Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 13
Geniposide Balances the Redox Signaling to Mediate Glucose-Stimulated Insulin Secretion in Pancreatic β-Cells
Authors Liu C, Hao Y, Yin F, Liu J
Received 2 December 2019
Accepted for publication 23 January 2020
Published 25 February 2020 Volume 2020:13 Pages 509—520
DOI https://doi.org/10.2147/DMSO.S240794
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Antonio Brunetti
Chunyan Liu, Yanan Hao, Fei Yin, Jianhui Liu
Chongqing Key Laboratory of Medicinal Chemistry & Molecular Pharmacology, Chongqing University of Technology, Chongqing 400054, People’s Republic of China
Correspondence: Jianhui Liu; Fei Yin
Chongqing Key Laboratory of Medicinal Chemistry & Molecular Pharmacology, Chongqing University of Technology, Hongguang Road 69, Ba’nan District, Chongqing 400054, People’s Republic of China
Tel/Fax +86-23-6256-3182
Email [email protected]; [email protected]
Purpose: To investigate the effect of geniposide on the biosynthesis of insulin and the expression protein disulfide isomerase (PDI) and endoplasmic reticulum oxidoreductin 1 (ERO1) in the presence of low (5 mM) and high (25 mM) glucose in pancreatic β cells.
Methods: The content of insulin was measured by ELISA, the number of SH groups was determined with the classical chromogenic reagent, 5,5ʹ-dithiobis-(2-nitrobenzoic) acid (DTNB; also known as Ellman’s reagent), the expressions of PDI and ERO1 were analyzed by Western blot.
Results: Geniposide played contrary roles on the accumulation of H 2O 2, the ratio of GSH/GSSG and the thiol–disulfide balance in the presence of low (5 mM) and high (25 mM) glucose in rat pancreatic INS-1 cells. Geniposide also regulated the protein levels of protein disulfide isomerase (PDI) and endoplasmic reticulum oxidoreductin1 (ERO1), the two key enzymes for the production of H 2O 2 during the biosynthesis of insulin in INS-1 cells.
Conclusion: Geniposide affects glucose-stimulated insulin secretion by modulating the thiol–disulfide balance that is controlled by the redox signaling in pancreatic β cells.
Keywords: endoplasmic reticulum oxidoreductin1, ERO1, geniposide, protein disulfide isomerase, PDI, glucose-stimulated insulin secretion, GSIS, type 2 diabetes mellitus, T2DM
Introduction
Type 2 diabetes mellitus (T2DM), characterized by progressive impairment of insulin secretion and insulin resistance, is a common metabolic disease. Due to its complex health and steadily increasing prevalence, T2DM represents one of the serious burdens of the twenty-first century.1 Although great progress has been made in understanding the molecular mechanisms of T2DM, the proposed beneficial strategies for patients of T2DM are limited.
A growing body of evidence suggests that normal pancreatic β cells exhibit a dramatic response to nutrients and obesity-associated insulin resistance through hypersecretion of insulin to maintain energy homeostasis.2 However, pancreatic β cells are not able to sustain the compensatory response over an extended period under the conditions of high glucose or over-nutrition, which lead to β cells dysfunction and death.3 Studies have revealed that, due to the low anti-oxidative capacity of pancreatic β cells, chronic exposure of β cells to high concentrations of glucose, resulting in increased glycolytic flux and subsequent production of reactive oxygen species (ROS), such as superoxide, hydrogen peroxide (H2O2) and hydroxyl radicals, which plays a critical role in the dysfunction and failure of β cells.4 Furthermore, nitric oxide, another kind of free radicals, which is formed from L-arginine by nitric oxide synthase (NOS), has been proposed to coordinate the release of insulin by regulating the oscillation of the cytoplasmic Ca2+ and the activity of nitric oxide synthase.5,6 It has been reported that the administration of antioxidant supplements may increase the defense capacity of β cells in response to oxidative stress.7 On the other hand, the oxidative environment might be beneficial to enhance sensitivity to insulin in some key tissues targeted by T2DM.8,9
During the biosynthesis of insulin, protein disulfide isomerase (PDI) and endoplasmic reticulum oxidoreductin 1 (ERO1) are the two major contributors to disulfide bond formation in the ER, with PDI directly inserts the disulfide bond into target polypeptide, PDI becomes reduced and unable to catalyze further S-S insertion until reoxidized by ERO1. By oxidizing PDI, ERO1 becomes reduced and resumes activity when it is reoxidized by passing electrons to molecular oxygen (O2) and generating H2O2.10,11 It was shown that mRNA of PDI in the diabetes-susceptible BTBR mouse strain was approximately 20-fold more abundant relative to the diabetes-resistant C56BL/l strain.12 Furthermore, a reductive shift in the redox status of diabetic rat liver microsomes had been observed with a significant increase of reduced thiol groups in polypeptide chain of PDI and more oxidized disulfide bonds in ERO1.10 Moreover, it was reported that the static oxidation-reduction potential, a biomarker of oxidative stress, was significantly higher in the patients with metabolic syndrome and T2DM compared to the controls.13 So keeping the balance of cellular redox state is very important to sustain the normal function of pancreatic β cells and to prevent the development of T2DM.
Gardenia jasminoides Ellis has been used as an important traditional Chinese medicine for many years in China and other Asian counties. Geniposide, isolated from its fruit, has a wide scope of therapeutic activities, such as anti-diabetic,14 anti-oxidative,15 anti–inflammatory,16 and hepatic disorders.17 Our previous works revealed that geniposide was a novel agonist for glucagon-like peptide receptor (GLP-1R), which enhanced acute glucose-stimulated insulin secretion (GSIS) in response to the stimulation of low or moderately high concentrations of glucose, and promoted glucose uptake and intracellular ATP levels in INS-1 cells.18–21 However, in the presence of a high concentration of glucose, geniposide exerted a contrary role on GSIS, and uptake and metabolism of glucose.22 Most importantly, we further demonstrated that the role of geniposide on GSIS was associated with its modulation on the expression of pyruvate carboxylase, a crucial enzyme for glucose metabolism in pancreatic β cells.22 But unfortunately, the molecular mechanisms of action are not well understood. Evidence is emerging to indicate that ROS derived from glucose metabolism, in particular H2O2, servers as additional metabolic signals to elicit GSIS.8,23 Multiple studies over the past decade also showed that H2O2 molecules played a role in glucose-stimulating insulin secretion (GSIS). So, we speculate that geniposide-regulating GSIS might be involved in its role in the pro-oxidant/antioxidant balance and the biosynthesis of insulin. Herein, we design to investigate the role of geniposide on the production of H2O2, the thiol–disulfide balance, the biosynthesis and secretion of insulin, and the expression of PDI/ERO1 in the presence of low and high concentrations of glucose in INS-1 cells.
Materials and Methods
Cell Culture
Rat INS-1 pancreatic β cell line, purchased from CCTCC (China Center for Type Culture Collection), was cultured at 37°C in a humidified atmosphere containing 5% CO2. The culture medium was RPMI medium 1640 containing 11 mM glucose and supplemented with 10% FBS, 10 mM HEPES, 100 U/mL penicillin, 100 μg/mL streptomycin, 2 mM L-glutamine, 1 mM sodium pyruvate and 50 μM mercaptoethanol.
Pancreatic Islet Isolation and Culture
Pancreatic islets were isolated from 2 to 3-month-old rats by the use of a collagenase inflation method.24 Generally, the pancreas of the anesthetized rat was perfused with a solution of collagenase type-V (1.0 mg/mL) in RPMI 1640 medium supplemented with 1% v/v penicillin/streptomycin and 1% L-glutamine via the common bile duct, the pancreas was digested for 17 mins at 37°C. Islets were purified by centrifugation with a Histopaque 1077. Islets were picked by hand in succession under a dissecting microscope and were cultured overnight in RPMI 1640 supplemented with 10% v/v FBS, 10 mM HEPES, 100 U/mL penicillin, 100 μg/mL streptomycin, 2 mM L-glutamine,1 mM sodium pyruvate and 50 μM mercaptoethanol in a humidified atmosphere of 5% CO2 and 95% air at 37°C. All procedures were performed by following the Institutional Guidelines for Animal Care at the Chongqing University of Technology and also approved by the Institutional Animal Care and Use Committee at Chongqing Science and Technology Committee.
Insulin Secretion Assay
An insulin secretion assay was performed as previously described.19,22 Briefly, INS-1 cells or primary islets were seeded onto 12-well plates and cultured overnight. Then, the cells were washed once with PBS and starved for 2 hrs in KRBB (129 mM NaCl, 4.8 mM KCl, 1.2 mM MgSO4, 1.2 mM KH2PO4, 2.5 mM CaCl2, 5 mM NaHCO3, 0.1% BSA, 10 mM HEPES, pH 7.4). The cells were incubated for 1 hr with or without geniposide in the fresh KRBB in the presence of different concentrations of glucose. The supernatants were collected to measure insulin contents using commercial kits according to the kit’s instructions.
Determination of H2O2 Content
The content of H2O2 was determined by using a fluorescence method reported earlier.25 Briefly, after the cells were incubated with indicated concentrations of glucose in the absence or presence of geniposide, the cells were washed once with PBS and the cell lysates were used to determine the content of H2O2 with the hydrogen peroxide fluorometric assay kit (Bio Vision, CA, USA) according to the manufacturer’s instructions.
Determination of Thiol Group
The number of thiol groups was determined with the classical chromogenic reagent, 5,5′-dithiobis-(2-nitrobenzoic) acid (DTNB; also known as Ellman’s reagent), as described by Nakano et al.26 In brief, the cells were washed once with PBS and starved for 2 hrs in fresh KRBB. The cells were then treated with different concentrations of glucose in the presence or absence of geniposide, and the cell lysates were obtained by using modified RIPA buffer, and samples were concentrated to around 10 mg/mL and suspended in 800 μL of 0.1 M Tris·HCl buffer (pH 8.0) containing 8 M urea and 1 mM EDTA. Fifty microliters of 0.1 M DTNB in 50 mM phosphate buffer (pH 7.0) was added to the solution. The reaction mixture was kept at room temperature for 10 min, and absorption at 412 nm was determined using a DTNB solution in 50 mM phosphate buffer as a control. The molar concentration of the thiol group was calculated using molar extinction coefficients of DTNB at 412 nm.
Protein Extraction
Cells were washed once with ice-cold PBS and whole-cell extracts were obtained by using modified RIPA buffer in the presence of 1% protease/phosphatase inhibitor cocktail, and protein concentrations were determined using BCA protein assay kit (Biyotime, Shanghai, China). All of the protein extracts were stored at −70°C.
Western Blot Analysis
After INS-1 cells were treated with geniposide in the presence of different concentrations of glucose and were washed with cold PBS and lysed in an RIPA buffer. Protein concentrations were calculated by BCA protein assay kit (Biyotime, Shanghai, China), 20–30 μg cell lysates were separated with a 10% SDS-PAGE gel and transferred to polyvinylidene difluoride (PVDF) membranes. After blocking, membranes were probed with an anti-PDI (1:2000 dilution) and anti-ERO1α/β (1:3000 dilution) followed by incubation with anti-horseradish-conjugated second antibodies (1:10,000 dilution). Excess antibody was washed off with 20 mM Tris-buffered saline containing Tween-20 (TBST, 20 mM Tris, 150 mM NaCl, and 0.1% Tween 20, pH 7.5–7.6). Immunoreactivity was detected using ECL Western blotting reagent from Millipore Corporation (Billerica, MA, USA). Signal bands were analyzed by Quantity One software (Bio-Rad, Hercules, CA, USA).
Statistical Analysis
Results are presented as mean ± SD from at least three independent experiments. Analysis of variance was carried out using the software of Origin Lab (Northampton, MA, USA). A one-way ANOVA followed with a post hoc Tukey’s or Dunnett’s test was used to detect and compare the differences among all the groups or selected groups to control. p < 0.05 was considered statistically significant.
Results
Geniposide Regulates GSIS
To confirm the role of geniposide on glucose-stimulated insulin secretion (GSIS), we first determined the influence of geniposide on GSIS in primary-cultured rat islets. The results demonstrated that similar with INS-1 cells, geniposide increased GSIS in the presence of low (5 mM) and moderate (11 mM) concentrations of glucose, but in the presence of high glucose (25 mM), geniposide decreased the insulin content in primary rat islets (Figure 1A).
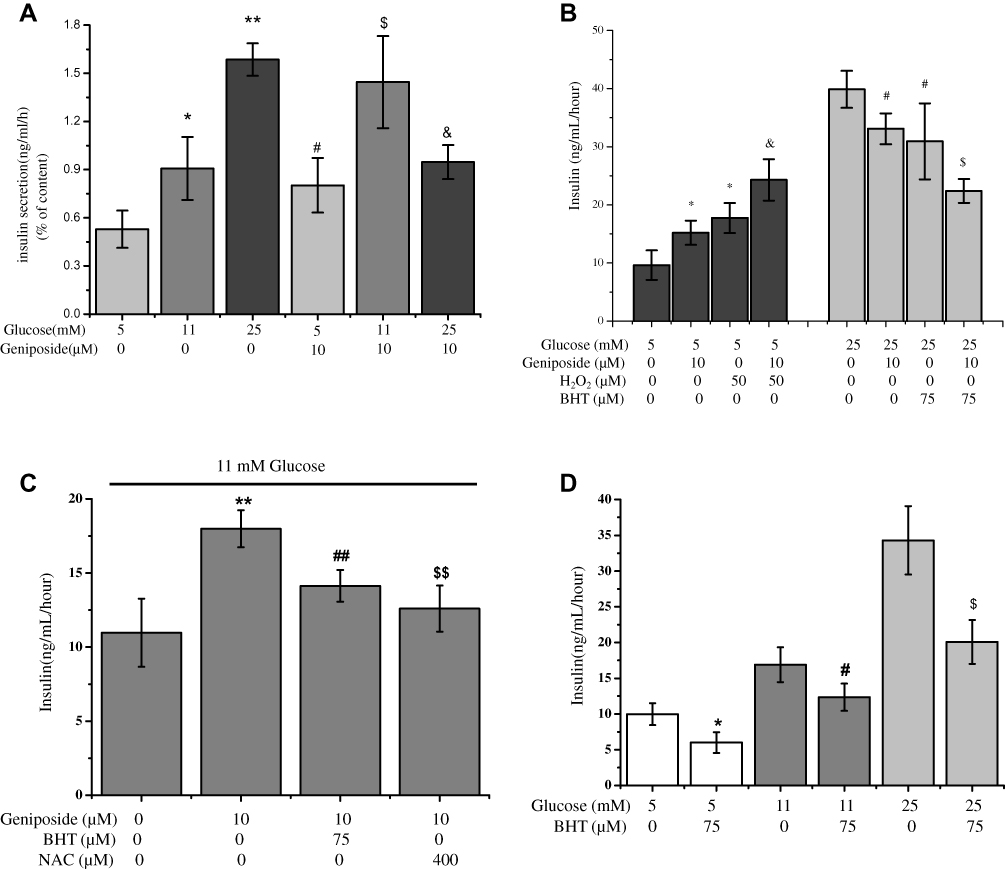 |
Figure 1 Geniposide regulates insulin secretion. (A) After primary-cultured rat islets were starved for 2 hrs in fresh KRBB, the media was changed with 5, 11 or 25 mM glucose completed media, and the cells were treated with 10 μM geniposide for 1 hr. The supernatants were collected to determine the content of insulin using commercial ELISA kits. Data are shown as mean ± SD (n = 6), *p < 0.05, **p < 0.01 vs 5 mM glucose alone, #,$,&p < 0.05 vs 5, 11 and 25 mM glucose, respectively. (B) After primary-cultured rat islets were starved for 2 hrs in fresh KRBB, the media was changed with 5 mM or 25 mM glucose completed media, and the cells were treated with 75 μM BHT or 50 µM H2O2in the presence or absence of 10 μM geniposide for 1 hr. The supernatants were collected to determine the content of insulin using commercial ELISA kits. Data are shown as mean ± SD (n=6), *p < 0.05 vs 5 mM glucose alone, &p < 0.05 vs 5 mM glucose plus 10 μM geniposide. #p < 0.05 vs 25 mM glucose alone, $p < 0.05 vs 25 mM glucose plus 10 μM geniposide. (C) After primary-cultured rat islets were starved for 2 hrs in fresh KRBB, the media was changed normal completed media (11 mM glucose), and then the cells were treated with 75 μM BHT or 400 µM NAC (N-acetyl-cysteine) in the presence or absence of 10 μM geniposide for 1 hr. The supernatants were collected to determine the content of insulin using commercial ELISA kits. Data are shown as mean ± SD (n = 6), **p < 0.051 vs 11 mM glucose alone, ##,&&p < 0.01 vs 10 μM geniposide alone. (D) After primary-cultured rat islets were starved for 2 hrs in fresh KRBB, the cells were treated with 75 μM BHT in the presence 5, 11 or 25 mM glucose for 1 hr. The supernatants were collected to determine the content of insulin using commercial ELISA kits. Data are shown as mean ± SD (n = 6), *p < 0.05 vs 5 mM glucose alone, #p < 0.05 vs 11 mM glucose alone, $p < 0.05 vs 25mM glucose alone. |
To probe the influence of geniposide on the redox status and its effect on GSIS, we determined the influence of antioxidants and oxidants on geniposide regulating GSIS in primary-cultured islets. The results showed that, in the presence of 5 mM glucose, geniposide increased GSIS and H2O2 potentiated the effect of geniposide on GSIS (p < 0.05). But in the presence of 25 mM glucose, geniposide attenuated GSIS, and butylated hydroxytoluene (BHT), enhanced the inhibiting effect of geniposide on GSIS (p < 0.05) (Figure 1B). Furthermore, BHT and N-acetyl-L-cysteine (NAC), two popular antioxidants, significantly prevented the effect of geniposide on GSIS in the presence of 11 mM glucose in primary-cultured islets (Figure 1C). Meanwhile, it was noticed that different from geniposide, antioxidant (BHT) inhibited GSIS in the presence of low (5 mM), moderate (11 mM) and high (25 mM) glucose in primary-cultured islets, which was independent on the concentrations of glucose (Figure 1D).
Geniposide Regulates the Accumulation of intracellular H2O2
As shown in Figure 2A, when INS-1 cells were incubated with 5, 11 and 25 mM glucose for 1 hr, the level of intracellular H2O2 was increased in a dose-dependent manner, and geniposide markedly raised the accumulation of H2O2 in the presence of low (5 mM) and moderate (11 mM) concentrations of glucose. But, when the concentration of glucose achieved to 25 mM, geniposide significantly attenuated the level of H2O2 (p<0.01). Interestingly, BHT, one of the popular antioxidants, showed a totally different role on the level of H2O2 from geniposide, which considerably decreased the levels of H2O2 in 5, 11 and 25 mM glucose-treated INS-1 cells (Figure 2B).
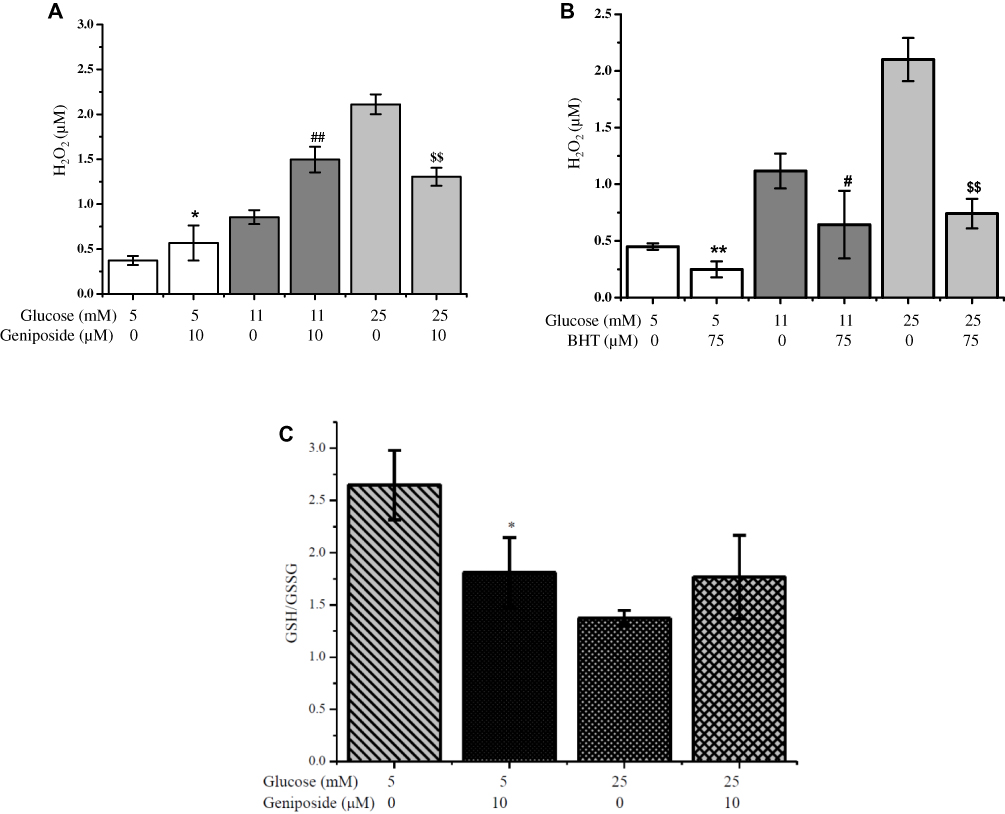 |
Figure 2 Effects of geniposide (A) and BHT (B) on the accumulation of H2O2. (A, B) After INS-1 cells were washed twice with KRBB, the cells were starved for 2 hrs in fresh KRBB, and the cells were continued to incubate with 10 µM geniposide (A) or 75 µM BHT (B) in the presence of indicated concentrations of glucose for 1 hr, the content of H2O2 in cell lysates was determined with hydrogen peroxide assay kit according to the instructions from supplier. Data are shown as mean ± SD (n =6). *p < 0.05, **p < 0.01 vs 5 mM glucose alone; #p < 0.05, ##p < 0.01 vs 11 mM glucose alone; $$p < 0.01 vs 25 mM glucose alone. (C) The contents of total GSH and reduced GSH in cell lysates were detected with commercial kits, and the ratio of GSH/GSSG was calculated according to the instructions from the supplier. Data are shown as mean ± SD from four independent experiments (three well for each replicate). *p < 0.05 vs the group of 5 mM glucose alone. |
Several reports indicate that the robust GSIS is accompanied by a significant increase in intracellular H2O2 levels and decreased the ratio of glutathione (GSH)/glutathione disulfide (GSSG). And the balance between GSH and GSSG has been considered as an important indicator of oxidative stress and redox signaling.27,28 To explore the influence of geniposide on the redox signals, we directly determined the effect of geniposide on the ratio of GSH/GSSG in the presence of low (5 mM) and high (25 mM) glucose in INS-1 cells. As shown in Figure 2C, geniposide remarkably decreased the ratio of GSH/GSSG (p < 0.05) in the presence of 5 mM glucose, but the presence of 25 mM glucose, geniposide increased the ratio of GSH/GSSG (p = 0.06), hinting that geniposide plays a different role on the redox signals in the presence of low and high concentrations of glucose in pancreatic β-cells.
Geniposide Regulates the Protein Level of PDI
Substantial evidence suggests that matured insulin has three S-S bonds, which is stabilized by van der Waals’ interactions and catalyzed by oxidoreductive thiol enzymes, protein disulfide isomerase (PDI) and endoplasmic reticulum oxidoreductin 1 (ERO1). PDI and ERO1 proteins harness the oxidizing power of molecular oxygen to create de novo disulfide bonds during the biosynthesis of insulin.11,29 In the present study, to probe the molecular mechanisms of geniposide regulating redox status to influence GSIS, we determined the effect of geniposide on the protein level of PDI with Western blot assay. The results indicated that, after starved for 2 hrs in KRBB, treatment with 10 μM geniposide for 1 hr (the condition is same as the determination of glucose-stimulated insulin secretion) significantly enhanced the protein level of PDI in the presence of 5 mM glucose, and geniposide remarkably attenuated the level of PDI protein in the presence of 25 mM glucose in INS-1 cells (Figure 3A). However, if the cells had not been starved, treatment with 10 μM geniposide for 1 hr had no noticeable effect on the expression of PDI (Figure 3B), but if the treating time was extended to 24 hrs, geniposide significantly enhanced the protein level of PDI in the presence of 5 mM glucose (Figure 3C) and attenuated the protein level of PDI in the presence of 25 mM glucose (Figure 3D). Furthermore, we also found that, although geniposide had no significant role to potentiate the effect of H2O2 on PDI protein INS-1 cells (p < 0.05) (Figure 3E), but in the presence of high glucose (25 mM), geniposide worked as an antioxidant to decrease the protein level of PDI (p < 0.05) (Figure 3F).
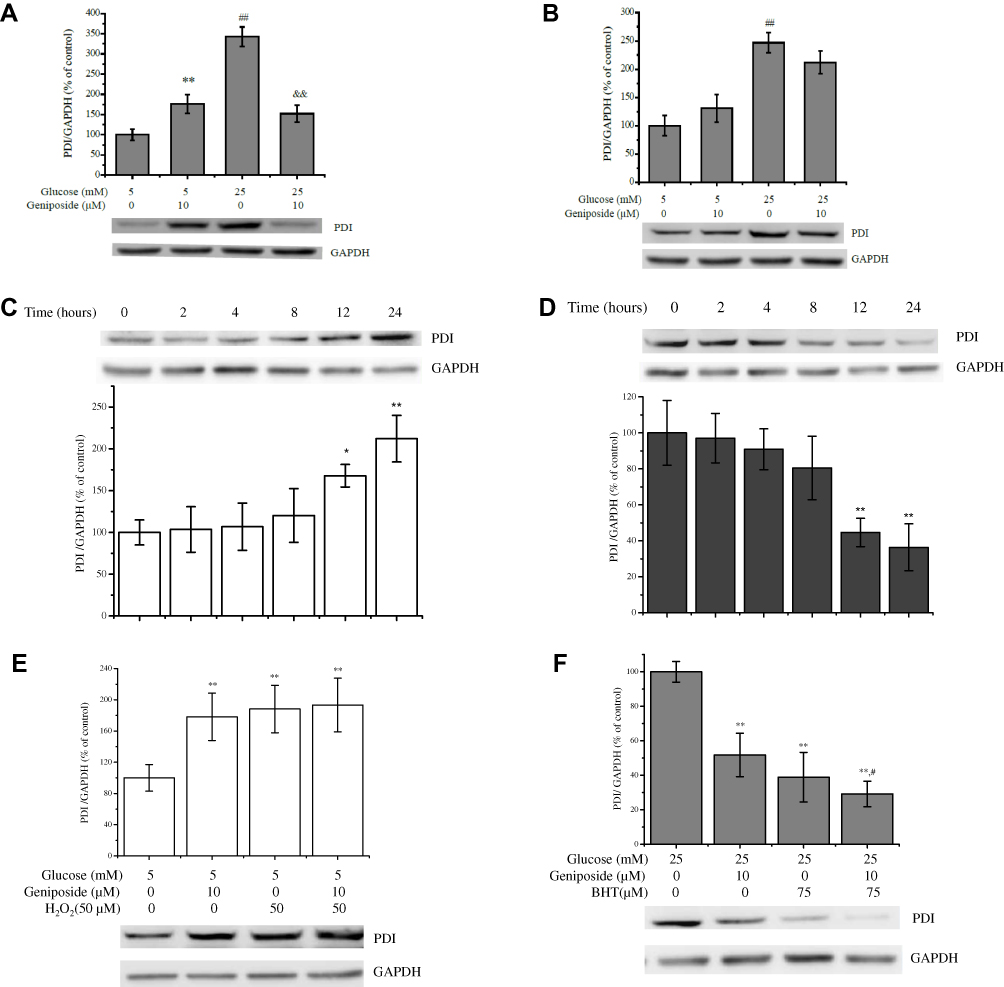 |
Figure 3 Geniposide regulates the protein level of PDI. (A) After INS-1 cells were starved for 2 hrs and then treated with 10 μM geniposide for 1 hr in the presence of 5 or 25 mM glucose the protein level of PDI was determined by Western blot. Data are shown as mean ± SD from three independent experiments. **,##p < 0.01 vs the group of 5 mM glucose alone, &&p < 0.01 vs the group of 25 mM glucose alone. (B) After INS-1 cells were incubated with 10 μM geniposide for 1 hr without starvation in the presence of 5 or 25 mM glucose. ##p < 0.01 vs the group of 5 mM glucose alone. (C) After INS-1 cells were incubated with 10 μM geniposide for 24 hrs without starvation in the presence of 5 mM glucose. *p < 0.05, **p < 0.01 vs control (0 hr). (D) After INS-1 cells were treated with 10 μM geniposide for 24 hrs without starvation, the effects of geniposide on the protein levels of PDI in the presence of 25 mM glucose were determined with Western blot. **p < 0.01 vs control (0 hr). (E) After the cells were incubated with 10 μM geniposide and/or 50 μM H2O2 in the presence of 5 mM glucose for 24 hrs, the PDI protein was detected by Western blot, data are shown as mean ± SD (n = 3). **p < 0.01 vs 5 mM glucose alone; (F) After the cells were incubated with 10 μM geniposide and/or 75 μM BHT in the presence of 25 mM glucose for 24 hrs, the PDI protein was detected by Western blot, data are shown as mean ± SD (n = 3). **p < 0.01 vs 25 mM glucose alone, #p < 0.05 vs 25 mM glucose plus 75 μM BHT. |
Geniposide Regulates the Protein Level of ERO1
As another oxidoreductive thiol enzyme, ERO1 plays an essential role to reoxidize PDI and resume its catalytic capacity for further S-S insertion.11 At present, we also measured the effect of geniposide on the protein level of ERO1α with the Western blot method. After starved for 2 hrs in KRBB, treatment with 10 μM geniposide for 1 hr (the condition is same as the determination of glucose-stimulated insulin secretion) significantly enhanced the protein level of ERO1α in the presence of 5 mM glucose, and geniposide remarkably attenuated the level of ERO1α protein in the presence of 25 mM glucose in INS-1 cells (Figure 4A). However, if the cells had not been starved, treatment with 10 μM geniposide for 1 hr had no noticeable effect on the expression of ERO1α (Figure 4B), and if the treating time was extended to 24 hrs, geniposide increased the protein level of ERO1α in the presence of 5 mM glucose, and treatment with 10 μM geniposide for 24 hrs increased the protein of ERO1α about 2.1 folds in INS-1 cells (Figure 4C). But in the presence of high glucose (25 mM), geniposide attenuated the protein level of ERO1α, and treatment with 10 μM geniposide for 24 hrs could attenuate the protein level of ERO1α to around 50% in INS-1 cells. Furthermore, when the cells were incubated with 75 µM BHT in the presence of high glucose (25 mM), geniposide could further attenuate the protein level of ERO1α in INS-1 cells (Figure 4D). But unfortunately, geniposide had no significant role on the protein level of ERO1β in high glucose (25 mM) cultured INS-1 cells (Figure 4E).
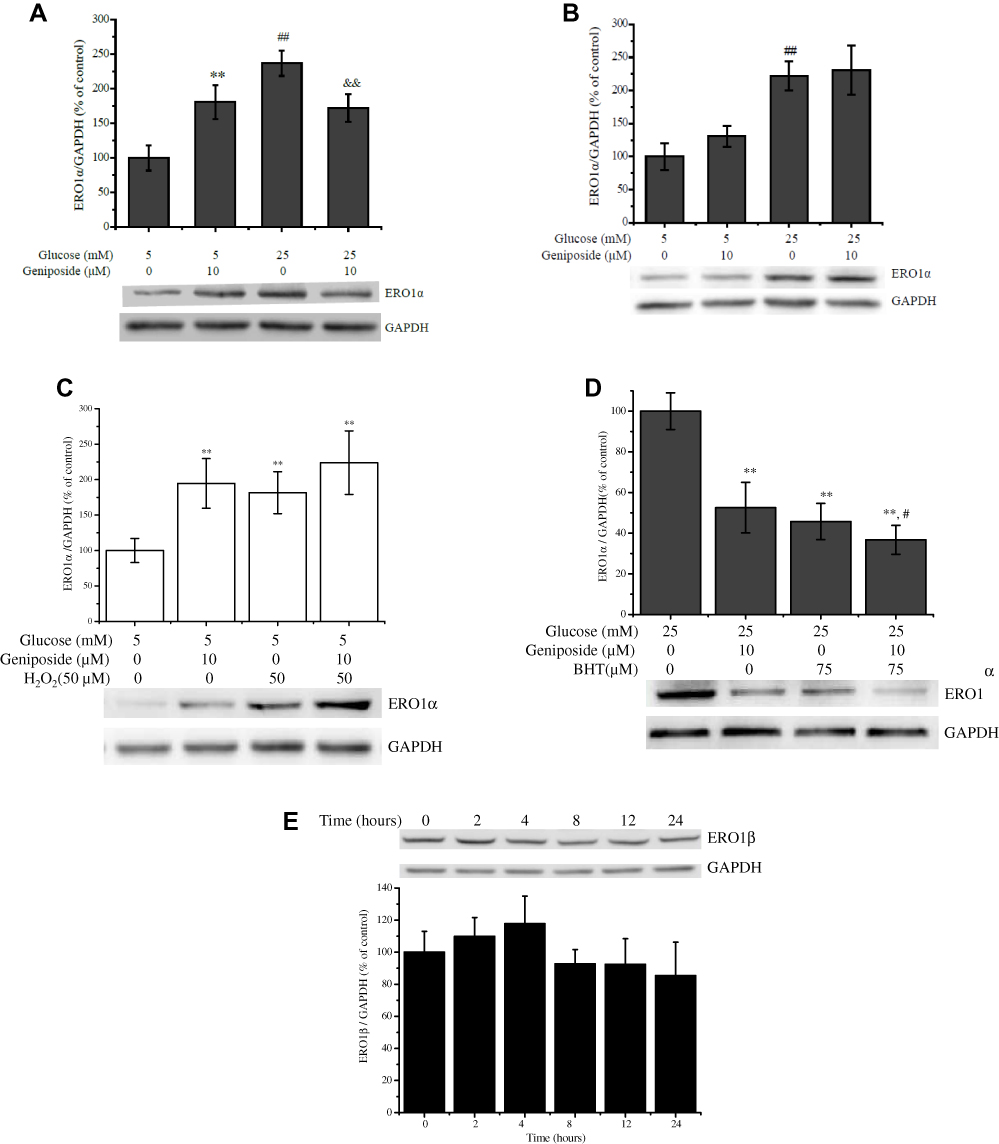 |
Figure 4 Geniposide regulates the protein level of ERO1. (A) After INS-1 cells were starved for 2 hrs and then treated with 10 μM geniposide for 1 hr in the presence of 5 or 25 mM glucose the protein level of ERO1α was determined by Western blot. Data are shown as mean ± SD from three independent experiments. **,##p < 0.01 vs the group of 5 mM glucose alone, &&p < 0.01 vs the group of 25 mM glucose alone. (B) After INS-1 cells were incubated with 10 μM geniposide without starvation in the presence of 5 or 25 mM glucose. ##p < 0.01 vs the group of 5 mM glucose alone. (C) After INS-1 cells were incubated with 10 μM geniposide and/or 50 μM H2O2 in the presence of 5 mM glucose for 24 hrs, the ERO1α protein was detected by Western blot. Data are shown as mean ± SD (n = 3). **p < 0.01 vs 5 mM glucose alone; (D) After INS-1 cells were incubated with 10 μM geniposide and/or 75 μM BHT in the presence of 25 mM glucose for 24 hrs, the ERO1α proteins were detected by Western blot. Data are shown as mean ± SD (n = 3). **p < 0.01 vs 25 mM glucose alone, #p < 0.05 vs 25 mM glucose plus 75 μM BHT. (E) After INS-1 cells were incubated with 10 μM geniposide in the presence of 25 mM glucose for 24 hrs, the ERO1β proteins were detected by Western blot. Data are shown as mean ± SD (n = 3). |
Geniposide Regulates the Content of the Thiol Group
Insulin consists of two individual chains, A-chain and B-chain, which are locked together by three disulfide bonds. During the biosynthesis of insulin, the formation of disulfide, which is usually derived by the coupling of two thiol groups (SH), is the key step.30 To investigate the effect of geniposide on the formation of disulfide bond, we measured the content of thiol in the geniposide-treated INS-1 cells. As shown in Figure 5, geniposide significantly decreased the content of thiol in the presence of low glucose (5 mM), as well as the presence of 50 µM H2O2, hinted that treatment with geniposide might be beneficial to the formation of disulfide in the presence of 5 mM glucose in INS-1 cells. But, in the presence of high glucose (25 mM), geniposide increased the content of thiol, as well as the presence of 75 µM BHT, in INS-1 cells, suggesting that geniposide might attenuate the formation of disulfide in the presence of high concentrations of glucose.
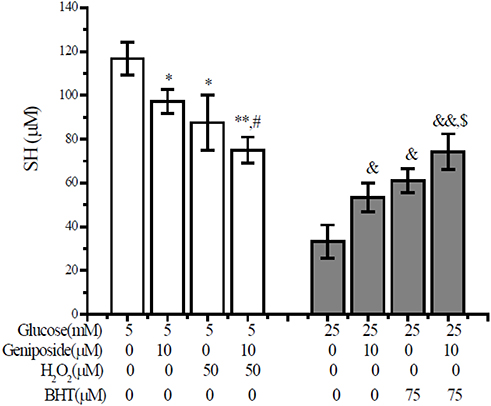 |
Figure 5 Geniposide regulates the content of thiol group in INS-1 cells. After INS-1 cells were washed twice with KRBB, the cells were starved for 2 hrs in fresh KRBB. And then, the cells were incubated with 10 µM geniposide together with 75 μM BHT (in the presence of 5 mM glucose) or 50 μM H2O2 (in the presence of 25 mM glucose) for 1 hr, the content of thiol was measured with DTNB assay. Data are expressed as mean ± SD (n= 6). *p < 0.05, **p < 0.01 vs 5 mM glucose alone; #p < 0.05 vs 5 mM glucose plus 50 μM H2O2; &p < 0.05, &&p < 0.01 vs 25 mM glucose alone, and $p < 0.05 vs 25 mM glucose plus 75 µM BHT. |
Discussion
In this study, we identified that in addition to regulating insulin secretion, geniposide played a contrary role on the production of H2O2 in the presence of low (5 mM) and high (25 mM) concentrations of glucose, which was different from the role of antioxidants on GSIS and the production of H2O2 both in INS-1 cells and primary-cultured islets. Exogenous antioxidants, such as BHT, decreased the level of H2O2 and attenuated GSIS distinctively, which was independent of the concentrations of glucose. Geniposide, however, increased the level of H2O2 and accelerated GSIS in the presence of low concentration of glucose (5 mM), and in the presence of a high concentration of glucose (25 mM), geniposide suppressed GSIS and decreased the production of H2O2 in INS-1 cells. All these data suggest that geniposide-regulating GSIS might be associated with its role on the redox balance in pancreatic β cells.
Mounting evidence shows that ERO1 (including the two isoforms, α and β) plays a critical role in promoting the oxidative folding of proinsulin in the endoplasmic reticulum. ERO1 couples disulfide transfer to PDI with reduction of molecular oxygen, in turn, PDI can shuttle disulfide bonds to substrate to catalyze the folding of the newly synthesized secretory protein, such as proinsulin.31 Moreover, ERO1 is the best-known source of disulfide bonds in the ER lumen, and ERO1 deficiency impairs proinsulin maturation.32,33 To further investigate the molecular mechanisms of geniposide-regulating GSIS and the level of H2O2, we determined the effect of geniposide on the expression of PDI and ERO1, which were the two key oxidoreductive thiol enzymes for the formation of S-S bond and the production of H2O2. The results indicated that geniposide inhibited the expression of PDI and ERO1 in the presence of high glucose (25 mM), but significantly increased the protein level of PDI and ERO1 in the presence of low glucose (5 mM) in INS-1 cells. We also observed that the role of oxidant (H2O2) and anti-oxidant (BHT) on the protein levels of ERO-1 and PDI was different from geniposide. Although the reasons and relative mechanisms about those need to be further clarified, a couple of references demonstrated that the ERO1-PDI pathway for disulfide bond formation was sensitive to the redox state of the endoplasmic reticulum (ER) and may recruit different regulators during the redox cycle, and ERO-1α-PDI dimer was lost under oxidizing conditions when there is less need for de novo disulfide bond formation.31 Moreover, PDI did not facilitate oxidative folding of proinsulin in the ER of pancreatic β cells and had been implicated in retro-translocation of certain ER-associated degradation (ERAD) substrates.34
Substantial evidence suggested glutathione was a fundamental function in cellular redox homeostasis,35,36 and cytosol derived reduced glutathione (GSH) could enter endoplasmic reticulum (ER), which did not only as an indicator of oxidative stress but also played an essential role on the rearrangement of aberrant disulfide bonds.37 Additionally, reactive oxygen species (ROS) plays an essential role in GSIS, and induction of endogenous antioxidative enzymes in β cells abrogates ROS signaling and reduces GSIS.23 Intriguingly, one of the unique features of pancreatic β cells is their relatively low expression and activity of antioxidative defense enzymes, such as superoxide dismutase (SOD), catalase, and glutathione peroxide (GPx), which renders β cell susceptible to oxidative damage but provides a system sensitive to ROS signaling.4 So, the balance between ROS signaling and antioxidant defense can be implicated in the dysfunction and failure of β cells. At present, our data indicated that geniposide played a contrary role on the ratio of GSH/GSSG, thiol–disulfide balance and the biosynthesis and release of insulin in the presence of low glucose (5 mM) and high glucose (25 mM) in INS-1 cells, suggesting that geniposide might be useful to keep the balance of redox status in pancreatic β-cells.
Mounting evidence shows that there is a delicate balance between oxidants and antioxidants in health, the proper balance of which is essential for the functions and survival of cells.23 When challenged by the high concentration of glucose, pancreatic β cells need to secrete more insulin to compensate for glucose intolerance. But during the process of GSIS, the oxidative phosphorylation of glucose, which is accompanied by the production of ROS, is necessary for increasing the ratio of ATP/ADP in pancreatic β-cells.8 It is increasingly understood that continuous exposure to high levels of glucose gradually leads to the diminution or complete loss of functions and even induce pancreatic β cells death.23 At present, geniposide was found to decrease the level of H2O2 in the presence of a high concentration of glucose, and thus might be beneficial for keeping the normal functions of pancreatic β cells. Together with decreasing GSIS in an acute stimulation of a high concentration of glucose, geniposide also significantly improved the impairment of insulin secretion induced by a chronic high concentration of glucose in INS-1 cells.18,21,38 So, geniposide might be a promising β cell protectant to prevent pancreatic β cells from exhaustion and injury resulted from over insulin secretion under high glucose burden.
Together with the effects of geniposide on the production of H2O2, the ratio of GSH/GSSG, the balance of thiol-disulfide, and the expression of PDI/ERO1, it should be reasonable to conclude that geniposide regulating GSIS is associated with its role on the biosynthesis of insulin. And the possible way of geniposide balancing the redox of pancreaticβ cells is summarized in Figure 6.
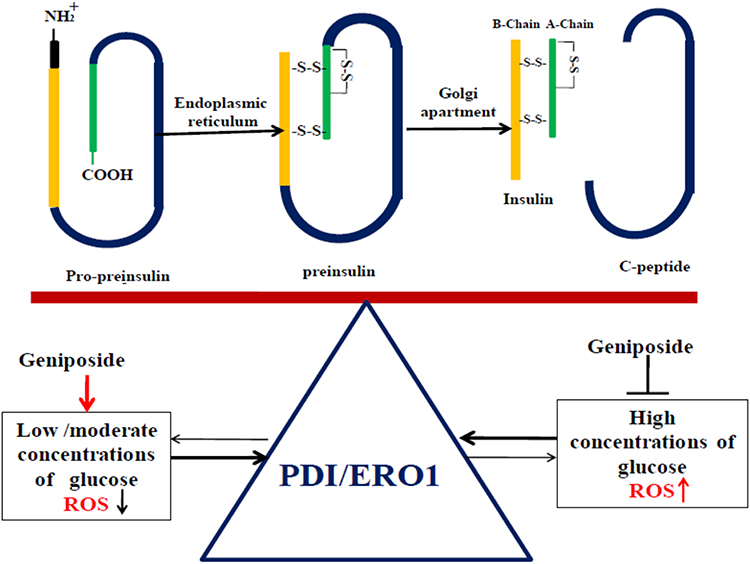 |
Figure 6 The possible ways of geniposide balance the redox of pancreatic β cells by regulating the protein levels of PDI and ERO1. Evidence is emerging to indicate that during the biosynthesis of insulin, protein disulfide isomerase (PDI) and endoplasmic reticulum oxidoreductin 1 (ERO1) are the two major contributors to disulfide bond formation in the ER. ERO1 couples disulfide transfer to PDI with reduction of molecular oxygen, in turn, PDI can shuttle disulfide bonds to the substrate to catalyze the folding of newly synthesized proinsulin. So in the presence of low glucose, geniposide enhances the expression of PDI and ERO1, and accelerates the biosynthesis of proinsulin at the same time, but increases the level of H2O2. Once the balance was broken, geniposide plays a feature to decrease the levels of PDI and ERO1, which in turn to reduce the biosynthesis of proinsulin and the production of H2O2, and then help pancreatic β cells restoring the oxidative redox. |
Acknowledgments
The work was supported by grants from the Chongqing Science and Technology Committee (2017jcyjBX0077 and 2018jcyjAX0529) and the Innovative Research Team Development Program at the University of Chongqing (CXTDX201601031).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Zimmet P, Alberti KG, Shaw J. Global and societal implications of the diabetes epidemic. Nature. 2001;414(6865):782–787. doi:10.1038/414782a
2. Zhang E, Mohammed Al-Amily I, Mohammed S, et al. Preserving insulin secretion in diabetes by inhibiting VDAC1 overexpression and surface translocation in beta cells. Cell Metab. 2019;29(1):64–77 e66. doi:10.1016/j.cmet.2018.09.008
3. Shah GN, Morofuji Y, Banks WA, Price TO. High glucose-induced mitochondrial respiration and reactive oxygen species in mouse cerebral pericytes is reversed by pharmacological inhibition of mitochondrial carbonic anhydrases: implications for cerebral microvascular disease in diabetes. Biochem Biophys Res Commun. 2013;440(2):354–358. doi:10.1016/j.bbrc.2013.09.086
4. Lenzen S, Drinkgern J, Tiedge M. Low antioxidant enzyme gene expression in pancreatic islets compared with various other mouse tissues. Free Radic Biol Med. 1996;20(3):463–466. doi:10.1016/0891-5849(96)02051-5
5. Akesson B, Henningsson R, Salehi A, Lundquist I. Islet constitutive nitric oxide synthase and glucose regulation of insulin release in mice. J Endocrinol. 1999;163(1):39–48. doi:10.1677/joe.0.1630039
6. Lundquist I, Alm P, Salehi A, Henningsson R, Grapengiesser E, Hellman B. Carbon monoxide stimulates insulin release and propagates Ca2+ signals between pancreatic beta-cells. Am J Physiol Endocrinol Metab. 2003;285(5):E1055–1063. doi:10.1152/ajpendo.00498.2002
7. Kaneto H, Kajimoto Y, Miyagawa J, et al. Beneficial effects of antioxidants in diabetes: possible protection of pancreatic beta-cells against glucose toxicity. Diabetes. 1999;48(12):2398–2406. doi:10.2337/diabetes.48.12.2398
8. Pi J, Bai Y, Zhang Q, et al. Reactive oxygen species as a signal in glucose-stimulated insulin secretion. Diabetes. 2007;56(7):1783–1791. doi:10.2337/db06-1601
9. Rebelato E, Abdulkader F, Curi R, Carpinelli AR. Control of the intracellular redox state by glucose participates in the insulin secretion mechanism. PLoS One. 2011;6(8):e24507. doi:10.1371/journal.pone.0024507
10. Nardai G, Stadler K, Papp E, Korcsmaros T, Jakus J, Csermely P. Diabetic changes in the redox status of the microsomal protein folding machinery. Biochem Biophys Res Commun. 2005;334(3):787–795. doi:10.1016/j.bbrc.2005.06.172
11. Watson JD. Type 2 diabetes as a redox disease. Lancet. 2014;383(9919):841–843. doi:10.1016/S0140-6736(13)62365-X
12. Lan H, Rabaglia ME, Schueler KL, Mata C, Yandell BS, Attie AD. Distinguishing covariation from causation in diabetes: a lesson from the protein disulfide isomerase mRNA abundance trait. Diabetes. 2004;53(1):240–244. doi:10.2337/diabetes.53.1.240
13. Spanidis Y, Mpesios A, Stagos D, et al. Assessment of the redox status in patients with metabolic syndrome and type 2 diabetes reveals great variations. Exp Ther Med. 2016;11(3):895–903. doi:10.3892/etm.2016.2968
14. Kimura Y, Okuda H, Arichi S. Effects of geniposide isolated from Gardenia jasminoides on metabolic alterations in high sugar diet-fed rats. Chem Pharm Bull (Tokyo). 1982;30(12):4444–4447. doi:10.1248/cpb.30.4444
15. Liu J, Yin F, Zheng X, Jing J, Hu Y. Geniposide, a novel agonist for GLP-1 receptor, prevents PC12 cells from oxidative damage via MAP kinase pathway. Neurochem Int. 2007;51(6–7):361–369. doi:10.1016/j.neuint.2007.04.021
16. Xu B, Li YL, Xu M, et al. Geniposide ameliorates TNBS-induced experimental colitis in rats via reducing inflammatory cytokine release and restoring impaired intestinal barrier function. Acta Pharmacol Sin. 2017;38(5):688–698. doi:10.1038/aps.2016.168
17. Wu SY, Wang GF, Liu ZQ, et al. Effect of geniposide, a hypoglycemic glucoside, on hepatic regulating enzymes in diabetic mice induced by a high-fat diet and streptozotocin. Acta Pharmacol Sin. 2009;30(2):202–208. doi:10.1038/aps.2008.17
18. Guo LX, Liu JH, Zheng XX, Yin ZY, Kosaraju J, Tam KY. Geniposide improves insulin production and reduces apoptosis in high glucose-induced glucotoxic insulinoma cells. Eur J Pharm Sci. 2017;110:70–76. doi:10.1016/j.ejps.2017.03.038
19. Guo LX, Xia ZN, Gao X, Yin F, Liu JH. Glucagon-like peptide 1 receptor plays a critical role in geniposide-regulated insulin secretion in INS-1 cells. Acta Pharmacol Sin. 2012;33(2):237–241. doi:10.1038/aps.2011.146
20. Hao Y, Liu C, Yin F, Zhang Y, Liu J. 5ʹ-AMP-activated protein kinase plays an essential role in geniposide-regulated glucose-stimulated insulin secretion in rat pancreatic INS-1 beta cells. J Nat Med. 2017;71(1):123–130. doi:10.1007/s11418-016-1038-5
21. Liu CY, Hao YN, Yin F, Zhang YL, Liu JH. Geniposide accelerates proteasome degradation of Txnip to inhibit insulin secretion in pancreatic beta-cells. J Endocrinol Invest. 2017;40(5):505–512. doi:10.1007/s40618-016-0591-9
22. Liu J, Guo L, Yin F, Zhang Y, Liu Z, Wang Y. Geniposide regulates glucose-stimulated insulin secretion possibly through controlling glucose metabolism in INS-1 cells. PLoS One. 2013;8(10):e78315. doi:10.1371/journal.pone.0078315
23. Fridlyand LE, Philipson LH. Does the glucose-dependent insulin secretion mechanism itself cause oxidative stress in pancreatic beta-cells? Diabetes. 2004;53(8):1942–1948. doi:10.2337/diabetes.53.8.1942
24. Al-Amily IM, Duner P, Groop L, Salehi A. The functional impact of G protein-coupled receptor 142 (Gpr142) on pancreatic beta-cell in rodent. Pflugers Arch. 2019;471(4):633–645. doi:10.1007/s00424-019-02262-7
25. Panus PC, Radi R, Chumley PH, Lillard RH, Freeman BA. Detection of H2O2 release from vascular endothelial cells. Free Radic Biol Med. 1993;14(2):217–223. doi:10.1016/0891-5849(93)90013-K
26. Nakano T, Watarai H, Liu YC, Oyama Y, Mikayama T, Ishizaka K. Conversion of inactive glycosylation inhibiting factor to bioactive derivatives by modification of a SH group. Proc Natl Acad Sci U S A. 1997;94(1):202–207. doi:10.1073/pnas.94.1.202
27. Jones DP. Redox potential of GSH/GSSG couple: assay and biological significance. Methods Enzymol. 2002;348:93–112.
28. Schafer FQ, Buettner GR. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic Biol Med. 2001;30(11):1191–1212. doi:10.1016/S0891-5849(01)00480-4
29. Eletto D, Eletto D, Boyle S, Argon Y. PDIA6 regulates insulin secretion by selectively inhibiting the RIDD activity of IRE1. FASEB J. 2016;30(2):653–665. doi:10.1096/fj.15-275883
30. Karas JA, Scanlon DB, Forbes BE, et al. 2-nitroveratryl as a photocleavable thiol-protecting group for directed disulfide bond formation in the chemical synthesis of insulin. Chemistry. 2014;20(31):9549–9552. doi:10.1002/chem.201403574
31. Benham AM, van Lith M, Sitia R, Braakman I. Ero1-PDI interactions, the response to redox flux and the implications for disulfide bond formation in the mammalian endoplasmic reticulum. Philos Trans R Soc Lond B Biol Sci. 2013;368(1617):20110403. doi:10.1098/rstb.2011.0403
32. Zito E. ERO1: a protein disulfide oxidase and H2O2 producer. Free Radic Biol Med. 2015;83:299–304. doi:10.1016/j.freeradbiomed.2015.01.011
33. Zito E, Chin KT, Blais J, Harding HP, Ron D. ERO1-beta, a pancreas-specific disulfide oxidase, promotes insulin biogenesis and glucose homeostasis. J Cell Biol. 2010;188(6):821–832. doi:10.1083/jcb.200911086
34. Rajpal G, Schuiki I, Liu M, Volchuk A, Arvan P. Action of protein disulfide isomerase on proinsulin exit from endoplasmic reticulum of pancreatic beta-cells. J Biol Chem. 2012;287(1):43–47. doi:10.1074/jbc.C111.279927
35. Mailloux RJ, Fu A, Robson-Doucette C, et al. Glutathionylation state of uncoupling protein-2 and the control of glucose-stimulated insulin secretion. J Biol Chem. 2012;287(47):39673–39685. doi:10.1074/jbc.M112.393538
36. Liu X, Han S, Yang Y, Kang J, Wu J. Glucose-induced glutathione reduction in mitochondria is involved in the first phase of pancreatic beta-cell insulin secretion. Biochem Biophys Res Commun. 2015;464(3):730–736.
37. Chakravarthi S, Bulleid NJ. Glutathione is required to regulate the formation of native disulfide bonds within proteins entering the secretory pathway. J Biol Chem. 2004;279(38):39872–39879. doi:10.1074/jbc.M406912200
38. Hao Y, Shen S, Yin F, Zhang Y, Liu J. Unfolded protein response is involved in geniposide-regulating glucose-stimulated insulin secretion in INS-1 cells. Cell Biochem Funct. 2019;37(5):368–376. doi:10.1002/cbf.v37.5
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.