study, we observed that elevated Eicosenedioate levels may increase the risk of EC, mediated through CD38 on PB PC. This finding aligns with existing evidence indicating that Eicosenedioate acts as a pro-EC factor. Its downstream COX-2 pathway has been shown to drive EC inflammation, angiogenesis and proliferative invasion. This pathway is also directly associated with CD38-positive cell differentiation and patient prognosis. Regarding the 5-LOX pathway, current studies on its relationship with EC are limited, but there are reports showing that elevated 5-LOX expression correlates with adverse EC prognosis.22 5-LOX is typically absent in normal epithelial tissues; however, its expression can be induced by proinflammatory stimuli and it may be overexpressed in various epithelial carcinomas. The expression of 5-LOX mRNA was significantly elevated in type II endometrial carcinoma compared to normal endometrial tissue. The proportion of its product, 5-hydroxyeicosatetraenoic acid (5-HETE), was also increased. Elevated 5-LOX mRNA expression was associated with poor outcomes. This may be another potential pathway for this metabolite to induce EC. However, the mechanism by which 5-LOX affects EC through immune cells still requires further investigation. In conclusion, the increase in EC risk by eicosenedioate may be related to the influence of the COX-2 and 5-LOX pathways and their related products on immune cells CD38. However, the precise mechanism underlying eicosenedioate’s interaction with EC requires further investigation.
GPC, a key intermediate metabolite in glycerophospholipid metabolism, may influence tumor progression. Eicosenedioate, a crucial component of cellular membrane glycerophospholipids, suggests a potential correlation between glycerophospholipid metabolism and the incidence of EC. Empirical evidence indicates that glycerophospholipids facilitate cell proliferation, tumorigenesis, and cancer survival, with an observed upregulation of glycerophospholipids in EC, thereby reinforcing this hypothesis.9 However, the precise relationship among GPC, eicosenedioate, and glycerophospholipid metabolism in the context of EC requires further experimental elucidation.
Presently, some studies have been executed to delineate the potential association between EC and diverse exposure factors utilizing MR analysis.28 By positing immune cells as intermediaries that modulate the potential association between glycerophospholipid metabolites and EC, a rigorously defined and statistically robust TSMR integrated with a two-step MR approach has elucidated the specific mechanisms through which glycerophospholipid metabolites influence EC. This method has also identified the crucial role of immune cells in this process and effectively avoided the confounding factor interference and reverse causality existing in the observational experiments. Nevertheless, there are some limitations to this study. Initially, concentrating on a European-descended population might constrain the extrapolation of the findings to diverse racial and ethnic populations. Secondly, the absence of patient demographic data in the GWAS clinical trial dataset precluded a thorough examination of the influence of age and gender on the observed associations. Lastly, notwithstanding the exclusion of potential confounding factors, the determination of association strength thresholds remains somewhat arbitrary, with the risk that overly strict or lenient criteria could introduce bias. Furthermore, although MR-Egger regression and MR-PRESSO analyses did not detect significant horizontal pleiotropy, the possibility of residual pleiotropic effects cannot be completely excluded. In addition, when the number of instrumental variables is limited, these sensitivity analyses may have reduced statistical power to detect potential violations. Future research should extend to multi-ethnic cohorts to validate these findings, integrate single-cell sequencing and metabolomics data to comprehensively analyze molecular interactions between immune cell subsets and metabolites, and confirm key pathways via in vitro experiments. These steps will further verify and assess the potential of these glycerophospholipid metabolites as cancer-preventive agents or lifestyle intervention targets.
Conclusion
This study concentrated on five glycerophospholipid metabolites and four immunophenotypes associated with EC risk. For the first time, the study quantified the proportion of immunophenotypes potentially involved in the glycerophospholipid metabolites–EC association pathway. This provides a theoretical foundation for non-invasive biomarker screening and immune-targeted therapy in EC. Nevertheless, additional research is necessary to elucidate the mechanisms underlying the relationship between glycerophospholipid metabolites and EC.
Data Sharing Statement
The dataset utilized in this study was obtained from the GWAS Catalog (https://www.ebi.ac.uk/gwas/), accessed on 6 February 2025. The GWAS summary statistics for serum metabolites are available from the GWAS Catalog (https://www.ebi.ac.uk/gwas/) under accession IDs GCST90199621–GCST90201020. The immunophenotype GWAS data are available from the IEU OpenGWAS database (https://gwas.mrcieu.ac.uk/), derived from the UK Biobank. The endometrial cancer GWAS data were obtained from the Endometrial Cancer Association Consortium (ECAC), with dataset ID GCST006464. All data used in this study are publicly available.
Ethics Statement
This study used only publicly available, de-identified GWAS summary statistics and did not involve human participant recruitment or identifiable personal data. According to Article 32, Items 1 and 2 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (China, February 18, 2023), ethical review is exempt for research using legally obtained public data that does not involve personal privacy. Therefore, this study was exempt from additional ethical approval.
Acknowledgments
Yichen Shen, Yijing Yan, and Peizhe Lin are co-first authors for this study. We extend our sincere gratitude to the authors of the original research and the reviewers for their invaluable time and generosity in providing a critical evaluation of this manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study received support from the Science and Technology Development Fund of the Tianjin Education Commission for Higher Education (No. 2023ZD024).
Disclosure
The authors declare that they have no discernible financial or personal interests that could have influenced the results of this research.
References
1. Zhang S, Gong TT, Liu FH, et al. Global, regional, and national burden of endometrial cancer, 1990-2017: results from the global burden of disease study. Front Oncol. 2019;9:1440. doi:10.3389/fonc.2019.01440
2. Bray F, Laversanne M, Sung H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024;74(3):229–17. doi:10.3322/caac.21834
3. Gaber C, Meza R, Ruterbusch JJ, Cote ML. Endometrial cancer trends by race and histology in the USA: projecting the number of new cases from 2015 to 2040. J Racial Ethn Health Disparities. 2016.
4. Crosbie EJ, Kitson SJ, McAlpine JN, Mukhopadhyay A, Powell ME, Singh N. Endometrial cancer. Lancet. 2022;399(10333):1412–1428. doi:10.1016/S0140-6736(22)00323-3
5. Raffone A, Troisi J, Boccia D, et al. Metabolomics in endometrial cancer diagnosis: a systematic review. Acta Obstet Gynecol Scand. 2020;99(9):1135–1146. doi:10.1111/aogs.13847
6. Wu Z, Nie Y, Kong D, et al. Lipid-metabolism-related gene signature predicts prognosis and immune microenvironment alterations in endometrial cancer. Biomedicines. 2025;13(5):1050. doi:10.3390/biomedicines13051050
7. Yang QJ, Zhao JR, Hao J, et al. Serum and urine metabolomics study reveals a distinct diagnostic model for cancer cachexia. J Cachexia, Sarcopenia Muscle. 2018;9(1):71–85. doi:10.1002/jcsm.12246
8. Troisi J, Mollo A, Lombardi M, et al. The metabolomic approach for the screening of endometrial cancer: validation from a large cohort of women scheduled for gynecological surgery. Biomolecules. 2022;12(9). doi:10.3390/biom12091229
9. Lorentzen GM, Laniewski P, Cui HY, et al. Cervicovaginal metabolome and tumor characteristics for endometrial cancer detection and risk stratification. Clin Cancer Res. 2024;30(14):3073–3087. doi:10.1158/1078-0432.CCR-23-2934
10. Sekula P, Greco MFD, Pattaro C, Kottgen A. Mendelian randomization as an approach to assess causality using observational data. J Am Soc Nephrol. 2016;27(11):3253–3265. doi:10.1681/ASN.2016010098
11. Wei C, Li C, Zhang G, Li H, Li J, Zeng J. Causality between immune cells, metabolites and breast cancer: mendelian randomization and mediation analysis. Biochem Genet. 2025;63(5):4790–4803. doi:10.1007/s10528-024-10966-4
12. Yang Q, Duan B, Yue J, et al. Causal effects and metabolites mediators between immune cell and risk of colorectal cancer: a Mendelian randomization study. Front Immunol. 2024;15:1444222. doi:10.3389/fimmu.2024.1444222
13. Zhou W, Ding J, Shen Y, Huang F, Xu D. Plasma metabolites mediate the effects of circulating immune cell phenotypes on prostate cancer: a 2-step Mendelian randomization study. Medicine. 2025;104(36):e44282. doi:10.1097/MD.0000000000044282
14. Glunde K, Bhujwalla ZM, Ronen SM. Choline metabolism in malignant transformation. Nat Rev Cancer. 2011;11(12):835–848. doi:10.1038/nrc3162
15. Sonkar K, Ayyappan V, Tressler CM, et al. Focus on the glycerophosphocholine pathway in choline phospholipid metabolism of cancer. NMR Biomed. 2019;32(10):e4112. doi:10.1002/nbm.4112
16. Bi Y, Ying X, Chen W, et al. Glycerophospholipid-driven lipid metabolic reprogramming as a common key mechanism in the progression of human primary hepatocellular carcinoma and cholangiocarcinoma. Lipids Health Dis. 2024;23(1):326. doi:10.1186/s12944-024-02298-4
17. Moestue SA, Giskeodegard GF, Cao MD, Bathen TF, Gribbestad IS. Glycerophosphocholine (GPC) is a poorly understood biomarker in breast cancer. Proc Natl Acad Sci U S A. 2012;109(38). doi:10.1073/pnas.1208226109
18. Yan D, Adeshakin AO, Xu M, et al. Lipid metabolic pathways confer the immunosuppressive function of myeloid-derived suppressor cells in tumor. Front Immunol. 2019;10:1399. doi:10.3389/fimmu.2019.01399
19. Al-Khami AA, Rodriguez PC, Ochoa AC. Metabolic reprogramming of myeloid-derived suppressor cells (MDSC) in cancer. Oncoimmunology. 2016;5(8):e1200771. doi:10.1080/2162402X.2016.1200771
20. Marshall AJ, Niiro H, Yun TJ, Clark EA. Regulation of B-cell activation and differentiation by the phosphatidylinositol 3-kinase and phospholipase Cγ pathway. Immunol Rev. 2000;176:30–46.
21. Peeters R, Jellusova J. Lipid metabolism in B cell biology. Mol Oncol. 2024;18(7):1795–1813. doi:10.1002/1878-0261.13560
22. Smith PG, Roque D, Ching MM, Fulton A, Rao G, Reader JC. The role of eicosanoids in gynecological malignancies. Front Pharmacol. 2020;11:1233. doi:10.3389/fphar.2020.01233
23. Wallace AE, Sales KJ, Catalano RD, et al. Prostaglandin F2alpha-F-prostanoid receptor signaling promotes neutrophil chemotaxis via chemokine (C-X-C motif) ligand 1 in endometrial adenocarcinoma. Cancer Res. 2009;69(14):5726–5733. doi:10.1158/0008-5472.CAN-09-0390
24. Sales KJ, List T, Boddy SC, et al. A novel angiogenic role for prostaglandin F2alpha-FP receptor interaction in human endometrial adenocarcinomas. Cancer Res. 2005;65(17):7707–7716. doi:10.1158/0008-5472.CAN-05-0101
25. Novikov VV, Mamaeva ME, Aliasova AV, et al. the serum level of total and oligomeric fractions of soluble molecules cd38 under malignant tumors of cervix and uterine body. Klin Lab Diagn. 2015;60(5):32–36.
26. Fan Y, Li X, Tian L, Wang J. Identification of a metabolism-related signature for the prediction of survival in endometrial cancer patients. Front Oncol. 2021;11:630905
27. Bernard MP, Phipps RP. Inhibition of cyclooxygenase-2 impairs the expression of essential plasma cell transcription factors and human B-lymphocyte differentiation. Immunology. 2010;129(1):87–96. doi:10.1111/j.1365-2567.2009.03152.x
28. Guo JZ, Wu QJ, Liu FH, Gao C, Gong TT, Li G. Review of mendelian randomization studies on endometrial cancer. Front Endocrinol. 2022;13:783150
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.