Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 7
Human ABC transporter ABCG2/BCRP expression in chemoresistance: basic and clinical perspectives for molecular cancer therapeutics
Authors Noguchi K, Katayama K, Sugimoto Y
Received 2 December 2013
Accepted for publication 16 January 2014
Published 5 February 2014 Volume 2014:7 Pages 53—64
DOI https://doi.org/10.2147/PGPM.S38295
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 6
Kohji Noguchi, Kazuhiro Katayama, Yoshikazu Sugimoto
Division of Chemotherapy, Faculty of Pharmacy, Keio University, Tokyo, Japan
Abstract: Adenine triphosphate (ATP)-binding cassette (ABC) transporter proteins, such as ABCB1/P-glycoprotein (P-gp) and ABCG2/breast cancer resistance protein (BCRP), transport various structurally unrelated compounds out of cells. ABCG2/BCRP is referred to as a “half-type” ABC transporter, functioning as a homodimer, and transports anticancer agents such as irinotecan, 7-ethyl-10-hydroxycamptothecin (SN-38), gefitinib, imatinib, methotrexate, and mitoxantrone from cells. The expression of ABCG2/BCRP can confer a multidrug-resistant phenotype on cancer cells and affect drug absorption, distribution, metabolism, and excretion in normal tissues, thus modulating the in vivo efficacy of chemotherapeutic agents. Clarification of the substrate preferences and structural relationships of ABCG2/BCRP is essential for our understanding of the molecular mechanisms underlying its effects in vivo during chemotherapy. Its single-nucleotide polymorphisms are also involved in determining the efficacy of chemotherapeutics, and those that reduce the functional activity of ABCG2/BCRP might be associated with unexpected adverse effects from normal doses of anticancer drugs that are ABCG2/BCRP substrates. Importantly, many recently developed molecular-targeted cancer drugs, such as the tyrosine kinase inhisbitors, imatinib mesylate, gefitinib, and others, can also interact with ABCG2/BCRP. Both functional single-nucleotide polymorphisms and inhibitory agents of ABCG2/BCRP modulate the in vivo pharmacokinetics and pharmacodynamics of these molecular cancer treatments, so the pharmacogenetics of ABCG2/BCRP is an important consideration in the application of molecular-targeted chemotherapies.
Keywords: kinase inhibitor, SNP, single-nucleotide polymorphisms, molecular target
Introduction
Various mechanisms are involved in the multidrug resistance of cancer cells, including reduced drug uptake, the efflux of intracellular drugs, the activation of deoxyribonucleic acid (DNA) repair pathways, and the induction of the antiapoptotic machinery.1 The adenine triphosphate (ATP)-binding cassette (ABC) transporter proteins, particularly P-glycoprotein (P-gp) (P-gp/MDR1 [multidrug resistance 1]/ABCB1), multidrug resistance protein 1 (MRP1) (MRP1/ABCC1), and breast cancer resistance protein (BCRP) (BCRP/MXR [mitoxantrone resistance]/ABCP/ABCG2), function as key molecules in the multidrug-resistant phenotype of cancer cells.2–4 They mediate the ATP-dependent unidirectional efflux of various compounds, both endogenous and exogenous, from cells.2–5
ABCG2/BCRP, a “half transporter” member of the ABCG subfamily,6 has been found in cancer-drug-resistant human cancer cell lines isolated by in vitro selection.7–9 The overexpression of ABCG2/BCRP confers resistance to various chemotherapeutic drugs, such as the topoisomerase I inhibitor topotecan and the antifolate agent methotrexate,9,10 and ABCG2/BCRP is probably associated with clinical drug resistance, including that in patients with acute myelogenous leukemia or acute lymphocytic leukemia.11–17 Because ABCB1/P-gp is quite an important factor in the drug resistance observed in clinical leukemia, a large-scale analysis that assesses both ABCG2/BCRP and ABCB1/P-gp is essential to properly delineating the contribution of ABCG2/BCRP to drug resistance in cancer patients.
ABCG2/BCRP structure and activity
ABCG2/BCRP is a 655 amino acid, 72 kDa protein with a single ABC signature domain within the nucleotide-binding domain and six transmembrane domains, as shown in Figure 1. Structural and functional studies of ABCG2/BCRP have provided valuable insight into the molecular mechanisms of ABCG2/BCRP-mediated transport. The cloning of ABCG2/BCRP cDNAs (complementary DNAs) from drug-selected clone cells and normal tissues has revealed functional variations in the amino acid substitutions in the ABCG2/BCRP protein and altered substrate preferences. ABCG2/BCRP proteins in drug-selected cells, such as the S1-M1-80 and MCF7/AdVp3000 cell lines, are mutant forms, and unique mutations in ABCG2/BCRP have been identified at amino acid position 482.18 MCF7/AdVp3000 and S1-M1-80 cells express R482T and R482G variants of ABCG2/BCRP, respectively, and are highly resistant to both mitoxantrone and doxorubicin. Anthracyline resistance and a rhodamine efflux capacity are also unique phenotypes of these two ABCG2/BCRP-overexpressing cell lines.18,19 The substitution of Arg at position 482 in ABCG2/BCRP with Gly or Thr confers additional efflux activity for rhodamine 123, doxorubicin, and other anthracyclines, which are usually not good substrates of wild-type ABCG2/BCRP.18,20 In contrast, the ABCG2/BCRP variants R482G and R482T lose their methotrexate-transporting activity but, at the same time, confer increased mitoxantrone resistance.19,21–23 The COOH terminus of the transmembrane 3 region, in close proximity to position 482, is involved in the substrate-binding pocket of ABCG2/BCRP, and the Arg residue at position 482 affects drug–ABCG2/BCRP interactions.23–25 Moreover, 13 variant ABCG2/BCRP proteins with substitutions at R482 (R482N, C, M, S, T, V, A, G, E, W, D, Q, and H, but not Y or K) confer strong resistance to doxorubicin and mitoxantrone in PA317 cells, as in S1-M1-80 and MCF7/AdVp3000 cells.26 Thus, structural variations in ABCG2/BCRP affect its drug efflux activity. In addition, mutations in ABCG2/BCRP at N557 and H630 severely affect this resistant phenotype. Cells expressing either N557D or H630E mutant ABCG2/BCRP displayed lower resistance to SN-38 (7-ethyl-10-hydroxycamptothecin), although the mitoxantrone resistance of these cells was similar to that of cells expressing wild-type ABCG2/BCRP.26 Structural studies using three-dimensional homology modeling of ABCG2/BCRP have also indicated that the transmembrane domains of ABCG2/BCRP function as drug-recognition interfaces24,25,27 and suggest that R482 is located in the central cavity of the protein, with its side chain pointing toward the drug translocation pathway.28 Therefore, the drug efflux activity of ABCG2/BCRP is influenced by various mutations, which will necessarily affect the clinical efficacy of ABCG2/BCRP-transportable anticancer drugs.
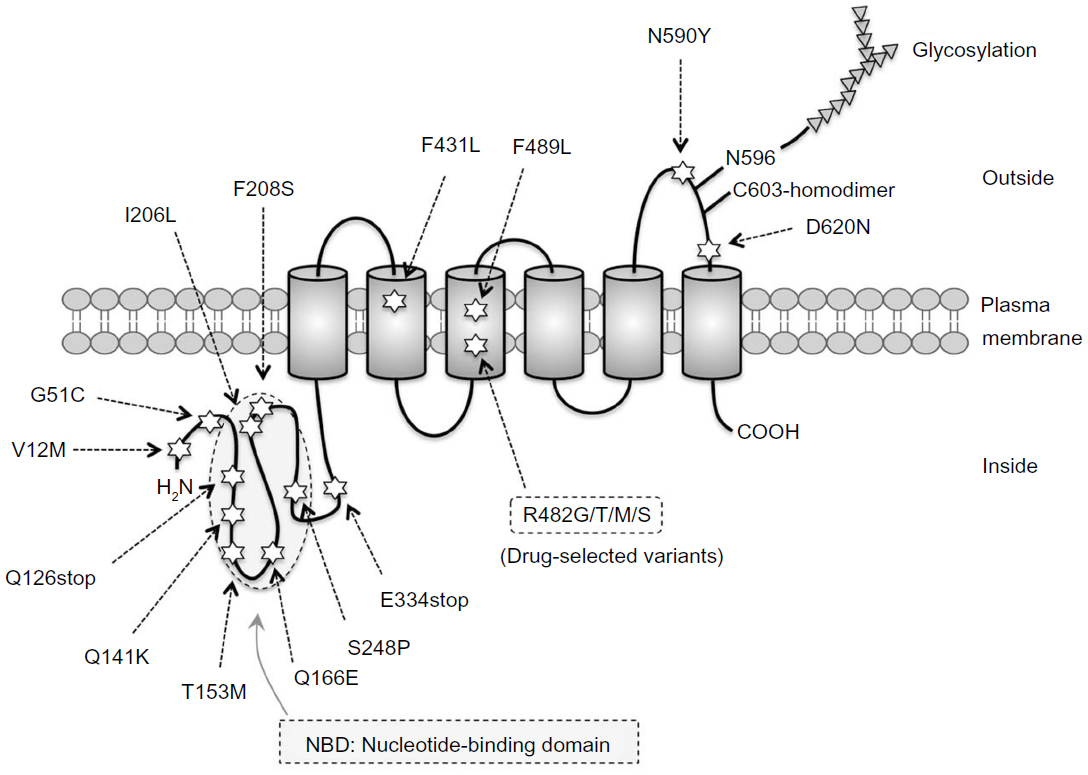 | Figure 1 Schematic diagram of ABCG2/BCRP protein and its amino acid variations. Amino acid variations by single-nucleotide polymorphisms are marked with stars. Transmembrane domains are shown as cylinders. |
ABCG2/BCRP functions as a homodimer.29,30 A disulfide bridge at C603 is involved in homodimer formation,31 and crystallization studies suggest a tetrameric complex composed of four ABCG2/BCRP homodimers.32 ABC transporter research has also demonstrated that the activity of these proteins is affected by both posttranslational regulation33,34 and genetic polymorphisms.35–37 ABCG2/BCRP is targeted to the plasma membrane via the endoplasmic reticulum–Golgi pathway, and during its passage, undergoes N-linked glycosylation at N59638,39 and the formation of an intramolecular disulfide bond between C592 and C608, which affects its stability.40 Oligomeric glycosylated ABCG2/BCRP is degraded in the lysosome, whereas underglycosylated, misfolded ABCG2/BCRP lacking the intramolecular disulfide bond is targeted to the proteasomal system for degradation.39
Single-nucleotide polymorphisms (SNPs) of ABCG2/BCRP
Primary structural variations of ABCG2/BCRP are associated with its drug-transporter function, as described above. Numerous germ-line mutations in the ABCG2/BCRP gene have been found in ethnically diverse populations,36,41–43 as shown in Figure 1. Therefore, SNPs in the ABCG2/BCRP gene would influence the pharmacological effects of ABCG2/BCRP differently in different patients.
421C>A (Q141K) ABCG2/BCRP SNP
Japanese population and human cancer cell lines have been shown to have three variant ABCG2/BCRP cDNAs with the substitutions: 34G>A (V12M), 421C>A (Q141K), and a nucleotide 944–949 deletion, removing A315 and T316 (Δ315–316).44 The 34G>A and 421C>A variants are SNPs. The frequency of the 421C>A SNP in a normal Japanese population demonstrated that 57 of 124 samples had the A421 allele, and nine of these were homozygous for this polymorphism.36,45 These data suggest that some Japanese individuals probably express low levels of ABCG2/BCRP. The 421C>A SNP appears to be very common in Asian populations, with reported allelic frequencies between 27% and 34%,35,44,46 whereas this SNP is rare in sub-Saharan African and African-American populations, with frequencies of <5%.47 Its frequency in Caucasian populations is approximately 10%.48 The physiological significance of the 421C>A ABCG2/BCRP SNP has been analyzed in relation to the pharmacokinetics of diflomotecan, a new camptothecin derivative anticancer agent, during a Phase I study.49 This study showed that five patients who were heterozygous for the A421 allele had much higher plasma levels of diflomotecan after its intravenous administration than 15 homozygous wild-type individuals (mean values of 138 ng h/mL/mg versus 46.1 ng h/mL/mg, respectively). Consistent with this, 421C>A ABCG2/BCRP-transfected murine fibroblast PA317 (PA/Q141K) cells expressed less exogenous ABCG2/BCRP protein levels than wild-type ABCG2/BCRP-transfected cells.44 Drug accumulation was higher in PA/Q141K cells than in other ABCG2/BCRP transfectants, suggesting that the SNP 421C>A (Q141K) reduces ABCG2/BCRP function. These observations from laboratory and clinical studies suggest that the levels and functions of ABCG2/BCRP expressed from the 421C>A ABCG2/BCRP allele are reduced compared with those of the wild-type protein. The Q141K mutation is located within the functionally important ATP-binding region between the Walker A and B motifs of ABCG2/BCRP and thus is likely to affect the ATPase activity of the protein.35,50
In terms of the genetic polymorphisms of ABCG2/BCRP and the expression of the protein, other non-synonymous SNPs, F208S and S441N, have been shown to affect ABCG2/BCRP protein levels at the plasma membrane, indicating that these non-synonymous SNPs reduce the stability of ABCG2/BCRP by enhancing its ubiquitin-mediated proteasomal proteolysis, resulting in reduced ABCG2/BCRP function.51
376C>T (Q126stop) ABCG2/BCRP SNP
Another SNP within the ABCG2/BCRP gene, 376C>T, is present at low frequencies in healthy Japanese samples as a heterozygosity (reported frequencies of 3/124 and 2/120 in two studies).44,47 The frequency of the T376 allele of ABCG2/BCRP is low and is not observed in Caucasian or African-American groups. A combination of the 376C>T and 421C>A SNPs would be expected to occur in a considerable proportion of the Japanese population. Because these SNPs would have negative effects on ABCG2/BCRP activity, the combined 376C>T 421T/C>A variant is expected to show severely reduced ABCG2/BCRP activity.
Additional ABCG2/BCRP SNPs
Other ABCG2/BCRP SNPs, such as 34G>A, 151G>T, 376C>T, 421C>A, 458C>T, 496C>T, 616A>C, 623T>C, 742T>C, 1000G>T, 1291T>C, 1465T>C, 1768A>T, and 1858G>A, cause amino acid substitutions. Synonymous mutations, such as 114T>C, 369C>T, 474C>T, 564A>G, 1098G>A, and 1425A>G, have been identified in the coding region of ABCG2/BCRP. The highest allele frequency for the 34G>A SNP is observed in Mexican-Indians, and there are significant differences in the frequencies of this SNP in Caucasian, Japanese, and Swedish populations.52,53 Transfection studies of ABCG2/BCRP (V12M) encoding the 34G>A SNP showed that the expression levels and drug-resistance of this variant are similar to those of wild-type ABCG2/BCRP, suggesting that V12M does not affect ABCG2/BCRP protein activity.44 However, a recent report suggested a possible association between this polymorphism and the alternative splicing of ABCG2/BCRP mRNA (messenger ribonucleic acid), particularly the liver-specific splicing of polymorphic exon 2 in these transcripts.54 ABCG2/BCRP mRNA expression was significantly lower in Hispanic livers with the 34G>A variant genotype, and individuals with the 34G>A allele displayed reduced ABCG2/BCRP expression in their liver cells. Although these observations suggest that chemotherapy regimens containing ABCG2/BCRP-substrate anticancer drugs may have increased efficacy in these patients because the expression of ABCG2/BCRP is reduced, another study has shown that Chinese acute myelogenous leukemia patients with the wild-type 34GG genotype had longer disease-free survival and longer overall survival than those with the 34GA/AA genotypes.55 The reason for the unpredictable effects of the 32G>A SNP on ABCG2/BCRP is unknown. The polymorphic and differential expression of another splicing variant of ABCG2/BCRP mRNA, involving exon 1b in the liver, also seems to be associated with lower ABCG2/BCRP expression.56 Approximately 90% of patients with the 34G>A SNP of ABCG2/BCRP show exon 2 skipping in the liver, and the lower level of ABCG2/BCRP mRNA in the liver may be associated with this SNP in the Hispanic population.54
ABCG2/BCRP expression and physiological functions
ABCG2/BCRP is widely expressed in the placenta, blood–brain barrier, gastrointestinal tract, liver, kidney, testis, and lactating breast. ABCG2/BCRP localizes apically in the epithelia of the small intestine and colon,57 on the luminal surfaces of the endothelial cells of the human brain,58 and on the luminal surfaces of the kidney tubules.59 Experimental inhibition of ABCG2/BCRP activity affected drug distribution in various in vivo experiments.60,61 These observations suggest a possible role for ABCG2/BCRP in controlling the absorption/distribution of its substrate compounds. Various cancers also contain subpopulations of stem cells that are characterized by the expression of ABCG2/BCRP and other ABC transporters.62 These transporters have been suggested to play an important role in the multidrug resistance of these cancer stem-like cells during chemotherapy.63 Indeed, the experimental inhibition of ABCG2/BCRP has been shown to suppress the proliferation of side population cells in cancer cell lines.64
ABCG2/BCRP extrudes various types of compounds among its functional substrates, including sulfated hormone metabolites, the chlorophyll metabolite pheophorbide a, fluorescent dyes such as Hoechst 33342 and BODIPY (boron-dipyrromethene)-prazosin, cimetidine, various flavonoids, and some antibiotics.30,65 Porphyrin/heme was the first-identified endogenous ABCG2/BCRP substrate,66 and ABCG2/BCRP regulates heme homeostasis under hypoxic conditions.67 ABCG2/BCRP also transports endogenous folates, such as the mono-, di-, and triglutamates of folic acid, and participates in the energy-dependent efflux of certain folates and antifolates.22 Recently, genome-wide screening for the genetic determinants of gout found that an SNP of ABCG2/BCRP (Q141K) is associated with high uric acid levels, and demonstrated that uric acid is a natural substrate of ABCG2/BCRP.68,69
ABCG2/BCRP also appears to play a protective role against xenobiotics and their metabolites.30,70 The typical ABCG2/BCRP substrates, irinotecan and SN-38, are detoxified by glucuronidation with uridine-diphosphate–glucuronyltransferase, and ABCG2/BCRP can extrude SN-38–glucuronide.71 Interestingly, ABCG2/BCRP can transport another of the glucuronide conjugates, 17-β-estradiol 17-(β-d-glucuronide), and the sulfated conjugates estrone-3-sulfate and dehydroepiandrosterone are also substrates of ABCG2/BCRP.22,70 The apical localization of ABCG2/BCRP in the intestinal epithelium and the bile canalicular membrane also suggests the intestinal absorption and hepatobiliary excretion of ABCG2/BCRP substrates.57,72–74 ABCG2/BCRP may also play a protective role by transporting dietary carcinogens. A study by van Herwaarden et al72 showed that BCRP1 effectively restricts the exposure of mice to the ingested food carcinogen 2-amino-1-methyl-6-phenylimidazo(4,5-b)pyridine by reducing its uptake from the gut lumen and by mediating its hepatobiliary and intestinal elimination. Similarly, ABCG2/BCRP limits the intestinal uptake of the carcinogens 2-amino-3-methylimidazo(4,5-f)quinoline, 3-amino-1,4-dimethyl-5H-pyrido(4,3-b)indole, and aflatoxin B1 in a mouse model.75 Overall, ABCG2/BCRP may restrict the bioavailability of orally administered ABCG2/BCRP-substrate anticancer agents, such as topotecan (and its metabolite SN-38), irinotecan, camptothecin derivatives, methotrexate, flavopiridol, and others.65 Therefore, the functional activity of ABCG2/BCRP is an important consideration in ABCG2/BCRP-transportable drug absorption, distribution, metabolism, and excretion in patients.
Pharmacological interaction of ABCG2/BCRP with molecular-targeted drugs
ABCG2/BCRP and anticancer kinase inhibitors
A growing number of small-molecule inhibitors of oncogenic kinases have come into clinical use and have shown great potential as anticancer drugs.76,77 Imatinib mesylate, which targets BCR-ABL (B-cell receptor-Abelson), was the first approved protein kinase inhibitor.78 Imatinib is very effective against chronic myeloid leukemia and other cancers associated with the deregulation of these kinase pathways. Resistance to this drug is typically conferred by mutations in the target kinase within the region of the drug–kinase interaction,79–82 but another mechanism leading to imatinib resistance correlates with ABCB1/P-gp expression.83,84 As shown in Table 1, a number of recent studies have indicated possible interactions between many kinase inhibitors and ABC transporters, including ABCB1/P-gp and ABCG2/BCRP.85–90 ABCG2/BCRP has a potent ability to interact with numerous clinically important kinase inhibitors, including imatinib,87–89,91–99 nilotinib,95 dasatinib,96 lapatinib,97 gefitinib,42,98–105 canertinib,106 erlotinib,107–109 sorafenib,110 pazopanib,111 vandetanib,112 vemurafenib,113 axitinib,114 and ponatinib.115 In contrast, bosutinib is unlikely to be a substrate of ABCG2/BCRP but probably inhibits it.116 A recently developed anaplastic lymphoma kinase inhibitor, crizotinib, is also unlikely to interact with ABCG2/BCRP, although it is a good substrate of ABCB1/P-gp.117
 | Table 1 Interactions between tyrosine kinase inhibitors and ABCG2/BCRP |
A variety of kinase-inhibiting compounds appear to interact with ABC transporters.85,118–124 These protein kinase inhibitors are designed to compete with ATP in the kinase domain and thus show competitive suppressive effects.125 Therefore, most ABCG2/BCRP-interactive kinase inhibitors used at higher concentration were initially suspected to block the ATPase activity of this protein. However, Saito et al126 demonstrated that gefitinib binds to ATP-bound ABCG2/BCRP, indicating that the as yet undetermined gefitinib-binding site in ABCG2/BCRP is not in the ATP-binding domain. Using photoaffinity labeling technique with 125I-labeled iodoarylazidoprazosin, a typical substrate of ABCB1/P-gp and ABCG2/BCRP, Brendel et al95 demonstrated that imatinib and nilotinib bind to ABCG2/BCRP at the substrate-interaction site, whereas Shi et al107 showed that erlotinib does not compete with iodoarylazidoprazosin at the substrate-binding sites on ABCG2/BCRP or ABCB1/P-gp. Some interesting studies have proposed the presence of multiple drug-binding sites in this ABC transporter,127,128 suggesting that kinase inhibitors may target these substrate-binding pockets. Further studies are required to properly clarify the modes of interaction between kinase inhibitors and ABCG2/BCRP.
Pharmacological interactions affected by ABCG2/BCRP polymorphisms
Studies of the pharmacological interactions between ABCG2/BCRP and molecular-targeted kinase inhibitors have revealed that clinically used kinase inhibitors are in vivo substrates and/or inhibitors of this ABC transporter. Gefitinib is an orally administered, active, selective epidermal growth factor receptor tyrosine kinase inhibitor (TKI) used to treat patients with advanced non-small-cell lung cancer (NSCLC),129,130 and ABCG2/BCRP is expressed in intestinal epithelial cells and at the blood–brain and blood–cerebrospinal barriers, where it restricts the penetration of the brain by xenobiotics.65,131 Stewart et al98 showed that the oral bioavailability of irinotecan, a good substrate of ABCG2/BCRP, is affected by the oral administration of gefitinib. Zhuang et al132 have also shown that the oral administration of gefitinib increases the penetration of topotecan across the brain extracellular fluid but, conversely, reduces its penetration of the ventricular cerebrospinal fluid.
Importantly, recent studies have tentatively implicated some ABCG2/BCRP polymorphisms in the pharmacokinetics of molecular-targeted drugs,103 as shown in Table 2. A common functional SNP in the ABCG2/BCRP gene, 421C>A, was shown to be associated with diarrhea in 124 patients treated with oral gefitinib (250 mg once daily); 44% of the patients with a heterozygous 421C>A allele developed diarrhea, whereas 12% of the other patients were homozygous for the wild-type allele. Other polymorphisms at the ABCG2/BCRP locus appear to affect gefitinib-induced diarrhea. A significant number of patients carrying the ABCG2 (–15622C/T) polymorphism and the ABCG2 (1143C/T, –15622C/T) haplotype developed gefitinib-dependent moderate-to-severe diarrhea.133 These studies suggest that patients with reduced ABCG2/BCRP activity arising from a genetic variation might be at increased risk of gefitinib-induced diarrhea, and these genetic markers should be considered in the optimization of NSCLC treatments with gefitinib. However, no clear association was reported between the 421C>A SNP of ABCG2/BCRP and a susceptibility to gefitinib-induced adverse effects in a Japanese population.134 The skin toxicity of gefitinib is also reported to be unrelated to this SNP.103 More large-scale analyses are required to resolve these discrepancies.
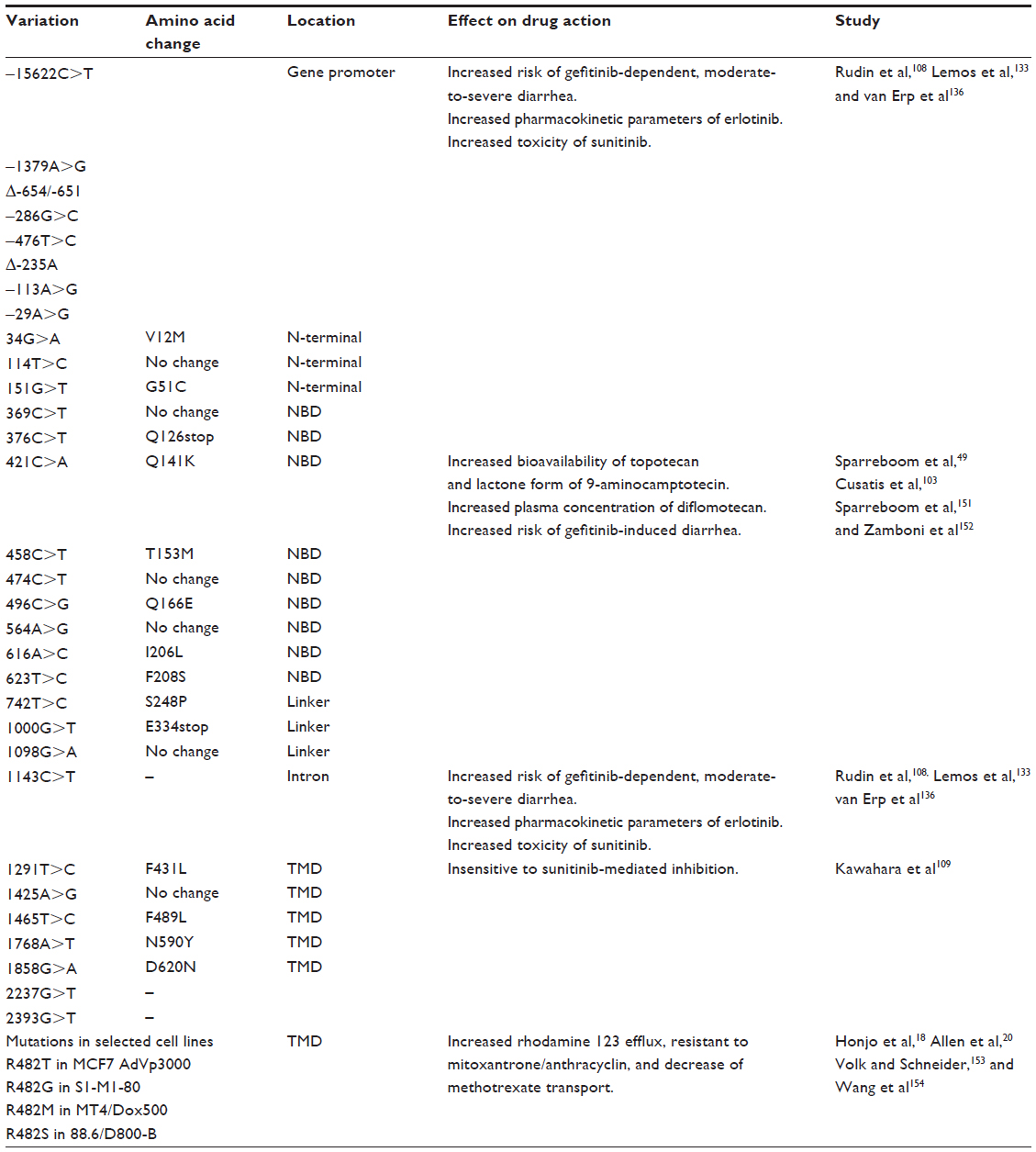 | Table 2 Single-nucleotide polymorphisms of the ABCG2/BCRP gene |
Like gefitinib, erlotinib has been shown to interact with ABCG2/BCRP.107,135 Two polymorphic loci identified in the ABCG2/BCRP promoter and intron, –15622C/T and 1143C/T, which reduce the protein’s expression, were reported to be associated with modulation of pharmacokinetic parameters for erlotinib.108 The area under the curve and maximum observed concentration (Cmax) were higher for the ABCG2 1143 C/T or T/T (lower expression) genotype, and Cmax was higher in patients with the –15622 C/T or T/T (lower expression) genotype than in those with the C/C genotype. These observations indicate that ABCG2/BCRP recognizes erlotinib as a substrate in vivo.
Sunitinib, an oral multitargeted TKI for vascular endothelial growth factor receptors 1, 2, and 3, platelet-derived growth factor receptor α and β, c-Kit, Fms-like tyrosine kinase 3 receptor, and the receptor encoded by the RET proto-oncogene, is used as a first-line treatment for metastatic renal cell carcinoma and imatinib-resistant metastatic gastrointestinal stromal tumors. The haplotype (–15622C/T, 1143C/T) at the ABCG2 locus that has been associated with gefitinib-associated adverse effects and increased erlotinib exposure, is also related to the development of increased sunitinib toxicity. The prevalence of toxicity higher than grade 2 increased when one or two copies of TT were present in the ABCG2 (–15622C/T, 1143C/T) haplotype.136 Another SNP in ABCG2/BCRP has also been shown to be associated with adverse effects similar to those attributed to sunitinib-related toxicity. Among 12 different genetic polymorphisms examined, the ABCG2/BCRP 421AA genotype correlated with the development of grade 3 or grade 4 thrombocytopenia and neutropenia in Korean patients suffering from metastatic renal cell carcinoma, and may be predominantly associated with the risk of sunitinib-related toxicity in those patients.137 Intriguingly, the 421C>A SNP of ABCG2/BCRP may have another effect on tumor lysis syndrome with hyperuricemia during TKI-based molecular-targeted therapy. Several studies have reported renal failure and tumor lysis syndrome during TKI-based molecular-targeted therapies, including with sorafenib for hepatocellular carcinoma, with imatinib for chronic myelogenous leukemia, and with flavopiridol for chronic lymphocytic leukemia.138–141 Because the Q141K variant of ABCG2/BCRP corresponding to SNP 421C>A reduces uric acid transport68 and those TKIs are substrates/inhibitors of ABCG2/BCRP, TKI therapy in patients with the SNP 421C>A (Q141K) may be at higher risk of tumor lysis syndrome.5
A defect in the pharmacological interaction has been suggested between sunitinib and another ABCG2/BCRP germ-line mutant allele, 1291T>C.109 Remarkably, sunitinib reversed wild-type ABCG2/BCRP-mediated drug resistance and competitively inhibited ABCG2/BCRP-mediated estrone 3-sulfate transport and the binding of 125I-iodoarylazidoprazosin to ABCG2/BCRP. The F431L variant of ABCG2/BCRP, which is expressed from a germ-line mutant allele 1291T>C, was insensitive to sunitinib-mediated inhibition. Thus, residue F431 of ABCG2/BCRP may have impact on the pharmacological interaction with sunitinib.
Perspectives
Molecular analyses of the functional interactions between novel molecular targeted drugs and the ABC transporter ABCG2/BCRP have demonstrable utility as indicators of the clinical efficacy of these anticancer agents in individual patients. Such pharmacological interactions might be influenced by personal genotypes, and the increased risk of adverse effects from putative ABCG2/BCRP substrates should be evaluated when considering combinations of protein kinase inhibitors, even at their clinically used doses.
Acknowledgments
Owing to space limitations, we apologize that we could not cite the excellent work of many investigators. This work was supported by a Grant-in-Aid for Cancer Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan.
Disclosure
The authors have no conflicts of interest to disclose.
References
Tsuruo T, Naito M, Tomida A, et al. Molecular targeting therapy of cancer: drug resistance, apoptosis and survival signal. Cancer Sci. 2003;94:15–21. | |
Ambudkar SV, Dey S, Hrycyna CA, Ramachandra M, Pastan I, Gottesman MM. Biochemical, cellular, and pharmacological aspects of the multidrug transporter. Annu Rev Pharmacol Toxicol. 1999;39:361–398. | |
Schinkel AH, Jonker JW. Mammalian drug efflux transporters of the ATP binding cassette (ABC) family: an overview. Adv Drug Deliv Rev. 2003;55:3–29. | |
Gottesman MM, Ling V. The molecular basis of multidrug resistance in cancer: the early years of P-glycoprotein research. FEBS Lett. 2006;580:998–1009. | |
Robey RW, Ierano C, Zhan Z, Bates SE. The challenge of exploiting ABCG2 in the clinic. Curr Pharm Biotechnol. 2010;12:595–608. | |
Allen JD, Schinkel AH. Multidrug resistance and pharmacological protection mediated by the breast cancer resistance protein (BCRP/ABCG2). Mol Cancer Ther. 2002;1:427–434. | |
Doyle LA, Yang W, Abruzzo LV, et al. A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc Natl Acad Sci U S A. 1998;95:15665–15670. | |
Allikmets R, Schriml LM, Hutchinson A, Romano-Spica V, Dean M. A human placenta-specific ATP-binding cassette gene (ABCP) on chromosome 4q22 that is involved in multidrug resistance. Cancer Res. 1998;58:5337–5339. | |
Brangi M, Litman T, Ciotti M, et al. Camptothecin resistance: role of the ATP-binding cassette (ABC), mitoxantrone-resistance half-transporter (MXR), and potential for glucuronidation in MXR-expressing cells. Cancer Res. 1999;59:5938–5946. | |
Maliepaard M, van Gastelen MA, de Jong LA, et al. Overexpression of the BCRP/MXR/ABCP gene in a topotecan-selected ovarian tumor cell line. Cancer Res. 1999;59:4559–4563. | |
Ross DD, Karp JE, Chen TT, Doyle LA. Expression of breast cancer resistance protein in blast cells from patients with acute leukemia. Blood. 2000;96:365–368. | |
Steinbach D, Sell W, Voigt A, Hermann J, Zintl F, Sauerbrey A. BCRP gene expression is associated with a poor response to remission induction therapy in childhood acute myeloid leukemia. Leukemia. 2002;16:1443–1447. | |
van den Heuvel-Eibrink MM, Wiemer EA, Prins A, et al. Increased expression of the breast cancer resistance protein (BCRP) in relapsed or refractory acute myeloid leukemia (AML). Leukemia. 2002;16:833–839. | |
Benderra Z, Faussat AM, Sayada L, et al. Breast cancer resistance protein and P-glycoprotein in 149 adult acute myeloid leukemias. Clin Cancer Res. 2004;10:7896–7902. | |
Plasschaert SL, Van Der Kolk DM, De Bont ES, Vellenga E, Kamps WA, De Vries EG. Breast cancer resistance protein (BCRP) in acute leukemia. Leuk Lymphoma. 2004;45:649–654. | |
Suvannasankha A, Minderman H, O’Loughlin KL, et al. Breast cancer resistance protein (BCRP/MXR/ABCG2) in acute myeloid leukemia: discordance between expression and function. Leukemia. 2004;18:1252–1257. | |
Uggla B, Stahl E, Wagsater D, et al. BCRP mRNA expression v. clinical outcome in 40 adult AML patients. Leuk Res. 2005;29:141–146. | |
Honjo Y, Hrycyna CA, Yan QW, et al. Acquired mutations in the MXR/BCRP/ABCP gene alter substrate specificity in MXR/BCRP/ABCP-overexpressing cells. Cancer Res. 2001;61:6635–6639. | |
Volk EL, Farley KM, Wu Y, Li F, Robey RW, Schneider E. Overexpression of wild-type breast cancer resistance protein mediates methotrexate resistance. Cancer Res. 2002;62:5035–5040. | |
Allen JD, Jackson SC, Schinkel AH. A mutation hot spot in the Bcrp1 (Abcg2) multidrug transporter in mouse cell lines selected for Doxorubicin resistance. Cancer Res. 2002;62:2294–2299. | |
Ozvegy C, Varadi A, Sarkadi B. Characterization of drug transport, ATP hydrolysis, and nucleotide trapping by the human ABCG2 multidrug transporter. Modulation of substrate specificity by a point mutation. J Biol Chem. 2002;277:47980–47990. | |
Chen ZS, Robey RW, Belinsky MG, et al. Transport of methotrexate, methotrexate polyglutamates, and 17beta-estradiol 17-(beta-D-glucuronide) by ABCG2: effects of acquired mutations at R482 on methotrexate transport. Cancer Res. 2003;63:4048–4054. | |
Ejendal KF, Diop NK, Schweiger LC, Hrycyna CA. The nature of amino acid 482 of human ABCG2 affects substrate transport and ATP hydrolysis but not substrate binding. Protein Sci. 2006;15:1597–1607. | |
Hazai E, Bikadi Z. Homology modeling of breast cancer resistance protein (ABCG2). J Struct Biol. 2008;162:63–74. | |
Li YF, Polgar O, Okada M, Esser L, Bates SE, Xia D. Towards understanding the mechanism of action of the multidrug resistance-linked half-ABC transporter ABCG2: a molecular modeling study. J Mol Graph Model. 2007;25:837–851. | |
Miwa M, Tsukahara S, Ishikawa E, Asada S, Imai Y, Sugimoto Y. Single amino acid substitutions in the transmembrane domains of breast cancer resistance protein (BCRP) alter cross resistance patterns in transfectants. Int J Cancer. 2003;107:757–763. | |
Rosenberg MF, Bikadi Z, Chan J, et al. The human breast cancer resistance protein (BCRP/ABCG2) shows conformational changes with mitoxantrone. Structure. 2010;18:482–493. | |
Cai X, Bikadi Z, Ni Z, et al. Role of basic residues within or near the predicted transmembrane helix 2 of the human breast cancer resistance protein in drug transport. J Pharmacol Exp Ther. 2010;333:670–681. | |
Kage K, Tsukahara S, Sugiyama T, et al. Dominant-negative inhibition of breast cancer resistance protein as drug efflux pump through the inhibition of S-S dependent homodimerization. Int J Cancer. 2002;97:626–630. | |
Doyle LA, Ross DD. Multidrug resistance mediated by the breast cancer resistance protein BCRP (ABCG2). Oncogene. 2003;22:7340–7358. | |
Henriksen U, Fog JU, Litman T, Gether U. Identification of intra- and intermolecular disulfide bridges in the multidrug resistance transporter ABCG2. J Biol Chem. 2005;280:36926–36934. | |
McDevitt CA, Collins RF, Conway M, et al. Purification and 3D structural analysis of oligomeric human multidrug transporter ABCG2. Structure. 2006;14:1623–1632. | |
Xie Y, Xu K, Linn DE, et al. The 44-kDa Pim-1 kinase phosphorylates BCRP/ABCG2 and thereby promotes its multimerization and drug-resistant activity in human prostate cancer cells. J Biol Chem. 2008;283:3349–3356. | |
Takada T, Suzuki H, Gotoh Y, Sugiyama Y. Regulation of the cell surface expression of human BCRP/ABCG2 by the phosphorylation state of Akt in polarized cells. Drug Metab Dispos. 2005;33:905–909. | |
Kondo C, Suzuki H, Itoda M, et al. Functional analysis of SNPs variants of BCRP/ABCG2. Pharm Res. 2004;21:1895–1903. | |
Yanase K, Tsukahara S, Mitsuhashi J, Sugimoto Y. Functional SNPs of the breast cancer resistance protein-therapeutic effects and inhibitor development. Cancer Lett. 2006;234:73–80. | |
Tamura A, Wakabayashi K, Onishi Y, et al. Re-evaluation and functional classification of non-synonymous single nucleotide polymorphisms of the human ATP-binding cassette transporter ABCG2. Cancer Sci. 2007;98:231–239. | |
Diop NK, Hrycyna CA. N-Linked glycosylation of the human ABC transporter ABCG2 on asparagine 596 is not essential for expression, transport activity, or trafficking to the plasma membrane. Biochemistry. 2005;44:5420–5429. | |
Wakabayashi-Nakao K, Tamura A, Furukawa T, Nakagawa H, Ishikawa T. Quality control of human ABCG2 protein in the endoplasmic reticulum: ubiquitination and proteasomal degradation. Adv Drug Deliv Rev. 2009;61:66–72. | |
Wakabayashi K, Nakagawa H, Tamura A, et al. Intramolecular disulfide bond is a critical check point determining degradative fates of ATP-binding cassette (ABC) transporter ABCG2 protein. J Biol Chem. 2007;282:27841–27846. | |
Honjo Y, Morisaki K, Huff LM, et al. Single-nucleotide polymorphism (SNP) analysis in the ABC half-transporter ABCG2 (MXR/BCRP/ABCP1). Cancer Biol Ther. 2002;1:696–702. | |
Sugimoto Y, Tsukahara S, Ishikawa E, Mitsuhashi J. Breast cancer resistance protein: molecular target for anticancer drug resistance and pharmacokinetics/pharmacodynamics. Cancer Sci. 2005;96:457–465. | |
Cascorbi I. Role of pharmacogenetics of ATP-binding cassette transporters in the pharmacokinetics of drugs. Pharmacol Ther. 2006;112:457–473. | |
Imai Y, Nakane M, Kage K, et al. C421A polymorphism in the human breast cancer resistance protein gene is associated with low expression of Q141K protein and low-level drug resistance. Mol Cancer Ther. 2002;1:611–616. | |
Yoshioka S, Katayama K, Okawa C, et al. The identification of two germ-line mutations in the human breast cancer resistance protein gene that result in the expression of a low/non-functional protein. Pharm Res. 2007;24:1108–1117. | |
Itoda M, Saito Y, Shirao K, et al. Eight novel single nucleotide polymorphisms in ABCG2/BCRP in Japanese cancer patients administered irinotacan. Drug Metab Pharmacokinet. 2003;18:212–217. | |
Kobayashi D, Ieiri I, Hirota T, et al. Functional assessment of ABCG2 (BCRP) gene polymorphisms to protein expression in human placenta. Drug Metab Dispos. 2005;33:94–101. | |
de Jong FA, Marsh S, Mathijssen RH, et al. ABCG2 pharmacogenetics: ethnic differences in allele frequency and assessment of influence on irinotecan disposition. Clin Cancer Res. 2004;10:5889–5894. | |
Sparreboom A, Gelderblom H, Marsh S, et al. Diflomotecan pharmacokinetics in relation to ABCG2 421C>A genotype. Clin Pharmacol Ther. 2004;76:38–44. | |
Mizuarai S, Aozasa N, Kotani H. Single nucleotide polymorphisms result in impaired membrane localization and reduced atpase activity in multidrug transporter ABCG2. Int J Cancer. 2004;109:238–246. | |
Nakagawa H, Tamura A, Wakabayashi K, et al. Ubiquitin-mediated proteasomal degradation of non-synonymous SNP variants of human ABC transporter ABCG2. Biochem J. 2008;411:623–631. | |
Zamber CP, Lamba JK, Yasuda K, et al. Natural allelic variants of breast cancer resistance protein (BCRP) and their relationship to BCRP expression in human intestine. Pharmacogenetics. 2003;13:19–28. | |
Backstrom G, Taipalensuu J, Melhus H, et al. Genetic variation in the ATP-binding cassette transporter gene ABCG2 (BCRP) in a Swedish population. Eur J Pharm Sci. 2003;18:359–364. | |
Poonkuzhali B, Lamba J, Strom S, et al. Association of breast cancer resistance protein/ABCG2 phenotypes and novel promoter and intron 1 single nucleotide polymorphisms. Drug Metab Dispos. 2008;36:780–795. | |
Wang F, Liang YJ, Wu XP, et al. Prognostic value of the multidrug resistance transporter ABCG2 gene polymorphisms in Chinese patients with de novo acute leukaemia. Eur J Cancer. 2011;47:1990–1999. | |
Nakanishi T, Bailey-Dell KJ, Hassel BA, et al. Novel 5′ untranslated region variants of BCRP mRNA are differentially expressed in drug-selected cancer cells and in normal human tissues: implications for drug resistance, tissue-specific expression, and alternative promoter usage. Cancer Res. 2006;66:5007–5011. | |
Maliepaard M, Scheffer GL, Faneyte IF, et al. Subcellular localization and distribution of the breast cancer resistance protein transporter in normal human tissues. Cancer Res. 2001;61:3458–3464. | |
Aronica E, Gorter JA, Redeker S, et al. Localization of breast cancer resistance protein (BCRP) in microvessel endothelium of human control and epileptic brain. Epilepsia. 2005;46:849–857. | |
Fetsch PA, Abati A, Litman T, et al. Localization of the ABCG2 mitoxantrone resistance-associated protein in normal tissues. Cancer Lett. 2006;235:84–92. | |
Marchetti S, de Vries NA, Buckle T, et al. Effect of the ATP-binding cassette drug transporters ABCB1, ABCG2, and ABCC2 on erlotinib hydrochloride (Tarceva) disposition in in vitro and in vivo pharmacokinetic studies employing Bcrp1-/-/Mdr1a/1b-/- (triple-knockout) and wild-type mice. Mol Cancer Ther. 2008;7:2280–2287. | |
Furman WL, Navid F, Daw NC, et al. Tyrosine kinase inhibitor enhances the bioavailability of oral irinotecan in pediatric patients with refractory solid tumors. J Clin Oncol. 2009;27:4599–4604. | |
Ding XW, Wu JH, Jiang CP. ABCG2: a potential marker of stem cells and novel target in stem cell and cancer therapy. Life Sci; 2010;86:631–637. | |
Zhou S, Morris JJ, Barnes Y, Lan L, Schuetz JD, Sorrentino BP. Bcrp1 gene expression is required for normal numbers of side population stem cells in mice, and confers relative protection to mitoxantrone in hematopoietic cells in vivo. Proc Natl Acad Sci U S A. 2002;99:12339–12344. | |
Katayama R, Koike S, Sato S, Sugimoto Y, Tsuruo T, Fujita N. Dofequidar fumarate sensitizes cancer stem-like side population cells to chemotherapeutic drugs by inhibiting ABCG2/BCRP-mediated drug export. Cancer Sci. 2009;100:2060–2068. | |
Sarkadi B, Homolya L, Szakacs G, Varadi A. Human multidrug resistance ABCB and ABCG transporters: participation in a chemoimmunity defense system. Physiol Rev. 2006;86:1179–1236. | |
Jonker JW, Buitelaar M, Wagenaar E, et al. The breast cancer resistance protein protects against a major chlorophyll-derived dietary phototoxin and protoporphyria. Proc Natl Acad Sci U S A. 2002;99:15649–15654. | |
Krishnamurthy P, Ross DD, Nakanishi T, et al. The stem cell marker Bcrp/ABCG2 enhances hypoxic cell survival through interactions with heme. J Biol Chem. 2004;279:24218–24225. | |
Woodward OM, Kottgen A, Coresh J, Boerwinkle E, Guggino WB, Kottgen M. Identification of a urate transporter, ABCG2, with a common functional polymorphism causing gout. Proc Natl Acad Sci U S A. 2009;106:10338–10342. | |
Matsuo H, Takada T, Ichida K, et al. Common defects of ABCG2, a high-capacity urate exporter, cause gout: a function-based genetic analysis in a Japanese population. Sci Transl Med. 2009;1:5ra11. | |
Suzuki M, Suzuki H, Sugimoto Y, Sugiyama Y. ABCG2 transports sulfated conjugates of steroids and xenobiotics. J Biol Chem. 2003;278:22644–22649. | |
Nakatomi K, Yoshikawa M, Oka M, et al. Transport of 7-ethyl-10-hydroxycamptothecin (SN-38) by breast cancer resistance protein ABCG2 in human lung cancer cells. Biochem Biophys Res Commun. 2001;288:827–832. | |
van Herwaarden AE, Jonker JW, Wagenaar E, et al. The breast cancer resistance protein (Bcrp1/Abcg2) restricts exposure to the dietary carcinogen 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine. Cancer Res. 2003;63:6447–6452. | |
Hirano M, Maeda K, Matsushima S, Nozaki Y, Kusuhara H, Sugiyama Y. Involvement of BCRP (ABCG2) in the biliary excretion of pitavastatin. Mol Pharmacol. 2005;68:800–807. | |
van Herwaarden AE, Schinkel AH. The function of breast cancer resistance protein in epithelial barriers, stem cells and milk secretion of drugs and xenotoxins. Trends Pharmacol Sci. 2006;27:10–16. | |
van Herwaarden AE, Wagenaar E, Karnekamp B, Merino G, Jonker JW, Schinkel AH. Breast cancer resistance protein (Bcrp1/Abcg2) reduces systemic exposure of the dietary carcinogens aflatoxin B1, IQ and Trp-P-1 but also mediates their secretion into breast milk. Carcinogenesis. 2006;27:123–130. | |
Sawyers C. Targeted cancer therapy. Nature. 2004;432:294–297. | |
Krause DS, Van Etten RA. Tyrosine kinases as targets for cancer therapy. N Engl J Med. 2005;353:172–187. | |
Capdeville R, Buchdunger E, Zimmermann J, Matter A. Glivec (STI571, imatinib), a rationally developed, targeted anticancer drug. Nat Rev Drug Discov. 2002;1:493–502. | |
Gorre ME, Mohammed M, Ellwood K, et al. Clinical resistance to STI-571 cancer therapy caused by BCR-ABL gene mutation or amplification. Science. 2001;293:876–880. | |
Shannon KM. Resistance in the land of molecular cancer therapeutics. Cancer Cell. 2002;2:99–102. | |
Azam M, Latek RR, Daley GQ. Mechanisms of autoinhibition and STI-571/imatinib resistance revealed by mutagenesis of BCR-ABL. Cell. 2003;112:831–843. | |
Daub H, Specht K, Ullrich A. Strategies to overcome resistance to targeted protein kinase inhibitors. Nat Rev Drug Discov. 2004;3:1001–1010. | |
Mahon FX, Belloc F, Lagarde V, et al. MDR1 gene overexpression confers resistance to imatinib mesylate in leukemia cell line models. Blood. 2003;101:2368–2373. | |
Illmer T, Schaich M, Platzbecker U, et al. P-glycoprotein-mediated drug efflux is a resistance mechanism of chronic myelogenous leukemia cells to treatment with imatinib mesylate. Leukemia. 2004;18:401–408. | |
Hegedus T, Orfi L, Seprodi A, Varadi A, Sarkadi B, Keri G. Interaction of tyrosine kinase inhibitors with the human multidrug transporter proteins, MDR1 and MRP1. Biochim Biophys Acta. 2002;1587:318–325. | |
Burger H, Nooter K. Pharmacokinetic resistance to imatinib mesylate: role of the ABC drug pumps ABCG2 (BCRP) and ABCB1 (MDR1) in the oral bioavailability of imatinib. Cell Cycle. 2004;3:1502–1505. | |
Ozvegy-Laczka C, Hegedus T, Varady G, et al. High-affinity interaction of tyrosine kinase inhibitors with the ABCG2 multidrug transporter. Mol Pharmacol. 2004;65:1485–1495. | |
Houghton PJ, Germain GS, Harwood FC, et al. Imatinib mesylate is a potent inhibitor of the ABCG2 (BCRP) transporter and reverses resistance to topotecan and SN-38 in vitro. Cancer Res. 2004;64:2333–2337. | |
Breedveld P, Pluim D, Cipriani G, et al. The effect of Bcrp1 (Abcg2) on the in vivo pharmacokinetics and brain penetration of imatinib mesylate (Gleevec): implications for the use of breast cancer resistance protein and P-glycoprotein inhibitors to enable the brain penetration of imatinib in patients. Cancer Res. 2005;65:2577–2582. | |
Lemos C, Jansen G, Peters GJ. Drug transporters: recent advances concerning BCRP and tyrosine kinase inhibitors. Br J Cancer. 2008;98:857–862. | |
Burger H, van Tol H, Boersma AW, et al. Imatinib mesylate (STI571) is a substrate for the breast cancer resistance protein (BCRP)/ABCG2 drug pump. Blood. 2004;104:2940–2942. | |
Thomas J, Wang L, Clark RE, Pirmohamed M. Active transport of imatinib into and out of cells: implications for drug resistance. Blood. 2004;104:3739–3745. | |
Jordanides NE, Jorgensen HG, Holyoake TL, Mountford JC. Functional ABCG2 is overexpressed on primary CML CD34+ cells and is inhibited by imatinib mesylate. Blood. 2006;108:1370–1373. | |
Liu W, Baer MR, Bowman MJ, et al. The tyrosine kinase inhibitor imatinib mesylate enhances the efficacy of photodynamic therapy by inhibiting ABCG2. Clin Cancer Res. 2007;13:2463–2470. | |
Brendel C, Scharenberg C, Dohse M, et al. Imatinib mesylate and nilotinib (AMN107) exhibit high-affinity interaction with ABCG2 on primitive hematopoietic stem cells. Leukemia. 2007;21:1267–1275. | |
Hiwase DK, Saunders V, Hewett D, et al. Dasatinib cellular uptake and efflux in chronic myeloid leukemia cells: therapeutic implications. Clin Cancer Res. 2008;14:3881–3888. | |
Polli JW, Humphreys JE, Harmon KA, et al. The role of efflux and uptake transporters in N-{3-chloro-4-[(3-fluorobenzyl)oxy]phenyl}-6-[5-({[2-(methylsulfonyl)ethyl]amino} methyl)-2-furyl]-4-quinazolinamine (GW572016, lapatinib) disposition and drug interactions. Drug Metab Dispos. 2008;36:695–701. | |
Stewart CF, Leggas M, Schuetz JD, et al. Gefitinib enhances the antitumor activity and oral bioavailability of irinotecan in mice. Cancer Res. 2004;64:7491–7499. | |
Yanase K, Tsukahara S, Asada S, Ishikawa E, Imai Y, Sugimoto Y. Gefitinib reverses breast cancer resistance protein-mediated drug resistance. Mol Cancer Ther. 2004;3:1119–1125. | |
Carter TA, Wodicka LM, Shah NP, et al. Inhibition of drug-resistant mutants of ABL, KIT, and EGF receptor kinases. Proc Natl Acad Sci U S A. 2005;102:11011–11016. | |
Elkind NB, Szentpetery Z, Apati A, et al. Multidrug transporter ABCG2 prevents tumor cell death induced by the epidermal growth factor receptor inhibitor Iressa (ZD1839, Gefitinib). Cancer Res. 2005;65:1770–1777. | |
Nakamura Y, Oka M, Soda H, et al. Gefitinib (“Iressa”, ZD1839), an epidermal growth factor receptor tyrosine kinase inhibitor, reverses breast cancer resistance protein/ABCG2-mediated drug resistance. Cancer Res. 2005;65:1541–1546. | |
Cusatis G, Gregorc V, Li J, et al. Pharmacogenetics of ABCG2 and ad-verse reactions to gefitinib. J Natl Cancer Inst. 2006;98:1739–1742. | |
Leggas M, Panetta JC, Zhuang Y, et al. Gefitinib modulates the function of multiple ATP-binding cassette transporters in vivo. Cancer Res. 2006;66:4802–4807. | |
Katayama K, Shibata K, Mitsuhashi J, Noguchi K, Sugimoto Y. Pharmacological interplay between breast cancer resistance protein and gefitinib in epidermal growth factor receptor signaling. Anticancer Res. 2009;29:1059–1065. | |
Erlichman C, Boerner SA, Hallgren CG, et al. The HER tyrosine kinase inhibitor CI1033 enhances cytotoxicity of 7-ethyl-10-hydroxycamptothecin and topotecan by inhibiting breast cancer resistance protein-mediated drug efflux. Cancer Res. 2001;61:739–748. | |
Shi Z, Peng XX, Kim IW, et al. Erlotinib (Tarceva, OSI-774) antagonizes ATP-binding cassette subfamily B member 1 and ATP-binding cassette subfamily G member 2-mediated drug resistance. Cancer Res. 2007;67:11012–11020. | |
Rudin CM, Liu W, Desai A, et al. Pharmacogenomic and pharmacokinetic determinants of erlotinib toxicity. J Clin Oncol. 2008;26:1119–1127. | |
Kawahara H, Noguchi K, Katayama K, Mitsuhashi J, Sugimoto Y. Pharmacological interaction with sunitinib is abolished by a germ-line mutation (1291T>C) of BCRP/ABCG2 gene. Cancer Sci. 2010;101:1493–1500. | |
Hu S, Chen Z, Franke R, et al. Interaction of the multikinase inhibitors sorafenib and sunitinib with solute carriers and ATP-binding cassette transporters. Clin Cancer Res. 2009;15:6062–6069. | |
Minocha M, Khurana V, Qin B, Pal D, Mitra AK. Enhanced brain accumulation of pazopanib by modulating P-gp and Bcrp1 mediated efflux with canertinib or erlotinib. Int J Pharm. 2012;436:127–134. | |
Hegedus C, Truta-Feles K, Antalffy G, et al. Interaction of the EGFR inhibitors gefitinib, vandetanib, pelitinib and neratinib with the ABCG2 multidrug transporter: implications for the emergence and reversal of cancer drug resistance. Biochem Pharmacol. 2012;84:260–267. | |
Mittapalli RK, Vaidhyanathan S, Sane R, Elmquist WF. Impact of P-glycoprotein (ABCB1) and breast cancer resistance protein (ABCG2) on the brain distribution of a novel BRAF inhibitor: vemurafenib (PLX4032). J Pharmacol Exp Ther. 2012;342:33–40. | |
Poller B, Iusuf D, Sparidans RW, Wagenaar E, Beijnen JH, Schinkel AH. Differential impact of P-glycoprotein (ABCB1) and breast cancer resistance protein (ABCG2) on axitinib brain accumulation and oral plasma pharmacokinetics. Drug Metab Dispos. 2011;39:729–735. | |
Sen R, Natarajan K, Bhullar J, et al. The novel BCR-ABL and FLT3 inhibitor ponatinib is a potent inhibitor of the MDR-associated ATP-binding cassette transporter ABCG2. Mol Cancer Ther. 2012;11:2033–2044. | |
Hegedus C, Ozvegy-Laczka C, Apati A, et al. Interaction of nilotinib, dasatinib and bosutinib with ABCB1 and ABCG2: implications for altered anti-cancer effects and pharmacological properties. Br J Pharmacol. 2009;158:1153–1164. | |
Tang SC, Nguyen LN, Sparidans RW, Wagenaar E, Beijnen JH, Schinkel AH. Increased oral availability and brain accumulation of the ALK inhibitor crizotinib by coadministration of the P-glycoprotein (ABCB1) and breast cancer resistance protein (ABCG2) inhibitor elacridar. Int J Cancer. 2014;134:1484–1494. | |
Hooijberg JH, Broxterman HJ, Scheffer GL, et al. Potent interaction of flavopiridol with MRP1. Br J Cancer. 1999;81:269–276. | |
Robey RW, Medina-Perez WY, Nishiyama K, et al. Overexpression of the ATP-binding cassette half-transporter, ABCG2 (Mxr/BCrp/ABCP1), in flavopiridol-resistant human breast cancer cells. Clin Cancer Res. 2001;7:145–152. | |
Boerner SA, Tourne ME, Kaufmann SH, Bible KC. Effect of P-glycoprotein on flavopiridol sensitivity. Br J Cancer. 2001;84:1391–1396. | |
Jager W, Gehring E, Hagenauer B, Aust S, Senderowicz A, Thalhammer T. The role of hepatic Mrp2 in the interaction of flavopiridol and bilirubin: impact on therapy. Int J Clin Pharmacol Ther. 2003;41:610–611. | |
Seamon JA, Rugg CA, Emanuel S, et al. Role of the ABCG2 drug transporter in the resistance and oral bioavailability of a potent cyclin-dependent kinase/Aurora kinase inhibitor. Mol Cancer Ther. 2006;5:2459–2467. | |
Yokota A, Kimura S, Masuda S, et al. INNO-406, a novel BCR-ABL/Lyn dual tyrosine kinase inhibitor, suppresses the growth of Ph+ leukemia cells in the central nervous system, and cyclosporine A augments its in vivo activity. Blood. 2007;109:306–314. | |
Robey RW, Shukla S, Steadman K, et al. Inhibition of ABCG2-mediated transport by protein kinase inhibitors with a bisindolylmaleimide or indolocarbazole structure. Mol Cancer Ther. 2007;6:1877–1885. | |
Noble ME, Endicott JA, Johnson LN. Protein kinase inhibitors: insights into drug design from structure. Science. 2004;303:1800–1805. | |
Saito H, Hirano H, Nakagawa H, et al. A new strategy of high-speed screening and quantitative structure-activity relationship analysis to evaluate human ATP-binding cassette transporter ABCG2-drug interactions. J Pharmacol Exp Ther. 2006;317:1114–1124. | |
Ejendal KF, Hrycyna CA. Differential sensitivities of the human ATP-binding cassette transporters ABCG2 and P-glycoprotein to cyclosporin A. Mol Pharmacol. 2005;67:902–911. | |
Clark R, Kerr ID, Callaghan R. Multiple drugbinding sites on the R482G isoform of the ABCG2 transporter. Br J Pharmacol. 2006;149:506–515. | |
Herbst RS, Fukuoka M, Baselga J. Gefitinib – a novel targeted approach to treating cancer. Nat Rev Cancer. 2004;4:956–965. | |
Cappuzzo F, Ligorio C, Janne PA, et al. Prospective study of gefitinib in epidermal growth factor receptor fluorescence in situ hybridization-positive/phospho-Akt-positive or never smoker patients with advanced non-small-cell lung cancer: the ONCOBELL trial. J Clin Oncol. 2007;25:2248–2255. | |
Robey RW, Polgar O, Deeken J, To KW, Bates SE. ABCG2: determining its relevance in clinical drug resistance. Cancer Metastasis Rev. 2007;26:39–57. | |
Zhuang Y, Fraga CH, Hubbard KE, et al. Topotecan central nervous system penetration is altered by a tyrosine kinase inhibitor. Cancer Res. 2006;66:11305–11313. | |
Lemos C, Giovannetti E, Zucali PA, et al. Impact of ABCG2 polymorphisms on the clinical outcome and toxicity of gefitinib in non-small-cell lung cancer patients. Pharmacogenomics. 2011;12:159–170. | |
Akasaka K, Kaburagi T, Yasuda S, et al. Impact of functional ABCG2 polymorphisms on the adverse effects of gefitinib in Japanese patients with non-small-cell lung cancer. Cancer Chemother Pharmacol. 2009;66:691–698. | |
Li J, Cusatis G, Brahmer J, et al. Association of variant ABCG2 and the pharmacokinetics of epidermal growth factor receptor tyrosine kinase inhibitors in cancer patients. Cancer Biol Ther. 2007;6:432–438. | |
van Erp NP, Eechoute K, van der Veldt AA, et al. Pharmacogenetic pathway analysis for determination of sunitinib-induced toxicity. J Clin Oncol. 2009;27:4406–4412. | |
Kim HR, Park HS, Kwon WS, et al. Pharmacogenetic determinants associated with sunitinib-induced toxicity and ethnic difference in Korean metastatic renal cell carcinoma patients. Cancer Chemother Pharmacol. 2013;72:825–835. | |
Gafter-Gvili A, Ram R, Gafter U, Shpilberg O, Raanani P. Renal failure associated with tyrosine kinase inhibitors – case report and review of the literature. Leuk Res. 2009;34:123–127. | |
Lin TS, Ruppert AS, Johnson AJ, et al. Phase II study of flavopiridol in relapsed chronic lymphocytic leukemia demonstrating high response rates in genetically high-risk disease. J Clin Oncol. 2009;27:6012–6018. | |
Huang WS, Yang CH. Sorafenib induced tumor lysis syndrome in an advanced hepatocellular carcinoma patient. World J Gastroenterol. 2009;15:4464–4466. | |
Al-Kali A, Farooq S, Tfayli A. Tumor lysis syndrome after starting treatment with Gleevec in a patient with chronic myelogenous leukemia. J Clin Pharm Ther. 2009;34:607–610. | |
Dohse M, Scharenberg C, Shukla S, et al. Comparison of ATP-binding cassette transporter interactions with the tyrosine kinase inhibitors imatinib, nilotinib, and dasatinib. Drug Metab Dispos. 2010;38:1371–1380. | |
Dai CL, Tiwari AK, Wu CP, et al. Lapatinib (Tykerb, GW572016) reverses multidrug resistance in cancer cells by inhibiting the activity of ATP-binding cassette subfamily B member 1 and G member 2. Cancer Res. 2008;68:7905–7914. | |
Shukla S, Robey RW, Bates SE, Ambudkar SV. Sunitinib (Sutent, SU11248), a small-molecule receptor tyrosine kinase inhibitor, blocks function of the ATP-binding cassette (ABC) transporters P-glycoprotein (ABCB1) and ABCG2. Drug Metab Dispos. 2009;37:359–365. | |
Zheng LS, Wang F, Li YH, et al. Vandetanib (Zactima, ZD6474) antagonizes ABCC1- and ABCG2-mediated multidrug resistance by inhibition of their transport function. PLoS One. 2009;4:e5172. | |
Jovelet C, Benard J, Forestier F, Farinotti R, Bidart JM, Gil S. Inhibition of P-glycoprotein functionality by vandetanib may reverse cancer cell resistance to doxorubicin. Eur J Pharm Sci. 2012;46:484–491. | |
Reyner EL, Sevidal S, West MA, et al. In vitro characterization of axitinib interactions with human efflux and hepatic uptake transporters: implications for disposition and drug interactions. Drug Metab Dispos. 2013;41:1575–1583. | |
Lagas JS, van Waterschoot RA, Sparidans RW, Wagenaar E, Beijnen JH, Schinkel AH. Breast cancer resistance protein and P-glycoprotein limit sorafenib brain accumulation. Mol Cancer Ther. 2010;9:319–326. | |
Durmus S, Sparidans RW, Wagenaar E, Beijnen JH, Schinkel AH. Oral availability and brain penetration of the B-RAFV600E inhibitor vemurafenib can be enhanced by the P-GLYCOprotein (ABCB1) and breast cancer resistance protein (ABCG2) inhibitor elacridar. Mol Pharm. 2012;9:3236–3245. | |
Wu CP, Sim HM, Huang YH, et al. Overexpression of ATP-binding cassette transporter ABCG2 as a potential mechanism of acquired resistance to vemurafenib in BRAF(V600E) mutant cancer cells. Biochem Pharmacol. 2012;85:325–334. | |
Sparreboom A, Loos WJ, Burger H, et al. Effect of ABCG2 genotype on the oral bioavailability of topotecan. Cancer Biol Ther. 2005;4:650–658. | |
Zamboni WC, Ramanathan RK, McLeod HL, et al. Disposition of 9-nitrocamptothecin and its 9-aminocamptothecin metabolite in relation to ABC transporter genotypes. Invest New Drugs. 2006;24:393–401. | |
Volk EL, Schneider E. Wild-type breast cancer resistance protein (BCRP/ABCG2) is a methotrexate polyglutamate transporter. Cancer Res. 2003;63:5538–5543. | |
Wang X, Furukawa T, Nitanda T, et al. Breast cancer resistance protein (BCRP/ABCG2) induces cellular resistance to HIV-1 nucleoside reverse transcriptase inhibitors. Mol Pharmacol. 2003;63:65–72. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.