Back to Journals » International Journal of Women's Health » Volume 17
Genetically Predicted Causal Risk Factors for Preeclampsia: A Comprehensive Mendelian Randomization Study
Authors Wu H, Sun D, Fang S, Pan Q, Yue C
Received 11 August 2025
Accepted for publication 21 November 2025
Published 3 December 2025 Volume 2025:17 Pages 5105—5116
DOI https://doi.org/10.2147/IJWH.S559901
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Vinay Kumar
Han Wu,1,* Dongmei Sun,2,* Sijia Fang,3 Qiuhui Pan,1,4– 6 Chaoyan Yue3
1Department of Clinical Laboratory, Shanghai Children’s Medical Center, School of Medicine, Shanghai Jiao Tong University, Shanghai, People’s Republic of China; 2Department of Obstetrics and Gynecology, Xuyi People’s Hospital, Huai’an, Jiangsu, People’s Republic of China; 3Obstetrics & Gynecology Hospital of Fudan University, Shanghai Key Laboratory of Reproduction and Development, Shanghai Key Laboratory of Female Reproductive Endocrine Related Diseases, Shanghai, People’s Republic of China; 4Shanghai Key Laboratory of Clinical Molecular Diagnostics for Pediatrics, Shanghai, People’s Republic of China; 5Hainan Branch, Shanghai Children’s Medical Center, Shanghai Jiao Tong University School of Medicine, Sanya, Hainan, People’s Republic of China; 6Faculty of Medical Laboratory Science, College of Health Science and Technology, Shanghai Jiao Tong University School of Medicine, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Chaoyan Yue, Obstetrics & Gynecology Hospital of Fudan University, Shanghai Key Laboratory of Reproduction and Development, Shanghai Key Laboratory of Female Reproductive Endocrine Related Diseases, Fangxie Road No. 419, Huangpu District, Shanghai, 200011, People’s Republic of China, Email [email protected] Qiuhui Pan, Department of Clinical Laboratory, Shanghai Children’s Medical Center, School of Medicine, Shanghai Jiao Tong University, Dongfang Road No. 1678, Pudong New District, Shanghai, 200127, People’s Republic of China, Email [email protected]
Background: Preeclampsia is a complex hypertensive disorder of pregnancy, significantly impacting maternal and fetal health worldwide. Despite extensive research, its pathogenesis, involving inflammatory, immune, microbiological, and metabolic factors, requires comprehensive elucidation.
Methods: This study applied Mendelian randomization (MR) to investigate causal relationships between multi-omics traits and the risk of preeclampsia. The genome-wide association studies (GWAS) datasets used consisted of immune cells (N = 3757), inflammatory factors (N = 14,824), gut microbiota (N = 7738), circulating metabolites (N 1 = 7824, N 2 = 8299), plasma proteins (N = 3301), and preeclampsia (7212 cases, 194,266 controls). The inverse variance-weighted method was used in the main analysis, and the weighted median, weighted mode, and MR Egger regression were used in sensitivity analyses.
Results: Our analysis identified 81 potential causal factors for preeclampsia. Among the most novel and clinically significant findings were several druggable plasma proteins, including Astacin-like metalloendopeptidase (ASTL) and Baculoviral IAP repeat-containing protein 3 (BIRC3), which exhibited strong causal evidence. Furthermore, we identified specific gut microbiota genera, notably Bifidobacterium, as potential protective factors. We also validated the causal roles of key metabolic disturbances, like cysteine and guanidinoacetate, and dysfunctions in specific immune cell populations, particularly regulatory T and B cells.
Conclusion: These findings highlight the intricate interplay of immune, inflammatory, microbiological, metabolic, and protein factors in preeclampsia, suggesting novel diagnostic and therapeutic targets. Further research is warranted to explore these associations in detail.
Keywords: causality, gut microbiome, inflammatory factor, Mendelian randomization, metabolome, multi-omics, preeclampsia
Introduction
Preeclampsia, a hypertensive complication of pregnancy affecting 2–8% of pregnancies,1 remains the leading cause of maternal and fetal morbidity and mortality worldwide, with substantial influence on the risk of long-term cardiovascular disease and cerebrovascular disease.2 In accordance with the definition of the American College of Obstetrics and Gynecology (ACOG), preeclampsia is characterized by the onset of hypertension and albuminuria or other end-organ damage emerging subsequent to the 20th week of gestation in individuals previously normotensive.3 The pathogenesis of preeclampsia involves various angiogenic, inflammatory, immune and metabolic pathways, encompassing spiral artery remodeling, oxidative stress, immune abnormality at the maternal-fetal interface, as well as the balance of angiogenic and antiangiogenic factors.4–6 Despite considerable research endeavors and substantial progress, the precise mechanisms underlying the pathogenesis of preeclampsia continue to be intricate and multifaceted and need further investigation.7
Impaired trophoblast invasion and incomplete spiral artery remodeling raise angiogenic marker levels, such as soluble fms-like tyrosine kinase-1 (sFlt-1) and soluble endoglin (sEng),8,9 which lower vascular endothelial growth factor (VEGF) and others, leading to endothelial dysfunction and complications like oxidative stress.10 Immune cell balance disruptions, especially with T helper cells and natural killer cells,11,12 and the imbalance between pro- and anti-inflammatory factors,13,14 can contribute to preeclampsia’s development. Moreover, proinflammatory cytokines11 and chemokines15,16 disrupt vascular stability, affecting pregnancies. Additionally, gut dysbiosis impacts metabolic syndrome and complicated pregnancies,17 with conditions like diabetes and obesity increasing preeclampsia risks.12,18 However, the majority of available findings are observational and incomplete, emphasizing the necessity for comprehensive and systematic studies to elucidate the multi-faceted factors contributing to the pathogenesis of preeclampsia.
The selection of specific exposures in this study, including immune cells, inflammatory factors, gut microbiome, metabolome, and proteome, was based on their established links to the pathophysiological pathways of preeclampsia outlined above. However, while observational evidence is abundant, and previous Mendelian randomization (MR) studies have investigated discrete risk factors for preeclampsia, these studies are often focused on a single domain or a limited set of traits.19–23 This fragmented approach makes it difficult to compare the relative importance of different pathways and to obtain a unified understanding of the disease’s causal architecture. Therefore, a comprehensive, multi-omics MR study that simultaneously interrogates these interconnected biological layers is needed to address this gap and systematically identify the most pivotal causal factors.
MR analysis is an epidemiologic method that utilizes genetic variations of risk factors as instrumental variables (IVs) to assess the causal relationship between exposures and outcomes.24,25 The principle underlying MR relies on the random assortment of genetic alleles during gamete formation, thereby minimizing biases from confounding factors and reverse causation.26 In the current study, we employed MR analysis as a tool to investigate the causal associations between preeclampsia and various risk factors, including immune cells, inflammatory factors, gut microbiome, metabolome, and proteome. By integrating evidence across these domains, this study aims to provide a comprehensive causal atlas of preeclampsia, which holds promise for advancing diagnostic precision and developing targeted therapeutic strategies, aiming to alleviate the impact of preeclampsia on maternal and fetal health.
Methods
Data Sources
The genetic instruments of 731 immune cells traits were procured from a prior genome-wide association study (GWAS) dataset derived from a cohort of 3757 individuals in Sardinia, Italy. This extensive dataset encompassed 539 immune traits (118 absolute cell counts, 389 median fluorescence intensities of surface antigens, and 32 morphological parameters) and 192 relative counts (ratios between cell levels).27,28 The donor’s peripheral blood was collected in heparin tubes and analyzed with standardized BD FACSCanto II flow cytometers and BD FACSDiva software (BD Biosciences).27 Single-nucleotide polymorphisms (SNPs) associated with inflammatory factors were extracted from GWAS statistics covering 91 plasma inflammatory proteins measured across 11 cohorts totaling 14,824 participants of European ancestry and meta-analyzed by Zhao et al.29 In the case of gut microbiota, IVs were derived from GWAS dataset generated in a previous study exploring 412 microbial compositions and functions (207 taxa and 205 pathways) from 7738 European participants in the Dutch Microbiome Project (DMP).30 Genetic instruments related to circulating metabolites were sourced from two distinct studies: one involving 486 circulating metabolites in 7824 European cases from the Cooperative Health Research in the Region of Augsburg (KORA) and TwinsUK in which the concentrations of metabolites were detected using ultrahigh performance liquid-phase chromatography and gas chromatography separation coupled with tandem mass spectrometry in either plasma or serum,31 and the other comprising 1091 plasma metabolites and 309 metabolite ratios in 8299 European subjects from the Canadian Longitudinal Study of Aging (CLSA) in which the levels of metabolites were quantified using Metabolon HD4 platform.32 SNPs for human plasma proteome were derived from GWAS data of previous research encompassing 3622 plasma proteins or protein complexes in 3301 British participants from the INTERVAL study.33 The GWAS summary-level data concerning preeclampsia (GWAS ID: O15_PRE_OR_ECLAMPSIA, N = 7212 cases, N = 194,266 controls) were obtained from the FinnGen consortium R9 release dataset (https://r9.risteys.finngen.fi/).
Genetic Instruments Selection
The conventional genome-wide significance threshold (P < 5 × 10−8) yielded an insufficient number of independent IVs for several exposure datasets with relatively limited GWAS sample sizes (notably, plasma proteins, immune cells, and inflammatory factors), often resulting in fewer than 10 usable SNPs per exposure. To ensure adequate statistical power for a robust MR analysis, we adopted slightly relaxed, exposure-specific P-value thresholds, a well-established practice in previous MR studies.34–36 Specifically, the threshold criteria at P < 1×10−5 for gut microbiota and metabolites from CLSA. Concurrently, the thresholds for immune cells, inflammatory factors, 486 metabolites, and proteins were set as P < 5×10−6. The removal of linkage disequilibrium (LD) among SNPs was conducted with a stringent clump threshold (r2 = 0.001, kb = 10 Mb). To rigorously mitigate potential bias introduced by the use of relaxed thresholds, the F-statistic for the chosen SNPs was calculated to ascertain the robustness of the genetic variants selected. All F-statistics above 10 indicated the absence of weak instrument bias. SNPs strongly associated with preeclampsia (P < 5×10−5) would be discarded to avoid reverse causalities preliminarily. In the process of harmonizing the effect allele of the exposures and outcome, SNPs exhibiting inconsistency or palindromic traits were excluded from the analysis.
Statistical Analysis and Sensitivity Analysis
The flowchart of this MR analysis was shown in Figure 1. In this investigation, the inverse variance weighted (IVW) statistical model served as the primary methodology for Mendelian randomization (MR) analysis, which estimates the summary-level effect by meta-analyzing the individual SNP Wald ratios.37 Besides, we employed the MR-Egger, weighted median estimator (WME), and weighted mode as supplementary techniques to address issues of heterogeneity and pleiotropy within genetic variants. Our investigation assessed horizontal pleiotropy utilizing the MR-Egger intercept, considering P > 0.05 as indicative of the absence of horizontal pleiotropy. The evaluation of heterogeneity among genetic variants was conducted utilizing Cochran’s Q-statistic test (MR-IVW). Depending on the significance level derived from the Cochran’s Q-statistic test, we employed either a multiplicative random-effects model (P < 0.05) or a fixed-effects model (P ≥ 0.05) of IVW. The causal estimations of immune cells, inflammatory factors, gut microbiota, metabolites, and proteins on preeclampsia were presented as odds ratios (ORs) with 95% confidence intervals (CIs) per one unit increase in the levels of exposure. We implemented a threshold of P < 0.0125 (0.05/4) to indicate statistical significance after correction. Meanwhile, a cutoff of P < 0.05, falling above the Bonferroni correction threshold, was designated as indicating a suggestive significant association. The statistical analysis was conducted utilizing R (4.3.0) (https://www.r-project.org/) and the TwoSampleMR package.
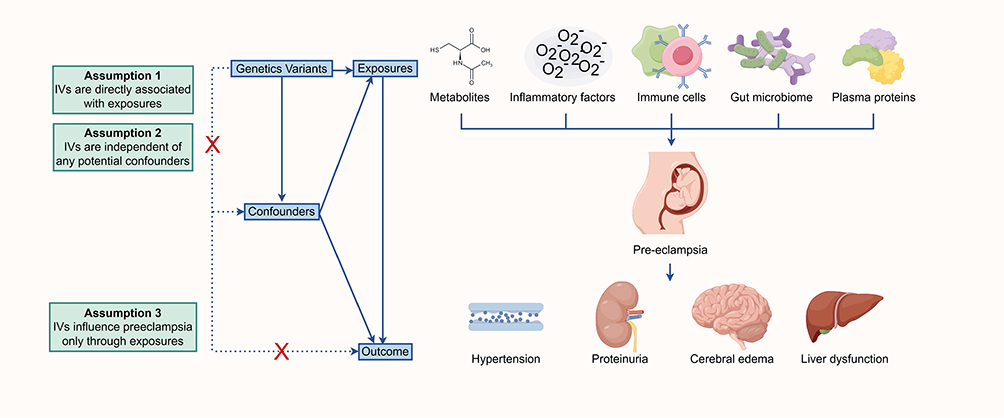 |
Figure 1 Study design and workflow of Mendelian randomization for exposures on preeclampsia. Created with Figdraw.com. Abbreviation: IVs, instrumental variables. |
Results
Causal Relationship Between Immune Cells and Preeclampsia
To search for possible associations between multiple types of immune cell indexes and the risk of preeclampsia, we performed MR analyses of 731 immune cells traits on the risk of preeclampsia (Supplementary Table 1). We identified 17 immune cell traits causally associated with the onset of preeclampsia by IVW method (Figure 2). To prioritize the most reliable findings, we focused on associations that were nominally significant (P < 0.05) and exhibited consistent direction of effect across all sensitivity analyses (MR-Egger, weighted median, and weighted mode). The results revealed that a one-standard deviation (SD) increase in CD62L− monocyte %monocyte was associated with the most substantial reduction in preeclampsia risk [OR = 0.90, 95% CI: 0.83–0.97, P = 0.004] among the protective factors. Conversely, a one-SD increase in BAFF-R on IgD− CD38+ B cell represented the strongest risk factor, associated with a 15% increase in risk [OR = 1.15, 95% CI: 1.05–1.25, P = 0.002].
 |
Figure 2 Mendelian randomization and sensitivity results for immune cell traits and inflammatory factors on preeclampsia in the form of forest plot. Abbreviations: SNP, single-nucleotide polymorphism; Beta, the effect size of the exposure on outcome; CI, confidence interval; IVW, inverse variance weighted. |
Notably, the association for CD62L− monocyte %monocyte showed significant heterogeneity (Cochran’s Q P < 0.05). While this heterogeneity prompted the use of a multiplicative random-effects model, which provides a more conservative effect estimate, it may also suggest that the causal effect varies across different populations or genetic backgrounds, a point that warrants further investigation. No significant horizontal pleiotropy was detected for this or any other immune cell trait (MR-Egger intercept P > 0.05), strengthening the credibility of a direct causal relationship.
Causal Relationship Between Inflammatory Factors and Preeclampsia
To search for possible associations between multiple types of inflammatory factors and the risk of preeclampsia, we performed MR analyses of 91 circulating inflammatory proteins on the risk of preeclampsia (Supplementary Table 2). In the result of IVW method, a one-SD increase in C-C motif chemokine 19 (CCL19) levels could reduce the risk of preeclampsia [OR = 0.89, 95% CI: 0.82–0.96, P = 0.003] (Figure 2). Cochran’s Q statistics (P > 0.05) and MR-Egger intercept (P > 0.05) demonstrated the nonexistence of heterogeneity and horizontal pleiotropy.
Causal Relationship Between Gut Microbiome and Preeclampsia
To search for possible associations between multiple types of gut microbiota and the risk of preeclampsia, we performed MR analyses of 412 microbial compositions and functions on the risk of preeclampsia (Supplementary Table 3). We identified 9 microbiological traits causally associated with the onset of preeclampsia by IVW method (Figure 3). A one-SD increase in the relative abundance of 7 microbial taxa was associated with a reduced risk of preeclampsia. Among these protective factors, the association with Coprococcus_catus demonstrated the strongest effect [OR = 0.80, 95% CI: 0.69–0.94, P = 0.006]. Notably, 3 taxa from the Bifidobacterium genus and its higher taxonomic orders (family Bifidobacteriaceae, order Bifidobacteriales) consistently showed protective effects with similar effect sizes [ORs approximately 0.82–0.83]. Conversely, 2 taxa were associated with an increased risk of preeclampsia. The largest effect observed across all findings was for Ruminococcaceae, which was associated with a 28% increase in risk [OR = 1.28, 95% CI: 1.07–1.53, P = 0.006]. Cochran’s Q statistics (P > 0.05) and MR-Egger intercept (P > 0.05) demonstrated no heterogeneity and horizontal pleiotropy.
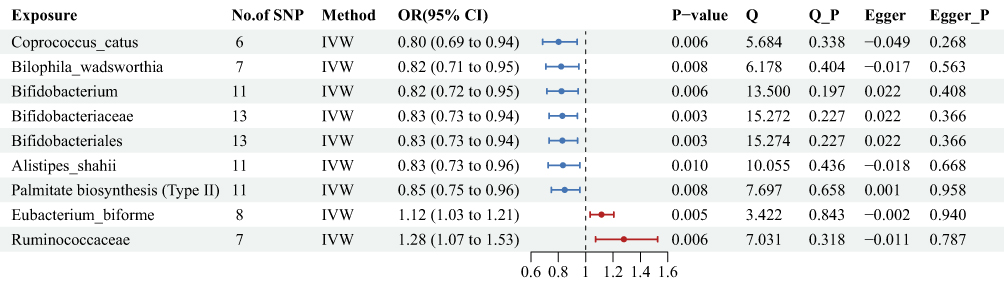 |
Figure 3 Mendelian randomization and sensitivity results for gut microbiome on preeclampsia in the form of forest plot. Abbreviations: SNP, single-nucleotide polymorphism; Beta, the effect size of the exposure on outcome; CI, confidence interval; IVW, inverse variance weighted. |
Causal Relationship Between Circulating Metabolites and Preeclampsia
To search for possible associations between multiple types of metabolites and the risk of preeclampsia, we performed MR analyses of 486 circulating metabolites as well as 1440 metabolites and ratios on the risk of preeclampsia, respectively.
The first analysis of 486 metabolites identified 6 significant associations (Figure 4, Supplementary Table 4). The most pronounced association was a 79% reduction in preeclampsia risk per log10 unit increase in the unidentified metabolite X-05907 [OR = 0.21, 95% CI: 0.09–0.49, P < 0.001], representing the strongest protective effect across all metabolite analyses. Conversely, 2-linoleoylglycerophosphocholine* was the strongest risk factor [OR = 2.25, 95% CI: 1.25–4.04, P = 0.007]. No significant heterogeneity (P > 0.05) or pleiotropy (P > 0.05) was detected for these associations.
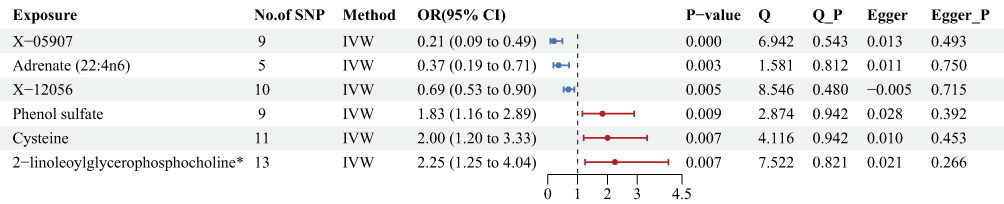 |
Figure 4 Mendelian randomization and sensitivity results for 486 metabolites on preeclampsia in the form of forest plot. * indicates metabolites for which reference spectra of the pure substances were not directly measured on the Metabolon platform. Abbreviations: SNP, single-nucleotide polymorphism; Beta, the effect size of the exposure on outcome; CI, confidence interval; IVW, inverse variance weighted. |
The second analysis of 1400 metabolic traits revealed 16 significant associations (11 metabolites and 5 ratios; Figure 5 and Supplementary Table 5). Among these, guanidinoacetate levels showed the strongest risk-increasing effect [OR = 1.15, 95% CI: 1.06–1.25, P = 0.001], while the serine to alpha-ketobutyrate ratio was the most robust protective factor [OR = 0.87, 95% CI: 0.78–0.96, P = 0.009]. A finding was the significant heterogeneity for hydroxypalmitoyl sphingomyelin (d18:1/16:0(OH)) levels (Cochran’s Q P < 0.05), which was appropriately analyzed using a multiplicative random-effects model. This heterogeneity suggests the causal effect of this metabolite may vary across populations. No horizontal pleiotropy (P > 0.05) was observed for any trait.
 |
Figure 5 Mendelian randomization and sensitivity results for 1400 metabolic traits on preeclampsia in the form of forest plot. Abbreviations: SNP, single-nucleotide polymorphism; Beta, the effect size of the exposure on outcome; CI, confidence interval; IVW, inverse variance weighted. |
Causal Relationship Between Circulating Proteins and Preeclampsia
To search for possible associations between multiple types of proteins and the risk of preeclampsia, we performed MR analyses of 3622 plasma proteins or protein complexes on the risk of preeclampsia. We identified 32 proteins causally associated with the risk of preeclampsia by IVW method (Figure 6 and Supplementary Table 6). A one natural log-transformed unit increase in genetically predicted levels of 17 proteins was associated with an elevated risk, while 15 proteins were associated with a reduced risk. Notably, the most substantial effect was observed for Astacin-like metalloendopeptidase (ASTL), which demonstrated the strongest protective association [OR = 0.84, 95% CI: 0.74–0.96, P = 0.009]. Among the risk factors, Baculoviral IAP repeat-containing protein 3 (BIRC3) showed the strongest effect [OR = 1.19, 95% CI: 1.08–1.31, P < 0.001]. Cochran’s Q statistics (P > 0.05) and MR-Egger intercept (P > 0.05) demonstrated no heterogeneity and horizontal pleiotropy.
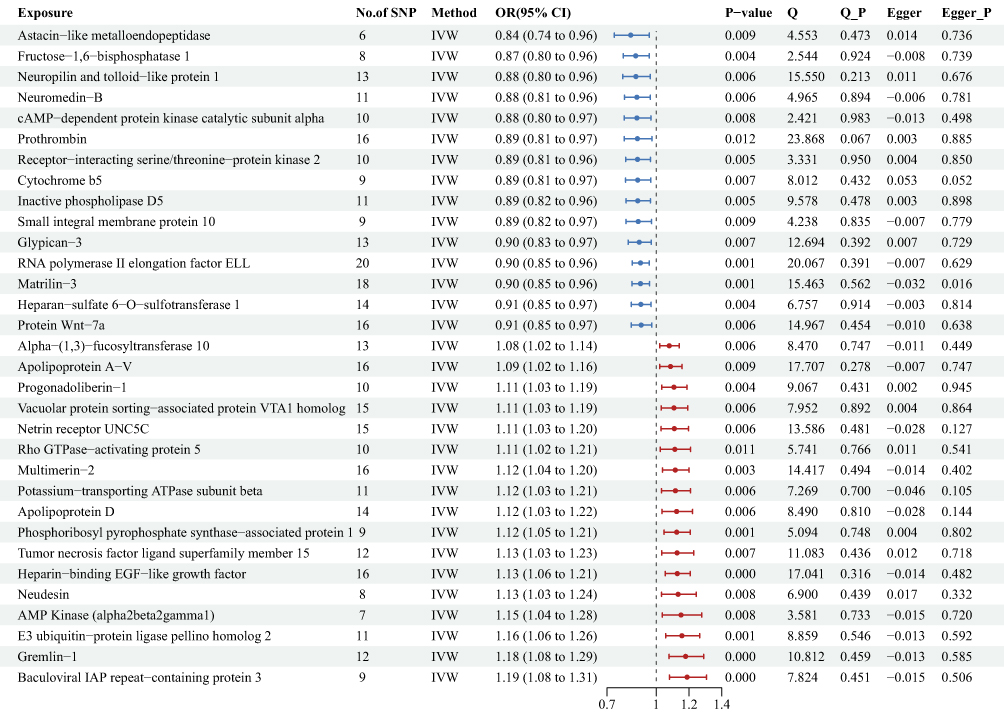 |
Figure 6 Mendelian randomization and sensitivity results for 3622 proteins on preeclampsia in the form of forest plot. Abbreviations: SNP, single-nucleotide polymorphism; Beta, the effect size of the exposure on outcome; CI, confidence interval; IVW, inverse variance weighted. |
Discussion
In our two-sample MR study, we conducted a thorough examination of the causal relationships between a diverse array of exposures (immune cells, inflammatory factors, gut microbiome, metabolome, and proteome) and the risk of preeclampsia. We identified 17 immune, 1 inflammatory, 9 gut microbiotic, 22 metabolic, and 32 proteinic traits that are potentially causally implicated in the disease. This multi-omics perspective allows for a more integrated understanding of preeclampsia’s pathophysiology. It is important to note, however, that while these findings provide robust evidence for a potential causal role, they do not constitute definitive proof.
Abnormal immune responses and pro-inflammatory cytokines are pivotal in the pathogenesis of preeclampsia. Regulatory B (Breg) cells, particularly CD24hi CD27+ cells,38 and CD4+ CD25+ regulatory T (Treg) cells,39 are vital in maintaining immune balance, with their dysregulation contributing to disease development. The protective effect associated with reduced CD62L expression likely stems from its function in impairing immune cell homing,40 which could attenuate pathogenic inflammation at the placenta. Observational studies have identified elevated serum B-cell-activating factor of the tumor necrosis factor family (BAFF) levels in preeclampsia patients during pregnancy, indicating its potential role as a risk factor for preeclampsia.41 CD3+ CD8+ CD28− inhibitory Treg cells exert inhibitory effects on T cell proliferation and cytotoxicity, contributing to immune tolerance.42 Human myeloid-derived suppressor cells (MDSCs), expressing CD33, possess the capability to induce Tregs, thereby indirectly suppressing immune responses. CD66b monocytes demonstrate a pro-inflammatory profile characterized by their ability to promote T cell proliferation and induce interferon-gamma (IFN-γ) secretion.43 Circulating natural killer (NK) cells and dendritic cells (DCs) may play a substantive role in the immune responses associated with preeclampsia. Plasmacytoid DCs (pDCs) are particularly noteworthy for their potential importance in maintaining immune tolerance and the induction of Treg cells.44 CCL19 plays a pivotal role in intensifying the immunosuppressive capacities of Treg, contributing to the potential amelioration of the atypical immune milieu associated with preeclampsia.45
The gut microbiota is intricately linked to host immune regulation, inflammatory state, metabolic syndrome, and pregnancy.46,47 Short-chain fatty acids (SCFAs) produced by specific gut bacteria, such as Coprococcus_catus48 and Bifidobacteriales,49 are linked to cardiovascular health benefits and potentially lower preeclampsia risk. SCFAs, particularly butyrate, are pivotal for maintaining immune homeostasis. They exert their protective effects by (1) promoting the differentiation and function of Tregs, which suppresses excessive inflammation, and (2) strengthening the intestinal epithelial barrier, thereby reducing the translocation of pro-inflammatory molecules into the systemic circulation. Since systemic maternal inflammation and immune dysregulation are hallmarks of preeclampsia, this SCFA-driven immunomodulatory effect represents a primary mechanism for risk reduction. Notably, despite being an opportunistic bacterium, some species in the genus Bilophila have been identified as symbiotic bacteria, potentially mitigating the risk of cardiovascular disease by averting trimethylamine-N-oxide (TMAO) production induced by animal diets.50 Besides, previous research has documented a noteworthy augmentation in the abundance of Alistipes_shahii in mice afflicted with hepatocellular carcinoma, subjected to probiotic treatment. Alistipes_shahii is posited to harbor potential immunosuppressive and anti-inflammatory properties. Ruminococcus2 shows a robust correlation with human leptin levels, a factor directly implicated in the pathogenesis of preeclampsia. Studies have ascertained a positive correlation between Ruminococcus2 and blood pressure levels in preeclampsia patients,49 suggesting a potential contributory role of Ruminococcaceae as a risk factor for preeclampsia.
Metabolic dysfunctions, including irregular glucose and lipid metabolism, increase preeclampsia risk, with diabetes mellitus as a known contributor.5 Serine is a key source of one-carbon units essential for nucleotide synthesis and methylation reactions, and its metabolic disruption is implicated in various vascular pathologies.51 Maintaining a specific balance in this amino acid metabolic pathway may be crucial for gestational vascular homeostasis. Phenyl sulfate, a uremic toxin originated from gut microbiota, exerts an inhibition effect on endothelial repair mechanisms subsequent to injury, and directly precipitates vascular damage. This process can escalate the risk of cardiovascular complications.52 Observational research has demonstrated that plasma levels of cysteine in pregnant women and fetuses diagnosed with preeclampsia are significantly elevated when compared to those in normal control groups. Cysteine is posited to be linked with the onset of cardiovascular disease and endothelial dysfunction.53 Moreover, research has substantiated that the accumulation of guanidinoacetate in substantial quantities functions as a potent pro-oxidant, thereby inducing oxidative stress.54
ASTL, a zinc-dependent metalloproteinase, is critically involved in embryo development through its proteolytic activity on the zona pellucida.55 The protective association of higher ASTL levels with preeclampsia risk found in our study implies its potential role in influencing key substrates related to placental development or vascular remodeling. BIRC3 is a key negative regulator of NF-κB signaling, a pathway critically linked to inflammation and preeclampsia risk. Dysregulation of BIRC3 may therefore lead to uncontrolled inflammation, directly contributing to the pathogenesis of preeclampsia.56 The enzyme fructose-1,6-bisphosphatase 1 (FBP1) is vital for glucose regulation,57 and the cAMP pathway is essential for cardiovascular health, affecting hypertension risk.58 The role of GPC3 in Wnt pathway activation is crucial for trophoblast invasions and a healthy pregnancy.5 Research highlights the significance of RNA polymerase II elongation factor (ELL) in blood supply59 and HS6ST-1 in endothelial function via VEGFA.60 Apolipoprotein A-V (APOA5) levels are associated with hypertriglyceridemia and atherosclerosis, indicating a preeclampsia risk.61 Neudesin (NENF) disrupts glucose metabolism in GDM,62 and Gremlin-1 (GREM1) overexpression in type II diabetes affects fat function and insulin resistance.63 Multimerin-2 (MMRN2) hinders angiogenesis and cell migration, potentially impairing placental blood flow.64 Tumor necrosis factor ligand superfamily member 15 (TNFSF15)65 and E3 ubiquitin-protein ligase pellino homolog 2 (PELI2)66 influence endothelial proliferation and inflammation, respectively, affecting preeclampsia development.
To present a balanced view, it is important to acknowledge that the literature contains contrasting findings. For example, while our analysis suggests a risk-increasing effect of Heparin-binding EGF-like growth factor (HB-EGF), this protein is generally considered vital for placental development.67 This apparent contradiction could be explained by its context-dependent dual roles, where excessive signaling in later gestation becomes pathological. This discrepancy may arise from differences in study design, cell-type specificity, or the limitations of MR in capturing complex cellular interactions present in observational data.
Different from conventional observational studies, our research exploits the GWAS datasets of large sample sizes, enhancing the reliability of our conclusions. Moreover, the use of MR analysis aids in minimizing confounding variables and the potential for reverse causality, thereby improving result accuracy. Nevertheless, several limitations must be acknowledged. First, the use of relaxed P-value thresholds, while necessary to secure sufficient IVs for exposures with limited GWAS sample sizes, increases the risk of false-positive findings. We mitigated this by employing a multi-tiered strategy: calculating F-statistics to ensure robust instruments (F > 10), requiring consistent effect directions across multiple MR methods, and applying stringent LD clumping. Second, a potential limitation is the multiple testing issue inherent in our multi-omics exploratory analysis. While a standard Bonferroni correction for all exposures would be overly conservative due to the high correlation among them, we adopted a significance threshold of P < 0.0125 (0.05/4) based on the number of MR methods used. Although we supplemented this with consistency and sensitivity analyses to enhance robustness, we acknowledge that this approach does not constitute a formal multiple testing correction and that the risk of false positives may be higher than if a more stringent, trait-wide correction had been applied. Third, despite sensitivity analyses showing no significant horizontal pleiotropy, we cannot completely rule out this potential bias, as MR-Egger regression can lack power. Fourth, the potential for population stratification exists, as our analysis primarily relied on GWAS data from European-ancestry populations, limiting the generalizability of our findings. Finally, the GWAS sample sizes for some exposures, particularly immune cells and plasma proteins, were relatively limited, which may affect the precision of our causal estimates.
The identification of putative causal factors across immunologic, microbial, metabolic, and proteomic domains highlights the potential of integrating these diverse layers of biology for superior risk prediction. A model combining indicators of immunological predisposition, metabolic status, and proteomic pathway activity would provide a more robust risk assessment than a single-modality approach. For example, it could more accurately stratify risk in an individual with only a subclinical immunological aberration by concurrently capturing their metabolic and proteomic profile. This multi-parametric strategy is promising for enabling earlier and more precise identification of high-risk pregnancies.
In conclusion, our integrated MR analysis uncovers a network of putative causal factors across multiple biological layers, offering novel insights into the complex etiology of preeclampsia. These findings not only reinforce the roles of immune and vascular pathways but also highlight the emerging significance of gut microbiota and specific metabolic disturbances. The putative causal proteins and metabolites identified serve as valuable candidates for future research into diagnostic biomarkers and therapeutic targets. Future studies should focus on validating these potential associations in diverse populations and elucidating the precise molecular mechanisms underlying these genetic associations.
Data Sharing Statement
The dataset analyzed during the current study are available from the IEU Open GWAS project (https://gwas.mrcieu.ac.uk/) and the FinnGen consortium R9 release dataset (https://r9.risteys.finngen.fi/).
Ethical Approval
Ethical exemption approval for this study was obtained from the Ethics Committee of Obstetrics & Gynecology Hospital of Fudan University. The Ethics Committee granted an exemption because this research involved the secondary analysis of existing, de-identified, and publicly available data. The ethical approvals and participant consents for the original GWAS can be found in their respective primary publications.
Acknowledgments
The authors are thankful for all the participants and investigators for their contributions to the GWAS data.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was financially supported by the Natural Science Foundation of Shanghai Basic Research Program [25ZR1402039] and the Special Project for Scientific Research Development of Off-campus Teaching Base, Jiangsu Medical College [20229150].
Disclosure
The authors report no conflicts of interest in this work.
References
1. Pre-eclampsia: World Health Organization. 2025. Available from: https://www.who.int/zh/news-room/fact-sheets/detail/pre-eclampsia.
2. Coutinho T, Lamai O, Nerenberg K. Hypertensive disorders of pregnancy and cardiovascular diseases: current knowledge and future directions. Curr Treat Options Cardiovasc Med. 2018;20:56. doi:10.1007/s11936-018-0653-8
3. Hypertension in pregnancy. Report of the American College of Obstetricians and Gynecologists’ Task Force on Hypertension in Pregnancy. Obstet Gynecol. 2013;122:1122–1131. doi:10.1097/01.AOG.0000437382.03963.88
4. Phipps EA, Thadhani R, Benzing T, Karumanchi SA. Pre-eclampsia: pathogenesis, novel diagnostics and therapies. Nat Rev Nephrol. 2019;15:275–289. doi:10.1038/s41581-019-0119-6
5. Rana S, Lemoine E, Granger JP, Karumanchi SA. Preeclampsia: pathophysiology, challenges, and perspectives. Circ Res. 2019;124:1094–1112. doi:10.1161/CIRCRESAHA.118.313276
6. Staff AC, Fjeldstad HE, Fosheim IK, et al. Failure of physiological transformation and spiral artery atherosis: their roles in preeclampsia. Am J Obstet Gynecol. 2022;226:S895–S906. doi:10.1016/j.ajog.2020.09.026
7. Ying W. Phenomic studies on diseases: potential and challenges. Phenomics. 2023;3:285–299. doi:10.1007/s43657-022-00089-4
8. Sircar M, Thadhani R, Karumanchi SA. Pathogenesis of preeclampsia. Curr Opin Nephrol Hypertens. 2015;24:131–138. doi:10.1097/MNH.0000000000000105
9. El-Sayed AAF. Preeclampsia: a review of the pathogenesis and possible management strategies based on its pathophysiological derangements. Taiwan J Obstet Gynecol. 2017;56:593–598.
10. Mustafa R, Ahmed S, Gupta A, Venuto RC. A comprehensive review of hypertension in pregnancy. J Pregnancy. 2012;2012:105918. doi:10.1155/2012/105918
11. Malik A, Jee B, Gupta SK. Preeclampsia: disease biology and burden, its management strategies with reference to India. Pregnancy Hypertens. 2019;15:23–31. doi:10.1016/j.preghy.2018.10.011
12. Ives CW, Sinkey R, Rajapreyar I, Tita ATN, Oparil S. Preeclampsia-pathophysiology and clinical presentations: JACC state-of-the-art review. J Am Coll Cardiol. 2020;76:1690–1702. doi:10.1016/j.jacc.2020.08.014
13. Mincheva-Nilsson L, Baranov V. Placenta-derived exosomes and syncytiotrophoblast microparticles and their role in human reproduction: immune modulation for pregnancy success. Am J Reprod Immunol. 2014;72:440–457. doi:10.1111/aji.12311
14. Tannetta DS, Dragovic RA, Gardiner C, Redman CW, Sargent IL. Characterisation of syncytiotrophoblast vesicles in normal pregnancy and pre-eclampsia: expression of Flt-1 and endoglin. PLoS One. 2013;8:e56754. doi:10.1371/journal.pone.0056754
15. Schanz A, Winn VD, Fisher SJ, et al. Pre-eclampsia is associated with elevated CXCL12 levels in placental syncytiotrophoblasts and maternal blood. Eur J Obstet Gynecol Reprod Biol. 2011;157:32–37. doi:10.1016/j.ejogrb.2011.02.023
16. Makikallio K, Kaukola T, Tuimala J, Kingsmore SF, Hallman M, Ojaniemi M. Umbilical artery chemokine CCL16 is associated with preterm preeclampsia and fetal growth restriction. Cytokine. 2012;60:377–384. doi:10.1016/j.cyto.2012.07.008
17. Chen X, Li P, Liu M, et al. Gut dysbiosis induces the development of pre-eclampsia through bacterial translocation. Gut. 2020;69:513–522. doi:10.1136/gutjnl-2019-319101
18. Wang H, Zhuang Y, Hua R, et al. Multi-omics exploration of obesity biomarkers in sedentary and weight loss cohorts. Phenomics. 2025;5:137–153. doi:10.1007/s43657-024-00165-x
19. Li P, Wang H, Guo L, et al. Association between gut microbiota and preeclampsia-eclampsia: a two-sample Mendelian randomization study. BMC Med. 2022;20:443. doi:10.1186/s12916-022-02657-x
20. Wang R, Liu C, Liu X, Liu L, Xiao Y, Huo Y. Association between cellular immune and preeclampsia and preterm birth: a Mendelian randomization study. J Reprod Immunol. 2024;166:104391. doi:10.1016/j.jri.2024.104391
21. Li C, Tian Y, Dougarem D, Sun L, Zhong Z. Systemic inflammatory regulators and preeclampsia: a two-sample bidirectional Mendelian randomization study. Front Genet. 2024;15:1359579. doi:10.3389/fgene.2024.1359579
22. Wei J, Huang L, Wu M, et al. The relationship between human blood metabolites and preeclampsia-eclampsia: a Mendelian randomization study. Medicine. 2024;103:e37505. doi:10.1097/MD.0000000000037505
23. Ardissino M, Truong B, Slob EAW, et al. Proteome- and transcriptome-wide genetic analysis identifies biological pathways and candidate drug targets for preeclampsia. Circ Genom Precis Med. 2024;17:e004755. doi:10.1161/CIRCGEN.124.004755
24. Smith GD, Ebrahim S. ‘Mendelian randomization’: can genetic epidemiology contribute to understanding environmental determinants of disease? Int J Epidemiol. 2003;32:1–22. doi:10.1093/ije/dyg070
25. Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013;37:658–665. doi:10.1002/gepi.21758
26. Davey Smith G, Hemani G. Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum Mol Genet. 2014;23:R89–98. doi:10.1093/hmg/ddu328
27. Orru V, Steri M, Sidore C, et al. Complex genetic signatures in immune cells underlie autoimmunity and inform therapy. Nat Genet. 2020;52:1036–1045. doi:10.1038/s41588-020-0684-4
28. Zhao J, Han X, Li H, et al. Analysis of the immune response by standardized whole-blood stimulation with metabolism modulation. Phenomics. 2024;4:81–89. doi:10.1007/s43657-023-00114-0
29. Zhao JH, Stacey D, Eriksson N, et al. Genetics of circulating inflammatory proteins identifies drivers of immune-mediated disease risk and therapeutic targets. Nat Immunol. 2023;24:1540–1551. doi:10.1038/s41590-023-01588-w
30. Lopera-Maya EA, Kurilshikov A, van der Graaf A, et al. Effect of host genetics on the gut microbiome in 7738 participants of the Dutch Microbiome Project. Nat Genet. 2022;54:143–151. doi:10.1038/s41588-021-00992-y
31. Shin SY, Fauman EB, Petersen AK, et al. An atlas of genetic influences on human blood metabolites. Nat Genet. 2014;46:543–550. doi:10.1038/ng.2982
32. Chen Y, Lu T, Pettersson-Kymmer U, et al. Genomic atlas of the plasma metabolome prioritizes metabolites implicated in human diseases. Nat Genet. 2023;55:44–53. doi:10.1038/s41588-022-01270-1
33. Sun BB, Maranville JC, Peters JE, et al. Genomic atlas of the human plasma proteome. Nature. 2018;558:73–79. doi:10.1038/s41586-018-0175-2
34. Wootton RE, Lawn RB, Millard LAC, et al. Evaluation of the causal effects between subjective wellbeing and cardiometabolic health: mendelian randomisation study. BMJ. 2018;362:k3788. doi:10.1136/bmj.k3788
35. Kwok MK, Kawachi I, Rehkopf D, Schooling CM. The role of cortisol in ischemic heart disease, ischemic stroke, type 2 diabetes, and cardiovascular disease risk factors: a bi-directional Mendelian randomization study. BMC Med. 2020;18:363. doi:10.1186/s12916-020-01831-3
36. Yang F, Wu Y, Hockey R, et al;. Evidence of shared genetic factors in the etiology of gastrointestinal disorders and endometriosis and clinical implications for disease management. Cell Rep Med. 2023;4:101250. doi:10.1016/j.xcrm.2023.101250
37. Hartley AE, Power GM, Sanderson E, Smith GD. A guide for understanding and designing mendelian randomization studies in the musculoskeletal field. JBMR Plus. 2022;6:e10675. doi:10.1002/jbm4.10675
38. Hasan MM, Thompson-Snipes L, Klintmalm G, et al. CD24(hi)CD38(hi) and CD24(hi)CD27(+) human regulatory B cells display common and distinct functional characteristics. J Immunol. 2019;203:2110–2120. doi:10.4049/jimmunol.1900488
39. Chen X, Du Y, Lin X, Qian Y, Zhou T, Huang Z. CD4+CD25+ regulatory T cells in tumor immunity. Int Immunopharmacol. 2016;34:244–249. doi:10.1016/j.intimp.2016.03.009
40. Bajnok A, Ivanova M, Rigo J Jr, Toldi G. The distribution of activation markers and selectins on peripheral T lymphocytes in preeclampsia. Mediators Inflamm. 2017;2017:8045161. doi:10.1155/2017/8045161
41. Stohl HE, Stohl W. Maternal and cord blood BAFF and APRIL levels during pregnancy. Am J Reprod Immunol. 2023; 89:e13654.
42. Filaci G, Fenoglio D, Fravega M, et al. CD8+ CD28- T regulatory lymphocytes inhibiting T cell proliferative and cytotoxic functions infiltrate human cancers. J Immunol. 2007;179:4323–4334. doi:10.4049/jimmunol.179.7.4323
43. Horzum U, Yoyen-Ermis D, Taskiran EZ, et al. CD66b(+) monocytes represent a proinflammatory myeloid subpopulation in cancer. Cancer Immunol Immunother. 2021;70:75–87. doi:10.1007/s00262-020-02656-y
44. Nagayama S, Shirasuna K, Nagayama M, et al. Decreased circulating levels of plasmacytoid dendritic cells in women with early-onset preeclampsia. J Reprod Immunol. 2020;141:103170. doi:10.1016/j.jri.2020.103170
45. Nishikawa H, Koyama S. Mechanisms of regulatory T cell infiltration in tumors: implications for innovative immune precision therapies. J Immunother Cancer. 2021;9: e002591.
46. Jin J, Gao L, Zou X, et al. Gut dysbiosis promotes preeclampsia by regulating macrophages and trophoblasts. Circ Res. 2022;131:492–506. doi:10.1161/CIRCRESAHA.122.320771
47. Zhao J, Pan X, Hao D, et al. Causal associations of gut microbiota and metabolites on sepsis: a two-sample Mendelian randomization study. Front Immunol. 2023;14:1190230. doi:10.3389/fimmu.2023.1190230
48. Liu J, Yang H, Yin Z, et al. Remodeling of the gut microbiota and structural shifts in Preeclampsia patients in South China. Eur J Clin Microbiol Infect Dis. 2017;36:713–719. doi:10.1007/s10096-016-2853-z
49. Lv LJ, Li SH, Li SC, et al. Early-onset preeclampsia is associated with gut microbial alterations in antepartum and postpartum women. Front Cell Infect Microbiol. 2019;9:224. doi:10.3389/fcimb.2019.00224
50. Kivenson V, Giovannoni SJ. An expanded genetic code enables trimethylamine metabolism in human gut bacteria. mSystems. 2020;5: 10–128.
51. Li M, Wu Y, Ye L. The role of amino acids in endothelial biology and function. Cells. 2022;11. doi:10.3390/cells12010011
52. Fiaccadori E, Cosola C, Sabatino A. Targeting the gut for early diagnosis, prevention, and cure of diabetic kidney disease: is the phenyl sulfate story another step forward? Am J Kidney Dis. 2020;75:144–147. doi:10.1053/j.ajkd.2019.07.001
53. Braekke K, Ueland PM, Harsem NK, Karlsen A, Blomhoff R, Staff AC. Homocysteine, cysteine, and related metabolites in maternal and fetal plasma in preeclampsia. Pediatr Res. 2007;62:319–324. doi:10.1203/PDR.0b013e318123fba2
54. Ostojic SM. Advanced physiological roles of guanidinoacetic acid. Eur J Nutr. 2015;54:1211–1215. doi:10.1007/s00394-015-1050-7
55. Maddirevula S, Coskun S, Al-Qahtani M, et al. ASTL is mutated in female infertility. Hum Genet. 2022;141:49–54. doi:10.1007/s00439-021-02388-8
56. Thorne A, Necker-Brown A, Bansal A, et al. Differential regulation of BIRC2 and BIRC3 expression by inflammatory cytokines and glucocorticoids in pulmonary epithelial cells. PLoS One. 2023;18:e0286783. doi:10.1371/journal.pone.0286783
57. Sugita G, Tsuyoshi H, Nishijima K, Yoshida Y. Fructose-1,6-bisphosphatase deficiency: a case of a successful pregnancy by closely monitoring metabolic control. JIMD Rep. 2014;14:115–118. doi:10.1007/8904_2013_290
58. Turnham RE, Scott JD. Protein kinase A catalytic subunit isoform PRKACA; history, function and physiology. Gene. 2016;577:101–108. doi:10.1016/j.gene.2015.11.052
59. Liu W, Lin S, Li L, Tai Z, Liu JX. Zebrafish ELL-associated factors Eaf1/2 modulate erythropoiesis via regulating gata1a expression and WNT signaling to facilitate hypoxia tolerance. Cell Regen. 2023;12:10. doi:10.1186/s13619-022-00154-3
60. Habuchi H, Nagai N, Sugaya N, Atsumi F, Stevens RL, Kimata K. Mice deficient in heparan sulfate 6-O-sulfotransferase-1 exhibit defective heparan sulfate biosynthesis, abnormal placentation, and late embryonic lethality. J Biol Chem. 2007;282:15578–15588. doi:10.1074/jbc.M607434200
61. Nilsson SK, Heeren J, Olivecrona G, Merkel M. Apolipoprotein A-V; a potent triglyceride reducer. Atherosclerosis. 2011;219:15–21. doi:10.1016/j.atherosclerosis.2011.07.019
62. Eren EC, Kaya S, Argun D. The assessment of maternal and umbilical cord neudesin levels in pregnancies with gestational diabetes mellitus. J Obstet Gynaecol. 2022;42:2941–2945. doi:10.1080/01443615.2022.2114328
63. Deischinger C, Bastian M, Leitner K, et al. Gremlin-1 in pregnancy and postpartum: relation to the fatty liver index, markers of bone health, glucose metabolism and gestational diabetes mellitus status. Acta Diabetol. 2023;60:1699–1707. doi:10.1007/s00592-023-02151-7
64. Andreuzzi E, Colladel R, Pellicani R, et al. The angiostatic molecule Multimerin 2 is processed by MMP-9 to allow sprouting angiogenesis. Matrix Biol. 2017;64:40–53. doi:10.1016/j.matbio.2017.04.002
65. Zhang Z, Li LY. TNFSF15 modulates neovascularization and inflammation. Cancer Microenviron. 2012;5:237–247. doi:10.1007/s12307-012-0117-8
66. Humphries F, Bergin R, Jackson R, et al. The E3 ubiquitin ligase Pellino2 mediates priming of the NLRP3 inflammasome. Nat Commun. 2018;9:1560. doi:10.1038/s41467-018-03669-z
67. Leach RE, Romero R, Kim YM, et al. Pre-eclampsia and expression of heparin-binding EGF-like growth factor. Lancet. 2002;360:1215–1219. doi:10.1016/S0140-6736(02)11283-9
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.