Back to Journals » OncoTargets and Therapy » Volume 12
Hypermethylated KLF9 Is An Independent Prognostic Factor For Favorable Outcome In Breast Cancer
Authors Wang L, Mao Q, Zhou S, Ji X ![]()
Received 6 August 2019
Accepted for publication 22 October 2019
Published 18 November 2019 Volume 2019:12 Pages 9915—9926
DOI https://doi.org/10.2147/OTT.S226121
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Takuya Aoki
Lei Wang,1,2 Qiqi Mao,1,2 Shaocheng Zhou,1,2 Xiaochun Ji1,2
1Department of Thyroid and Breast Surgery, Ningbo Medical Center Lihuili Hospital, Ningbo, Zhejiang, People’s Republic of China; 2Department of Thyroid and Breast Surgery, Lihuili Hospital Ningbo University, Ningbo, Zhejiang, People’s Republic of China
Correspondence: Xiaochun Ji
Department of Thyroid and Breast Surgery, Ningbo Medical Center Lihuili Hospital, Ningbo, Zhejiang, People’s Republic of China
Email [email protected]
Background and objective: Breast cancer (BC) is the most lethal human malignancy and is the leading cause of cancer-associated death in women worldwide. Krüppel-like factor 9 (KLF9) belongs to a family of transcriptional regulators and its role in BC has not been fully investigated.
Method: Data mining was used to analyze BC data from The Cancer Genome Atlas (TCGA) database, which was downloaded using the UCSC Xena browser. The differential expression and methylation level of KLF9 was analyzed in patients with BC and corresponding normal controls enrolled from our hospital. Besides, the correlation of KLF9 methylation and prognosis was explored, and gene set enrichment analysis (GSEA) was conducted to identify the potential signaling pathway of KLF9 involved.
Results: Both TCGA and BC tissues indicated hypermethylation of the KLF9 promoter region in patients with BC compared with normal controls, which might account for the dysregulation of KLF9 in patients with BC. Besides, hypermethylation of KLF9 was detected in patients with estrogen or progesterone receptor-positive and non-triple-negative disease. Further, hypermethylation of KLF9 was demonstrated to be a potential independent biomarker in obtaining favorable outcomes in BC. By GSEA, tumor-associated biological processes and signaling pathway were identified, which indicated that KLF9 might play a vital role in the carcinogenesis of BC.
Conclusion: KFL9 plays an important role in the carcinogenesis of BC through the multiple tumor-associated signaling pathway. The hypermethylation of KLF9 resulted in its reduced expression in BC, while the hypermethylation of KLF9 has potential in the prediction of favorable outcomes in BC.
Keywords: Krüppel-like factor 9, methylation, breast cancer, prognostic biomarker, GSEA
Introduction
Breast cancer (BC) is the most common cause of cancer-related death in women worldwide.1 Incidence rates are high in well-developed countries, whereas those in developing nations such as Africa and Asia, incidence rates have historically been relatively low, but have increased in recent decades.2 According to the report on the global burden of cancer by the International Agency for Research on Cancer, almost half of the cases and over half of the cancer deaths worldwide occurred in Asia in 2018.3 In 2018, there were approximately 2.1 million new cases of BC, comprising 25% of cancer cases among women, and almost 700,000 deaths, accounting for 16% of cancer-associated deaths among women.
Many established risk factors for BC are related to estrogen.4,5 Risk is increased by early age at menarche,6 later age at menopause,7 and obesity in postmenopausal women.8 Prospective studies have shown that high concentrations of endogenous estradiol are associated with increased risk of BC development.9 Childbearing reduces risk, with greater protection for early first birth and a larger number of births; breastfeeding also likely has a protective effect.10 Alcohol consumption increases BC risk, whereas physical activity is probably protective;11 however, these risk factors account for a minority of cases.
Transcription factors can contribute to carcinogenesis and the progression of various cancers.12 Transcriptomics-based screening of molecular signatures has identified multiple differentially expressed genes that are associated with decreasing in the overall survival (OS) of BC patients.13 The human positive cofactor 4 (PC4), initially identified as a transcriptional cofactor, exerted its oncogenic functions by directly binding to c-Myc promoters and inducing Warburg effect.14 The expression of PC4 has effects on sensitivity to paclitaxel of cancer cells and is associated with poor survival in patients with BC.15 Krüppel-like factors (KLFs) are DNA-binding transcription regulators that control essential cellular processes, such as proliferation, differentiation, migration, and maintenance of pluripotency.16–18 Previous studies showed that Krüppel-like factor 6 (KLF6) functions as a tumor suppressor in BC by reducing cell proliferation rate through increased c-Jun degradation and proliferating cell signaling.19,20 Krüppel-like factor 11 (KLF11) was hypermethylated in BC; this hypermethylation may be associated with low expression and cancer metastases.21 Krüppel-like factor 8 (KLF8) acts as an oncogene in lung adenocarcinoma, and Kaplan–Meier curves revealed that high expression of KLF8 was related to poor prognosis in patients with lung adenocarcinoma.22 Expression of Krüppel-like factor 15 (KLF15) was also found to be abnormally high in lung adenocarcinoma tissues and was correlated with tumor TNM stage, and has potential as a cancer prognostic marker.23 Krüppel-like factor 9 (KLF9) was previously reported as a basic transcription element-binding protein, due to its specific binding to the transcription element GC box in a gene promoter region.24 Further, KLF9 is implicated in the pathogenesis of several cancers, including endometrial cancer and other endocrine-responsive cancers of female reproductive tissues;25 however, the role of KLF9 in BC remains largely unknown. Here, we took advantage of public databases to explore the expression and methylation status of KLF9, and to identify the biological processes involving KLF9 and its co-expressed genes in the context of BC. Further, we also validated our findings using laboratory experiments.
Materials And Methods
Tissue Collection
A total of 144 BC and adjacent normal tissue samples were collected and stored in liquid nitrogen after surgery. All patients were enrolled in the thyroid and breast surgery department of Li Huili Affiliated Hospital of Ningbo University and they signed the informed consent forms before surgery and sample collection. All patients were pathologically diagnosed with BC by at least two experienced pathologists. The study was approved by the Human Research Ethical Committee of Ningbo Medical Center Lihuili Hospital.
DNA Extraction And Bisulfite Modification
Genomic DNA was extracted from patients with BC and normal tissue controls (n = 144) using DNA Mini Kit (Qiagen, Germany), according to the manufacturer’s protocol. The quality and concentration of DNA were measured. Extracted DNA was bisulfite-treated using the ZYMO EZ DNA Methylation-Gold Kit, as recommended by the manufacturer (Zymo Research, USA).
Quantitative Methylation-Specific Polymerase Chain Reaction (qMSP)
The promoter region of KLF9 was amplified by fluorescence-based qMSP, using SYBR Green Master Mix (Promega, USA). The qMSP primers for KLF9 were described in a previous study. The primer sequences of KLF9,26 along with those of primers for amplification of the control gene, ACTB27 are listed in Table 1.
 |
Table 1 Sequences Of KLF9 And ACTB Primers For qMSP |
Bioinformatic Analysis Using The UCSC Xena Browser
All microarray data (including methylation and expression information) from patients with BC and normal tissue controls, along with the clinical characteristics, were from the Cancer Genome Atlas (https://www.cancer.gov/, TCGA), which was downloaded using the UCSC Xena browser (https://xenabrowser.net/).28
Bioinformatic Analysis Using cBioPortal For Cancer Genomics And ClueGo
Genes co-expressed with KLF9 were identified using cBioPortal for Cancer Genomics (www.cbioportal.org/, |Pearson’s r| ≥ 0.4; P < 0.01).29 Then, the genes were loaded into ClueGo in Cytoscape version 3.71 for analysis of KEGG pathways.30
Gene Set Enrichment Analysis (GSEA) And Single-Sample GSEA (ssGSEA)
TCGA BC patients were classified into two groups, high expression and low expression, according to the median expression of KLF9. GSEA of KLF9 was performed using GSEA 3.0 software. An enrichment score (ES) >0.4 was obtained and false discovery rate (FDR) value <0.05 was regarded as statistically significant. To compare the activation degree of enriched pathways from GSEA, we used ssGSEA to generate activation pathway score.31 Using GSVA package and its ssGSEA method (http://www.bioconductor.org), the enrichment pathway score in each sample was calculated.32 The scatter plot of activated pathway score and KLF9 expression was generated by ggplots package on the R platform.
Statistical Analysis
Statistical analysis was performed using SPSS 13.0 (SPSS Inc., Chicago, IL, USA). Differences in KLF9 methylation and expression between BC and healthy control tissues, and associations between KLF9 methylation and clinicopathological features were evaluated by independent t-est or one-way ANOVA. Multivariate Cox regression models were constructed to analyze the prognostic power of clinical variables, using the “survival” package on the R platform (https://cran.r-project.org/web/packages/survival/survival.pdf). A heatmap of KLF9 methylation, clustered by sample type, was generated using the “pheatmap” package on the R platform (https://cran.r-project.org/web/packages/pheatmap/index.html). The ssGSEA analysis was performed using GSVA package, and the scatter plots were generated using ggplots package on the R platform. Other data were analyzed using GraphPad Prism 6 software (GraphPad, San Diego, CA). Two-tailed P values <0.05 were deemed statistically significant.
Results
KLF9 Is Significantly Downregulated In BC Tisuues Compared With Normal Control Tissues
In the initial data mining, we downloaded KLF9 expression data of 1108 BC and 139 normal control tissue samples using the UCSC Xena browser. After filtering out the samples with null values for KLF9 expression, a total of 1104 BC and 114 normal control tissue samples were included for further analysis. As shown in Figure 1, our results indicated that KLF9 expression was approximately 1.2-fold reduced in BC (cases vs. controls: 9.76 ± 0.87 vs. 11.56 ± 0.77, P = 4.96E-86).
 |
Figure 1 Downregulated expression of KLF9 in breast cancer. ***P < 0.001. |
Biological Processes Regulated By KLF9
By data mining using cBioPortal for Cancer Genomics, we identified genes co-expressed with KLF9 in BC (|Pearson’s r|≥0.4, P < 0.01; Table S1). A total of 13,010 genes were identified as co-expressed with KLF9 in BC. To further explore possible signaling pathways in which KLF9 may be involved, KLF9 co-expressed genes were subjected to pathway analysis. As shown in Figure 2, the co-expressed genes were enriched in multiple biological processes, including negative regulation of cellular processes, regulation of multicellular organization, cell migration, and cellular responses to growth factor stimuli.
 |
Figure 2 Biological processes involving genes co-expressed with KLF9. |
A GSEA was conducted between low and high expression of KLF9 to identify some associated KGEE signaling pathways. The most significant pathways were identified according to the ES and FDR q value. As shown in Table 2, the results showed 50 significantly enriched pathways (P value < 0.05). Figure 3 exhibited top 10 pathways correlated with cancer, such as MAPK signaling pathway, pathway in cancer, renal cell carcinoma and prostate cancer (ES > 0.45 and FDR < 0.05).
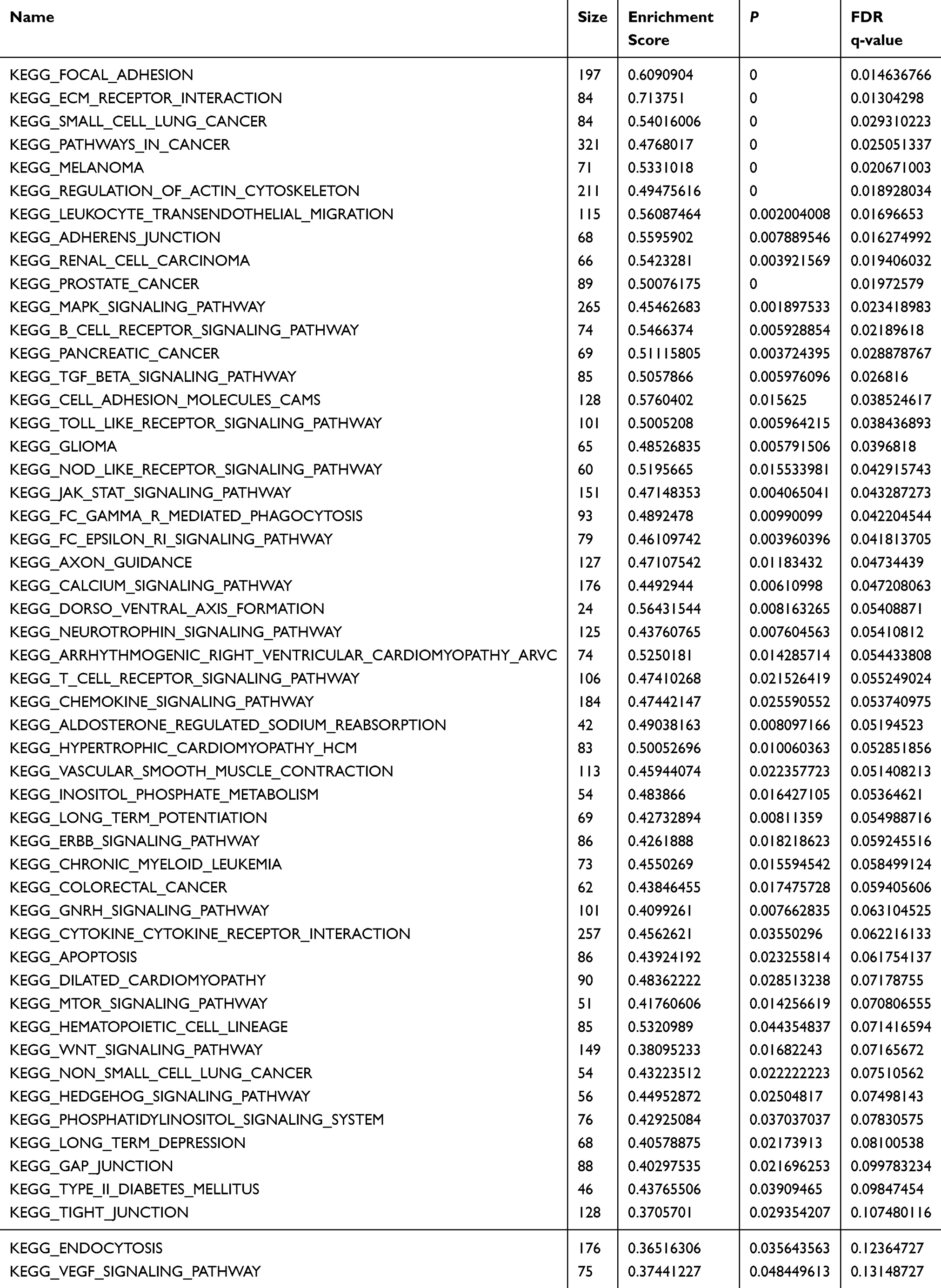 |
Table 2 Enriched KEGG Pathways From GSEA Results (P < 0.05) |
 |
Figure 3 The enrichment plots from GSEA results. |
Single-sample Gene Set Enrichment Analysis (ssGSEA) calculates separate ESs for each sample and pathway. Each ssGSEA ES represents the degree to which the genes in a particular pathway are coordinately up- or downregulated within a sample. Therefore, in order to validate the differentially enriched pathways associated with KLF9 expression from GSEA, we used the top 10 pathways shown in Figure 3 for ssGSEA. Then, the scatter plots about ssGSEA activation score of pathways and KLF9 levels were exhibited in Figure 4, and results showed most activated pathways score (especially MAPK signaling pathway) were significantly associated with KLF9 expression in BC.
 |
Figure 4 The scatter plots about ssGSEA activation score of pathways and KLF9 levels. |
Negative Correlation Of KLF9 Expression And Promoter Methylation
To determine the mechanism of downregulation of KLF9 in BC, we simultaneously obtained KLF9 methylation data from the UCSC Xena browser. First, the location of the KLF9 gene in the genome and the distribution of CpG sites at the locus were extracted from UCSC Genome Browser (http://genome.ucsc.edu/) (Figure 5). Subsequently, a heatmap of all KLF9 CpG methylation sites was constructed (Figure 6), which demonstrated that the methylation level of site cg00049440, located in the KLF9 promoter region was significantly higher in BC than normal control tissues (Figure 6). Finally, Pearson’s correlation analysis demonstrated a negative correlation between KLF9 expression and cg00049440 methylation, indicating that promoter methylation could be the mechanism underlying KLF9 dysregulation in BC (r = −0.34, P = 5.08E-23; Figure 7).
 |
Figure 5 KLF9 gene information from the UCSC Genome browser. |
 |
Figure 6 Heatmap of KLF9 methylation levels at CpG sites. |
 |
Figure 7 Negative correlation of KLF9 methylation and expression in the TCGA dataset. |
Associations Between KLF9 Methylation And Clinical Characteristics
Clinical features of BC were also downloaded using the UCSC Xena browser. There were 788 cases with both KLF9 methylation data and clinical features. Associations between KLF9 methylation and demographic and clinicopathological parameters in patients with BC are summarized in Table 3. The results showed that KLF9 methylation levels were significantly elevated in elderly patients (≥55 years), and patients with estrogen receptor-positive (ER+), progesterone receptor-positive (PR+), and non-triple-negative tumors.
 |
Table 3 Associations Of KLF9 Methylation With Clinical Characteristics Of BC In TCGA Dataset |
Prognostic Value Of KLF9 Expression And Methylation In BC
By performing multivariate analysis, we identified three clinical parameters associated with poor OS: age, lymph node metastasis, and distant organ metastasis (age, HR = 1.04, P = 3.3E-07; lymph node metastasis, HR = 1.79, P = 0.02; distant metastasis, HR = 4.42, P = 5.6E-05). Distant metastasis was associated with poor recurrence-free survival (RFS) (M stage: HR = 5.12, P = 3.66E-03). Moreover, methylation of KLF9 was an independent prognostic factor for superior OS (HR = 0.35, P = 0.015) and RFS (HR = 0.31, P = 0.035) of patients with BC (Table 4).
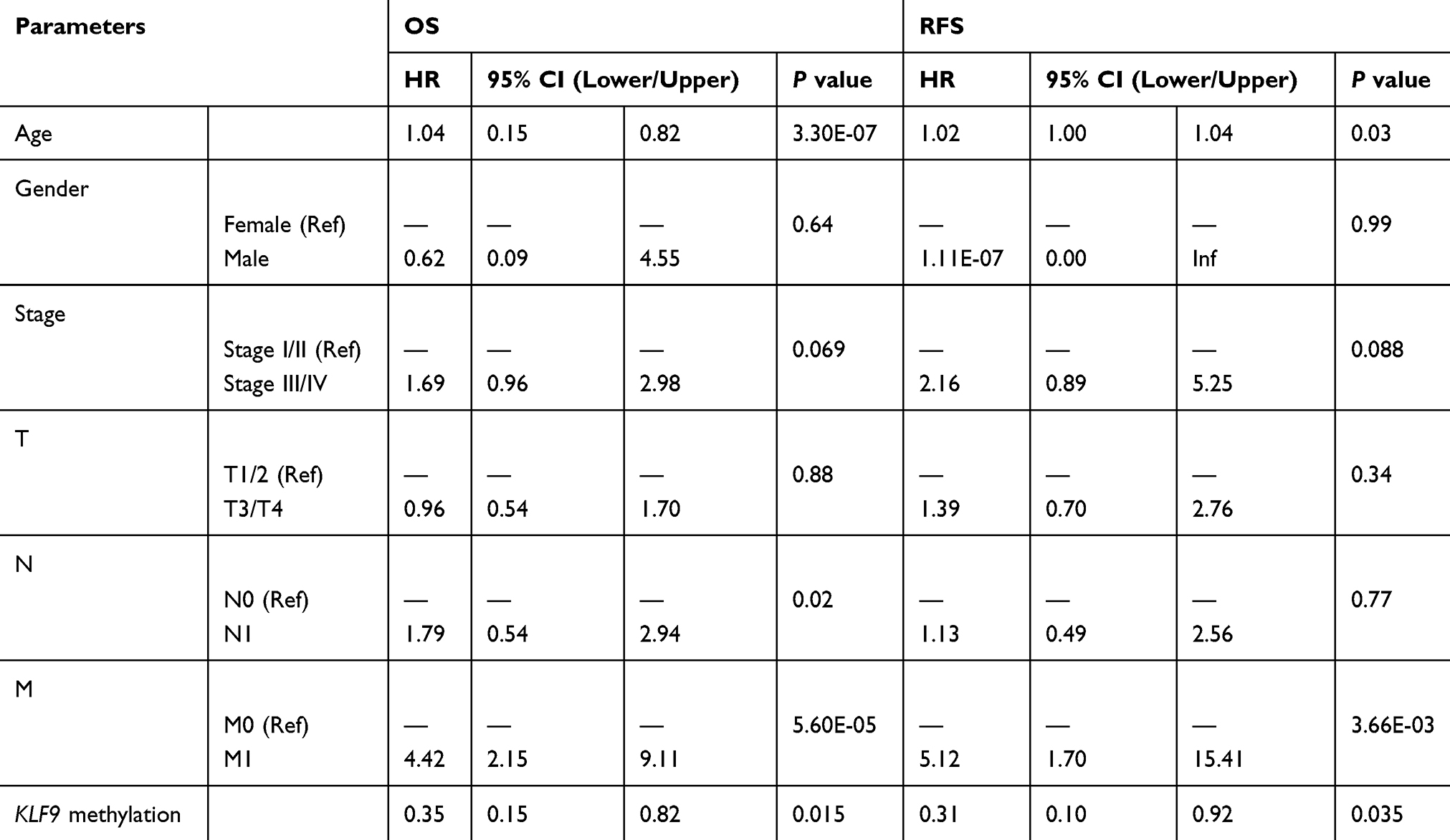 |
Table 4 Multivariate Analysis Of KLF9 Methylation In BC |
Hypermethylation Of KLF9 In A Validation Dataset
To verify the methylation levels of KLF9, a fragment of the KLF9 promoter region was amplified using qMSP. A total of 144 bisulfite-converted DNA residues from BC and adjacent normal control tissue samples were analyzed. The results demonstrated elevated KLF9 methylation levels in BC relative to controls (0.322 ± 0.193 vs. 0.113 ± 0.059, P = 3.89E-25; Figure 8). Next, subgroup analyses were conducted according to clinical features. The results were in accordance with our findings from analysis of TCGA dataset, showing elevated methylation level of KLF9 in patients with ER+, PR+, and non-triple-negative tumors (Table 5).
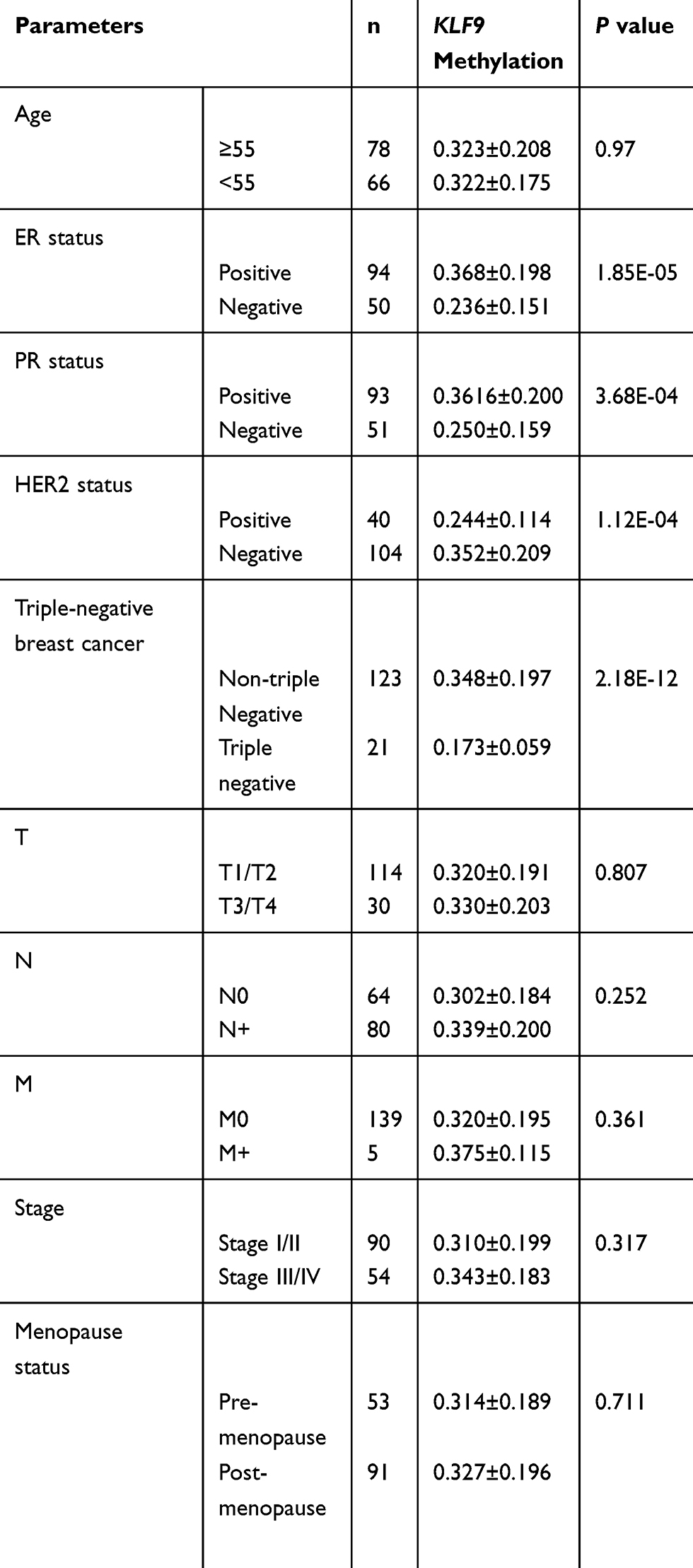 |
Table 5 Associations Of KLF9 Methylation With Clinical Characteristics Of BC In Our Validation Dataset |
 |
Figure 8 KLF9 methylation levels in our validation dataset. |
Discussion
BC is clinically heterogenous disease. Approximately 10–15% of patients with BC have aggressive disease and develop distant metastasis within 3 years after the initial detection of the primary tumor. Further, patients with BC are at risk of experiencing metastasis for their entire lifetime. The heterogenous nature of BC metastases hinders both the definition of cure for this disease and assessment of risk factors for metastasis.
Biomarkers that can predict the metastasis or prognosis of BC are also scarce. Although the risk of distant metastases increases with the presence of lymph node metastasis, larger-size primary tumor, and poor histopathological grade, which are established prognostic markers in BC,33–35 approximately one-third of women with BC that has not spread to the lymph nodes develop distant metastasis, and around one-third of those with BC that has spread to the lymph node remain free of distant metastasis 10 years after local therapy.35,36
KLFs are a family of transcriptional regulators characterized by a triple zinc finger DNA-binding domain with highly conserved C-terminal binding to GC-rich sequences.37–39 By influencing the expression of major regulatory factors, KLFs contribute to virtually all facets of cellular function, including cell proliferation, apoptosis, differentiation, and neoplastic transformation.37 An emerging body of evidence indicates that KLFs are associated with various types of cancers, and abnormal expression of KLF genes has been detected in multiple tumor types.40–42 The expression profile and functions of some KLFs are overlapping, while others are widely divergent. Reduction or loss of KLF6 has been reported in colorectal cancer.42 In BC, KLF5 expression is significantly reduced compared with that in matched normal tissues;40 however, KLF4 mRNA expression is increased in early and invasive BC,43 and KLF4 has attracted considerable attention for its opposing effects in carcinogenesis as tumor suppressor in gastrointestinal cancer44 or an oncoprotein in BC.45 As an important member of the KLF family, the role of KLF9 in BC remains largely unknown.
KLF9 was previously named basic transcription element-binding protein 1 (BTEB) and first identified as a trans-repressor of the CYP1A1 gene24 and then reported to induce CYP7A.46 KLF9 mRNA is most strongly expressed in the brain, kidney, lung, and testis.24 Previous studies have reported regulatory activity of KLF9 in the uterus during the development of BC and pregnancy,47 with KLF9 knockout mice exhibiting uterine hypoplasia, smaller little size, reduced numbers of implantation sites, partial progesterone resistance in the uterus, and delayed parturition.48,49 In addition to the roles of KLF9 in normal cells and tissues, it has important tumor suppressive and oncogenic functions in cancer. Well-characterized biological effects of KLF9 include a role in endometrial carcinogenesis.25 In the current study, our results also confirmed that KLF9 is mainly enriched in signaling pathway associated with carcinogenesis by GSEA analysis, including small cell lung cancer, prostate cancer, pancreatic cancer, pathway in cancer, and melanoma. In uterine endometrial cells, progesterone opposes the pro-proliferative effects of estrogen, and the absence of progesterone receptor (PR) signaling can promote cell proliferation.50 KLF9 can facilitate progesterone-induced effects on uterine gene expression by its co-recruitment to the PR51,52 and trans-inhibition of estrogen receptor α activity by promoting its estrogen-induced downregulation,53 suggesting that KLF9 inhibits proliferation in hormonally responsive cancers. Further, endometrial cancer cells with loss of KLF9 fail to be activated by estrogen, demonstrating that alteration of KLF9 expression may lead to escape from estrogen-mediated growth regulation.54 These findings suggest the intriguing likelihood that KLF9, regulated in concert with PR and ER, may serve as a prognostic predictor for hormonally responsive diseases.
In our study, we found that KLF9 mRNA expression was significantly reduced in patients with BC relative to normal controls, partly due to hypermethylation of the KLF9 gene promoter region. No significant difference in methylation was observed among different TNM stage tumors; however, an upward trend in methylation was detected in advanced TNM stage disease (T3–4 stage, N2 stage, stage III). The reverse phenomenon (lower KLF9 methylation) was exhibited in patients with T4, N3, M1, and clinical stage IV tumors, which may be attributable to the relatively small number of samples from these tumor stages (24 patients with T4, 58 patients with N3, 13 patients with M1, and 13 patients with stage IV). The results of subgroup analysis according to other clinical characteristics demonstrated higher methylation of KLF9 in patients with ER+, PR+, or non-triple-negative BC, which may be accounted for by the relationship of KLF9 expression with hormone levels in BC. These data suggest the possibility of acquired resistance to endocrine therapy following prolonged exposure to tamoxifen; that is, KLF9 expression may be re-induced in breast cells responsive to tamoxifen. This hypothesis requires confirmation by more rigorous and comprehensive prospective studies.
Additionally, we performed multivariate analysis demonstrating that age (HR for OS = 1.04, HR for RFS = 1.00), node metastasis status (HR for OS = 1.79, HR for RFS = 1.13), and distant metastasis (HR for OS = 4.42, HR for RFS = 5.12) are useful prognostic biomarkers of poor outcome in BC. Notably, hypermethylation of KLF9 exhibited the potential to act as a prognostic marker of favorable outcome in BC (HR for OS = 0.35, HR for RFS= 0.31).
Conclusion
In summary, our data demonstrate that hypermethylation of the KLF9 promoter region results in its downregulation in BC. Multivariate analysis showed that KLF9 methylation was associated with favorable prognosis in patients with BC. Further, the construction of an interaction network of genes co-expressed with KLF9 indicated that this factor may participate in breast carcinogenesis by contributing to cell migration and multiple growth regulation pathways. Moreover, we found that methylation of KLF9 is an independent prognostic factor for superior OS in patients with BC. These findings may inspire new clinical practices for patients with BC, including for diagnosis, treatment, and prognosis.
Disclosure
The authors declare that there are no conflicts of interest.
References
1. Ferlay J, Colombet M, Soerjomataram I, et al. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int J Cancer. 2019;144(8):1941–1953. doi:10.1002/ijc.v144.8
2. Bray F, McCarron P, Parkin DM. The changing global patterns of female breast cancer incidence and mortality. Breast Cancer Res. 2004;6(6):229–239. doi:10.1186/bcr932
3. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.v68.6
4. Clemons M, Goss P. Estrogen and the risk of breast cancer. N Engl J Med. 2001;344(4):276–285. doi:10.1056/NEJM200101253440407
5. Yager JD, Davidson NE. Estrogen carcinogenesis in breast cancer. N Engl J Med. 2006;354(3):270–282. doi:10.1056/NEJMra050776
6. Staszewski J. Age at menarche and breast cancer. J Natl Cancer Inst. 1971;47(5):935–940.
7. Trichopoulos D, MacMahon B, Cole P. Menopause and breast cancer risk. J Natl Cancer Inst. 1972;48(3):605–613.
8. Shieh Y, Scott CG, Jensen MR, et al. Body mass index, mammographic density, and breast cancer risk by estrogen receptor subtype. Breast Cancer Res. 2019;21(1):48.
9. Toniolo PG, Levitz M, Zeleniuch-Jacquotte A, et al. A prospective study of endogenous estrogens and breast cancer in postmenopausal women. J Natl Cancer Inst. 1995;87(3):190–197.
10. Collaborative Group on Hormonal Factors in Breast C. Breast cancer and breastfeeding: collaborative reanalysis of individual data from 47 epidemiological studies in 30 countries, including 50302 women with breast cancer and 96973 women without the disease. Lancet. 2002;360(9328):187–195. doi:10.1016/S0140-6736(02)09454-0
11. Dal Maso L, Zucchetto A, Talamini R, et al. Effect of obesity and other lifestyle factors on mortality in women with breast cancer. Int J Cancer. 2008;123(9):2188–2194. doi:10.1002/ijc.v123:9
12. Lee TI, Young RA. Transcriptional regulation and its misregulation in disease. Cell. 2013;152(6):1237–1251. doi:10.1016/j.cell.2013.02.014
13. Eskandari E, Motalebzadeh J. Transcriptomics-based screening of molecular signatures associated with patients overall survival and their key regulators in subtypes of breast cancer. Cancer Genet. 2019;239:62–74.
14. Luo P, Zhang C, Liao F, et al. Transcriptional positive cofactor 4 promotes breast cancer proliferation and metastasis through c-Myc mediated Warburg effect. Cell Commun Signal. 2019;17(1):36. doi:10.1186/s12964-019-0348-0
15. Camp NJ, Madsen MJ, Herranz J, et al. Re-interpretation of PAM50 gene expression as quantitative tumor dimensions shows utility for clinical trials: application to prognosis and response to paclitaxel in breast cancer. Breast Cancer Res Treat. 2019;175(1):129–139. doi:10.1007/s10549-018-05097-5
16. Jiang J, Chan YS, Loh YH, et al. A core Klf circuitry regulates self-renewal of embryonic stem cells. Nat Cell Biol. 2008;10(3):353–360. doi:10.1038/ncb1698
17. McConnell BB, Yang VW. Mammalian Kruppel-like factors in health and diseases. Physiol Rev. 2010;90(4):1337–1381. doi:10.1152/physrev.00058.2009
18. Yamanaka S. Strategies and new developments in the generation of patient-specific pluripotent stem cells. Cell Stem Cell. 2007;1(1):39–49. doi:10.1016/j.stem.2007.05.012
19. Gehrau RC, D’Astolfo DS, Dumur CI, Bocco JL, Koritschoner NP. Nuclear expression of KLF6 tumor suppressor factor is highly associated with overexpression of ERBB2 oncoprotein in ductal breast carcinomas. PLoS One. 2010;5(1):e8929. doi:10.1371/journal.pone.0008929
20. Slavin DA, Koritschoner NP, Prieto CC, Lopez-Diaz FJ, Chatton B, Bocco JL. A new role for the Kruppel-like transcription factor KLF6 as an inhibitor of c-Jun proto-oncoprotein function. Oncogene. 2004;23(50):8196–8205. doi:10.1038/sj.onc.1208020
21. Faryna M, Konermann C, Aulmann S, et al. Genome-wide methylation screen in low-grade breast cancer identifies novel epigenetically altered genes as potential biomarkers for tumor diagnosis. FASEB J. 2012;26(12):4937–4950. doi:10.1096/fj.12-209502
22. Liu Y, Yao X, Zhang Q, et al. Expression of Kruppel-like factor 8 and Ki67 in lung adenocarcinoma and prognosis. Exp Ther Med. 2017;14(2):1351–1356. doi:10.3892/etm.2017.4632
23. Gao L, Qiu H, Liu J, et al. KLF15 promotes the proliferation and metastasis of lung adenocarcinoma cells and has potential as a cancer prognostic marker. Oncotarget. 2017;8(66):109952–109961. doi:10.18632/oncotarget.v8i66
24. Imataka H, Sogawa K, Yasumoto K, et al. Two regulatory proteins that bind to the basic transcription element (BTE), a GC box sequence in the promoter region of the rat P-4501A1 gene. EMBO J. 1992;11(10):3663–3671. doi:10.1002/embj.1992.11.issue-10
25. Simmen RC, Pabona JM, Velarde MC, Simmons C, Rahal O, Simmen FA. The emerging role of Kruppel-like factors in endocrine-responsive cancers of female reproductive tissues. J Endocrinol. 2010;204(3):223–231. doi:10.1677/JOE-09-0329
26. Bethge N, Honne H, Hilden V, et al. Identification of highly methylated genes across various types of B-cell non-hodgkin lymphoma. PLoS One. 2013;8(11):e79602. doi:10.1371/journal.pone.0079602
27. Li J, Zhou C, Ni S, et al. Methylated claudin-11 associated with metastasis and poor survival of colorectal cancer. Oncotarget. 2017;8(56):96249–96262. doi:10.18632/oncotarget.21997
28. Cline MS, Craft B, Swatloski T, et al. Exploring TCGA pan-cancer data at the UCSC cancer genomics browser. Sci Rep. 2013;3:2652. doi:10.1038/srep02652
29. Zhou S, Wang P, Su X, et al. High ECT2 expression is an independent prognostic factor for poor overall survival and recurrence-free survival in non-small cell lung adenocarcinoma. PLoS One. 2017;12(10):e0187356. doi:10.1371/journal.pone.0187356
30. Bindea G, Galon J, Mlecnik B. CluePedia Cytoscape plugin: pathway insights using integrated experimental and in silico data. Bioinformatics. 2013;29(5):661–663. doi:10.1093/bioinformatics/btt019
31. Verhaak RG, Tamayo P, Yang JY, et al. Prognostically relevant gene signatures of high-grade serous ovarian carcinoma. J Clin Invest. 2013;123(1):517–525. doi:10.1172/JCI65833
32. Hanzelmann S, Castelo R, Guinney J. GSVA: gene set variation analysis for microarray and RNA-seq data. BMC Bioinformatics. 2013;14:7. doi:10.1186/1471-2105-14-7
33. Koscielny S, Tubiana M, Le MG, et al. Breast cancer: relationship between the size of the primary tumour and the probability of metastatic dissemination. Br J Cancer. 1984;49(6):709–715. doi:10.1038/bjc.1984.112
34. Page DL. Prognosis and breast cancer. Recognition of lethal and favorable prognostic types. Am J Surg Pathol. 1991;15(4):334–349. doi:10.1097/00000478-199104000-00002
35. Rosen PP, Groshen S, Saigo PE, Kinne DW, Hellman S. Pathological prognostic factors in stage I (T1N0M0) and stage II (T1N1M0) breast carcinoma: a study of 644 patients with median follow-up of 18 years. J Clin Oncol. 1989;7(9):1239–1251. doi:10.1200/JCO.1989.7.9.1239
36. Ponten J. Natural history of breast cancer. Acta Oncol. 1990;29(3):325–329. doi:10.3109/02841869009090008
37. Black AR, Black JD, Azizkhan-Clifford J. Sp1 and Kruppel-like factor family of transcription factors in cell growth regulation and cancer. J Cell Physiol. 2001;188(2):143–160. doi:10.1002/jcp.1111
38. Dang DT, Pevsner J, Yang VW. The biology of the mammalian Kruppel-like family of transcription factors. Int J Biochem Cell Biol. 2000;32(11–12):1103–1121. doi:10.1016/S1357-2725(00)00059-5
39. Tupler R, Perini G, Green MR. Expressing the human genome. Nature. 2001;409(6822):832–833. doi:10.1038/35057011
40. Chen C, Bhalala HV, Qiao H, Dong JT. A possible tumor suppressor role of the KLF5 transcription factor in human breast cancer. Oncogene. 2002;21(43):6567–6572. doi:10.1038/sj.onc.1205817
41. Chen CJ, Lin SE, Lin YM, Lin SH, Chen DR, Chen CL. Association of expression of Kruppel-like factor 4 and Kruppel-like factor 5 with the clinical manifestations of breast cancer. Pathol Oncol Res. 2012;18(2):161–168. doi:10.1007/s12253-011-9422-7
42. Cho YG, Choi BJ, Song JW, et al. Aberrant expression of Kruppel-like factor 6 protein in colorectal cancers. World J Gastroenterol. 2006;12(14):2250–2253. doi:10.3748/wjg.v12.i14.2250
43. Foster KW, Frost AR, McKie-Bell P, et al. Increase of GKLF messenger RNA and protein expression during progression of breast cancer. Cancer Res. 2000;60(22):6488–6495.
44. Wei D, Kanai M, Huang S, Xie K. Emerging role of KLF4 in human gastrointestinal cancer. Carcinogenesis. 2006;27(1):23–31. doi:10.1093/carcin/bgi243
45. Pandya AY, Talley LI, Frost AR, et al. Nuclear localization of KLF4 is associated with an aggressive phenotype in early-stage breast cancer. Clin Cancer Res. 2004;10(8):2709–2719. doi:10.1158/1078-0432.CCR-03-0484
46. Foti D, Stroup D, Chiang JY. Basic transcription element binding protein (BTEB) transactivates the cholesterol 7 alpha-hydroxylase gene (CYP7A). Biochem Biophys Res Commun. 1998;253(1):109–113. doi:10.1006/bbrc.1998.9759
47. Pabona JM, Zhang D, Ginsburg DS, Simmen FA, Simmen RC. Prolonged pregnancy in women is associated with attenuated myometrial expression of progesterone receptor co-regulator Kruppel-like Factor 9. J Clin Endocrinol Metab. 2015;100(1):166–174. doi:10.1210/jc.2014-2846
48. Simmen RC, Eason RR, McQuown JR, et al. Subfertility, uterine hypoplasia, and partial progesterone resistance in mice lacking the Kruppel-like factor 9/basic transcription element-binding protein-1 (Bteb1) gene. J Biol Chem. 2004;279(28):29286–29294. doi:10.1074/jbc.M403139200
49. Zeng Z, Velarde MC, Simmen FA, Simmen RC. Delayed parturition and altered myometrial progesterone receptor isoform A expression in mice null for Kruppel-like factor 9. Biol Reprod. 2008;78(6):1029–1037. doi:10.1095/biolreprod.107.065821
50. Kumar NS, Richer J, Owen G, Litman E, Horwitz KB, Leslie KK. Selective down-regulation of progesterone receptor isoform B in poorly differentiated human endometrial cancer cells: implications for unopposed estrogen action. Cancer Res. 1998;58(9):1860–1865.
51. Velarde MC, Iruthayanathan M, Eason RR, Zhang D, Simmen FA, Simmen RC. Progesterone receptor transactivation of the secretory leukocyte protease inhibitor gene in Ishikawa endometrial epithelial cells involves recruitment of Kruppel-like factor 9/basic transcription element binding protein-1. Endocrinology. 2006;147(4):1969–1978. doi:10.1210/en.2005-1419
52. Zhang XL, Zhang D, Michel FJ, Blum JL, Simmen FA, Simmen RC. Selective interactions of Kruppel-like factor 9/basic transcription element-binding protein with progesterone receptor isoforms A and B determine transcriptional activity of progesterone-responsive genes in endometrial epithelial cells. J Biol Chem. 2003;278(24):21474–21482. doi:10.1074/jbc.M212098200
53. Velarde MC, Zeng Z, McQuown JR, Simmen FA, Simmen RC. Kruppel-like factor 9 is a negative regulator of ligand-dependent estrogen receptor alpha signaling in Ishikawa endometrial adenocarcinoma cells. Mol Endocrinol. 2007;21(12):2988–3001. doi:10.1210/me.2007-0242
54. Simmons CD, Pabona JM, Heard ME, et al. Kruppel-like factor 9 loss-of-expression in human endometrial carcinoma links altered expression of growth-regulatory genes with aberrant proliferative response to estrogen. Biol Reprod. 2011;85(2):378–385. doi:10.1095/biolreprod.110.090654
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.