Back to Journals » OncoTargets and Therapy » Volume 10
HSD17B12 gene rs11037575 C>T polymorphism confers neuroblastoma susceptibility in a Southern Chinese population
Authors Zhang ZR, Zou Y, Zhu JH ![]() , Zhang RZ, Yang TY, Wang FH, Xia HM
, Zhang RZ, Yang TY, Wang FH, Xia HM ![]() , He J
, He J ![]() , Feng ZC
, Feng ZC
Received 2 March 2017
Accepted for publication 23 March 2017
Published 4 April 2017 Volume 2017:10 Pages 1969—1975
DOI https://doi.org/10.2147/OTT.S136006
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr William C. Cho
Zhuorong Zhang,1,2 Yan Zou,2 Jinhong Zhu,3 Ruizhong Zhang,2 Tianyou Yang,2 Fenghua Wang,2 Huimin Xia,1,2 Jing He,2 Zhichun Feng1,4–6
1Southern Medical University, Guangzhou, Guangdong, 2Department of Pediatric Surgery, Guangzhou Institute of Pediatrics, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, Guangzhou, Guangdong, 3Molecular Epidemiology Laboratory, Department of Laboratory Medicine, Harbin Medical University Cancer Hospital, Harbin, Heilongjiang, 4Division of Neonatology, Affiliated BaYi Children’s Hospital, Clinical Medical College in PLA Army General Hospital, Southern Medical University, 5National Engineering Laboratory for Birth Defects Prevention and Control of Key Technology, 6Beijing Key Laboratory of Pediatric Organ Failure, Beijing, People’s Republic of China
Abstract: A previous genome-wide association study (GWAS) identified four genetic polymorphisms (rs1027702 near DUSP12, rs10055201 in IL31RA, rs2619046 in DDX4, and rs11037575 in HSD17B12 gene) that were associated with neuroblastoma susceptibility, especially for low-risk subjects. The aim of this study was to examine the association between these four polymorphisms and neuroblastoma susceptibility in a Southern Chinese population composed of 256 cases and 531 controls. Overall, among all the polymorphisms, single-locus analysis only revealed significant association between the HSD17B12 rs11037575 C>T polymorphism and neuroblastoma susceptibility (CT vs CC: adjusted odds ratio [OR] =0.71, 95% confidence interval [CI] =0.51–0.97, P=0.030). Moreover, stratified analysis indicated that the rs11037575 T allele was associated with decreased neuroblastoma risk among the children aged 0–18 months (adjusted OR =0.60, 95% CI =0.37–0.97, P=0.036); regarding the tumor site, this polymorphism protected against tumor in the mediastinum (adjusted OR =0.59, 95% CI =0.37–0.94, P=0.025). When risk genotypes were combined, we found that girls with two to four risk genotypes were at a significantly increased risk of neuroblastoma (adjusted OR =1.65, 95% CI =1.03–2.64, P=0.039). In terms of clinical stages, individuals with two to four risk genotypes had a tendency toward the development of stage III/IV diseases (adjusted OR =1.69, 95% CI =1.12–2.54, P=0.012). In conclusion, we verified that the HSD17B12 rs11037575 T allele might negatively associate with neuroblastoma risk. These findings need further validation by prospective studies with larger sample size and different ethnicities.
Keywords: GWAS, HSD17B12, polymorphism, neuroblastoma, susceptibility
Introduction
Neuroblastoma is a pediatric cancer originating from the developing sympathetic nervous system. It is the most frequent peripheral nervous system tumor of infancy and childhood,1,2 which constitutes of approximately 7%–10% of all childhood cancers.3 Neuroblastoma is the third leading cause of cancer-related death in children.3 Despite of remarkable advances in the treatments of many childhood cancers, neuroblastoma remains a serious clinical problem, causing 15% of childhood cancer mortality.4 The median age at diagnosis of this disease is around 17 months.5 Neuroblastoma may occur throughout the sympathetic nervous system, mostly within the abdomen and adrenal medulla.6 The incidence rate of neuroblastoma is about 1 in 7000 live newborns worldwide.7 It is also one of the most commonly diagnosed solid tumors in the Chinese infants, with an incidence rate of ~7.7 per million.8 The majority of neuroblastomas are sporadic, and only about 1% of patients have a family history of this type of disease.9 Thus far, the etiology of neuroblastoma remains largely unidentified, especially for predisposing factors for neuroblastoma.10,11 Family studies and case–control studies have been often adopted to discover neuroblastoma susceptibility genetic variants.12–15 For instance, Diskin et al16 conducted a case–control study with 2,817 neuroblastoma cases and 7,473 controls. They recognized two loci at 6q16 region associated with the neuroblastoma susceptibility, one within the HACE1 (rs4336470), and the other within the LIN28B (rs17065417) gene.
Over the past years, at least five genome-wide association studies (GWASs) have been accomplished mainly in North American patients of European descent, and several loci were proven to be related with the risk of neuroblastoma, such as CASC15 in 2008, BARD1 in 2009, and LMO1 in 2011.11,16–19 A two-stage neuroblastoma GWAS by Nguyen et al18 included 1,627 cases and 2,575 controls at the first stage, as well as 398 cases and 1,507 controls in the replication stage. They found that dual-specificity phosphatase 12 (DUSP12) gene polymorphisms, at chromosome band 1q23.3, significantly conferred neuroblastoma susceptibility. When they limited the analysis to 574 low-risk neuroblastoma cases and 1,722 controls, DUSP12 and three novel genes were validated to be associated with low-risk neuroblastoma. They were IL31RA and DDX4 located on chromosome band 5q11.2 and HSD17B12 on chromosome band 11p11.2. Among all the significant polymorphisms in the four genes, the rs1027702 T>C, rs10055201 A>G, rs2619046 G>A, and rs11037575 C>T are most noteworthy. The association between these polymorphisms and neuroblastoma susceptibility has been replicated in the Italians20 and a Northern Chinese population.15 To scrutinize the association between the four most significant polymorphisms and neuroblastoma susceptibility in Southern Chinese population, we conducted this study including 256 neuroblastoma cases and 531 cancer-free controls.
Materials and methods
Study subjects
To investigate the association between chosen genetic polymorphisms and the risk of neuroblastoma, we included 256 neuroblastoma cases having received treatments from the Guangzhou Women and Children’s Medical Center as reported previously,21–25 and 531 age-, gender-, and race-matched cancer-free controls were randomly picked from children who visited the same hospital for a routine physical examination.26–28 This study received the approval of the Institutional Review Board of Guangzhou Women and Children’s Medical Center. Written informed consent was acquired from each participant or his/her guardian.
Polymorphism analysis
DNA samples were processed as we described elsewhere.29–32 Briefly, DNA samples were diluted to a stock concentration of 10 ng/μL and added to the 96-well plates. Genotyping for the four GWAS-identified gene single-nucleotide polymorphisms (SNPs) (rs1027702 T>C, rs10055201 A>G, rs2619046 G>A, and rs11037575 C>T)20 was carried out in the 384-well plate using Taqman method following a published protocol.29 Moreover, for the purpose of quality control and validation of the accuracy of genotyping results, ~10% of the samples were randomly selected for sequencing. The results were 100% concordant.
Statistical analysis
Differences in the demographics and genotypes between neuroblastoma cases and controls were compared by χ2 test. Hardy–Weinberg equilibrium for control subjects was calculated by goodness-of-fit χ2 test. Unconditional univariate and multivariate logistic regression analyses were performed. Odds ratios (ORs) and 95% confidence intervals (CIs), with adjustment for age and gender, were used to quantify the strength of associations between these four polymorphisms and neuroblastoma susceptibility. Stratified analysis was performed regarding age, gender, tumor sites, and clinical stages. A P<0.05 was considered as statistically significant.33 All statistical tests were two-sided, and were calculated using SAS software (Version 9.1; SAS Institute, Cary, NC, USA).
Results
Population characteristics
The demographic characteristics of participants are reviewed in Table S1. No statistically significant differences were detected between cases and controls with respect to age (P=0.239) and gender (P=0.333). Based on International Neuroblastoma Staging System criteria,2 54 (21.09%), 65 (25.39%), 44 (17.19%), 77 (30.08%), and 9 (3.52%) patients were diagnosed with clinical stage I, II, III, IV, and 4s neuroblastoma, respectively. Regarding tumor sites, 46 (17.97 %) neuroblastomas were found in the adrenal glands, 87 (33.98%) in retroperitoneal regions, 90 (35.16%) in the mediastinum, and 25 (9.77%) in other regions.
Association between selected polymorphisms and neuroblastoma susceptibility
The genotype counts of the polymorphisms and their associations with neuroblastoma susceptibility are presented in Table 1. We spotted significant association between the rs11037575 T allele and a decreased risk of neuroblastoma (CT vs CC: adjusted OR =0.71, 95% CI =0.51–0.97, P=0.030). However, we observed no significant association with neuroblastoma susceptibility for the rs1027702 T>C and rs10055201 A>G polymorphism. We then combined risk genotypes and found a significant trend toward an increased neuroblastoma risk with the accumulation of risk genotypes (adjusted OR =1.26, 95% CI =1.01–1.58, P=0.043). Participants harboring two to four risk genotypes had a borderline significantly increased neuroblastoma risk (adjusted OR =1.34, 95% CI =0.99–1.81, P=0.055) in comparison to those with zero to one risk genotype.
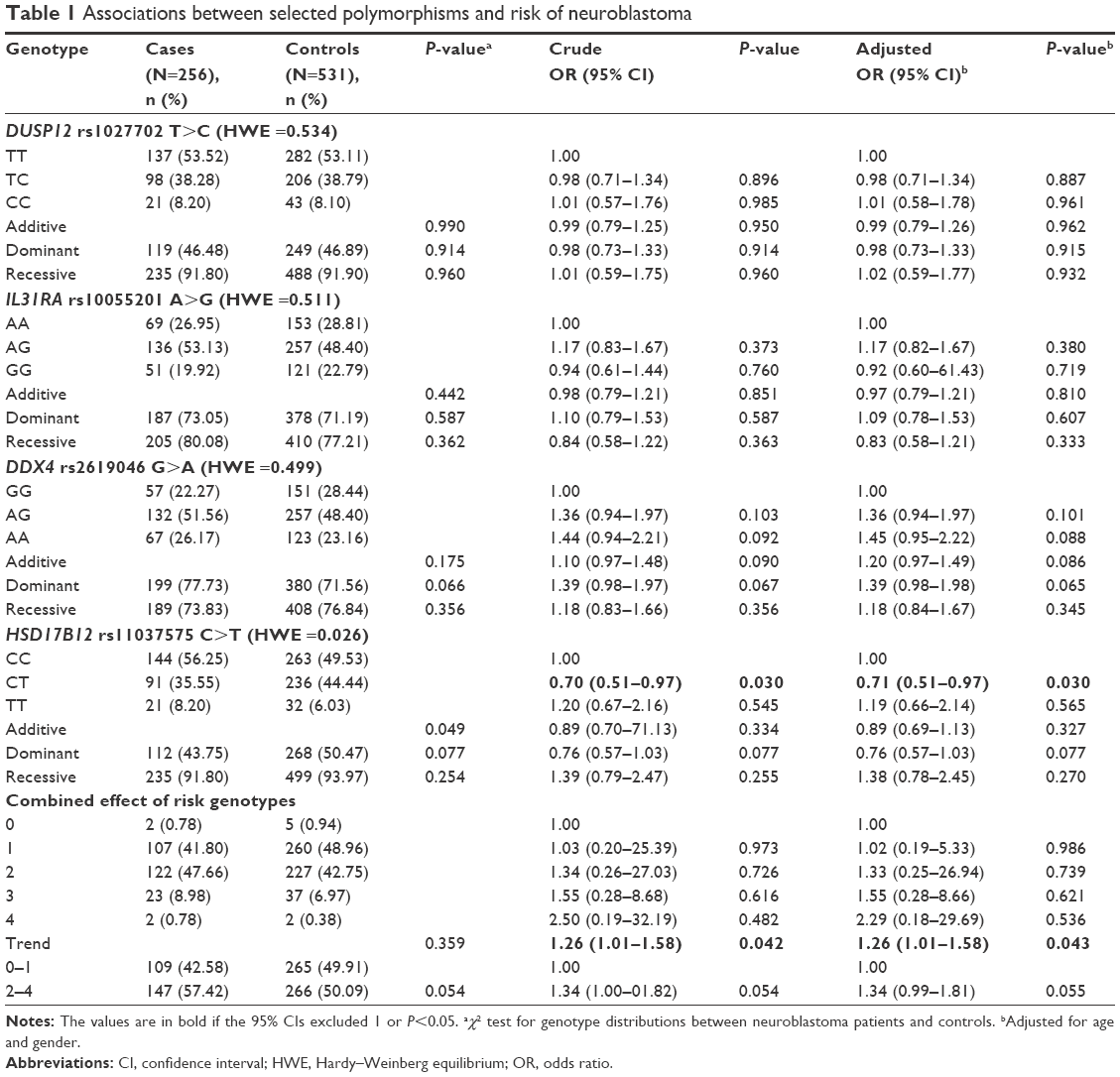 | Table 1 Associations between selected polymorphisms and risk of neuroblastoma |
Stratified analysis of selected polymorphisms and neuroblastoma susceptibility
Next, participants were stratified in terms of age, gender, sites of origin, and clinical stages. We further assessed the effects of variant genotypes of polymorphisms on the risk of neuroblastoma among the different strata (Table 2). We found that the rs11037575 T allele was associated with a decreased neuroblastoma risk among the children aged ≤18 months (CT/TT vs CC: adjusted OR =0.60, 95% CI =0.37–0.97, P=0.036), and those with tumor in mediastinum (CT/TT vs CC: adjusted OR =0.59, 95% CI =0.37–0.94, P=0.025). In the combined analysis, it was shown that the accumulation of risk genotypes (two to four) statistically significantly increased neuroblastoma risk in girls (adjusted OR =1.65, 95% CI =1.03–2.64, P=0.039). Moreover, those with more than one risk genotype tended to develop tumor originated from others (adjusted OR =2.55, 95% CI =1.05–06.23, P=0.039), and were also more likely to have clinical stage III/IV disease (adjusted OR =1.69, 95% CI =1.12–2.54, P=0.012).
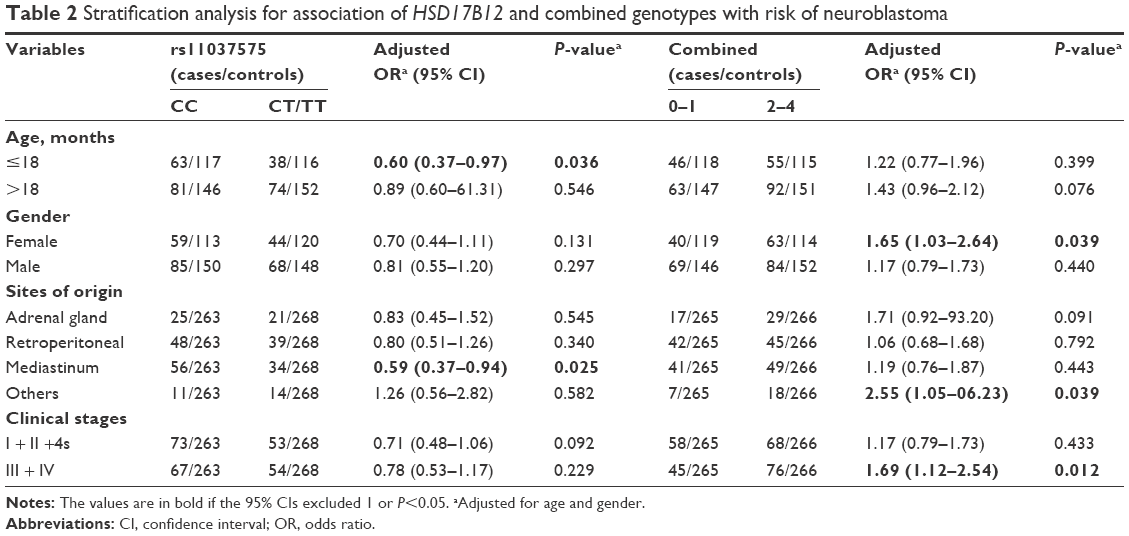 | Table 2 Stratification analysis for association of HSD17B12 and combined genotypes with risk of neuroblastoma |
Discussion
In this study, we evaluated the association of four GWAS-identified polymorphisms (rs1027702 T>C, rs10055201 A>G, rs2619046 G>A, and rs11037575 C>T) with the risk of neuroblastoma in 256 patients and 531 cancer-free controls. Our results demonstrated that rs11037575 T allele protected against neuroblastoma. Moreover, stratified analysis showed that the rs11037575 variants reduced the risk of neuroblastoma among younger subjects (0–18 months of age), and decreased the risk of mediastinal neuroblastoma. When we collectively analyzed risk genotypes, we found that girls carrying two to four risk genotypes had a statistically significantly increased neuroblastoma risk, and patients with two to four risk genotypes tended to develop advanced disease (clinical stage III + IV). The results from the current study suggested that rs11037575 T allele alone had negative effect on neuroblastoma, while combined risk genotypes conferred increased neuroblastoma susceptibility. The rs2619046 allele A (frequency of 0.52 and 0.47 in cases and controls, respectively) showed a trend toward the association with risk of neuroblastoma development (P=0.087, OR =1.202, 95% CI =0.973–1.484) as previously reported in Italians and American Europeans.18,20 These results were in accordance with the findings from previous GWAS study.18 To the best of our knowledge, this is the first investigation to validate the association of neuroblastoma risk with GWAS-identified SNPs within the DUSP12, IL31RA, DDX4, and HSD17B12 genes in a Southern Chinese population.
DUSP12 belongs to the family of dual specificity phosphatases (DUSPs), which function to regulate multiple critical signaling pathways.34 Misregulation of DUSPs contributes to the development of many diseases, including cancers.35,36 DDX4 (VASA) is an ATP-dependent RNA helicase, which fundamentally regulates proliferation and differentiation of germ cell.37 The mammalian HSD17B12 was originally recognized as a 3-ketoacyl-CoA reductase, engaged in the synthesis of long-chain fatty acid.38 Interestingly, decreased expression of HSD17B12 significantly inhibited breast cancer cell proliferation in vitro, which could be fully restored by the addition of arachidonic acid.39 Combined with our findings, we believe that further functional experiments could validate whether rs11037575 C>T polymorphism has a role in HSD17B12 expression. IL-31RA is a unique gp130-like receptor chain of interleukin-31 (IL-31). IL-31 is primarily synthesized by activated CD4 (+) T cells, and mediates activities of a wide spectrum of immune and nonimmune cells. Thus, this cytokine is potentially pleiotropic, which regulates hematopoiesis and immune response, and promotes the development of inflammatory bowel disease, airway hypersensitivity, and dermatitis.40 Nguyen et al completed a two-stage GWAS on neuroblastoma, 574 low-risk cases and 1,722 controls in the first stage and 124 cases and 496 controls in the second stage.18 They found that these four genes and their SNPs were associated with neuroblastoma susceptibility, especially for low-risk neuroblastoma.18 In the previous replication study, comprising 370 neuroblastoma patients and 809 controls, Capasso et al20 confirmed the association of neuroblastoma risk with two independent neuroblastoma-associated common genetic variants (rs1027702, rs11037575) in an Italian population.
Apparently, our findings were not totally consistent with the previous studies conducted among Caucasians, African-Americans, and Italians. The inconsistency may be ascribed to the environmental and genetic variations among different ethnicities. For instance, given the possible differences in the minor allele frequency27 and pattern of linkage disequilibrium of SNPs among Asians, African-Americans, and Caucasians, the effects of the studied SNPs on genetic susceptibility to neuroblastoma may vary. Besides this, the relatively small sample size of this study might have limited the statistical power.
Limitations
There were several possible limitations that should be addressed in this study. First, although it was the largest study in Chinese children, there were only 256 neuroblastoma patients and 531 cancer-free controls included. As a result, the statistical power may be limited. Replication studies from other centers with more sample size were encouraged to validate the association. Second, only four most significant polymorphisms were investigated in the present study, and more polymorphisms, especially the potentially functional SNPs not contained in GWASs, remain to be studied. Third, due to the nature of the retrospective study design, information bias and selection bias might not be avoidable. We could only reduce these biases by frequency-matching of cases and controls by age and gender, due to lack of information on living environment, dietary intake, and parental exposures. Finally, as participants were recruited only from Chinese Han ethnicity residing in Southern China, the findings should be extrapolated to different ethnic groups with great caution.
Conclusion
In conclusion, we verified significant association between HSD17B12 gene rs11037575 T allele and decreased neuroblastoma susceptibility in Southern Chinese children, especially for children aged ≤18 months, and those with tumor of mediastinum region. However, future well-designed prospective studies with larger sample size including different ethnic populations, detailed information (eg, parental exposures), and functional studies are warranted to strengthen our findings.
Acknowledgments
This work was supported by the grants of the State Clinical Key Specialty Construction Project (Pediatric Surgery) 2013 (No: GJLCZD1301), Clinical Medicine Research and Transformation Center of Brain Injury in Premature Infant in Guangzhou (No: 520101–2150092), Guangzhou Science Technology and Innovation Commission (201607010395), and Natural Science Foundation of Guangdong Province, China (2016A030313496). We thank Yanlu Tong and Hezhen Wang for their assistance in DNA extraction and medical histories information collection.
Disclosure
The authors report no conflicts of interest in this work.
References
Maris JM. Recent advances in neuroblastoma. N Engl J Med. 2010;362(23):2202–2211. | ||
Brodeur GM, Pritchard J, Berthold F, et al. Revisions of the international criteria for neuroblastoma diagnosis, staging, and response to treatment. J Clin Oncol. 1993;11(8):1466–1477. | ||
Smith MA, Seibel NL, Altekruse SF, et al. Outcomes for children and adolescents with cancer: challenges for the twenty-first century. J Clin Oncol. 2010;28(15):2625–2634. | ||
Maris JM, Hogarty MD, Bagatell R, Cohn SL. Neuroblastoma. Lancet. 2007;369(9579):2106–2120. | ||
London WB, Castleberry RP, Matthay KK, et al. Evidence for an age cutoff >365 days for neuroblastoma risk group stratification in the Children’s Oncology Group. J Clin Oncol. 2005;23(27):6459–6465. | ||
Deyell RJ, Attiyeh EF. Advances in the understanding of constitutional and somatic genomic alterations in neuroblastoma. Cancer Genet. 2011;204(3):113–121. | ||
Gurney JG, Ross JA, Wall DA, Bleyer WA, Severson RK, Robison LL. Infant cancer in the U.S.: histology-specific incidence and trends, 1973 to 1992. J Pediatr Hematol Oncol. 1997;19(5):428–432. | ||
Bao PP, Li K, Wu CX, et al. [Recent incidences and trends of childhood malignant solid tumors in Shanghai, 2002–2010]. Zhonghua Er Ke Za Zhi. 2013;51(4):288–294. Chinese. | ||
Shojaei-Brosseau T, Chompret A, Abel A, et al. Genetic epidemiology of neuroblastoma: a study of 426 cases at the Institut Gustave-Roussy in France. Pediatr Blood Cancer. 2004;42(1):99–105. | ||
Capasso M, Diskin SJ. Genetics and genomics of neuroblastoma. Cancer Treat Res. 2010;155:65–84. | ||
Maris JM, Mosse YP, Bradfield JP, et al. Chromosome 6p22 locus associated with clinically aggressive neuroblastoma. N Engl J Med. 2008;358(24):2585–2593. | ||
Sridhar S, Al-Moallem B, Kamal H, Terrile M, Stallings RL. New insights into the genetics of neuroblastoma. Mol Diagn Ther. 2013;17(2):63–69. | ||
Han W, Zhou Y, Zhong R, et al. Functional polymorphisms in FAS/FASL system increase the risk of neuroblastoma in Chinese population. PLoS One. 2013;8(8):e71656. | ||
Capasso M, Diskin S, Cimmino F, et al. Common genetic variants in NEFL influence gene expression and neuroblastoma risk. Cancer Res. 2014;74(23):6913–6924. | ||
Lu J, Chu P, Wang H, et al. Candidate Gene Association Analysis of Neuroblastoma in Chinese Children Strengthens the Role of LMO1. PLoS One. 2015;10(6):e0127856. | ||
Diskin SJ, Capasso M, Schnepp RW, et al. Common variation at 6q16 within HACE1 and LIN28B influences susceptibility to neuroblastoma. Nat Genet. 2012;44(10):1126–1130. | ||
Capasso M, Devoto M, Hou C, et al. Common variations in BARD1 influence susceptibility to high-risk neuroblastoma. Nat Genet. 2009;41(6):718–723. | ||
Nguyen le B, Diskin SJ, Capasso M, et al. Phenotype restricted genome-wide association study using a gene-centric approach identifies three low-risk neuroblastoma susceptibility Loci. PLoS Genet. 2011;7(3):e1002026. | ||
Wang K, Diskin SJ, Zhang H, et al. Integrative genomics identifies LMO1 as a neuroblastoma oncogene. Nature. 2011;469(7329):216–220. | ||
Capasso M, Diskin SJ, Totaro F, et al. Replication of GWAS-identified neuroblastoma risk loci strengthens the role of BARD1 and affirms the cumulative effect of genetic variations on disease susceptibility. Carcinogenesis. 2013;34(3):605–611. | ||
He J, Wang F, Zhu J, et al. Association of potentially functional variants in the XPG gene with neuroblastoma risk in a Chinese population. J Cell Mol Med. 2016;20(8):1481–1490. | ||
He J, Yang T, Zhang R, et al. Potentially functional polymorphisms in the LIN28B gene contribute to neuroblastoma susceptibility in Chinese children. J Cell Mol Med. 2016;20(8):1534–1541. | ||
He J, Zhong W, Zeng J, et al. LMO1 gene polymorphisms contribute to decreased neuroblastoma susceptibility in a Southern Chinese population. Oncotarget. 2016;7(16):22770–22778. | ||
Zheng J, Zhang R, Zhu J, et al. Lack of Associations between XPC Gene Polymorphisms and Neuroblastoma Susceptibility in a Chinese Population. Biomed Res Int. 2016;2016:2932049. | ||
Zhang Z, Zhang R, Zhu J, et al. Common variations within HACE1 gene and neuroblastoma susceptibility in a Southern Chinese population. Onco Targets Ther. 2017;10:703–709. | ||
He J, Zhang R, Zou Y, et al. Evaluation of GWAS-identified SNPs at 6p22 with neuroblastoma susceptibility in a Chinese population. Tumour Biol. 2016;37(2):1635–1639. | ||
Zhang R, Zou Y, Zhu J, et al. The Association between GWAS-identified BARD1 Gene SNPs and Neuroblastoma Susceptibility in a Southern Chinese Population. Int J Med Sci. 2016;13(2):133–138. | ||
Fu W, Zhu J, Xiong SW, et al. BARD1 Gene Polymorphisms Confer Nephroblastoma Susceptibility. EBioMedicine. 2017;16:101–105. | ||
He J, Qiu LX, Wang MY, et al. Polymorphisms in the XPG gene and risk of gastric cancer in Chinese populations. Hum Genet. 2012;131(7):1235–1244. | ||
He J, Wang MY, Qiu LX, et al. Genetic variations of mTORC1 genes and risk of gastric cancer in an Eastern Chinese population. Mol Carcinog. 2013;52 (Suppl 1):E70–E79. | ||
Hua RX, Zhuo ZJ, Shen GP, et al. Polymorphisms in the XPC gene and gastric cancer susceptibility in a Southern Chinese population. Onco Targets Ther. 2016;9:5513–5519. | ||
Hua RX, Zhuo ZJ, Zhu J, et al. Association between genetic variants in the XPG gene and gastric cancer risk in a Southern Chinese population. Aging (Albany NY). 2016;8(12):3311–3320. | ||
Zhang Y, Yang D, Zhu JH, Chen MB, Shen WX, He J. The association between NQO1 Pro187Ser polymorphism and urinary system cancer susceptibility: a meta-analysis of 22 studies. Cancer Invest. 2015;33(2):39–40. | ||
Patterson KI, Brummer T, O’Brien PM, Daly RJ. Dual-specificity phosphatases: critical regulators with diverse cellular targets. Biochem J. 2009;418(3):475–489. | ||
Elchebly M, Payette P, Michaliszyn E, et al. Increased insulin sensitivity and obesity resistance in mice lacking the protein tyrosine phosphatase-1B gene. Science. 1999;283(5407):1544–1548. | ||
Pulido R, Hooft van Huijsduijnen R. Protein tyrosine phosphatases: dual-specificity phosphatases in health and disease. FEBS J. 2008;275(5):848–866. | ||
Linder P. Dead-box proteins: a family affair – active and passive players in RNP-remodeling. Nucleic Acids Res. 2006;34(15):4168–4180. | ||
Moon YA, Horton JD. Identification of two mammalian reductases involved in the two-carbon fatty acyl elongation cascade. J Biol Chem. 2003;278(9):7335–7343. | ||
Nagasaki S, Miki Y, Akahira J, Suzuki T, Sasano H. Transcriptional regulation of 17beta-hydroxysteroid dehydrogenase type 12 by SREBP-1. Mol Cell Endocrinol. 2009;307(1–2):163–168. | ||
Zhang Q, Putheti P, Zhou Q, Liu Q, Gao W. Structures and biological functions of IL-31 and IL-31 receptors. Cytokine Growth Factor Rev. 2008;19(5–6):347–356. |
Supplementary material
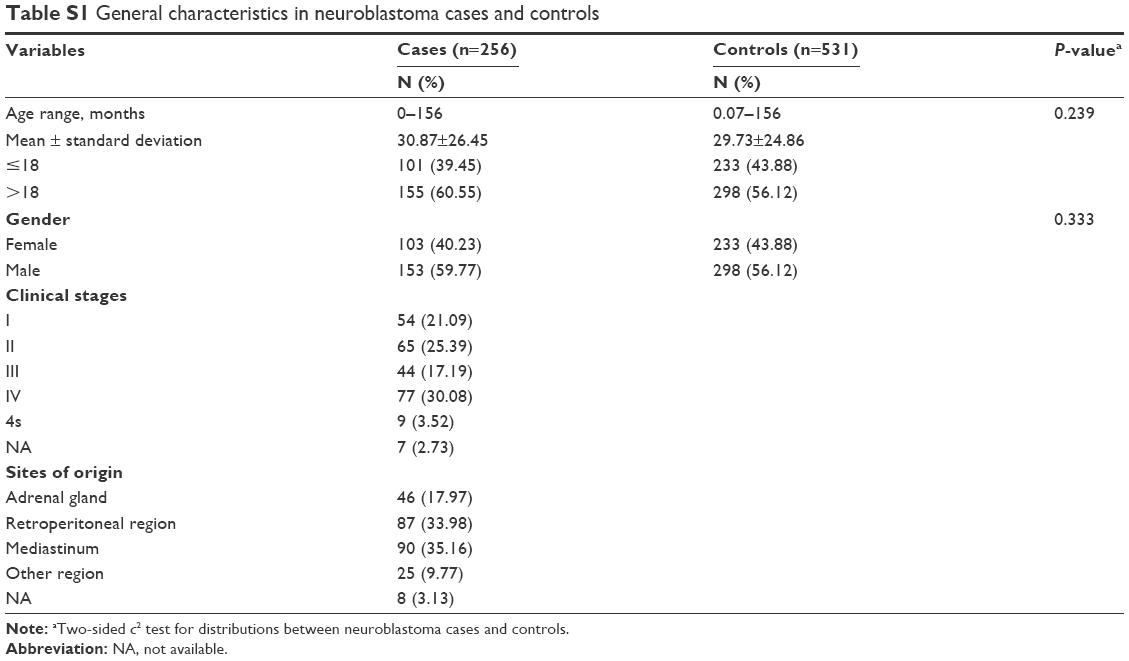 | Table S1 General characteristics in neuroblastoma cases and controls |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.