Back to Journals » Drug Design, Development and Therapy » Volume 19
Identification and Validation of Antidepressant Small Molecules Using Bioinformatics and Mouse Depression Models
Authors Qiao Y ![]() , Zhang X
, Zhang X ![]() , Chen H, Liang X, Guo J, Wang Q, Ding Y
, Chen H, Liang X, Guo J, Wang Q, Ding Y ![]() , Wei L
, Wei L ![]() , Bi H
, Bi H ![]() , Gao T
, Gao T
Received 1 May 2025
Accepted for publication 13 August 2025
Published 20 August 2025 Volume 2025:19 Pages 7161—7183
DOI https://doi.org/10.2147/DDDT.S537918
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Tuo Deng
Yajun Qiao,1– 4,* Xingfang Zhang,2,5,6,* Hanxi Chen,2,5 Xinxin Liang,1– 4 Juan Guo,1,3 Qiannan Wang,2,5 Yi Ding,6 Lixin Wei,2,5 Hongtao Bi,2,4,5 Tingting Gao1,3
1School of Psychology, Chengdu Medical College, Chengdu, Sichuan, People’s Republic of China; 2CAS Key Laboratory of Tibetan Medicine Research, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining, Qinghai, People’s Republic of China; 3Department of Psychiatry, the People’s Hospital of Jiangmen, Southern Medical University, Jiangmen, Guangdong, People’s Republic of China; 4University of Chinese Academy of Sciences, Chinese Academy of Sciences, Beijing, People’s Republic of China; 5Qinghai Provincial Key Laboratory of Tibetan Medicine Pharmacology and Safety Evaluation, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining, Qinghai, People’s Republic of China; 6Department of Pharmacy, Xijing Hospital, Fourth Military Medical University, Xi’an, Shanxi, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hongtao Bi, Qinghai Provincial Key Laboratory of Tibetan Medicine Pharmacology and Safety Evaluation, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, 23 Xinning Road, Xining, 810008, People’s Republic of China, Tel +86-971-6143765, Fax +86-971-6143765, Email [email protected] Tingting Gao, School of Psychology, Chengdu Medical College, 783 Xindu Road, Chengdu, 610500, People’s Republic of China, Tel +86-28-62739705, Fax +86-28-62739705, Email [email protected]
Background: Depression, a prevalent psychiatric disorder with limited effective treatments, can be addressed by repurposing existing small molecules via bioinformatics as a promising approach, though previous studies using tools like CMAP and GEO have successfully identified candidate drugs for neuropsychiatric disorders, few have combined in silico predictions with in vivo validation for it.
Objective: The aim of this study was to employ bioinformatics and in vivo experimental validation to mine potential antidepressant small molecule compounds.
Methods: This study utilized data from the GEO database, employing bioinformatics analysis methods to analyze the dataset. The CMAP platform was used to deeply explore potential antidepressant small-molecule compounds. In vivo experiments validated the antidepressant effects of the small-molecule compounds on a chronic restraint stress mouse model.
Results: This study identified 311 differentially expressed genes (DEGs) from GSE182193—associated with the PI3K-Akt, MAPK, and neurotrophic factor signaling pathways, with key genes identified via Weighted Gene Co-expression Network Analysis (WGCNA) and immune correlation analysis—and screened 5 candidate compounds via CMAP, among which pyrimethamine, pifithrin-mu, and mibefradil significantly improved depressive behaviors in chronic restraint stress (CRS) model mice by regulating key protein expression in the PI3K-Akt and neurotrophic factor signaling pathways, as shown by a 33.44%– 60.32% increase in movement distance in the open field test (P < 0.01 to P < 0.001) and a 20.25%– 30.19% decrease in immobility time in the forced swim test (P < 0.01 to P < 0.001).
Conclusion: This study shows that pyrimethamine, pifithrin-mu, and mibefradil can regulate key proteins in the PI3K-Akt and neurotrophic factor pathways, improving depressive behaviors in mice and indicating their potential in alleviating depression; additionally, bioinformatics-driven repurposing of existing drugs for antidepressant discovery is more efficient than de novo development, and this study provides an exploratory demonstration of this.
Keywords: depression, bioinformatics, pyrimethamine, pifithrin-mu, mibefradil
Introduction
Depression is a common mental disorder that can often manifest as a depressed mood, impaired cognitive functioning, and loss of enjoyment. Its physical manifestations often include symptoms such as fatigue and sleep disturbance.1,2 In the worst cases, depression can lead to suicide,3,4 which has tragically claimed more than 700,000 lives.5 Especially in recent years, owing to the COVID-19 outbreak, reasons such as company bankruptcy and collapse, worker unemployment, and home isolation have led to a breakdown of psychological defenses, which has resulted in a dramatic increase in the incidence of depression.6 A study published in The Lancet entitled “Global prevalence and burden of depressive and anxiety disorders in 204 countries and territories in 2020 due to the COVID-19 pandemic” reported that the number of people with depression increased by 27.6% globally following the COVID-19 pandemic.7 This means that the pandemic has increased the number of depressed people worldwide by a quarter. As a result, depression has now become a serious mental disorder worldwide and has attracted much attention from scientists.
In research on the pathogenesis of depression,8 although several hypotheses have been proposed—including the monoamine neurotransmitter hypothesis,9 oxidative stress dysfunction,10 cytokine abnormalities,11 neurotrophic deficiency,12,13 HPA axis abnormalities, and the ‘gut–brain axis’14,15—their molecular links remain poorly defined. Notably, HPA axis hyperactivity is known to suppress neurotrophic factor expression,16 while gut-brain axis dysfunction may impair PI3K-Akt signaling via inflammatory cytokines,15 both of which overlap with the pathways targeted in this study. Thus, exploring small molecules that regulate these intersecting pathways (PI3K-Akt, MAPK, neurotrophins) could bridge these fragmented hypotheses. The onset of depression is closely related to the physiological, psychological, and social environment,17 but further research is needed. Currently, many types of antidepressants are available, such as monoamine oxidase inhibitors, tricyclic antidepressants, 5-hydroxytryptamine reuptake inhibitors, and 5-hydroxytryptamine and norepinephrine reuptake inhibitors. However, many antidepressants are ineffective and are even associated with serious adverse effects.18 According to the literature, current antidepressants provide relief in only 30%–40% of patients.19,20 More effective drugs with few side effects are needed to treat depression, but developing new drugs is time-consuming and costly. Repurposing old drugs plays an important role in the treatment of many diseases. For example, arsenic trioxide was used in ancient China to treat maladies such as hemorrhoids; current research has shown that it can be used to treat acute promyelocytic leukemia.21 Thalidomide was originally used to treat pregnancy vomiting in pregnant women, but recent studies have shown that its inhibitory effect on the inflammatory response can be applied in many diseases.22 In view of the abovementioned examples, the “new use of old drugs” provides new ideas and directions for the development of medicine.23,24 Among such repurposable drugs, pyrimethamine, pifithrin-mu, and mibefradil were selected for in vivo validation based on three criteria: (1) their predicted ability to reverse depression-related gene signatures (via CMAP, raw connectivity score < −0.6);25,26 (2) limited prior association with depression, unlike scopolamine (already in Phase II trials27) and estradiol (gender-dependent effects28); and (3) reported roles in inflammation or neuroprotection—pyrimethamine modulates T cell activity,29 pifithrin-mu inhibits microglial activation,30 and mibefradil regulates calcium signaling,31 all of which intersect with depression-related pathways.
Bioinformatics is the study of all aspects of acquisition, processing, storage, dissemination, analysis, and interpretation of biological information via computers as tools.32 The main research areas of bioinformatics are genomics, proteomics, and systems biology.33 Currently, bioinformatics techniques have been applied in many fields, such as gene identification analysis, genetic coding, and drug design.34–36 In recent years, studies have been conducted to identify new therapeutic drugs for diseases via bioinformatics, which has contributed greatly to research into related diseases.37–39 However, current strategies for repurposing antidepressant drugs often exhibit limited conversion efficiency, primarily due to the reliance on single-database screening approaches or the absence of in vivo validation. Numerous candidate compounds identified through bioinformatics analyses remain unverified in animal models, underscoring the urgent need to establish a systematic framework encompassing “screening – prediction – validation”. To address this gap, the present study investigates key targets and signaling pathways associated with depression using bioinformatics tools, aiming to identify potential therapeutic agents. Furthermore, the study integrates molecular docking techniques with animal experiments to validate the efficacy of these candidates, with the objective of offering novel insights into depression research and treatment. By combining multi-omics data (GEO, CMAP) with WGCNA and immune-related pathway analysis, complemented by rigorous in vivo experiments, this study addresses a critical research gap. This approach not only enhances the credibility of candidate compounds but also establishes a reproducible model for antidepressant drug discovery.
Materials and Methods
Data Resources
The GEO database was created in 2000 and is a gene expression database (https://www.ncbi.nlm.nih.gov/geo/).40 We obtained gene expression profile data (GSE182193) from the GEO database.41 GSE182193 contains 12 samples of peripheral venous blood samples from six depressed patients and six healthy volunteers derived from the GPL22120 platform of the GEO database.
PCA and OPLS-DA
PCA, a common multivariate statistical method, converts many related variables into uncorrelated ones via linear transformation to simplify sample analysis.42 OPLS-DA, a supervised discriminant method, uses partial least squares regression to predict sample categories.43 Based on differential gene expression matrices, we performed both for dimensionality reduction and identifying group differences.44
Screening of DEGs
GEO2R is an online analytics platform for the GEO database and is based on the Bioconductor project GEOquery and limma package for differential analysis of GEO high-throughput data. We performed the analysis via the GEO2R platform. After the analysis was completed, the gene symbols of the DEGs were obtained by removing replicates and setting the P value <0.05, |log2FC|≥1.
Protein‒Protein Interaction (PPI) and Pathway Enrichment Analysis
STRING 11.5 (medium confidence, hidden disconnected nodes) processed DEGs to get PPI networks.45 Nodes were uploaded to Metascape for KEGG/GO enrichment (P < 0.05), with PPI and protein-pathway networks built via Cytoscape 3.9.0.46,47
WGCNA
The GEO data were processed using the R package. The median absolute deviation (MAD) was calculated, and data with MAD values greater than 50% were selected for further analysis. A soft threshold of 5 was applied to construct the co-expression modules. The main clusters associated with the PPI network modules were subsequently visualized and analyzed using Cytoscape.48
Immune Correlation Analysis
The relative proportions of various immune cell types were estimated using the CIBERSORT algorithm. Key genes enriched through KEGG pathway analysis were selected, and the correlations between their expression levels and the proportions of immune cell infiltration were evaluated using the R package.49
Identification of Candidate Small Molecule Compounds
The DEGs obtained from the PPI network were uploaded to the second-generation CMAP platform (https://clue.io/). Prediction parameters were set to identify potential compounds. Candidate small molecules with potential antidepressant effects were selected based on prediction results and literature review.25,26
Molecular Docking
Molecular interactions were validated using AutoDock Vina. The small molecules identified by CMAP were used as ligands (with structures obtained from PubChem), while the key protein targets derived from KEGG pathway analysis were used as receptors (with structures obtained from UniProt).50–52
Calculation of Drug Similarity
RDKit is an open-source toolkit for cheminformatics based on 2D and 3D molecular manipulation of compounds via machine learning methods for compound descriptor generation, fingerprint generation, compound structure similarity calculation, and 2D and 3D molecular display.53 It is based on the Python language for fetching and use. We obtained several classes of current antidepressants through the PubChem platform, including monoamine oxidase inhibitors (iproniazid and phenelazine), tricyclic antidepressants (imipramine and amitriptyline), 5-hydroxytryptamine reuptake inhibitors (fluoxetine and sertraline), 5-hydroxytryptamine and norepinephrine reuptake inhibitors (venlafaxine and duloxetine), and noradrenergic and selective serotonergic antidepressants (mirtazapine). Furthermore, we used the RDKit package in Python to calculate the similarity between potential compounds obtained via CMAP and the abovementioned representative antidepressants obtained from PubChem and present them in the form of heatmaps.
In vivo Validation
Animals
Male Kunming (KM) mice aged 5--6 weeks (22–28 g) were purchased from SiPeiFu (Beijing) Biotechnology Co., Ltd. [license number: SCXK (Jing) 2019--0010)]. The mice were housed in standard cages containing wood shavings in a room with a carefully controlled ambient temperature (22 ± 1°C) and artificial lighting from 7:00 to 19:00 pm and were given standard laboratory chow and distilled water ad libitum. Test sample sizes (n=8 for behavioral tests, n=5 for biochemical assays) were determined based on power analysis conducted using G*Power 3.1, assuming a medium effect size (f = 0.25), α = 0.05, and a desired power of 0.8. This approach is consistent with established practices in preclinical depression research,54 where group sizes ranging from 5 to 8 mice are commonly adequate to detect significant behavioral and molecular alterations.
Drugs and Reagents
Fluoxetine HCl (item number: S27345-5g), Pyrimethamine (Item number: B27266), pifithrin-mu (item number: S88152) and mibefradil (item number: T23817) were procured from the Shanghai Yuanye Bioengineering Institute, situated in Shanghai, China. ELISA (enzyme-linked immunosorbent assay) kits for CDK2 (cyclin-dependent kinase 2), GHR (growth hormone receptor), GH (growth hormone), BDNF (brain-derived neurotrophic factor) and TrkB (tropomyosin receptor kinase B) were obtained from Wuhan Gene Beauty Biotechnology Co., Ltd. (Wuhan, China).
Stress Model Establishment and Drug Intervention
After one week of acclimation, the mice were randomly (Mice were randomly assigned to 6 groups (n = 8 per group) using a computer-generated random number table (https://www.random.org)) divided into the following six groups of eight mice each: (1) the control group, (2) the model group, (3) the fluoxetine group, (4) the pyrimethamine group, (5) the pifithrin-mu group, and (6) the mibefradil group. The mice in the control and model groups received intragastric injections of 10% DMSO (dimethyl sulfoxide). Small-molecule drugs were dissolved in 10% DMSO and then administered intragastrically (fluoxetine: 3.33 mg.kg−1/day; pyrimethamine: 4.17 mg.kg−1/day; pifithrin-mu: 8 mg.kg−1/day; mibefradil: 8.33 mg.kg−1/day).55–58 In addition, all mice except those in the control group were exposed to chronic restraint stress (CRS) on a daily basis for one week, with simultaneous administration of the drug intervention and restraint procedures. CRS consists of placing the mice in tubes from 8:30 am to 14:30 pm each day.59 The one week CRS protocol was selected based on prior research indicating that this duration reliably induces depressive-like behaviors in mice, such as decreased locomotor activity and increased immobility in behavioral assessments.59 This experimental model is extensively utilized in preclinical studies to simulate depression induced by chronic stress.
Behavioral Tests
Behavioral tests, including the open field test (OFT), the tail suspension test (TST), and the forced swim test (FST), were performed after 2 weeks of stress. The behavioral tests were performed every day according to the damage intensity of the behavioral tests, from weak to strong (7 d: OFT; 8 d: TST; 9 d: FST). The OFT was performed following the procedures described by Choleris et al.60 The TST was performed following the procedures described by Andreasen and Redrobe.61 Finally, the FST was carried out following the procedures described by Porsolt et al.62
Western Blot Analysis
After the behavioral study, the hippocampus of each mouse was homogenized in RlPA buffer containing PMSF and phosphatase inhibitors (time), and the supernatant was collected at 10000 RPM at 4°C for 10 min. The protein concentration was determined via a BCA protein detection kit (Beyotime). After electrophoresis on a 10% gel, the isolated proteins were transferred to a 0.45 µm PVDF membrane, which was blocked with 5% skim milk in TBST at room temperature for 1 hour and incubated with primary antibodies overnight. The primary antibodies used included rabbit polyclonal antibodies against GH1 (25KD; 1:500; Boster, BA14245), rabbit polyclonal antibodies against GHR (72–130KD; 1:500; Boster, BM5255), rabbit monoclonal antibodies against CDK2 (30–34KD; 1:2000; Boster, BM3926), rabbit monoclonal antibodies against BDNF (28KD; 1:2000, Boster, BM4113), rabbit monoclonal antibodies against TrkB/NTRK2 (92/140KD; 1:2000; Boster, M01388-3), and rabbit monoclonal antibodies against GAPDH (36KD; 1:2000; Boster, BM1623). The samples and molecular marker (10–180KD; Boster, AR1113) were incubated with a secondary antibody (1:5000, Boster) at room temperature for 2 hours. ECL chemiluminescence reagent (Boster, AR1196) was added to observe the imprinted protein, and the gray values of the protein bands were analyzed with ImageJ software (The original Western blot data is provided in the Supplementary Figure 1).
ELISA Test
After behavioral testing, the mouse hippocampus was collected and mixed with phosphate-buffered saline (pH 7.4). The tissue homogenizer was subsequently used (frequency 60 Hz, rotation rate 1800 times/min, duration 2 min) to homogenize the tissue, and the supernatant was collected at 4°C, 5000 RPM, 10 min, and stored at −80°C. Mouse hippocampus samples were tested via ELISA according to the kit manufacturer’s instructions. An enzyme label (EnSpire 2300, PerkinElmer, USA) was used. GH, GHR, CDK2, TrkB and BDNF were detected at 450 nm.
Statistical Analysis
All the data are expressed as the mean ± standard deviation of the mean (SD), and one-way ANOVA was performed via a fully randomized trial design. For multiple comparisons across groups (eg, behavioral tests and Western blot), Bonferroni correction was applied to adjust p-values, reducing the risk of false positives. Adjusted P < 0.05 was considered statistically significant. SPSS 22.0 software was used for one-way ANOVA to determine differences between the means and for Person correlation analysis. P < 0.05 was considered to indicate statistical significance. The effect size was quantified using Cohen’s d. The criteria for interpreting Cohen’s d are as follows: d ≈ 0.2 indicates a small effect, d ≈ 0.5 indicates a medium effect, and d > 0.8 indicates a large effect.63 The data were visualized via Origin 2022 software.
Results
GEO2R Analysis of Differentially Expressed Genes in Patients with Depression
Through GEO2R analysis and a series of processing steps, we ultimately obtained 311 DEGs, as shown in Figure 1A. Among these genes, 125 were upregulated, and the remaining 186 were downregulated. We collated the expression data of the DEGs and performed PCA and OPLS-DA; the results are shown in Figure 1B and C. There were two obvious groups, which indicates that these differentially expressed genes are well represented.
 |
Figure 1 GEO2R analysis of differentially expressed genes in patients with depression. Notes: (A) Volcano plot analysis of DEGs; (B) Results of PCA of DEGs; (C) Results of OPLS-DA of DEGs. In Figure 1A red dots represent genes upregulated in depressed patients compared with normal controls, and blue dots represent downregulated genes. P<0.05, |log2FC|≥1. |
PPI Network and Enrichment Analysis
PPI networks identified 92 nodes, which were enriched in the PI3K-Akt, MAPK, and neurotrophin signaling pathways (Figure 2A–C). The results of the enrichment analysis are shown in Figure 2A and B. The results of the KEGG pathway enrichment analysis revealed several signaling pathways closely related to depression: the PI3K-Akt signaling pathway,64 the MAPK signaling pathway,65 and the neurotrophin signaling pathway.66 GO results revealed an association with circadian rhythms67 and neurotrophins.66 There were 122 nodes in this network, 30 pathways and 92 proteins, and 382 edges: 235 for protein–protein and 147 for protein-pathway (Figure 2C).
 |
Figure 2 Results of KEGG pathway enrichment analysis, results of GO enrichment analysis, and protein‒protein and protein‒pathway network diagrams. Abbreviations: BP, biological process; CC, cellular component; MF, molecular function. Notes: (A) Results of KEGG pathway enrichment analysis; (B) results of GO enrichment analysis; (C) protein‒protein and protein‒pathway network diagrams, with circular nodes representing pathways and V-shaped nodes representing proteins. The colors from light to dark represent the degree of change from small to large. |
WGCNA Results and Cluster Construction
Using WGCNA, we obtained a total of 20 coexpressed gene modules; the results are shown in Figure 3A and B. Nodes in the PPI network were distributed mainly in the turquoise and blue modules of the WGCNA results. Therefore, we constructed these groups, as shown in Figure 3C. This finding suggests, to some extent, that there may be synergistic effects between different genes in the same module during the development and progression of depression.
 |
Figure 3 Analysis of network topology for soft powers, gene dendrogram and module colors, clusters of genes coexpressed in DEGs. Notes: (A) Analysis of network topology for soft powers; (B) gene dendrogram and module colors; (C) clusters of genes coexpressed in DEGs. Nodes with the same color indicate genes of the same coexpression module. |
Immune Correlation Analysis
According to the relevant literature, the immune system in depressed patients is abnormal.49 The expression of 12 key genes in the PI3K‒Akt signaling pathway, MAPK signaling pathway, and neurotrophin signaling pathway was correlated with the proportion of immune cells, as shown in Figure 4A and B. The results revealed good correlations among them. These findings also suggest that these genes affect the immune system of the organism.
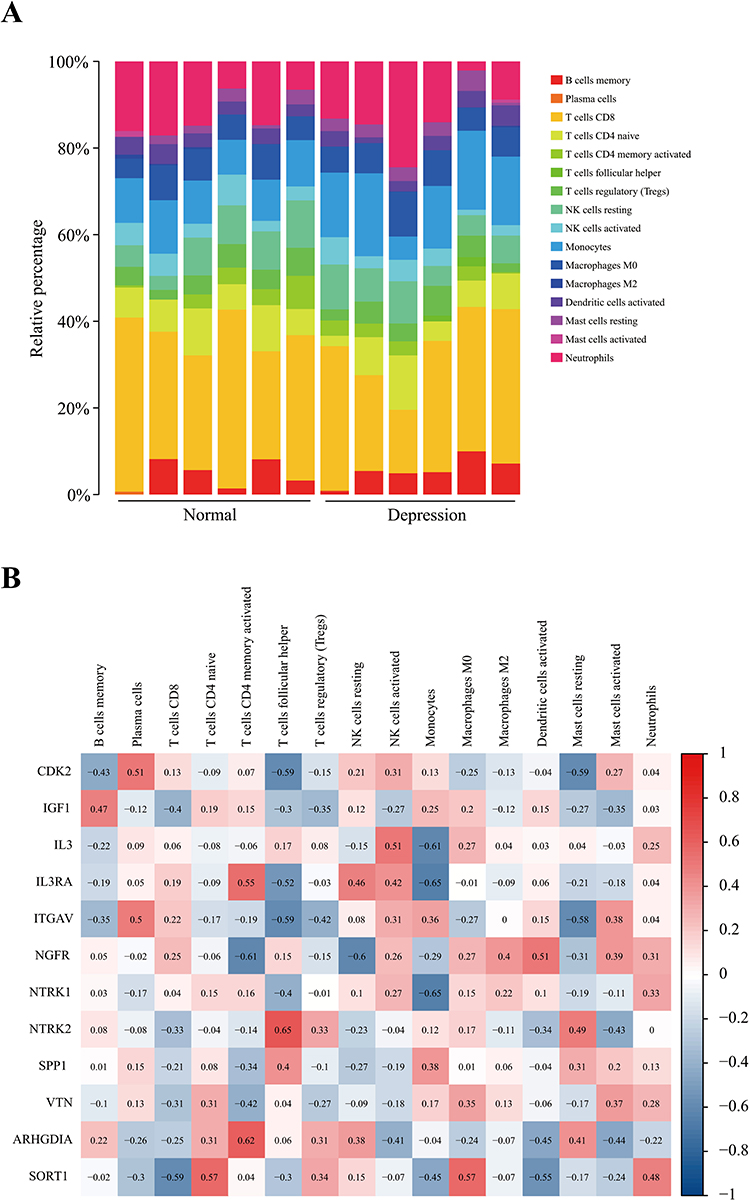 |
Figure 4 Proportion of immune cells in each individual and correlation between key gene expression in important pathways and proportions of different immune cells. Notes: (A) Proportion of immune cells in each individual; (B) correlation between the expression of key genes in important pathways and the proportions of different immune cells. |
Screening of Potential Drugs
The “raw connectivity score” is an important parameter in the results provided via the CMAP database analysis platform. A negative value indicates that the expression profile of the drug perturbation is negatively correlated with the expression profile of the disease perturbation, tentatively suggesting the drug can alleviate or even reverse the disease state. By searching the literature, five drugs were screened as potential small-molecule compounds for the treatment of depression in the next step of the analysis, as shown in Table 1.
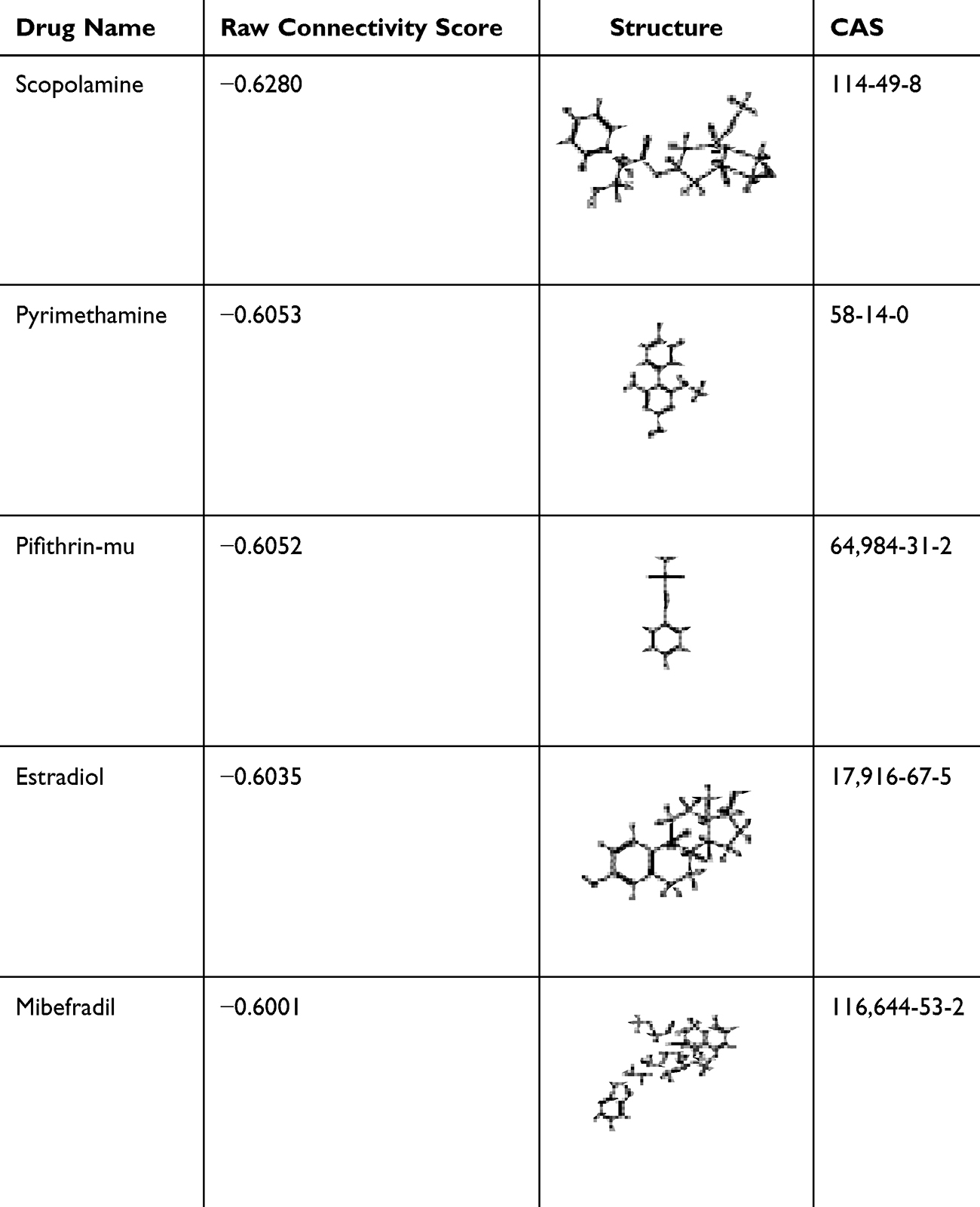 |
Table 1 Specific Information on the Five Small-Molecule Compounds in the CMAP Screening Results |
Molecular Docking
We used the compounds listed in Table 1 (scopolamine, pyrimethamine, pifithrin-mu, estradiol, and mibefradil) as ligands and fluoxetine as a control. Twelve targets enriched in important pathways (PI3K-Akt signaling pathway, MAPK signaling pathway, and neurotrophin signaling pathway) were used as receptors (CDK2, IGF1, IL3, IL3RA, ITGAV, NGFR NTRK1, NTRK2, SPP1, VTN, ARHGDIA, and SORT1). The results revealed that these compounds exhibited good binding (Table 2). For molecular docking, a binding energy ≤ −7 kcal/mol is generally considered a ‘strong’ interaction, indicating stable binding between the compound and target protein. Most of the tested compounds (eg, estradiol with CDK2: −8.8 kcal/mol, pyrimethamine with IL3RA: −8.2 kcal/mol) met this criterion, suggesting high affinity. We plotted the binding energy of the molecular docking results as a heatmap (Figure 5A) and visualized the lowest binding energy results with PyMOL. Figure 5B shows the results of the molecular docking of estradiol with CDK2, and Figure 5C shows the results of the molecular docking of estradiol with NTRK2.
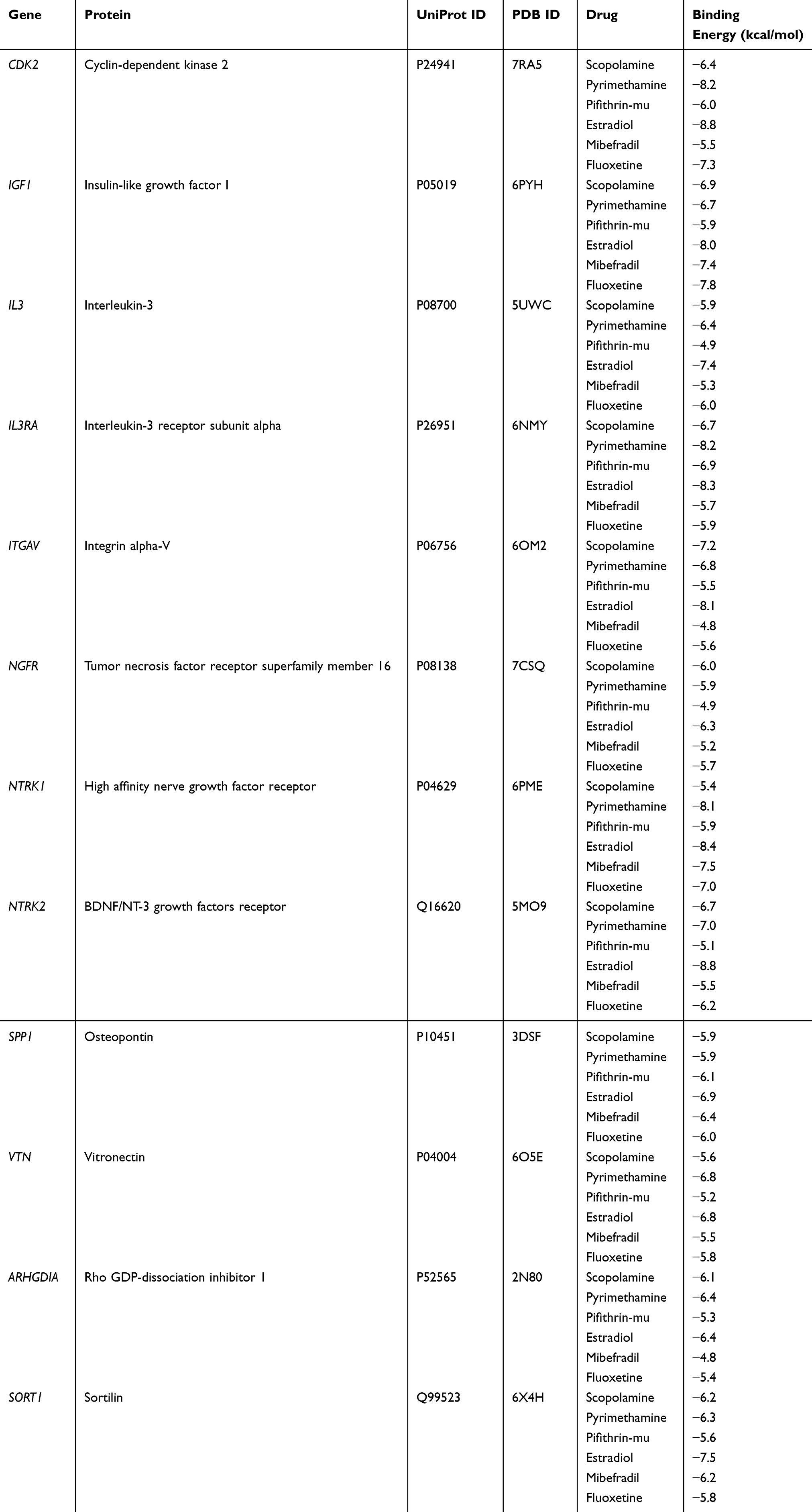 |
Table 2 Molecular Docking Results |
 |
Figure 5 Results of the molecular docking and drug similarity calculations. Notes: (A) Binding energy heatmap of molecular docking; (B) visualization of the molecular docking of estradiol with CDK2; (C) visualization of the molecular docking of estradiol with NTRK2. (D) Heatmap of similarity between predicted potential antidepressants and representative antidepressant drugs. |
Drug Similarity Calculation Results
We calculated the similarity of the drugs via the RDKit package, which is based on the method described previously; the results are shown in Figure 5D. The predicted antidepressant drugs presented some structural similarity with various types of current antidepressant representatives, which supports the results obtained from our predictions.
In vivo Validation Assays
Among the five predicted compounds, scopolamine and estradiol were excluded from in vivo testing. Scopolamine has already advanced to phase II clinical trials for depression,27 and estradiol’s antidepressant effects are highly dependent on gender,28 which limits its applicability in our male mouse model. Pyrimethamine, pifithrin-mu, and mibefradil were selected for in vivo validation due to their limited prior association with depression, thereby emphasizing the novelty of our findings. Following the bioinformatics analysis above, the validation test adopted the CRS model for stress binding, simulated a depressive environment to induce depression-like behavior in mice and administered small molecule drugs (pyrimethamine, pifithrin-mu and mibefradil) for intervention (Figure 6A).
 |
Figure 6 The impact of stress and small-molecule drug intervention on mouse behavior. Notes: (A) Behavioral tests; (B) OFT (open field test); (C) TST (tail suspension test); (D) FST (forced swim test). The data are shown as the means ± SDs (n = 8). *P < 0.05, **P < 0.01 and ***P < 0.001 denote significant differences from the control group. #P < 0.05, ##P < 0.01 and ###P < 0.001 denote significant differences from the Model group. |
Behavioral Test Results
The effects of small-molecule drugs on depressive behavior and recovery were assessed through the open field test (OFT), the forced swim test (FST) and the tail suspension test (TST). The results of the aforementioned tests are presented in Figure 6B and D. Compared with the control group, the model group exhibited significant changes in behavioral parameters: immobile time increased by 29.62% (P < 0.05; d = 0.69); struggling time increased markedly in both TST (72.76%, P < 0.001; d = 1.92) and FST (65.86%, P < 0.001, d = 1.78); conversely, movement distance decreased by 23.38% (P < 0.05; d = 0.68), center movement distance decreased by 94.21% (P < 0.001; d = 2.31), and center residence time decreased by 89.16% (P < 0.001; d = 2.15). Following the administration of the small-molecule drugs, the movement distance (fluoxetine: 24.67%, P < 0.05, d = 0.82; pyrimethamine: 33.44%, P < 0.01, d = 1.05; pifithrin-mu: 34.62%, P < 0.01, d = 1.08; mibefradil: 60.32%, P < 0.001, d = 1.89), center residence time (fluoxetine: 399.44%, P < 0.05, d = 0.76; pyrimethamine: 381.02%, P < 0.01, d = 0.75; pifithrin-mu: 446.43%, P < 0.01, d = 0.85; mibefradil: 409.02%, P < 0.001, d = 0.78) and center movement distance (fluoxetine: 594.86%, P < 0.01, d = 1.20; pyrimethamine: 633.59%, P < 0.01, d = 1.25; pifithrin-mu: 804.52%, P < 0.01, d = 1.50; mibefradil: 843.97%, P < 0.01, d = 1.55) of the mice in each group were found to be significantly greater than those of the Model group. Conversely, the immobile time (fluoxetine: −17.04%, P < 0.05, d = 0.60; pyrimethamine: −20.25%, P < 0.01, d = 0.70; pifithrin-mu: −23.24%, P < 0.01, d = 0.80; mibefradil: −30.19%, P < 0.001, d = 0.95), TST-struggling time (fluoxetine: −25.00%, P < 0.05, d = 0.70; pyrimethamine: −26.54%, P < 0.05, d = 0.75; pifithrin-mu: −21.03%, P < 0.05, d = 0.65; mibefradil: −20.21%, P < 0.05, d = 0.60) and FST-struggling time (fluoxetine: −19.60%, P < 0.05, d = 0.60; pyrimethamine: −15.88%, P < 0.05, d = 0.55; pifithrin-mu: −25.23%, P < 0.01, d = 0.80; mibefradil: −17.16%, P < 0.05, d = 0.58) significantly decreased.
WB Verification Results
In accordance with the aforementioned relationship between small-molecule drugs and pathways, target proteins in the PI3K‒Akt signaling pathway and neurotrophic factor signaling pathway were selected for WB verification (Figure 7A). The results, as illustrated in Figure 7B and C, demonstrated that the expression levels of GH (P < 0.001, d = 2.10), GHR (P < 0.01, d = 1.65) and CDK2 (P < 0.001, d = 1.95) in the Model group were markedly elevated in comparison with those in the Control group, whereas the expression levels of TrkB (P < 0.001, d = 2.20) and BDNF (P < 0.001, d = 2.05) were significantly reduced. Following the administration of a small molecule drug, the expression levels of GH (fluoxetine: P < 0.01, d = 1.40; pyrimethamine: P < 0.001, d = 1.85; pifithrin-mu: P < 0.001, d = 1.70; mibefradil: P < 0.05, d = 0.75), GHR (fluoxetine: P < 0.01, d = 1.30; pifithrin-mu: P < 0.05, d = 0.65) and CDK2 (fluoxetine: P < 0.001, d = 1.60; pifithrin-mu: P < 0.05, d = 0.70; mibefradil: P < 0.05, d = 0.60) were significantly lower than those in the control group. Conversely, the expression levels of TrkB (fluoxetine: P < 0.001, d = 2.15; pyrimethamine: P < 0.001, d = 2.00; pifithrin-mu: P < 0.001, d = 1.90; mibefradil: P < 0.05, d = 0.80) and BDNF (fluoxetine: P < 0.001, d = 1.80; pyrimethamine: P < 0.01, d = 1.25; mibefradil: P < 0.05, d = 0.70) were significantly elevated.
 |
Figure 7 Verification results of the WB and ELISA test. Notes: (A) Test procedure; (B) WB strip map; (C) WB gray value; (D) ELISA test results. The data are shown as the means ± SDs (n = 5). *P < 0.05, **P < 0.01 and ***P < 0.001 denote significant differences from the control group. #P < 0.05, ##P < 0.01 and ###P < 0.001 denote significant differences from the Model group. |
ELISA Test Results
The ELISA results are presented in Figure 7D. Compared with those in the control group, the expression levels of GH (P < 0.01, d = 1.45), GHR (P < 0.01, d = 1.35) and CDK2 (P < 0.001, d = 1.80) in the model group were markedly elevated, whereas the expression levels of TrkB (P < 0.05, d = 0.75) and BDNF (P < 0.05, d = 0.70) were significantly reduced. Following the administration of the small molecule drug, there was a notable reduction in the expression levels of GH (fluoxetine: P < 0.01, d = 1.20; pyrimethamine: P < 0.01, d = 1.25; pifithrin-mu: P < 0.001, d = 1.50; mibefradil: P < 0.01, d = 1.20), GHR (fluoxetine: P < 0.001, d = 1.60; pyrimethamine: P < 0.01, d = 1.30; pifithrin-mu: P < 0.001, d = 1.55; mibefradil: P < 0.01, d = 1.30), CDK2 (fluoxetine: P < 0.001, d = 1.70; pyrimethamine: P < 0.001, d = 1.65; pifithrin-mu: P < 0.001, d = 1.75; mibefradil: P < 0.001, d = 1.60), while the expression levels of TrkB (fluoxetine: P < 0.001, d = 1.60; pyrimethamine: P < 0.01, d = 1.30; pifithrin-mu: P < 0.001, d = 1.55; mibefradil: P < 0.001, d = 1.50), and BDNF (fluoxetine: P < 0.01, d = 1.10; mibefradil: P < 0.01, d = 1.05) markedly increased in comparison with the levels observed in the Model group.
Correlation Analysis
A Pearson association analysis was conducted to examine the relationships between target proteins in the PI3K‒Akt signaling pathway and the neurotrophin signaling pathway and behavioral traits. As shown in Figure 8, CDK2, GHR, and GH were markedly positively correlated with immobility time (P < 0.05; P < 0.05; P < 0.05), TST-struggle time (P < 0.05; P < 0.05; P < 0.05), and FST-struggle time (P < 0.05; P < 0.05; P < 0.05) and negatively correlated with movement distance (P < 0.05; P < 0.05; P < 0.05), center movement distance (P < 0.05; P < 0.05; P < 0.05) and center residence time (P < 0.05; P < 0.05; P < 0.05). Conversely, the data indicate a positive correlation between BDNF and center motion distance (P < 0.05) and center residence time (P < 0.05), and a significant negative correlation was observed with FST-struggle time (P < 0.05). In addition, a significant positive correlation was observed between TrkB and movement distance (P < 0.05). Note that the correlation analysis was conducted using a relatively small sample size (n = 5), which may affect statistical power. Further studies involving larger cohorts are warranted to confirm these associations.
 |
Figure 8 Pearson correlation analysis (n = 5). |
Discussion
Depression, a prevalent mental disorder with a rising incidence, poses substantial threats to public health, including life-threatening outcomes.68 Two critical challenges persist in depression research: elucidating its complex pathogenesis and developing efficient therapeutic agents. To address these, our study integrated bioinformatics tools—encompassing differential gene analysis, PPI network construction, enrichment analysis, WGCNA, and immune correlation analysis—with molecular docking and in vivo experiments, aiming to systematically identify potential antidepressant targets and compounds.
Through GEO2R analysis of the GSE182193 dataset, we identified 311 DEGs. Subsequent PCA and OPLS-DA clearly distinguished the normal and depressed groups, validating the robustness of these DEGs as reliable biomarkers. Uploading these DEGs to the STRING database yielded a PPI network with 92 nodes, which were further analyzed via WGCNA to identify 20 coexpression modules. Notably, PPI nodes were concentrated in two main modules, suggesting that coexpressed genes within these modules may exert synergistic effects in driving the onset and progression of depression. KEGG pathway enrichment analysis highlighted the PI3K-Akt, MAPK, and neurotrophin signaling pathways as core nodes associated with depression16,69–72—a finding consistent with broader frameworks of depression-related molecular mechanisms but uniquely substantiated herein through multi-layered validation, combining human transcriptome data with murine experimental evidence.
Given the well-documented immune dysregulation in depressed patients,49 we further explored the interplay between key pathways and immunity. Consistent with prior reports linking the PI3K-Akt,73 MAPK,74 and neurotrophin66 pathways to immune and inflammatory processes, our immune correlation analysis revealed significant associations between the expression of key genes in these pathways and the proportions of various immune cells (eg, CDK2 with T cell subsets and macrophages). These associations are reinforced by evidence that pathway-mediated mechanisms—such as TREM2 activation via PI3K/Akt,75 baicalein’s regulation of PI3K/AKT/FoxO1,64 Astragalus polysaccharides’ modulation of NF-κB/MAPK,76 and Magnolol’s activation of MAPK77—link neuroinflammation to depression. Collectively, these findings provide novel insights into the crosstalk between neuroinflammation and depression, extending previous research by demonstrating pathway-specific immune modulation and reinforcing the role of neurotrophin synthesis/secretion mechanisms in inflammatory processes.78 This establishes a mechanistic basis for targeting these genes to concurrently regulate neuronal dysfunction and immune imbalance in depression.
To translate these pathway insights into potential therapeutics, we uploaded DEGs from the PPI network to the CMAP II CLUE platform, identifying five candidate small-molecule compounds (scopolamine, pyrimethamine, pifithrin-mu, estradiol, and mibefradil) with a “raw connectivity score” < −0.6. Existing literature supports their relevance to depression: scopolamine exhibits rapid, long-lasting antidepressant effects;27 pyrimethamine reverses depressive symptoms in chronic toxoplasmosis,29 aligning with links between toxoplasmosis and depression;79 pifithrin-mu reduces proinflammatory factors and reverses behavioral deficits in LPS-treated mice;30 estradiol alleviates anxiety and depressive behavior in aged female mice,80 consistent with its reported correlation with depression;28,81 and mibefradil mimics antidepressant effects.31 Among these, pyrimethamine, pifithrin-mu, and mibefradil were selected for in vivo validation, whereas scopolamine and estradiol were excluded: scopolamine is already in Phase II clinical trials for depression,27 and estradiol’s antidepressant effects are highly gender-dependent,28 limiting its applicability in our male mouse model. Importantly, these three compounds had limited prior association with depression, underscoring the novelty of our validation. In CRS model mice, they exhibited significant antidepressant efficacy comparable to fluoxetine. Focusing on proteins involved in the neurotrophin and PI3K-Akt signaling pathways82,83—which are critical to depression—WB and ELISA results demonstrated that these compounds restore stress-induced metabolic abnormalities by increasing BDNF and TrkB expression and decreasing GH and CDK2 levels. Molecular docking further confirmed their strong binding to key proteins in these pathways (eg, estradiol with CDK2 and NTRK2, binding energies < −8 kcal/mol).
Pearson correlation analysis clarified the mechanistic relevance: overexpression of CDK2, GHR, and GH correlated positively with depressive-like behaviors (eg, immobility time in the forced swim test), while BDNF and TrkB upregulation associated with behavioral improvement. These findings align with reports that excessive GH expression links to depressive symptoms84 and support CDK2 and GH as potential therapeutic targets. Notably, our data suggest CDK2 inhibition may protect hippocampal neurons by preventing excessive neurogenic depletion—consistent with evidence that CDK inhibitors regulate neural progenitor proliferation in neurogenic niches85 and counteract antidepressant-induced neurogenetic depletion,86 a mechanism not previously reported in depression research. This represents the first validation of their antidepressant efficacy through a systematic “screening-prediction-validation” framework, distinguishing it from studies relying solely on single-database screening or in silico predictions, and confirms that regulating abnormal GH and CDK2 expression in the PI3K-Akt pathway is key to their therapeutic effects.
Study Limitations
(1) The small sample size in animal experiments (n=8 per group) may limit the statistical power of the results; (2) the exclusive use of male mice restricts the generalizability of findings to female populations; (3) the short treatment duration (2 weeks) may not reflect long-term therapeutic efficacy; and (4) the lack of human in vivo data necessitates cautious interpretation of the clinical relevance of these findings. (5) The molecular docking results showed good binding properties, suggesting potential interactions. However, these in silico findings were not validated by experimental methods such as surface plasmon resonance (SPR) or thermal shift assay, which would confirm the binding affinity in vitro. Future studies should include these experiments to strengthen the mechanistic evidence.
Future Directions
(1) Expanding the experimental model to include female and aged mice to investigate sex- and age-specific effects; (2) conducting long-term toxicity evaluations and dose-response curve analyses to optimize therapeutic windows; (3) exploring the synergistic effects of combined administration of pyrimethamine, pifithrin-mu, and mibefradil; and (4) validating these findings in human clinical samples to enhance translational relevance (For example, tests are performed using human neural models or patient-derived induced pluripotent stem cell-based systems to improve clinical relevance).
In summary, this study identifies pyrimethamine, pifithrin-mu, and mibefradil as promising antidepressant candidates that act by regulating key proteins in the PI3K-Akt and neurotrophin signaling pathways. It also validates a robust framework for drug repurposing and identifies novel targets, advancing both depression research and therapeutic development.
Conclusion
In this study, we employed bioinformatics techniques to investigate key signaling pathways and potential antidepressant small-molecule compounds, validating our findings through molecular docking, similarity calculations, and in vivo experiments. Our results indicate that scopolamine, pyrimethamine, pifithrin-mu, estradiol, and mibefradil may exert antidepressant effects by modulating the PI3K-Akt, MAPK, and neurotrophin signaling pathways. Animal experiments confirmed that pyrimethamine, pifithrin-mu, and mibefradil can regulate the expression of key proteins in the PI3K-Akt and neurotrophin signaling pathways, thereby alleviating depressive behaviors in mice. These data support the potential of these three compounds as antidepressant candidates, though further preclinical evaluations—including toxicity tests, dose-response curve analyses, and validation in female mouse models—are required to confirm their safety and efficacy. Subsequent steps should prioritize these preclinical assessments to lay the groundwork for eventual translational research, including human clinical trials, to evaluate their therapeutic potential in clinical settings.
Abbreviations
PCA, Principal component analysis; WB, Western blot; ELISA, Enzyme-linked immunosorbent assay; CRS, Chronic restraint stress; HPA, Hypothalamic pituitary adrenal; PPI, Protein–protein interaction; PMSF, Phenylmethanesulfonyl fluoride; TBST, Tris-Borate-Sodium Tween-20; ECL, Electrochemiluminescence; GEO, GENE EXPRESSION OMNIBUS; OPLS-DA, Orthogonal partial least-squares discrimination analysis; KM, Male Kunming; ARRIVE, Animal Research: Reporting of In vivo Experiments; CDK2, Cyclin-dependent kinase 2; GHR, Growth hormone receptor; GH, Growth hormone; BDNF, Brain-derived neurotrophic factor; TrkB, Tropomyosin receptor kinase B; DMSO, Dimethyl sulfoxide; OFT, Open field test; TST, Tail suspension test; FST, Forced swim test; SD, Mean ± standard deviation of the mean; MDD, Major Depressive Disorder; LPS, Lipopolysaccharide; KEGG, Kyoto Encyclopedia of Genes and Genomes; GO, Gene Ontology; WGCNA, Weighted gene coexpression network analysis; CMAP, Connectivity map.
Data Sharing Statement
The original data that support the conclusions of this article will be made available by the corresponding author upon reasonable request, without any reservation.
Ethics Statement
All studies involving human data, including the GEO database data (GSE182193) used in this research, have been reviewed and approved by the Ethics Review Committee of the Northwest Institute of Plateau Biology (Approval No. NWIPB20171106-01) and adhere to the Helsinki Declaration and relevant ethical standards; all animal experiments, in compliance with the ARRIVE guidelines and the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 8023, revised 1978), were approved by the Animal Experiment Committee of the Northwest Plateau Institute of Biology, CAS with the same approval number.
Acknowledgments
We thank AJE (https://www.aje.cn/) for its linguistic assistance during the preparation of this manuscript.
Funding
This work was supported by the Natural Science Foundation of China (Grant No. 82171863), the Innovation Platform Program of Qinghai Province (2021-ZJ-T02), and the Tianfu Emei Project of Sichuan Province.
Disclosure
The authors have no competing interests.
References
1. Touloumis C. Treatment resistant depression: challenges and therapeutic choices. Psychiatriki. 2021;32(Supplement 1):15–31. doi:10.22365/jpsych.2021.047
2. Wang Y-Q, Li R, Zhang M-Q, Zhang Z, Qu W-M, Huang Z-L. The Neurobiological Mechanisms and Treatments of REM Sleep Disturbances in Depression. Curr Neuropharmacol. 2015;13(4):543–553. doi:10.2174/1570159x13666150310002540
3. Roca M, Del Amo AR-L, Riera-Serra P, et al. Suicidal risk and executive functions in major depressive disorder: a study protocol. BMC Psychiatry. 2019;19(1):253. doi:10.1186/s12888-019-2233-1
4. Hennings JM, Ising M, Uhr M, Holsboer F, Lucae S. Effects of weariness of life, suicide ideations and suicide attempt on HPA axis regulation in depression. Psychoneuroendocrinology. 2021;131:105286. doi:10.1016/j.psyneuen.2021.105286
5. Wisłowska-Stanek A, Kołosowska K, Maciejak P. Neurobiological Basis of Increased Risk for Suicidal Behaviour. Cells. 2021;10(10):2519. doi:10.3390/cells10102519
6. Qi T, Hu T, Ge -Q-Q, et al. COVID-19 pandemic related long-term chronic stress on the prevalence of depression and anxiety in the general population. BMC Psychiatry. 2021;21(1):380. doi:10.1186/s12888-021-03385-x
7. Santomauro DF, Mantilla Herrera AM, Shadid J. COVID-19 Mental Disorders Collaborators. Global prevalence and burden of depressive and anxiety disorders in 204 countries and territories in 2020 due to the COVID-19 pandemic. Lancet. 2021;398(10312):1700–1712. doi:10.1016/S0140-6736(21)02143-7
8. Yirmiya R, Rimmerman N, Reshef R. Depression as a microglial disease. Trends Neurosci. 2015;38(10):637–658. doi:10.1016/j.tins.2015.08.001
9. Hirschfeld RM. History and evolution of the monoamine hypothesis of depression. J Clin Psychiatry. 2000;61(6):4–6. PMID: 10775017.
10. Tobe EH. Mitochondrial dysfunction, oxidative stress, and major depressive disorder. Neuropsychiatr Dis Treat. 2013;9:567–573. doi:10.2147/NDT.S44282
11. Schiepers OJ, Wichers MC, Maes M. Cytokines and major depression. Prog Neuropsychopharmacol Biol Psychiatry. 2005;29(2):201–217. doi:10.1016/j.pnpbp.2004.11.003
12. Castrén E, Antila H. Neuronal plasticity and neurotrophic factors in drug responses. Mol Psychiatry. 2017;22(8):1085–1095. doi:10.1038/mp.2017.61
13. Groves JO. Is it time to reassess the BDNF hypothesis of depression? Mol Psychiatry. 2007;12(12):1079–1088. doi:10.1038/sj.mp.4002075
14. Góralczyk-Bińkowska A, Szmajda-Krygier D, Kozłowska E. The Microbiota-Gut-Brain Axis in Psychiatric Disorders. Int J Mol Sci. 2022;23(19):11245. doi:10.3390/ijms231911245
15. Mayer EA, Nance K, Chen S. The Gut-Brain Axis. Annu Rev Med. 2022;73:439–453. doi:10.1146/annurev-med-042320-014032
16. Bódizs R, Purebl G, Rihmer Z. Hangulat, hangulatingadozások és depresszió: a cirkadián ritmusok szerepe [Mood, mood fluctuations and depression: role of the circadian rhythms]. Neuropsychopharmacol Hung. 2010;12(1):277–287. Hungarian. PMID: 20305303.
17. Remes O, Mendes JF, Biological TP. Psychological, and Social Determinants of Depression: a Review of Recent Literature. Brain Sci. 2021;11(12):1633. doi:10.3390/brainsci11121633
18. Andrade C. Relative Efficacy and Acceptability of Antidepressant Drugs in Adults With Major Depressive Disorder: commentary on a Network Meta-Analysis. J Clin Psychiatry. 2018;79(2):18f12254. doi:10.4088/JCP.18f12254
19. Yuan Z, Chen Z, Xue M, Zhang J, Leng L. Application of antidepressants in depression: a systematic review and meta-analysis. J Clin Neurosci. 2020;80:169–181. doi:10.1016/j.jocn.2020.08.013
20. Hong R, Luo L, Hu ZL, et al. Qi SH. Lepidium meyenii Walp (Maca)-derived extracellular vesicles ameliorate depression by promoting 5-HT synthesis via the modulation of gut-brain axis. Imeta. 2023;2(3):e116. doi:10.1002/imt2.116
21. Yousefnia S. Mechanistic effects of arsenic trioxide on acute promyelocytic leukemia and other types of leukemias. Cell Biol Int. 2021;45(6):1148–1157. doi:10.1002/cbin.11563
22. Koch HP. Thalidomide and congeners as anti-inflammatory agents. Prog Med Chem. 1985;22:165–242. doi:10.1016/s0079-6468(08)70231-5
23. Pantziarka P, Pirmohamed M, Mirza N. New uses for old drugs. BMJ. 2018;361:k2701. doi:10.1136/bmj.k2701
24. Sachs RE, Ginsburg PB, Goldman DP. Encouraging New Uses for Old Drugs. JAMA. 2017;318(24):2421–2422. doi:10.1001/jama.2017.17535
25. Lamb J, Crawford ED, Modell JW, et al. The Connectivity Map: using gene-expression signatures to connect small molecules, genes, and disease. Science. 2006;313(5795):1929–1935. doi:10.1126/science.1132939
26. Subramanian A, Narayan R, Corsello SM, et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell. 2017;171(6):1437–1452.e17. doi:10.1016/j.cell.2017.10.049. PMID: 29195078; PMCID: PMC5990023.
27. Hasselmann H. Scopolamine and depression: a role for muscarinic antagonism? CNS Neurol Disord Drug Targets. 2014;13(4):673–683.
28. Balzer BW, Duke SA, Hawke CI, Steinbeck KS. The effects of estradiol on mood and behavior in human female adolescents: a systematic review. Eur J Pediatr. 2015;174(3):289–298. doi:10.1007/s00431-014-2475-3
29. Castaño BL, Silva AA, Hernandez-Velasco LL, et al. Sulfadiazine Plus Pyrimethamine Therapy Reversed Multiple Behavioral and Neurocognitive Changes in Long-Term Chronic Toxoplasmosis by Reducing Brain Cyst Load and Inflammation-Related Alterations. Front Immunol. 2022;13:822567. doi:10.3389/fimmu.2022.822567
30. Zhang R, Wang J, Hu Y, et al. Pifithrin-μ Attenuates Acute Sickness Response to Lipopolysaccharide in C57BL/6J Mice. Pharmacology. 2016;97(5–6):245–250. doi:10.1159/000444187
31. Boselli C, Barbone MS, Lucchelli A. Older versus newer antidepressants: substance P or calcium antagonism? Can J Physiol Pharmacol. 2007;85(10):1004–1011. doi:10.1139/y07-089
32. Tao Z, Shi A, Li R, Zhao J. Microarray bioinformatics in cancer- a review. J BUON. 2017;22(4):838–843. PMID: 29155508.
33. Brewis IA, Brennan P. Proteomics technologies for the global identification and quantification of proteins. Adv Protein Chem Struct Biol. 2010;80:1–44. doi:10.1016/B978-0-12-381264-3.00001-1
34. Zhou Q, Hong L, Wang J. Identification of key genes and pathways in pelvic organ prolapse based on gene expression profiling by bioinformatics analysis. Arch Gynecol Obstet. 2018;297(5):1323–1332. doi:10.1007/s00404-018-4745-1
35. Ling J, O’Donoghue P, Söll D. Genetic code flexibility in microorganisms: novel mechanisms and impact on physiology. Nat Rev Microbiol. 2015;13(11):707–721. doi:10.1038/nrmicro3568
36. Yang Y, Wang J. Rational drug design with the aid of molecular modeling and bioinformatics. Curr Pharm Des. 2013;19(12):2245. doi:10.2174/1381612811319120011
37. Liu Z, Zhang R, Chen X, et al. Identification of hub genes and small-molecule compounds related to intracerebral hemorrhage with bioinformatics analysis. PeerJ. 2019;7:e7782. doi:10.7717/peerj.7782
38. Geng L, Zhang X, Tang Y, Gu W. Identification of Potential Key Biomarkers and Immune Infiltration in Oral Lichen Planus. Dis Markers. 2022;2022:7386895. doi:10.1155/2022/7386895
39. Luo D, Liang XZ, Xu B, Liu JB, Wei CF, Li G. Rapid Discovery of Potential Drugs for Osteonecrosis of Femoral Head Based on Gene Expression Omnibus Database and Connectivity Map. Orthop Surg. 2019;11(6):1209–1219. doi:10.1111/os.12533
40. Barrett T, Wilhite SE, Ledoux P, et al. NCBI GEO: archive for functional genomics data sets--update. Nucleic Acids Res. 2013;41(Database issue):D991–5. doi:10.1093/nar/gks1193
41. Zhou T, Li M, Xiao Z, et al. Chronic Stress-Induced Gene Changes In Vitro and In vivo: potential Biomarkers Associated With Depression and Cancer Based on circRNA- and lncRNA-Associated ceRNA Networks. Front Oncol. 2021;11:744251. doi:10.3389/fonc.2021.744251
42. Ben salem K, Ben Abdelaziz A. Principal Component Analysis (PCA). Tunis Med. 2021;99(4):383–389. English. PMID: 35244921.
43. Boccard J, Rutledge DN. A consensus orthogonal partial least squares discriminant analysis (OPLS-DA) strategy for multiblock Omics data fusion. Anal Chim Acta. 2013;769:30–39. doi:10.1016/j.aca.2013.01.022
44. Vitale R, Marini F, Ruckebusch C. SIMCA Modeling for Overlapping Classes: fixed or Optimized Decision Threshold? Anal Chem. 2018;90(18):10738–10747. doi:10.1021/acs.analchem.8b01270
45. Szklarczyk D, Gable AL, Nastou KC, et al. The STRING database in 2021: customizable protein‒protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021;49(D1):D605–D612. doi:10.1093/nar/gkaa1074. Erratum in: Nucleic Acids Res. 2021;49(18):10800. doi: 10.1093/nar/gkab835.
46. Zhou Y, Zhou B, Pache L, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. 2019;10(1):1523. doi:10.1038/s41467-019-09234-6
47. Shannon P, Markiel A, Ozier O, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13(11):2498–2504. doi:10.1101/gr.1239303
48. Langfelder P, Horvath S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinf. 2008;9:559. doi:10.1186/1471-2105-9-559
49. Troubat R, Barone P, Leman S, et al. Neuroinflammation and depression: a review. Eur J Neurosci. 2021;53(1):151–171. doi:10.1111/ejn.14720
50. Trott O, Olson AJ. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. 2010;31(2):455–461. doi:10.1002/jcc.21334
51. Kim S, Chen J, Cheng T, et al. PubChem in 2021: new data content and improved web interfaces. Nucleic Acids Res. 2021;49(D1):D1388–D1395. doi:10.1093/nar/gkaa971
52. Consortium U. UniProt: the universal protein knowledgebase in 2021. Nucleic Acids Res. 2021;49(D1):D480–D489. doi:10.1093/nar/gkaa1100
53. Lovrić M, Molero JM, Kern R. PySpark and RDKit: moving towards Big Data in Cheminformatics. Mol Inform. 2019;38(6):e1800082. doi:10.1002/minf.201800082
54. Faul F, Erdfelder E, Buchner A, Lang AG. Statistical power analyses using G*Power 3.1: tests for correlation and regression analyses. Behav Res Methods. 2009;41(4):1149–1160. doi:10.3758/BRM.41.4.1149
55. Rossi A, Barraco A, Donda P. Fluoxetine: a review on evidence based medicine. Ann Gen Hosp Psychiatry. 2004;3(1):2. doi:10.1186/1475-2832-3-2
56. Baharivand N, Mahdavifard A, Fouladi RF. Intravitreal clindamycin plus dexamethasone versus classic oral therapy in toxoplasmic retinochoroiditis: a prospective randomized clinical trial. Int Ophthalmol. 2013;33(1):39–46. doi:10.1007/s10792-012-9634-1
57. Krukowski K, Nijboer CH, Huo X, Kavelaars A, Heijnen CJ. Prevention of chemotherapy-induced peripheral neuropathy by the small-molecule inhibitor pifithrin-μ. Pain. 2015;156(11):2184–2192. doi:10.1097/j.pain.0000000000000290
58. Welker HA, Wiltshire H, Bullingham R. Clinical pharmacokinetics of mibefradil. Clin Pharmacokinet. 1998;35(6):405–423. doi:10.2165/00003088-199835060-00001
59. Qiao Y, Zhao J, Li C, et al. Effect of combined chronic predictable and unpredictable stress on depression-like symptoms in mice. Ann Transl Med. 2020;8(15):942. doi:10.21037/atm-20-5168
60. Choleris E, Thomas AW, Kavaliers M, Prato FS. A detailed ethological analysis of the mouse open field test: effects of diazepam, chlordiazepoxide and an extremely low frequency pulsed magnetic field. Neurosci Biobehav Rev. 2001;25(3):235–260. doi:10.1016/s0149-7634(01)00011-2
61. Andreasen JT, Redrobe JP. Antidepressant-like effects of nicotine and mecamylamine in the mouse forced swim and tail suspension tests: role of strain, test and sex. Behav Pharmacol. 2009;20(3):286–295. doi:10.1097/FBP.0b013e32832c713e
62. Porsolt RD, Le Pichon M, Jalfre M. Depression: a new animal model sensitive to antidepressant treatments. Nature. 1977;266(5604):730–732. doi:10.1038/266730a0
63. Ialongo C. Understanding the effect size and its measures. Biochem Med. 2016;26(2):150–163. doi:10.11613/BM.2016.015
64. Guo LT, Wang SQ, Su J, et al. Baicalin ameliorates neuroinflammation-induced depressive-like behavior through inhibition of toll-like receptor 4 expression via the PI3K/AKT/FoxO1 pathway. J Neuroinflammation. 2019;16(1):95. doi:10.1186/s12974-019-1474-8
65. Humo M, Ayazgök B, Becker LJ, Waltisperger E, Rantamäki T, Yalcin I. Ketamine induces rapid and sustained antidepressant-like effects in chronic pain induced depression: role of MAPK signaling pathway. Prog Neuropsychopharmacol Biol Psychiatry. 2020;100:109898. doi:10.1016/j.pnpbp.2020.109898
66. Levy MJF, Boulle F, Steinbusch HW, van den Hove DLA, Kenis G, Lanfumey L. Neurotrophic factors and neuroplasticity pathways in the pathophysiology and treatment of depression. Psychopharmacology. 2018;235(8):2195–2220. doi:10.1007/s00213-018-4950-4
67. Wittmann M, Schreiber W, Landgrebe M, Hajak G. Störung zirkadianer Rhythmen im Kontext depressiver Erkrankungen [Circadian rhythms and depression]. Fortschr Neurol Psychiatr. 2018;86(5):308–318. doi:10.1055/s-0043-123069
68. Rotenstein LS, Ramos MA, Torre M, et al. Prevalence of Depression, Depressive Symptoms, and Suicidal Ideation Among Medical Students: a Systematic Review and Meta-Analysis. JAMA. 2016;316(21):2214–2236. doi:10.1001/jama.2016.17324
69. Ludka FK, Constantino LC, Dal-Cim T, et al. Involvement of PI3K/Akt/GSK-3β and mTOR in the antidepressant-like effect of atorvastatin in mice. J Psychiatr Res. 2016;82:50–57. doi:10.1016/j.jpsychires.2016.07.004
70. Zhang S, Lu Y, Chen W, et al. Network Pharmacology and Experimental Evidence: PI3K/AKT Signaling Pathway is Involved in the Antidepressive Roles of Chaihu Shugan San. Drug Des Devel Ther. 2021;15:3425–3441. doi:10.2147/DDDT.S315060
71. Behl T, Alotaibi GH, Shamsuzzaman M, et al. Polyphenols inhibiting MAPK signalling pathway mediated oxidative stress and inflammation in depression. Biomed Pharmacother. 2022;146:112545. doi:10.1016/j.biopha.2021.112545
72. Chan RF, Turecki G, Shabalin AA, et al. Cell Type-Specific Methylome-wide Association Studies Implicate Neurotrophin and Innate Immune Signaling in Major Depressive Disorder. Biol Psychiatry. 2020;87(5):431–442. doi:10.1016/j.biopsych.2019.10.014
73. Zeng J, Ji Y, Luan F, et al. Xiaoyaosan ethyl acetate fraction alleviates depression-like behaviors in CUMS mice by promoting hippocampal neurogenesis via modulating the IGF-1Rβ/PI3K/Akt signaling pathway. J Ethnopharmacol. 2022;288:115005. doi:10.1016/j.jep.2022.115005
74. Li X, Teng T, Yan W, et al. AKT and MAPK signaling pathways in hippocampus reveals the pathogenesis of depression in four stress-induced models. Transl Psychiatry. 2023;13(1):200. doi:10.1038/s41398-023-02486-3
75. Chen S, Peng J, Sherchan P, et al. TREM2 activation attenuates neuroinflammation and neuronal apoptosis via PI3K/Akt pathway after intracerebral hemorrhage in mice. J Neuroinflammation. 2020;17(1):168. doi:10.1186/s12974-020-01853-x
76. Dong N, Li X, Xue C, Zhang L, Wang C, Shan A. Astragalus polysaccharides alleviates LPS-induced inflammation via the NF-κB/MAPK signaling pathway. J Cell Physiol. 2020;235(7–8):5525–5540. doi:10.1002/jcp.29452
77. Chen H, Fu W, Chen H, et al. Magnolol attenuates the inflammation and enhances phagocytosis through the activation of MAPK, NF-κB signal pathways in vitro and in vivo. Mol Immunol. 2019;105:96–106. doi:10.1016/j.molimm.2018.11.008
78. Al-Qudah MA, Al-Dwairi A. Mechanisms and regulation of neurotrophin synthesis and secretion. Neurosciences. 2016;21(4):306–313. doi:10.17712/nsj.2016.4.20160080
79. Kar N, Misra B. Toxoplasma seropositivity and depression: a case report. BMC Psychiatry. 2004;4:1. doi:10.1186/1471-244X-4-1
80. Walf AA, Frye CA. Estradiol reduces anxiety- and depression-like behavior of aged female mice. Physiol Behav. 2010;99(2):169–174. doi:10.1016/j.physbeh.2009.09.017
81. Parry BL. Estradiol fluctuations and depressive history as risk factors for menopausal depression. Menopause. 2021;28(3):234–236. doi:10.1097/GME.0000000000001736
82. Zhang JC, Yao W, Hashimoto K. Brain-derived Neurotrophic Factor (BDNF)-TrkB Signaling in Inflammation-related Depression and Potential Therapeutic Targets. Curr Neuropharmacol. 2016;14(7):721–731. doi:10.2174/1570159x14666160119094646
83. Guo N, Wang X, Xu M, Bai J, Yu H, Zhang L. PI3K/AKT signaling pathway: molecular mechanisms and therapeutic potential in depression. Pharmacol Res. 2024;206:107300. doi:10.1016/j.phrs.2024.107300
84. Algahtany M, Sharma S, Fahoum K, et al. The Role of Growth Hormone in Depression: a Human Model. Front Neurosci. 2021;15:661819. doi:10.3389/fnins.2021.661819
85. Chesnokova V, Pechnick RN. Antidepressants and Cdk inhibitors: releasing the brake on neurogenesis? Cell Cycle. 2008;7(15):2321–2326. doi:10.4161/cc.6446
86. Rush AJ, Trivedi MH, Wisniewski SR, et al. Acute and longer-term outcomes in depressed outpatients requiring one or several treatment steps: a STAR*D report. Am J Psychiatry. 2006;163(11):1905–1917. doi:10.1176/ajp.2006.163.11.1905
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.