Back to Journals » Journal of Experimental Pharmacology » Volume 18
Hesperidin Mitigates Bisphenol-A Induced Oxidative Stress, Endocrine Disruption and Testicular Damage in Adult Male Wistar Rats
Authors Oviosun A ![]() , Oviosun EC, Usman IM, Anyanwu CN
, Oviosun EC, Usman IM, Anyanwu CN ![]() , Ojiakor VO, Esom EA, Anyanwu EG
, Ojiakor VO, Esom EA, Anyanwu EG ![]()
Received 12 January 2026
Accepted for publication 18 May 2026
Published 22 May 2026 Volume 2026:18 595476
DOI https://doi.org/10.2147/JEP.S595476
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Abdelwahab Omri
Augustine Oviosun,1 Ezinne Chidinma Oviosun,1,2 Ibe Micheal Usman,3 Chinyere Nkemjika Anyanwu,4 Vivian Onyinye Ojiakor,1 Emmanuel Anayochukwu Esom,5 Emeka Godson Anyanwu1,5
1Department of Anatomy, Faculty of Biomedical Sciences, Kampala International University, Western Campus, Ishaka-Bushenyi, Uganda; 2Department of Anatomy, Faculty of Basic Medical Sciences, Ambrose Alli University, Ekpoma, Edo State, Nigeria; 3Department of Human Anatomy, Faculty Medicine and Pharmacy, University of Rwanda, Butare, Rwanda; 4Department of Microbiology and Immunology, Faculty of Biomedical Sciences, Kampala International University, Ishaka-Bushenyi, Uganda; 5Department of Anatomy, Faculty of Basic Medical Sciences, University of Nigeria, Enugu Campus, Enugu, Nigeria
Correspondence: Augustine Oviosun, Department of Anatomy, Faculty of Biomedical Sciences, Kampala International University, Western Campus, Ishaka-Bushenyi, Uganda, Tel +234-8068259663, Email [email protected]
Introduction: Bisphenol A (BPA), a well-known endocrine disruptor, is newly emerging as causing male infertility by endocrine disturbances, induction of oxidative stress and testicular toxicity. This study investigates the protective role of hesperidin against BPA-induced testicular toxicity in male Wistar rats.
Methodology: The rats were divided into six groups (n=7): control (normal saline); BPA-only (50 mg/kg); hesperidin-only at 50 mg/kg and 100 mg/kg; and two pre-treated groups receiving hesperidin (50 and 100 mg/kg) followed by BPA (50 mg/kg). All treatments were administered orally for 8 weeks. Post-treatment analyses included testicular weight, volume, and diameter measurements; serum testosterone, FSH, and LH levels; oxidative stress markers (MDA, SOD, CAT, GPx, and GSH); and histological and morphometric evaluation of the testis.
Result and Discussion: BPA exposure significantly reduced testicular dimensions, decreased serum testosterone, FSH, and LH. Histopathological examination revealed degeneration of seminiferous tubules, germ-cell depletion, reduced counts of spermatogonia, spermatocytes, spermatids, Sertoli and Leydig cells. BPA significantly induced oxidative stress, evidenced by increased MDA and decreased SOD, CAT, GPx, and GSH. Hesperidin significantly mitigated these effects by restoring testicular morphology, hormonal profiles, enhancing oxidative stress markers, improving histology and cell populations.
Conclusion: Hesperidin demonstrates protective properties against BPA-induced testicular and endocrine toxicity in rats, likely as a result of its antioxidant properties.
Keywords: hesperidin, bisphenol A, oxidative stress, testicular toxicity, endocrine
Introduction
Increasing attention has been directed toward the harmful effects of environmental toxins on male fertility, especially their capacity to disrupt the testicular microenvironment, impairing spermatogenesis and hormonal regulation. Despite the relatively low oxygen tension in the testicular microenvironment, testicular tissue remains highly susceptible to oxidative stress1 Environmental toxins can cause an excessive increase in Reactive oxygen species (ROS), which can lead to oxidative stress, causing damage to sperm DNA, impairing steroid hormone production, and disrupting cell signalling pathways, ultimately affecting male fertility.2,3 However, exposure to environmental pollutants like Bisphenol A (BPA) disrupts the delicate balance between oxidants and antioxidants, thereby compromising testicular function and potentially leading to male infertility.2,4,5 Bisphenol A (BPA) is an industrial chemical compound widely used as a monomer in the production of polycarbonate plastics, epoxy resins, thermal paper, and food-contact materials such as can linings and reusable containers.4,6 Its unique properties, durability, clarity, heat resistance, and adhesive strength have made BPA indispensable in industries ranging from consumer electronics and compact discs to cosmetics, toys, food containers, including reusable beverage bottles, and medical devices.7–9
The primary mechanism by which the general population is exposed to BPA is through their diet; however, dermal absorption, especially from handling thermal-printing receipts, contributes significantly and is particularly important for those who are exposed in the workplace, such as cashiers.7,10 Studies have shown that BPA is an endocrine-disrupting chemical capable of interfering with hormonal homeostasis, particularly reproductive hormone regulation.11,12
It has been proven that BPA is harmful to the kidney, liver, testis, and brain, mostly due to oxidative damage, mitochondrial instability, inflammation and apoptosis.13–17 BPA has been related in studies to disturbed spermatogenesis, decreased sperm counts, decreased motility, and sperm DNA damage, all of which are detrimental to male fertility.18,19
There is a growing body of research patronising medicinal plants in the management of male infertility.14,20–22 Citrus fruits and vegetables contain a polyphenolic compound called hesperidin, which is the primary flavonoid found in oranges and lemons.23,24 Hesperidin is known to have antioxidant, anti-inflammatory, anticancer, and anti-lipidemic activities.16,23 Its potent anti-apoptotic and reactive oxygen species (ROS)-scavenging properties have demonstrated protective effects against a range of toxicant-induced injuries, including hepatotoxicity, nephrotoxicity, and testicular dysfunction.25,26 Hesperidin’s special effectiveness against BPA-induced testicular and endocrine disruption is still insufficiently studied, despite strong evidence of its protective benefits against general reproductive toxins. Current literature remains limited in providing systematic dose–response evaluations of hesperidin, along with detailed mechanistic evidence linking oxidative stress to steroidogenic dysfunction and broader endocrine axis disruption following bisphenol A (BPA) exposure. Therefore, this study investigated the dose-dependent protective effects of hesperidin against BPA-induced reproductive toxicity in adult male Wistar rats, with emphasis on oxidative stress biomarkers, endocrine hormone profiles, and testicular histopathological alterations. Unlike previous reports that primarily focused on sperm parameters and apoptosis, the present study expands the scope by integrating comprehensive endocrine disruption analysis and mechanistic biochemical assessment of hormonal levels. Collectively, these findings further support hesperidin as a potential natural gonadoprotective agent against BPA-induced testicular damage.
Materials and Methods
Chemical/Reagents
We purchased Hesperidin (CAS No. 520–26-3) and Bisphenol A (CAS No. 80–05-7) from Sigma-Aldrich (St. Louis, MO, USA); all chemicals and reagents used for this study were of high analytical grade.
Experimental Design
Forty-two male Wistar rats weighing 200–220 g were obtained from the University of Nigeria, Veterinary Medicine Department, and randomly assigned to six equal groups (n = 7). The sample size was determined using power analysis with an expected effect size of 0.8, α = 0.05, and power (1−β) of 0.8, based on preliminary studies. They were kept in standard laboratory settings, which included a 12-hour light-dark cycle, a constant temperature of 22 ± 2 °C, and a humidity of 55 ± 5%. They were also given unrestricted access to clean water and a standard mouse feed. All animal procedures were conducted in strict compliance with European Directive 2010/63/EU on the protection of animals for scientific purposes, and performed under protocols approved by the Department of Anatomy and Faculty of Basic Medical Science (FBMS), University of Nigeria, Enugu Campus, Nigeria (UNN/ FBMS/COMHRECR/2022/01). Following the 3Rs principle, every effort was made to minimise the number of animals used and to alleviate any potential pain. The experimental design included pre-treatment protocol. The animal grouping and administration design are shown in Table 1. The BPA dose used in this study (50 mg/kg) was selected based on toxicological studies indicating that this level reliably induces reproducible testicular damage and oxidative stress, thereby enabling evaluation of protective interventions such as hesperidin.27,28 However, this dose is higher than typical human environmental exposure and may represent a subchronic toxicity model rather than real-life conditions; therefore, systemic effects beyond the testes cannot be excluded. Hesperidin is a naturally occurring flavonoid with a generally favourable safety profile in experimental studies. The doses of Hesperidin and Bisphenol-A used in this study were from previous studies.27–30
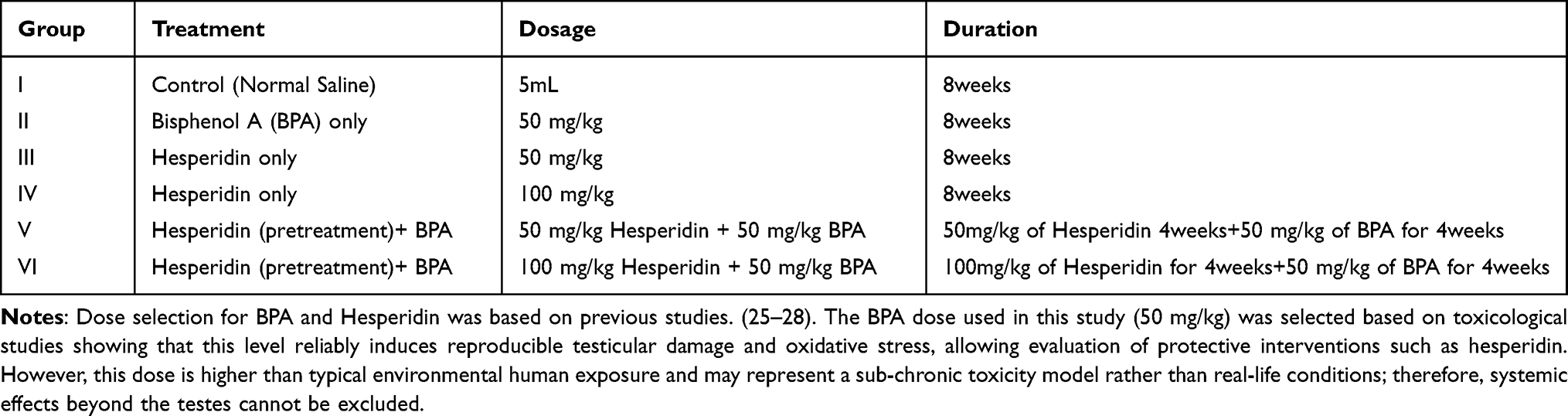 |
Table 1 Experimental Grouping |
Animal Sacrifice, Sample Collection and Testicular Measurement
Following eight weeks of treatment, rats were anaesthetized via inhalation and euthanized through cervical dislocation. Blood samples were collected via cardiac puncture; blood samples were obtained and placed into siliconized glass tubes containing EDTA as an anticoagulant. These samples were centrifuged for 15 minutes at 4 °C, and the resulting plasma was stored at −20 °C until analysis. The testis was harvested, blotted dry and weighed using an electronic scale. One testis was rinsed with an icy cold normal saline solution (0.9%, w/v), divided into minute pieces before being homogenised in 3 volumes of ice-cold suitable buffer (TBS, pH 7.4) using an Ultra Turrax homogenizer T25. Until used for enzyme assays and lipid peroxidation, supernatants were collected, aliquoted, and kept at 80 °C. The standard water displacement method, which was based on Archimedes’ principle, was used to measure testicular volume. Each testis was gently submerged in a graduated cylinder that contained a known volume of water, and the rise in water level, the difference between the initial and final volume, was recorded as the actual volume of the testis.
Estimation of the Plasma Levels of Follicle-Stimulating Hormone (FSH), Luteinizing Hormone (LH) and Testosterone
Plasma FSH and LH levels were measured using ELISA (AccuBind ELISA Microwells, Monobind Inc., Lake Forest, CA, USA), and testosterone levels were measured by competitive radioimmunoassay (RIA) using a ^125I-labelled testosterone analogue (Immunotech, Beckman Coulter, France). The sensitivity of the ELISA kits was 0.1 mIU/mL for FSH and 0.05 mIU/mL for LH, with intra-assay and inter-assay coefficients of variation of 5% and 8%, respectively. The RIA for testosterone had a sensitivity of 0.05 ng/mL, with intra-assay and inter-assay CVs of 6% and 9%, respectively. All samples were measured in duplicate, and mean values were used for statistical analysis.
Estimation of Antioxidant Enzymes and Lipid Peroxidation
Catalase (CAT) Activity
Catalase activity was quantified following the method outlined by Aebi (1984) and previously reported by.31 This method is based on the ability of catalase to decompose hydrogen peroxide (H2O2) into water and oxygen. The reaction mixture containing the sample homogenate and phosphate buffer was initiated by the addition of hydrogen peroxide, and the decrease in absorbance due to H2O2 decomposition was monitored spectrophotometrically at 240 nm over a fixed time interval. Catalase activity was assumed to be proportional to the rate of absorbance reduction. The activity of the enzymes was then reported in units/mg of protein.
Superoxide Dismutase (SOD) Activity
Superoxide dismutase (SOD) activity was measured spectrophotometrically using a modified version of the pyrogallol autoxidation assay originally described by.32 The principle of the assay relies on the ability of SOD to suppress autoxidation of pyrogallol in alkaline conditions. Absorbance was measured at 420 nm and enzyme activity was determined. One unit of SOD activity was defined as the amount of enzyme required to inhibit pyrogallol autoxidation by 50% under the assay conditions. SOD activity was calculated and expressed as units per mg protein (U/mg protein).
Glutathione Peroxidase (GPx) Activity
Glutathione peroxidase (GPx) activity was determined using the fixed-time Trichloroacetic acid (TCA) method of.33 The assay is based on the enzyme-catalyzed oxidation of reduced glutathione (GSH) by GPx in the presence of hydrogen peroxide where the remaining GSH is measured after precipitation with TCA. The activity of GPx was determined by the consumption rate of GSH and expressed in units per milligram of protein (U/mg protein).
Reduced Glutathione (GSH) Concentration
Reduced glutathione (GSH) was quantified by the classic Ellman assay.34 This method is based on the reaction of free sulfhydryl groups in GSH with 5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB) to produce the yellow-colored chromophore, 5-thio-2-nitrobenzoic acid (TNB). Absorbance was spectrophotometrically measured at 412 nm, and GSH concentration was calculated using the molar extinction coefficient of TNB. The results were expressed as nmol GSH/mg protein.
Malondialdehyde Activity
Malondialdehyde (MDA) levels were quantified using the thiobarbituric acid reactive substances (TBARS) assay, as described by.35 And results were expressed as nmol/mg protein. The TBARS method was selected because it is a simple, sensitive, and widely accepted assay for assessing lipid peroxidation and oxidative stress. Although widely used as an index of lipid peroxidation, the TBARS assay lacks absolute specificity for MDA because thiobarbituric acid can react with other aldehydes and oxidised biomolecules, potentially leading to overestimation of lipid peroxidation levels.
Histological and Histomorphometric Analysis
One of the excised testes was quickly fixed in Bouin’s fluid for routine tissue processing as previously described31 The tissue was then dehydrated, embedded in paraffin, and sectioned at 5 µm thickness. Hematoxylin and eosin (H&E) staining was performed to visualize cellular and structural details. Histological examination was conducted using a Leica DM 750 light microscope.
Histomorphometric analysis was performed using digital photomicrographs and ImageJ software (NIH, USA). For each animal, three non-serial sections were selected, and 10 randomly chosen non-overlapping microscopic fields per section were analyzed at ×40 magnification. Only round or nearly round seminiferous tubule cross-sections were included to avoid bias due to tangential sectioning. A minimum of 30 seminiferous tubules per animal were evaluated. All measurements and cell counting were performed by an investigator blinded to the experimental groups.
Seminiferous tubule diameter was calculated as the mean of two perpendicular diameters, while epithelial height was measured from the basement membrane to the luminal border. Germ cell populations (spermatogonia, spermatocytes, and spermatids) and Sertoli cells were identified within the seminiferous epithelium, while Leydig cells were identified in the interstitial tissue. Cells were counted only when nuclei were clearly visible. Damaged or folded tissue areas were excluded from analysis to avoid counting bias.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism version 10 (GraphPad Software, San Diego, CA, USA). Data normality was assessed using the Shapiro–Wilk test, and homogeneity of variances was evaluated using Levene’s test. Comparisons among multiple groups were conducted using one-way analysis of variance (ANOVA). When a significant overall difference was detected, Tukey’s multiple comparisons post hoc test was applied to identify pairwise differences among groups. Exact p-values were reported, and statistical significance was defined as p < 0.05. Data are presented as mean ± standard error of the mean (SEM) and displayed using bar charts and summary tables.
Results
Effects of Hesperidin on Testicular Weight and Volume
The effects of Hesperidin on testicular weight and volume following BPA exposure are shown in Table 2. The exposure of rats to 50 mg/kg of BPA for 8 weeks caused a significant (p < 0.05) decrease in testis weight and volume compared with the control group and the hesperidin-treated groups. However, pretreatment with (50mg/kg and 100mg/kg) hesperidin significantly improved (p < 0.05) these parameters.
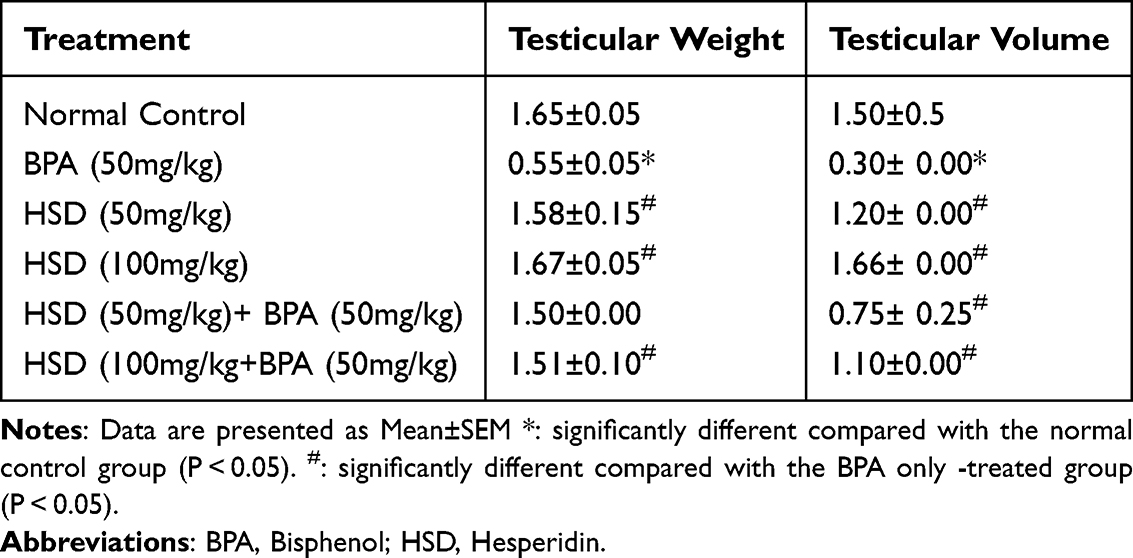 |
Table 2 Effects of Hesperidin on Testicular Measurement Following BPA Exposure |
Effect of Hesperidin on Hormonal Assay Results of Serum FSH, LH and Testosterone Levels
(Figure 1a–c) illustrates how BPA affects plasma levels of FSH, LH, and testosterone. Rats administered 50 mg/kg of BPA for eight weeks showed a significant (p < 0.05) decrease in these hormone levels compared with the control group. Alterations of these hormones were significantly reversed (p < 0.05) by pretreatment with hesperidin (50mg/kg and 100mg/kg) compared to BPA-intoxicated groups.
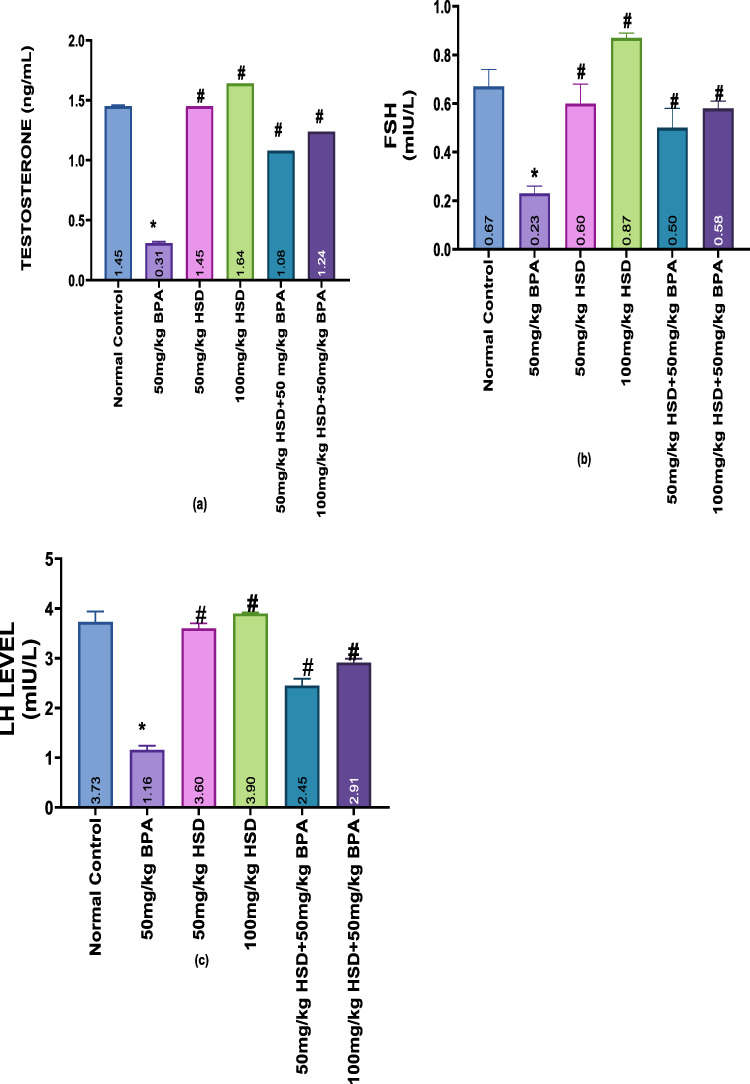 |
Figure 1 (a–c) Bar charts showing Hormonal Assay (a) Testosterone Level (b) FSH Level (c) LH Level. Data are presented as Mean±SEM. Data are presented as Mean±SEM. *Significantly different from the normal control group (P < 0.05). #: significantly different from the BPA-only-treated group (P < 0.05) n=5. Abbreviations: BPA, Bisphenol; HSD, Hesperidin. |
Effect of Hesperidin on Antioxidant Status and Lipid Peroxidation Levels
The results of lipid peroxidation and testicular antioxidant status are shown in (Figure 2a–e). The testes’ SOD, CAT, GPx activity, and GSH levels significantly (p < 0.05) decreased after 8 weeks of exposure to 50 mg/kg of BPA, whereas the MDA levels increased in comparison to the control group, even in the hesperidin-only groups. However, pretreatment with hesperidin after exposure to 50mg/kg of BPA significantly improved (p < 0.05) these parameters.
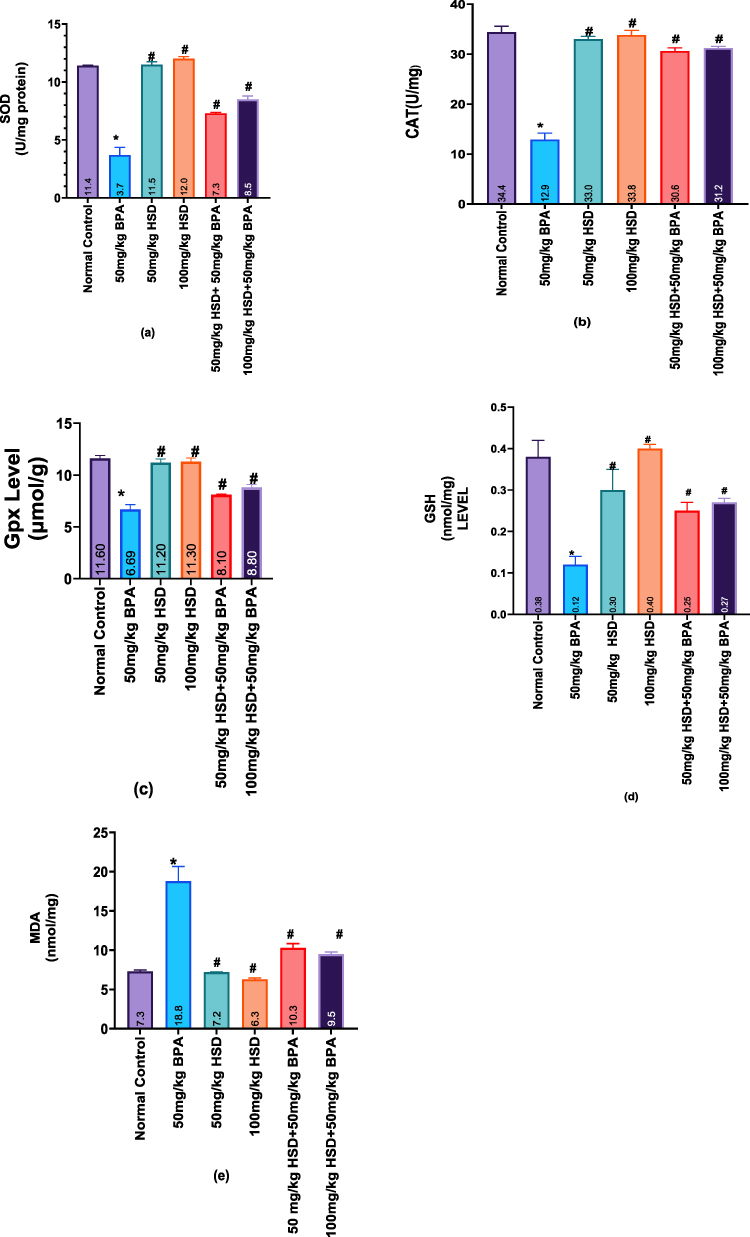 |
Figure 2 (a–e) Bar Charts showing Values for anti-oxidative stress markers. (a) SOD level (b), CAT level (c) Gpx level (d), GSH level (e), and MDA level. Data are presented as Mean±SEM. Data are presented as Mean±SEM. *: significantly different from the normal control group (P < 0.05). #: significantly different from the BPA-only-treated group (P < 0.05). n=5. |
Effect of Hesperidin on Histological Examination of the Testis
As shown in (Figure 3A–F), in contrast to hesperidin alone (50 and 100 mg/kg), which preserved normal architecture, testicular histology showed that BPA (50 mg/kg) caused significant disorganization of the germinal epithelium, vacuolization, germ-cell sloughing, widened interstitial spaces, vascular congestion, and focal tubule wall damage. Co-administration with Hesperidin (100 mg/kg) fully restored tubule structure and spermatogenesis, with almost total reversal of BPA-induced lesions, whereas the 50 mg/kg co-treatment only partially recovered.
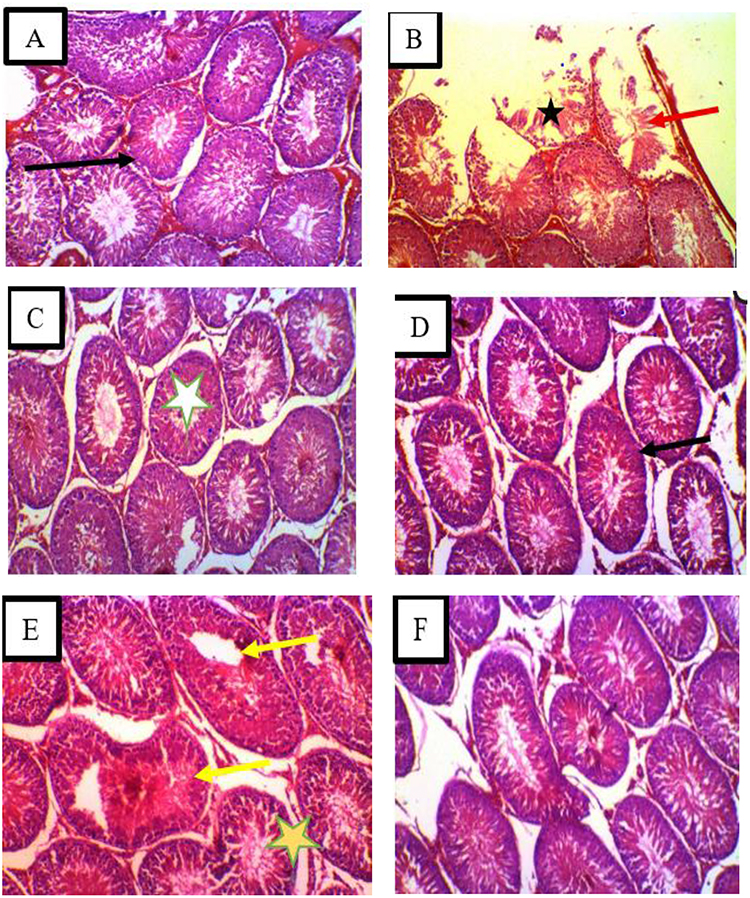 |
Figure 3 (A–F) Testicular Histology in Wistar Rats Following Bisphenol A (BPA) Exposure and Hesperidin Treatment. Photomicrographs of Haematoxylin & Eosin-stained testicular sections from Wistar rats (X200). (A) Normal control – Seminiferous tubules are oval with complete spermatogenic layers and abundant spermatozoa in the lumen. Interstitial connective tissue appears normal. (B) BPA (50 mg/kg) – Marked histopathological changes: disorganisation of the germinal epithelium (red arrows), vacuolization, sloughing of germ cells, widened interstitial spaces, vascular congestion, and focal destruction of seminiferous tubule walls (indicated by star). (C) Hesperidin (50 mg/kg) and (D) Hesperidin (100 mg/kg) – Histology resembles that of the normal control: seminiferous tubules are uniform and well-organised, with intact germ cell layers, complete spermatogenesis, and normal interstitial tissue. (E) BPA + Hesperidin (50 mg/kg) – Partial recovery: minor interstitial edema and slight vacuolization remain; a few seminiferous tubules show necrotic germinal epithelium (yellow arrows). (F) BPA + Hesperidin (100 mg/kg) – signs of restoration: tubule structure normalized, active spermatogenesis evident (elongated spermatids and spermatozoa present), interstitial spaces returned to baseline, and inflammation significantly reduced. Stars indicate the lumen of a seminiferous tubule. Scale bar = 50 µm. |
Effects of Hesperidin on Histomorphometric Parameters, Following BPA Exposure
The result in (Figure 4a and b) shows that rats exposed to 50 mg/kg of BPA for 8 weeks caused a significant (p < 0.05) decrease in seminiferous tubule height and diameter, compared with those of the control group and hesperidin only. However, pre-treatment with hesperidin following BPA exposure significantly improved (p < 0.05) the seminiferous tubule height and diameter.
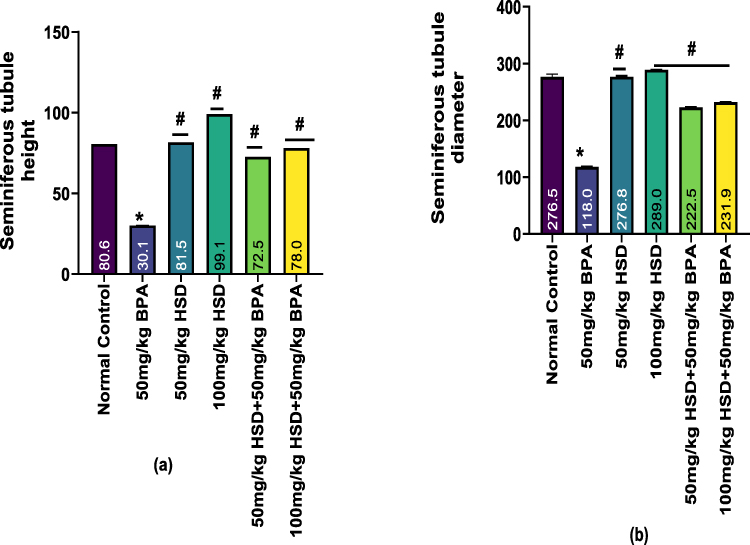 |
Figure 4 (a and b) Bar charts showing histomorphometric parameters (a) Seminiferous tubule height, (b) Seminiferous tubule diameter. Data are presented as Mean±SEM. *Significantly different from the normal control group (P < 0.05). #: significantly different compared with the BPA only -treated group (P < 0.05). n=5. Abbreviations: BPA, Bisphenol; HSD, Hesperidin. |
Effects of Hesperidin on Testicular Germ Cells Following BPA Exposure
Results of the Effects of Hesperidin on the number of spermatogonia, spermatocytes and spermatids, Leydig and Sertoli cells following BPA exposure are shown in (Figure 5a–c and 6a and b). The exposure of rats to 50mg/kg of BPA for 8 weeks caused a significant (p < 0.05) decrease in the number of spermatogonia, spermatocytes, spermatids, Sertoli and Leydig cells compared with those of the control group and hesperidin-only groups. However, pretreatment with hesperidin following exposure to 50mg/kg of BPA significantly improved (p < 0.05) these parameters.
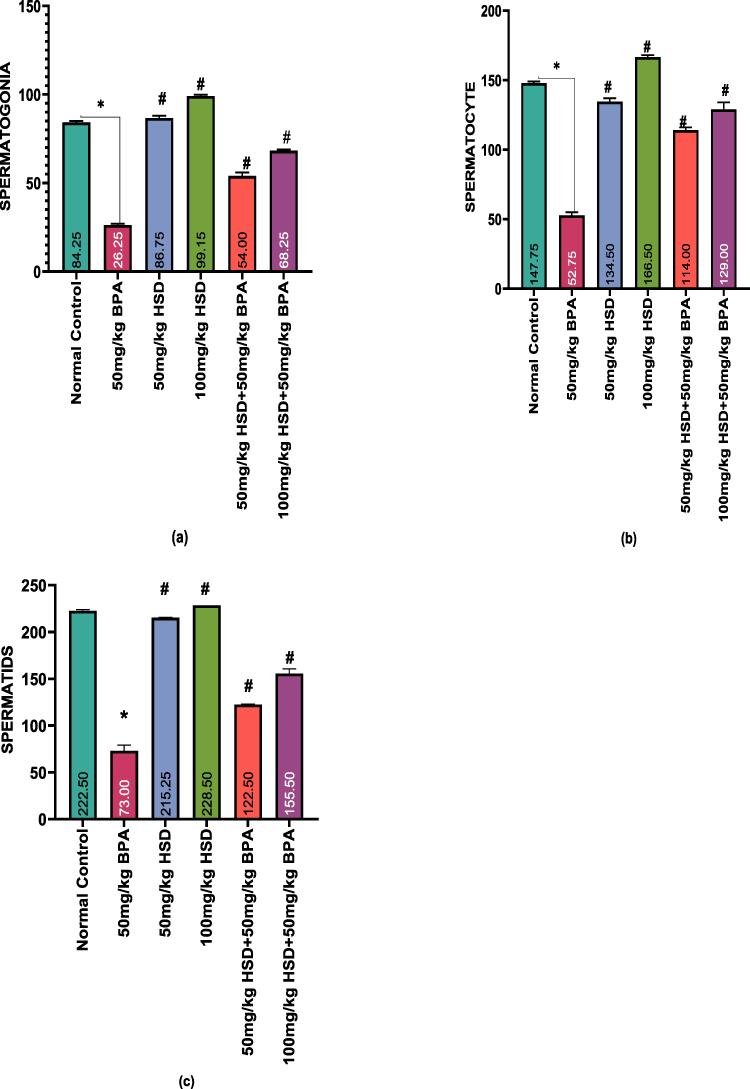 |
Figure 5 (a–c) Bar charts showing Effects of Hesperidin on Testicular Germ Cells following BPA exposure. Data are presented as Mean±SEM *Significantly different compared with the normal control group (P < 0.05). #: significantly different compared with the BPA only -treated group (P < 0.05). n=5. Abbreviations: BPA, Bisphenol; HSD, Hesperidin. |
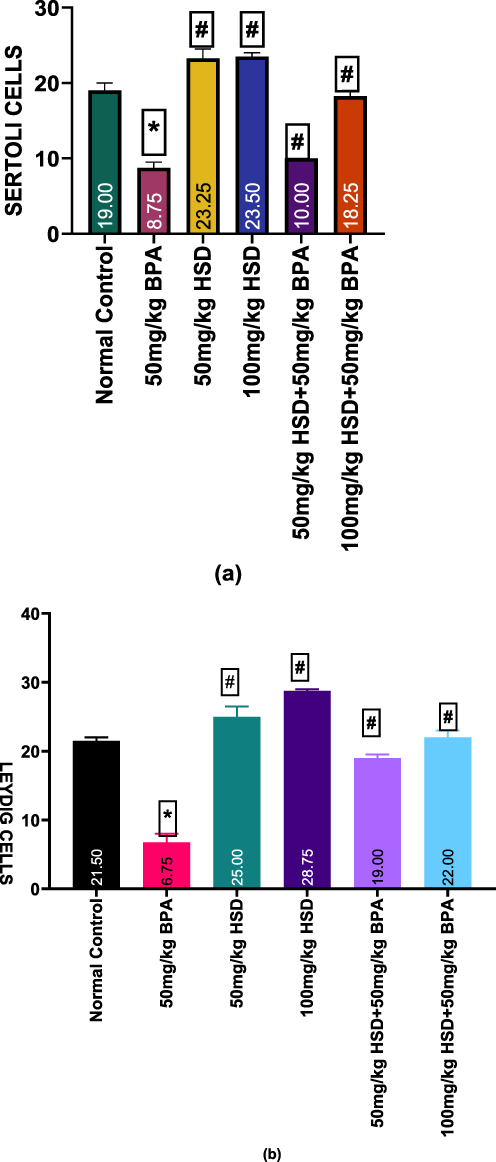 |
Figure 6 (a and b) Bar charts showing Effects of Hesperidin on Sertoli and Leydig cells following BPA exposure. Data are presented as Mean±SEM *Significantly different compared with the normal control group (P <0.0001). #: significantly different compared with the BPA only -treated group (P <0.0001). n=5. Abbreviations: BPA, Bisphenol; HSD, Hesperidin. |
Discussion
In the present study, oral exposure to 50 mg/kg BPA significantly decreased testicular weight and volume, purportedly due to derangement of the seminiferous tubules and Leydig cells. Both low and high concentrations of BPA have been observed to consistently decrease testicular weight in experimental animals.36 The decline in testicular size may reflect impairment of seminiferous tubule integrity, germ cell depletion, and reduced interstitial cell populations, all of which contribute to overall tissue degeneration. The noted decrease in testicular volume can thus be correlated to the reduction of the spermatogenic cells as well as structural disfigurement of seminiferous tubules, which has been previously reported in previous models of BPA toxicity.36,37
Interestingly, the administration of 50mg/kg and 100mg/kg of hesperidin significantly enhanced the testicular weight and volume compared to the BPA-treated animals. This finding confirms the previous research indicating that hesperidin could prevent the reproductive toxicity of chemically induced testicular tissue damage. The improvement in organ weight and volume may reflect the ability of hesperidin to preserve testicular cellular organisation and reduce tissue degeneration.38,39 Hesperidin administration has been occasionally shown to augment or even restore testicular weight to normal levels when compared to control groups.
In this study, BPA exposure significantly reduced serum testosterone levels, indicating impaired Leydig cell function and compromised androgen production. This result is consistent with previous studies that revealed BPA upsets androgen production by negatively affecting Leydig cell structure and functionality.40,41 This observation is consistent with previous studies that revealed that BPA interferes with androgen production by negatively affecting the structure and functionality of Leydig cells. These findings were further corroborated by histopathological results of rats treated with BPA, where degenerative alterations and lowered integrity of Leydig cells were observed. Testosterone plays a crucial role in ensuring spermatogenesis, and therefore, the decreased testosterone levels induced by BPA could be one of the causes of the impaired spermatogenic process and structural defects found in the testes.42,43 In addition to testosterone reduction, BPA administration caused a significant decline in serum LH and FSH levels. The present study is in line with the earlier reports that BPA may disrupt hormonal regulation at the hypothalamic-pituitary-gonadal axis.4,44 Although the present study did not assess the hypothalamic or pituitary molecular markers, the observed reduction in gonadotropins indicates that BPA exposure can affect hormonal signalling that is central to normal testicular functioning.45,46
Notably, hesperidin treatment (50mg/kg and 100mg/kg), either alone or as pretreatment before BPA exposure, significantly restored testosterone, LH, and FSH levels toward normal values. The higher dose of hesperidin showed a stronger protective effect on hormonal recovery, suggesting a dose-dependent effect. This is in agreement with other studies.46,47 Although the current result is clear in terms of functional recovery in endocrine parameters, the exact mechanisms by which this recovery was achieved could not be conclusively determined, since the steroidogenic enzymes and regulatory proteins were not assessed. Therefore, the observed hormonal normalization should be interpreted as an improvement in reproductive endocrine function rather than direct evidence of steroidogenic pathway restoration.
Oxidative stress has been implicated extensively as one of the primary contributors to reproductive toxicity caused by BPA. In the current experiment, exposure to BPA caused a dramatic decrease in activities of testicular antioxidant enzymes, GSH levels, and a dramatic increase in lipid peroxidation, which validated the onset of oxidative stress. Antioxidant enzymes, including SOD and CAT, are major defense mechanisms against reactive oxygen species, and their loss indicates a loss of testicular tissue capacity to counteract oxidative damage. The decrease in the activity of GPx and in GSH could be another indication of more active exploitation of the antioxidant stores due to overproduction of ROS. The high increase in MDA levels in BPA-treated rats is indicative of higher lipid peroxidation, which means damage to the membrane and cellular impairment. These results are consistent with other studies that have documented oxidative imbalance and increased lipid peroxidation in testes exposed to BPA.17,48–50
Hesperidin administration significantly improved antioxidant enzyme activities, including SOD, CAT, GPx, and glutathione reductase, while markedly reducing MDA levels. These findings hint that hesperidin was capable of reducing the effects of BPA on oxidative damage, probably by improving antioxidant defense mechanisms and suppression of lipid peroxidation. Previous toxicological research has reported similar antioxidant protective effects of hesperidin. Combined, the biochemical results considerably prove the protective effect of hesperidin in the present study in connection with the antioxidant activity.51,52
Histopathological examination further confirmed the biochemical and endocrine results. Exposure to BPA resulted in severe testicular injury with degeneration of seminiferous tubules, vacuolization, and disrupted germinal epithelium architecture. These morphological defects align with previous findings that indicate that BPA leads to severe degeneration of spermatogenic cells and disruption of testicular differentiation.37,53 Conversely, treatment with hesperidin maintained testicular architecture and enhanced seminiferous tubule structure in a dose-dependent fashion. The higher hesperidin dose produced more pronounced improvement in spermatogenic progression and seminiferous tubule integrity, indicating stronger protective activity.
According to other studies, BPA have a serious impact on the testicles and reproductive tract in male rats. The improvement of histoarchitecture in the group treated with hesperidin after BPA exposure can be attributed to a decrease in oxidative stress.38,52
Morphometric measurements also indicated that BPA exposure caused a considerable decrease in seminiferous tubule diameter and epithelial height, which indicates damage to spermatogenic activity. The observed decrease could be due to the depletion and degeneration of the spermatogenic epithelium and the fact that the seminiferous tube sizes are positively correlated with spermatogenic efficiency. Exposure to BPA also had a significant effect of lowering the count of spermatogonia, spermatocytes and spermatids, which signifies interference with the development of germ cells. These findings are consistent with other studies that have shown that BPA has negative effects on germ cell populations and spermatogenesis.54 Hesperidin treatment considerably increased these spermatogenic indices, which implies improved recovery of germinal epithelium and restoration of spermatogenic capacity, which is consistent with other studies.18,55
Although the present findings indicate that hesperidin effectively mitigated BPA-induced reproductive toxicity, the study has certain limitations. Inflammatory markers, apoptosis-related proteins, and steroidogenic enzyme expression were not assessed in this work; however, these parameters have been evaluated in previous related studies.17
The protective effects observed with hesperidin are consistent with those reported for other well-known plant-derived bioactive compounds, such as curcumin, which has also demonstrated significant antioxidant and reproductive protective properties against BPA-induced toxicity.21 This similarity suggests that dietary flavonoids and polyphenols may share common protective mechanisms, primarily through attenuation of oxidative stress and preservation of testicular function, supporting their potential role as natural therapeutic agents against environmental toxicant-induced reproductive damage.
Conclusion
This study demonstrates that BPA induces significant testicular toxicity, evidenced by reduced testicular weight and volume, hormonal disruption, oxidative stress, histological damage, and impaired spermatogenesis. These effects are associated with degeneration of testicular cellular organization and compromised seminiferous tubule integrity. Hesperidin treatment, particularly at higher dose (100mg/kg), significantly alleviated BPA-induced damage by improving antioxidant defences, reducing lipid peroxidation, preserving testicular histoarchitecture, restoring endocrine parameters, and enhancing spermatogenesis. The protective effect of hesperidin in this study is most strongly supported by its antioxidant activity, although additional mechanistic studies are required to confirm its involvement in other pathways.
Abbreviation
HSD, Hesperidin; BPA, Bisphenol-A; CAT, Catalase; SOD, Superoxide dismutase; Gpx, Glutathione peroxidase; GSH, Glutathione; MDA, Malondialdehyde; LH, Luteinizing Hormone; FSH, Follicle-Stimulating Hormone; ROS, Reactive Oxygen Species.
Data Sharing Statement
The data set or material used for this study will be available on request from the corresponding author.
Ethical Statement
All animal procedures were conducted in strict compliance with European Directive 2010/63/EU on the protection of animals for scientific purposes, and performed under protocols approved by the Department of Anatomy and Faculty of Basic Medical Science (FBMS), University of Nigeria, Enugu Campus, Nigeria (UNN/ FBMS/COMHREC/2022/01).
Acknowledgment
The authors wish to appreciate the Department of Anatomy, University of Nigeria, Nsukka and all technical staff who assisted in the technical support during the experimental phase of this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
No funding was received for this study.
Disclosure
The authors declared that there is no conflict of interest.
References
1. Dutta S, Sengupta P, Slama P. Oxidative stress, testicular inflammatory pathways, and male reproduction. Int J Mol Sci. 2021;22(Figure 1):1–15.
2. Bardaweel SK, Gul M, Alzweiri M, Ishaqat A, Alsalamat HA, Bashatwah RM. Reactive oxygen species: the dual role in physiological and pathological conditions of the human body. Eurasian J Med. 2018;50(3):193–201. doi:10.5152/eurasianjmed.2018.17397
3. Darbandi M, Darbandi S, Agarwal A, et al. Reactive oxygen species and male reproductive hormones. Reprod Biol Endocrinol. 2018;16(1):1–14. doi:10.1186/s12958-018-0406-2
4. Krzastek SC, Farhi J, Gray M, Smith RP. Impact of environmental toxin exposure on male fertility potential. Transl Androl Urol. 2021;9(6):2797–2813. doi:10.21037/tau-20-685
5. Chidinma E, Oviosun A, Johnson N. Phytomedicine Plus Hesperidin protects against spermatological damages and testicular apoptosis induced by Bisphenol-A in adult male Wistar rats. Phytomed Plus. 2025;5(2):100797. doi:10.1016/j.phyplu.2025.100797
6. Maniradhan M, Calivarathan L. Bisphenol A-induced endocrine dysfunction and its associated metabolic disorders. Endocrine Metab Immune Disord Drug Targets. 2022;23(4):515–529. doi:10.2174/1871530322666220928144043
7. Cimmino I, Fiory F, Perruolo G, et al. Potential mechanisms of bisphenol a (BPA) contributing to human disease. Int J Mol Sci. 2020;21(16):1–22. doi:10.3390/ijms21165761
8. Manzoor MF, Tariq T, Fatima B, et al. An insight into bisphenol A, food exposure and its adverse effects on health: a review. Front Nutr. 2022;9(November):1–16. doi:10.3389/fnut.2022.1047827
9. Agarwal A, Gandhi S, Tripathi AD, Gupta A, Iammarino M, Sidhu JK. Critical reviews in biotechnology food contamination from packaging material with special focus on the bisphenol-A. Crit Rev Biotechnol. 2025;45(1):69–79. doi:10.1080/07388551.2024.2344571
10. Lee I, Kim S, tae KK, Kim S, Park S, Lee H. Bisphenol A exposure through receipt handling and its association with insulin resistance among female cashiers. Environ Int. 2018;117(May):268–275. doi:10.1016/j.envint.2018.05.013
11. Oladipe TT, Uadia PO. Occupational exposure to bisphenol A (BPA) is a risk factor for coronary heart disease. J Health Sci. 2022;12(1):1–6. doi:10.5923/j.health.20221201.01
12. Bousoumah R, Leso V, Iavicoli I, et al. Science of the total environment biomonitoring of occupational exposure to bisphenol A, bisphenol S and bisphenol F: a systematic review. Sci Total Environ. 2021;783:146905. doi:10.1016/j.scitotenv.2021.146905
13. Abdou HM, Abd Elkader HTAE, El-Gendy AH, Eweda SM. Neurotoxicity and neuroinflammatory effects of bisphenol A in male rats: the neuroprotective role of grape seed proanthocyanidins. Environ Sci Pollut Res. 2022;29(6):9257–9268. doi:10.1007/s11356-021-16311-1
14. Adeniyi IA, Onaadepo O, Owu D, et al. Exploring the male fertility potential of medicinal plants from central and west african countries: a systematic review. Phytomed Plus. 2025:100786. 10.1016/j.phyplu.2025.100786.
15. Costa HE, Cairrao E. Effect of Bisphenol a on the Neurological System: A Review Update. Vol. 98. Berlin Heidelberg: Springer; 2024. doi:10.1007/s00204-023-03614-0
16. Oviosun EC, Oviosun A, Nto NJ, Okwara BO, Anyanwu EG. Hesperidin protects against bisphenol-A-induced renal damage in adult male wistar rats. Trop J Nat Prod Res. 2025;9(June):2802–2808.
17. Oviosun EC, Oviosun A, Nto NJ, Anyanwu EG. Hesperidin protects against spermatological damages and testicular apoptosis induced by Bisphenol-A in adult male Wistar rats. Phytomed Plus. 2025;5(2):100797. doi:10.1016/j.phyplu.2025.100797
18. Presunto M, Mariana M, Lorigo M, Cairrao E. The effects of bisphenol A on human male infertility: a review of current epidemiological studies. Int J Mol Sci. 2023;24(15). doi:10.3390/ijms241512417
19. Salami EA, Rotimi OA. The impact of Bisphenol-A on human reproductive health. Toxicol Rep. 2024;13(June):101773. doi:10.1016/j.toxrep.2024.101773
20. Roozbeh N, Amirian A, Abdi F, Haghdoost S. A systematic review on use of medicinal plants for male infertility treatment. J Fam Reprod Heal. 2021;15(2):74–81. doi:10.18502/jfrh.v15i2.6447
21. Bakeer MR, Rashad MM, Ahmed O, et al. Antioxidant and anti ‑ apoptotic effects of curcumin ‑ loaded nanoliposomes against di ‑ n ‑ butyl phthalate ‑ induced hepatorenal damage in rats. Naunyn Schmiedebergs Arch Pharmacol. 2026;(0123456789). doi:10.1007/s00210-026-05212-0
22. Soliman SS, Ahmed O, Youssef FS, et al. Astragalus polysaccharides protect against Di-n-butyl phthalate-induced testicular damage by modulating oxidative stress, apoptosis, and the PI3K / Akt / mTOR pathway in rats. Front Vet Sci. 2025;(July):1–13. doi:10.3389/fvets.2025.1616186
23. Singh N, Saini S, Nagpal N, et al. Pharmacological and therapeutic potential of hesperidin-A comprehensive review. Eur J Org Chem. 2023;2023(8):4499–4537.
24. Parasuraman S, Huey LY, Jee YS. Effect of hesperidin in the prevention of aluminum chloride-induced testicular dysfunction in rats. J Adv Pharm Technol Res. 2024;15(3):194–199. doi:10.4103/JAPTR.JAPTR_458_23
25. Hussein R, Khalaf M, Mohamed W. Hesperidin and eugenol attenuate cadmium-induced nephrotoxicity via regulation of oxidative stress, Bax/Bcl2 and cleaved caspase 3 expression. Turk J Biochem. 2020;45(6):767–775. doi:10.1515/tjb-2020-0161
26. Kasem SE, Abdelnaby AA, Mohammed PA, Hemdan SB, Abd El-Fattah RMZ, Elsayed RM. Protective effect of hesperidin on kidneys and testes of adult male rats exposed to bisphenol A. Egypt J Hosp Med. 2022;88(1):3005–3013. doi:10.21608/ejhm.2022.244529
27. Shaukat S, Umbreen F, Ali H, Khokhar A. Protective effect of soya bean oil against Bisphenol A induced interstitial lung fibrosis in mice. Prof Med J. 2021;28(10):1457–1462. doi:10.29309/tpmj/2021.28.10.6391
28. Cao T, Cao Y, Wang H, et al. The effect of exposure to bisphenol A on spermatozoon and the expression of tight junction protein occludin in male mice. Dose Response. 2020;18(2):1–6. doi:10.1177/1559325820926745
29. Carballo-Villalobos AI, González-Trujano ME, Pellicer F, López-Muñoz FJ. Antihyperalgesic effect of hesperidin improves with diosmin in experimental neuropathic pain. Biomed Res Int. 2016;2016. 10.1155/2016/8263463.
30. Celik E, Oguzturk H, Sahin N, Turtay MG, Oguz F, Ciftci O. Protective effects of hesperidin in experimental testicular ischemia/reperfusion injury in rats. Arch Med Sci. 2016;12(5):928–934. doi:10.5114/aoms.2015.47697
31. Emeka AG, Augustine O, Chidinma OE, Nto NJ. Zingerone improves memory impairment in Wistar rats exposed to cadmium via modulation of redox imbalance. J Krishna Inst Med Sci Univ. 2023;12(1):3–16.
32. Marklund S, Marklund G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur J Biochem. 1974;474:469–474.
33. Flohé L, Günzler WA. Assays of gluthathione peroxidase. Methods Enzymol. 1984;105:114–120.
34. Ellman GL. Tissue Su ~ yd ~ l Groups. 1959;70–77.
35. Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues thiobarbituric acid reaction. Anal Biochem. 1979;358:351–358.
36. Behmanesh MA, Najafzadehvarzi H, Poormoosavi SM. Protective effect of aloe vera extract against bisphenol a induced testicular toxicity in wistar rats. Cell J. 2018;20(2):278–283. doi:10.22074/cellj.2018.5256
37. Munir B, Qadir A, Tahir M. Negative effects of bisphenol A on testicular functions in albino rats and their abolitions with Tribulus terristeris L. Braz J Pharm Sci. 2017;53(3):1–9. doi:10.1590/s2175-97902017000300104
38. Abu-Khudir R, Almutairi HH, Abd El-Rahman SS, El-Said KS. The palliative and antioxidant effects of hesperidin against lead-acetate-induced testicular injury in male wistar rats. Biomedicines. 2023;11(9):1–17. doi:10.3390/biomedicines11092390
39. Khamis T, Hegazy AA, El-Fatah SSA, et al. Hesperidin mitigates cyclophosphamide-induced testicular dysfunction via altering the hypothalamic pituitary gonadal axis and testicular steroidogenesis, inflammation, and apoptosis in male rats. Pharmaceuticals. 2023;16(2):1–20. doi:10.3390/ph16020301
40. Hammad AY, Hammad AY, Adam SIY, Abdelgadir WS. The impact of bisphenol A on gonadal hormones and histological structure of wistar rats. Clin Trials Case Stud. 2024;1–7. doi:10.31579/2835-835X/082
41. Pivonello C, Muscogiuri G, Nardone A, et al. Bisphenol A: an emerging threat to female fertility. Reprod Biol Endocrinol. 2020;18(1):4–11. doi:10.1186/s12958-019-0558-8
42. Ryu DY, Pang WK, Rahman MS, Park YJ, Pang MG. Differential susceptibility of Leydig and Sertoli cells to bisphenol A. Toxicology. 2025;516(May):154182. doi:10.1016/j.tox.2025.154182
43. Adegoke EO, Rahman MS, Pang MG. Bisphenols threaten male reproductive health via testicular cells. Front Endocrinol. 2020;11(September):1–18. doi:10.3389/fendo.2020.00624
44. Yesildemir O, Celik MN. The effect of various environmental pollutants on the reproductive health in children: a brief review of the literature. Curr Nutr Rep. 2024;13(3):382–392. doi:10.1007/s13668-024-00557-5
45. Pironti C, Ricciardi M, Proto A, Bianco PM, Montano L, Motta O. Endocrine-disrupting compounds: an overview on their occurrence in the aquatic environment and human exposure. Water. 2021;13(10):1–32. doi:10.3390/w13101347
46. Deiab NS, Kodous AS, Mahfouz MK, Said AM, Ghobashy MM, Abozaid OAR. Smart Hesperidin/Chitosan nanogel mitigates apoptosis and endoplasmic reticulum stress in fluoride and aluminum-induced testicular injury. Biol Trace Elem Res. 2024;202(9):4106–4124. doi:10.1007/s12011-023-03991-8
47. Hari Priya B, Ramya B, Bora S, et al. Mitigating cyclophosphamide-associated gonadotoxicity in male Wistar rats: exploring the therapeutic potential of hesperidin. Front Vet Sci. 2024;11(May):1–13. doi:10.3389/fvets.2024.1376225
48. Meli R, Monnolo A, Annunziata C, Pirozzi C, Ferrante MC. Oxidative stress and BPA toxicity: an antioxidant approach for male and female reproductive dysfunction. Antioxidants. 2020;9(5). doi:10.3390/antiox9050405
49. Amjad S, Rahman MS, Pang MG. role of antioxidants in alleviating bisphenol A toxicity. Biomol. 2020;10(8):1105. doi:10.3390/BIOM10081105
50. Zhao Z, Yao J, Guo Z, et al. Zinc alleviates oxidative stress injury of the testis induced by Zearalenone through Sirt3-Foxo3 pathway in mice. Biol Trace Elem Res. 2025. doi:10.1007/s12011-025-04707-w
51. Adefegha SA, Rosa Leal DB, Olabiyi AA, Oboh G, Castilhos LG. Hesperidin attenuates inflammation and oxidative damage in pleural exudates and liver of rat model of pleurisy. Redox Rep. 2017;22(6):563–571. doi:10.1080/13510002.2017.1344013
52. Yan L, Wang J, Dai D, Zhang Y, Li Y, Xiao W. Testicular protective effects of hesperidin against chemical and biological toxicants. Toxicol Res. 2024;13(3):1–9. doi:10.1093/toxres/tfae078
53. Asma M, Fouzia T, Lazhari T, Khireddine O. Protective effects of apigenin against Bisphenol A-induced testis toxicity in Wistar rats through modulating hepatic biochemical biomarkers and histological changes. Comp Clin Path. 2020;29(5):1041–1049. doi:10.1007/s00580-020-03158-0
54. Nawaz F, Asmatullah, Ara C. Assessment of deleterious effects of bisphenol a (Bpa) on steroidogenesis, sperm count, and spermatogenesis in a mammalian model. J Anim Plant Sci. 2021;31(6):1568–1574. doi:10.36899/JAPS.2021.6.0360
55. Jahan S, Ain QU, Ullah H. Therapeutic effects of quercetin against bisphenol A induced testicular damage in male Sprague Dawley rats. Syst Biol Reprod Med. 2016;62(2):114–124. doi:10.3109/19396368.2015.1115139
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Assessment of the Impact of Bidens pilosa on Behavioral, Oxidative Stress and Cerebellar Cortical Histoarchitectural Alterations During Bisphenol A Exposure in Mice
Jama IA, Usman IM, Oviosun A, Eze ED, Adeniyi IA, Etukudo EM, Kwizera E, Owembabazi E, Anyanwu E
Journal of Experimental Pharmacology 2025, 17:403-416
Published Date: 27 June 2025