Back to Journals » OncoTargets and Therapy » Volume 11
Increased expression of antisense lncRNA SPINT1-AS1 predicts a poor prognosis in colorectal cancer and is negatively correlated with its sense transcript
Authors Li C, Li W ![]() , Zhang YL, Zhang X
, Zhang YL, Zhang X ![]() , Liu T
, Liu T ![]() , Zhang Y, Yang YM, Wang LL
, Zhang Y, Yang YM, Wang LL ![]() , Pan HW, Ji J, Wang CX
, Pan HW, Ji J, Wang CX ![]()
Received 29 January 2018
Accepted for publication 14 March 2018
Published 10 July 2018 Volume 2018:11 Pages 3969—3978
DOI https://doi.org/10.2147/OTT.S163883
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Carlos E Vigil
Chen Li,1,* Wei Li,2,* Yanli Zhang,3 Xin Zhang,2 Tong Liu,1 Yi Zhang,2 Yongmei Yang,2 Lili Wang,2 Hongwei Pan,2 Jian Ji,2 Chuanxin Wang1
1Department of Clinical Laboratory, The Second Hospital of Shandong University, Jinan 250033, China; 2Department of Clinical Laboratory, Qilu Hospital, Shandong University, Jinan, 250012, China; 3Department of Clinical Laboratory, Shandong Provincial Third Hospital, Jinan, 250031, China
*These authors contributed equally to this work
Purpose: Colorectal cancer (CRC) is a leading cause of cancer-associated mortality worldwide. Natural antisense transcripts (NATs) are pervasively expressed in human genome and have been confirmed to contribute to cancer progression. In our study, we aimed to investigate the expression and clinical pertinence of serine peptidase inhibitor, Kunitz type 1 antisense RNA1 (SPINT1-AS1) in CRC.
Materials and methods: The expression levels of SPINT1-AS1 and the corresponding sense transcript SPINT1 mRNA were analyzed in 150 pairs of CRC tissues and adjacent normal (AN) tissues, along with 45 pairs of preoperative and postoperative serum exosome samples by the strand-specific real-time quantitative polymerase chain reaction.
Results: Compared with AN tissues, the expression of SPINT1-AS1 was increased (P<0.001, 3.771 vs 0.980) in CRC tissues, while SPINT1 mRNA expression was decreased in CRC (P<0.001, 0.927 vs 1.165), and there was an obviously negative correlation between SPINT1-AS1 expression and its sense transcript (r=–0.701, P<0.001). SPINT1-AS1 yielded an area under the receiver operating characteristic curve value of 0.865 (95% confidence interval, 0.821–0.902) for discriminating CRC tissues from AN tissues. Moreover, high SPINT1-AS1 expression was correlated with regional lymph node metastasis (P<0.001), distant metastasis (P<0.001), and shorter relapse-free survival (RFS) time (P<0.001), and Cox regression analysis indicated that SPINT1-AS1 was an independent prognostic factor for RFS. Meanwhile, significant reduction of SPINT1-AS1 expression level (P=0.001) was observed in CRC serum exosomes after surgical resection.
Conclusion: SPINT1-AS1 is upregulated in CRC tissues and plays an essential role in CRC progression and prognosis. Thereby, SPINT1-AS1 may serve as a candidate prognostic biomarker and molecular therapy target for CRC.
Keywords: colorectal cancer, natural antisense transcripts, SPINT1-AS1, SPINT1 mRNA, prognosis
Introduction
Colorectal cancer (CRC) is the third-ranking cancer among males and the second among females worldwide, with an estimated 1.4 million new cases and 693,900 deaths per year.1 Despite the great progress made in CRC screening and therapeutic methods, the 5-year survival rate of CRC patients remains unsatisfactory. Until now, multiple genetic and epigenetic alterations have been confirmed to exert critical functions in CRC tumorigenesis and progression,2 whereas they are still underutilized for precise prediction of CRC course. Accordingly, it is of imminent importance to explore the appropriate molecular markers associated with clinical outcome and identify effective molecular targets for gene therapies.
Natural antisense transcripts (NATs) are transcribed from the DNA strands complementary to that of their endogenous sense counterparts.3 NATs, with limited protein-coding ability, exist ubiquitously in mammalian genome.4 They can regulate the expression of complementary sense transcripts through various mechanisms, such as epigenetic regulation of transcription, pre-mRNA alternative splicing, mRNA transport, mRNA stability, translation, generation of endogenous siRNA, and masking miRNA-binding sites.5 A growing body of research shows that NATs participate in multiple important biological processes, such as neurogenesis,6 female X-chromosome inactivation,7,8 and circadian clock function.9 Recently, NATs have been found to play important roles in the occurrence and development of various types of cancers. NAT KRT7-AS promotes gastric cancer cell proliferation and migration through stabilizing KRT7 mRNA by forming an RNA–RNA duplex.10 LCT13, antisense to TFPI-2, is associated with deposition of repressive histone marks and then reduces expression of in breast and colon cancer cell lines.11 Moreover, high gene locus-specific combinations of sense/antisense (S/AS) pairs make NATs promising precise therapeutic targets.12,13 However, among the enormous amount of non-coding RNAs (ncRNAs), NATs are less studied compared with other types of ncRNA, especially another subclass of long non-coding RNA (lncRNA), long-intergenic non-coding RNAs (lincRNAs);12,13 hence, the functional roles of the vast majority of NATs in CRC remain to be explored.
In the current study, we focused on one of the NATs termed serine peptidase inhibitor, Kunitz type 1 antisense RNA1 (SPINT1-AS1). Until now, the status of SPINT1-AS1 in CRC has not been explored; thus, we evaluated the expression of SPINT1-AS1 and analyzed the relationship with its corresponding sense transcript serine peptidase inhibitor, Kunitz type 1 (SPINT1) mRNA. The fact that SPINT1 has anti-cancer properties through its inhibitory effects on cell proliferation, migration, invasion, and decreased expression has been confirmed in breast cancer,14 ovarian cancer,15 cervical cancer,16 prostate cancer,17 gastric cancer,18 pancreatic cancer,19 and endometrial cancer.20 The same trend held true in CRC; the expression of SPINT1 obviously decreased in cancer cells compared with adjacent normal (AN) or adenoma cells.18,21 It has also been proved in mouse models that SPINT1 has a critical role as a suppressor of tumorigenesis in the intestinal tract.22 Then, we investigated the relationship between SPINT1-AS1 and clinicopathological characteristics and further confirmed whether its expression can be used for prognostic and therapeutic evaluation in CRC.
Materials and methods
Patients and sample collection
Written informed consent was obtained from all patients, and ethical approval for the study was obtained from the Ethics Committee of Qilu Hospital, Shandong University (protocol number: KYLL-2013-081). A total of 150 pairs of CRC tissues and AN tissues from patients who underwent radical surgical cancer resection were collected in this study between June 2009 and April 2011. CRC diagnosis was confirmed by pathological method, and noncancerous tissues were at least 5 cm away from the tumor margin. All tissues were snap-frozen in liquid nitrogen immediately after surgical resection and then preserved at −80°C until used for total RNA extraction. Venous blood samples were collected before and 7 days after surgical treatment from another 45 CRC patients. All blood samples were centrifuged at 3,000 rpm for 10 min to collect serum, followed by 110,000 rpm for 10 min to remove the remaining cell debris, and finally stored at −80°C until exosome extraction. Patients who received radiotherapy and chemotherapy before surgical treatment were excluded. The postoperative pathological staging was defined according to the tumor-node-metastasis grading system of the American Joint Committee on Cancer.
All patients enrolled in this study have been followed up at intervals of 3 months in the first 2 years and at 6 months’ interval up to the 5th year, and the follow-up was completed in July 30, 2016. Post-surgery follow-up was implemented through imageological examination, colonoscopy, pathological examination, and/or telephone follow-up. The overall survival (OS) time was defined as the period from diagnosis to death or the last follow-up date. The relapse-free survival (RFS) time was defined as the period from diagnosis to the date of relapse or the last follow-up date if no relapse.
RNA extraction and strand-specific reverse transcription quantitative polymerase chain reaction (RT-qPCR)
Total RNAs of the tissue samples were isolated by using TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA) in accordance with standard methodology. Exosomes were isolated by using ExoQuick Exosome Precipitation Solution (SBI, Mountain View, CA, USA), and exosomal RNA was extracted by using miRNeasy Mini Kit (Qiagen NV, Venlo, the Netherlands). Total RNAs from the tissue samples or exosomes were denatured at 65°C for 5 min and then reversely transcribed into cDNA with specific primers by using a TaKaRa Reverse Transcription Kit (TaKaRa, Dalian, China). DNase I was applied to treat total RNAs to remove potential chromosomal DNA contamination before reverse transcription. The conditions for reverse transcription were 37°C for 15 min and 85°C for 5 s. Then, qPCRs were performed for expression analysis on 10-fold diluted first-strand cDNA by using Power SYBR Green (TaKaRa). The amplification was performed as follows: 95°C for 30 s, 40 cycles of 95°C for 5 s, and 60°C for 34 s, and melting curve analysis. The expression of individual genes was normalized to the expression of two reference genes, GAPDH and UBC. The primer sequences used were listed in Table 1. The relative expressions of target genes were calculated by using the comparative delta-delta CT method (2−ΔΔCt).
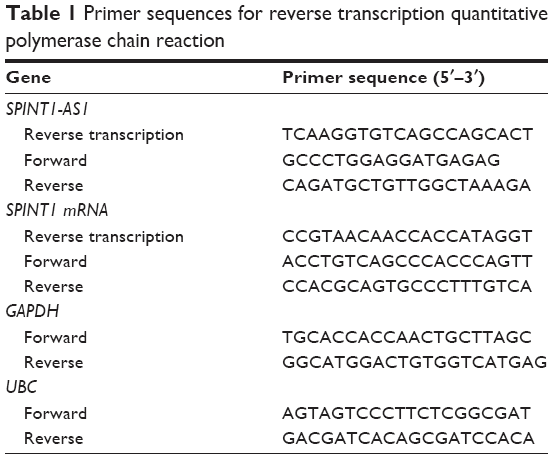 | Table 1 Primer sequences for reverse transcription quantitative polymerase chain reaction |
Dual luciferase reporter assay
Luciferase reporter gene vectors were constructed with SPINT1 mRNA 5′UTR sequences or mutant 5′UTR sequences in pGL3 plasmids (RiboBio, Guangzhou, China). The SPINT1-AS1 sequence was subcloned into the pcDNA3.1 vector (RiboBio), and an empty pcDNA3.1 vector was used as a control. SW480 cells were co-transfected with wide-type (or mutant-type) pGL3 plasmid and pcDNA3.1 plasmid containing SPINT1-AS1 sequence (or control pcDNA3 plasmid) by using Lipofectamine 2000 (Thermo Fisher Scientific). The activities of firefly luciferase and Renilla luciferase were measured, and firefly luciferase was standardized to the value of Renilla luciferase.
Statistical analysis
The data were analyzed by using SPSS 17.0 software (SPSS Inc., Chicago, IL, USA). Distribution of the data was determined by the Kolmogorov–Smirnov test. Data are presented as median (interquartile range). Expression differences between groups were analyzed by using Mann–Whitney U-test or Kruskal–Wallis test. Receiver operating characteristic (ROC) curve and area under the ROC curve (AUC) were employed to illustrate the performance of SPINT1-AS1 to discriminate CRC tissues from AN tissues. MedCalc 9.3.9.0 (MedCalc, Mariakerke, Belgium) was used to construct the ROC curve. OS and RFS curves were plotted by using the Kaplan–Meier method with the log-rank test performed to evaluate the statistical difference between the curves. Spearman’s correlation analysis was used to investigate the correlation between the expression of SPINT1-AS1 and SPINT1 mRNA. The hazard ratio (HR) and the 95% confidence interval (95% CI) were calculated by using univariate Cox proportional hazards regression. The final multivariate Cox regression was conducted with all covariates controlled. A P-value of <0.05 was considered statistically significant for all analyses.
Results
Expression of SPINT1-AS1 in CRC tissues
The result of lncRNA microarray analysis suggested that certain NATs were dysregulated in CRC tissues compared with AN tissues. Among these differentially expressed transcripts, SPINT1-AS1 was one of the upregulated NATs demonstrated by human genome-wide lncRNA microarray analysis performed on six pairs of CRC tissues and AN tissues (Figure 1A). By performing strand-specific RT-qPCR on these six pairs of tissues, a consistent result was presented, and the expression level of SPINT1-AS1 was increased in CRC tissues compared with AN tissues (P=0.015, Figure 1B). We further detected SPINT1-AS1 expression in 150 pairs of CRC tissues and AN tissues. As shown in Figure 1C, the expression level of SPINT1-AS1 was significantly upregulated in CRC tissues compared with AN tissues (P<0.001, Figure 1C) and was upregulated more than twofold in 116 of the 150 CRC tissues (Figure 1D). The ROC curve analysis illustrated that the expression level of SPINT1-AS1 was able to distinguish CRC tissues from AN tissues, with an AUC value of 0.865 (95% CI, 0.821–0.902, Figure 1E).
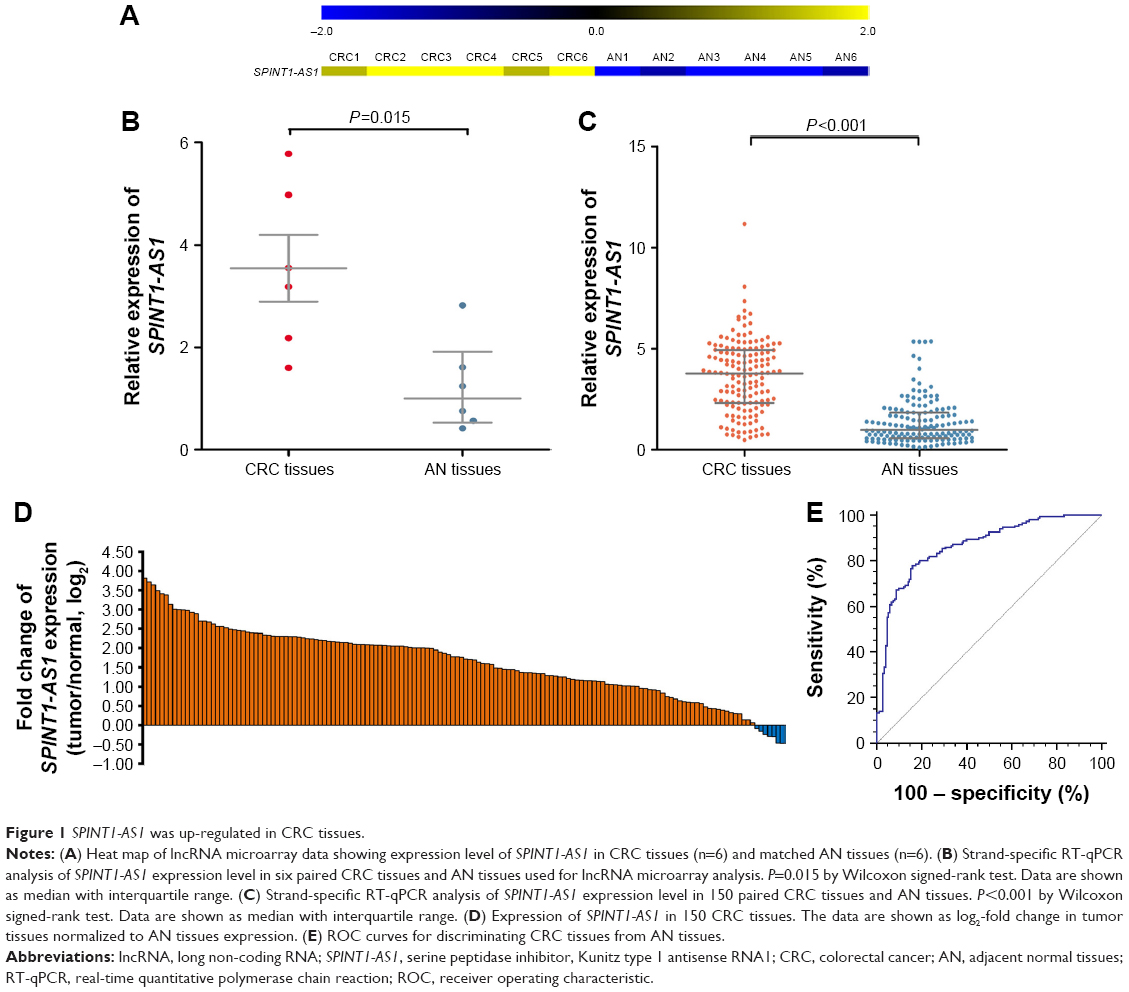 | Figure 1 SPINT1-AS1 was up-regulated in CRC tissues. |
Correlation between SPINT1-AS1 and its sense transcript
Information from the National Center for Biotechnology Information Gene database (http://www.ncbi.nlm.nih.gov/gene) shows SPINT1-AS1 and SPINT1 mRNA overlap at the 5′UTR in a head-to-head manner (Figure 2A). To study the relationship between SPINT1-AS1 and SPINT1 mRNA, we also assessed the expression of SPINT1 mRNA in the same 150 pairs of CRC tissues and AN tissues. SPINT1 mRNA expression level was significantly downregulated in CRC tissues compared with AN tissues (P<0.001, Figure 2B). Correlation analysis was performed, and the Spearman’s correlation coefficients showed a statistically negative correlation between SPINT1-AS1 and SPINT1 mRNA expression in CRC tissue samples (r=−0.701, P<0.001, Figure 2C). To test whether the regulation between SPINT1-AS1 and SPINT1 mRNA was mediated through their overlapping 5′UTR, we performed dual luciferase reporter assay, and the result showed that SPINT1-AS1 overexpression decreased the overlapping region of luciferase activity (P=0.020), while mutant overlapping sequence (Figure 2D) could abrogate this effect (Figure 2E).
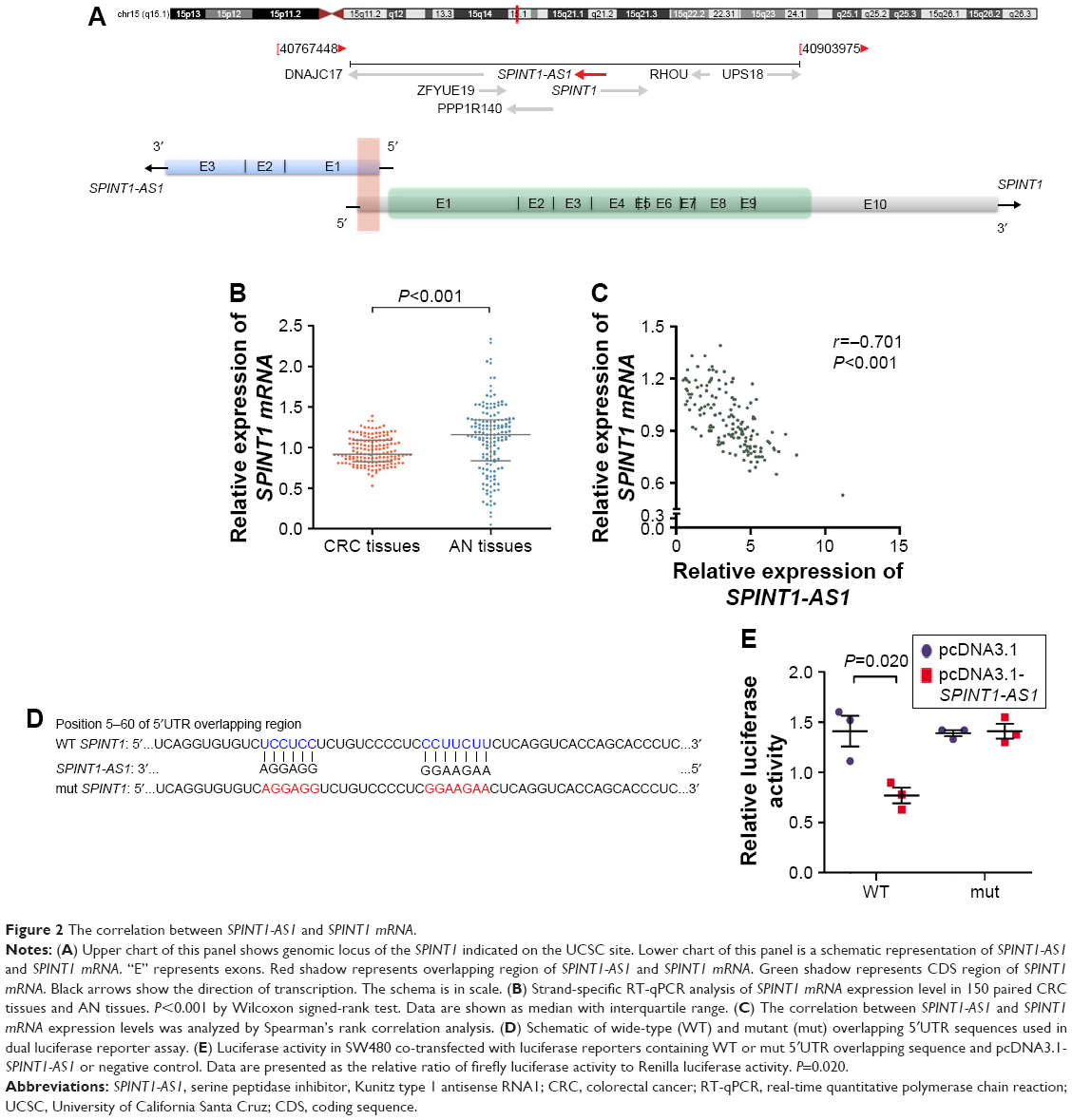 | Figure 2 The correlation between SPINT1-AS1 and SPINT1 mRNA. |
Relationship between SPINT1-AS1 expression and clinicopathological characteristics in CRC
To determine the correlation between SPINT1-AS1 and the progression of CRC, we analyzed whether SPINT1-AS1 expression was associated with clinicopathological features of CRC patients. As presented in Table 2, high SPINT1-AS1 expression level was significantly correlated with adverse clinicopathological characteristics of CRC, including regional lymph node metastasis (P<0.001, Mann–Whitney U-test) and distant metastasis (P<0.001, Mann–Whitney U-test). However, no significant relevance was found between SPINT1-AS1 expression level and other clinicopathological factors such as age (P=0.747, Mann–Whitney U-test), gender (P=0.218, Mann–Whitney U-test), tumor location (P=0.959, Mann–Whitney U-test), tumor differentiation (P=0.647, Kruskal–Wallis test), T stage (P=0.430, Mann–Whitney U-test), and tumor size (P=0.410, Mann–Whitney U-test).
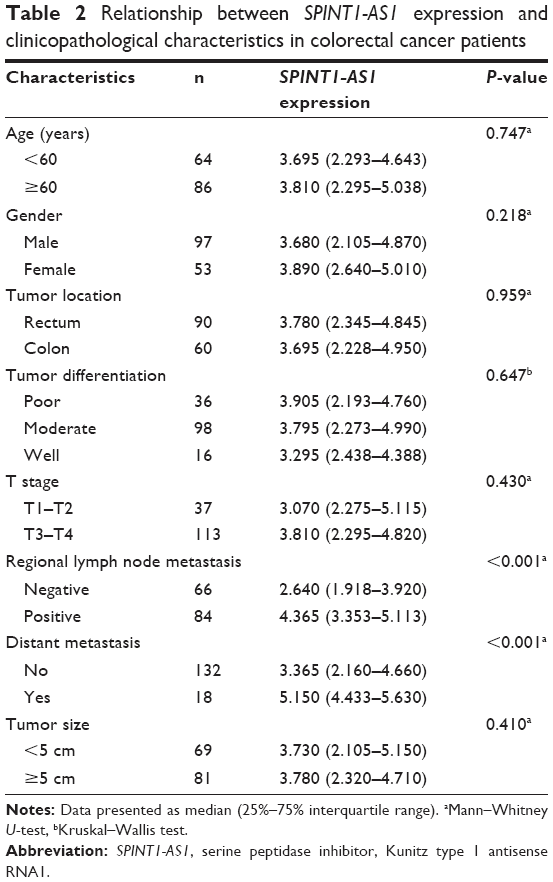 | Table 2 Relationship between SPINT1-AS1 expression and clinicopathological characteristics in colorectal cancer patients |
The prognostic value of SPINT1-AS1 expression for CRC patients
To determine the prognostic value of SPINT1-AS1, survival analysis was performed in the 150 CRC patients by using the Kaplan–Meier method along with log-rank test. According to the median value of SPINT1-AS1 expression level, patients with CRC were categorized into high-expression group and low-expression group. The results revealed that SPINT1-AS1 expression level had little to do with the OS time (P=0.152, HR =1.717; 95% CI, 0.811–3.637, Figure 3A), whereas high SPINT1-AS1 expression level was a significant prognostic factor for a shorter RFS time (P<0.001, HR =4.325; 95% CI, 2.204–8.489, Figure 3B). We further estimated the significance of several clinical parameters that might influence RFS in the subjects, and the univariate Cox proportional hazards regression model analysis showed that decreased RFS was correlated with advanced T stage classification (HR =2.259; 95% CI, 1.013–5.036; P=0.046), regional lymph node metastasis (HR =3.657; 95% CI, 1.820–7.348; P<0.001), distant metastasis (HR =5.606; 95% CI, 3.010–10.439; P<0.001), and high expression level of SPINT1-AS1 (HR =4.355; 95% CI, 2.219–8.549; P<0.001). Further multivariate Cox analysis revealed that the expression level of SPINT1-AS1 (HR =2.519; 95% CI, 1.180–5.378; P=0.017), along with distant metastasis (HR =2.770; 95% CI, 1.398–5.489; P=0.003) could act as independent predictors of RFS (Table 3).
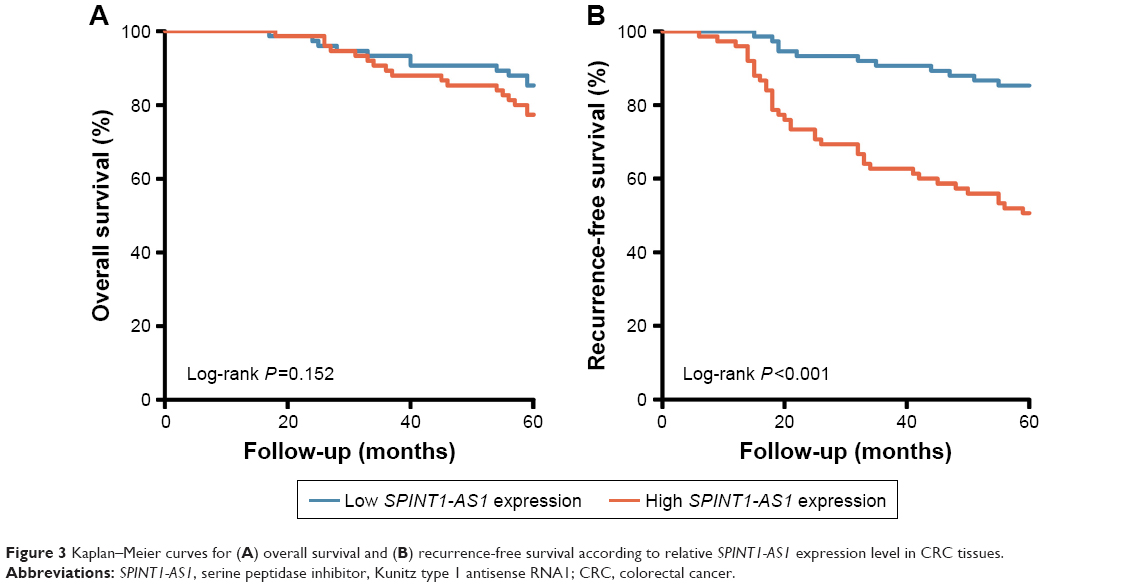 | Figure 3 Kaplan–Meier curves for (A) overall survival and (B) recurrence-free survival according to relative SPINT1-AS1 expression level in CRC tissues. |
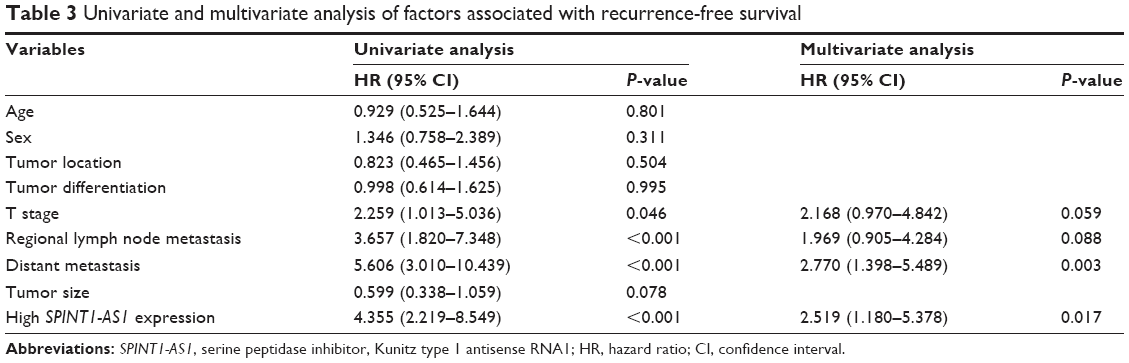 | Table 3 Univariate and multivariate analysis of factors associated with recurrence-free survival |
SPINT1-AS1 expression level in CRC serum exosomes
To further consolidate the vital function of this S/AS pair, SPINT1-AS1 and SPINT1 mRNA expression levels in serum exosomes were measured in 45 pairs of preoperative and postoperative serum samples. Decreased SPINT1-AS1 expression was observed after surgery (P=0.001, Figure 4A), while SPINT1 mRNA expression showed no comparable change (P=0.062, Figure 4B). These results suggested a promising role of SPINT1-AS1 in CRC monitoring and prompted us to focus on SPINT1-AS1 in the follow-up study.
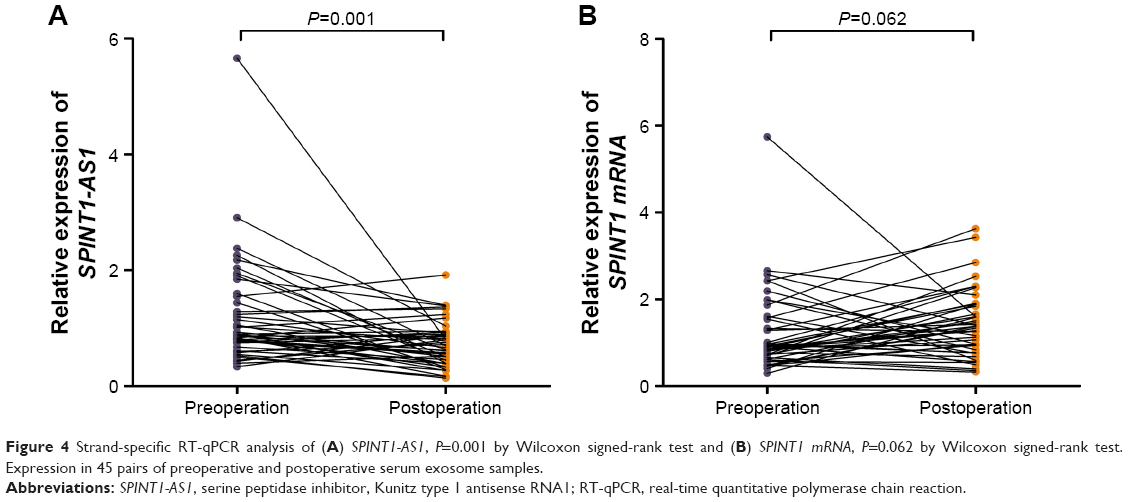 | Figure 4 Strand-specific RT-qPCR analysis of (A) SPINT1-AS1, P=0.001 by Wilcoxon signed-rank test and (B) SPINT1 mRNA, P=0.062 by Wilcoxon signed-rank test. Expression in 45 pairs of preoperative and postoperative serum exosome samples. |
Discussion
In this study, NAT, SPINT1-AS1 was first found in CRC patients by genome-wide lncRNA microarray and strand-specific RT-qPCR methods. And, its expression was negatively correlated with its sense transcript expression. Based on the analysis of a large number of specimens, SPINT1-AS1 showed an increased expression in CRC tissues and had a function to differentiate CRC from AN tissues. Moreover, high SPINT1-AS1 expression level was significantly correlated with regional lymph node metastasis, distant metastasis, and short RFS. Meanwhile, exosomal SPINT1-AS1 in serum of CRC patients might be used as a noninvasive biomarker for therapeutic evaluation.
Recently, growing evidences suggests that dysregulation of lncRNAs is involved in the complexity of the CRC process by influencing various aspects of cell biology, including proliferation, differentiation, apoptosis, angiogenensis, epithelial–mesenchymal transition, invasion, and metastasis.23,24 Certain lncRNAs are uniquely expressed in CRC, and their expression levels are associated with CRC state; therefore, lncRNAs have potential clinical application as biomarkers for diagnosis, prognosis, or as therapeutic targets.23,24 For these reasons, intensive investigations in recent years have focused on the role of lncRNAs in the pathogenesis of CRC. In our study, strand-specific RT-qPCR performed with specific reverse transcription primers was implemented to eliminate detection interference caused by complementary strands. The results of the quantitative studies showed that SPINT1-AS1 expression was significantly increased in CRC tissues compared with AN tissues while SPINT1 mRNA expression was downregulated.
Over the past few years, mounting evidence pointed to the significance of NATs in regulating gene expression at multiple levels, including epigenetic modifications and transcriptional and post-transcriptional regulation. BDNF-AS serves a critical role in the guidance, introduction, and maintenance of H3K27met3 at the BDNF locus by recruiting polycomb repressive complex 2 to the BDNF promoter region.25 Lrp1-AS directly combines with high mobility group box 2 (Hmgb2) and blocks the function of Hmgb2 to enhance Srebp1a-dependent transcription of Lrp1.26 FOXM1-AS regulates FOXM1 post-transcriptionally by forming RNA hybridization to increase FOXM1 pre-mRNA stability. FOXM1-AS elevates FOXM1 expression via carrying ALKBH5 to weaken N6-methyladenosine (m6A) RNA modification of FOXM1 pre-mRNA.27 Based on the site of biogenesis, NATs are separated into cis-NATs or trans-NATs. Cis-NATs are produced from opposite DNA strands on the same genomic loci as their sense counterparts, whereas trans-NATs are transcripts originating from different genomic loci.28,29 Cis-NATs prevailingly form long perfect complementary regions with a specific sense transcript from the same generation locus, while trans-NATs usually form short imperfect matches with several sense transcripts from different genomic loci.28 Cis-NAT pairs are further classified as head-to-head (5′-regions overlap), tail-to-tail (3′-regions overlap), embedded (one transcript is fully contained within the other), or intronic pairs.30 SPINT1-AS1 is transcribed from the reverse strand of the SPINT1 locus, located at 15q15.1, and consists of three exons. The sense transcript SPINT1 mRNA is transcribed from the forward strand of the same locus and consists of 10 exons. The 5′UTR of SPINT1-AS1 directly overlaps the 5′UTR of SPINT1 mRNA by 80 bp. In our study, a negative correlation between SPINT1-AS1 and SPINT1 mRNA expression in CRC tissues was observed. Dual luciferase reporter assay showed decreased SPINT1 mRNA 5′UTR luciferase activity in SPINT1-AS1 overexpression cells. These phenomena may indicate a regulatory role of SPINT1-AS1 to SPINT1 mRNA.
Serine peptidase inhibitor, Kunitz type 1 (SPINT1), also known as hepatocyte growth factor (HGF) activator (HGFA) inhibitor type 1 (HAI-1), encoded by the SPINT1 gene is a specific cellular inhibitor of active HGFA.31,32 HGFA is the most potent proteinase to activate HGF which is secreted in an inactive single-chain precursor form.33 The activation step of HGF is a critical limiting step in the HGF-induced signaling pathway.34 To date, a growing amount of evidence has accumulated in favor of the notion that HGF gets involved in the invasive growth of tumor cells through a sequence of steps, including dissociation of the tumor cells from their neighbors, migration through the extracellular matrix, and proliferation.33 The HGFA/HAI-1 ratio is of great significance to the regulation of HGF activity, and the unbalanced ratio is related to tumorigenesis.35 Clinical implications of SPINT1-AS1 in CRC were demonstrated in our study. SPINT1-AS1 might participate in tumorigenesis and progression of CRC, as SPINT1-AS1 expression level in CRC tissues was significantly related to regional lymph node metastasis and distant metastasis. According to the Kaplan–Meier curve, patients with high SPINT1-AS1 expression level had a markedly higher tumor recurrence rate than those with low level. Moreover, Cox proportional hazards regression model indicated that elevated SPINT1-AS1 level was an independent prognostic factor of CRC.
It is of interest to note that besides differential expression of SPINT1-AS1 between CRC tissues and AN tissues, dynamic expression was also observed in pre- and postoperative serum exosomes. SPINT1-AS1 expression level significantly declined after surgical resection. This observation positions SPINT1-AS1 as a potential noninvasive biomarker for monitoring changes in the course of CRC treatment.
In the current study, several obstacles are unclear. Although SPINT1-AS1 expression level is not closely tied with OS, we are not sure if this is due to the limited follow-up time. Whether SPINT1-AS1 could serve as a noninvasive biomarker for early diagnosis needs more empirical evidence for further confirmation. CRC screening can greatly reduce CRC-related mortality. Improvement of screening methods has become a main point of concern, such as fecal occult blood testing has been replaced by fecal immunochemical testing (FIT) to increase sensitivity36,37 and FIT combined with microbiome constituents may improve the screening results.38 In our future study, we could measure SPINT1-AS1 level in feces of CRC patients and controls and analyze the microbial diversity and composition to explore the correlation between SPINT1-AS1 level and the dysbiosis of gut microbiota. Although the regulative role of SPINT1-AS1 on SPINT1 mRNA expression level has been confirmed in our study, whether SPINT1-AS1 could regulate the alternative splicing of SPINT1 mRNA warrants further study. Previous study has demonstrated that antisense lncRNA TPM1-AS could regulate the alternative splicing of TPM1 through an interaction with RNA-binding mofit protein 4 (RBM4).39 The online RNA-binding protein (RBP) database RBPDB predicts that both SPINT1-AS1 and SPINT1 pre-mRNA have binding sites of RBM4. Thus, there is a possibility that SPINT1-AS1 regulates the process of CRC by changing the expression level and influencing the alternative splicing of SPINT1 mRNA simultaneously.
Conclusion
Our study evaluated the expression pattern of SPINT1-AS1 in the context of CRC and demonstrated that upregulated SPINT1-AS1 expression was a common event in CRC. High expression of SPINT1-AS1 is associated with poor clinical outcome and SPINT1-AS1 is a promising biomarker for the prognostic prediction and treatment monitoring of CRC patients.
Acknowledgments
This study was supported by National Natural Science Foundation of China (81472025, 81772271, 81401709, 81501819, 81702066), Shandong Key Research and Development Program (2016CYJS01A02, 2015GSF118167), Fundamental Research Funds of Shandong University (2014QLKY03), Shandong Provincial Natural Science Foundation (ZR2015PH049, ZR2014HP049), and Taishan Scholar Foundation (ts201511094).
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel RL, Miller KD, Fedewa SA, et al. Colorectal cancer statistics, 2017. CA Cancer J Clin. 2017;67(3):177–193. | ||
Yamauchi M, Lochhead P, Morikawa T, et al. Colorectal cancer: a tale of two sides or a continuum? Gut. 2012;61(6):794–797. | ||
Pelechano V, Steinmetz LM. Gene regulation by antisense transcription. Nat Rev Genet. 2013;14(12):880–893. | ||
Chen J, Sun M, Kent WJ, et al. Over 20% of human transcripts might form sense-antisense pairs. Nucleic Acids Res. 2004;32(16):4812–4820. | ||
Faghihi MA, Wahlestedt C. Regulatory roles of natural antisense transcripts. Nat Rev Mol Cell Biol. 2009;10(9):637–643. | ||
Fatica A, Bozzoni I. Long non-coding RNAs: new players in cell differentiation and development. Nat Rev Genet. 2014;15(1):7–21. | ||
Maclary E, Buttigieg E, Hinten M, et al. Differentiation-dependent requirement of Tsix long non-coding RNA in imprinted X-chromosome inactivation. Nat Commun. 2014;5:4209. | ||
Gayen S, Maclary E, Buttigieg E, Hinten M, Kalantry S. A primary role for the Tsix lncRNA in maintaining random X-chromosome inactivation. Cell Rep. 2015;11(8):1251–1265. | ||
Xue Z, Ye Q, Anson SR, et al. Transcriptional interference by antisense RNA is required for circadian clock function. Nature. 2014;514(7524):650–653. | ||
Huang B, Song JH, Cheng Y, et al. Long non-coding antisense RNA KRT7-AS is activated in gastric cancers and supports cancer cell progression by increasing KRT7 expression. Oncogene. 2016;35(37):4927–4936. | ||
Cruickshanks HA, Vafadar-Isfahani N, Dunican DS, et al. Expression of a large LINE-1-driven antisense RNA is linked to epigenetic silencing of the metastasis suppressor gene TFPI-2 in cancer. Nucleic Acids Res. 2013;41(14):6857–6869. | ||
Piatek MJ, Henderson V, Zynad HS, Werner A. Natural antisense transcription from a comparative perspective. Genomics. 2016;108(2):56–63. | ||
Khorkova O, Myers AJ, Hsiao J, Wahlestedt C. Natural antisense transcripts. Hum Mol Genet. 2014;23(R1):R54–R63. | ||
Parr C, Jiang WG. Hepatocyte growth factor activation inhibitors (HAI-1 and HAI-2) regulate HGF-induced invasion of human breast cancer cells. Int J Cancer. 2006;119(5):1176–1183. | ||
Oberst MD, Johnson MD, Dickson RB. Expression of the serine protease matriptase and its inhibitor HAI-1 in epithelial ovarian cancer: correlation with clinical outcome and tumor clinicopathological parameters. Clin Cancer Res. 2002;8(4):1101–1107. | ||
Nakamura K, Abarzua F, Hongo A, et al. The role of hepatocyte growth factor activator inhibitor-1 (HAI-1) as a prognostic indicator in cervical cancer. Int J Oncol. 2009;35(2):239–248. | ||
Saleem M, Adhami VM, Zhong W, et al. A novel biomarker for staging human prostate adenocarcinoma: overexpression of matriptase with concomitant loss of its inhibitor, hepatocyte growth factor activator inhibitor-1. Cancer Epidemiol Biomarkers Prev. 2006;15(2):217–227. | ||
Zeng L, Cao J, Zhang X. Expression of serine protease SNC19/matriptase and its inhibitor hepatocyte growth factor activator inhibitor type 1 in normal and malignant tissues of gastrointestinal tract. World J Gastroenterol. 2005;11(39):6202–6207. | ||
Ye J, Kawaguchi M, Haruyama Y, et al. Loss of hepatocyte growth factor activator inhibitor type 1 participates in metastatic spreading of human pancreatic cancer cells in a mouse orthotopic transplantation model. Cancer Sci. 2014;105(1):44–51. | ||
Nakamura K, Hongo A, Kodama J, Hiramatsu Y. The role of hepatocyte growth factor activator inhibitor (HAI)-1 and HAI-2 in endometrial cancer. Int J Cancer. 2011;128(11):2613–2624. | ||
Kataoka H, Hamasuna R, Itoh H, Kitamura N, Koono M. Activation of hepatocyte growth factor/scatter factor in colorectal carcinoma. Cancer Res. 2000;60(21):6148–6159. | ||
Hoshiko S, Kawaguchi M, Fukushima T, et al. Hepatocyte growth factor activator inhibitor type 1 is a suppressor of intestinal tumorigenesis. Cancer Res. 2013;73(8):2659–2670. | ||
Xie X, Tang B, Xiao Y, et al. Long non-coding RNAs in colorectal cancer. Oncotarget. 2015;7(5):5226–5239. | ||
Yang Y, Zhao L, Lei L, et al. LncRNAs: the bridge linking RNA and colorectal cancer. Oncotarget. 2017;8(7):12517–12532. | ||
Modarresi F, Faghihi MA, Lopez-Toledano MA, et al. Inhibition of natural antisense transcripts in vivo results in gene-specific transcriptional upregulation. Nat Biotechnol. 2012;30(5):453–459. | ||
Yamanaka Y, Faghihi MA, Magistri M, Alvarez-Garcia O, Lotz M, Wahlestedt C. Antisense RNA controls LRP1 Sense transcript expression through interaction with a chromatin-associated protein, HMGB2. Cell Rep. 2015;11(6):967–976. | ||
Zhang S, Zhao BS, Zhou A, et al. m6A demethylase ALKBH5 maintains tumorigenicity of glioblastoma stem-like cells by sustaining FOXM1 expression and cell proliferation program. Cancer Cell. 2017; 31(4):591–606.e6. | ||
Zhang X, Lii Y, Wu Z, et al. Mechanisms of small RNA generation from cis-NATs in response to environmental and developmental cues. Mol Plant. 2013;6(3):704–715. | ||
Rosikiewicz W, Makalowska I. Biological functions of natural antisense transcripts. Acta Biochim Pol. 2016;63(4):665–673. | ||
Balbin OA, Malik R, Dhanasekaran SM, et al. The landscape of antisense gene expression in human cancers. Genome Res. 2015;25(7):1068–1079. | ||
Shimomura T, Denda K, Kitamura A, et al. Hepatocyte growth factor activator inhibitor, a novel Kunitz-type serine protease inhibitor. J Biol Chem. 1997;272(10):6370–6376. | ||
Kataoka H, Kawaguchi M. Hepatocyte growth factor activator (HGFA): pathophysiological functions in vivo. FEBS J. 2010;277(10):2230–2237. | ||
Kataoka H, Miyata S, Uchinokura S, Itoh H. Roles of hepatocyte growth factor (HGF) activator and HGF activator inhibitor in the pericellular activation of HGFscatter factor. Cancer Metastasis Rev. 2003;22(2–3):223–236. | ||
Gak E, Taylor WG, Chan AM, Rubin JS. Processing of hepatocyte growth factor to the heterodimeric form is required for biological activity. FEBS Lett. 1992;311(1):17–21. | ||
Zheng Q, Wu H, Cao J, Ye J. Hepatocyte growth factor activator inhibitor type1 in cancer: advances and perspectives (review). Mol Med Rep. 2014;10(6):2779–2785. | ||
Kuipers EJ. Colorectal cancer: screening-one small step for mankind, one giant leap for man. Nat Rev Clin Oncol. 2014;11(1):5–6. | ||
Grobbee EJ, Schreuders EH, Hansen BE, et al. Association between concentrations of hemoglobin determined by fecal immunochemical tests and long-term development of advanced colorectal neoplasia. Gastroenterology. 2017;153(5):1251–1259.e2. | ||
Li S, Konstantinov SR, Smits R, Peppelenbosch MP. Bacterial biofilms in colorectal cancer initiation and progression. Trends Mol Med. 2017;23(1):18–30. | ||
Huang GW, Zhang YL, Liao LD, Li EM, Xu LY. Natural antisense transcript TPM1-AS regulates the alternative splicing of tropomyosin I through an interaction with RNA-binding motif protein 4. Int J Biochem Cell Biol. 2017;90:59–67. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.