Back to Journals » International Journal of Women's Health » Volume 18
Gut Microbiota-Immune Interactions in Endometrial Cancer: Causal Mediation and Subtype-Specific Mechanisms
Authors Yu S, Shu W, Zhang J, Cheng S ![]() , Shen X, Chen G, Zhang T, Dong K, Zhang J, Wang H
, Shen X, Chen G, Zhang T, Dong K, Zhang J, Wang H
Received 21 November 2025
Accepted for publication 11 March 2026
Published 17 March 2026 Volume 2026:18 583327
DOI https://doi.org/10.2147/IJWH.S583327
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Everett Magann
Shuyang Yu,1 Wan Shu,1 Jiarui Zhang,1 Shuangshuang Cheng,1 Xiaoyu Shen,1 Guanxiao Chen,1 Tangansu Zhang,1 Kejun Dong,1 Jun Zhang,1 Hongbo Wang1,2
1Department of Obstetrics and Gynecology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, People’s Republic of China; 2Clinical Research Center of Cancer Immunotherapy, Union Hospital, Wuhan, Hubei, People’s Republic of China
Correspondence: Hongbo Wang, Department of Obstetrics and Gynecology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, People’s Republic of China, Email [email protected]
Purpose: In this study, we applied two-sample Mendelian randomization (MR) to explore the causal effects between gut microbiota (GM), immune cells, and endometrial cancer (EC) subtypes and to assess whether immune cells mediate the impact of GM on EC.
Patients and Methods: Using two-sample Mendelian randomization and mediation analysis, we analyzed GWAS data: GM (Dutch Microbiome Project; N=7738), EC subtypes (IEU Open GWAS; N=331,588), and immune traits (N=3757). We assessed the effects of the causal gut microbiota on EC subtypes and immune trait mediation.
Results: Subtype-specific causal relationships were identified.
Overall EC: Four positive (e.g. genus Erysipelotrichaceae noname) and three negative (e.g. species Bacteroides faecis) microbial causal effects; three mediated by immune traits (e.g. Ruminococcus obeum via CD86+ myeloid DC AC). Endometrioid EC: Five negative (e.g. class Bacilli) and two positive (e.g. species Aspergillus senegalensis) effects; three immune-mediated (e.g. Bacilli via IgD+ CD38br % lymphocytes). Non-endometrioid EC: Two positive (e.g. species Bacteroides stercoris) and one negative (species Ruminococcus bromii) effect; one mediated (Ruminococcus bromii via CD8br NKT % lymphocytes).
Conclusion: Immune traits significantly mediated causal pathways from GM to EC development. It also highlighted the distinct causal relationships and immune-mediated mechanisms across the three major EC subtypes (overall, endometrioid, and non-endometrioid). These subtype-specific insights into the gut-immune-cancer axis provide novel perspectives for developing therapeutic strategies targeting GM and the immune microenvironment in different EC subtypes.
Keywords: causal mediation analysis, gut microbiota, immunity, mendelian randomization analysis, uterine neoplasms
Introduction
Endometrial cancer (EC) is one of the most common gynecological malignancies, and its incidence has increased progressively in high-income regions.1 According to their histological features, grade, and hormone receptor (ER and PR) expression, EC can generally be classified into types I and II.1–3
Type I EC is the most frequent subtype, presenting as low-grade, endometrioid, and hormone receptor-positive with a good prognosis (85% 5-year OS rate).1,2 Conversely, Type II EC is non-endometrioid, high-grade, and hormone receptor-negative, accompanied by high-risked metastasis and poor prognosis (5-year OS rate of ~55%).2,4 Genetic and environmental factors, including obesity and metabolic and reproductive factors, are the primary risk factors for the occurrence and development of EC.1,5 Notably, these risks are strongly linked to the gut and vaginal microbiota; for instance, the GM may affect endometrial carcinogenesis through altering the systemic and uterine cavity microenvironment, providing a novel therapy for EC.2,6,7 In addition, EC, especially type II EC, is regarded as an immunogenic disease, and its infiltrating immune cells affect anti-cancer therapy to facilitate tumor progression, ultimately leading to the development of immune tolerance.8–10 Malignant and non-malignant immune cells and signaling molecules constitute the tumor microenvironment surrounded by blood vessels, disrupting therapeutic efficacy.11–13 Immunotherapy is a prospective therapeutic intervention for carcinostasis by activating the immune system.14,15 Recently, immunotherapy has demonstrated benefits in treating recurrent and advanced EC.14–17
Microbiota refers to the collection of microorganisms in a particular community, whereas the genome of microorganisms is defined as a microbiome.18–20 Evidence suggests that crosstalk between microbiota and the gut immune system is crucial.21 With further exploration of the gut microbiota (GM), the disrupted balance of the GM may influence various disease states, such as obesity, mental disorders, and autoimmune diseases.22–24 GM can impact human health by regulating host immunity and metabolism.25–28 Therefore, regulating host immunity by GM can prominently affect a variety of treatment efficacies and toxicities in cancer.18
Furthermore, it is recognized that the gut microbiota typically functions within a community rather than as individual bacteria. Increasing evidence suggested that microbial communities as regulated networks, reshape host immunity and tumor progression. Multi-omics analyses indicated that certain microbial communities are associated with immune infiltration in tumors. For instance, an integrated tumor-immune-microbiota network in colorectal cancer performed that, Firmicutes enriched communities are accompanied by enhanced cytotoxic T cell responses, while the Proteobacteria characteristics are related with immunosuppressive myeloid infiltration.29 Moreover, microbiota derived metabolites such as inosine and bile acids, have been proven to regulate anti-tumor immunity through T cells and NKT cells.25,30 Clinical studies further supported that the enrichment of Ruminococcaceae and Bacteroidetes species are correlated with improved responses to anti-PD-1 treatment in melanoma patients.31,32 These findings demonstrated that microbiota induced immune regulation represents a conserved mechanism across various tumors. However, it is unclear that whether there are subtype-specific microbiota-immune pathways in EC.
Mendelian randomization (MR), a data analysis tool used in epidemiological studies to evaluate causal inferences, uses genetic variants strongly correlated with exposure factors as instrumental variables to assess the causal relationship between exposure factors and outcomes.23–35 MR generally presents single nucleotide polymorphisms (SNPs) that serve as genetic variants to estimate the causal effect of exposure on outcomes with less susceptibility to environmental confounders and reverse causality.33 Single-sample MR data are derived from the same individual, whereas two-sample MR uses large-scale GWAS data from independent study populations.36,37 Large-scale summary statistics can be used to analyze the relationship between the GM, immune cells, and EC, enhancing the statistical power of the two-sample MR Analysis.
In this study, comprehensive MR Analysis was performed to explore the causal effects of GM, immune cells, and various EC types, including EC and EC with endometrioid and non-endometrioid histology. Next, we discuss whether immune cells serve as mediators from GM to EC.
Materials and Methods
Study Design
Bidirectional two-sample univariable Mendelian randomization (UVMR) was applied to explore the causal association between GM and the three types of EC: EC overall, endometrioid histology, and non-endometrioid histology. Subsequently, two-step MR analysis was performed to determine whether immune cell traits mediated these causal associations (Figure 1).
 |
Figure 1 Overview flow of the two-sample univariable Mendelian randomization (UVMR) and mediation analysis design. GM, gut microbiota; EC, endometrial cancer; SNPs, single nucleotide polymorphisms; IVs, instrumental variables. β0 (total effect): The causal function of GM on EC; β1 (direct effect A): The causal function of GM on immune cell traits; β2 (direct effect B): The causal function of immune cell traits on EC; β (mediating effect) = β1 (Direct effect A) × β2 (Direct effect B); mediated proportion = β (mediating effect) / β0 (total effect). All the bolds present the summary or generic terms of MR analysis. GWAS summary data (in bold): The summary of data sources; Gut microbiota (LifeLines Cohort Study) (in bold): The summary of gut microbiota; Mediator (in bold): The data sources of immune cell traits as mediators; Endometrial cancer (GWAS) (in bold): The summary of 3 types endometrial cancer from GWAS; Exposure variables (in bold): The exposure of MR study; Outcome variables (in bold): The outcomes of MR study; Mediating variables (in bold): The mediators of MR study; Mendelian randomized analysis (in bold): The methods used for MR analysis; Sensitivity analysis (in bold): The methods used for assessing sensitivity. |
First, the causal relationship between the GM and EC was assessed with UVMR using three filters: SNPs should (1) be strongly associated with EC, (2) affect EC only through the causal effect of the GM, and (3) remove other potential confounders.38 Subsequently, reverse MR analysis was conducted to screen for GMs with reverse functions in the EC. Second, the mediating effects of various immune cells in the corresponding GM and EC were evaluated and quantified using UVMR and Mendelian randomization. The above analysis followed the guidelines for Strengthening the Reporting of Observational Studies in Epidemiology using Mendelian Randomization (STRBOE-MR) (Table S1).39
Data Sources
GM data were summarized from the Dutch Microbiome Project (DMP), a GWAS of 7738 European individuals.40 The project applied shotgun metagenomic sequencing of fecal samples, which identified 207 microbial taxa (5 phyla, 10 classes, 13 orders, 26 families, 48 genera, and 105 species).
A total of 731 immune-related genome-wide features with approximately 22 million variants in 3757 individuals of European ancestry were identified using GWAS Summary Statistics.41 Integrated GWAS summary Statistics, with classified numbers from GCST0001391 to GCST0002121 published in the GWAS directory.
Genetic summary data for the two-sample and mediated MR analyses were primarily obtained from the IEU Open GWAS project and included endometrial cancer (ebi-a-GCST90018838), endometrial cancer with endometrioid histology (ebi-a-GCST006465), and non-endometrioid histology (ebi-a-GCST006466). All data are publicly available as GWAS summary data and received ethical approval (Table 1). This Mendelian randomization study is based solely on publicly available summary statistics from previously published genome-wide association studies. All original studies obtained informed consent and ethical approval from their respective institutional review boards. This study is exempt from approval based on Article 32, Item 1, of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects, issued by China on 18 February 2023. As no new human participants were involved and no individual-level data were analyzed in the present study, ethical approval for this specific analysis was not required.
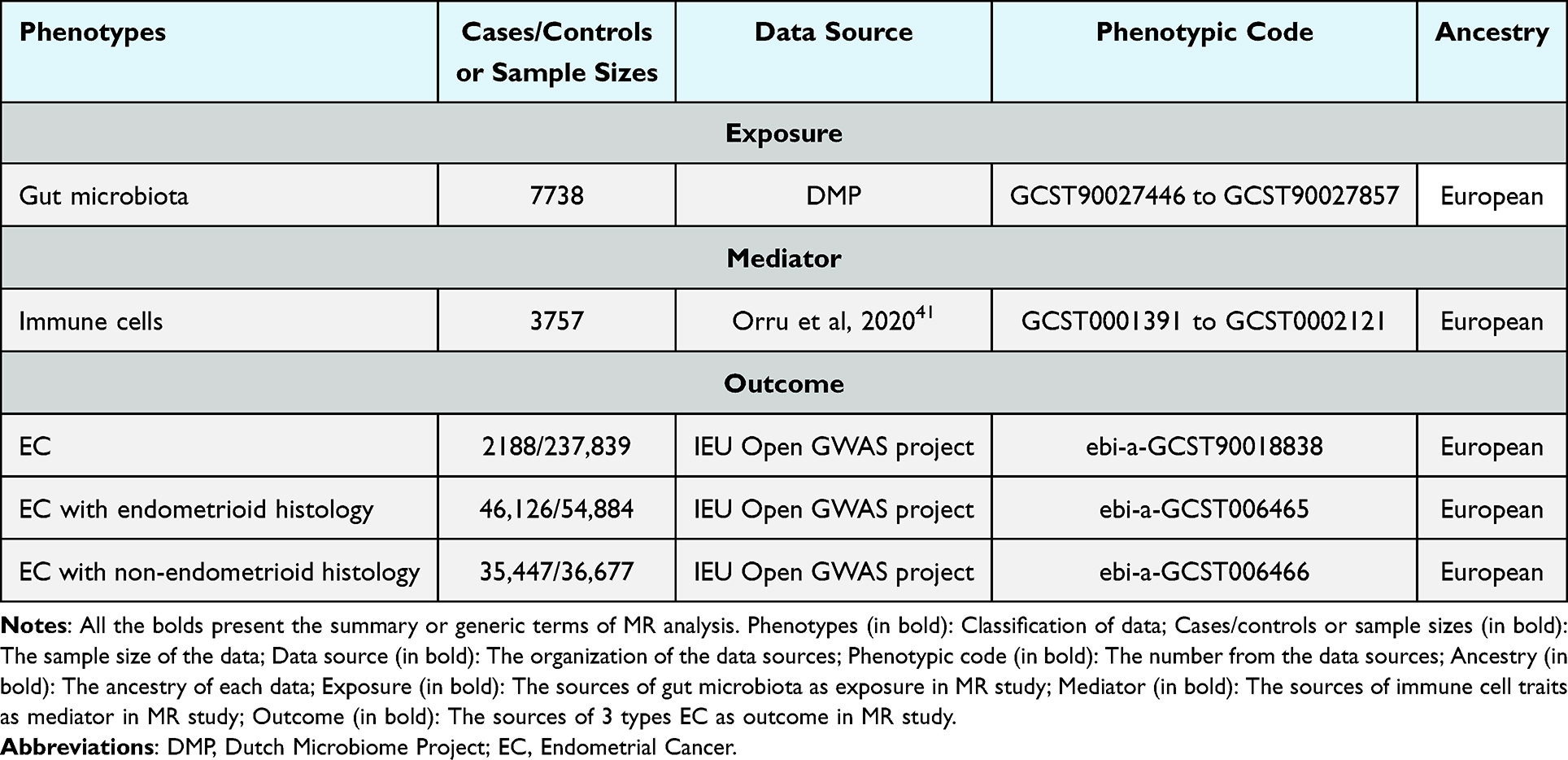 |
Table 1 Summary of Data Sources |
Genetic Instrumental Variable (IVs) Selection
Effective MR analysis relies on three foundations: IVs should (1) be strongly associated with EC, (2) affect EC only through the causal effect of GM, and (3) remove other potential confounders.38 The first step was to select SNPs associated with exposure with a significance threshold of 5e-08, which were further screened via linkage disequilibrium analysis (r2 < 0.001; distance within 10,000 kb).42 Finally, F values [F=β/r2 (square of the standard error)] were calculated to identify the strength of IV, retaining SNPs with an F value of >10.43 The study assumptions were presented in Figure 2, and selected SNPs would view in Table S2.
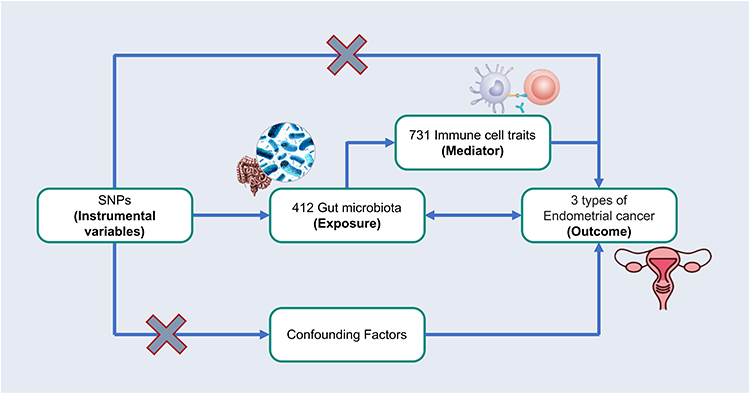 |
Figure 2 Assumptions of the two-sample UVMR and mediation analysis. SNPs, single nucleotide polymorphisms. All the bolds present the summary or generic terms of MR analysis. Instrumental variables (in bold): SNPs as the instrumental variables in MR study; Exposure (in bold): Gut microbiota as the exposure of MR study; Outcome (in bold): Endometrial cancer as the outcomes of MR study; Mediator (in bold): Immune cell traits as the mediators of MR study. |
Statistical Analysis
All statistical analyses were performed using the R 4.4.0 (https://www.r-project.org). The “VariantAnnotation” package, “ieugwasr” package, and “TwoSampleMR” package were used to perform UVMR. Among the five methods (“MR Egger,” “Weighted median,” “Inverse variance weighted (IVW),” “Simple mode,” and “Weighted mode”), IVW is the primary method of causal estimation; P < 0.05 was identified as a significant causal association. Moreover, heterogeneity and horizontal pleiotropy were based on the Cochran Q statistic and MR-Egger intercept of the IVW and MR-Egger methods. Heterogeneity and pleiotropy were identified by a p-value less than 0.05, which indicated unsustainable causality. The “leave-one-out” test was used to investigate the effects of possible uncorrelated SNPs.
Mediation Analysis
After a two-sample UVMR, GM and immune cells with significant causal relationships to various EC were selected for further mediation analysis. We determined whether the GM had causal effects on immune cells; if so, multiple MR analyses were conducted to explore the mediating effect of immune cells from GM to EC.
Results
Instrumental Variable Selection
Initially, 4031 GM-associated SNPs were identified by P < 1×10−5 with closely linked SNPs removed (single nucleotide polymorphisms), which served as the IV of 412 GM. R2 and F values were calculated for the screened SNPs (r2 < 0.001; F-values > 10), which were unlikely to be affected by weak instrument bias (Table S2). The same process was used to select SNPs of immune cell traits (Table S2).
Causal Effects of GM on Different Types of EC
The IVW method was used to evaluate the potential causal relationship between GM and the invasion of different EC (Figure 3B, D and F). Although other methods, including MRE, WMed, SMod, and WMod, did not show statistical significance, the estimated causal effect demonstrated a tendency similar to that obtained using the IVW method. There was no heterogeneity or horizontal pleiotropy in this MR analysis, as shown by Cochran’s Q statistic, MR-Egger intercept test, and MR-PRESSO test (Table S3). Furthermore, none of the SNP severely interfered with the overall effects of GM on EC.
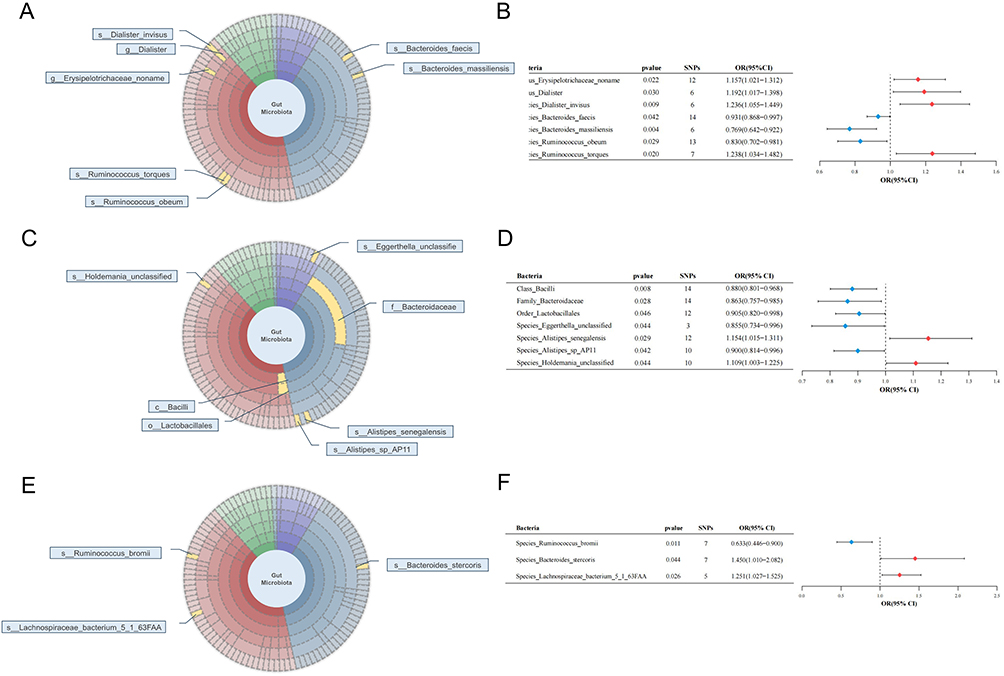 |
Figure 3 Various microorganisms of 412 GM presented causal relationships with 3 different types of EC. (A) MR result of GM on overall EC; (B) MR-IVW result of GM on overall EC; (C) MR result of GM on endometrioid EC (type I EC); (D) MR-IVW result of GM on endometrioid EC (type I EC); (E) MR result of GM on non-endometrioid EC (type II EC); (F) MR-IVW result of GM on non-endometrioid EC (type II EC). The symbol “c_/o_/f_/g_/s_” demonstrated the class, order, family, genus, and species. All the bolds present the summary or generic terms of MR analysis. Bacteria (in bold): the microbiota that can impact EC progression; pvalue (in bold): the p value of each bacterium on EC through MR analysis, which less than 0.05; SNPs (in bold): the significant SNPs used for MR analysis; OR (95% CI) (in bold): odds ratio with 95% confidence interval. Abbreviations: MR, mendelian randomization; GM, gut microbiota; EC, endometrial cancer; IVW, inverse variance weighted method; OR, odds ratio; 95% CI, 95% confidence interval. |
Overall EC
Genetic predictions of the genus Erysipelotrichaceae_noname (odds ratio (OR) = 1.157, 95% Confidence Interval (CI) [1.021, 1.316], P = 0.021), genus Dialister (OR = 1.192, 95% CI [1.017, 1.398], P = 0.030), species Dialister_invisus (OR = 1.236, 95% CI [1.055, 1.450], P = 0.009) and species Ruminococcus_torques (OR = 1.238, 95% CI [1.034, 1.482], P = 0.020) were associated with an increased risk of overall EC. Species Bacteroides faecis (OR = 0.931, 95% CI [0.868, 0.997], P = 0.042), species Bacteroides massiliensis (OR = 0.769, 95% CI [0.642, 0.922], P = 0.004), and species Ruminococcus obeum (OR = 0.830, 95% CI [0.702, 0.981], P = 0.029) were associated with a reduced risk of EC (Figure 3A and B).
Endometrioid EC (Type I EC)
For endometrioid EC, class Bacilli (OR = 0.880, 95% CI [0.801, 0.968], P = 0.008), family Bacteroidaceae (OR = 0.863, 95% CI [0.757, 0.985], P = 0.028), order Lactobacillales (OR = 0.905, 95% CI [0.820, 0.998], P = 0.046), species Eggerthella_unclassified (OR = 0.855, 95% CI [0.734, 0.996], P = 0.044), and species Alistipes_sp_AP11 (OR = 0.900, 95% CI [0.814, 0.996], P = 0.042) were protective factors for endometrioid EC. Species Aspergillus senegalensis (OR = 1.154, 95% CI [1.015, 1.311], P = 0.029) and species Holdemania_unclassified (OR = 1.109, 95% CI [1.003, 1.225], P = 0.044) were risk factors for endometrioid EC (Figure 3C and D).
Non-Endometrioid EC (Type II EC)
Focusing on non-endometrioid EC, species Bacteroides_stercoris (OR = 1.450, 95% CI [1.010, 2.082], P = 0.044) and species Lachnospiraceae_bacterium_5_1_63FAA (OR = 1.251, 95% CI [1.027, 1.525], P = 0.026) were associated with an increased risk of developing this type of EC, whereas species Ruminococcus_bromii (OR = 0.633, 95% CI [0.446, 0.900], P = 0.011) was associated with a decreased risk of EC (Figure 3E and F).
Reverse MR Analysis of GM on Different Types of EC
Next, a reverse MR analysis was conducted on the three EC subtypes and their causal GM. The results showed no obvious causal effects (p > 0.05, Table 2), except for species Lachnospiraceae_bacterium_5_1_63FAA, between the three EC types and the relevant GM using the IVW method of reverse analysis (Table 2). Therefore, species Lachnospiraceae_bacterium_5_1_63FAA should not be further analyzed in non-endometrioid EC.
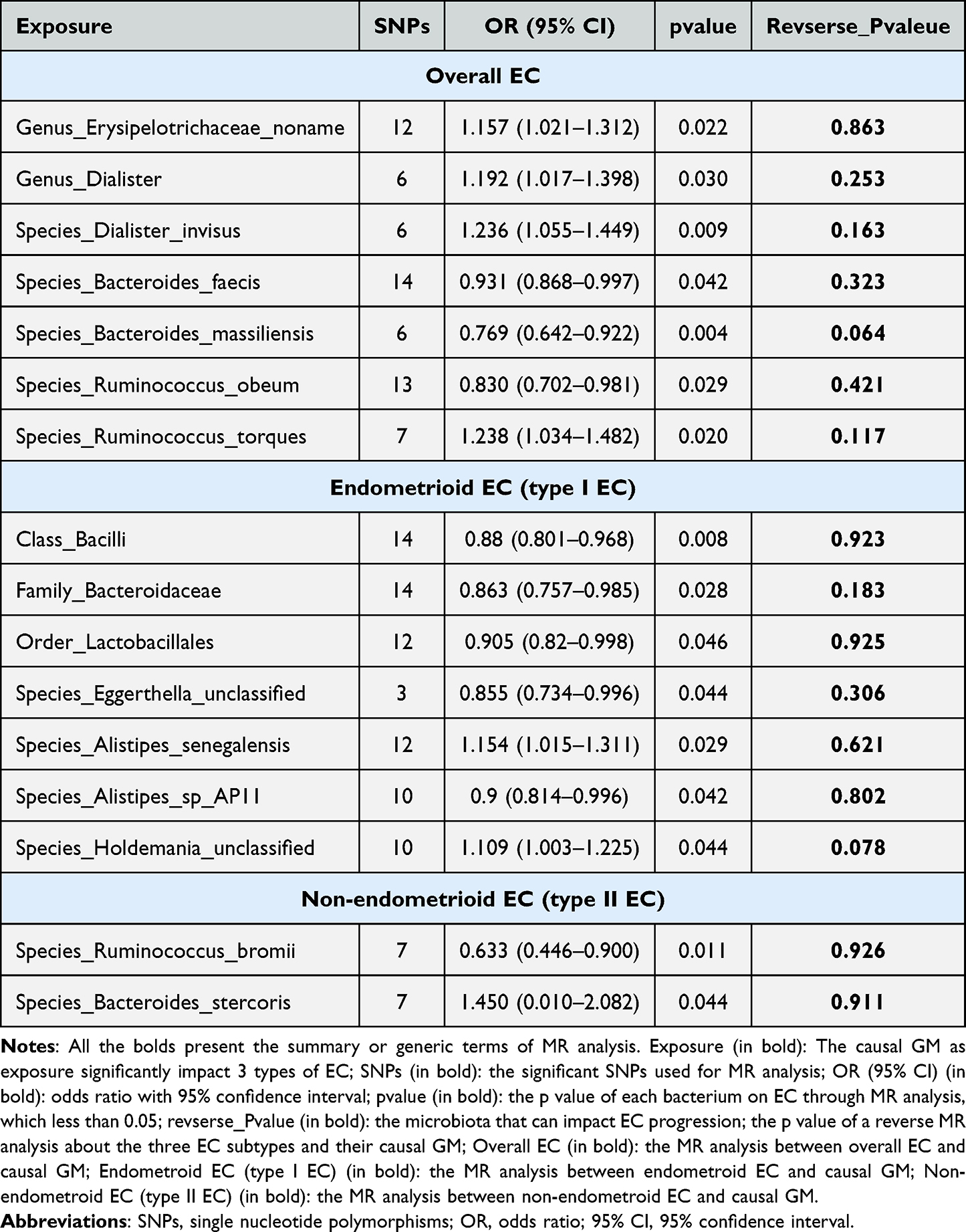 |
Table 2 Reverse MR_IVW Analysis of Gut Microbiota on 3 Types of EC |
Effect of Immune Cell Traits on Different Types of EC
The IVW method, as the primary assessment method, was used to explore the effect of immune cells on different types of EC, with causal effects similar to those of MRE, WMed, SMod, and WMod. In addition, the MR-Egger intercept test, MR-PRESSO test, and leave-one-out sensitivity analysis showed no heterogeneity or horizontal pleiotropy, and the removal of any single SNP did not significantly affect the overall influence of immune cells on EC (Table S3).
The analysis also revealed that protecting 6 immune cell traits and 14 genetically predicted immune cell traits enhanced the overall risk of EC (Figure 4A). Thirteen immune cell traits were associated with increased risk, whereas 14 traits were associated with reduced risk of endometrioid EC (Figure 4B). In addition, MR analysis showed that non-endometrioid EC were amplified by 19 immune cell traits and suppressed by 19 other immune cell traits (Figure 4C).
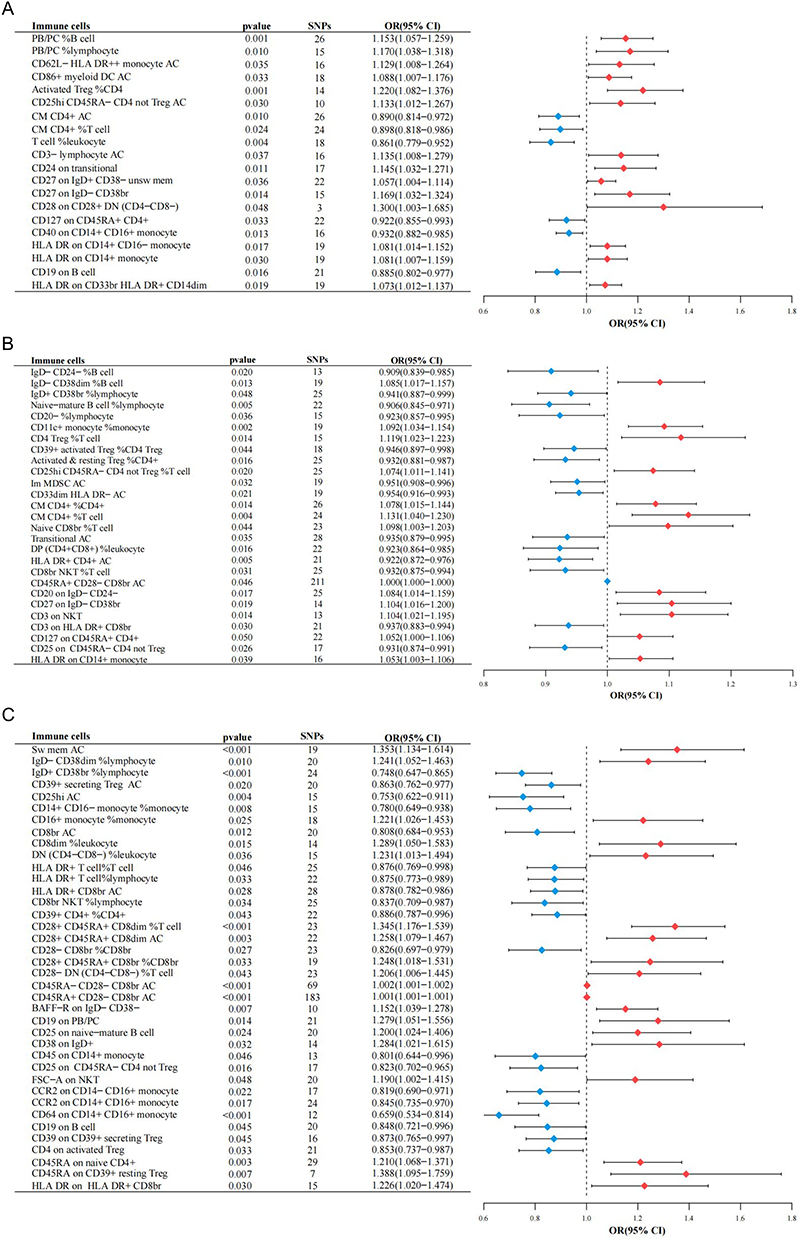 |
Figure 4 Various immune cell traits of 713 traits presented causal relationships with 3 different types of endometrioid cancer. (A) Results of MR with IVW method on causality between immune cells and overall EC. (B) Results of MR with IVW method on causality between immune cells and endometrioid EC. (C) Results of MR with IVW method on causality between immune cells and non-endometrioid EC. All the bolds present the summary or generic terms of MR analysis. Immune cells (in bold): the immune cell traits that can impact EC progression; pvalue (in bold): the p value of each bacterium on EC through MR analysis, which less than 0.05; SNPs (in bold): the significant SNPs used for MR analysis; OR (95% CI) (in bold): odds ratio with 95% confidence interval. Abbreviations: MR, mendelian randomization; EC, endometrioid cancer; IVW, inverse variance weighted method; AC, absolute cell counts; Treg, regulatory T cell; NKT, natural killer T cells; OR, odds ratio; 95% CI, 95% confidence interval. |
Effect of GM on Immune Cell Traits
We have demonstrated the role of GM and immune cell traits in various EC subtypes, and the causation of GM on immune cells in the three EC subtypes was explored through MR Analysis (Table S4).
Overall EC
Overall, 7 GM and 20 immune cell traits were analyzed, of which species Dialister_invisus was a risk factor for CD25hi CD45RA-CD4 but not for Treg AC (OR = 1.222, 95% CI [1.026, 1.456], P = 0.089). Species Bacteroides_massiliensis was protected against CD62L-HLA Dr + monocyte AC (OR = 0.761, 95% CI [0.617, 0.939], P = 0.011). In addition, species Ruminococcus_obeum was protective against CD86+ myeloid DC AC (OR = 0.817; 95% CI [0.687, 0.971], P = 0.222). However, it is also dangerous for CD27 on IgD+ CD38-unsw mem (OR = 1.356, 95% CI [1.081, 1.701], P = 0.008) and CD27 on IgD-CD38BR (OR = 1.222, 95% CI [1.036, 1.443], P = 0.018).
Endometrioid EC (Type I EC)
MR analysis revealed seven GM and 27 immune cell traits. The results showed that class Bacilli was harmful to IgD+ CD38br % lymphocytes (OR = 1.172, 95% CI [1.034, 1.327], P = 0.013) and Transitional AC (OR = 1.175, 95% CI [1.038, 1.329], P = 0.011), whereas species Holdemania_unclassified was also dangerous relative to Transitional AC (OR = 1.201, 95% CI [1.055, 1.369], P = 0.006), and order Lactobacillales was a risk factor for CD39+ activated Treg %CD4 Treg (OR = 1.121, 95% CI [1.007, 1.247], P = 0.036).
Non-Endometrioid EC (Type II EC)
For non-endometrioid EC with three GM and 38 immune cell traits, species Ruminococcus_bromii for IgD-CD38dim % lymphocytes (OR = 1.217, 95% CI [1.008, 1.468], P = 0.041), CD8dim % leukocytes (OR = 1.243, 95% CI [1.025, 1.508], P = 0.027), and CD8br NKT % lymphocytes (OR = 1.223, 95% CI [1.010, P = 0.027) 1.482], P = 0.039) were detrimental; however, they were protective against CD25hi AC (OR = 0.772, 95% CI [0.632, 0.943], P = 0.011).
Mediation Analysis
The proportion of mediating effects was quantified by determining the ratio of indirect to direct effects after confirming the cause-effect relationship between GM, immune cell traits, and various EC types (Table 3).
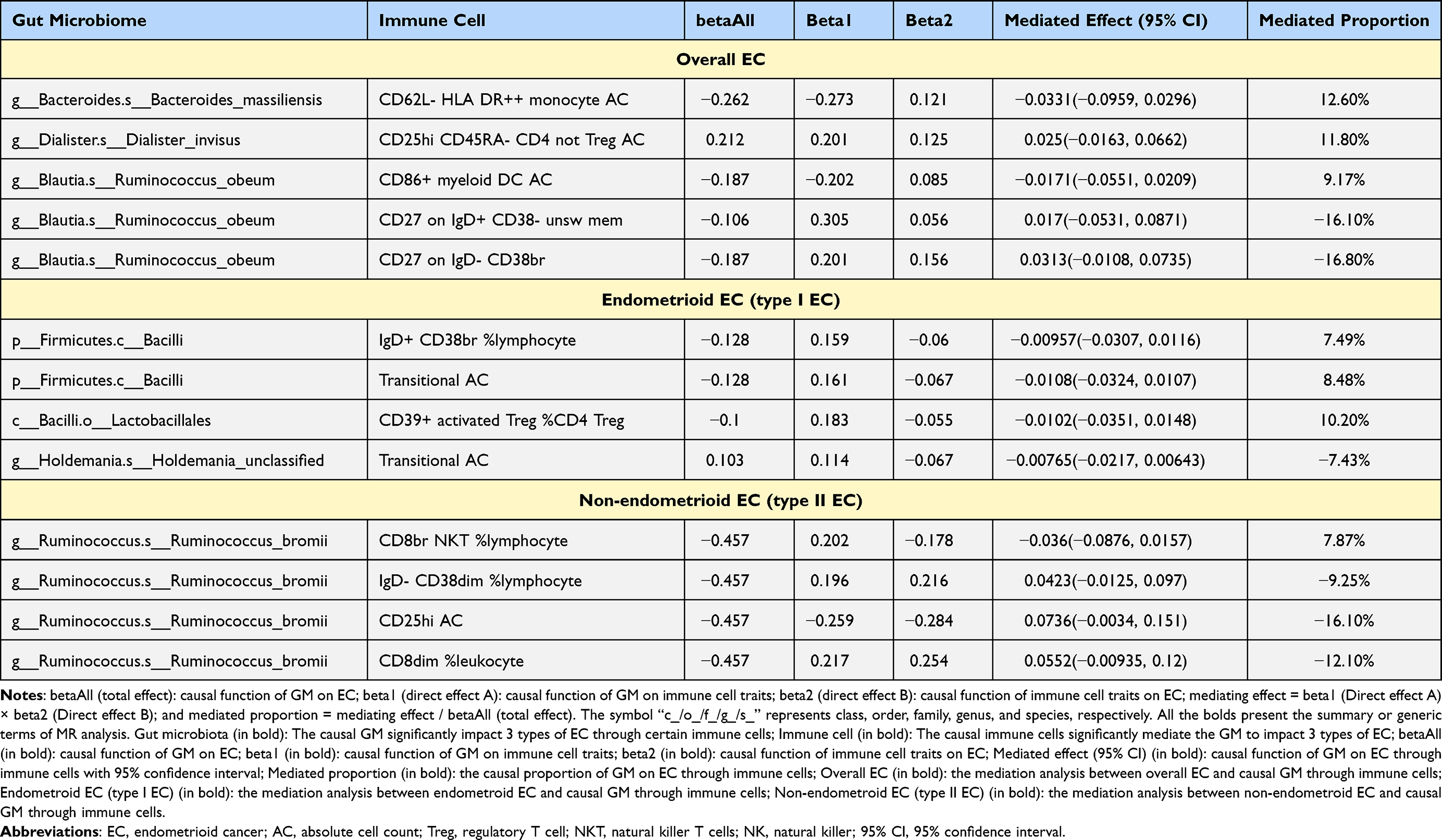 |
Table 3 Mediation Effect of Various GM on 3 Types of EC via Immune Cell Traits |
Overall EC
Species Ruminococcus_obeum affected EC through three different mediators: CD86+ myeloid DC AC (mediated effect β = −0.017, mediated proportion = 9.17%), CD27 on IgD+ CD38- unsw mem (β = 0.017, mediated proportion = −16.10%), and CD27 on IgD- CD38br (β = 0.031, mediated proportion = −16.80%) (Figure 5A). Notably, the mediated proportion of the mediator CD27 on IgD+ CD38- unsw mem and CD27 on IgD- CD38br was negative, indicating that exposure had the opposite effect on the outcome, as expected, through the mediator. This may be owing to 1) the reverse effect of the mediating variables. The mediating variable may have a reverse effect, and its regulation may lead to a reverse change in the outcome owing to various factors such as biological mechanisms, environmental factors, or individual differences. 2) Other unconsidered variables: Other unidentified variables may influence the mediating effect. In addition, statistical errors during calculations may have contributed to this result.
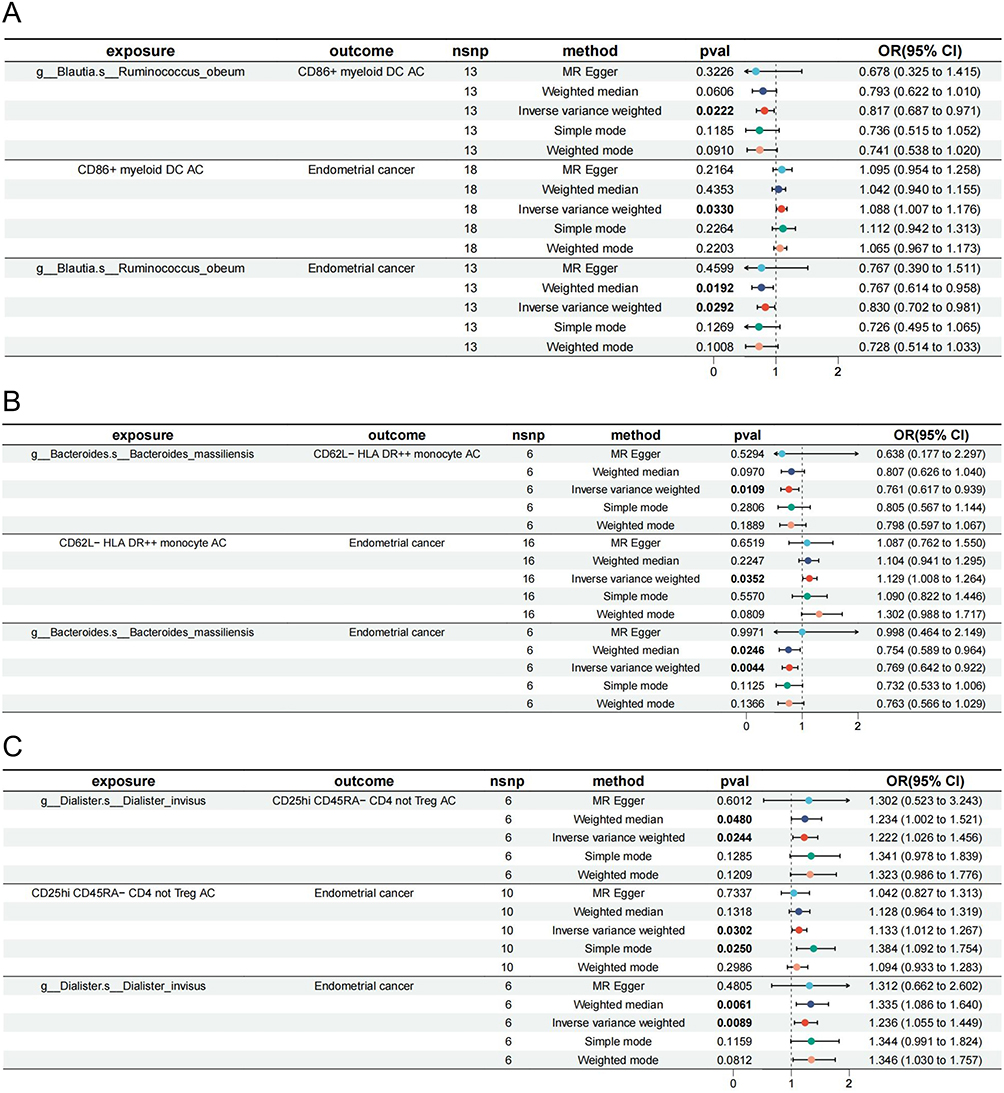 |
Figure 5 Mediation effect of various GM on EC with traditional definition via immune cell traits. (A) Mediation analysis of Dialister_invisus on EC via CD25hi CD45RA- CD4 not Treg AC. (B) Mediation analysis of Bacteroides_massiliensis on EC via CD62L- HLA DR++ monocyte AC. (C) Mediation analysis of Ruminococcus_obeum on EC via CD86+ myeloid DC AC. The symbol “c_/o_/f_/g_/s_” demonstrated the class, order, family, genus, and species. All the bolds present the summary or generic terms of MR analysis. Exposure (in bold): The exposures impact the outcomes in MR study; outcome (in bold): The outcomes impacted by exposures in MR study; nsnp (in bold): the significant SNPs used for MR analysis; method (in bold): The methods used for MR analysis; pval (in bold): the p value of each exposure on outcomes through 5 different methods, values less than 0.05 will be marked in bold; OR (95% CI) (in bold): odds ratio with 95% confidence interval. Abbreviations: EC, endometrioid cancer; AC, absolute cell counts; Treg, regulatory T cell; OR, odds ratio; 95% CI, 95% confidence interval. |
The results showed that species Bacteroides_massiliensis affected EC via CD62L- HLA DR++ monocyte AC (β = −0.033, mediated proportion = 12.60%) with a negative mediating role (Figure 5B). CD25hi CD45RA- CD4, not Treg AC, positively mediated species Dialister_invisus acting on EC (β = 0.025, mediated proportion = 11.80%) (Figure 5C).
Endometrioid EC (Type I EC)
The Class Bacilli protected endometrioid EC through IgD+ CD38br % lymphocytes (β = −0.010, mediated proportion = 7.49%) and transitional AC (β = −0.011, mediated proportion = 8.48%) with negative mediations (Figure 6A and B). Transitional AC also mediated the influence of species Holdemania_unclassified influence on endometrioid EC (β = −0.008, mediated proportion = −7.43%), which was illogical. In addition, the order Lactobacillales was protective against endometrioid EC by CD39+ activated Treg %CD4 Treg (β = −0.010, mediated proportion = 10.20%) (Figure 6C).
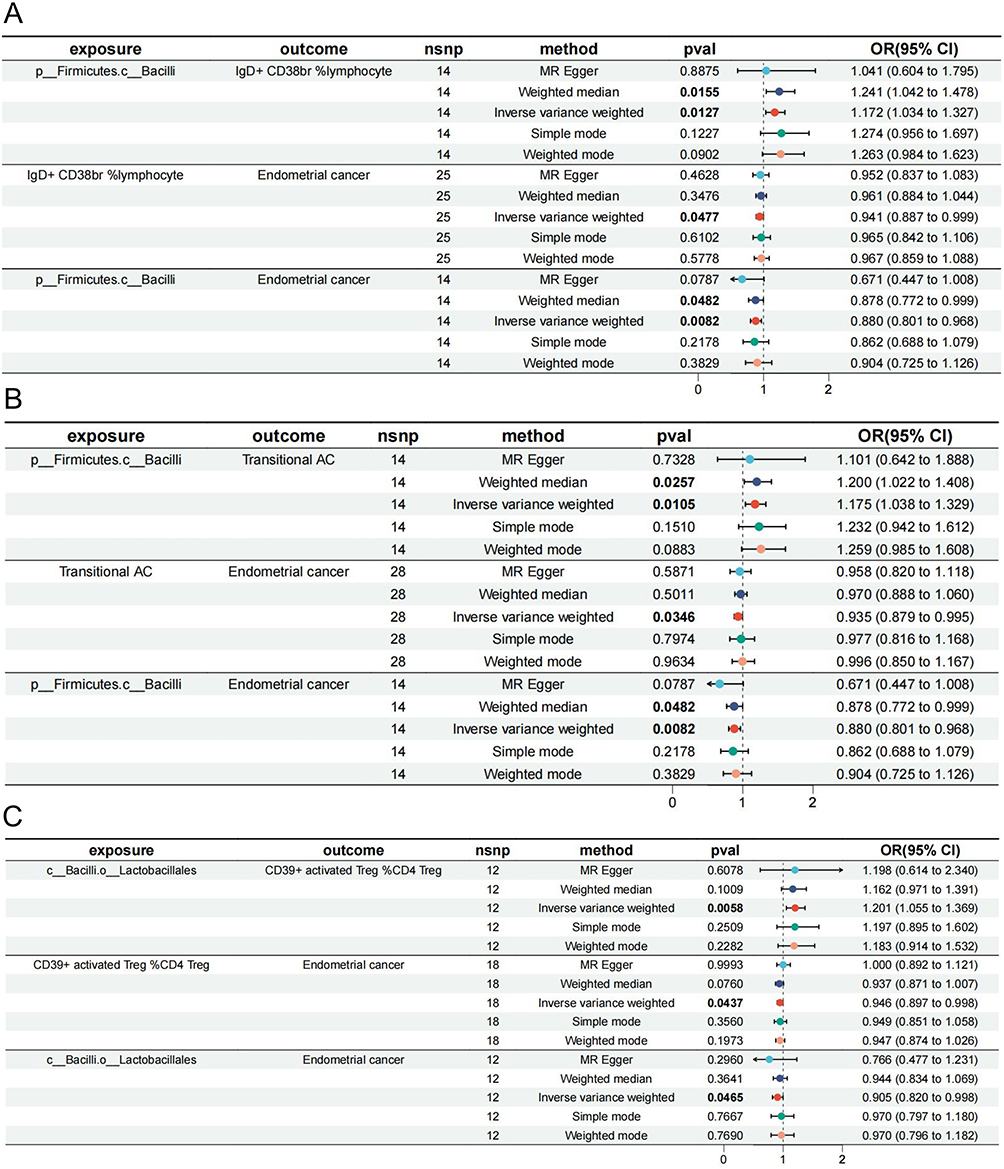 |
Figure 6 Mediation effect of various GM on EC with endometrioid histology via immune cell traits. (A) Mediation analysis of Bacilli on EC via IgD+ CD38br %lymphocyte. (B) Mediation analysis of Bacilli on EC via Transitional AC. (C) Mediation analysis of Lactobacillales on EC via CD39+ activated Treg %CD4 Treg. The symbol “c_/o_/f_/g_/s_” demonstrated the class, order, family, genus, and species. All the bolds present the summary or generic terms of MR analysis. Exposure (in bold): The exposures impact the outcomes in MR study; outcome (in bold): The outcomes impacted by exposures in MR study; nsnp (in bold): the significant SNPs used for MR analysis; method (in bold): The methods used for MR analysis; pval (in bold): the p value of each exposure on outcomes through 5 different methods, values less than 0.05 will be marked in bold; OR (95% CI) (in bold): odds ratio with 95% confidence interval. Abbreviations: EC, endometrioid cancer; AC, absolute cell counts; Treg, regulatory T cell; OR, odds ratio; 95% CI, 95% confidence interval. |
Non-Endometrioid EC (Type II EC)
Species Ruminococcus_bromii reduced the risk of non-endometrioid EC by IgD- CD38dim %lymphocyte (β = 0.042, mediated proportion = −9.25%), CD25hi AC (β = 0.074, mediated proportion = −16.10%), CD8dim % leukocytes (β = 0.055, mediated proportion = −12.10%), and CD8br NKT % lymphocytes (β = −0.036, mediated proportion = 7.87%), although only CD8br NKT % lymphocytes were logical (Figure 7).
 |
Figure 7 Mediation effect of Ruminococcus_bromii on EC with non-endometrioid histology via CD8br NKT %lymphocyte. The symbol “c_/o_/f_/g_/s_” demonstrated the class, order, family, genus, and species. All the bolds present the summary or generic terms of MR analysis. Exposure (in bold): The exposures impact the outcomes in MR study; outcome (in bold): The outcomes impacted by exposures in MR study; nsnp (in bold): the significant SNPs used for MR analysis; method (in bold): The methods used for MR analysis; pval (in bold): the p value of each exposure on outcomes through 5 different methods, values less than 0.05 will be marked in bold; OR (95% CI) (in bold): odds ratio with 95% confidence interval. Abbreviations: EC, endometrioid cancer; AC, absolute cell counts; NKT, natural killer T cells; OR, odds ratio; 95% CI, 95% confidence interval. |
Discussion
Intestinal microbiota and its metabolites affect several pathophysiological processes covering host metabolism and immune response, known as the “second endocrine organ”.44 Previous studies have verified that the regulation of host immunity is closely interrelated with the microbiota, and a disrupted balance would motivate the occurrence and deterioration of diseases;45,46 hence, it is crucial to equilibrate the complex communions between them. In this study, we conducted a comprehensive large-scale two-sample MR analysis based on the LifeLine Biobank and GWAS databases, followed by reverse MR analysis for each of the three types of EC. There were 17 causal relationships, including 7 GM species and overall EC, 7 species and endometrioid EC, and 3 species and non-endometrioid EC, which were evaluated by predicting 158 SNP loci. Therefore, the three EC subtypes were primarily causally related to the phyla Firmicutes and Bacteroidetes, and type I EC presented a causal correlation with the Phylum Actinobacteria. This indicates that there are differences in the microbiome-associated profiles among different EC subtypes. Type I (estrogen-related, highly differentiated endometrial adenocarcinoma) may be influenced by specific Actinobacteria, whereas type II (non-endometrial, high-grade cancer) does not show such a relationship.
Next, a two-sample MR analysis was conducted on immune cell traits and different types of EC, and causal relationships based on various SNPs are discussed. Twenty immune traits were associated with typical EC, 27 were causally linked to endometrioid EC, and non-endometrioid EC showed a causal correlation with 38 immune cells, further demonstrating the heterogeneity of the immune microenvironment among the different EC subtypes.
Finally, through two-step MR mediation analysis, we identified seven significant “GM-immune-EC” causal chains and quantified the proportion of the immune mediation effect in the total effect. These results support the hypothesis from a genetic perspective that GM may affect EC occurrence by regulating the immune system.
Notably, types I and II may have unique pathways in their pathogenic mechanisms. Owing to the differences in hormone dependence and molecular characteristics between type I and type II EC, it is impossible to present a common “ GM-immune-EC” pathway for both subtypes. The partially common associations observed in this study were more likely the result of indirect immune regulation. Therefore, caution should be exercised when discussing these results, emphasizing that this study is a hypothesis generating exploration. The causal inferences from this study require further biological validation and in-depth mechanistic exploration. Moreover, targeted research and clinical studies should be conducted on different EC subtypes with different pathologies to clarify their GM and immune mechanisms.2
Overall EC
Bacteroides massiliensis can reduce the risk of EC by repressing the immune cell trait “CD62L HLA DR++ monocyte AC”. Bacteroides regulate the immune system to maintain homeostasis, and their metabolites maintain immune system stability.47 Bacteroides are the major producers of short-chain fatty acids (SCFAs) in the gut,48 which are important for maintaining microecological equilibrium, mainly in the form of acetic and propionic acids. Acetate and propionate are potent anti-inflammatory mediators that inhibit the release of pro-inflammatory cytokines from neutrophils and macrophages.49 The anticancer effects of propionic acid on apoptosis have been described in human colon cancer cells.50 The anti-tumor effects of cytotoxic T lymphocyte antigen 4 (CTLA-4) blockers depend on different Bacteroides species, with specific T cell responses to B. thetaiotaomicron or B. Fragilis affecting the efficacy of CTLA-4 blockers in mice and patients.51 In addition, the HLA-DR phenotype is closely related to anti-tumor immunotherapy, with CD62L as a marker of monocyte activation,52,53 and CD62L HLA DR++ monocyte were analyzed to facilitate EC progression. Nevertheless, the immunosuppressive cytokines interleukin (IL)-10 and pro-inflammatory factors IL-6 and TNFα are negatively correlated with HLA-DR in B-cell non-Hodgkin lymphoma.54,55 Mengos et al found that monocytes with reduced or no HLA-DR expression are crucial mediators of tumor-induced immunosuppression and negatively affect programmed death-1 (PD-1) and CTLA-4 checkpoint inhibition,56–62 chimeric antigen receptor T-cell (CAR-T) immunotherapy,53–65 cancer vaccines,66–70 and hematopoietic stem cell transplantation.71–74 Previous studies have presented conflicting views on our analysis; therefore, further exploration is necessary to investigate the effect of CD62L-HLA DR++ monocytes on EC.
Ruminococcus obeum, which belongs to the genus Blautia, protects EC via the trait “CD86+ myeloid DC AC”. GM analysis of immune checkpoint inhibitor (ICI)-treated patients showed that the high diversity and presence of immunogenic bacteria, such as Ruminococcus, resulted in more significant CD8+ T cells and CD4+ Th1-dependent anti-tumor responses, leading to better outcomes.31,32,75,76 However, Xu et al found that Blautia obeum was enriched in patients who were nonreactive to anti-PD-1 and ICI treatments and may have antibiotic properties along with antibiotic resistance.77,78 Therefore, the distinct mechanism of action of Ruminococcus obeum on EC warrants further investigation. In addition, CD86 expression is considered a poor prognostic indicator and is down-regulated by transendocytosis of CTLA-4.78,79 Blocking CD28: CD80/CD86 in vivo re-sensitizes multiple myeloma cells to chemotherapy and significantly reduces the tumor load.80 Combined with MR analysis, Ruminococcus obeum may inhibit CD86+ on myeloid DC to enhance immunotherapy, thereby restraining EC.
Dialister invisus is a risk factor for EC functioning through the immune cell trait “CD25hi CD45RA- CD4, not Treg AC.” Enriching Dialister invisus, isolated from the human oral cavity, indicates a high risk and rapid progression of tumors in colorectal cancer and increases the risk of HPV-infected cervical cancer in female reproductive system tumors.81–85 However, Byrd et al found that dialister status was associated with higher survival in MSI-H colorectal tumors.86 CD4+ CD25hi T cells maintain immune tolerance to autoantigens in various cancers, including ovarian and cervical.87–89 Typically, T cells can be divided into CD45RA+ initial and CD45RA- memory subtypes. Activated CD45RA- T cells inhibit the anti-tumor function of CD8+ T cells via IL-10 secretion and intercellular contacts. This facilitates immunosuppression and gastric cancer progression.90–92 Tassi et al confirmed that CD45RA-T lymphocytes were increased in the epithelial ovarian tumor microenvironment.93 Therefore, together with our analysis, Dialister invisus down-regulates the anti-tumor function of CD8+ T cells by stimulating the immunosuppressive trait “CD25hi CD45RA-CD4 not Treg,” following accelerated tumor progression of EC.
Endometrioid EC (Type I EC)
Class Bacilli play a protective role in endometrioid EC by colonizing the gastrointestinal tract.94 The regulates tumor pathophysiological processes, and their secretions exhibit bacteriological and anti-tumor properties. Typically, SCFAs from Bacilli are beneficial for gut peristalsis and secretion and inhibit tumor proliferation by inducing apoptosis and controlling epigenetic modification.94–98 Bacteriocins and other secretions derived from bacilli can suppress phospholipase A2, downregulate pro-inflammatory cytokines, and upregulate anti-inflammatory cytokines.99,100 Extracellular vesicles (EVs) of bacilli induce apoptosis in HepG2 cells by increasing the expression ratio of bax/bcl-2.101 Bacilli stimulate the production of insulin-like growth factor 1 (IGF-1), participates in the regulation of blood glucose and lipids, and is closely associated with the pathogenesis of type I EC.97 In a clinical study on colorectal cancer, the abundance of bacilli in the metastatic group was significantly decreased compared with that in tumor patients,102 indicating that it may restrain tumor invasion. CD38, a type II transmembrane glycoprotein, is a crucial metabolic enzyme located on the cell surface and its products are vital during immune regulation.103,104 CD38 is an anti-tumor therapeutic target involved in adenosine formation and exerts remarkable immunosuppressive effects on the solid tumor microenvironment.103,105 Thavaneswaran et al reported that patients with EC with high CD38 + expression in peripheral tertiary lymphoid structures (TLSs) had favorable survival outcomes during chemotherapy.106,107 Our MR analysis showed that lymphocytes with lgD+ and bright CD38 are protective against endometrioid EC. Bacilli play a protective role by enhancing lymphocyte function. However, their specific mechanisms require further investigation.
In particular, the order Lactobacillales belongs to the Class Bacilli; hence, its metabolites and secretions, such as SCFAs, exhibit similar antitumor mechanisms. Bactericin mainly disrupts the membrane intimal potential, leading to uncontrolled ion leakage and cell death.108,109 Moreover, these bacteria can reduce the concentration of soluble bile salts in faeces, thereby neutralizing the cancer-promoting effects of bile acids, including DNA damage and apoptosis.109–112 Importantly, the Lactobacillales is the main component of a healthy vaginal microecosystem, and its disturbance promotes inflammation and cancer progression.113–115 In contrast, CD39+-activated Treg %CD4 Treg mainly play an anti-inflammatory role. CD39, an extranuclear triphosphate diphosphate hydrolase-1, can hydrolyze ATP into AMP, obstructing inflammatory signal transduction of extracellular ATP.116–118 Maddaloni et al showed that lactic acid bacteria can deliver IL-35 in collagen-induced arthritis in mice, thus stimulating CD39+ CD4+ Tregs to release increased IL-10, exerting anti-inflammatory and immune effects.118 Furthermore, lactic acid bacteria increase the proliferation of CD39+ Tregs in mouse allergic asthma models, following the regulation of the inflammatory response induced by immune disorders.119 Therefore, we speculate that Lactobacillales activates and promotes the proliferation of CD39+-activated Treg %CD4 Tregs by delivering special inflammatory mediators, such as IL-35, in endometrioid EC, indicating their protective role.
Non-Endometrioid EC (Type II EC)
Ruminococcus bromii is beneficial for patients with non-endometrioid EC. A colon cancer study proposed the microbiota characteristics of R. bromii as a reliable indicator, combined with the immune rejection constant, to create a score for evaluating survival prognosis, which contributed to the discovery of personalized therapy.29 Notably, colon cancer has a genetic phenotype similar to that of non-endometrioid EC: DNA mismatch repair (MMR) defects or microsatellite instability (MSI).120 The increased abundance of R. bromii following symbiotic treatment (MS-20) combined with anti-PD-1 treatment positively correlated with reduced tumor load and CD8+ T cell infiltration in xenograft mouse models.121 Additionally, rumen cocci can bind to castalagin and promote anti-cancer responses; castalagin rich in R. bromii influences the efficacy of anti-PD-L1 immunotherapy and increases the ratio of CD8 + / FOXP3 + CD4 + in the tumor microenvironment.122 NKT cells specifically assemble T-cell receptor (TCR) and natural killer (NK) cell receptor on the surface, which produce numerous cytokines and play a similar cytotoxic role as NK cells.123–128 This enhances the immune response and mediates the inhibition of tumor growth in various cancers, including liver, ovarian, and colon cancers.30,129–132 In mismatch repair-deficient EC, a large infiltration of CD8+ and NKT cells is a marker for better survival prognosis, indicating a favorable immune microenvironment.132,133 Therefore, combined with the MR analysis, R. bromii may improve the tumor immune response and the efficacy of immunotherapy by increasing CD8+ NKT cells, thus playing a protective role. Recently, universal CAR-engineered NKT (UCAR-NKT) cells were developed and demonstrated powerful anti-tumor efficacy against blood and solid tumors in vitro and in vivo, with multiple tumor-targeting mechanisms.134,135 These cells regulate the tumor microenvironment by selectively depleting immunosuppressive macrophages. Therefore, based on this analysis and previous research, targeting R. bromii and the immune microenvironment may help improve the prognosis of non-endometrioid EC.
Clinical Implication
However, it is necessary to consider whether the alteration of the gut microbiota actively drive the EC progression, or instead present as a consequence of the EC presence.136 If specific microbial dysbiosis is the cause of the EC development, the microbiota can serve as a therapeutic target. For instance, interventions including probiotics and diet could be attempted to prevent the progression of EC.136,137 On the other hand, if the tumors induce the observed microbiota changes, the gut microbiota may tend to serve as a diagnostic or prognostic marker for the disease state, rather than a treatment therapy. Our bidirectional MR results support the former point, we found non-significant evidence of reverse causality from EC to microbiota, while significant causal effects were shown from gut microbiota to EC. It indicated that gut microbiota perform a directional impact on the development of EC, rather than a resulting phenomenon. And other recent studies revealed that altering the unfavorable microbiota can affect tumor outcomes.122,138 Therefore, the gut microbiota is not only a biomarker, also a potential therapeutic target for EC, which need experimental supports.
Besides, our analysis emphasized the gut microbiota-immune axis as a promising new therapy or biomarker for improving the diagnosis and risk forecast of EC. Firstly, from the perspective of prevention strategies, many high-risk factors for EC are closely related to intestinal microbiota, such as obesity and metabolic disorders. Hence, the gut microbiota regulation may become a new therapy to reducing the EC risk. For example, correcting the microbiota imbalance through dietary intervention and probiotics, is expected to improve the pro-tumor inflammation and metabolism, thereby preventing especially type I EC.139–141 Secondly, in terms of immunotherapy, the immunogenicity of EC especially type II suggests that patients may benefit from immune checkpoint inhibitors (ICB), and our results further supported that the gut microbiota may affect the efficacy of immunotherapy.139 Studies have shown that patients with different compositions of the gut microbiota respond significantly differently to immunotherapy. Therefore, it is prospective to incorporate microecological regulation into EC treatment. And it is effective to enhance the efficacy of immunotherapy in patients with advanced or recurrent EC, combining with probiotic preparations or fecal microbiota transplantation (FMT) to improve the microbiota composition.139 At the same time, the gut microbiota and its metabolites have the potential to serve as biomarkers for predicting the efficacy of immunotherapy in EC patients.140,142,143 In summary, the gut microbiota-immune axis provides a new perspective for the prevention and treatment of EC, microecological interventions can both regulate the EC progression and improve the immune status to enhance the therapeutic effect.143
Strengths of This Study
The advantage of this study lies in the first application of a comprehensive causal mediation analysis of the EC subtypes. It systematically integrated bidirectional two-sample MR and mediation analyses to explore the potential causality between GM and EC mediated by immune cells. We performed comparative analyses for overall, type I, and type II EC, revealing similarities and differences in the risks of GM and immune-mediated patterns among the different subtypes. Moreover, our study utilized large-scale GWAS data and a strict two-sample MR design to identify multiple robust causal associations between microbiota, immunity, and EC, and excluded major confounding factors through multiple sensitivity tests. These findings provide new insights into the biological basis of EC heterogeneity and support evidence for the development of treatments targeting the GM and the immune microenvironment.
Limitations of This Study
This study has some limitations. First, the data were derived mainly from large-scale European population databases. The homogeneity of race and region may have limited the results to other populations. Second, EC occurrence and development are the outcomes of the combined effects of multiple factors. Our genetic instrumental variable analysis cannot consider the influence of non-genetic factors such as the environment and lifestyle. In addition, causal inference in the MR analysis relies on this hypothesis. Although we controlled for horizontal pleiotropy, unobserved biases may still be present. Finally, our inference lacks in vivo and in vitro verification and the mechanistic explanation remains speculative. Further research should combine clinical cohorts and functional experiments to confirm and deepen our understanding of microbiota-immune interactions, especially for different EC subtypes.
Conclusion
Through a two-step MR and mediation analysis, our findings provide robust evidence that immune traits act as significant mediators in connecting the causal pathway of GM to EC development. Importantly, this study highlights distinct causal relationships and immune-mediated mechanisms across the three major EC subtypes (overall, endometrioid, and non-endometrioid). These subtype-specific insights into the gut-immune-cancer axis provide novel perspectives for developing therapeutic strategies targeting the GM and immune microenvironment in different forms of EC.
Abbreviations
EC, Endometrioid cancer; GWAS, Genome-wide association studies; DMP, Dutch Microbiome Project; ER, Estrogen receptor; PR, Progesterone receptor; GM, Gut microbiota; MR, Mendelian randomization; SNPs, Single nucleotide polymorphisms; UVMR, Univariable Mendelian randomization; IVs, instrumental variables; IVW, Inverse variance weighted; OR, Odds ratio; CI, Confidence Interval; AC, Absolute cell counts; Treg, Regulatory T cell; NKT, Natural killer T cells; CTLA-4, Cytotoxic T lymphocyte antigen 4; IL-10, Interleukin 10; PD-1, Programmed death-1; ICI, Immune checkpoint inhibitors; CAR-T, chimeric antigen receptor T-cell; SCFAs, Short-chain fatty acids; EVs, Extracellular vesicles; IGF-1, Insulin-like growth factor 1; TLSs, Tertiary lymphoid structures; MMR, Mismatch repair; MSI, Microsatellite instability; TCR, T-cell receptor; FMT, fecal microbiota transplantation.
Data Sharing Statement
The datasets used in this study can be found in the NHGRI-EBI GWAS Catalog (https://www.ebi.ac.uk/gwas/). The names and numbers of the datasets used are listed in Table 1.
Ethics Approval and Consent to Participate
This research has been conducted relied on publicly available de-identification data from participant studies authorized by the Ethical Standards Committee. All original studies were approved by the corresponding ethical review board, and informed consent was obtained from all the participants. In addition, no individual-level data were used in this study. Therefore, no new ethical review board approval was required.
Acknowledgments
The authors appreciate the Dutch Microbiome Project for releasing the GM-related GWAS summary data. The authors also wish to acknowledge the participants and investigators of the IEU Open GWAS project and the immune cell study by Orru et al.41 This paper has been uploaded to ResearchSquare as a preprint: https://www.researchsquare.com/article/rs-6189084/v1.
Funding
The authors declare that they received financial support for the research, authorship, and/or publication of this article. This research was funded by the National Key Research and Development Plan (No. 2023YFC2705400), National Natural Science Foundation of China (82472965), and Wuhan Knowledge Innovation Special Project (202202080101045).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–22. doi:10.3322/caac.21660
2. Makker V, MacKay H, Ray-Coquard I, et al. Endometrial cancer. Nat Rev Dis Primers. 2021;7(1):88. doi:10.1038/s41572-021-00324-8
3. Bokhman JV. Two pathogenetic types of endometrial carcinoma. Gynecol Oncol. 1983;15(1):10–17.
4. Brinton LA, Felix AS, McMeekin DS, et al. Etiologic heterogeneity in endometrial cancer: evidence from a Gynecologic Oncology Group trial. Gynecol Oncol. 2013;129(2):277–284. doi:10.1016/j.ygyno.2013.02.023
5. Łaniewski P, Ilhan ZE, Herbst-Kralovetz MM. The microbiome and gynaecological cancer development, prevention and therapy. Nat Rev Urol. 2020;17(4):232–250. doi:10.1038/s41585-020-0286-z
6. Baker JM, Al-Nakkash L, Herbst-Kralovetz MM. Estrogen-gut microbiome axis: physiological and clinical implications. Maturitas. 2017;103:45–53. doi:10.1016/j.maturitas.2017.06.025
7. Sobstyl M, Brecht P, Sobstyl A, Mertowska P, Grywalska E. The Role of Microbiota in the Immunopathogenesis of Endometrial Cancer. Int J Mol Sci. 2022;23(10). doi:10.3390/ijms23105756
8. Suemori T, Susumu N, Iwata T, et al. Intratumoral CD8+ lymphocyte infiltration as a prognostic factor and its relationship with cyclooxygenase 2 expression and microsatellite instability in endometrial cancer. Int J Gynecol Cancer. 2015;25(7):1165–1172. doi:10.1097/IGC.0000000000000482
9. Pakish JB, Zhang Q, Chen Z, et al. Immune microenvironment in microsatellite-instable endometrial cancers: hereditary or sporadic origin matters. Clin Cancer Res. 2017;23(15):4473–4481. doi:10.1158/1078-0432.CCR-16-2655
10. Zhang P, Liu M, Cui Y, Zheng P, Liu Y. Microsatellite instability status differentially associates with intratumoral immune microenvironment in human cancers. Brief Bioinform. 2021;22(3). doi:10.1093/bib/bbaa180
11. Bu L, Baba H, Yoshida N, et al. Biological heterogeneity and versatility of cancer-associated fibroblasts in the tumor microenvironment. Oncogene. 2019;38(25):4887–4901. doi:10.1038/s41388-019-0765-y
12. Pitt JM, Marabelle A, Eggermont A, Soria JC, Kroemer G, Zitvogel L. Targeting the tumor microenvironment: removing obstruction to anticancer immune responses and immunotherapy. Ann Oncol. 2016;27(8):1482–1492. doi:10.1093/annonc/mdw168
13. Xiao Y, Yu D. Tumor microenvironment as a therapeutic target in cancer. Pharmacol Ther. 2021;221:107753. doi:10.1016/j.pharmthera.2020.107753
14. O’Malley DM, Bariani GM, Cassier PA, et al. Pembrolizumab in patients with microsatellite instability-high advanced endometrial cancer: results from the KEYNOTE-158 study. J Clin Oncol. 2022;40(7):752–761. doi:10.1200/JCO.21.01874
15. Dey DK, Krause D, Rai R, Choudhary S, Dockery LE, Chandra V. The role and participation of immune cells in the endometrial tumor microenvironment. Pharmacol Ther. 2023;251:108526. doi:10.1016/j.pharmthera.2023.108526
16. Eskander RN, Sill MW, Beffa L, et al. Pembrolizumab plus chemotherapy in advanced endometrial cancer. N Engl J Med. 2023;388(23):2159–2170. doi:10.1056/NEJMoa2302312
17. Mirza MR, Chase DM, Slomovitz BM, et al. Dostarlimab for primary advanced or recurrent endometrial cancer. N Engl J Med. 2023;388(23):2145–2158. doi:10.1056/NEJMoa2216334
18. Gopalakrishnan V, Helmink BA, Spencer CN, Reuben A, Wargo JA. The influence of the gut microbiome on cancer, immunity, and cancer immunotherapy. Cancer Cell. 2018;33(4):570–580. doi:10.1016/j.ccell.2018.03.015
19. Sender R, Fuchs S, Milo R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 2016;14(8):e1002533. doi:10.1371/journal.pbio.1002533
20. Morgan XC, Huttenhower C. Chapter 12: human microbiome analysis. PLoS Comput Biol. 2012;8(12):e1002808. doi:10.1371/journal.pcbi.1002808
21. Honda K, Littman DR. The microbiota in adaptive immune homeostasis and disease. Nature. 2016;535(7610):75–84. doi:10.1038/nature18848
22. Patterson E, Ryan PM, Cryan JF, et al. Gut microbiota, obesity and diabetes. Postgrad Med J. 2016;92(1087):286–300. doi:10.1136/postgradmedj-2015-133285
23. Wang L, Christophersen CT, Sorich MJ, Gerber JP, Angley MT, Conlon MA. Low relative abundances of the mucolytic bacterium Akkermansia muciniphila and Bifidobacterium spp. in feces of children with autism. Appl Environ Microbiol. 2011;77(18):6718–6721. doi:10.1128/AEM.05212-11
24. De Luca F, Shoenfeld Y. The microbiome in autoimmune diseases. Clin Exp Immunol. 2019;195(1):74–85. doi:10.1111/cei.13158
25. Mager LF, Burkhard R, Pett N, et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science. 2020;369(6510):1481–1489. doi:10.1126/science.abc3421
26. Matson V, Chervin CS, Gajewski TF. Cancer and the microbiome-influence of the commensal microbiota on cancer, immune responses, and immunotherapy. Gastroenterology. 2021;160(2):600–613. doi:10.1053/j.gastro.2020.11.041
27. Soares PMG, Mota JMSC, Souza EP, et al. Inflammatory intestinal damage induced by 5-fluorouracil requires IL-4. Cytokine. 2013;61(1):46–49. doi:10.1016/j.cyto.2012.10.003
28. Frosali S, Pagliari D, Gambassi G, Landolfi R, Pandolfi F, Cianci R. How the intricate interaction among toll-like receptors, microbiota, and intestinal immunity can influence gastrointestinal pathology. J Immunol Res. 2015;2015:489821. doi:10.1155/2015/489821
29. Roelands J, Kuppen PJK, Ahmed EI, et al. An integrated tumor, immune and microbiome atlas of colon cancer. Nat Med. 2023;29(5):1273–1286. doi:10.1038/s41591-023-02324-5
30. Ma C, Han M, Heinrich B, et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science. 2018;360(6391). doi:10.1126/science.aan5931
31. Routy B, Le Chatelier E, Derosa L, et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science. 2018;359(6371):91–97. doi:10.1126/science.aan3706
32. Gopalakrishnan V, Spencer CN, Nezi L, et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science. 2018;359(6371). doi:10.1126/science.aan4236
33. Smith GD, Ebrahim S. “Mendelian randomization”: can genetic epidemiology contribute to understanding environmental determinants of disease? Int J Epidemiol. 2003;32(1): 1–22.
34. Lawlor DA, Harbord RM, Sterne JAC, Timpson N, Davey Smith G. Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat Med. 2008;27(8):1133–1163.
35. Davies NM, Holmes MV, Davey Smith G. Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ. 2018;362:k601. doi:10.1136/bmj.k601
36. Pierce BL, Burgess S. Efficient design for Mendelian randomization studies: subsample and 2-sample instrumental variable estimators. Am J Epidemiol. 2013;178(7):1177–1184. doi:10.1093/aje/kwt084
37. Magnus MC, Miliku K, Bauer A, et al. Vitamin D and risk of pregnancy related hypertensive disorders: mendelian randomisation study. BMJ. 2018;361:k2167. doi:10.1136/bmj.k2167
38. Hingorani A, Humphries S. Nature’s randomised trials. Lancet. 2005;366(9501):1906–1908.
39. Skrivankova VW, Richmond RC, Woolf BAR, et al. Strengthening the reporting of observational studies in epidemiology using mendelian randomization: the STROBE-MR statement. JAMA. 2021;326(16):1614–1621. doi:10.1001/jama.2021.18236
40. Lopera-Maya EA, Kurilshikov A, van der Graaf A, et al. Effect of host genetics on the gut microbiome in 7738 participants of the Dutch Microbiome Project. Nat Genet. 2022;54(2):143–151. doi:10.1038/s41588-021-00992-y
41. Orrù V, Steri M, Sidore C, et al. Complex genetic signatures in immune cells underlie autoimmunity and inform therapy. Nat Genet. 2020;52(10):1036–1045. doi:10.1038/s41588-020-0684-4
42. Auton A, Brooks LD, Durbin RM, et al. A global reference for human genetic variation. Nature. 2015;526(7571):68–74. doi:10.1038/nature15393
43. Burgess S, Davey Smith G, Davies NM, et al. Guidelines for performing Mendelian randomization investigations: update for summer 2023. Wellcome Open Res. 2019;4:186. doi:10.12688/wellcomeopenres.15555.3
44. Miyauchi E, Kim S-W, Suda W, et al. Gut microorganisms act together to exacerbate inflammation in spinal cords. Nature. 2020;585(7823):102–106. doi:10.1038/s41586-020-2634-9
45. Zheng D, Liwinski T, Elinav E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020;30(6):492–506. doi:10.1038/s41422-020-0332-7
46. Xu H, Liu M, Cao J, et al. The dynamic interplay between the gut microbiota and autoimmune diseases. J Immunol Res. 2019;2019:7546047. doi:10.1155/2019/7546047
47. Zafar H, Saier MH. Gut Bacteroides species in health and disease. Gut Microbes. 2021;13(1). doi:10.1080/19490976.2020.1848158
48. Shimizu J, Kubota T, Takada E, et al. Propionate-producing bacteria in the intestine may associate with skewed responses of IL10-producing regulatory T cells in patients with relapsing polychondritis. PLoS One. 2018;13(9):e0203657. doi:10.1371/journal.pone.0203657
49. Vinolo MAR, Rodrigues HG, Nachbar RT, Curi R. Regulation of inflammation by short chain fatty acids. Nutrients. 2011;3(10):858–876. doi:10.3390/nu3100858
50. Cruz-Bravo RK, Guevara-González RG, Ramos-Gómez M, et al. The fermented non-digestible fraction of common bean (Phaseolus vulgaris L.) triggers cell cycle arrest and apoptosis in human colon adenocarcinoma cells. Genes Nutr. 2014;9(1):359. doi:10.1007/s12263-013-0359-1
51. Vétizou M, Pitt JM, Daillère R, et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science. 2015;350(6264):1079–1084. doi:10.1126/science.aad1329
52. Hopkins D, Sanchez H, Berwin B, Wilkinson-Ryan I. Cisplatin increases immune activity of monocytes and cytotoxic T-cells in a murine model of epithelial ovarian cancer. Transl Oncol. 2021;14(12):101217. doi:10.1016/j.tranon.2021.101217
53. Mengos AE, Gastineau DA, Gustafson MP. The CD14+HLA-DRlo/neg monocyte: an immunosuppressive phenotype that restrains responses to cancer immunotherapy. Front Immunol. 2019;10:1147. doi:10.3389/fimmu.2019.01147
54. Bourbon E, Sesques P, Gossez M, et al. HLA-DR expression on monocytes and outcome of anti-CD19 CAR T-cell therapy for large B-cell lymphoma. Blood Adv. 2023;7(5):744–755. doi:10.1182/bloodadvances.2021006563
55. Amrane K, Le Meur C, Besse B, et al. HLA-DR expression in melanoma: from misleading therapeutic target to potential immunotherapy biomarker. Front Immunol. 2023;14:1285895. doi:10.3389/fimmu.2023.1285895
56. Meyer C, Cagnon L, Costa-Nunes CM, et al. Frequencies of circulating MDSC correlate with clinical outcome of melanoma patients treated with ipilimumab. Cancer Immunol Immunother. 2014;63(3):247–257. doi:10.1007/s00262-013-1508-5
57. Kitano S, Postow MA, Ziegler CGK, et al. Computational algorithm-driven evaluation of monocytic myeloid-derived suppressor cell frequency for prediction of clinical outcomes. Cancer Immunol Res. 2014;2(8):812–821. doi:10.1158/2326-6066.CIR-14-0013
58. Tarhini AA, Edington H, Butterfield LH, et al. Immune monitoring of the circulation and the tumor microenvironment in patients with regionally advanced melanoma receiving neoadjuvant ipilimumab. PLoS One. 2014;9(2):e87705. doi:10.1371/journal.pone.0087705
59. Martens A, Wistuba-Hamprecht K, Geukes Foppen M, et al. Baseline peripheral blood biomarkers associated with clinical outcome of advanced melanoma patients treated with ipilimumab. Clin Cancer Res. 2016;22(12):2908–2918. doi:10.1158/1078-0432.CCR-15-2412
60. Gebhardt C, Sevko A, Jiang H, et al. Myeloid cells and related chronic inflammatory factors as novel predictive markers in melanoma treatment with ipilimumab. Clin Cancer Res. 2015;21(24):5453–5459. doi:10.1158/1078-0432.CCR-15-0676
61. Eissler N, Mao Y, Brodin D, et al. Regulation of myeloid cells by activated T cells determines the efficacy of PD-1 blockade. Oncoimmunology. 2016;5(12):e1232222. doi:10.1080/2162402X.2016.1232222
62. Weber J, Gibney G, Kudchadkar R, et al. Phase I/II study of metastatic melanoma patients treated with nivolumab who had progressed after ipilimumab. Cancer Immunol Res. 2016;4(4):345–353. doi:10.1158/2326-6066.CIR-15-0193
63. Stroncek DF, Ren J, Lee DW, et al. Myeloid cells in peripheral blood mononuclear cell concentrates inhibit the expansion of chimeric antigen receptor T cells. Cytotherapy. 2016;18(7):893–901. doi:10.1016/j.jcyt.2016.04.003
64. Burga RA, Thorn M, Point GR, et al. Liver myeloid-derived suppressor cells expand in response to liver metastases in mice and inhibit the anti-tumor efficacy of anti-CEA CAR-T. Cancer Immunol Immunother. 2015;64(7):817–829. doi:10.1007/s00262-015-1692-6
65. Hosoi A, Matsushita H, Shimizu K, et al. Adoptive cytotoxic T lymphocyte therapy triggers a counter-regulatory immunosuppressive mechanism via recruitment of myeloid-derived suppressor cells. Int J Cancer. 2014;134(8):1810–1822. doi:10.1002/ijc.28506
66. Santegoets SJ, Stam AG, Lougheed SM, et al. Myeloid derived suppressor and dendritic cell subsets are related to clinical outcome in prostate cancer patients treated with prostate GVAX and ipilimumab. J Immunother Cancer. 2014;2:31. doi:10.1186/s40425-014-0031-3
67. Hansen GL, Gaudernack G, Brunsvig PF, Cvancarova M, Kyte JA. Immunological factors influencing clinical outcome in lung cancer patients after telomerase peptide vaccination. Cancer Immunol Immunother. 2015;64(12):1609–1621. doi:10.1007/s00262-015-1766-5
68. Olin MR, Low W, McKenna DH, et al. Vaccination with dendritic cells loaded with allogeneic brain tumor cells for recurrent malignant brain tumors induces a CD4(+)IL17(+) response. J Immunother Cancer. 2014;2:4. doi:10.1186/2051-1426-2-4
69. Poschke I, Mao Y, Adamson L, Salazar-Onfray F, Masucci G, Kiessling R. Myeloid-derived suppressor cells impair the quality of dendritic cell vaccines. Cancer Immunol Immunother. 2012;61(6):827–838. doi:10.1007/s00262-011-1143-y
70. Bakdash G, Buschow SI, Gorris MAJ, et al. Expansion of a BDCA1+CD14+ myeloid cell population in melanoma patients may attenuate the efficacy of dendritic cell vaccines. Cancer Res. 2016;76(15):4332–4346. doi:10.1158/0008-5472.CAN-15-1695
71. Kansagra A, Inwards DJ, Ansell SM, et al. Infusion of autograft natural killer cell/CD14+HLA-DRDIM cell ratio predicts survival in lymphoma post autologous stem cell transplantation. Bone Marrow Transplant. 2018;53(2):146–154. doi:10.1038/bmt.2017.225
72. Guan Q, Blankstein AR, Anjos K, et al. Functional myeloid-derived suppressor cell subsets recover rapidly after allogeneic hematopoietic stem/progenitor cell transplantation. Biol Blood Marrow Transplant. 2015;21(7):1205–1214. doi:10.1016/j.bbmt.2015.04.015
73. Mougiakakos D, Jitschin R, von Bahr L, et al. Immunosuppressive CD14+HLA-DRlow/neg IDO+ myeloid cells in patients following allogeneic hematopoietic stem cell transplantation. Leukemia. 2013;27(2):377–388. doi:10.1038/leu.2012.215
74. Vendramin A, Gimondi S, Bermema A, et al. Graft monocytic myeloid-derived suppressor cell content predicts the risk of acute graft-versus-host disease after allogeneic transplantation of granulocyte colony-stimulating factor-mobilized peripheral blood stem cells. Biol Blood Marrow Transplant. 2014;20(12):2049–2055. doi:10.1016/j.bbmt.2014.09.011
75. Matson V, Fessler J, Bao R, et al. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science. 2018;359:6371):104–108. doi:10.1126/science.aao3290
76. Derosa L, Routy B, Fidelle M, et al. Gut bacteria composition drives primary resistance to cancer immunotherapy in renal cell carcinoma patients. Eur Urol. 2020;78(2):195–206. doi:10.1016/j.eururo.2020.04.044
77. Xu L, Ma Y, Fang C, et al. Genomic and microbial factors affect the prognosis of anti-pd-1 immunotherapy in nasopharyngeal carcinoma. Front Oncol. 2022;12:953884. doi:10.3389/fonc.2022.953884
78. Zeriouh M, Raskov H, Kvich L, Gögenur I, Bennedsen ALB. Checkpoint inhibitor responses can be regulated by the gut microbiota - A systematic review. Neoplasia. 2023;43:100923. doi:10.1016/j.neo.2023.100923
79. Chen L, Flies DB. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat Rev Immunol. 2013;13(4):227–242. doi:10.1038/nri3405
80. Murray ME, Gavile CM, Nair JR, et al. CD28-mediated pro-survival signaling induces chemotherapeutic resistance in multiple myeloma. Blood. 2014;123(24):3770–3779. doi:10.1182/blood-2013-10-530964
81. Huh J-W, Kim MJ, Kim J, et al. Enterotypical Prevotella and three novel bacterial biomarkers in preoperative stool predict the clinical outcome of colorectal cancer. Microbiome. 2022;10(1):203. doi:10.1186/s40168-022-01388-8
82. Downes J, Munson M, Wade WG. Dialister invisus sp. nov. isolated from the human oral cavity. Int J Syst Evol Microbiol. 2003;53(Pt 6):1937–1940.
83. Ritu W, Enqi W, Zheng S, Wang J, Ling Y, Wang Y. Evaluation of the associations between cervical microbiota and HPV infection, clearance, and persistence in cytologically normal women. Cancer Prev Res. 2019;12(1):43–56. doi:10.1158/1940-6207.CAPR-18-0233
84. So KA, Yang EJ, Kim NR, et al. Changes of vaginal microbiota during cervical carcinogenesis in women with human papillomavirus infection. PLoS One. 2020;15(9):e0238705. doi:10.1371/journal.pone.0238705
85. Walther-António MRS, Chen J, Multinu F, et al. Potential contribution of the uterine microbiome in the development of endometrial cancer. Genome Med. 2016;8(1):122.
86. Byrd DA, Fan W, Greathouse KL, Wu MC, Xie H, Wang X. The intratumor microbiome is associated with microsatellite instability. J Natl Cancer Inst. 2023;115(8):989–993. doi:10.1093/jnci/djad083
87. Beyer M, Kochanek M, Darabi K, et al. Reduced frequencies and suppressive function of CD4+CD25hi regulatory T cells in patients with chronic lymphocytic leukemia after therapy with fludarabine. Blood. 2005;106(6):2018–2025.
88. Curiel TJ, Coukos G, Zou L, et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat Med. 2004;10(9):942–949.
89. Molling JW, de Gruijl TD, Glim J, et al. CD4(+)CD25hi regulatory T-cell frequency correlates with persistence of human papillomavirus type 16 and T helper cell responses in patients with cervical intraepithelial neoplasia. Int J Cancer. 2007;121(8):1749–1755.
90. Champagne P, Ogg GS, King AS, et al. Skewed maturation of memory HIV-specific CD8 T lymphocytes. Nature. 2001;410(6824):106–111.
91. Kurobe H, Liu C, Ueno T, et al. CCR7-dependent cortex-to-medulla migration of positively selected thymocytes is essential for establishing central tolerance. Immunity. 2006;24(2):165–177.
92. Mao F-Y, Kong H, Zhao Y-L, et al. Increased tumor-infiltrating CD45RA-CCR7- regulatory T-cell subset with immunosuppressive properties foster gastric cancer progress. Cell Death Dis. 2017;8(8):e3002. doi:10.1038/cddis.2017.388
93. Tassi E, Bergamini A, Wignall J, et al. Epithelial ovarian cancer is infiltrated by activated effector T cells co-expressing CD39, PD-1, TIM-3, CD137 and interacting with cancer cells and myeloid cells. Front Immunol. 2023;14:1212444. doi:10.3389/fimmu.2023.1212444
94. Derrien M, van Hylckama Vlieg JET. Fate, activity, and impact of ingested bacteria within the human gut microbiota. Trends Microbiol. 2015;23(6):354–366. doi:10.1016/j.tim.2015.03.002
95. Yang NJ, Chiu IM. Bacterial signaling to the nervous system through toxins and metabolites. J Mol Biol. 2017;429(5):587–605. doi:10.1016/j.jmb.2016.12.023
96. Bhattarai Y, Schmidt BA, Linden DR, et al. Human-derived gut microbiota modulates colonic secretion in mice by regulating 5-HT3 receptor expression via acetate production. Am J Physiol Gastrointest Liver Physiol. 2017;313(1):G80–G87. doi:10.1152/ajpgi.00448.2016
97. Yan J, Herzog JW, Tsang K, et al. Gut microbiota induce IGF-1 and promote bone formation and growth. Proc Natl Acad Sci U S A. 2016;113(47):E7554–E7563.
98. Natarajan N, Pluznick JL. From microbe to man: the role of microbial short chain fatty acid metabolites in host cell biology. Am J Physiol Cell Physiol. 2014;307(11):C979–C985. doi:10.1152/ajpcell.00228.2014
99. Ilinskaya ON, Ulyanova VV, Yarullina DR, Gataullin IG. Secretome of intestinal bacilli: a natural guard against pathologies. Front Microbiol. 2017;8:1666. doi:10.3389/fmicb.2017.01666
100. Selvam R, Maheswari P, Kavitha P, Ravichandran M, Sas B, Ramchand CN. Effect of Bacillus subtilis PB6, a natural probiotic on colon mucosal inflammation and plasma cytokines levels in inflammatory bowel disease. Indian J Biochem Biophys. 2009;46(1):79–85.
101. Behzadi E, Mahmoodzadeh Hosseini H, Imani Fooladi AA. The inhibitory impacts of Lactobacillus rhamnosus GG-derived extracellular vesicles on the growth of hepatic cancer cells. Microb Pathog. 2017;110:1–6. doi:10.1016/j.micpath.2017.06.016
102. Sun L, Zhu Z, Jia X, et al. The difference of human gut microbiome in colorectal cancer with and without metastases. Front Oncol. 2022;12:982744. doi:10.3389/fonc.2022.982744
103. Navas LE, Carnero A. NAD+ metabolism, stemness, the immune response, and cancer. Signal Transduct Target Ther. 2021;6(1):2. doi:10.1038/s41392-020-00354-w
104. van de Donk NWCJ, Richardson PG, Malavasi F. CD38 antibodies in multiple myeloma: back to the future. Blood. 2018;131(1):13–29. doi:10.1182/blood-2017-06-740944
105. Chini CCS, Peclat TR, Warner GM, et al. CD38 ecto-enzyme in immune cells is induced during aging and regulates NAD+ and NMN levels. Nat Metab. 2020;2(11):1284–1304. doi:10.1038/s42255-020-00298-z
106. Qin M, Hamanishi J, Ukita M, et al. Tertiary lymphoid structures are associated with favorable survival outcomes in patients with endometrial cancer. Cancer Immunol Immunother. 2022;71(6):1431–1442. doi:10.1007/s00262-021-03093-1
107. Thavaneswaran S, Kansara M, Lin F, et al. A signal-seeking Phase 2 study of olaparib and durvalumab in advanced solid cancers with homologous recombination repair gene alterations. Br J Cancer. 2023;129(3):475–485. doi:10.1038/s41416-023-02311-0
108. De Vuyst L, Leroy F. Bacteriocins from lactic acid bacteria: production, purification, and food applications. J Mol Microbiol Biotechnol. 2007;13(4):194–199.
109. Garbacz K. Anticancer activity of lactic acid bacteria. Semin Cancer Biol. 2022;86(Pt 3):356–366. doi:10.1016/j.semcancer.2021.12.013
110. Biasco G, Paganelli GM, Brandi G, et al. Effect of lactobacillus acidophilus and bifidobacterium bifidum on rectal cell kinetics and fecal pH. Ital J Gastroenterol. 1991;23(3):142.
111. Gill CIR, Rowland IR. Diet and cancer: assessing the risk. Br J Nutr. 2002;88(Suppl 1):S73–S87.
112. Jia W, Xie G, Jia W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat Rev Gastroenterol Hepatol. 2018;15(2):111–128. doi:10.1038/nrgastro.2017.119
113. Kaluanga bwanga P, Tremblay-Lemoine P-L, Timmermans M, et al. The endometrial microbiota: challenges and prospects. Medicina. 2023;59(9). doi:10.3390/medicina59091540
114. Chen C, Song X, Wei W, et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat Commun. 2017;8(1):875. doi:10.1038/s41467-017-00901-0
115. Reschini M, Benaglia L, Ceriotti F, et al. Endometrial microbiome: sampling, assessment, and possible impact on embryo implantation. Sci Rep. 2022;12(1):8467. doi:10.1038/s41598-022-12095-7
116. Borsellino G, Kleinewietfeld M, Di Mitri D, et al. Expression of ectonucleotidase CD39 by Foxp3+ Treg cells: hydrolysis of extracellular ATP and immune suppression. Blood. 2007;110(4):1225–1232.
117. Deaglio S, Dwyer KM, Gao W, et al. Adenosine generation catalyzed by CD39 and CD73 expressed on regulatory T cells mediates immune suppression. J Exp Med. 2007;204(6):1257–1265.
118. Maddaloni M, Kochetkova I, Hoffman C, Pascual DW. Delivery of IL-35 by lactococcus lactis ameliorates collagen-induced arthritis in mice. Front Immunol. 2018;9:2691. doi:10.3389/fimmu.2018.02691
119. Montuori-Andrade ACM, Nolasco AE, Malacco NLSO, et al. Lactobacillus delbrueckii UFV-H2b20 increases IFN-γ production and CD39+CD73+ Treg cell numbers in lungs, and protects mice against experimental allergic asthma. Immunobiology. 2022;227(6):152284. doi:10.1016/j.imbio.2022.152284
120. Argilés G, Tabernero J, Labianca R, et al. Localised colon cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann Oncol. 2020;31(10):1291–1305. doi:10.1016/j.annonc.2020.06.022
121. Lee P-J, Hung C-M, Yang A-J, et al. MS-20 enhances the gut microbiota-associated antitumor effects of anti-PD1 antibody. Gut Microbes. 2024;16(1):2380061. doi:10.1080/19490976.2024.2380061
122. Messaoudene M, Pidgeon R, Richard C, et al. A natural polyphenol exerts antitumor activity and circumvents anti-PD-1 resistance through effects on the gut microbiota. Cancer Discov. 2022;12(4):1070–1087. doi:10.1158/2159-8290.CD-21-0808
123. Taniguchi M, Harada M, Kojo S, Nakayama T, Wakao H. The regulatory role of Valpha14 NKT cells in innate and acquired immune response. Annu Rev Immunol. 2003;21:483–513.
124. Godfrey DI, MacDonald HR, Kronenberg M, Smyth MJ, Van Kaer L. NKT cells: what’s in a name? Nat Rev Immunol. 2004;4(3):231–237. doi:10.1038/nri1309
125. Kronenberg M. Toward an understanding of NKT cell biology: progress and paradoxes. Annu Rev Immunol. 2005;23:877–900.
126. Bendelac A, Savage PB, Teyton L. The biology of NKT cells. Annu Rev Immunol. 2007;25:297–336.
127. Rossjohn J, Pellicci DG, Patel O, Gapin L, Godfrey DI. Recognition of CD1d-restricted antigens by natural killer T cells. Nat Rev Immunol. 2012;12(12):845–857. doi:10.1038/nri3328
128. Stetson DB, Mohrs M, Reinhardt RL, et al. Constitutive cytokine mRNAs mark natural killer (NK) and NK T cells poised for rapid effector function. J Exp Med. 2003;198(7):1069–1076.
129. Krijgsman D, de Vries NL, Skovbo A, et al. Characterization of circulating T-, NK-, and NKT cell subsets in patients with colorectal cancer: the peripheral blood immune cell profile. Cancer Immunol Immunother. 2019;68(6):1011–1024. doi:10.1007/s00262-019-02343-7
130. Li J, Moresco P, Fearon DT. Intratumoral NKT cell accumulation promotes antitumor immunity in pancreatic cancer. Proc Natl Acad Sci U S A. 2024;121(29):e2403917121. doi:10.1073/pnas.2403917121
131. Wang K, Hou H, Zhang Y, Ao M, Luo H, Li B. Ovarian cancer-associated immune exhaustion involves SPP1+ T cell and NKT cell, symbolizing more malignant progression. Front Endocrinol. 2023;14:1168245. doi:10.3389/fendo.2023.1168245
132. Li B-L, Wan X-P. Prognostic significance of immune landscape in tumour microenvironment of endometrial cancer. J Cell Mol Med. 2020;24(14):7767–7777. doi:10.1111/jcmm.15408
133. Ma J, Lin J, Lin X, et al. Assessment of immune status in patients with mismatch repair deficiency endometrial cancer. J Inflamm Res. 2024;17:2039–2050. doi:10.2147/JIR.S453337
134. Li Y-R, Zhou Y, Yu J, et al. Engineering allorejection-resistant CAR-NKT cells from hematopoietic stem cells for off-the-shelf cancer immunotherapy. Mol Ther. 2024;32(6):1849–1874. doi:10.1016/j.ymthe.2024.04.005
135. Heczey A, Courtney AN, Montalbano A, et al. Anti-GD2 CAR-NKT cells in patients with relapsed or refractory neuroblastoma: an interim analysis. Nat Med. 2020;26(11):1686–1690. doi:10.1038/s41591-020-1074-2
136. Stabile G, Doria A, Bruno M, et al. The role of the endometrial microbiota in endometrial cancer: a systematic review of the literature. J Clin Med. 2024;13(23). doi:10.3390/jcm13237135
137. Wang L, Yang J, Su H, Shi L, Chen B, Zhang S. Endometrial microbiota from endometrial cancer and paired pericancer tissues in postmenopausal women: differences and clinical relevance. Menopause. 2022;29(10):1168–1175. doi:10.1097/GME.0000000000002053
138. Routy B, Lenehan JG, Miller WH, et al. Fecal microbiota transplantation plus anti-PD-1 immunotherapy in advanced melanoma: a Phase I trial. Nat Med. 2023;29(8):2121–2132. doi:10.1038/s41591-023-02453-x
139. Wu J, Li J, Yan M, Xiang Z. Gut and oral microbiota in gynecological cancers: interaction, mechanism, and therapeutic value. NPJ Biofilms Microbiomes. 2024;10(1):104. doi:10.1038/s41522-024-00577-7
140. Ciernikova S, Sevcikova A, Mladosievicova B, Mego M. Microbiome in cancer development and treatment. Microorganisms. 2023;12(1). doi:10.3390/microorganisms12010024
141. Spalinger MR, Scharl M. Microbiota manipulation as an emerging concept in cancer therapy. Visc Med. 2023;40(1). doi:10.1159/000534810
142. Knippel RJ, Drewes JL, Sears CL. The cancer microbiome: recent highlights and knowledge gaps. Cancer Discov. 2021;11(10):2378–2395. doi:10.1158/2159-8290.CD-21-0324
143. Sun J, Song S, Liu J, Chen F, Li X, Wu G. Gut microbiota as a new target for anticancer therapy: from mechanism to means of regulation. NPJ Biofilms Microbiomes. 2025;11(1):43. doi:10.1038/s41522-025-00678-x
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.