Back to Journals » International Journal of Nanomedicine » Volume 14
In vitro comparison of liposomal drug delivery systems targeting the oxytocin receptor: a potential novel treatment for obstetric complications
Received 13 December 2018
Accepted for publication 9 February 2019
Published 27 March 2019 Volume 2019:14 Pages 2191—2206
DOI https://doi.org/10.2147/IJN.S198116
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Thomas Webster
Susan Hua,1,2 Benjamin Vaughan3
1Therapeutic Targeting Research Group, School of Biomedical Sciences and Pharmacy, University of Newcastle, Callaghan, NSW, Australia; 2Hunter Medical Research Institute, New Lambton Heights, NSW, Australia; 3Centre for Organic Electronics, University of Newcastle, Callaghan, NSW, Australia
Introduction: Targeted intervention to the uterus has great potential for the treatment of obstetric complications (eg, preterm birth, dysfunctional labor, and postpartum hemorrhage) by improving the effectiveness and safety of therapeutic compounds. In particular, targeting the oxytocin receptor (OTR) is a novel approach for drug delivery to the uterus. The aim of this study was to report the complete data set for the pharmaceutical synthesis and in vitro characterization of PEGylated liposomes conjugated with anti-OTR monoclonal antibodies (OTR-Lipo) or atosiban (ATO-Lipo, OTR antagonist).
Methods: OTR-targeted liposomal platforms composed of 1,2-distearoyl-sn-glycero-2-phosphocholine and cholesterol were prepared according to the method of dried lipid film hydration. Ligands were conjugated with the surface of liposomes using optimized methods to maximize conjugation efficiency. The liposomes were characterized for particle size, ligand conjugation, drug encapsulation, liposome stability, specificity of binding, cellular internalization, mechanistic pathway of cellular uptake, and cellular toxicity.
Results: Both OTR-Lipo and ATO-Lipo showed significant and specific binding to OTRs in a concentration-dependent manner compared to all control groups. There was no significant difference in binding values between OTR-Lipo and ATO-Lipo across all concentrations evaluated. In addition, OTR-Lipo (81.61%±7.84%) and ATO-Lipo (85.59%±8.28%) demonstrated significantly increased cellular internalization in comparison with rabbit IgG immunoliposomes (9.14%±1.71%) and conventional liposomes (4.09%±0.78%) at 2.02 mM phospholipid concentration. Cellular association following liposome incubation at 4.05 mM resulted in similar findings. Evaluation of the mechanistic pathway of cellular uptake indicated that they undergo internalization through both clathrin- and caveolin-mediated mechanisms. Furthermore, cellular toxicity studies have shown no significant effect of both liposomal platforms on cell viability.
Conclusion: This study further supports OTRs as a novel pharmaceutical target for drug delivery. OTR-targeted liposomal platforms may provide an effective way to deliver existing therapies directly to myometrial tissue and avoid adverse effects by circumventing non-target tissues.
Keywords: liposomes, nanoparticles, targeted drug delivery, oxytocin receptor, uterus, obstetric complications
Background
Targeted drug delivery of nanomedicines has shown to be beneficial for increasing the therapeutic index of compounds by improving drug targeting to specific sites of disease and/or attenuating localization in healthy non-target tissues. Obstetrics is an area of clinical medicine that has recently gained attention for drug targeting. Current treatment for obstetric complications (eg, preterm labor, dysfunctional labor, and postpartum hemorrhage) is challenging, as there are limited effective and safe therapeutic options available.1–5 Preterm labor (birth before 37 weeks of gestation) is the most important cause of perinatal morbidity and mortality,6–8 whereas postpartum hemorrhage represents the most important cause of maternal morbidity and mortality worldwide.9,10 In addition, up to one third of women may require induction of labor, primarily due to concerns for the health of the mother or fetus.11,12
The development of novel strategies to deliver therapeutic agents specifically to the uterus would address the clinical challenge of effectively managing these obstetric complications. Ligand-targeted nanomedicines have shown enormous potential for site-specific delivery of therapeutic compounds to designated cell types, which selectively express or overexpress specific receptors at the site of action. For obstetric complications, the smooth muscle layer of the uterus (myometrium) represents an ideal target for pharmacological interventions by controlling the contractility state of the uterus. In this way, therapeutic agents can be directed to the uterus to inhibit myometrial contractions (tocolysis) for the management of preterm labor or enhance myometrial contractions (uterotonic) for the management of dysfunctional labor and postpartum hemorrhage.13
More specifically, the oxytocin receptor (OTR) is considered one of the most important molecules in the myometrium and thus a promising target for drug delivery. OTR agonists and antagonists have been clinically used to modify the contractility of the uterus; however, the clinical efficacy of the available agents has been disappointing. For example, oxytocin was reported to be effective in ~50% of patients,12 and atosiban (ATO-Lipo, OTR antagonist) showed lack of efficacy over other conventional treatments and no evidence of improving neonatal outcomes.14,15 Despite this, OTR density in the uterus significantly increases with the progression of pregnancy.16,17 Low OTR expression has been reported in early gestation, but this rises significantly over the course of pregnancy to ~12-fold by 37–41 weeks.17 Maximal expression is seen after the onset of labor, which is assumed to mediate an increase in sensitivity of the myometrium to oxytocin at term.18,19 Importantly, OTR numbers in the myometrium have also been shown to be upregulated in preterm labor.16,17,20 Therefore, designing a drug delivery system to target this moiety would provide a logical means to enhance local drug delivery to the uterus.
We have recently developed a targeted drug delivery system for the uterus by conjugating anti-OTR polyclonal antibodies to a liposomal platform.21,22 Liposomes have the advantage of having flexible physicochemical and biophysical properties, which allow easy modifications to address different delivery considerations.23–26 The functionality of the drug delivery system was evaluated on murine and human myometrial tissues, as well as in vivo in a murine model of preterm labor.21 However, the polyclonal nature of the antibody posed a significant limitation to specificity of binding to OTRs.22 In order to improve the clinical translation of this technology, it is important to determine the effectiveness of other potential anti-OTR ligands for the targeted delivery system – in particular, anti-OTR monoclonal antibodies and atosiban (OTR antagonist). Therefore, the aim of this study was to report the complete data set for the pharmaceutical synthesis and characterization of PEGylated immunoliposomes conjugated with anti-OTR monoclonal antibodies (OTR-Lipo) and atosiban-conjugated PEGylated liposomes (ATO-Lipo), and compare their specific cellular interaction with OTR-expressing myometrial cells in vitro.
Materials and methods
Liposome-related materials
1,2-Distearoyl-sn-glycero-2-phosphocholine (DSPC), 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[maleimide (polyethylene glycol)-2000] (DSPE-PEG(2000) maleimide), and 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[carboxy (polyethylene glycol)-2000] (DSPE-PEG(2000) carboxylic acid) were purchased from Avanti Polar Lipids (Alabaster, AL, USA). N-Succinimidyl-3-(2-pyridyldithio) propionate (SPDP), cholesterol, atosiban, and tris(2-carboxyethyl) phosphine hydrochloride (TCEP) were from Sigma-Aldrich Co. (St Louis, MO, USA). 1-Ethyl-3-(3-dimethylaminopropyl)carbodiimide (EDC), CBQCA protein assay kit and N-hydroxysuccinimide (NHS) were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Rabbit IgG antibody and anti-OTR monoclonal antibody (ab181077) were obtained from Abcam (Cambridge, UK). PD-10 column was from VWR International (Radnor, PA, USA). All other chemicals and solvents were of at least analytical grade.
Cell-related materials
FBS, trypsin-EDTA (1:250), trypan blue solution 0.4%, dansylcadaverine (D4008), chlorpromazine hydrochloride (C8138), genistein (G6649), filipin (F9765), methyl-β-cyclodextrin (C4555), nocodazole (M1404), cytochalasin D (C8273), MTT-based assay kit (CGD1), and all buffer reagents were from Sigma-Aldrich Co. L-Glutamine, DMEM, Gibco® Antibiotic-Antimycotic (penicillin, streptomycin, amphotericin), sodium pyruvate, Alexa Fluor™ 488 conjugated cholera toxin subunit B (C34775), and Alexa Fluor 488 conjugated transferrin (T13342) were obtained from Thermo Fisher Scientific. Fluoresbrite® YG carboxylate microspheres (size 1 μm) was from Polysciences Inc. (Warrington, PA, USA). hTERT-immortalized myometrial (hTERT-myo) cells were obtained from Prof Roger Smith (University of Newcastle, Australia). All other reagents were of analytical grade. The POLARstar Optima™ fluorescent plate reader and its corresponding software were obtained from BMG Labtech (Ortenberg, Germany). The Institutional Biosafety Committee at the University of Newcastle approved all experiments and the use of the cell line.
Preparation of immunoliposomes
Immunoliposomes composed of DSPC and cholesterol (molar ratio 2:1) containing DSPE-PEG(2000) maleimide at 1.5 mol% of DSPC as a coupling lipid were prepared according to the method of dried lipid film hydration in PBS (pH 7.4) as previously described.27 DiI (fluorescent lipophilic dye) was added at 0.2 mol% of DSPC, and drugs were incorporated based on their hydrophilicity. The resulting multilamellar dispersions were reduced in size and lamellarity by ultrasonication at 60% amplitude for 5 minutes at 65°C. The activated liposome suspension was immediately mixed with thiolated antibody at room temperature. Thiolated antibodies were prepared by conjugating anti-OTR monoclonal antibodies (25 μg) or non-specific rabbit IgG (25 μg) with SPDP (6.25 mg/mL; SPDP/mAb molar ratio =10:1). PD-10 column equilibrated with distilled water was used to remove excess SPDP and fractions containing pyridyldithiopropionated-Ab (PDP-Ab) conjugates (assessed by absorbance at 280 nm) were lyophilized and stored at 4°C under nitrogen gas. PDP-Ab was reduced with 5 mM TCEP for 5 minutes to produce thiolated-Ab (Ab-SH), and absorbance was checked at 280 nm (protein concentration) and 343 nm (SPDP modification) to ensure stability of the compound. Thiolated antibody was mixed immediately with liposomes for 1 hour at room temperature with stirring in the dark. TLX ultracentrifugation (Optima™) was used to remove unconjugated Ab and non-encapsulated drug (100,000× g; 45 minutes). Liposomes were resuspended in PBS (pH 7.4) and stored at 4°C under nitrogen gas and in the dark, and were used within 2 weeks.
Preparation of atosiban-conjugated liposomes
Liposomes were composed of DSPC, cholesterol, and DSPE-PEG(2000) carboxylic acid at a molar ratio of 2:1:0.06 according to the method of dried lipid film hydration in MES buffer (pH 4.7). The fluorescent lipophilic dye DiI was added at 0.2 mol% of DSPC. Drugs were incorporated based on their hydrophilicity. The resulting multilamellar dispersions were reduced in size and lamellarity by ultrasonication at 60% amplitude for 5 minutes at 65°C. NHS and EDC (molar ratio 2:1) were then added to the liposome suspension and incubated for 15 minutes. Prior to the addition of atosiban, the pH of the liposome suspension was raised to 7.2–7.5 for optimal reaction with amine-containing molecules. The liposome suspension was incubated overnight at room temperature with stirring in the dark. Excess reagents were removed by TLX ultracentrifugation (Optima) (100,000× g; 45 minutes). Liposomes were resuspended in PBS (pH 7.4) and stored at 4°C under nitrogen gas and in the dark and were used within 2 weeks.
Liposome characterization
Size distribution of the liposomes was determined by dynamic laser light scattering (Zetasizer Nano ZS™; ATA Scientific, Taren Point, Australia). Phospholipid concentration was determined indirectly by measuring DiI concentration within liposomes. CBQCA protein assay was used to quantify the amount of antibody associated with the liposomes, using bovine serum albumin for the preparation of the standard curve. Atosiban concentration was evaluated using HPLC.
HPLC analysis
Drug concentration was determined by HPLC using an Agilent Technologies 1200 series HPLC system (Agilent Technologies, Santa Clara, CA, USA), consisting of an autoinjector, column oven, binary pump and UV-Vis detector. Data were integrated using Agilent Chemstation software. Separation was performed using a Thermo Scientific BDS Hypersil C18 column (150×4.6 mm, 5 μm). Drugs were dissolved in distilled water (hydrophilic drugs) or ethanol (hydrophobic drugs), with subsequent dilutions made in mobile phase for the calibration curves.
HPLC mobile phase and settings for nifedipine
Mobile phase consisted of 60% acetonitrile and 40% 10 mM KH2PO4 buffer (pH 2.5), which was pumped through the column at 1 mL/min. The column was maintained at 25°C and the wavelength of detection was 208 nm.
HPLC mobile phase and settings for salbutamol hemisulfate
Mobile phase consisted of 30% methanol and 70% 0.05 M KH2PO4 buffer (pH 3.5), which was pumped through the column at 1 mL/min. The column was maintained at 25°C and the wavelength of detection was 230 nm.
HPLC mobile phase and settings for atosiban
Mobile phase consisted of 30% acetonitrile and 70% 10 mM KH2PO4 (pH 2.5), which was pumped through the column at 1 mL/min. The column was maintained at 25°C and the wavelength of detection was 220 nm.
Atomic force microscopy (AFM)
AFM was performed on a Cypher Scanning Probe Microscope (Asylum Research; Oxford Instruments, Abingdon, UK). Commercial pyramidal silicon tips (TAPAl300-G; BudgetSensors, Sofia, Bulgaria) with a radius <10 nm, a resonance frequency of ~300 kHz and a nominal force constant of 40 (20–75) N/m were used. All measurements were performed in tapping mode to avoid damage of the sample surface. The scan speed was proportional to the scan size and the scan rate was between 1.95 and 2.44 Hz. The results were visualized in amplitude mode. Liposomes were directly transferred onto a silicon chip by dipping it into the liposome suspension. The silicon chip was then dried at room temperature before analysis.
Liposome stability assay
The dialysis technique was used to evaluate the in vitro stability of the liposomal formulations in PBS pH 7.4% and 50% FBS at 37°C over a 48 hours duration, as previously described.22 A modified assay was used to evaluate the true release of each drug from the liposomes without surpassing saturation point, which addressed the potential solubility issues across the dialysis membrane. Drug retention percentage was determined by: Drug retention (%) = 100% − [(Dt/D0) × 100%], where Dt and D0 indicate the amount of drug released from the liposomes at certain intervals and the total amount of drug in the liposome suspension, respectively. Liposome samples were collected at the end of the study and lysed with ethanol for analysis of drug content by HPLC.
Specificity of binding to OTRs on myometrial cells
Specificity of binding was evaluated on hTERT-myo cells as previously described.22 In brief, cells were seeded at 4×104 cells per well in 96-well tissue culture plates in complete medium (DMEM containing 1% L-glutamine, 1% sodium pyruvate, 1% Gibco Antibiotic-Antimycotic, 10% FBS) at 37°C in 5% CO2. At confluence, the plates were exchanged with serum and supplement-free DMEM to avoid binding of serum proteins to the liposome surfaces which might induce agglomeration. For competitive inhibition studies, cells were preincubated with a saturating concentration of anti-OTR monoclonal antibodies for 30 minutes. Liposomes were added to the cells for 1 hour at 4°C to determine exclusively cell binding.28,29 A fluorescent dye (DiI) was incorporated into the phospholipid bilayer as a marker for the liposomes. At the end of incubation, ice-cold PBS was used to wash the cells three times to remove unbound liposomes and fluorimetric detection (POLARstar Optima) was used to assess binding at excitation/emission wavelengths corresponding to DiI (549/565 nm). Background reading was assessed with cells incubated in medium alone.
Cellular uptake of OTR-targeted liposomes
Cellular association of OTR-targeted liposomes was assessed on hTERT-myo cells as previously described.22 In brief, cells were seeded in 96-well tissue culture plates (4×104 cells per well) and grown until confluent under the conditions described above. At confluence, the plates were exchanged with serum and supplement-free DMEM prior to the addition of fluorescent-labeled liposomes for 1 hour at 37°C. It should be noted that both receptor binding and internalization takes place at this temperature.28,29 At the end of incubation, ice-cold PBS was used to wash the cells three times before fluorimetric detection to assess extracellularly bound liposomes. Acid wash (0.1 M HCl for 10 minutes)30 was used to release surface-bound fluorescence before lysis of the cells with the addition of 1% Triton X-100 in PBS for 10 minutes at 37°C. Results from the acid wash were expressed as extracellularly bound fluorescence, whereas fluorescence in the lysis fractions was expressed as internalized fluorescence. Background reading was evaluated with solubilized cells without prior incubation with liposomes.
Endocytotic mechanisms of OTR-targeted liposomes
Mechanisms of cellular uptake were determined in the presence of specific inhibitory agents for different types of endocytosis, as previously described.22 In brief, hTERT-myo cells were preincubated with specific inhibitory agents for 30 minutes at 37°C and were then incubated with liposomes for 1 hour at 37°C. Chlorpromazine hydrochloride and monodansylcadaverine were used as inhibitiors of clathrin-mediated endocytosis, whereas genistein and filipin were used to inhibit caveolin-mediated endocytosis. Methyl-β-cyclodextrin is able to inhibit both clathrin- and caveolin-mediated endocytosis by depleting cholesterol. Nocodazole was used to inhibit macropinocytosis, and cytochalasin D was used to inhibit both phagocytosis and macropinocytosis. Positive controls were used together with specific inhibitors to investigate clathrin-mediated endocytosis (Alexa Fluor 488 conjugated transferrin at 120 μg/mL), caveolin-mediated endocytosis (Alexa Fluor 488 conjugated cholera toxin subunit B at 0.6 μg/mL), and phagocytosis and macropinocytosis (Fluoresbrite YG carboxylate microspheres of 1 μm diameter at 100 μg/mL). At the end of incubation, cells were washed three times with ice-cold PBS before fluorimetric detection as described above.
Cellular toxicity
Cell viability following exposure of hTERT-myo cells to liposomes and specific endocytotic inhibitors at various concentrations for 24 and 1.5 hour, respectively, was measured using an MTT-based assay, as previously described.22 Triton X-100 (1% in PBS) was used as a positive control. At the end of incubation, the medium was removed and MTT solution (5 mg/mL MTT in supplemented DMEM) was added to the cultures (final concentration of 0.5 mg/mL) and incubated for 4 hours at 37°C. The medium was then discarded and formazan crystals were solubilized using 150 μL dimethylsulfoxide for 15 minutes under light protection and at room temperature. Absorbance was measured at 550 nm in a microplate spectrophotometer (POLARstar Optima), which is directly proportional to cellular metabolism. Background reading was assessed with untreated cells.
Statistical analysis
All data are expressed as mean ± standard error of the mean (SEM) or SD. GraphPad Prism 7.01 software was used for statistical analysis. One-way ANOVA with Tukey’s multiple comparison test was used to evaluate differences between formulation groups or between time points (one independent variable). Comparisons between the different formulation groups over various concentrations were made using two-way ANOVA with Tukey’s multiple comparison test (two independent variables). Differences were considered significant when P<0.05.
Results
Dispersion properties of the liposome formulations
OTR-Lipo and ATO-Lipo had a mean particle size of 201±2.9 and 201±3.8 nm with a polydispersity index of 0.240±0.072 and 0.220±0.056, respectively. The size and polydispersity index of the control and drug-encapsulated liposome formulations were similar (Table 1). The choice of particle size was based on the results from our previous studies.21,22 The low polydispersity indexes indicate that the mean particle size is a reasonable indicator of the size of the majority of the particles in the dispersions. All liposome formulations had an approximate neutral net charge, as they were composed of neutral phospholipids. Both nifedipine-loaded liposomes and salbutamol-loaded liposomes had drug encapsulation efficiencies of >98%, which equated to ~4 mg of drug per milliliter of liposome suspension (Table 1). Mean atosiban coupling ratio for the ATO-Lipo was 5.970±0.232 μg of peptide per μmol of phospholipid, which equated to ~2,017±78.39 molecules per liposome. Mean antibody coupling ratio for the OTR-Lipo was 1.227±0.023 μg of antibody per μmol of phospholipid. With a starting antibody concentration of 25 μg and a phospholipid concentration of 2.026×10−5 mol, this equated to a conjugation efficiency of >99% (Table 1). Experiments were conducted to monitor size and size distribution of the liposome formulations under storage conditions of 4°C in PBS pH 7.4 over 12 weeks to ensure stability over time. The size and polydispersity index of the liposome formulations were stable over this period (Figure 1).
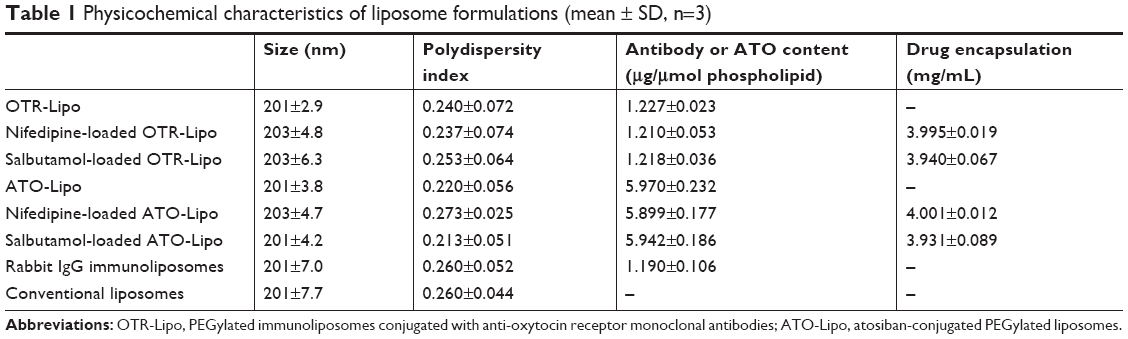 | Table 1 Physicochemical characteristics of liposome formulations (mean ± SD, n=3) |
 | Figure 1 Particle size and polydispersity index of OTR-Lipo (A, B) and ATO-Lipo (C, D) over a period of 12 weeks. |
Stability of drug-encapsulated OTR-targeted liposomes
Nifedipine (hydrophobic drug) and salbutamol hemisulfate (hydrophilic drug) were used to evaluate the in vitro stability of OTR-Lipo and ATO-Lipo. Nifedipine is a hydrophobic compound; therefore, its solubility in PBS pH 7.4 was initially assessed to ensure that the parameters of the dialysis assay did not surpass the saturation point for the drug. This determined that 70 μg of nifedipine in 10 mL PBS solution within the dialysis tubing was able to dialyze into 30 mL of the PBS release volume. This was not required for salbutamol as it is readily soluble in an aqueous phase, thereby allowing 1 mg of the drug to be placed in 10 mL PBS solution within the dialysis tubing. Minimal drug release was detected for both salbutamol-loaded OTR-targeted liposomes and nifedipine-loaded OTR-targeted liposomes in PBS at 37°C across the 48 hours duration of the experiment (Figure 2). In serum, there was a significant difference in stability at 24 and 48 hours compared to baseline values for both the liposome formulations (Figure 2). This equated to a decrease of 19.7% for nifedipine-loaded OTR-Lipo (P<0.01), 23.9% for salbutamol-loaded OTR-Lipo (P<0.01), 26.2% for nifedipine-loaded ATO-Lipo (P<0.0001), and 19.2% for salbutamol-loaded ATO-Lipo (P<0.01) at 48 hours. Drug release was not limited by the diffusion of free drug through the dialysis membrane, as shown by complete recovery of the drug solutions in the dialysis medium within 6 hours. Therefore, both of the drugs evaluated were able to pass through the cellulose membrane freely.
 | Figure 2 Stability of nifedipine-loaded OTR-Lipo (A), nifedipine-loaded ATO-Lipo (B), salbutamol-loaded OTR-Lipo (C), and salbutamol-loaded ATO-Lipo (D) in PBS pH 7.4 and 50% FBS at 37°C. |
Liposomal imaging with AFM
AFM was used to visualize all liposomal formulations when dried to confirm particle size and morphology. For size determination, all visible particles within a representative scan area were individually evaluated. OTR-Lipo, ATO-Lipo, rabbit IgG immunoliposomes, and conventional liposomes showed approximate vesicle sizes of 200 nm (Figure 3), which corresponds with the results from dynamic laser light scattering. The surface of conventional liposomes was typically smooth (Figure 3D), whereas small surface structures can be detected at the membrane border for OTR-Lipo (Figure 3A) and rabbit IgG immunoliposomes (Figure 3C) and only slightly for ATO-Lipo (Figure 3B). Surface coverage in the height mode was difficult to evaluate due to the collapsed vesicular structure when dried.
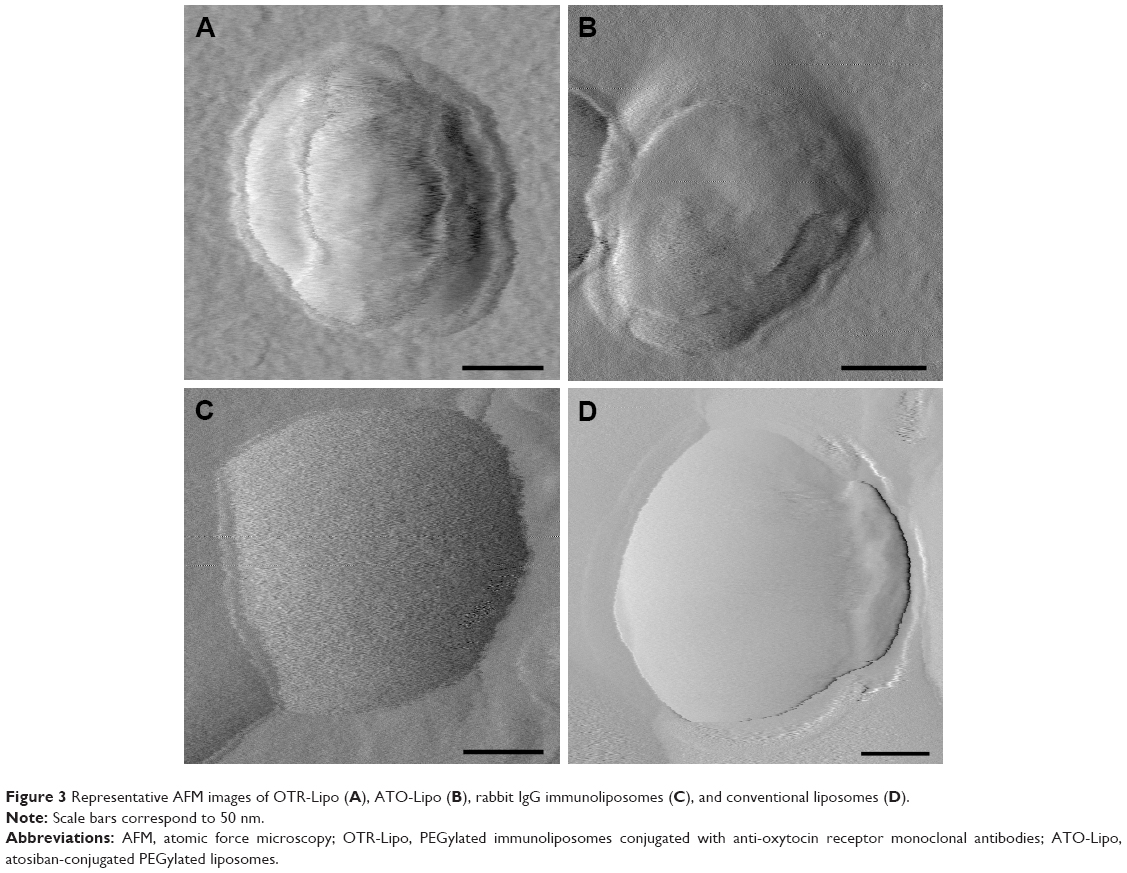 | Figure 3 Representative AFM images of OTR-Lipo (A), ATO-Lipo (B), rabbit IgG immunoliposomes (C), and conventional liposomes (D). |
Specificity of binding
OTR-Lipo and ATO-Lipo bound significantly to hTERT-myo cells in a concentration-dependent manner in comparison with all control groups (P<0.0001) (Figure 4). There was no significant difference in binding values between OTR-Lipo and ATO-Lipo across all concentrations evaluated (P>0.05). Negative control binding experiments using non-specific rabbit-IgG immunoliposomes and conventional liposomes were assessed to demonstrate the specificity of the binding toward OTR expressing cells. The results showed very low cell binding values across all concentrations evaluated and were not significantly different (P>0.05). In another set of experiments, hTERT-myo cells were pretreated with excess monoclonal antibody directed against OTRs to further demonstrate the specific nature of the OTR-Lipo and ATO-Lipo interaction. Figure 4 shows significant inhibition of binding of both OTR-Lipo and ATO-Lipo to hTERT-myo cells pretreated with excess anti-OTR monoclonal antibody (1 mg/mL) by up to 95.0% (P<0.0001) and 92.1% (P<0.0001), respectively.
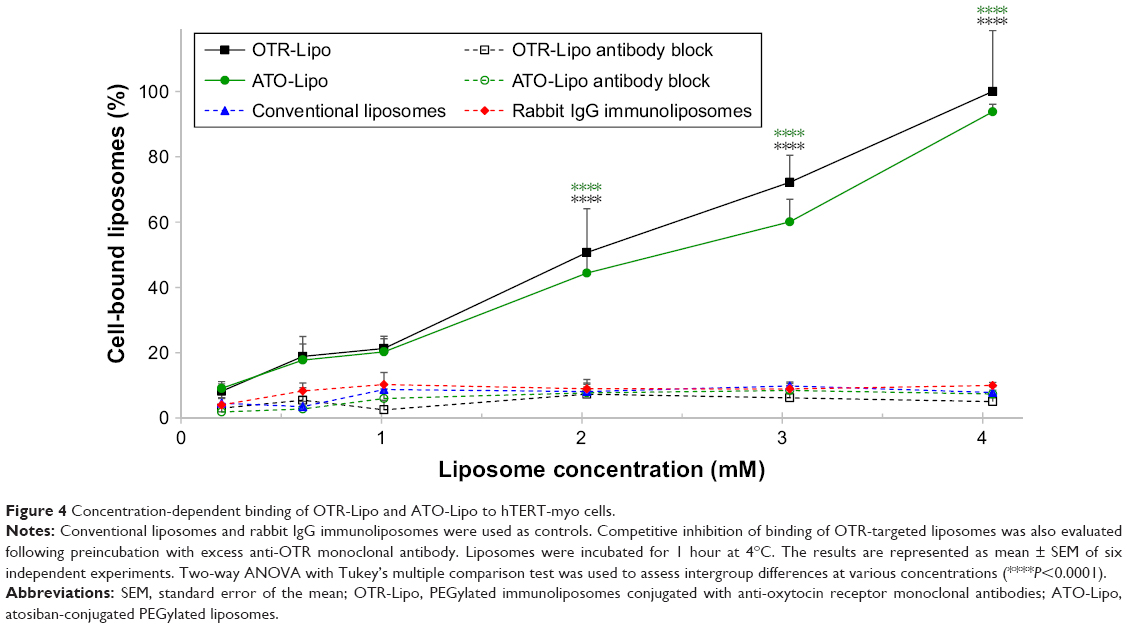 | Figure 4 Concentration-dependent binding of OTR-Lipo and ATO-Lipo to hTERT-myo cells. |
Degree of cellular uptake
OTR-Lipo (81.61%±7.84%) and ATO-Lipo (85.59%±8.28%) demonstrated significantly increased cellular internalization compared with rabbit IgG immunoliposomes (9.14%±1.71%) and conventional liposomes (4.09%±0.78%) at 2.02 mM phospholipid concentration after 1 hour of incubation at 37°C (Figure 5A, P<0.0001). Similar results were shown following liposome incubation at 4.05 mM (Figure 5B), with cell-bound liposomes of 15.95%±2.45% and 9.21%±0.74% and cellular internalization of 84.05%±12.30% and 85.60%±11.00% for OTR-Lipo and ATO-Lipo, respectively. Low levels of cell binding were shown for non-specific rabbit-IgG immunoliposomes and conventional liposomes at both concentrations, which were not significantly different from cell-bound OTR-Lipo and ATO-Lipo (P>0.05). No significant difference was also detected between cellular binding and uptake for rabbit-IgG immunoliposomes and conventional liposomes (P>0.05).
 | Figure 5 Cellular association of OTR-Lipo, ATO-Lipo, rabbit IgG immunoliposomes and conventional liposomes by hTERT-myo cells. |
Mechanisms of cellular uptake
All three pharmacological inhibitors of clathrin-mediated endocytosis (chlorpromazine, monodansylcadaverine, and methyl-β-cyclodextrin) were able to significantly block the internalization of OTR-Lipo and ATO-Lipo (Figure 6, P<0.0001). As further support of the specificity for inhibition of clathrin-mediated endocytosis, the results also showed significant inhibition of uptake for fluorescent-labeled transferrin with all three compounds (positive control, P<0.0001). However, methyl-β-cyclodextrin is also known to block caveolin-mediated endocytosis, which is supported by significant inhibition of uptake of the positive control – cholera toxin subunit B (Figure 6C, P<0.0001). Hence, further investigations were conducted with more specific pharmacological inhibitors of caveolin-mediated endocytosis (genistein and filipin). Figure 7A shows significant inhibition of cellular uptake for OTR-Lipo (P<0.01), ATO-Lipo (P<0.0001), and fluorescent-labeled cholera toxin subunit B (positive control, P<0.001) with genistein. Filipin did not affect the uptake of both OTR-targeted liposomes or cholera toxin subunit B on hTERT-myo cells (Figure 7B, P>0.05). Furthermore, no significant inhibition of cellular uptake was observed for OTR-Lipo and ATO-Lipo with cytochalasin D (Figure 7C, P>0.05), a pharmacological inhibitor of both phagocytosis and macropinocytosis. Cytochalasin D was, however, able to inhibit the uptake of fluorescent microspheres of 1 μm diameter (positive control, P<0.0001). Nocodazole (pharmacological inhibitor of macropinocytosis) was unable to block the cellular uptake of both OTR-targeted liposomes and positive control (fluorescent microspheres) across all concentrations evaluated (Figure 7D, P>0.05).
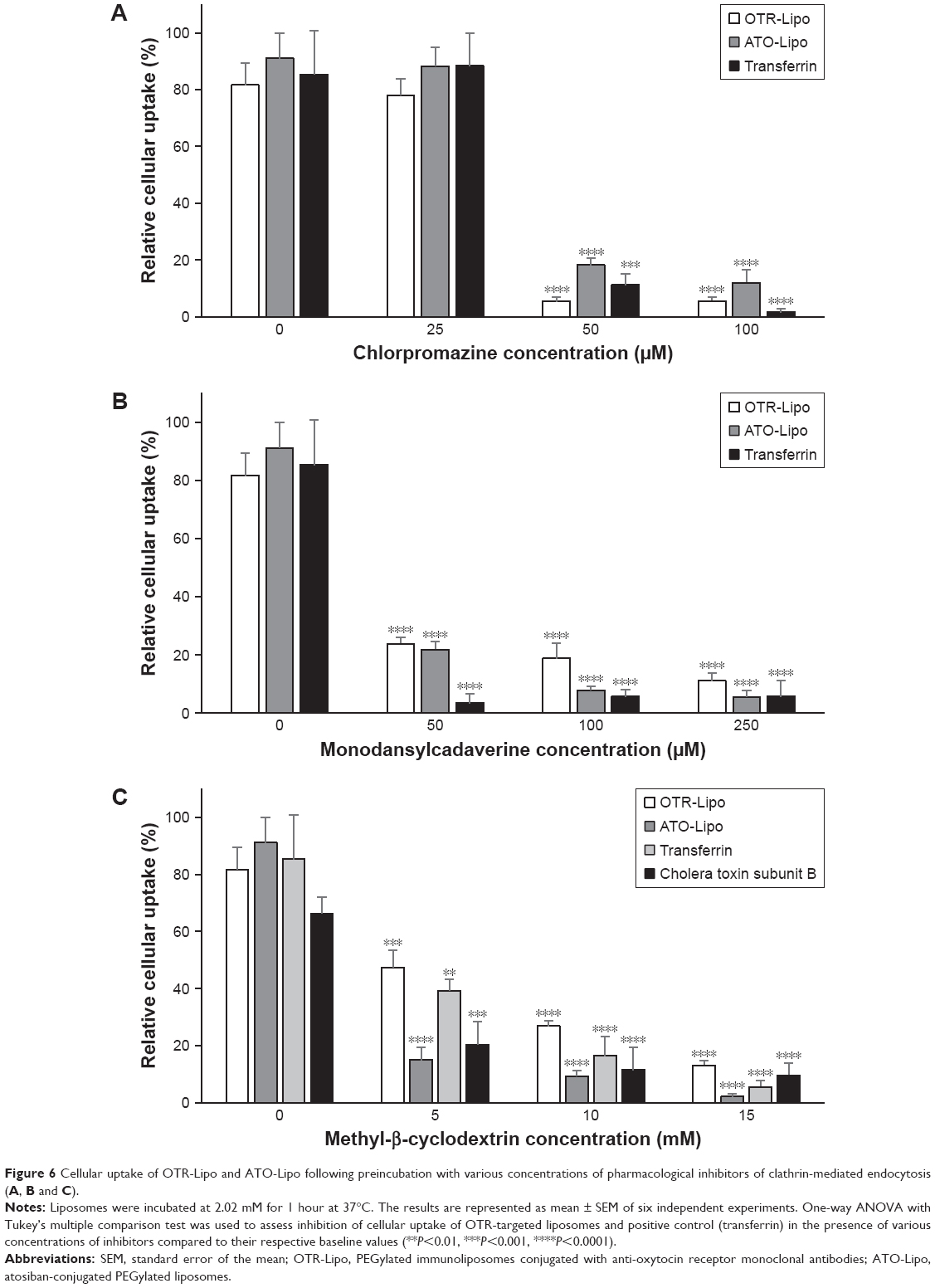 | Figure 6 Cellular uptake of OTR-Lipo and ATO-Lipo following preincubation with various concentrations of pharmacological inhibitors of clathrin-mediated endocytosis (A, B and C). |
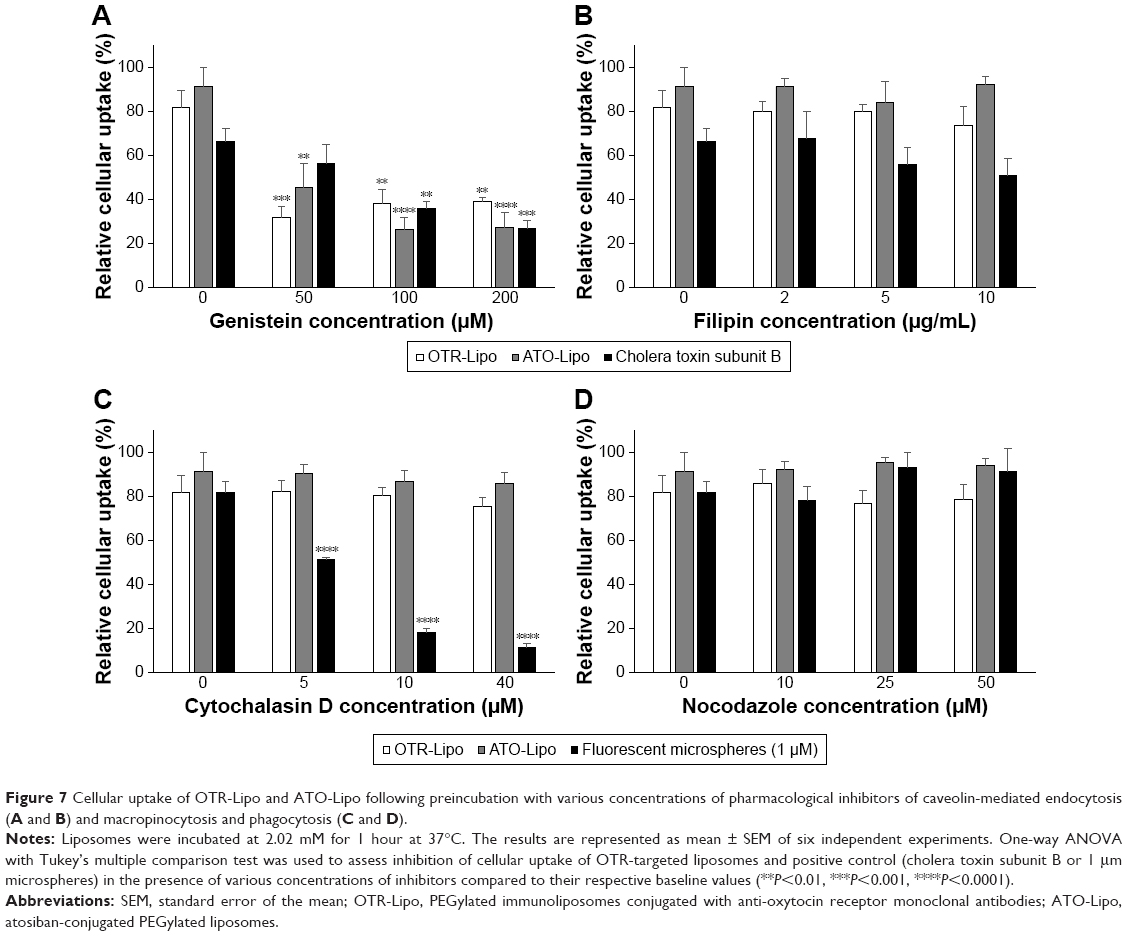 | Figure 7 Cellular uptake of OTR-Lipo and ATO-Lipo following preincubation with various concentrations of pharmacological inhibitors of caveolin-mediated endocytosis (A and B) and macropinocytosis and phagocytosis (C and D). |
Cellular toxicity
Cell viability studies were conducted following incubation with liposomes and the endocytotic inhibitors to rule out cellular toxicity as a potential reason for decreased liposome internalization during these experiments (Figure 8). The results showed no significant effect of OTR-Lipo and ATO-Lipo on the viability of hTERT-myo cells following 24 hours of incubation at both the concentrations evaluated (2.02 and 4.05 mM) (P>0.05). Furthermore, incubation with the endocytic inhibitors at the concentrations used in the in vitro experiments for 1.5 hour (cellular incubation time) did not impair cell viability (P>0.05). Significant cell death was demonstrated for hTERT-myo cells treated with Triton X-100 (positive control) (P<0.0001).
 | Figure 8 Cell viability following exposure to OTR-targeted liposomes and specific endocytotic inhibitors at various concentrations for 24 and 1.5 hour, respectively. |
Discussion
We have engineered liposomal platforms that are able to specifically target OTRs expressed on myometrial cells. OTRs are cell surface receptors containing seven transmembrane domains and belong to the class I family of G protein-coupled receptors.31 They are regulated by changes in receptor expression and desensitization, as well as local changes in oxytocin concentration.11,31–33 The expression of OTRs in the uterus is significantly upregulated during gestation and undergoes rapid downregulation after parturition.11,16,17 Importantly, OTR levels were shown to be maximal and significantly higher after the onset of labor, either preterm or term, than before the onset of labor.17 This tissue-specific regulation of OTR expression is ideal for specific and local accumulation of OTR-targeted nanoparticles to the uterus in obstetric complications such as preterm labor, dysfunctional labor and postpartum hemorrhage21 – in which the management of these conditions is challenging due to limited effective and safe treatment options.1–5 In the uterus, the expression of OTRs is not confined to the myometrium. Chorio-decidual tissue expression of OTRs also increases during parturition34 and spatial differences exist between OTR expression within uterine tissues, which is suggested to aid fetal decent and passage during labor.11
When anti-OTR monoclonal antibodies or rabbit IgG molecules were conjugated to the liposomes, small surface structures were visible along the membrane border. Based on molecular weights of the targeting ligands, we would not expect to see the conjugation of atosiban with the surface of liposomes as clearly as compared with monoclonal antibodies or IgG molecules. Studies in the literature have attempted to visualize the conjugation of targeting ligands to the surface of liposomes using AFM with varying results.35,36 For example, Bendas et al demonstrated that when antibodies are attached to liposomes via a PEG 2000 anchor, it can cause high protein mobility that affects the ability to image the proteins,36 whereas Anabousi et al reported the detection of small globular structures at the surface of such liposomes.35 Similar to Bendas et al,36 we found it difficult to clearly image the anti-OTR antibodies attached to the liposomes via a PEG 2000 anchor. Data obtained from AFM can also deviate slightly from the results of dynamic laser light scattering because of an interaction of the soft and flexible liposomes with the surface of the silicon chip substrate, which we also observed. This is particularly relevant when the liposomes are dried onto the silicon chip prior to analysis, thereby resulting in liposomes flattening and/or spreading on the silicon support.
Conjugation of OTR targeting ligands to the surface of liposomes increased cellular interaction with myometrial uterine cells. hTERT-immortalized myometrial cells were used to evaluate specificity of OTR targeting, as they exhibit the phenotype and properties of highly differentiated smooth muscle cells, without the high variability of responses seen with primary myometrial cells.37,38 Human myometrial cells express OTRs39–41 and have been reported to acquire additional OTRs in culture,42–44 with upregulation likely due to the presence of fetal bovine serum in the culture medium.45 It should be noted that serum and supplement free media were used in the in vitro cellular association studies to determine binding and cellular uptake of the OTR-targeted liposomal platforms in a reductionist in vitro system, which would mimic the in vivo situation of when the liposomes reached the myometrium of the uterus. We have demonstrated highly specific binding of OTR-Lipo and ATO-Lipo to OTR-expressing hTERT-myo cells using concentration-dependent binding studies, competitive inhibition experiments, and negative control binding studies. No significant difference was detected in binding efficiency between liposomes conjugated with anti-OTR monoclonal antibodies and atosiban. Similarly, Refuerzo et al showed improved attachment, retention, and targeting efficiency with atosiban-conjugated liposomes compared with non-targeted liposomes on smooth muscle cells isolated from pregnant humans and mice.46 The use of monoclonal antibodies is preferential to polyclonal antibodies for translational purposes to improve the specificity of binding. This is due to monoclonal antibodies being able to specifically detect a particular epitope on the antigen, which means they are less likely to cross-react with other proteins.
Cellular association experiments showed significant cellular uptake for both OTR-Lipo and ATO-Lipo following binding, which is necessary for subsequent intracellular processing and release of encapsulated therapeutic agents in myometrial cells. Interestingly, anti-OTR monoclonal antibodies and atosiban in the free drug form typically act by blocking natural ligands from binding to myometrial OTRs on the membrane surface.14,15 However, conjugation of these ligands to the surface of liposomes enhances its uptake into myometrial cells. The cellular interaction of liposomes is highly dependent on several liposomal characteristics including particle size, charge, composition, sterical stabilization, and specific properties of the conjugated ligands.23,27 OTR-targeted liposomes themselves are unlikely to impact on OTR desensitization rates, as the concentration of ligand conjugated to the surface of the liposomes is low. Usually much higher doses and continuous dosing of oxytocin are required to cause downregulation of OTRs.47–49
Evaluation of the mechanistic pathway of cellular uptake of the different OTR-targeted liposomal platforms indicates that they both undergo internalization through clathrin- and caveolin-mediated mechanisms. OTRs have previously been shown to congregate with β-arrestin following activation into defined punctuated regions of the plasma membrane, which suggest that they are targeted into clathrin-coated pits for internalization.31,50 OTRs are also highly expressed in the cholesterol-rich, caveolin-containing membrane domains of the plasma membrane.51,52 Furthermore, results from cellular toxicity experiments showed no significant effects of both OTR-targeted liposomal platforms and the endocytotic inhibitors on cell viability at 24 and 1.5 hour (cellular incubation time), respectively. It should be noted that utilization of pharmacological inhibitors is a common approach for studying endocytotic pathways. Generally, a number of pharmacological inhibitors should be evaluated as they can lack specificity for defined pathways and have been shown to be cell type dependent.53 Therefore, a balance is needed between the concentration of inhibitor high enough to inhibit endocytosis but not to induce cytotoxicity.53
OTR-Lipo and ATO-Lipo were shown to be highly stable upon dilution in both an aqueous phase (PBS pH 7.4) and serum (50% FBS). Drug retention was only significantly lower than baseline values when the liposomes were dispersed in serum at 24 and 48 hours. This is likely due to the bioactive substance in FBS, including plasma proteins which can lead to opsonization, destabilization, and lipid exchange.23 The amount of encapsulated drug released was between 19.2% and 26.2% at 48 hours. This is unlikely to affect the performance of the OTR-targeted liposomes, as human serum generally contains 7% proteins (50% FBS was used in the experiment), and the expected application for the technology would require good stability in the circulation particularly following the first few hours after initial administration. Therefore, the targeted nanoparticles are suitable for administration to sites of high dilution, which occurs following intravenous administration. Stability against leakage has been achieved by using phospholipids (eg, DSPC) that remain in the solid phase at physiological temperatures and incorporating cholesterol in the lipid bilayer to minimize lipid exchange.54 PEGylation also improves the circulation half-life of the formulation by evading recognition by the immune system.23,55,56 Salbutamol and nifedipine were chosen for the stability study due to their difference in hydrophobicity, which allows evaluation of the delivery system to potentially encapsulate a variety of therapeutic agents. Hydrophobic molecules are inserted into the bilayer membrane, and hydrophilic molecules can be entrapped in the aqueous center.23 In addition, salbutamol and nifedipine are currently used to treat preterm labor and have been associated with significant maternal and fetal adverse effects, due to the high doses needed to achieve therapeutic effects in the uterus.2–4
The functionality of the OTR-targeted liposomal platforms has previously been evaluated on murine and human myometrial tissues as well as in vivo in a murine model of preterm labor.21,46 We have shown that liposomes conjugated with anti-OTR polyclonal antibodies (200 nm) and loaded with salbutamol, nifedipine, or rolipram (tocolytic agents) inhibited myometrial contractions ex vivo, while those loaded with dofetilide (uterotonic agent) increased contraction duration.21 Empty OTR-targeted liposomes and non-targeted control liposomes loaded with these agents had no effect.21 Similar findings have also been demonstrated for ATO-Lipo (124 nm).46 Furthermore, liposomes loaded with indomethacin (tocolytic agent) and conjugated with anti-OTR polyclonal antibodies21 or atosiban46 significantly reduced the rates of preterm birth in mice compared with non-targeted liposomes and showed no detectable accumulation in the maternal brain or fetus.21,46 Parturition is considered an inflammatory process,23,57 which indicates that the enhanced permeability and retention (EPR) effect may also play a role in liposome accumulation into the uterus. To what extent EPR occurs in parturition as well as in obstetric complications will need to be evaluated in future studies. It should be noted, however, that OTR-targeted liposomes are unlikely to be effective in patients previously exposed to continuous high doses of oxytocin, as this typically leads to desensitization of OTRs for hours or even days,47,48 thereby reducing the targeting effectiveness of the drug delivery system. This phenomenon is accompanied by a downregulation of OTRs at the protein and mRNA level.47,49
Conclusion
This study further supports OTRs as a novel pharmaceutical target for drug delivery. OTR-targeted liposomes may provide an effective way to deliver existing therapies directly to myometrial tissue for the treatment of obstetric complications and avoid adverse effects by circumventing non-target tissues. Achieving targeted delivery of therapeutic agents to the myometrium would potentially increase therapeutic efficacy, decrease the therapeutically effective dose, and/or reduce the risk of systemic side effects. This study shows that when all physicochemical parameters of the liposomes are the same, both OTR-Lipo and ATO-Lipo have similar stability, specificity in OTR binding, degree of cellular uptake, mechanism of endocytotic uptake, and effect on cell viability. Translational development will depend on comprehensive preclinical and clinical studies, as well as other factors specific to the commercialization of nanomedicines.58
In addition to the uterus, OTRs have been reported to be expressed in a multitude of tissues, including breast, pituitary, kidney, ovary, thymus, heart, vascular endothelium, osteoclasts, myoblasts, pancreatic islet cells, and adipocytes.11,31 However, their clinical relevance in humans has not been fully established. Further studies will be required to quantify the biodistribution of OTR-targeted liposomes in these tissues in relevant animal models of these obstetric complications. OTRs are also structurally similar to the vasopressin receptor subtypes.11,20,59 It will be important to determine the degree of cross-reactivity of the different OTR-targeted liposomal platforms with vasopressin receptors and the potential clinical implications of this interaction with regard to biodistribution of the delivery system. It should be noted that vasopressin receptors (V1a receptors) are also expressed in the uterus and are functionally coupled to myometrial contraction.60,61 Interestingly, OTRs have also been detected in several human cancer tissues and cell lines,31 including human breast cancer,62,63 uterine leiomyoma,41,64 adenocarcinoma of the endometrium,65 neuroblastoma, and glioma.66 Therefore, OTR-targeted drug delivery systems may also be adapted to assist in the treatment of other disease pathologies characterized by an upregulation of OTR expression.
Acknowledgments
This work was supported by the National Health and Medical Research Council, Hunter Medical Research Institute, Global Alliance to Prevent Prematurity and Stillbirth, Gladys M Brawn Fellowship, and University of Newcastle. This work was performed in part at the Materials node of the Australian National Fabrication Facility, which is a company established under the National Collaborative Research Infrastructure Strategy to provide nano and microfabrication facilities for Australian researchers.
Author contributions
SH contributed to study concept and design, liposome manufacturing and characterization, liposome imaging, cellular studies, analysis and interpretation of data, and drafting of manuscript. BV contributed to liposome imaging and analysis and interpretation of data. Both authors were involved in revising the article critically for important intellectual content and gave final approval of the version to be published. The authors agree to be accountable for all aspects of the work.
Disclosure
SH has a patent through the University of Newcastle related to the use of targeted liposomes. BV reports no conflicts of interest in this work.
References
Butwick AJ, Carvalho B, Blumenfeld YJ, El-Sayed YY, Nelson LM, Bateman BT. Second-line uterotonics and the risk of hemorrhage-related morbidity. Am J Obstet Gynecol. 2015;212(5):642.e1–642.e7. | ||
de Heus R, Mol BW, Erwich JJ, et al. Adverse drug reactions to tocolytic treatment for preterm labour: prospective cohort study. BMJ. 2009;338:b744. | ||
Haas DM, Caldwell DM, Kirkpatrick P, Mcintosh JJ, Welton NJ. Tocolytic therapy for preterm delivery: systematic review and network meta-analysis. BMJ. 2012;345:e6226. | ||
Jørgensen JS, Weile LK, Lamont RF. Preterm labor: current tocolytic options for the treatment of preterm labor. Expert Opin Pharmacother. 2014;15(5):585–588. | ||
Mousa HA, Blum J, Abou El Senoun G, Shakur H, Alfirevic Z. Treatment for primary postpartum haemorrhage. Cochrane Database Syst Rev. 2014;(2):CD003249. | ||
Liu L, Oza S, Hogan D, et al. Global, regional, and national causes of child mortality in 2000-13, with projections to inform post-2015 priorities: an updated systematic analysis. Lancet. 2015;385(9966):430–440. | ||
Manuck TA, Rice MM, Bailit JL, et al. Preterm neonatal morbidity and mortality by gestational age: a contemporary cohort. Am J Obstet Gynecol. 2016;215(1):103.e1–103.e14. | ||
Romero R, Dey SK, Fisher SJ. Preterm labor: one syndrome, many causes. Science. 2014;345(6198):760–765. | ||
Al-Zirqi I, Vangen S, Forsen L, Stray-Pedersen B. Prevalence and risk factors of severe obstetric haemorrhage. BJOG. 2008;115(10):1265–1272. | ||
Khan KS, Wojdyla D, Say L, Gülmezoglu AM, van Look PF. Who analysis of causes of maternal death: a systematic review. Lancet. 2006;367(9516):1066–1074. | ||
Arrowsmith S, Wray S. Oxytocin: its mechanism of action and receptor signalling in the myometrium. J Neuroendocrinol. 2014;26(6):356–369. | ||
Blanch G, Lavender T, Walkinshaw S, Alfirevic Z. Dysfunctional labour: a randomised trial. Br J Obstet Gynaecol. 1998;105(1):117–120. | ||
Arrowsmith S, Kendrick A, Wray S. Drugs acting on the pregnant uterus. Obstet Gynaecol Reprod Med. 2010;20(8):241–247. | ||
Goodwin TM, Paul R, Silver H, et al. The effect of the oxytocin antagonist atosiban on preterm uterine activity in the human. Am J Obstet Gynecol. 1994;170(2):474–478. | ||
Romero R, Sibai BM, Sanchez-Ramos L, et al. An oxytocin receptor antagonist (atosiban) in the treatment of preterm labor: a randomized, double-blind, placebo-controlled trial with tocolytic rescue. Am J Obstet Gynecol. 2000;182(5):1173–1183. | ||
Wathes DC, Borwick SC, Timmons PM, Leung ST, Thornton S. Oxytocin receptor expression in human term and preterm gestational tissues prior to and following the onset of labour. J Endocrinol. 1999;161(1):143–151. | ||
Fuchs AR, Fuchs F, Husslein P, Soloff MS. Oxytocin receptors in the human uterus during pregnancy and parturition. Am J Obstet Gynecol. 1984;150(6):734–741. | ||
Fuchs AR, Fields MJ, Freidman S, Shemesh M, Ivell R. Oxytocin and the timing of parturition. Influence of oxytocin receptor gene expression, oxytocin secretion, and oxytocin-induced prostaglandin F2 alpha and E2 release. Adv Exp Med Biol. 1995;395:405–420. | ||
Kimura T, Takemura M, Nomura S, et al. Expression of oxytocin receptor in human pregnant myometrium. Endocrinology. 1996;137(2):780–785. | ||
Akerlund M. Targeting the oxytocin receptor to relax the myometrium. Expert Opin Ther Targets. 2006;10(3):423–427. | ||
Paul JW, Hua S, Ilicic M, et al. Drug delivery to the human and mouse uterus using immunoliposomes targeted to the oxytocin receptor. Am J Obstet Gynecol. 2017;216(3):283.e1–283.e14. | ||
Hua S. Synthesis and in vitro characterization of oxytocin receptor targeted PEGylated immunoliposomes for drug delivery to the uterus. J Liposome Res. 2019:1–11. | ||
Sercombe L, Veerati T, Moheimani F, Wu SY, Sood AK, Hua S. Advances and challenges of liposome assisted drug delivery. Front Pharmacol. 2015;6:286. | ||
Ulrich AS. Biophysical aspects of using liposomes as delivery vehicles. Biosci Rep. 2002;22(2):129–150. | ||
Majumder P, Bhunia S, Bhattacharyya J, Chaudhuri A. Inhibiting tumor growth by targeting liposomally encapsulated CDC20siRNA to tumor vasculature: therapeutic RNA interference. J Control Release. 2014;180:100–108. | ||
Reddy JA, Abburi C, Hofland H, et al. Folate-targeted, cationic liposome-mediated gene transfer into disseminated peritoneal tumors. Gene Ther. 2002;9(22):1542–1550. | ||
Hua S, Chang HI, Davies NM, Cabot PJ. Targeting of ICAM-1-directed immunoliposomes specifically to activated endothelial cells with low cellular uptake: use of an optimized procedure for the coupling of low concentrations of antibody to liposomes. J Liposome Res. 2011;21(2):95–105. | ||
Lopes de Menezes DE, Pilarski LM, Allen TM. In vitro and in vivo targeting of immunoliposomal doxorubicin to human B-cell lymphoma. Cancer Res. 1998;58(15):3320–3330. | ||
Voinea M, Manduteanu I, Dragomir E, Capraru M, Simionescu M. Immunoliposomes directed toward VCAM-1 interact specifically with activated endothelial cells-a potential tool for specific drug delivery. Pharm Res. 2005;22(11):1906–1917. | ||
Everts M, Koning GA, Kok RJ, et al. In vitro cellular handling and in vivo targeting of E-selectin-directed immunoconjugates and immunoliposomes used for drug delivery to inflamed endothelium. Pharm Res. 2003;20(1):64–72. | ||
Zingg HH, Laporte SA. The oxytocin receptor. Trends Endocrinol Metab. 2003;14(5):222–227. | ||
Szukiewicz D, Bilska A, Mittal TK, et al. Myometrial contractility influences oxytocin receptor (OXTR) expression in term trophoblast cells obtained from the maternal surface of the human placenta. BMC Pregnancy Childbirth. 2015;15:220. | ||
Gimpl G, Fahrenholz F. The oxytocin receptor system: structure, function, and regulation. Physiol Rev. 2001;81(2):629–683. | ||
Takemura M, Kimura T, Nomura S, et al. Expression and localization of human oxytocin receptor mRNA and its protein in chorion and decidua during parturition. J Clin Invest. 1994;93(6):2319–2323. | ||
Anabousi S, Laue M, Lehr C-M, Bakowsky U, Ehrhardt C. Assessing transferrin modification of liposomes by atomic force microscopy and transmission electron microscopy. Eur J Pharm Biopharm. 2005;60(2):295–303. | ||
Bendas G, Krause A, Bakowsky U, Vogel J, Rothe U. Targetability of novel immunoliposomes prepared by a new antibody conjugation technique. Int J Pharm. 1999;181(1):79–93. | ||
Sharkey J, Olcese J. Transcriptional inhibition of oxytocin receptor expression in human myometrial cells by melatonin involves protein kinase C signaling. J Clin Endocrinol Metab. 2007;92(10):4015–4019. | ||
Soloff MS, Jeng YJ, Ilies M, et al. Immortalization and characterization of human myometrial cells from term-pregnant patients using a telomerase expression vector. Mol Hum Reprod. 2004;10(9):685–695. | ||
Maggi M, Peri A, Baldi E, et al. Interferon-alpha downregulates expression of the oxytocin receptor in cultured human myometrial cells. Am J Physiol. 1996;271(5 Pt 1):E840–E846. | ||
Tahara A, Tsukada J, Tomura Y, et al. Pharmacologic characterization of the oxytocin receptor in human uterine smooth muscle cells. Br J Pharmacol. 2000;129(1):131–139. | ||
Busnelli M, Rimoldi V, Viganò P, Persani L, Di Blasio AM, Chini B. Oxytocin-induced cell growth proliferation in human myometrial cells and leiomyomas. Fertil Steril. 2010;94(5):1869–1874. | ||
Molnár M, Rigo J Jr, Romero R, Hertelendy F. Oxytocin activates mitogen-activated protein kinase and up-regulates cyclooxygenase-2 and prostaglandin production in human myometrial cells. Am J Obstet Gynecol. 1999;181(1):42–49. | ||
Ohmichi M, Koike K, Nohara A, et al. Oxytocin stimulates mitogen-activated protein kinase activity in cultured human puerperal uterine myometrial cells. Endocrinology. 1995;136(5):2082–2087. | ||
Phaneuf S, Europe-Finner GN, Varney M, Mackenzie IZ, Watson SP, López Bernal A. Oxytocin-stimulated phosphoinositide hydrolysis in human myometrial cells: involvement of pertussis toxin-sensitive and -insensitive G-proteins. J Endocrinol. 1993;136(3):497–509. | ||
Jeng YJ, Soloff SL, Anderson GD, Soloff MS. Regulation of oxytocin receptor expression in cultured human myometrial cells by fetal bovine serum and lysophospholipids. Endocrinology. 2003;144(1):61–68. | ||
Refuerzo JS, Leonard F, Bulayeva N, et al. Uterus-targeted liposomes for preterm labor management: studies in pregnant mice. Sci Rep. 2016;6:34710. | ||
Vrachnis N, Malamas FM, Sifakis S, Deligeoroglou E, Iliodromiti Z. The oxytocin-oxytocin receptor system and its antagonists as tocolytic agents. Int J Endocrinol. 2011;2011:350546. | ||
Plested CP, Bernal AL. Desensitisation of the oxytocin receptor and other G-protein coupled receptors in the human myometrium. Exp Physiol. 2001;86(2):303–312. | ||
Phaneuf S, Rodríguez Liñares B, Tambyraja RL, Mackenzie IZ, López Bernal A. Loss of myometrial oxytocin receptors during oxytocin-induced and oxytocin-augmented labour. J Reprod Fertil. 2000;120(1):91–97. | ||
Oakley RH, Laporte SA, Holt JA, Barak LS, Caron MG. Molecular determinants underlying the formation of stable intracellular G protein-coupled receptor-beta-arrestin complexes after receptor endocytosis*. J Biol Chem. 2001;276(22):19452–19460. | ||
Gimpl G, Burger K, Politowska E, Ciarkowski J, Fahrenholz F. Oxytocin receptors and cholesterol: interaction and regulation. Exp Physiol. 2000;85 Spec No:41S–49S. | ||
Gimpl G, Fahrenholz F. Human oxytocin receptors in cholesterol-rich vs cholesterol-poor microdomains of the plasma membrane. Eur J Biochem. 2000;267(9):2483–2497. | ||
Vercauteren D, Vandenbroucke RE, Jones AT, et al. The use of inhibitors to study endocytic pathways of gene carriers: optimization and pitfalls. Mol Ther. 2010;18(3):561–569. | ||
Anderson M, Omri A. The effect of different lipid components on the in vitro stability and release kinetics of liposome formulations. Drug Deliv. 2004;11(1):33–39. | ||
Allen TM. Long-circulating (sterically stabilized) liposomes for targeted drug delivery. Trends Pharmacol Sci. 1994;15(7):215–220. | ||
Torchilin VP. Immunoliposomes and PEGylated immunoliposomes: possible use for targeted delivery of imaging agents. Immunomethods. 1994;4(3):244–258. | ||
Norman JE, Bollapragada S, Yuan M, Nelson SM. Inflammatory pathways in the mechanism of parturition. BMC Pregnancy Childbirth. 2007;7(Suppl 1):S7. | ||
Hua S, de Matos MBC, Metselaar JM, Storm G. Current trends and challenges in the clinical translation of nanoparticulate nanomedicines: pathways for translational development and Commercialization. Front Pharmacol. 2018;9:790. | ||
Shojo H, Kaneko Y. Characterization and expression of oxytocin and the oxytocin receptor. Mol Genet Metab. 2000;71(4):552–558. | ||
Bossmar T, Akerlund M, Fantoni G, Szamatowicz J, Melin P, Maggi M. Receptors for and myometrial responses to oxytocin and vasopressin in preterm and term human pregnancy: effects of the oxytocin antagonist atosiban. Am J Obstet Gynecol. 1994;171(6):1634–1642. | ||
Maggi M, Del Carlo P, Fantoni G, et al. Human myometrium during pregnancy contains and responds to V1 vasopressin receptors as well as oxytocin receptors. J Clin Endocrinol Metab. 1990;70(4):1142–1154. | ||
Cassoni P, Sapino A, Fortunati N, Munaron L, Chini B, Bussolati G. Oxytocin inhibits the proliferation of MDA-MB231 human breast-cancer cells via cyclic adenosine monophosphate and protein kinase A. Int J Cancer. 1997;72(2):340–344. | ||
Ito Y, Kobayashi T, Kimura T, et al. Investigation of the oxytocin receptor expression in human breast cancer tissue using newly established monoclonal antibodies. Endocrinology. 1996;137(2):773–779. | ||
Lee KH, Khan-Dawood FS, Dawood MY. Oxytocin receptor and its messenger ribonucleic acid in human leiomyoma and myometrium. Am J Obstet Gynecol. 1998;179(3 Pt 1):620–627. | ||
Cassoni P, Fulcheri E, Carcangiu ML, Stella A, Deaglio S, Bussolati G. Oxytocin receptors in human adenocarcinomas of the endometrium: presence and biological significance. J Pathol. 2000;190(4):470–477. | ||
Cassoni P, Sapino A, Stella A, Fortunati N, Bussolati G. Presence and significance of oxytocin receptors in human neuroblastomas and glial tumors. Int J Cancer. 1998;77(5):695–700. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.