Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 13
Genetic Polymorphisms in Transforming Growth Factor-β, Interferon-γ and Interleukin-6 Genes and Susceptibility to Behcet’s Disease in Saudi Population
Authors Al Okaily F, Alrashidi S, Mustafa M ![]() , Alrashdan FB
, Alrashdan FB
Received 13 February 2020
Accepted for publication 28 May 2020
Published 31 July 2020 Volume 2020:13 Pages 253—259
DOI https://doi.org/10.2147/PGPM.S249361
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Fahdah Al Okaily,1 Seham Alrashidi,1 Mohammad Mustafa,2 Faris Bander Alrashdan3
1Department of Rheumatology, Prince Sultan Military Medical City, Riyadh, Kingdom of Saudi Arabia; 2Scientific Research Center, Prince Sultan Military Medical City, Riyadh, Kingdom of Saudi Arabia; 3Department of Internal Medicine, Prince Sultan Military Medical City, Riyadh, Kingdom of Saudi Arabia
Correspondence: Fahdah Al Okaily
Department of Rheumatology, Prince Sultan Military Medical City, Riyadh, Kingdom of Saudi Arabia
Email [email protected]
Background: Behcet’s disease (BD) is a complex, inflammatory, immune-mediated multi-systemic disease of unknown etiology. Cytokines play major roles in the pathophysiology of BD, and its production may be affected by polymorphism in cytokine genes. Hence, the present study was planned to investigate any possible association between the polymorphism in TGF-β, IFN-γ, and IL-6 genes and BD in the Saudi population.
Materials and Methods: The present study includes 79 BD cases and 117 age-matched controls. Polymerase chain reaction with restriction fragment length polymorphism (PCR-RFLP) and amplification refractory mutation systems (ARMS) PCR methods were used for polymorphic analysis.
Results: The heterozygous (CT) and (CT+TT) combined genotypes of TGF-β (509-C/T), heterozygous (AT), variant (TT), and (AT+TT) combined genotypes of IFN-γ (874-A/T) were significantly (P< 0.05) associated with BD in the Saudi population. No significant differences were observed for IL-6 (174-G/C) genotypes and alleles between BD cases and controls. Gender does not reflect any significant genotypic and allelic association with males and females.
Conclusion: CT genotype of TGF-β, and AT and TT genotypes of IFN-γ could be associative genetic risk factors for BD in Saudis. Regulatory region polymorphism in cytokines gene can increase inflammation and deregulated immune response, which could be risk factor for BD.
Keywords: tumor growth factor-β, Interferon-γ, Interleukin-6, Behcet’s disease, Saudis, genetic polymorphism
Introduction
Behcet’s disease (BD) is a multi-systemic, chronic inflammatory disorder with the strong genetic background, characterized by recurrent oral aphthous, genital ulcer, uveitis, and skin lessons.1,2 Prevalence of this disease is higher in ancient silk route countries extending from eastern Asia to Mediterranian and middle east countries.3,7 The highest incidence rate of BD is reported in Turkey (420/100,000), while the lowest rates were in the UK, Spain, Sweden, Portugal, and the US, ranging from 0.3–6.4 per 100,000 persons.5,8,9 In the Saudi population, the incidence rate of BD is 20/100,000.10 It affects, without sex predilection, mostly individuals in the age range 20–40 years.8,11 Despite substantial basic scientific and clinical investigations, BD is still a clinical and public health problem. Several studies have suggested the role of genetic, environmental factors, and deregulated immune for the development of BD.1,12,13,14
The identification of genetic susceptibility variants in BD will lead to better understanding of the pathophysiology of disease and also to screen the high risk patients at the early stage of disease, development of new diagnostic markers, and treatment strategies. Previous studies, including those from our laboratory, have reported human leukocyte antigen (HLA)-B*51 as the strongest genetic risk factor for BD.12,13,15 Previous studies reported the significant association of tumor growth factor (TGF-β) variants (CC) at codon 10 and GG at codon 25 with BD in Iranian and Turkish populations.16,17 The TGF-β1 is an effective immunosuppressive cytokine. Its functional promoter SNP (509C/T, rs1800469) has been shown to cause a binding site for the transcription factor Yin-Yang1 (YY-1), thereby increasing TGF-β1 gene transcription.18 Interferon gamma (IFN-γ) is an important activator of macrophage and has shown its involvement in host defense and immune regulation.19 TGF-β and IFN-γ often function in opposition, which can modulate the inflammatory response.20 The high level of IFN-γ in Behçet uveitis and also genetic susceptibility of IFN-γ +874A allele and A/A genotype to BD in the Turkish population suggest a possible genetic association of IFN-γ (874A/T, rs2430561) with the Saudi population.21,22
Interleukin (IL)-6 is another important mediator of inflammatory immune responses, and it’s polymorphisms are well known for their role in chronic inflammatory state and autoimmune disorders.23–25 The significant association of IL-6 (174-C/G) genotype with BD has been reported in an Iranian population.26 However, plasma protein analyses of IL-6 in BD have shown inconsistent results in different populations.27,28 The polymorphism in the regulatory region of cytokines gene can affect the gene transcription, resulting in a variation of many of the effector and regulatory functions of immune and inflammatory responses.1,28 So far, no studies have reported a genetic association of these genes with BD in a Saudi population. Hence, the present study aimed to investigate the genetic polymorphism in TGF-β, IFN-γ, and IL-6 (rs1800795), and susceptibility to BD in a Saudi population.
Materials and Methods
In the present study, a total of 79 BD patients and 117 age, sex, and ethnicity (Saudis) matched controls were recruited from the Department of Rheumatology, Prince Sultan Military Medical City (PSMMC), Riyadh, Kingdom of Saudi Arabia. The ethical clearance was obtained from the institutional ethical clearance committee, PSMMC, Riyadh to conduct this research work. A properly filled and duly signed consent form was obtained from each of the participants prior to sample collection. The patient and control samples were collected after following the strict exclusion and inclusion criteria. The past medical records for all the subjects were collected to know drug in use, demography, and life style. The patients having a history of inflammatory/autoimmune diseases were excluded from this study. None of the subjects had any malignancies or other chronic illnesses in the past. The diagnosis has been done as per the International Study Group criteria (ISG 1990) for BD.29 The principles of the Declaration of Helsinki were followed to complete this research work. Our skilled physician examined the patients for clinical features such as oral ulcers, genital ulcer, ocular inflammation, musculoskeletal, cutaneous, gastrointestinal lesions, nervous, pulmonary, cardiovascular manifestations, and vascular lesions. Venous blood (3 mL) was drawn in an EDTA coated vial from each subjects, brought to the laboratory in an ice box and stored at −80°C before extraction of genomic DNA.
Genotyping Analysis
Genomic DNA for genotyping was isolated from whole blood by using a commercially available QIAampDNA mini kit (Qiagen, Venlo, Limburg, the Netherlands) as per the manufacturer protocol. The quality of DNA was checked on agarose gel and quantitation was done by using Nano Drop-2000 at 260/280 nm (ThermoFisherScientific Inc, Waltham, MA, USA). The gradient PCR (Eppendorf, Hamburg, Germany) was used to amplify the genomic DNA for polymorphic analysis. Amplification refractory mutation systems (ARMS)-PCR methodology was used to detect polymorphisms at position 874 of IFN-γ. In this PCR, we used a generic antisense primer and one of the two allele specific sense primers. Human growth hormone (hGH) primer was used as an internal (426 bp) control to check the success of PCR amplification in both the reactions. Restriction fragment length polymorphism (PCR-RFLP) method was used for the polymorphic analysis of TGF-β (509 C/T) and IL-6 (174-G/C). PCR amplification was carried out using 5× FIREPol Master Mix (Solis Biodyne, Tartu, Estonia) with specific optimized reaction conditions. The primer sequences and annealing temperatures used for TGF-β, IFN-γ, and IL-6 genes amplification are mentioned in Table 1.
 |
Table 1 Oligonucleotide Primers Used for Amplification of TGF-β, IFN-γ, IL-6, hGH Genes, and Annealing Temperatures |
DNA amplification was done in a 20 µL reaction mixture consisting of 50 ng of genomic DNA, 10 µM of each primer, 200 µM of dNTPs mixture, 1.5 mM MgCl2, 2.5 µL of 10x PCR buffer (10x500 mM KCl, 100 mM Tris-HCl, pH 9.0) and 1U Taq DNA polymerase. The thermal profile for IFN-γ gene consisted of initial denaturation for 1 minute at 95°C, 40 cycles of denaturation for 30 seconds at 95°C, annealing for 30 seconds at 55°C and extension for 30 seconds at 72°C. In the present study, an amplification program was similar for TGF-β and IL-6 genes except that annealing temperatures were kept at 50°C and 59°C, respectively. Amplification of genomic DNA using a specific protocol for IL-6 and TGF-β1 genes yielded 198 bp and 441 bp PCR products of DNA, respectively. The PCR products were digested with SfaNI and Bsu36 I restriction enzyme (New England BioLabs, Beverly, MA) at 37°C for 12 hours. The digested products for various samples were separated on the 2.5% agarose gel, stained with ethidium bromide and gel pictures were taken using a gel documentation system. Digestion of TGF-β1 PCR product gave two fragments of 251 and 190 bp for common genotype (CC), three fragments of 441, 251, and 190 bp for heterozygous (CT) and uncut DNA of 441 bp for variants (TT) genotype. As a result of digestion, of IL-6 PCR product two fragments of 140 bp and 58 bp indicated a common (GG) genotype, while three fragment of 198, 140, and 58 bp indicated a heterozygous (GC) genotype, and an uncut single band indicated variant (CC) type for IL-6 gene.
Statistical Analysis
The statistical analysis was carried out by using SSPS software, windows version 21.0 (Chicago, IL, USA). The χ2-test was used to compare genotypic and allelic frequencies between case and controls and values were presented as crude odds ratio (OR) and 95% confidence intervals. P<0.05 was considered as significant.
Results
In our study, 54/79 (68.35%) were male and 25/79 (31.64%) were female, the age of the BD cases was ranged between 20–64 years and for controls 20–60 years. Clinically, all BD patients (100%) had oral ulcers, 80.32% genital ulcers, 70.49% ocular, 67.21% musculoskeletal, 60.65% cutaneous, 36.06% gastrointestinal, and 22.95% of patients had nervous system involvement. The distribution of the TGF-β-(509 C/T), IFN-γ-(874 A/T), and IL-6-(174 C/T) genotypes and allele frequencies in BD cases and controls are shown in Tables 2, 3, and 4. The representative gel pictures for different genes are shown in Figures 1, 2, and 3.
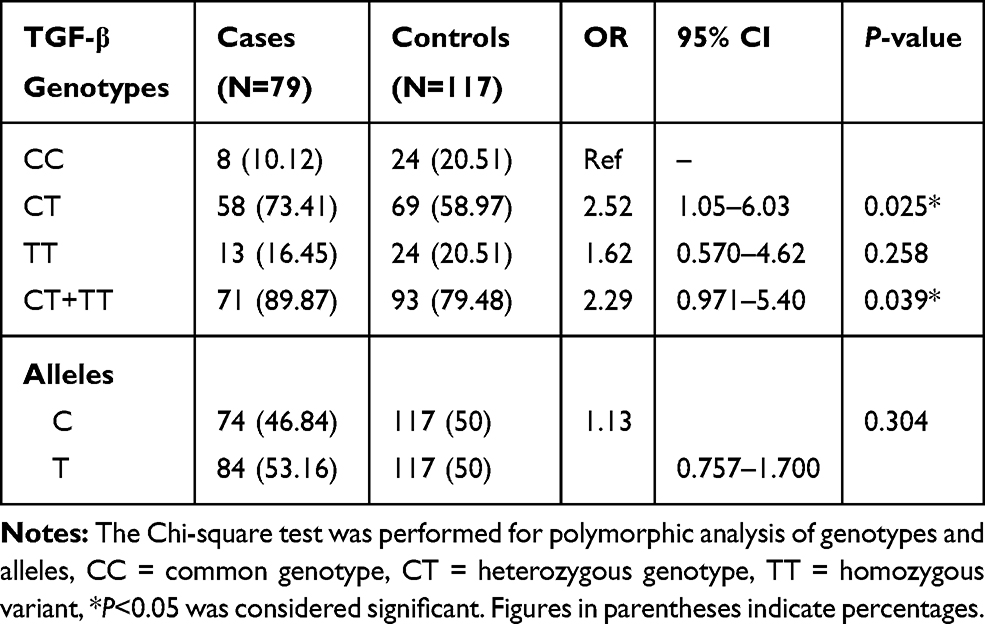 |
Table 2 Genotypic and Allelic Distribution of TGF-β (509-C/T) Gene in BD Cases and Controls |
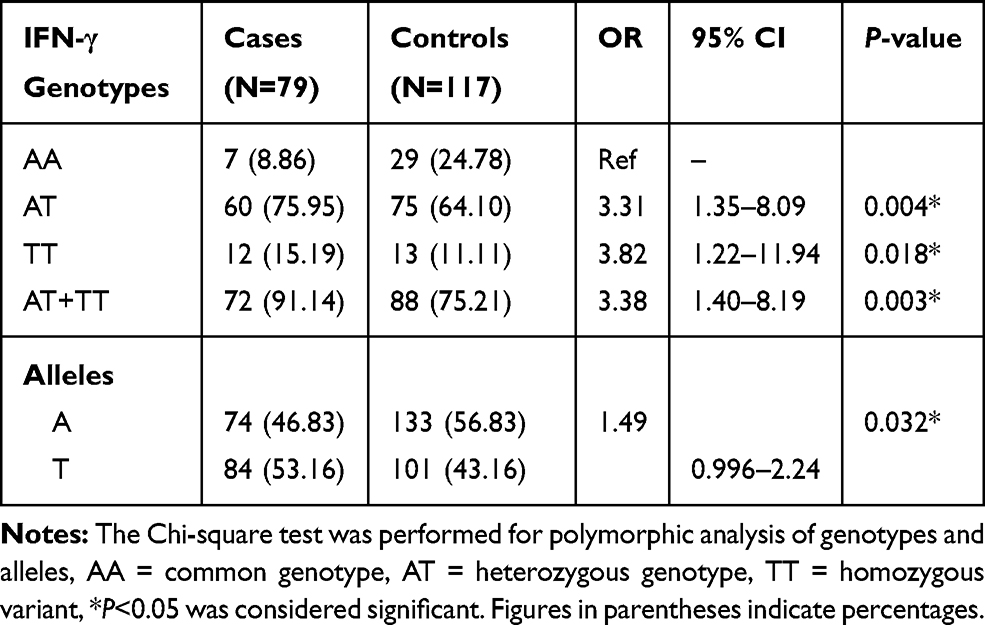 |
Table 3 Genotypic and Allelic Distribution of IFN-γ (874 -A/T) Gene in BD Cases and Controls |
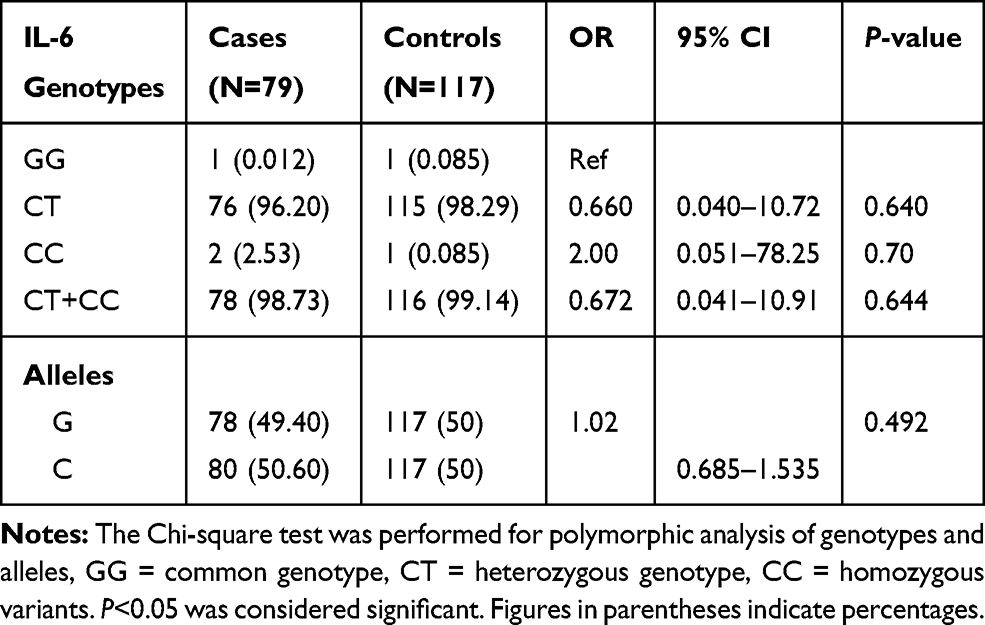 |
Table 4 Genotypic and Allelic Distribution of IL-6 (174-G/C) Gene in BD Cases and Controls |
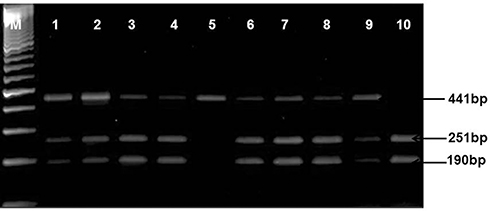 |
Figure 1 Amplified and digested PCR product of TGF-β1 (509-C/T) gene. Lane M: 100 bp DNA marker, Lanes 1, 2, 3, 4, 6, 7, 8, 9: heterozygous genotypes (CT: bands of 441, 251, and 190 bp), Lane 5: homozygous mutant (TT: uncut DNA of 441 bp), and lane 10: wild type genotype (CC: bands of 251 and 190 bp). |
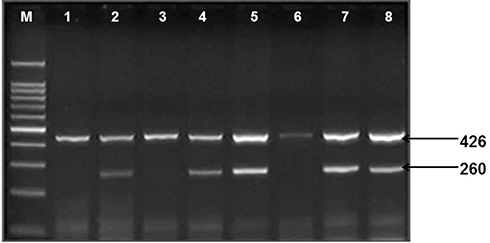 |
Figure 2 Shows IFN-γ (874-A/T) gene amplification. Lane M: 100 bp DNA marker, Lanes 2.4 and 8: amplification of allele T, Lanes 5 and 7: amplification of allele A. Lanes 1.2 and 3.4: Homozygous mutant, Lanes 5 and 6: wild type genotypes, Lanes 7 and 8: heterozygous genotype, 426 bp band for human growth factor gene (internal control). |
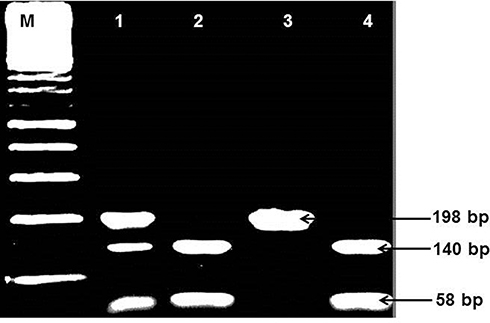 |
Figure 3 Amplified and digested PCR product of IL-6 (174-G/C). Lane M: 100 bp DNA marker: Lane 1: GC (bands of 198, 140, and 58 bp), Lanes 2 and 4: GG genotype (bands of 140 and 58 bp), Lane 3: CC (uncut DNA of 198 bp). |
Our study demonstrates the frequency of CT genotype of TGF-β gene was significantly higher (OR=2.52, 95% CI=1.05–6.03, P=0.025), while TT genotype frequency was lower in BD patients as compared to controls. The allelic data analysis for TGF-β revealed no significance difference (OR=1.13, 95% CI=0.757–1.700, P=0.304) between BD cases and controls. When risk factors containing genotypes were combined (CT+TT) and data were analyzed between BD cases and controls, we found combined genotype increases significant risk (OR=2.29, 95% CI=0.971–5.40, P=0.039) of BD as compared to individual genotypes (Table 2). The analysis of IFN-γ (874-A/T) SNP showing significantly higher frequency of AT and TT genotypes (OR=3.31, 95% CI=1.35–8.09, P=0.004) and (OR=3.82, 95% CI=1.22–11.94, P=0.018) in BD patients as compared to controls. Additionally, the combined (AT+TT) genotypes analysis of IFN-γ between BD cases and controls reflects a significantly higher risk of BD (OR=3.38, 95% CI=1.40–8.19, P=0.003) as compared to controls. The frequency of allele T of IFN-γ gene has shown significant higher risk (OR=1.49, 95% CI=0.996–2.24, P=0.032) of BD cases in comparison to allele A (Table 3). When we analyzed the genotypic and allelic data of IL-6 no significant difference was observed for any of the genotypes and alleles between BD cases and controls (Table 4). The genotypic data stratification for IFN-γ, TGF-β, and IL-6 between male and females has shown no significant difference. The IFN-γ and TGF-β genotypes are shown, with a higher frequency in males as compared to females (Tables 5 and 6).
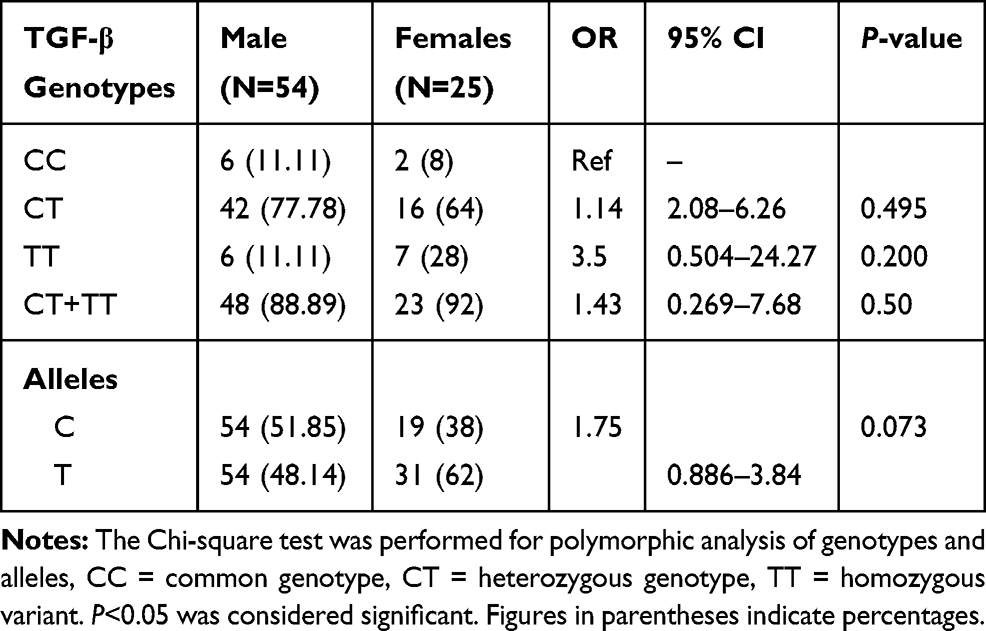 |
Table 5 Genotypic and Allelic Distribution of TGF-β (509-C/T) Gene in Male and Female Cases |
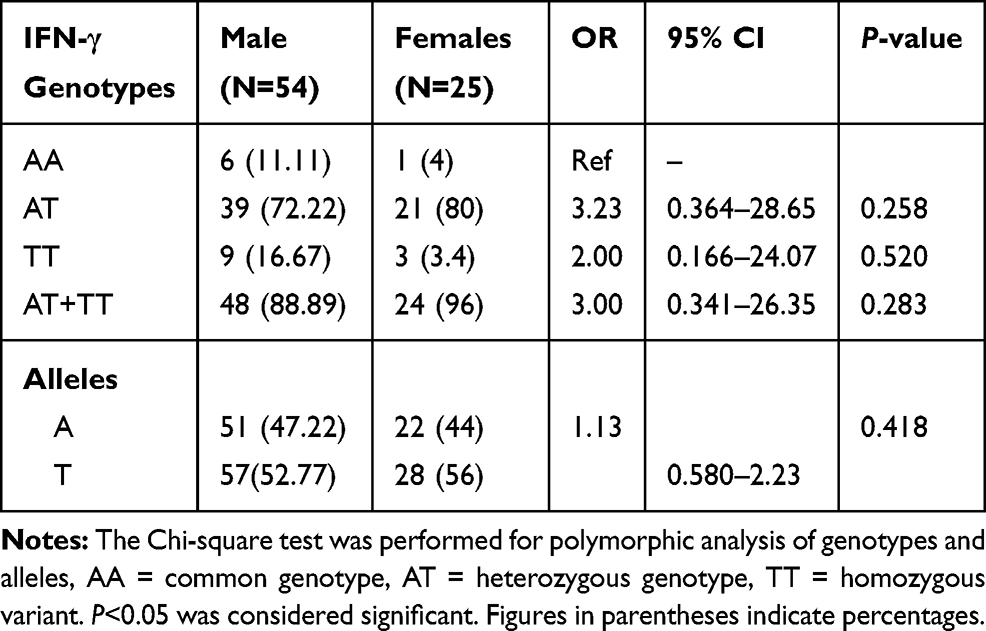 |
Table 6 Genotypic and Allelic Distribution of IFN-γ (874-A/T) Gene in Male and Female Cases |
Discussion
BD is a multifactorial complex human disease with unknown etiology. The role of genetic predisposition, infectious agents, and immune dis-regulation seems to be critical in its pathogenesis. The inter-individual variations in cytokines production can be attributed by mutations within cytokine coding regions and nucleotide variations within more pronounced regulatory regions which could be an important determinants of genetic susceptibility to BD.22,30 To strengthen the genetic association of cytokines with BD among Saudis, it is important to genotype TGF-β, IFN-γ, and IL-6 genes in BD cases and controls.
Our study demonstrates a significant (P=0.025) association of CT genotype of TGF-β in BD cases as compared to controls (Table 2). However, the frequency of CC genotype is lower in BD cases compared to controls, suggesting a common genotype of TGF-β may play a protective role in the development of BD among Saudis (Table 2). Similar to our findings, higher frequency of CT and lower frequency of CC genotypes of TGF-β in BD were reported earlier in Iranian and Turkish populations.16,17 The combined genotypes (CT+TT) of TGF-β confers significantly more risk of BD as compared to individual genotypes. Our data reflects the risk factors aggregation heightened the risk of BD in a Saudi population.
TGF-β and IFN-γ often act opposite to modulate mucosal immunity and inflammation.31 TGF-β is mainly produced by activated macrophages which regulates the production of IFN-γ by natural killer cell in response to tissue injury.17 The underlying mechanism includes transcriptional inhibition of expression factors T-bet and STAT4 by TGF-β which regulate IFN-γ expression. It can also regulate the expression of IFN-γ by binding of TGF-β molecule to the TGF-β receptor which induces phosphorylation type-I receptor by type-II receptor kinase. Then activated type-1 receptor phosphorylates SMAD3 leading to heterodimer complex formation with SMAD4. This can bind to the IFN-γ promoter and repress its transcription.32–34 In contrast to this, inhibition of the TGF-β pathway in the lymphocytes may induce chronic inflammation.35 Thus, the balance in level of TGF-β and IFN-γ is important in signaling to determine the immunological status and could be targeted for the therapeutic intervention in BD patients.
In the present study, AT and TT genotypes and T allele of IFN-γ gene may be associative risk factors for BD in a Saudi population (Table 3). The frequency of AA genotype of IFN-γ is significantly lower in BD cases as compared to controls reflecting its possible protective role in the BD development. When we combined genotypic risk factors (AT+TT) of IFN-γ, data analysis revealed individuals with combined genotypes may be more susceptible to the disease. Alayli et al22 reported strong susceptibility of A allele and AA genotype of IFN-γ, while AT was not significantly different in BD cases as compared to controls in their study. IFN-γ (874 A/T) polymorphism affects its serum protein levels since this polymorphism is located within a putative nuclear factor-kB (NF-kB) binding site. It has been suggested that the high production of IFN-γ may be because of its T allele.31
We have not observed any evidence for genetic association of IL-6 (174- G/C) genotypes with BD (Table 4). Our findings for IL-6 genotypes are corroborated with Turkish, Tunisian, and Korean populations.36–38 In contrast to this, a significant association of CG genotype of IL-6 (174-C/G) with BD was reported in an Iranian population.26 IL-6 is a pleiotropic cytokine and it plays a significant role in inflammation and immune response.39 It has been demonstrated that IL-6 promoter polymorphisms are key regulators of IL-6 levels in vitro and in vivo.24 The elevated plasma protein and increased mRNA expression of IL-6 were reported in BD patients.28,40 Thus, genetic susceptibility of IL-6 (174-C/G) to BD is still illusive as some of the studies have presented as a risk factor while in other studies reported its protective role. Upon stratification of genotypic data of different cytokine genes based on gender, we did not find any significant differences (Tables 5 and 6). However, many studies have suggested gender could influence the genetic susceptibility to BD, while others oppose and certified no significant association.37,41
The limitation of our studies are small sample size and genotyping of only three SNPs in three cytokines gene. Our findings need to be confirmed and validated in a much larger sample size by whole genome/whole exome sequencing that will also enable us to investigate all the other crucial SNPs associated with BD in a Saudi population. The estimation of TGF-β, IL-6, and IFN-γ activity and other inflammatory markers and gene environment interaction will further strengthen the observation obtained in the present study.
Conclusion
Our findings supported the genetic basis of BD. The present study clearly demonstrates the association of TGF-β and IFN-γ polymorphism with BD and it could be used as a genetic marker for screening BD patient among Saudis. The promoter polymorphisms could be an activator of gene expression; therefore, our findings may be useful in exploring the molecular mechanism underlying the onset of BD.
Ethical Approval
Institutional ethical approval has been taken to conduct this Research study.
Acknowledgment
The authors are thankful to Maha Al-onazi for their help with laboratory work.
Disclosure
The authors declare that they have no conflicts of interest for this work.
References
1. Greco A, De Virgilio A, Ralli M, et al. Behçet’s disease: new insights into pathophysiology, clinical features and treatment options. Autoimmun Rev. 2018;17(6):567–575. doi:10.1016/j.autrev.2017.12.006
2. Saleh Z, Arayssi T. Update on the therapy of Behçet disease. Ther Adv Chronic Dis. 2014;5(3):112–134. doi:10.1177/2040622314523062
3. Jang WC, Park SB, Nam YH, et al. Interleukin-18 gene polymorphisms in Korean patients with Behçet’s disease. Clin Exp Rheumatol. 2005;23:S59–63.
4. Wang LY, Zhao DB, Gu J, Dai SM. Clinical characteristics of Behçet’s disease in China. Rheumatol Int. 2010;30(9):1191–1196. doi:10.1007/s00296-009-1127-9
5. Al-Dalaan AN, Al Balaa SR, El Ramahi K, et al. Behçet’s disease in Saudi Arabia. J Rheumatol. 1994;21(4):658–661.
6. Azizlerli G, Köse AA, Sarica R, et al. Prevalence of Behçet’s disease in Istanbul, Turkey. Int J Dermatol. 2003;42(10):803–806. doi:10.1046/j.1365-4362.2003.01893.x
7. Krause I, Yankevich A, Fraser A, et al. Prevalence and clinical aspects of Behcet’s disease in the north of Israel. Clin Rheumatol. 2007;26(4):555–560. doi:10.1007/s10067-006-0349-4
8. Davatchi F, Shahram F, Chams-Davatchi C, et al. Behcet’s disease: from East to West. Clin Rheumatol. 2010;29:823–833.
9. Cho SB, Cho S, Bang D. New insights in the clinical understanding of Behçet’s disease. Yonsei Med J. 2012;53(1):35–42. doi:10.3349/ymj.2012.53.1.35
10. Al Zahrani FM, Alsultan B, Alahmari H, et al. Behçet’s disease presentations and care outcomes in a tertiary hospital in south‐western Saudi Arabia. Int J Rheum Dis. 2019;22(8):1466–1473. doi:10.1111/1756-185X.13598
11. Hamzaoui A, Jaziri F, Ben Salem T, et al. Comparison of clinical features of Behcet disease according to age in a Tunisian cohort. Acta Med Iran. 2014;52:748–751.
12. Al-Okaily F, Al-Rashidi S, Al-Balawi M, Mustafa M, Arfin M, Al-Asmari A. Genetic association of HLA-A*26, -A*31, and -B*51 with Behcet’s disease in Saudi patients. Clin Med Insights Arthritis Musculoskelet Disord. 2016;9:167–173. doi:10.4137/CMAMD.S39879
13. Xavier JM, Davatchi F, Abade O, et al. Characterization of the major histocompatibility complex locus association with Behçet’s disease in Iran. Arthritis Res Ther. 2015;17:81. doi:10.1186/s13075-015-0585-6
14. Hou S, Qi J, Zhang Q, et al. Genetic variants in the JAK1 gene confer higher risk of Behcet’s disease with ocular involvement in Han Chinese. Hum Genet. 2013;132(9):1049–1058. doi:10.1007/s00439-013-1312-5
15. Tong B, Liu X, Xiao J, Su G. Immunopathogenesis of Behcet’s disease. Front Immunol. 2019;10:665. doi:10.3389/fimmu.2019.00665
16. Shahram F, Nikoopour E, Rezaei N, et al. Association of interleukin-2, interleukin-4 and transforming growth factor-beta gene polymorphisms with Behcet’s disease. Clin Exp Rheumatol. 2011;29:S28–31.
17. Dilek K, Ozçimen AA, Saricaoğlu H, et al. Cytokine gene polymorphisms in Behçet’s disease and their association with clinical and laboratory findings. Clin Exp Rheumatol. 2009;27:S73–8.
18. Deng Z, Cao P, Wan M, Sui G. A multifaceted protein beyond a transcription factor. Transcription. 2010;1(2):81–84. doi:10.4161/trns.1.2.12375
19. Calvo J, Martínez N, Etxagibel A, et al. Allelic frequencies of polymorphic variants of cytokine genes (IL1A, IL1B, IL1RN, IL6, IL10, IL12p40, and IFNG) in a Spanish population. Inmunologia. 2002;21(2):76–86.
20. Sanjabi S, Oh SA, Li MO. Regulation of the immune response by TGF-β: from conception to autoimmunity and infection. Cold Spring Harb Perspect Biol. 2017;9(6):a022236. doi:10.1101/cshperspect.a022236
21. Ahn JK, Yu HG, Chung H, et al. Intraocular cytokine environment in active Behçet uveitis. Am J Ophthalmol. 2006;142:429–434. doi:10.1016/j.ajo.2006.04.016
22. Alayli G, Aydin F, Coban AY, et al. T helper 1 type cytokines polymorphisms: association with susceptibility to Behçet’s disease. Clin Rheumatol. 2007;26(8):299–305. doi:10.1007/s10067-006-0503-z
23. Linker-Israeli M, Deans RJ, Wallace DJ, Prehn J, Ozeri-Chen T, Klinenberg JR. Elevated levels of endogenous IL-6 in systemic lupus erythematosus. J Immunol. 1991;147(1):117–123.
24. Fishman D, Faulds G, Jeffery R, et al. The effect of novel polymorphisms in the interleukin-6 (IL-6) gene on IL-6 transcription and plasma IL-6 levels, and an association with systemic-onset juvenile chronic arthritis. J Clin Invest. 1998;102(7):1369–1376. doi:10.1172/JCI2629
25. Pascual M, Nieto A, Matarán L, Balsa A, Pascual-Salcedo D, Martín J. IL-6 promoter polymorphisms in rheumatoid arthritis. Genes Immun. 2000;1(5):338–340. doi:10.1038/sj.gene.6363677
26. Amirzargar A, Shahram F, Nikoopour E, et al. Proinflammatory cytokine gene polymorphisms in Behçet’s disease. Eur Cytokine Netw. 2010;21(4):292–296. doi:10.1684/ecn.2009.0209
27. al-Dalaan A, al-Sedairy S, al-Balaa S, et al. Enhanced interleukin 8 secretion in circulation of patients with Behçet’s disease. J Rheumatol. 1995;22(5):904–907.
28. Talaat RM, Ashour ME, Bassyouni IH, Raouf AA. Polymorphisms of interleukin 6 and interleukin 10 in Egyptian people with Behcet’s disease. Immunol Biol. 2014;219(8):573–582. doi:10.1016/j.imbio.2014.03.004
29. International Study Group for Behcet’s Disease. Criteria for diagnosis of Behcet’s disease. Lancet. 1990;335(8697):1078–1080.
30. Gholijani N, Ataollahi MR, Samiei A, et al. An elevated pro-inflammatory cytokines profile in Behcet’s disease: a multiplex analysis. Immunol Lett. 2017;186:46–51.
31. Pravica V, Perrey C, Stevens A, et al. A single nucleotide polymorphism in the first intron of the human IFN-gamma gene: absolute correlation with a polymorphic CA microsatellite marker of high IFN-gamma production. Hum Immunol. 2000;61(9):863–866. doi:10.1016/S0198-8859(00)00167-1
32. Wahl SM, McCartney-Francis N, Mergenhagen SE. Inflammatory and immunomodulatory roles of TGF-beta. Immunol Today. 1989;10(8):258–261. doi:10.1016/0167-5699(89)90136-9
33. Trotta R, Dal Col J, Yu J, et al. TGF-beta utilizes SMAD3 to inhibit CD16-mediated IFN-gamma production and antibody-dependent cellular cytotoxicity in human NK cells. J Immunol. 2008;181(6):3784–3792. doi:10.4049/jimmunol.181.6.3784
34. Yu J, Wei M, Becknell B, et al. Pro- and anti-inflammatory cytokine signaling: reciprocal antagonism regulates interferon-gamma production by human natural killer cells. Immunity. 2006;24(5):575–590. doi:10.1016/j.immuni.2006.03.016
35. Lin JT, Martin SL, Xia L, Gorham JD. TGF-beta 1 uses distinct mechanisms to inhibit IFN-gamma expression in CD4+ T cells at priming and at recall: differential involvement of Stat4 and T-bet. J Immunol. 2005;174(10):5950–5958. doi:10.4049/jimmunol.174.10.5950
36. Barış S, Akyürek Ö, Dursun A, Akyol M. The impact of the IL-1β, IL-1Ra, IL-2, IL-6 and IL-10 gene polymorphisms on the development of Behcet’s disease and their association with the phenotype. Med Clin (Barc). 2016;146(9):379–383. doi:10.1016/j.medcli.2015.09.017
37. Hamzaoui A, Klii R, Harzallah O, Mahjoub T, Mahjoub S. Polymorphism of interleukin 6-174 G/C in Behcet disease: case series and review of literature. Acta Med Iran. 2014;52(11):811–815.
38. Chang HK, Jang WC, Park SB, et al. Association between interleukin 6 gene polymorphisms and Behcet’s disease in Korean people. Ann Rheum Dis. 2005;64(2):339–340. doi:10.1136/ard.2004.024208
39. Yamakawa Y, Sugita Y, Nagatani T, et al. Interleukin-6 (IL-6) in patients with Behçet’s disease. J Dermatol Sci. 1996;11(3):189–195. doi:10.1016/0923-1811(95)00439-4
40. Shim J, Byun HO, Lee YD, et al. Interleukin-6 small interfering RNA improved the herpes simplex virus induced systemic inflammation in vivo Behcet’s disease like mouse model. Gene Ther. 2009;16(3):415–425. doi:10.1038/gt.2008.180
41. Ohno S, Ohguchi M, Hirose S, et al. Close association of HLA-Bw51 with Behçet’s disease. Arch Ophthalmol. 1982;100(9):1455–1458. doi:10.1001/archopht.1982.01030040433013
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.