Back to Journals » Journal of Asthma and Allergy » Volume 18
Identification of Co-Diagnostic Genes and Potential Therapeutic Targets for Asthma and Sepsis Through Mendelian Randomization and Immune Infiltration Analysis
Authors Chen C, Li X, Zhang X, Hu M, Yang M ![]() , Yuan Z, Yao S, Qin S, Qin Y, Xiao Y
, Yuan Z, Yao S, Qin S, Qin Y, Xiao Y ![]()
Received 30 June 2025
Accepted for publication 28 November 2025
Published 10 December 2025 Volume 2025:18 Pages 1689—1711
DOI https://doi.org/10.2147/JAA.S550648
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Luis Garcia-Marcos
Changhan Chen,1,2,* Xinyi Li,3,* Xupeng Zhang,3 Manlin Hu,3 Meng Yang,3 Zhangchi Yuan,3 Shanhu Yao,4 Sha Qin,5 Yuexiang Qin,1,6 Yuyang Xiao1,3
1Department of Otolaryngology and Head and Neck Surgery, Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China; 2National Medical Metabolomics International Collaborative Research Center, Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China; 3Xiangya School of Medicine, Central South University, Changsha, Hunan, People’s Republic of China; 4Department of Radiology, the Third Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China; 5Clinical Laboratory of Shenzhen Children’s Hospital, Shenzhen, Guangdong, People’s Republic of China; 6Health Management Medicine Center, the Third Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yuexiang Qin, Department of Otolaryngology and Head and Neck Surgery, Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China, Email [email protected] Yuyang Xiao, Department of Otolaryngology and Head and Neck Surgery, Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China, Email [email protected]
Background: Evidence suggests a bidirectional relationship between asthma and sepsis, but the mechanisms are unclear. This study explores the co-diagnostic genes and molecular links between asthma and sepsis using Mendelian Randomization (MR) and bioinformatics methods, and further validates the findings through clinical data, aiming to identify potential therapeutic targets and drugs.
Methods: Protein Quantitative Trait Loci (pQTL) data from an Icelandic population were used for two-sample MR analysis. Differential expression analysis identified common genes. Gene Ontology and KEGG enrichment analyses explored biological functions. Receiver Operating Characteristic (ROC) curves validated gene performance, and immune cell infiltration was analyzed using CIBERSORT. Drug targets were predicted using DrugSigDB and validated by molecular docking. Clinical data of the three groups of people were collected, and baseline analysis, ROC curve analysis, and comparison of the expression levels of CXCL8 were carried out.
Results: At the genetic level, 435 genes related to asthma and 1,385 genes related to sepsis were identified, with 141 common genes. Further findings showed that there were 247 differentially expressed genes in asthma and 2,878 differentially expressed genes in sepsis, and 65 common differentially expressed genes were enriched in immune and inflammatory pathways. The key gene CXCL8 exhibited excellent diagnostic performance and was closely related to immune cell subsets. Based on the research results, ten potential therapeutic drugs were screened, and seven of them were verified by molecular docking. Clinical sample testing results show that CXCL8 is closely related to asthma and sepsis.
Conclusion: CXCL8 may be a biomarker for both asthma and sepsis. Immune cells like monocytes, macrophages, and T cells may drive comorbid progression. Potential drug candidates were identified, offering new insights for future research on these diseases.
Keywords: MR, machine learning, molecular docking, asthma, sepsis, CXCL8
Introduction
Asthma is recognized as a heterogeneous disease1 and remains a widespread global health issue, imposing a significant economic burden worldwide.2 An estimated 241 million people are affected by asthma, with both its morbidity and mortality rates remaining high.3 The disease is characterized by airway inflammation and hyperresponsiveness.4 Genetic, epigenetic, and environmental factors all contribute to the development of asthma.5 Patients typically present with expiratory dyspnea and wheezing, often accompanied by shortness of breath, chest tightness, or cough.6,7 Furthermore, asthma is frequently associated with comorbidities such as gastroesophageal reflux disease, allergic rhinitis, obesity, depression, diabetes, and cardiovascular disease.8 Currently, asthma diagnosis primarily relies on pulmonary function tests, exhaled nitric oxide measurements, sputum eosinophil counts, allergen tests, chest imaging, and blood gas analysis. Its treatment mainly uses glucocorticoid therapy, in addition to β receptor agonists, leukotriene modulators, theophylline and other drugs.6,7 Topical and systemic glucocorticoid therapy is effective in controlling asthma; however, it is associated with several side effects, including central weight gain, hypertension, insulin resistance, type 2 diabetes, and osteoporosis.9 Additionally, current methods for evaluating and treating severe asthma have notable limitations.10 Consequently, there is an urgent need to explore novel approaches for the diagnosis and treatment of asthma.
Sepsis is a clinical syndrome triggered by an abnormal immune response to infection,11 characterized by life-threatening multi-organ dysfunction.12 The progression of sepsis can be divided into two stages: (i) an initial systemic inflammatory response syndrome, which may lead to multi-organ failure and septic shock; (ii) the subsequent compensatory anti-inflammatory response syndrome, a stage that results in sepsis-induced immunosuppression, heightening the risk of secondary infections and contributing to mortality.13 Annually, approximately 48.9 million people worldwide are diagnosed with sepsis, with around 11 million deaths, accounting for 19.7% of global mortality.14 It is also the leading cause of death among pediatric patients.15 Sepsis is a heterogeneous condition, causing a variety of infections from diverse pathogens, each resulting in distinct patterns of organ damage.16 Currently, the treatment of sepsis primarily relies on bundled management, including rapid cardiopulmonary resuscitation, intravenous antibiotic therapy, and source control.17,18 While these approaches have shown some effectiveness, they are also fraught with limitations.19 The timely selection and administration of antibiotics are crucial, as delayed or inappropriate antibiotic use is associated with higher mortality.20 Emerging therapies, such as glucocorticoids and immunomodulation, are under investigation, but robust evidence supporting their efficacy remains lacking. Despite the gradual standardization of sepsis treatment protocols, significant challenges persist, and further research is essential to enhance therapeutic outcomes.21
Although asthma and sepsis differ significantly in terms of pathogenesis and clinical manifestations, there is an underlying intrinsic connection between the two, primarily involving immune system dysfunction and inflammatory responses. Macrophage activation and polarization are thought to play a pivotal role in the onset and progression of both diseases,22,23 while the complement system is extensively implicated in their pathogenesis.24 Additionally, the structure and function of gut microbes are closely linked to the development of asthma,25 and the gut microbiota also plays a crucial role in the onset and progression of sepsis.26 Both asthma and sepsis are known to disrupt the body’s acid-base and electrolyte balance.27,28 In this study, we integrated the gene expression data from both diseases, systematically analyzed their shared Differentially Expressed Genes (DEGs) using bioinformatics methods, and employed Protein Quantitative Trait Loci (pQTL) colocalization recognition sites as Single Nucleotide Polymorphisms (SNPs) for Mendelian Randomization (MR) analysis to identify common genes. By examining these shared genes, we aim to uncover their roles in immune responses and inflammatory processes, as well as explore potential co-regulatory mechanisms. Finally, we further validated the associations between the common genes and the diseases through the analysis of clinical patient data. This study highlights the connections and common factors influencing both asthma and sepsis, offering significant insights into their pathogenesis and suggesting new directions for potential therapeutic targets.
Materials and Methods
Study Design
In this study, we identified co-diagnostic genes for asthma and sepsis through pQTL MR analysis and comprehensive bioinformatics approaches. We evaluated the expression levels and diagnostic potential of these biomarkers. In our MR analysis, SNPs associated with pQTLs were selected as instrumental variables (IVs).29,30 Subsequently, immune infiltration analysis and candidate drug evaluations were conducted. Figure 1 is the flow chart of this study. At the same time, this study included 295 research subjects, among whom there were 99 patients with sepsis and 106 patients with asthma. Through a systematic analysis of the clinical cohort data, the associated characteristics of asthma and sepsis were explored from an actual clinical perspective.
 |
Figure 1 The flow chart of the study. |
Data Sources
pQTL data for human plasma proteins were sourced from a previous study,31 which conducted a Genome-wide Association Study (GWAS) on plasma protein levels measured using 4907 aptamers in 35,559 Icelandic individuals. This study identified 18,084 associations between genetic variants and plasma protein levels (denoted as pQTLs), with 19% involving rare variants (Minor Allele Frequency < 1%).
For the MR analysis, we used summary-level data from the largest publicly available GWAS database (https://gwas.mrcieu.ac.uk/) for each trait (Table 1). Asthma and sepsis data were extracted from the MRC Integrative Epidemiology Unit database and the UK Biobank. All participants were of European ancestry to minimize potential race-related biases in the analysis.
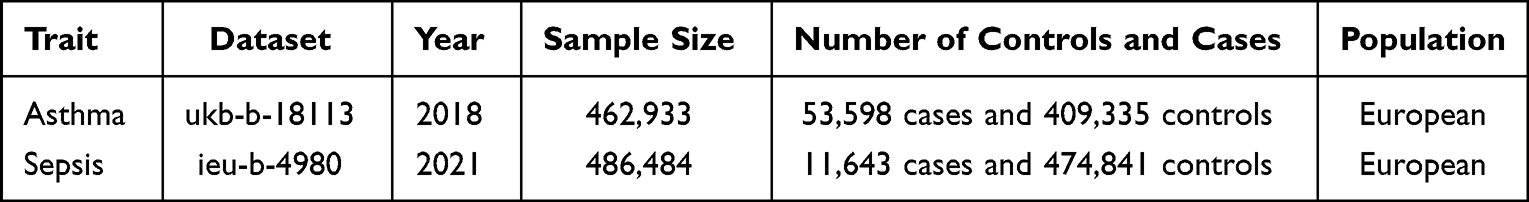 |
Table 1 The Study Details of the Asthma and Sepsis Datasets in the Mendelian Randomization Analysis |
The gene expression datasets for asthma and sepsis were obtained from the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/).32 The following criteria were applied to guide the search process:1 The samples were sourced from Homo sapiens;2 The experimental data were of the microarray type;3 The datasets included both control groups and disease groups;4 The total number of samples in each cohort was at least 10. Eventually, four GEO datasets, namely GSE143303, GSE28750, GSE147878 and GSE57065 were selected as the datasets for asthma and sepsis. Among them, GSE143303 related to asthma and GSE28750 related to sepsis were designated as the discovery cohorts. GSE147878 and GSE57065 were used to validate the hub genes of asthma and sepsis respectively. The detailed information of the above-mentioned datasets is presented in Table 2.
 |
Table 2 The Basic Information of Gene Expression Omnibus Datasets Used in This Study |
The clinical validation part of this study was conducted in the health management centers of three tertiary general hospitals in southern China, including 90 healthy individuals, 106 asthma patients, and 99 sepsis patients. Demographic data and clinical indicators were systematically collected, and the expression of CXCL8 in serum was detected by enzyme-linked immunosorbent assay (ELISA). This study was approved by the Institutional Review Board of The Third Xiangya Hospital of Central South University (Ethics Approval No.23911) and adhered to the principles of the Declaration of Helsinki. All necessary approvals were obtained from the government and the Health and Wellness Committee. A broad consent form was used in this study, which was signed by each participant before undergoing the physical examination. All personal information was anonymized during the analysis and reporting process to ensure confidentiality and privacy.
Selection of IVs and Data Coordination in MR Analysis
We included genome-wide significant SNPs with a P value < 5 × 10−8. The SNPs were clustered based on Linkage Disequilibrium (LD) (window size = 10,000 kilobases, r² < 0.001). The level of LD was estimated based on the 1000 Genomes Project for European samples.33 If a specific exposure-related SNP was not present in the outcome dataset, a proxy SNP was used as the LD marker. In MR analysis, palindromic and ambiguous SNPs were excluded from the IVs.34 The F-statistic was calculated from the variance explained by the SNPs in each exposure, using the formula [(N - K – 1)/K]/[R²/(1 – R²)]], where K is the number of genetic variants and N is the sample size. In this study, weak IVs with an F-statistic < 10 were removed.35,36 The minor allele frequency was required to be > 1%.
MR Analysis of pQTL
We utilized pQTL datasets as exposure factors and GWAS for asthma and sepsis as outcomes to identify causal proteins and genes associated with both conditions. Inverse Variance Weighting (IVW) was applied to meta-analyze and combine the Wald ratios of causal effects for each SNP.34,37 Then, the MR-Egger38 and weighted median39 methods are used to supplement IVW. According to different validity assumptions, different methods are used to obtain MR Estimates. The application of the IVW method assumes that all SNPs are valid IVs, allowing for accurate estimation of causal effects. The MR-Egger method, on the other hand, evaluates directed pleiotropy by interpreting the intercept as an estimate of the average pleiotropy of the genetic variation. In contrast, the weighted median method offers higher accuracy, with smaller standard deviation, and remains robust in the presence of horizontal pleiotropy. Even when up to 50% of the genetic variations are invalid IVs, the weighted median provides consistent estimates.40
Sensitivity Analysis
The causal direction of each extracted SNP with respect to the exposure and the outcome was tested using the MR Steiger filtering method.41 This method calculates the variance explained by the instrumental SNPs in both the exposure and the outcome, and tests whether the variance in the outcome is less than that in the exposure. A “TRUE” MR Steiger indicates that the causal direction is consistent with the hypothesis, whereas a “FALSE” result implies the reverse causality, ie, the SNP influences the outcome rather than the exposure; such SNPs are therefore excluded.
Cochran’s Q statistic and funnel plots were used to assess the heterogeneity among SNPs.42,43 The MR-Egger intercept method38 and the MRPRESSO method44 were employed to detect horizontal pleiotropy. If outliers were identified and removed, the MR causal estimates were recalculated. If heterogeneity persisted despite their removal, a random effects model was applied to assess the stability of the results, as this model is less sensitive to the influence of weaker SNP-exposure associations. A leave-one-out analysis was conducted to evaluate the impact of each SNP on the overall causal estimate.
Identification of DEGs
All raw datasets were preprocessed, including background adjustment and normalization, using the “affy” package in R software (version 4.3.0). Probes were mapped to gene symbols based on the GPL10558 and GPL570 platforms (Affymetrix Human Genome U133 Plus 2.0 Array). For genes matched by multiple probes, the mean value was selected as the gene expression level. The “Limma” R package was then used to identify DEGs in both the asthma and sepsis datasets. The thresholds for DEGs were set as an adjusted p-value < 0.05 and |log2 fold change (FC)| ≥ 0.5. The “ggplot2” and “pheatmap” R packages were employed to create volcano plots and clustering heatmaps for DEGs. Venn diagrams were generated using the “Venn” package to identify common DEGs between asthma and sepsis.
2.7 Gene Ontology (GO) annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) cg pathway enrichment analysis.
DAVID (Database for Annotation, Visualization and Integrated Discovery) 6.8 (https://david.ncifcrf.gov/tools.jsp) is a gene function classification tool used to perform enrichment analysis on common genes (cg) for implementing GO.45 The enriched GO terms are categorized into Biological Process (BP), Cellular Component (CC), and Molecular Function (MF) ontologies. To understand the biological functions of common genes (cg), we uploaded the common genes (cg) to the DAVID database (a gene function classification tool, https://david.ncifcrf.gov/tools.jsp).46 The significant enrichment thresholds for GO and KEGG analyses were set as p < 0.05 and a count ≥ 2.
Expression Analysis and Diagnostic Evaluation of Candidate Biomarkers
The “ggplot2” package was employed to visualize the expression levels of candidate biomarkers in both the control and disease groups (p < 0.05). The Area Under Curve (AUC) of the Receiver Operating Characteristic (ROC) curve was calculated, and the “pROC” package was used to assess the diagnostic accuracy of the candidate biomarkers.
Immune Infiltration Analysis
The datasets we used were GSE143303 (asthma) and GSE28750 (sepsis). CIBERSORT, a deconvolution algorithm, was employed to determine the composition of 22 immune cell types based on gene expression data. The “CIBERSORT” software package was used to analyze immune cell infiltration, while the “ggplot2”, “corplot”, and “vioplot” packages were utilized for visualizing the results. Spearman correlation analysis was performed to assess the relationships between immune cells and candidate biomarkers.
Evaluation of Candidate Drugs
The enrichment platform was used to explore the relationships between drug molecules and hub genes, with data sourced from the Drug Signature Database (DSigDB, https://tanlab.ucdenver.edu/DSigDB). Currently, this database contains 22,527 gene sets, including 17,389 drugs, covering 19,531 genes.47 The three-dimensional structures of drug target molecules were obtained from PubChem (https://pubchem.ncbi.nlm.nih.gov/)48 and saved in the SDF file format. The 3D structures of key gene targets were retrieved from the Protein Data Bank (PDB; https://www.rcsb.org/) database49 and saved as files in the PDB format. AutoDock Vina50 and PyMol 1.8.1 (Schrödinger, http://www.pymol.org/) were used for molecular docking and visualization analysis to display the interaction forces between drug molecules and gene targets.
Methodology of Clinical Characteristics Analysis
The demographic characteristics of the study population were systematically collected and analyzed, including age, sex, weight, and BMI, along with vital signs (blood pressure) measured at admission. For continuous variables, all data were presented in the form of mean ± standard deviation (SD), and for categorical variables, they were presented in the form of counts (percentages). One-way analysis of variance (ANOVA) was used to evaluate the differences of continuous variables among the three groups, and the chi-square test was applied to categorical variables.
Assessment of Diagnostic Biomarkers via ROC Curves
ROC curves were generated to assess the diagnostic performance of candidate biomarkers. Sensitivity (true - positive rate) was plotted on the y - axis, and 1 - Specificity (false - positive rate) was plotted on the x - axis. The Area Under the Curve (AUC) values were calculated to quantify the overall diagnostic accuracy of the biomarkers. For each biomarker, multiple threshold settings were considered to obtain different pairs of sensitivity and 1 - Specificity values, which were then connected to form the ROC curves.
CXCL8 Levels in Different Patient Groups
Samples were collected from normal subjects, asthma patients, and sepsis patients. We measured CXCL8 concentrations in samples from the three groups using an enzyme-linked immunosorbent assay (ELISA) kit. The data were presented as a combination of scatter plots and box plots. The box plots display the median, inter - quartile range (IQR), and the whiskers typically extend to 1.5 times the IQR. Dots and triangles represent individual data points for each group. One - way ANOVA was used to compare the CXCL8 levels among the three groups.
Statistical Analysis
Statistical analyses were performed using R software (version 4.3.0), with a p-value threshold of < 0.05 considered statistically significant. The “TwoSampleMR” package was utilized for conducting the MR analysis.51 MR-Pleiotropy RESidualS and Outliers and Robust Adjusted Profile Score were computed using the R packages “MRPRESSO” and “MR.RAPS” respectively. Additionally, we performed a PhenoScanner search to evaluate all known phenotypes associated with the genetic instruments used in our analysis. For the screening of DEGs, module genes, enrichment analysis, transcription factor-gene networks, gene-microRNA networks, and immune infiltration analysis, a p-value threshold of less than 0.05 was considered statistically significant. We utilized SPSS software for descriptive statistics of the data and comparison of differences between groups. Prism software, on the other hand, was employed to create intuitive graphs to better present the analysis results.
Results
MR Analysis of pQTL in Relation to Asthma and Sepsis
MR analysis was conducted to investigate the relationship between pQTLs and asthma in an Icelandic population. This analysis identified 435 genetically determined proteins significantly associated with asthma, using a significance threshold of P < 0.05. The detailed results are provided in Supplementary Table S1. Similarly, an MR analysis was performed to examine the association between pQTLs and sepsis in the same population, revealing 1,385 genetically determined proteins significantly linked to sepsis, also with a P < 0.05 threshold. The comprehensive findings are outlined in Supplementary Table S2. To identify overlapping genetic factors between asthma and sepsis, the intersection of genes significantly associated with both conditions was determined, which comprised 141 genes, offering insights into potential shared genetic mechanisms between asthma and sepsis. The detailed list of these intersecting genes is available in Supplementary Table S3.
Sensitivity Analysis
Sensitivity analyses were conducted to examine and address the potential pleiotropy in causal estimates. Cochran’s Q test and funnel plots revealed no significant heterogeneity or asymmetry in the causal relationships among these SNPs (Supplementary Table S4, Supplementary Figure S1, and S2). The influence of each SNP on the overall causal estimate was validated through leave-one-out analysis (Supplementary Figure S3 and S4). By systematically removing each SNP and re-performing the MR analysis on the remaining SNPs, we observed consistent results, which indicate that the inclusion of all SNPs contributed to a significant causal association.
Functional Enrichment Analysis of Intersected Genes from pQTL Analysis
Using GO and KEGG pathway enrichment analyses, we explored the biological characteristics and pathways of the 141 intersected genes identified through pQTL analysis. The GO analysis yielded 122 terms, including 71 BP, 21 CC, and 30 MF. The top 15 most significant terms, comprising 5 each from BP, CC, and MF, are presented here. In BP, the main enrichments were observed in multicellular organism development (P<0.001), neutrophil chemotaxis (P<0.001), positive regulation of kinase activity (P<0.001), positive regulation of cell migration (P<0.001), and response to virus (P<0.001). For CC, the primary enrichments were in extracellular region (P<0.001), extracellular space (P<0.001), cell surface (P<0.001), plasma membrane (P<0.001), and receptor complex (P<0.001). In MF, the main enrichments included transmembrane receptor protein tyrosine kinase activity (P<0.001), chemokine activity (P<0.001), CCR chemokine receptor binding (P<0.001), insulin-like growth factor binding (P<0.01), and serine-type endopeptidase activity (P<0.01).
The KEGG pathway enrichment analysis identified 28 pathways, with the top 5 most significant ones: cytokine-cytokine receptor interaction (P<0.001), viral protein interaction with cytokine and cytokine receptor (P<0.001), JAK-STAT signaling pathway (P<0.001), Measles (P<0.01), and Toll-like receptor signaling pathway (P<0.01). The results from the GO and KEGG enrichment analyses are depicted in Figure 2, with complete details provided in Supplementary Table S5. These findings suggest a strong association between asthma and sepsis in relation to inflammatory responses and immune processes.
 |
Figure 2 The functional enrichment analysis of intersection genes obtained from Protein Quantitative Trait Loci analysis between asthma and sepsis. |
Identification of DEGs
In the asthma dataset (GSE143303), a total of 247 DEGs were identified, comprising 127 upregulated genes and 120 downregulated genes (Figure 3A). In the sepsis dataset (GSE28750), a total of 2878 DEGs were identified, including 1376 upregulated genes and 1482 downregulated genes (Figure 3B). Cluster heatmaps based on all DEGs in asthma and sepsis, respectively (Figure 3C and D). Using a Venn diagram, a total of 65 common DEGs were identified between asthma and sepsis (Figure 3E).
 |
Figure 3 The identification of DEGs. (A and B) Volcano plots of all DEGs in asthma (GSE143303) and sepsis (GSE28750). (C and D) Heatmaps of all DEGs in asthma (GSE143303) and sepsis (GSE28750). (E) Venn diagram of the intersection of DEGs in asthma and sepsis. |
Functional Enrichment Analysis of Common DEGs
Functional enrichment analysis using GO and KEGG pathways was performed to explore the biological characteristics and pathways of the 65 common DEGs. The GO analysis identified 55 terms, including 35 BP, 11 CC, and 9 MF. The 15 most significant terms, comprising 5 BP, 5 CC, and 5 MF, were presented. BP was primarily enriched in inflammatory responses (P<0.001), response to lipopolysaccharide (P<0.001), neutrophil activation (P<0.01), apoptotic processes (P<0.01), and cellular response to lipopolysaccharide (P<0.01). CC was mainly enriched in extracellular region (P<0.001), extracellular space (P<0.01), extracellular exosome (P<0.01), collagen-containing extracellular matrix (P<0.01), and external side of the plasma membrane (P<0.01). MF was enriched in protein binding (P<0.001), calcium-dependent protein binding (P<0.01), identical protein binding (P<0.01), signaling receptor binding (P<0.01), and protein homodimerization activity (P<0.01).
The KEGG pathway enrichment analysis highlighted 12 pathways, with the five most significant presented here: Cell cycle (P<0.01), Hepatitis B (P<0.01), Diabetic cardiomyopathy (P<0.05), Human T-cell leukemia virus 1 infection (P<0.05), and Oocyte meiosis (P<0.05). The results of the GO and KEGG enrichment analyses are shown in Figure 4, with the complete findings listed in Supplementary Table S6. These findings suggest a strong association between asthma and sepsis in relation to inflammatory responses and immune processes.
 |
Figure 4 The functional enrichment analysis of common DEGs between asthma and sepsis. |
Diagnostic Value and Validation of Candidate Biomarkers
By intersecting the 141 genes identified from the pQTL analysis with the 65 DEGs, we identified CXCL8 as the overlapping gene, as shown in Figure 5. To validate whether CXCL8 functions as a diagnostic biomarker for both asthma and sepsis, we further analyzed its expression levels and diagnostic performance. As illustrated in Figure 6A, the expression level of CXCL8 was significantly higher in the asthma disease group compared to the control group. In contrast, in the sepsis dataset, the expression level of CXCL8 was significantly lower in the disease group than in the control group (Figure 6B). ROC curve analysis demonstrated that CXCL8 exhibits excellent predictive performance for both asthma and sepsis. (Figure 6C and D): for the asthma dataset (GSE143303), CXCL8 achieved an AUC of 0.833 (95% CI: 0.707–0.930); for the sepsis dataset (GSE28750), CXCL8 achieved an AUC of 0.840 (95% CI: 0.670–0.960).
 |
Figure 5 The intersection of results obtained from Protein Quantitative Trait Loci analysis and differential expression gene analysis. |
 |
Figure 6 Continued. |
 |
Figure 6 The evaluation of the diagnostic value of the candidate biomarkers in the discovery and validation datasets. (A) The expression difference of the shared gene in the asthma (GSE143303) discovery dataset. (B) The expression difference of the shared gene in the sepsis (GSE28750) discovery dataset. (C) ROC curve of the shared gene in the asthma (GSE143303) discovery dataset, and (D) The ROC curve of the shared gene in the sepsis (GSE28750) discovery dataset. (E) The expression difference of the shared gene in the asthma (GSE147878) validation dataset. (F) The expression difference of the shared gene in the sepsis (GSE57065) validation dataset, (G) The ROC curve of the shared gene in the asthma (GSE147878) validation dataset. (H) The ROC curve of the shared gene in the sepsis (GSE57065) validation dataset. |
To further assess the accuracy of the candidate biomarkers, we validated them in two external datasets: GSE147878 for asthma and GSE57065 for sepsis. Consistent with previous findings, the expression of this biomarker was significantly upregulated in the disease group of the validated asthma dataset (Figure 6E). In the validated sepsis dataset, the biomarker expression was significantly lower in the disease group compared to the control group (Figure 6F). ROC curve analysis further confirming its strong diagnostic value (Figure 6G and H). In summary, these findings suggest that CXCL8 holds promise as a diagnostic biomarker for both asthma and sepsis.
Immune Cell Infiltration Analysis
Asthma and sepsis are both immune-mediated diseases; therefore, CIBERSORT was employed to analyze the proportions of immune infiltrating cells in these conditions. Figure 7A and B depict the distribution of 22 types of immune cells in each sample for asthma and sepsis, respectively. The correlation heatmaps between individual immune cells are shown in Figure 7C and D, with Figure 7C representing asthma and Figure 7D representing sepsis. Figure 7 Continued. Figure 7 The immune cell infiltration analysis. (A–D) The percentages of 22 immune cells identified by the CIBERSORT algorithm in the asthma dataset (GSE143303) and the sepsis dataset (GSE28750). (E and F) Box plots showing the proportions of immune cells in disease and control groups of asthma and sepsis patients. (G) The correlation between CXCL8 and infiltrating immune cells in asthma and normal samples. (H) The correlation between CXCL8 and infiltrating immune cells in sepsis and normal samples.

Furthermore, we observed that T cells CD4 memory resting, T cells regulatory (Tregs), Macrophages M2, and Mast cells resting exhibited dysregulated levels between normal and asthma samples (Figure 7E). Similarly, B cells memory, Plasma cells, T cells CD8, T cells CD4 naïve, T cells follicular helper, Macrophages M0, Macrophages M1, Dendritic cells resting, Mast cells resting, and Neutrophils showed dysregulated levels between normal and sepsis samples (Figure 7F).
Additionally, we conducted a correlation analysis between the shared biomarker and immune cells. The results revealed that in asthma samples, CXCL8 was negatively correlated with T regulatory cells (Tregs) and resting mast cells, while showing a positive correlation with activated memory CD4+ T cells and neutrophils (Figure 7G). In sepsis samples, CXCL8 was negatively correlated with neutrophils, Macrophages M2, and Macrophages M0, and positively correlated with T cells CD8, T cells CD4 naïve, and T cells CD4 memory activated (Figure 7H). These findings indicate that CXCL8 is closely related to immune cells such as macrophages.
Identification of Candidate Drugs
We utilized the DSigDB database on the Enrichr platform to identify potential drugs targeting the CXCL8 gene, resulting in 608 gene-drug associations, with detailed findings provided in Supplementary Table S7. We extracted the top 10 drugs ranked by adjusted p-values, including GLYOXAL CTD 00006046, Leupeptin CTD 00001574, 6401–97-4 CTD 00000925, Chromium (III) Chloride CTD 00001073, glutaraldehyde CTD 00006033, amoxicillin CTD 00005390, Diatrizoic acid CTD 00005787, Beclomethasone 17-monopropionate CTD 00002649, proscillaridin CTD 00006639, and ibuprofen TTD 00008500 (Table 3).
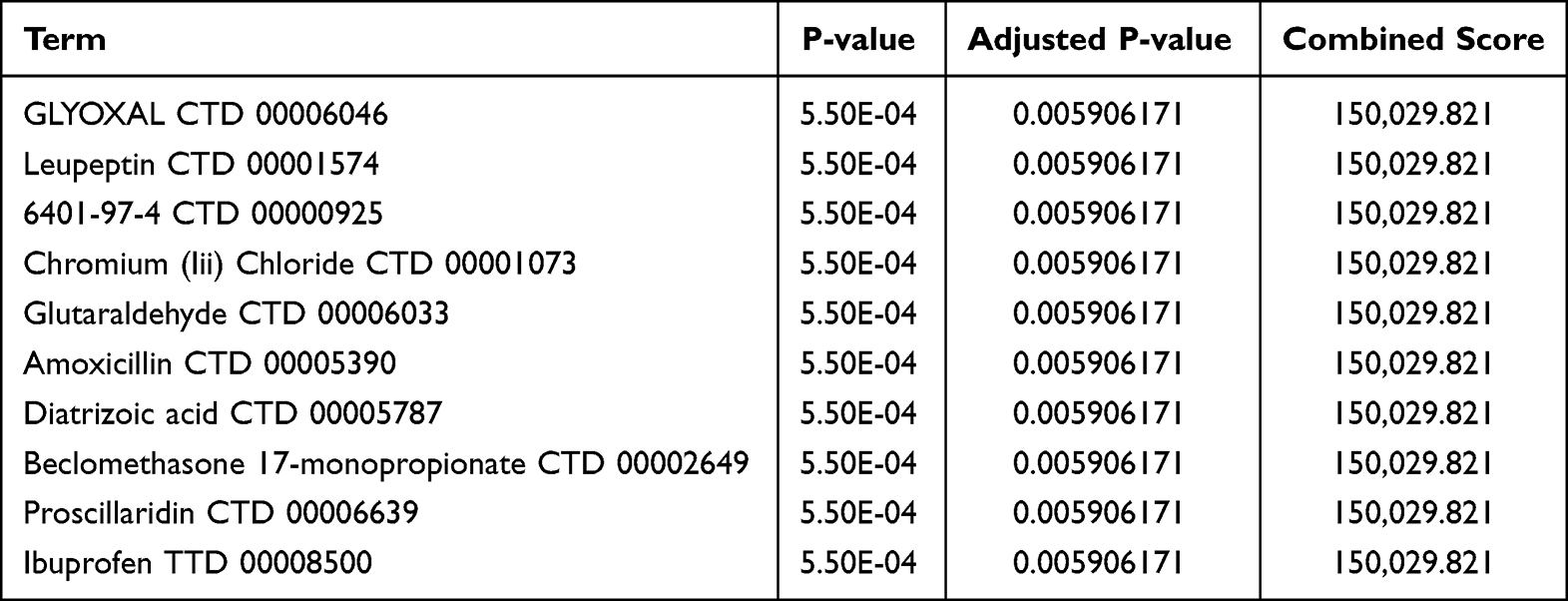 |
Table 3 The Top 10 Predicted Drug Compounds |
The molecular targets of the screened drugs and their corresponding genes were subjected to molecular docking. Protein structures of the gene targets were retrieved from the PDB database, while drug molecule ligands were obtained from PubChem for docking analysis. The molecular docking results showed that the binding energies of the ligands 6401–97-4 CTD 00000925, amoxicillin CTD 00005390, Diatrizoic acid CTD 00005787, proscillaridin CTD 00006639, and ibuprofen TTD 00008500 with potential gene targets were less than −4.5 KJ/mol, indicating low conformational energy and strong binding activity of these drug ligands with the gene targets (Table 4). Notably, Leupeptin CTD 00001574 and Chromium (III) Chloride CTD 00001073 did not have their three-dimensional structures found in PubChem, and the docking results for Beclomethasone 17-monopropionate CTD 00002649 showed that the drug molecule did not form hydrogen bonds with the corresponding gene. Therefore, only 7 drug targets were validated in the molecular docking process, as shown in Figure 8.
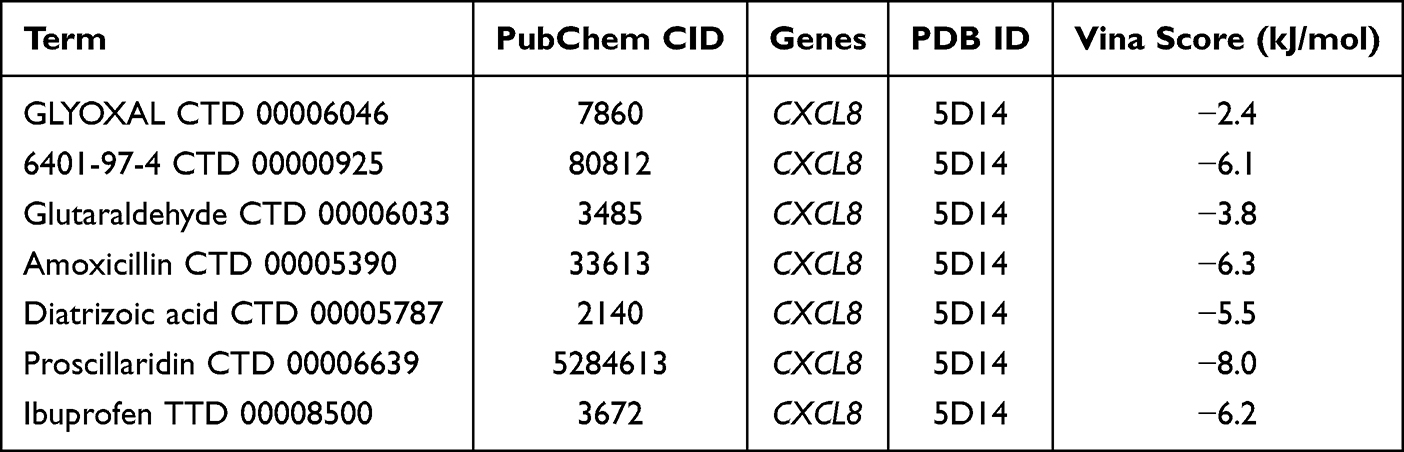 |
Table 4 The Corresponding IDs and Docking Binding Energies of Each Drug Ligand with Potential Gene Targets |
 |
Figure 8 The schematic diagrams of molecular docking for the 7 drugs. |
Demographic and Clinical Feature Differences Among Groups
The study compared clinical characteristics among normal subjects (n=90), asthma patients (n=106), and sepsis patients (n=99). The specific results of clinical data analysis are shown in Table 5. Significant differences were observed across groups. Age showed that normal subjects had a mean age of 35.21±7.804 years, younger than asthma patients (54.34±14.546 years) and sepsis patients (48.63±11.353 years) (p<0.001). The proportion of individuals aged ≥ 65 years was 0% in normal subjects, 25.47% in asthma patients, and 14.14% in sepsis patients (p<0.001). Regarding gender, the percentage of females was 70% in normal subjects, 37.74% in asthma patients, and 76.77% in sepsis patients (p<0.001). Mean weight and BMI also varied significantly among the three groups (p<0.001). For vital signs upon admission, both systolic and diastolic blood pressure were significantly different across the groups (p<0.001).
 |
Table 5 Clinical Characteristics and Biomarker Levels of Normal Subjects, Asthma Patients, and Sepsis Patients |
Diagnostic Performance of Biomarkers and CXCL8 Expression in Sepsis and Asthma
The diagnostic performance of biomarkers was evaluated via ROC curves. Curve A exhibited an Area Under the Curve of 0.999, with a specificity of 0.99 and a sensitivity of 1 (Figure 9A). Curve B had an AUC of 0.998, a specificity of 1, and a sensitivity of 0.022 (Figure 9B). These findings indicated that the biomarkers demonstrated high diagnostic accuracy under different performance metrics.
 |
Figure 9 Analysis of ROC curves of diagnostic biomarkers for sepsis (A) and asthma (B). |
Furthermore, the expression levels of CXCL8 were measured. As depicted in the box - and - whisker plot, normal subjects had the lowest CXCL8 levels (22.85±3.622 pg/mL), followed by asthma patients (59.33±36.99 pg/mL), and sepsis patients had the highest levels (147.40±99.82 pg/mL) (Figure 10). There were significant differences in CXCL8 levels among the three groups (p<0.001).
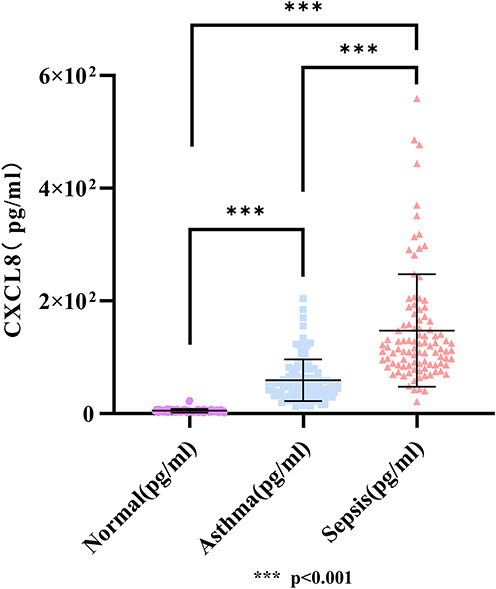 |
Figure 10 CXCL8 expression levels in normal subjects, asthma patients, and sepsis patients. |
Discussion
There are about 251 million patients suffering from asthma worldwide, with high morbidity and mortality,3 imposing a huge burden on the world economy.2 Sepsis is a life-threatening infectious disease with an extremely high mortality rate, closely associated with septic shock and multiple organ dysfunction.52 In recent years, the incidence of sepsis has increased and been consumed a large amount of medical and health resources.53 Research exploring the relationship between asthma and sepsis is currently limited, yet evidence suggests a connection between the two diseases, with a complex and not fully understood pathophysiological mechanism. Previous studies have identified potential mechanisms for asthma and sepsis individually; however, no study has yet examined their common underlying mechanisms. In this paper, we employ pQTL MR analysis, along with comprehensive bioinformatics and immune infiltration analyses, to investigate the shared transcriptional features between asthma and sepsis, as well as the role of immune cells in this relationship.
Initially, we conducted MR analysis of pQTLs in the Icelandic population, identifying 435 and 1,385 proteins significantly associated with asthma and sepsis, respectively, and pinpointing 141 intersection genes. We then identified 247 DEGs in the asthma dataset and 2,878 DEGs in the sepsis dataset, with 65 common DEGs. Functional enrichment analysis revealed that these genes were primarily involved in innate and adaptive immune responses, as well as inflammatory processes. Overall, these findings suggest that the regulation of immune responses and cytokine secretion may serve as a key link between asthma and sepsis. Through analysis of the intersection genes and DEGs identified in the MR study, we identified CXCL8 as a key diagnostic gene shared between asthma and sepsis. Compared to the control group, CXCL8 expression was significantly elevated in asthma patients, while its expression was markedly reduced in sepsis patients. Furthermore, ROC analysis demonstrated the potential of CXCL8 as a diagnostic biomarker for both diseases, with all AUC values exceeding 0.75, emphasizing its crucial role in the pathogenesis and diagnosis of asthma and sepsis. Additionally, based on the results of the clinical validation, we found that CXCL8 is indeed closely related to both asthma and sepsis. Using DSigDB, we predicted 10 potential drug compounds targeting this hub gene, seven of which exhibited stable binding in molecular docking, supporting the feasibility of these drugs as treatment options. These predicted drugs may serve as therapeutic targets for asthma and sepsis, though their functions and molecular pathways require further validation through cell experiments and clinical studies.
Our study identified the CXCL8 gene as a key intersection gene for both asthma and sepsis, suggesting its potential as a diagnostic biomarker for these conditions. Also known as the IL-8 gene, CXCL8 encodes a CXC chemokine, initially recognized as a low molecular weight protein with potent neutrophil chemotactic activity.54 Its product is a precursor protein containing 99 amino acids. After treatment, it produces an active protein, 77 amino acids in non-immune cells, and 72 amino acids in monocytes and macrophages.55 CXCL8 transmits signals by binding to CXCR1 and CXCR2 G protein-coupled receptors. The CXCL8 gene is expressed in a variety of cells and is closely related to inflammatory responses, tumor development, and immune regulation.55 Previous studies have shown that the CXCL8-CXCR1/2 axis plays an important role in the pathogenesis of inflammatory diseases such as asthma.56 Some small molecule CXCR1/2 inhibitors, CXCL8 release inhibitors, and CXCL8 and CXCR1/2 neutralizing antibodies are in advanced clinical trials for asthma treatment,56 providing new ideas for the clinical treatment of asthma. For neonatal asthma, a decrease in CXCL8 is associated with an increased risk of early asthma endpoints.57 Recent studies have found that IL-8/CXCL8 is overexpressed in lung tissues of patients with severe asthma and lung tissues of asthmatic mouse models, which is a potential target for the treatment of severe asthma.58 The CXCL8 gene is also closely related to sepsis, and IL-8/CXCL8 is significantly associated with the risk of sepsis.59,60 Currently, numerous studies have investigated the relationship between CXCL8 expression, its associated pathways, and sepsis-related complications. Specifically, CXCL8 expression has been identified as a risk factor for heart failure in patients with sepsis;61 Inhibition of CXCL8 expression in serum and kidney can prevent sepsis-induced acute kidney injury;62 CXCL8 is a key mediator in the progression of septic hepatitis and liver failure.63 The above results suggest that the CXCL8 gene is associated with both asthma and sepsis, highlighting its potential as a common diagnostic marker for these conditions and an important target for their treatment. In our study, GO-term, KEGG-pathway and immune-infiltration analyses revealed that both asthma and sepsis are tightly linked to immune-related cascades. Functional enrichment analyses revealed that the two datasets significantly share the core CC terms “extracellular region” and “extracellular space”. This indicates that, irrespective of disease context, CXCL8-mediated effects are exerted preferentially within the extracellular micro-environment, consistent with its role in regulating cytokine expression and providing a spatial basis for the shared pathogenesis of the two disorders. As a cytokine-encoding gene, CXCL8 plays a pivotal role in initiating and modulating these immune pathways.55 Taken together with the published evidence, it is reasonable to predict that CXCL8—identified here as a shared disease gene—exerts its effects on both disorders by regulating immunological processes and immune-cell functions (eg, macrophage), thereby representing a critical diagnostic and therapeutic target.
Macrophages are key immune cells whose primary functions include recognizing, engulfing, and digesting foreign microorganisms, dead cells, and other particulate matter. They play a crucial role in immune defense and the maintenance of tissue homeostasis.64 Macrophage polarization refers to the different functions and phenotypes of macrophages under different microenvironment stimulation, which are usually divided into M1 and M2 types.65 The former is involved in combating pathogens and mediating inflammatory responses through the secretion of pro-inflammatory factors and phagocytosis, while the latter promotes tissue repair and the resolution of inflammation by releasing anti-inflammatory factors. Macrophage polarization plays a crucial role in maintaining immune homeostasis, participating in immune regulation and tissue repair, and its dysregulation may be linked to the onset and progression of various diseases.66 Our study found that Macrophages M2 showed a dysregulated level between normal samples and asthma samples, and Macrophages M0 and Macrophages M1 showed a dysregulated level between normal samples and sepsis samples through immune cell infiltration analysis, indicating that asthma and sepsis are closely related to macrophage activation and polarization. Some relative studies have confirmed these results.22,23 As mentioned above, allergic asthma is related to airway inflammation and airway hyperresponsiveness, in which macrophages play an important role. Under different microenvironment stimulation, macrophages will polarize into M1 and M2 types and play different roles, which has a profound impact on the pathogenesis of asthma.67 The activation and polarization of macrophages also play an important role in the occurrence and development of sepsis, which can provide antibacterial defense for the body.68,69 In addition, the activation and polarization of macrophages are closely related to multiple sepsis complications, including acute lung injury,70,71 cardiomyopathy,72 liver injury73,74 and so on, suggesting macrophage activation and polarization may serve as common factors in the pathogenesis of both asthma and sepsis.
Additionally, our study identified the CXCL8 gene as a common diagnostic biomarker for both asthma and sepsis, which was negatively correlated with M0 and M2 macrophages. This finding further corroborates the role of macrophage activation and polarization in the pathophysiology of both diseases. CXCL8 has been shown to induce M2 macrophage polarization, and this polarization, in turn, promotes the continued expression and secretion of CXCL8, forming a positive feedback, which is closely related to gastric cancer,75 colorectal cancer,76 cervical cancer,77 pancreatic cancer,78 periodontitis79 and other systemic diseases. Based on our research findings, we hypothesize that this process may also be implicated in the onset and progression of asthma and sepsis.
The strength of this study lies in the use of Icelandic human plasma-derived pQTL data for MR analysis. This novel dataset minimizes the risk of errors associated with population duplication. Additionally, bioinformatics methods were employed to screen relevant genes, assess their expression levels, and evaluate their diagnostic efficacy. Potential therapeutic drugs targeting these genes were identified, and molecular simulations were performed, yielding high data accuracy and strong clinical relevance. Subsequently, we confirmed the accuracy and reliability of the aforementioned analysis results through clinical validation. However, some limitations exist. Firstly, the sample size of the dataset used is relatively small. Moreover, this study lacks data on patients with both asthma and sepsis. Future research should aim to collect larger datasets and incorporate comorbidity data. Additionally, the results require further validation through subsequent in vivo and in vitro experiments. More importantly, as a widely expressed cytokine-encoding gene, CXCL8 is markedly up- or down-regulated in many other disorders and can influence their onset and progression through multiple pathways, including COPD80–82 and pneumonia83,84 in the respiratory system. Serum IL-8 levels also serve as a shared diagnostic biomarker for both COPD and asthma.82 This overlap inevitably reduces its diagnostic specificity, so clinical signs and symptoms must be used to rule out confounding diseases before CXCL8 can be considered a reliable biomarker. Despite these limitations, the preliminary identification of these biomarkers provides a foundation for generating new hypotheses to guide future investigations.
This study is the first to explore the relationship between asthma and sepsis using bioinformatics approaches, yielding promising statistical results that align with existing evidence and demonstrate significant innovation. By employing a variety of analytical methods, this research explores genes and regulatory factors linked to asthma, sepsis, and immune cells, while predicting relevant cellular signaling pathways and potential therapeutic targets. These findings offer valuable research directions for both basic and clinical medical research in the future, provide bioinformatics evidence for asthma and sepsis-related studies, and propose new avenues for future research and drug development in this field.
Conclusion
In the MR analysis, we established a causal relationship between asthma and sepsis; however, further investigation is required to explore additional risk factors as potential mediators. Through GEO analysis, we identified two diagnostic genes, CXCL8, along with co-regulatory pathways and immune characteristics shared by asthma and sepsis. Additionally, we screened potential therapeutic drugs targeting these diagnostic genes, offering a novel strategy for the clinical management of asthma and sepsis. Immune cell infiltration analysis revealed a strong association between macrophage activation and polarization with both asthma and sepsis. Clinical validation has confirmed that CXCL8 is indeed closely related to both asthma and sepsis. These findings provide a theoretical framework for understanding the molecular mechanisms underlying asthma complicated by sepsis, encompassing genetic factors, signaling pathways, and immune infiltration.
Data Sharing Statement
The datasets generated and/or analysed during the current study are available in the GWAS database (https://gwas.mrcieu.ac.uk/), the UK Biobank (https://www.nature.com/collections/bpthhnywqk), GEO repository (https://www.ncbi.nlm.nih.gov/gds/?term=), PubChem (https://pubchem.ncbi.nlm.nih.gov/) and PDB (https://www.rcsb.org/).
Statement of Ethics
The study protocol and consent forms were approved by the Ethics Committee of the Third Xiangya Hospital (No. 23911), with all procedures followed by the World Medical Association Declaration of Helsinki, and all essential permissions were obtained from the government and health commission. All personal information was anonymized during analysis and reporting to ensure confidentiality and privacy.
Acknowledgment
The authors gratefully acknowledge the voluntary participation of all the study subjects.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by the National Natural Science Foundation of China (No.82303133, No. 82400873); China Postdoctoral Science Foundation (No. 2022M723559, No. GZB20240866); Natural Science Foundation of Hunan Province (No.2021JJ40951, No. 2025JJ60551); Project of Hunan Health Commission (No. B202307017799, No. B202309018525).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
References
1. Gans MD, Gavrilova T. Understanding the immunology of asthma: pathophysiology, biomarkers, and treatments for asthma endotypes. Paediatr Respir Rev. 2020;36:118–127. doi:10.1016/j.prrv.2019.08.002
2. Hu LR, Scanlon P, Miller K, et al. National center for health statistics’ 2019 research and development survey, RANDS 3. Vital Health Stat. 2023;(65):1–55.
3. Stern J, Pier J, Litonjua AA. Asthma epidemiology and risk factors. Semin Immunopathol. 2020;42(1):5–15. doi:10.1007/s00281-020-00785-1
4. Agache I, Eguiluz‐Gracia I, Cojanu C, et al. Advances and highlights in asthma in 2021. Allergy. 2021;76(11):3390–3407. doi:10.1111/all.15054
5. Ntontsi P, Photiades A, Zervas E, Xanthou G, Samitas K. Genetics and epigenetics in asthma. Int J Mol Sci. 2021;22(5):2412. doi:10.3390/ijms22052412
6. Wenzel SE. Severe adult asthmas: integrating clinical features, biology, and therapeutics to improve outcomes. Am J Respir Crit Care Med. 2021;203(7):809–821. doi:10.1164/rccm.202009-3631CI
7. Miller RL, Grayson MH, Strothman K. Advances in asthma: new understandings of asthma’s natural history, risk factors, underlying mechanisms, and clinical management. J Allergy Clin Immunol. 2021;148(6):1430–1441. doi:10.1016/j.jaci.2021.10.001
8. Cazzola M, Rogliani P, Ora J, Calzetta L, Matera MG. Asthma and comorbidities: recent advances. Pol Arch Intern Med. 2022;132(4). doi:10.20452/pamw.16250
9. Pofi R, Caratti G, Ray DW, Tomlinson JW. Treating the side effects of exogenous glucocorticoids; can we separate the good from the bad? Endocr Rev. 2023;44(6):975–1011. doi:10.1210/endrev/bnad016
10. Schoettler N, Strek ME. Recent advances in severe asthma: from phenotypes to personalized medicine. Chest. 2020;157(3):516–528. doi:10.1016/j.chest.2019.10.009
11. Liu AC, Patel K, Vunikili RD, et al. Sepsis in the era of data-driven medicine: personalizing risks, diagnoses, treatments and prognoses. Brief Bioinform. 2020;21(4):1182–1195. doi:10.1093/bib/bbz059
12. Singer M, Deutschman CS, Seymour CW, et al. The third international consensus definitions for sepsis and septic shock (sepsis-3). JAMA. 2016;315(8):801–810. doi:10.1001/jama.2016.0287
13. Clere-Jehl R, Mariotte A, Meziani F, Bahram S, Georgel P, Helms J. JAK-STAT targeting offers novel therapeutic opportunities in sepsis. Trends Mol Med. 2020;26(11):987–1002. doi:10.1016/j.molmed.2020.06.007
14. Rudd KE, Johnson SC, Agesa KM, et al. Global, regional, and national sepsis incidence and mortality, 1990-2017: analysis for the global burden of disease study. Lancet. 2020;395(10219):200–211. doi:10.1016/S0140-6736(19)32989-7
15. Klick B, Guins T. Sepsis in the urgent care setting. Curr Probl Pediatr Adolesc Health Care. 2021;51(2):100968. doi:10.1016/j.cppeds.2021.100968
16. Maslove DM, Tang B, Shankar-Hari M, et al. Redefining critical illness. Nat Med. 2022;28(6):1141–1148. doi:10.1038/s41591-022-01843-x
17. Ferrer R, Artigas A, Levy MM, et al. Improvement in process of care and outcome after a multicenter severe sepsis educational program in Spain. JAMA. 2008;299(19):2294–2303. doi:10.1001/jama.299.19.2294
18. Levy MM, Dellinger RP, Townsend SR, et al. The Surviving Sepsis Campaign: results of an international guideline-based performance improvement program targeting severe sepsis. Intensive Care Med. 2010;36(2):222–231. doi:10.1007/s00134-009-1738-3
19. Dellinger RP, Levy MM, Rhodes A, et al. Surviving sepsis campaign: international guidelines for management of severe sepsis and septic shock: 2012. Crit Care Med. 2013;41(2):580–637. doi:10.1097/CCM.0b013e31827e83af
20. Angus DC, van der Poll T. Severe sepsis and septic shock. N Engl J Med. 2013;369(9):840–851. doi:10.1056/NEJMra1208623
21. Laupland KB, Kirkpatrick AW, Delaney A. Polyclonal intravenous immunoglobulin for the treatment of severe sepsis and septic shock in critically ill adults: a systematic review and meta-analysis. Crit Care Med. 2007;35(12):2686–2692.
22. Arora S, Dev K, Agarwal B, Das P, Syed MA. Macrophages: their role, activation and polarization in pulmonary diseases. Immunobiology. 2018;223(4–5):383–396. doi:10.1016/j.imbio.2017.11.001
23. Liu Y-C, Zou X-B, Chai Y-F, Yao Y-M. Macrophage polarization in inflammatory diseases. Int J Biol Sci. 2014;10(5):520–529. doi:10.7150/ijbs.8879
24. Guo R-F, Ward PA. Role of C5a in inflammatory responses. Annu Rev Immunol. 2005;23(1):821–852. doi:10.1146/annurev.immunol.23.021704.115835
25. Hufnagl K, Pali-Schöll I, Roth-Walter F, Jensen-Jarolim E. Dysbiosis of the gut and lung microbiome has a role in asthma. Semin Immunopathol. 2020;42(1):75–93. doi:10.1007/s00281-019-00775-y
26. Adelman MW, Woodworth MH, Langelier C, et al. The gut microbiome’s role in the development, maintenance, and outcomes of sepsis. Crit Care. 2020;24(1):278. doi:10.1186/s13054-020-02989-1
27. Edwards SL. Pathophysiology of acid base balance: the theory practice relationship. Inten Criti Care Nurs. 2008;24(1):28–40. doi:10.1016/j.iccn.2007.05.003
28. Yang S, Guo J, Kong Z, et al. Causal effects of gut microbiota on sepsis and sepsis-related death: insights from genome-wide Mendelian randomization, single-cell RNA, bulk RNA sequencing, and network pharmacology. J Transl Med. 2024;22(1):10. doi:10.1186/s12967-023-04835-8
29. King EA, Davis JW, Degner JF, Marchini J. Are drug targets with genetic support twice as likely to be approved? Revised estimates of the impact of genetic support for drug mechanisms on the probability of drug approval. PLoS Genet. 2019;15(12):e1008489. doi:10.1371/journal.pgen.1008489
30. Nelson MR, Tipney H, Painter JL, et al. The support of human genetic evidence for approved drug indications. Nat Genet. 2015;47(8):856–860. doi:10.1038/ng.3314
31. Ferkingstad E, Sulem P, Atlason BA, et al. Large-scale integration of the plasma proteome with genetics and disease. Nat Genet. 2021;53(12):1712–1721. doi:10.1038/s41588-021-00978-w
32. Edgar R. Gene expression omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002;30(1):207–210. doi:10.1093/nar/30.1.207
33. Abecasis GR, Altshuler D, Auton A, et al. A map of human genome variation from population-scale sequencing. Nature. 2010;467(7319):1061–1073.
34. Hemani G, Zheng J, Elsworth B, et al. The MR-base platform supports systematic causal inference across the human phenome. Elife. 2018;7.
35. Li BB, Martin EB. An approximation to the F distribution using the chi-square distribution. Comput Stat Data Anal. 2002;40(1):21–26. doi:10.1016/S0167-9473(01)00097-4
36. Burgess S, Thompson SG. Avoiding bias from weak instruments in Mendelian randomization studies. Int J Epidemiol. 2011;40(3):755–764. doi:10.1093/ije/dyr036
37. Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013;37(7):658–665. doi:10.1002/gepi.21758
38. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32(5):377–389. doi:10.1007/s10654-017-0255-x
39. Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent estimation in mendelian randomization with some invalid instruments using a weighted median estimator. Genet Epidemiol. 2016;40(4):304–314. doi:10.1002/gepi.21965
40. Zhang Y, Liu Z, Choudhury T, Cornelis MC, Liu W. Habitual coffee intake and risk for nonalcoholic fatty liver disease: a two-sample Mendelian randomization study. Eur J Nutr. 2021;60(4):1761–1767. doi:10.1007/s00394-020-02369-z
41. Hemani G, Tilling K, Davey Smith G, Li J. Orienting the causal relationship between imprecisely measured traits using GWAS summary data. PLoS Genet. 2017;13(11):e1007081. doi:10.1371/journal.pgen.1007081
42. Tan J-S, Liu -N-N, Guo -T-T, Hu S, Hua L. Genetically predicted obesity and risk of deep vein thrombosis. Thromb Res. 2021;207:16–24. doi:10.1016/j.thromres.2021.08.026
43. Tan J-S, Ren J-M, Fan L, et al. Genetic predisposition of anti-cytomegalovirus immunoglobulin g levels and the risk of 9 cardiovascular diseases. Front Cell Infect Microbiol. 2022;12:884298. doi:10.3389/fcimb.2022.884298
44. Verbanck M, Chen C-Y, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. 2018;50(5):693–698. doi:10.1038/s41588-018-0099-7
45. Huang da W, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009;37(1):1–13. doi:10.1093/nar/gkn923
46. Huang da W, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4(1):44–57. doi:10.1038/nprot.2008.211
47. Yoo M, Shin J, Kim J, et al. DSigDB: drug signatures database for gene set analysis. Bioinformatics. 2015;31(18):3069–3071. doi:10.1093/bioinformatics/btv313
48. Wang Y, Suzek T, Zhang J, et al. PubChem BioAssay: 2014 update. Nucleic Acids Res. 2014;42(D1):D1075–82. doi:10.1093/nar/gkt978
49. Berman HM, Westbrook J, Feng Z, et al. The protein data bank. Nucleic Acids Res. 2000;28(1):235–242. doi:10.1093/nar/28.1.235
50. Trott O, Olson AJ. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. 2010;31(2):455–461. doi:10.1002/jcc.21334
51. Broadbent JR, Foley CN, Grant AJ, Mason AM, Staley JR, Burgess S. MendelianRandomization v0.5.0: updates to an R package for performing Mendelian randomization analyses using summarized data. Wellcome Open Res. 2020;5:252. doi:10.12688/wellcomeopenres.16374.2
52. Esposito S, De Simone G, Boccia G, De Caro F, Pagliano P. Sepsis and septic shock: new definitions, new diagnostic and therapeutic approaches. J Glob Antimicrob Resist. 2017;10:204–212. doi:10.1016/j.jgar.2017.06.013
53. Srzić I, Nesek Adam V, Tunjić Pejak D. SEPSIS DEFINITION: ’WHAT’S NEW IN THE TREATMENT GUIDELINES. Acta Clin Croat. 2022;61(Suppl 1):67–72. doi:10.20471/acc.2022.61.s1.11
54. Baggiolini M. CXCL8 — the first chemokine. Front Immunol. 2015;6:285. doi:10.3389/fimmu.2015.00285
55. Cambier S, Gouwy M, Proost P. The chemokines CXCL8 and CXCL12: molecular and functional properties, role in disease and efforts towards pharmacological intervention. Cell Mol Immunol. 2023;20(3):217–251. doi:10.1038/s41423-023-00974-6
56. Ha H, Debnath B, Neamati N. Role of the CXCL8-CXCR1/2 axis in cancer and inflammatory diseases. Theranostics. 2017;7(6):1543–1588. doi:10.7150/thno.15625
57. Chawes BL, Wolsk HM, Carlsson CJ, et al. Neonatal airway immune profiles and asthma and allergy endpoints in childhood. Allergy. 2021;76(12):3713–3722. doi:10.1111/all.14862
58. Yuliani FS, Chen J-Y, Cheng W-H, Wen H-C, Chen B-C, Lin C-H. Thrombin induces IL-8/CXCL8 expression by DCLK1-dependent RhoA and YAP activation in human lung epithelial cells. J Biomed Sci. 2022;29(1):95. doi:10.1186/s12929-022-00877-0
59. Han T, Liang T, Liu R, Quan Y. The association between interleukin-8 gene polymorphism and the risk of sepsis in older adults. J Orthop Surg Res. 2024;19(1):804. doi:10.1186/s13018-024-05296-5
60. Machado JR, Soave DF, da Silva MV, et al. Neonatal sepsis and inflammatory mediators. Mediators Inflamm. 2014;2014:269681. doi:10.1155/2014/269681
61. Zhu J, Song C, Cai T, et al. The relationship between serum CXCL8 and ET-1 expression levels and sepsis complicated with heart failure. Cardiol Res Pract. 2022;2022:8570486. doi:10.1155/2022/8570486
62. Zhou Y, Xu W, Zhu H. CXCL8(3–72) K11R/G31P protects against sepsis-induced acute kidney injury via NF-κB and JAK2/STAT3 pathway. Biol Res. 2019;52(1):29. doi:10.1186/s40659-019-0236-5
63. Liu N, Bauer M, Press AT. The immunological function of CXCR2 in the liver during sepsis. J Inflamm. 2022;19(1):23. doi:10.1186/s12950-022-00321-y
64. Varol C, Mildner A, Jung S. Macrophages: development and tissue specialization. Annu Rev Immunol. 2015;33(1):643–675. doi:10.1146/annurev-immunol-032414-112220
65. Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. 2008;8(12):958–969. doi:10.1038/nri2448
66. Anderson NR, Minutolo NG, Gill S, Klichinsky M. Macrophage-based approaches for cancer immunotherapy. Cancer Res. 2021;81(5):1201–1208. doi:10.1158/0008-5472.CAN-20-2990
67. Saradna A, Do DC, Kumar S, Fu Q-L, Gao P. Macrophage polarization and allergic asthma. Transl Res. 2018;191:1–14. doi:10.1016/j.trsl.2017.09.002
68. Patoli D, Mignotte F, Deckert V, et al. Inhibition of mitophagy drives macrophage activation and antibacterial defense during sepsis. J Clin Invest. 2020;130(11):5858–5874. doi:10.1172/JCI130996
69. Essandoh K, Li Y, Huo J, Fan G-C. MiRNA-mediated macrophage polarization and its potential role in the regulation of inflammatory response. Shock. 2016;46(2):122–131. doi:10.1097/SHK.0000000000000604
70. Jiao Y, Zhang T, Zhang C, et al. Exosomal miR-30d-5p of neutrophils induces M1 macrophage polarization and primes macrophage pyroptosis in sepsis-related acute lung injury. Crit Care. 2021;25(1):356. doi:10.1186/s13054-021-03775-3
71. Luo J, Wang J, Zhang J, et al. Nrf2 deficiency exacerbated CLP-induced pulmonary injury and inflammation through autophagy- and NF-κB/PPARγ-mediated macrophage polarization. Cells. 2022;11(23):3927. doi:10.3390/cells11233927
72. Chen X-S, Wang S-H, Liu C-Y, et al. Losartan attenuates sepsis-induced cardiomyopathy by regulating macrophage polarization via TLR4-mediated NF-κB and MAPK signaling. Pharmacol Res. 2022;185:106473. doi:10.1016/j.phrs.2022.106473
73. Jin G-L, Liu H-P, Huang Y-X, et al. Koumine regulates macrophage M1/M2 polarization via TSPO, alleviating sepsis-associated liver injury in mice. Phytomedicine. 2022;107:154484. doi:10.1016/j.phymed.2022.154484
74. Chen Y, Yang L, Li X. Advances in Mesenchymal stem cells regulating macrophage polarization and treatment of sepsis-induced liver injury. Front Immunol. 2023;14:1238972. doi:10.3389/fimmu.2023.1238972
75. Piao H, Fu L, Wang Y, et al. A positive feedback loop between gastric cancer cells and tumor-associated macrophage induces malignancy progression. J Exp Clin Cancer Res. 2022;41(1):174. doi:10.1186/s13046-022-02366-6
76. Shao Y, Lan Y, Chai X, et al. CXCL8 induces M2 macrophage polarization and inhibits CD8 + T cell infiltration to generate an immunosuppressive microenvironment in colorectal cancer. THE FASEB journal. 2023;37(10):e23173. doi:10.1096/fj.202201982RRR
77. Chen L, Shi V, Wang S, et al. SCCA1/SERPINB3 suppresses antitumor immunity and blunts therapy-induced T cell responses via STAT-dependent chemokine production. J Clin Invest. 2023;133(15). doi:10.1172/JCI163841
78. Liao C-Y, Li G, Kang F-P, et al. Necroptosis enhances ‘don’t eat me’ signal and induces macrophage extracellular traps to promote pancreatic cancer liver metastasis. Nat Commun. 2024;15(1):6043. doi:10.1038/s41467-024-50450-6
79. Gonzalez OA, Kirakodu SS, Nguyen LM, Ebersole JL. Gingival transcriptomic patterns of macrophage polarization during initiation, progression, and resolution of periodontitis. Clin Exp Immunol. 2023;211(3):248–268. doi:10.1093/cei/uxac122
80. Eyraud E, Maurat E, Sac-Epée JM, et al. Short-range interactions between fibrocytes and CD8(+) T cells in COPD bronchial inflammatory response. Elife. 2023;12.
81. Li X, Li J, Zhang Y, Zhang L. The role of IL-8 in the chronic airway inflammation and its research progress. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2021;35(12):1144–1148. doi:10.13201/j.issn.2096-7993.2021.12.020
82. Ding Q, Sun S, Zhang Y, et al. Serum IL-8 and VEGFA are two promising diagnostic biomarkers of asthma-COPD overlap syndrome. Int J Chron Obstruct Pulmon Dis. 2020;15:357–365. doi:10.2147/COPD.S233461
83. Chang Y, Bai M, You Q, Abdalla SS. Associations between serum interleukins (IL-1 β, IL-2, IL-4, IL-6, IL-8, and IL-10) and disease severity of COVID-19: a systematic review and meta-analysis. Biomed Res Int. 2022;2022(1):2755246. doi:10.1155/2022/2755246
84. Pius-Sadowska E, Niedźwiedź A, Kulig P, et al. CXCL8, CCL2, and CMV seropositivity as new prognostic factors for a severe COVID-19 course. Int J Mol Sci. 2022;23(19):11338. doi:10.3390/ijms231911338
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.