Back to Journals » Drug Design, Development and Therapy » Volume 20
From Phytochemistry to Drug Discovery: A Systematic Review of Multitarget Pharmacology and Lead Compound Prioritization of Piper crocatum Ruiz & Pav
Authors Apriyanti E ![]() , Kurnia D
, Kurnia D ![]() , Al-Anshori J
, Al-Anshori J ![]() , Takaya A
, Takaya A
Received 24 March 2026
Accepted for publication 10 June 2026
Published 20 June 2026 Volume 2026:20 611626
DOI https://doi.org/10.2147/DDDT.S611626
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Anastasios Lymperopoulos
Eti Apriyanti,1,2 Dikdik Kurnia,2 Jamaludin Al-Anshori,2 Akiko Takaya3,4
1Doctoral Program of Chemistry, Department of Chemistry, Faculty of Mathematics and Natural Sciences, Universitas Padjadjaran, Sumedang, West Java, 45363, Indonesia; 2Department of Chemistry, Faculty of Mathematics and Natural Sciences, Universitas Padjadjaran, Sumedang, West Java, 45363, Indonesia; 3Department of Infection Control Science, Graduate School of Pharmaceutical Sciences, Chiba University, Chiba, 260-8675, Japan; 4Medical Mycology Research Center, Chiba University, Chiba, 260-8673, Japan
Correspondence: Dikdik Kurnia, Department of Chemistry, Faculty of Mathematics and Natural Sciences, Universitas Padjadjaran, Sumedang, West Java, 45363, Indonesia, Email [email protected]
Abstract: Piper crocatum Ruiz & Pav. (Piperaceae), commonly known as red betel, is a medicinal plant widely used in Southeast Asian traditional medicine and has attracted increasing interest due to its diverse pharmacological properties. However, the relationship between its phytochemical constituents, biological activities, molecular mechanisms, and drug discovery potential remains insufficiently integrated. This Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA)-guided review aimed to summarize the phytochemical constituents and pharmacological activities of P. crocatum and evaluate their relevance as potential lead structures for drug development. Literature published between 2000 and 2026 was retrieved from Scopus and PubMed. A total of 180 records were identified, and 102 eligible original studies reporting phytochemical and/or biological investigations of P. crocatum were included in the final analysis. Forty compounds have been reported from P. crocatum, including phenolic compounds, neolignans, flavonoids, terpenoids, and phytosterols. These constituents are associated with antimicrobial, antibiofilm, antioxidant, anti-inflammatory, antidiabetic, hepatoprotective, anticancer, and immunomodulatory activities. Mechanistic evidence suggests that these effects involve modulation of oxidative stress, inflammatory signaling, microbial sterol biosynthesis, biofilm-related proteins, and metabolic enzymes such as α-glucosidase and pancreatic lipase. Among the reported compounds, pachypodol, crocatin derivatives, β-sitosterol, and stigmasterol show promising pharmacological profiles and may serve as lead structures for further development. Overall, the chemical diversity and multitarget activities of P. crocatum support its potential as a source of drug discovery candidates. Future studies should prioritize bioassay-guided isolation, molecular target validation, pharmacokinetic evaluation, and structure–activity relationship analysis. The flowchart begins with an image of Piper crocatum, leading to ′Phytochemical Classes′ which include Phenolics, Neolignans, Flavonoids, Terpenoids and Phytosterols. This connects to ′Molecular Targets′ with enzymatic targets: CYP51, alpha-glucosidase; signaling pathways: NF-kB, iNOS, COX-2; cellular processes: ROS; and virulence/biofilm target: GbpC. Next, ′Therapeutic Effects′ are listed: Infection Control (Antibacterial, Antifungal, Antibiofilm), Inflammation (decreased TNF-alpha, IL-6, excessive NO production), Metabolic Disorders (Antidiabetic, Anti-obesity) and Hepatoprotection (decreased ROS, ALT/AST). ′Lead Compounds′ prioritized for development are Pc-1 (Immunomodulator), Pachypodol (Antioxidant), beta-Sitosterol (Antimicrobial) and Apigenin (Anti-inflammatory). These lead to ′Drug Discovery Potential′ for multi-target therapeutic candidates. Structures of Pachypodol, Apigenin and beta-Sitosterol are shown. Note: downward arrow indicates decreased levels or activity.Flowchart of Piper crocatum′s phytochemicals, targets and therapeutic effects.
Keywords: Piper crocatum, phytochemistry, pharmacological activity, natural product drug discovery, multitarget therapy, molecular mechanisms
Introduction
Natural products continue to play a vital role in modern drug discovery by providing structurally diverse molecules that may serve as scaffolds for the development of new therapeutic agents. Most currently approved drugs are derived from natural products or inspired by natural product scaffolds, primarily for the treatment of anti-infective, anticancer, and metabolic diseases.1,2 Thus, medicinal plants rich in chemically diverse secondary metabolites represent a valuable source of bioactive compounds with potential pharmaceutical applications.
Piper crocatum Ruiz & Pav., commonly known as red betel, is a medicinal plant of the Piperaceae family that has attracted increasing scientific attention owing to its diverse applications in traditional medicine. P. crocatum is widely used in traditional medicine in Southeast Asia, particularly in Indonesia, where its leaves are used to manage infections, inflammation, metabolic disorders, oral conditions, and gynecological complaints.3–6 These traditional uses have encouraged pharmacological research aimed at identifying the bioactive compounds responsible for its therapeutic effects.
Phytochemical studies have shown that P. crocatum contains various secondary metabolites, including phenolic compounds, lignans, flavonoids, terpenoids, and phytosterols. These constituents have been associated with antimicrobial, antioxidant, anti-inflammatory, antidiabetic, hepatoprotective, anticancer, immunomodulatory, and analgesic activities.7–13 Many of these effects involve molecular targets related to oxidative stress, inflammatory signaling, microbial growth, and metabolic enzyme regulation, supporting the relevance of these constituents in drug discovery and therapeutic development.14
Although studies on the biological properties of P. crocatum have increased, the available evidence remains fragmented across phytochemical, pharmacological, and computational studies. Most previous reports have focused on individual extracts, isolated compounds, or specific biological activities, without linking these findings into a broader drug discovery context. As a result, the relationship between the chemical constituents of P. crocatum, their biological activities, and their molecular targets remains insufficiently synthesized. Unlike previous reviews that mainly summarized traditional uses, phytochemical profiles, or selected pharmacological effects, this review integrates phytochemistry, multitarget pharmacology, molecular mechanisms, and lead compound prioritization within a drug discovery framework.
Therefore, following a PRISMA-guided literature screening approach, this review aimed to systematically summarize the phytochemical constituents and pharmacological activities of P. crocatum and to integrate evidence from in vitro, in vivo, and in silico studies. Specifically, this review sought to: (i) compile the major classes of compounds reported from P. crocatum; (ii) summarize their reported biological activities and possible mechanisms of action; (iii) identify links between selected compounds and molecular targets; and (iv) evaluate their relevance as potential lead structures for future drug discovery and development.
Methods
This systematic review was conducted in accordance with the PRISMA 2020 guidelines.15 We conducted a systematic literature search in Scopus and PubMed to identify studies published between 2000 and 2026 related to the phytochemical constituents and pharmacological activities of P. crocatum. The search terms included combinations of keywords related to the plant name (Piper crocatum, red betel, red betel leaf/leaves), phytochemical classes (phenolic compounds, flavonoids, lignans, neolignans, terpenoids, steroids, phytosterols, alkaloids, tannins, quinones, and saponins), biological activities (antibacterial, antifungal, antidiabetic, antioxidant, anti-inflammatory, and anticancer), and drug discovery-related approaches (in vitro, in vivo, in silico, molecular docking, drug design, pharmacological mechanism, absorption, distribution, metabolism, excretion, and toxicity [ADMET], and structure–activity relationship [SAR]).
A total of 180 records were initially identified from Scopus (n = 161) and PubMed (n = 19). Before manual screening, 19 duplicate records were removed, leaving 161 records for title and abstract screening. After title and abstract screening, 44 records were excluded because they did not meet the inclusion criteria, lacked relevance to P. crocatum, or did not report relevant phytochemical or pharmacological information, leaving 117 reports for full-text retrieval. Of these, one report could not be retrieved, leaving 116 full-text articles for eligibility assessment. Full-text evaluation resulted in the exclusion of 14 articles, including seven articles that were not relevant to the research scope and seven review articles. Consequently, 102 studies were included in the final analysis.
The inclusion criteria were as follows: (1) peer-reviewed original research articles published in English; (2) studies investigating extracts, fractions, or isolated compounds from P. crocatum; (3) studies reporting biological activities supported by in vitro, in vivo, or in silico approaches; and (4) studies providing phytochemical or compound identification data, where applicable, such as nuclear magnetic resonance (NMR), mass spectrometry (MS), fourier-transform infrared spectroscopy (FTIR), or gas chromatography–mass spectrometry (GC–MS). The exclusion criteria included duplicate records, review articles, non-scientific publications, studies lacking clear species identification, and reports without relevant phytochemical or pharmacological data, such as purely ethnobotanical descriptions. For several compounds reported from P. crocatum, additional bioactivity data were retrieved from studies in which the same compounds were isolated from other natural sources or synthesized. These complementary data were used to assess the pharmacological and drug development potential of the compounds rather than to represent the direct activity of P. crocatum extracts alone. A detailed overview of the literature search and study selection process is presented in the PRISMA 2020 flow diagram (Figure 1).
 |
Figure 1 PRISMA 2020 flow diagram of the study selection process. |
Phytochemistry of P. crocatum Ruiz & Pav
P. crocatum Ruiz & Pav. is a plant native to Peru, South America, that has spread to several regions of the world, including Indonesia.16 It is a climbing shrub with elliptical leaves, dark green upper surfaces, and reddish-purple lower surfaces (Figure 2).17 Phytochemical screening revealed that red betel contains various secondary metabolites including alkaloids, tannins, phenolics, flavonoids, quinones, terpenoids, steroids, and saponins.18 According to previous studies, forty compounds have been isolated from red betel and characterized using spectroscopic and spectrometric techniques, including ultraviolet–visible (UV–Vis), FTIR, NMR, and mass spectrometry. These compounds comprise seventeen phenolic compounds, ten lignans, seven terpenes, three flavonoids, two steroids, and one hydroxylamine.
 |
Figure 2 P. crocatum plants. |
Phenolic Compounds
Most phenolic compounds isolated from red betel are phenolic glycosides (1–15) with C6-C1 (5, 6, 12–15), C6-C2 (4, 9–11), and C6-C3 (1–3, 7, and 8) skeletons. These structural types indicate the contribution of simple phenolic and phenylpropanoid-derived metabolites to the phytochemical profile of this species. In addition to phenolic glycosides, phenolic amide derivatives have also been reported. Compound 16 was isolated from the methanol extract, whereas piperyamide A (17) was detected in the ethyl acetate extract of red betel leaves.19,20 The structures of the phenolic compounds isolated from P. crocatum are shown in Figure 3.
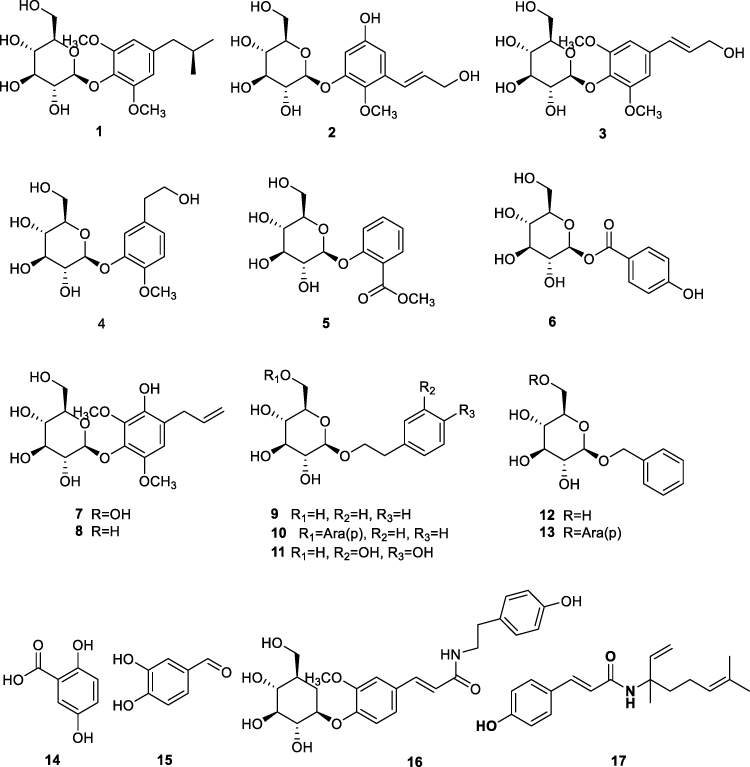 |
Figure 3 Phenolic compounds of P. crocatum: (8R)-8-(4-hydroxy-3,5- dimethoxy)-propane-8-ol-4-O-β-D-glucopyranoside (1); 3-[(1E)-3-hydroxy-1-propen-1-yl]-2,5-dimethoxyphenyl-D-glucopyranoside (2); syringin (3); cimidahurinin (4); methylsalicylate-2-O-β-D-glucopyranoside (5); 4-Hydroxybenzoic acid-D-glucosylester (6); 4-Allyl-2,6-dimethoxy-3-hydroxy-1-D-glucopyranoside (7); erigeside II (8); β-phenylethyl-β-D-glucoside (9); icariside D1 (10); Hydroxytyrosol-1glucopyranoside (11); benzyl-β-D-glucoside (12); phenylmethyl-6-O-α-L-arabinofuranosyl-β-D-glucopyranoside (13); gentisic acid (14); catechaldehyde (15); N-trans-feruloyltyramine-4′-O-β-D-glucopyranoside (16); and piperyamide A (17). |
Lignans
In addition to phenolic compounds, red betel leaves contain lignans.21 Lignans are natural products formed through the oxidative dimerization of two phenylpropanoid units.22,23 Based on the linkage pattern between the phenylpropanoid units, lignans can be broadly classified into classical lignans and neolignans. Two classical lignans belonging to the 2,6-diarylfurofuran subtype (18 and 19) have been isolated from the methanol extract of red betel leaves.24,25 In addition, five bicyclo[3.2.1]octane-type neolignans (20–24) and three other lignan-related compounds (25–27) were reported from the methanol extract.26–28 These findings indicate that lignans and neolignans represent one of the major compound groups in P. crocatum. Their structures are shown in Figure 4.
 |
Figure 4 Structure of isolated lignans from P. crocatum; syringaresinol-β-D-glucoside (18); 2-(5’,6’-dimethoxy-3’,4’-methylenedioxyphenyl)-6-(3’,4’,5-trimethoxyphenyl)-dioxabiclo [3,3,0] octane (19); pipcroside A (20); pipcroside B (21); pipcroside C (22); crocatin A (23); crocatin B (24); pipercrocatin (25); deacethyl pipercrocatin (26), and nectamazin A (27). |
Flavonoids
Although phytochemical screening has indicated the presence of flavonoids in red betel, only three flavonoids (28–30) have been successfully isolated and structurally characterized.24,29 These compounds belong to the flavone subclass (Figure 5). The relatively small number of isolated flavonoids compared with phenolic compounds and lignans suggests that flavonoids are present as minor but pharmacologically relevant constituents in P. crocatum.
 |
Figure 5 Flavonoid of P. crocatum; vitexin 2″- O-rhamnoside (28); pachypodol [4′,5-dihydroxy-3,3′,7- trimethoxyflavone] (29); and kumatakenin [4′,5-dihydroxy-3,7- dimethoxyflavone] (30). |
Terpenoids
Terpenoid-related compounds have also been identified from P. crocatum. These include one monoterpenoid derivative (31), five norisoprenoids (32–36), and one diterpenoid (37) which were reported from the ethyl acetate fraction of red betel leaves.19,30 Their structures are shown in Figure 6. The presence of these terpenoid derivatives expands the chemical diversity of P. crocatum beyond phenolic and lignan-based metabolites.
 |
Figure 6 Monoterpens and sesquiterpenes of P. crocatum; (S)-menthiafolic acid (31); 5β,6β-dihydroxy-3α-(β-D-glucopyranosyloxy)-7E-megastigmen-9-one (32); (3E)-4- [(1S,2S,4S)-4-(β-D-glucopyranosyloxy)-1,2-dihydroxy-2,6,6-tri-methylcyclohexyl] 3-buten-2-one (33); (6S,9S)-roseoside (34); cuneataside E (35), ioliolide (36); and cassipourol (37). |
Steroids
Two phytosterols, β-sitosterol (38) and stigmasterol (39), have been isolated from red betel (Figure 7).20,24 Steroids are characterized by a tetracyclic carbon framework consisting of three six-membered rings and one five-membered ring. The identification of these phytosterols is important because sterol-type compounds are commonly associated with membrane-related biological effects and may contribute to the pharmacological profile of P. crocatum.
 |
Figure 7 Steroid structure of P. crocatum; β-sitosterol (38) and stigmasterol (39). |
Other Compounds
A hydroxylamine derivative, piperyamine A (40) was isolated from the methanol extract of red betel leaves (Figure 8).20 This compound has a unique structure with an unsaturated side chain. Although its biosynthetic origin has not been fully established, the presence of this hydroxylamine derivative highlights the structural diversity of metabolites produced by P. crocatum. Hydroxylamine-containing natural products are relatively uncommon, and their occurrence in red betel provides an additional example of the chemical uniqueness of this species.
 |
Figure 8 Structure of Piperyamine A (40). |
Overall, these compound classes provide different structural and physicochemical features that may influence their biological behavior. Phenolic compounds and flavonoids contain hydroxylated and methoxylated aromatic systems that may support redox-related and enzyme-modulating activities, whereas lignans and neolignans provide phenylpropanoid-derived frameworks relevant to antimicrobial and anti-inflammatory potential. In contrast, terpenoids and phytosterols contribute more lipophilic scaffolds that may influence membrane-related interactions but may also present limitations in solubility and bioavailability.
Pharmacological Activities of P. crocatum Ruiz & Pav
Antimicrobial and Antibiofilm Activity
P. crocatum has shown antimicrobial activity against a wide range of Gram-positive and Gram-negative bacteria, fungi, and selected protozoa, indicating its potential relevance in the management of infectious diseases. The reported activities included crude extracts, fractions, and isolated compounds, with varying degrees of effectiveness, depending on the chemical class and purity level (Table 1).
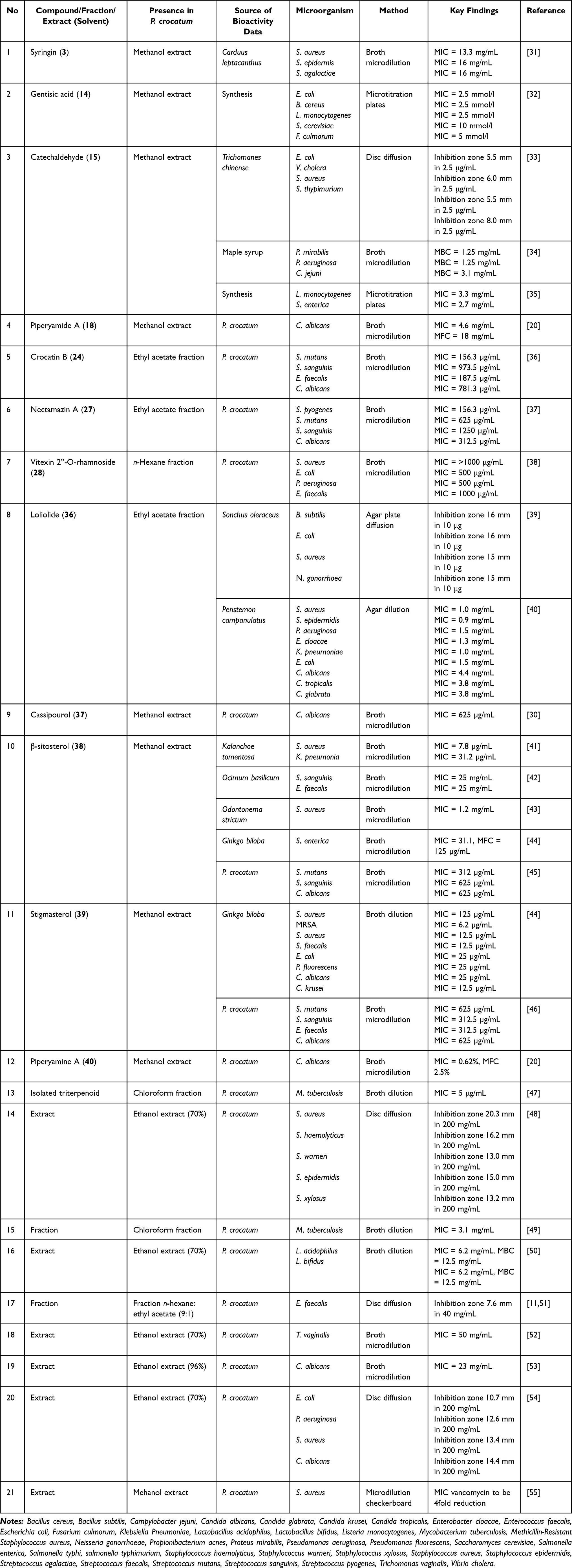 |
Table 1 Antibacterial and/or Antifungal Activity of Extract, Fraction, and Isolated Compounds |
Antibacterial Activity
The ethanolic extract showed inhibitory effects against S. aureus, S. epidermidis, S. haemolyticus, S. warneri, and S. xylosus with inhibition zones ranging from 13.0 to 20.3 mm at a concentration of 200 mg/mL.48 Activity was also observed against gram-negative pathogens including E. coli, P. aeruginosa, and K. pneumoniae.54 Despite the relatively high concentration of this extract, this indicates the presence of bioactive constituents capable of inhibiting bacterial growth. More potent activity was observed for the isolated compounds. β-Sitosterol (38) showed significant activity against S. aureus (minimum inhibitory concentration [MIC] = 7.8 µg/mL) and K. pneumoniae (MIC 31.2 µg/mL) and showed moderate inhibition against oral pathogens such as S. mutans and S. sanguinis (MIC 312–625 µg/mL).41,45 Crocatin B (24) also showed inhibitory activity against S. mutans and E. faecalis with MIC of 156.3 and 187.5 µg/mL, respectively.36 In addition, nectamazin A (27) exhibited MIC of 156.3 µg/mL against S. pyogenes.37 Stigmasterol (39) showed activity against S. aureus, methicillin-resistant S. aureus (MRSA), and E. coli, with MIC values as low as 6.2 µg/mL in some models.44 These findings suggested that the phytosterol skeleton may significantly contribute to its antibacterial potential. Phenolic compounds such as gentisic acid (14) and catechaldehyde (15) exhibit moderate inhibitory activity against several microorganisms, including E. coli, B. cereus, L. monocytogenes, S. enterica, P. aeruginosa, and V. cholerae.32–35,44 The presence of hydroxyl and aldehyde groups may facilitate interactions with membranes and induce oxidative stress, thereby contributing to the bacteriostatic or bactericidal effects. The triterpenoid isolated from the chloroform fraction is of particular interest, as it exhibits activity against M. tuberculosis (MIC 5 µg/mL),47 highlighting its potential relevance in the discovery of antituberculosis drugs. Considering the urgent need for new anti-TB agents, these findings require further mechanistic and in vivo validation.
Significantly, the methanol extract showed a synergistic interaction with vancomycin, resulting in a fourfold decrease in the MIC values against S. aureus.55 This indicates potential resistance-modifying properties, possibly through membrane disruption or efflux pump interference. Combination therapies of this type may be useful in overcoming antimicrobial resistance.
Antibiofilm and Oral Pathogen Relevance
The antimicrobial profile of P. crocatum is particularly prominent in oral pathogens. β-Sitosterol and stigmasterol have been shown to inhibit S. mutans, S. sanguinis, and E. faecalis, which are closely associated with dental plaque formation and endodontic infections.45,46 The results of clinical applications further reinforce this relevance. Water extracts have been evaluated as root canal irrigants, and 30% extracts effectively removed residual layers in a clinical setting. Red betel leaf gel increased clinical attachment levels by 31.4% when utilized as adjunctive periodontal therapy, and a mouthwash formulation containing ethanol extract reduced dental plaque accumulation (Table 2).7,56–58 These findings indicate not only antimicrobial properties but also antibiofilm properties, which are crucial for treating chronic oral infections.
 |
Table 2 Application of P. crocatum Extract |
Molecular docking studies have provided a possible mechanism for the antibiofilm potential. β-sitosterol was predicted to bind to glucan-binding protein C (GbpC) and sortase C (SrtC), both of which are involved in biofilm formation and bacterial adhesion.45 Suppression of these targets could interrupt extracellular polysaccharide synthesis and bacterial colonization, suggesting a plausible explanation for the observed reduction in plaque formation.
Antifungal Activity
Several compounds exhibit antifungal activity, particularly against C. albicans and other related species. Among the compounds that showed inhibitory activity against C. albicans, piperyamide A, piperyamine A, cassipourol, stigmasterol, β-sitosterol, and nectamazin A showed the same MIC value of 625 μg/mL, whereas crocatin B showed an MIC of 312 μg/mL. In silico studies suggest that several of these compounds may interfere with fungal membrane sterol formation by targeting enzymes involved in ergosterol biosynthesis. Piperyamide A, stigmasterol, and piperyamine A interact with ERG1, ERG2, ERG11, and ERG24 with inhibition constants (Ki) in the submicromolar to micromolar range.20 This mechanism is consistent with antifungal strategies targeting lanosterol 14α-demethylase (CYP51) as summarized in Table 3. Cassipourol, β-sitosterol, crocatin A, crocatin B, and nectamazin A have been predicted to bind to CYP51, further supporting their multitarget antifungal potential.30,36,37,45 Although ADMET predictions indicate limitations in the oral bioavailability of some compounds, the convergence of ergosterol biosynthesis highlights a promising route for targeted antifungal agents.
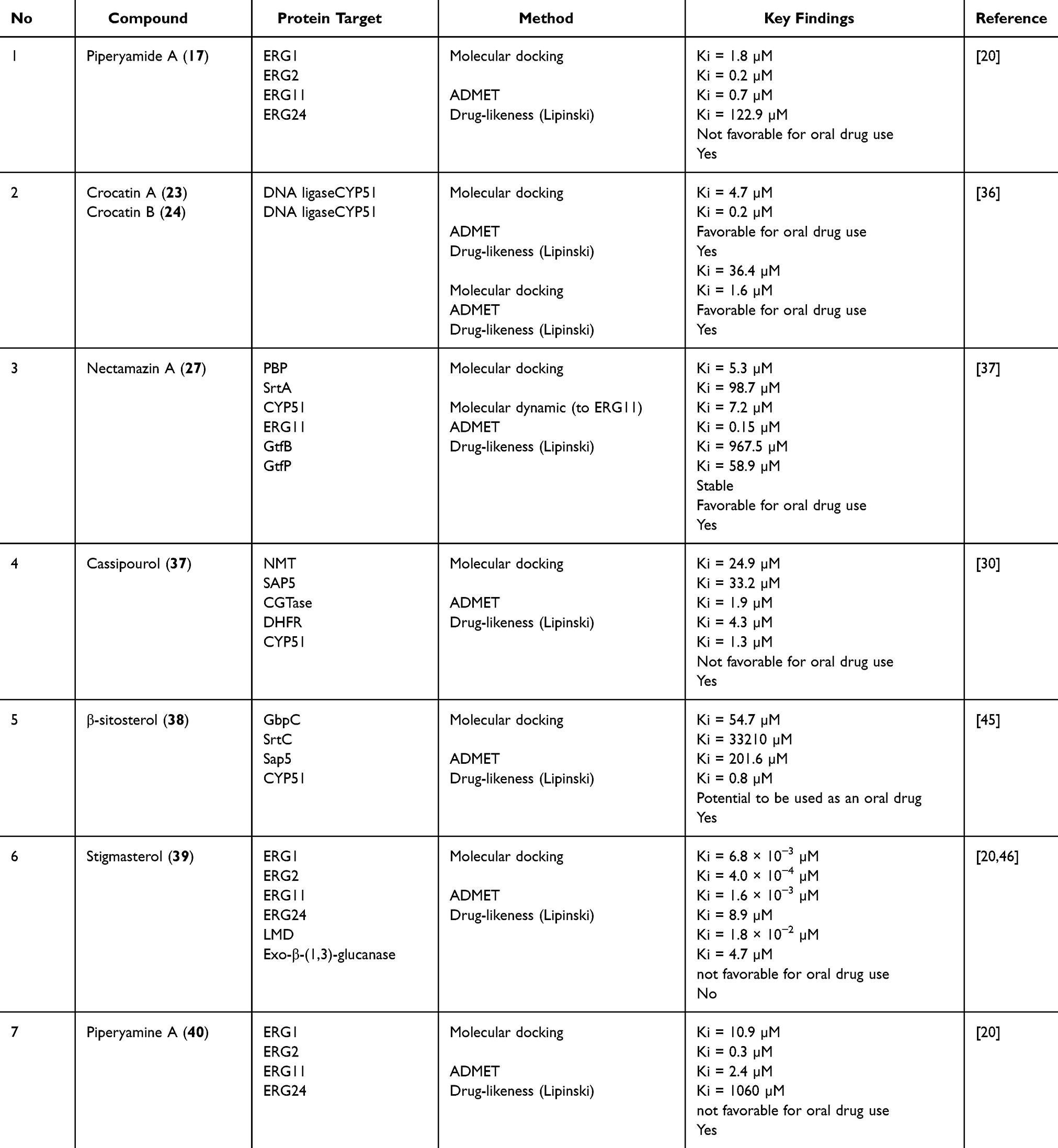 |
Table 3 In silico Study of P. crocatum Compounds and Protein-Related Bacteria or Fungi |
Overall, these findings highlight ergosterol synthesis as a potential molecular target for mediating antifungal activity of P. crocatum. This mechanism-based insight is particularly relevant for traditional therapeutic practices where red betel leaf preparations have been used to manage symptoms related to vaginal infections, including fluor albus. A decoction of P. crocatum leaves effectively reduced the symptoms of Fluor albus in women of reproductive age, both in adults and adolescents. There were significant differences in pH levels before and after the application of boiled red betel leaf water, with pH averaging 2.00 before and 1.21 after treatment.3,60
However, despite its broad spectrum of activity, the following limitations must be acknowledged: the majority of studies on the extract have been carried out using high concentrations (>100 mg/mL), which directly limits its clinical relevance (1); inconsistent standardization of extract composition (2); poor reporting of bacterial killing kinetics or post-antibiotic effects (3); inconsistency in biofilm inhibition testing (4); and lack of pharmacokinetic data (5). Thus, despite the emergence of sterols and triterpenoids as promising frameworks, most antimicrobial evidence remains at the preliminary level.
Redox-Related and Enzyme-Modulating Bioactivity
P. crocatum leaves have demonstrated antioxidant activity through various experimental approaches, including free-radical chemistry systems, lipid peroxidation models, enzyme inhibition assays, and in silico studies of target proteins associated with oxidative stress. In general, extracts and compounds isolated from this plant can scavenge free radicals, inhibit lipid peroxidation, reduce transition metal ions, and modulate the activity of enzymes associated with oxidative stress.
The 70% ethanol extract exhibited 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity with an IC50 value of 85.2 µg/mL. Another study comparing several solvents reported that water extract had the strongest activity (IC50 81.2 µg/mL), followed by ethanol extract (185.3 µg/mL), ethyl acetate extract (202.8 µg/mL), and n-hexane extract (552.2 µg/mL).61,62 This indicates that more polar components likely contribute dominantly to the antioxidant activity. At the pure compound level, pachypodol (29) and kumatakenin (30) showed stronger activities than the extracts. In the DPPH assay, pachypodol had an IC50 of 58.3 µg/mL and kumatakenin 204.8 µg/mL (Table 4). In the superoxide dismutase (SOD) mimetic activity test, both showed excellent activity, with pachypodol reaching an IC50 of 7.1 µg/mL and kumatakenin 42.3 µg/mL. Density Functional Theory (DFT) studies showed that the hydrogen atom transfer (HAT) mechanism is thermodynamically favored by both flavonoids, with the C4′–OH group identified as the most reactive site for hydrogen donation.29 These data indicate that flavonoids play an important role in the free radical scavenging capacity of P. crocatum.
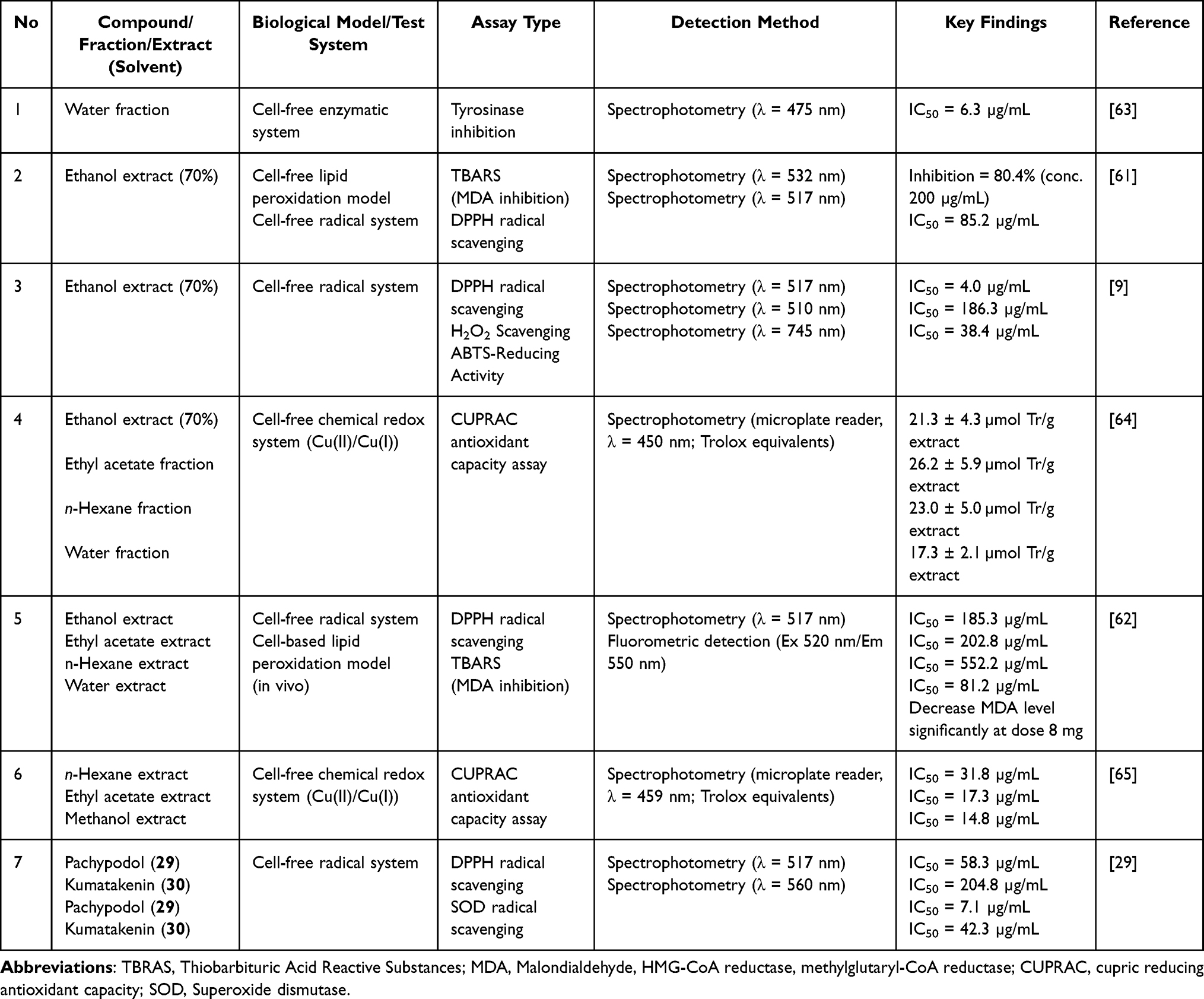 |
Table 4 Redox-Related and Enzyme-Modulating Bioactivities of P. crocatum Extracts, Fractions, and Isolated Compounds |
In addition, P. crocatum extract was also tested in a lipid peroxidation model using the Thiobarbituric acid reactive substances (TBARS) method, which measures malondialdehyde (MDA) levels as an indicator of lipid oxidative damage. The 70% ethanol extract inhibited lipid peroxidation by 80.4% at 200 µg/mL.61 In the TBARS-based in vivo model, the administration of 8 mg significantly reduced MDA levels.62 These results indicate that the antioxidant activity of P. crocatum is not limited to the chemical radical system, but is also capable of suppressing the formation of lipid oxidation products.
Based on the results of the cupric reducing antioxidant capacity (CUPRAC) method, 70% ethanol extract, ethyl acetate fraction, n-hexane fraction, and water fraction showed reduction capacities in the range of 17–26 µmol Trolox equivalent per gram of extract.64 These findings indicate that P. crocatum compounds contribute significantly to metal reduction capacity. The antioxidant activity related to enzyme modulation was reported using a tyrosinase inhibition test. The water fraction showed strong tyrosinase inhibitory activity (IC50 of 6.3 µg/mL).63 Tyrosinase is an enzyme that plays a role in the oxidation of phenols and the formation of melanin; therefore, its inhibition is often associated with antioxidant activity and potential applications in dermatology. As shown in Table 5, in silico studies showed that P. crocatum compounds, such as piperine, piperamide-C7:2, and trichostachine, interact with tyrosinase with fairly good binding affinity (range −6.8 −7.0 kcal/mol). Molecular dynamics simulations revealed the stability of the ligand–protein complex, and ADMET analysis and drug-likeness parameters (Lipinski) indicated a profile supporting oral utilization.63 These results confirm the experimental data that, in addition to radical-scavenging activity, several P. crocatum compounds can inhibit enzyme activity related to oxidative processes.
 |
Table 5 In silico Study of P. crocatum Compounds and Protein-Related Antioxidant Activity |
Although P. crocatum exhibits strong antioxidant activity in vitro, current evidence is largely derived from chemical assays. Without mechanistic validation of cellular systems and pharmacokinetic characterization, classification as a viable source of antioxidant drugs remains premature. Nevertheless, pachypodol has emerged as a promising redox-active framework with the potential for further optimization.
Immunomodulatory and Anti-Inflammatory Activity
Modulation of Innate Immune Response
The immunomodulatory activity of P. crocatum has been demonstrated through its ability to modulate macrophage functions. In an L. monocytogenes infection model in BALB/c mice, the macrophage phagocytic index and nitric oxide (NO) production were used as indicators of innate immune activation. Administration of the methanol extract at a dose of 450 mg/kg significantly improved macrophage activity and NO levels, showing an efficacy equivalent to that of isolated neolignans (Pc-1 and Pc-2) delivered at a dose of 5 mg/kg.28 This approximately 90-fold difference in dose indicates a much higher potential for the pure compound to stimulate innate immune responses.
Nitric oxide production in stimulated macrophages is regulated by inducible nitric oxide synthase (iNOS), implying that the immunostimulatory effect may involve classical macrophage signaling pathways, such as nuclear factor-kappa B (NF-κB) or the mitogen-activated protein kinase (MAPK) cascade. During microbial infection, these pathways are typically triggered by pattern recognition receptor signals and mediate the transcription of genes associated with antimicrobial defense. However, upstream receptor interactions, particularly the possible involvement of Toll-like receptors (TLRs), have not been directly investigated, resulting in unresolved molecular mechanisms.
Although they had similar immunostimulatory efficacy, significant toxicological differences were observed between the isolated compounds and the crude extracts. Pc-2 caused hydropic degeneration in the liver tissue, indicating hepatocellular stress, whereas Pc-1 and crude extracts did not cause significant histopathological changes and maintained the alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels within acceptable limits. These differences highlight an essential translational concern: higher potency does not necessarily translate into a greater therapeutic potential. Crude extracts may offer a buffering effect that reduces toxicity through antagonistic or balanced synergistic interactions between the phytochemicals. Such interactions may stabilize the immune response and prevent overstimulation. For drug development, Pc-1 appeared to offer the best balance of efficacy and safety, whereas Pc-2 had stronger efficacy, but caused concerns about hepatotoxicity. The crude extract, which was less potent, showed better bioavailability.
Suppression of Pro-Inflammatory Signaling
Most anti-inflammatory studies of P. crocatum use lipopolysaccharide (LPS)-induced RAW 264.7 macrophages, which represents activation of the TLR4–NF-κB pathway. LPS stimulates TLR4; induces IκB kinase (IKK) phosphorylation, IκB degradation, and NF-κB translocation to the nucleus; and subsequently increases the transcription of tumor necrosis factor-alpha (TNF-α), interleukin-1β (IL-1β), IL-6, and iNOS, which generates NO. Experimental data showed that extracts and isolated compounds from P. crocatum suppressed mediators at several points in this pathway, particularly at cytokine and iNOS levels (Table 6). The ethanol extract reduced TNF-α by 51.6%, IL-1β by 21.4%, IL-6 by 68.7%, and NO by 81.2% at 50 µg/mL, suggesting the upstream modulation of inflammatory signaling.14 However, direct measurements of key molecular events, such as IKK phosphorylation or NF-κB nuclear translocation, have rarely been performed; therefore, the exact intervention site remains unknown.
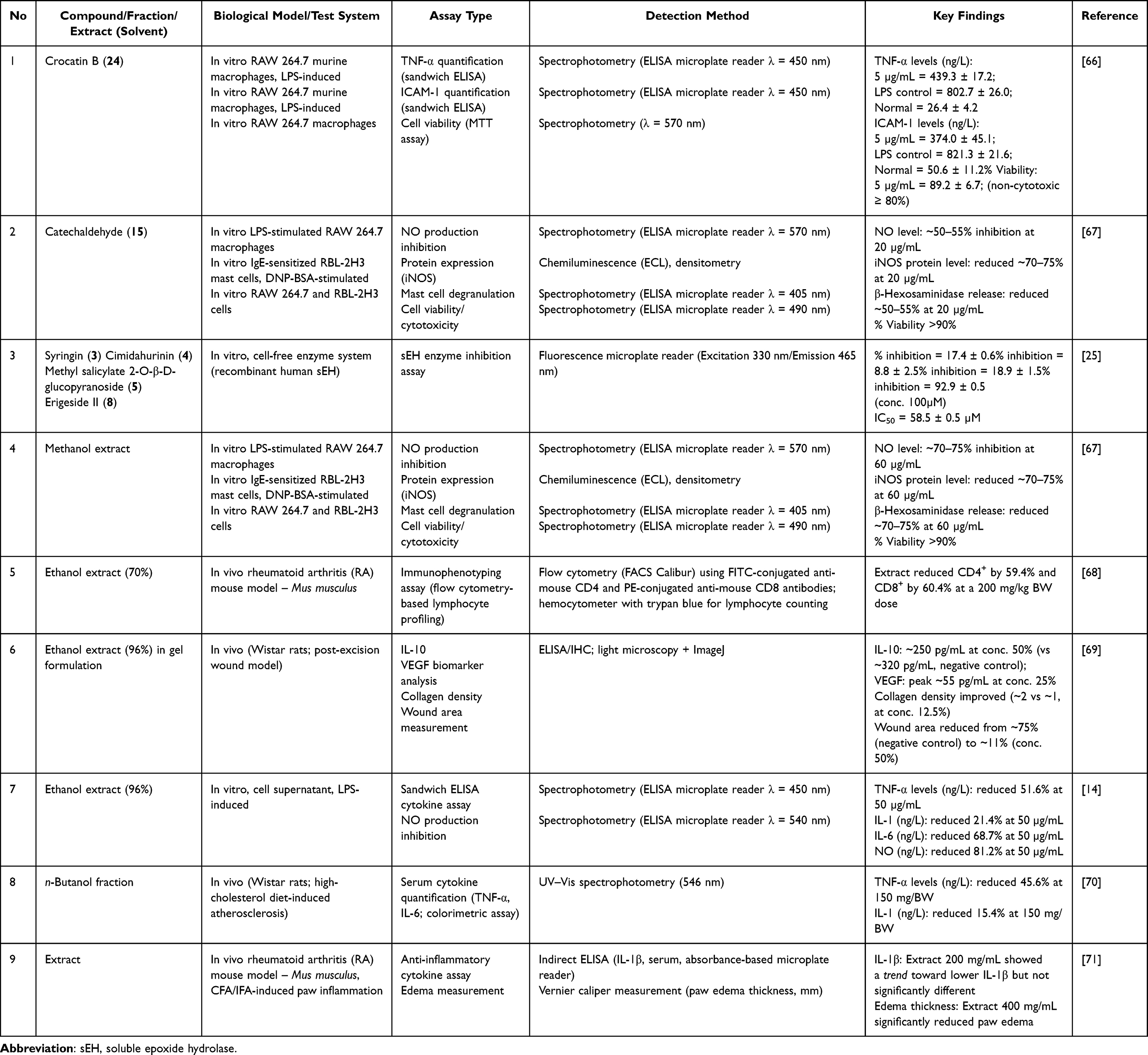 |
Table 6 Anti-Inflammation Activity of P. crocatum Extracts, Fractions, and Isolated Compounds |
At a single compound level, Lely et al reported that a neolignan, crocatin B (24), significantly attenuated inflammatory responses in LPS-stimulated RAW 264.7 cells at 5 µg/mL.66 Measurements using the sandwich enzyme-linked immunosorbent assay (ELISA) method demonstrated a reduction in TNF-α and intercellular adhesion molecule-1 (ICAM-1) concentrations compared with those in the control LPS-treated group, reflecting the inhibition of pro-inflammatory cytokine secretion and adhesion molecule expression. These dual activities imply modulation of upstream regulatory pathways, such as the NF-κB signaling cascade, which controls the transcriptional regulation of TNF-α and ICAM-1 in activated macrophages. Cell toxicity assessments showed that the viable cell counts remained above 80% over the tested concentration range, indicating that the observed biological activity was not due to general cellular toxicity. Preliminary safety data highlighted the selectivity of the compound toward inflammatory targets. In terms of drug design, the neolignan skeleton, which is rich in phenolic and methoxy moieties, likely contributes to key molecular interactions through hydrogen bonding and improved membrane permeability. Such structural features may underlie the observed bioactivity and offer a rational approach for further structure-activity relationship analysis. Overall, these findings suggest that neolignans from P. crocatum are prospective multitarget anti-inflammatory agents. However, further in vivo and in silico validation is required to confirm their target engagement and translational potential.
NO and iNOS Regulation
P. crocatum methanol extract inhibited NO production by approximately 70–75% and decreased iNOS protein expression by approximately 70–75% at 60 µg/mL. Meanwhile, cell viability remained above 90%, indicating that the inhibition of inflammatory mediators was not caused by cellular toxicity. Catechaldehyde (15) showed similar effects at lower concentrations (20 µg/mL), indicating that the potential of the single compounds was stronger than that of the extracts.67 The decrease in iNOS expression indicated that the activity was not limited to NO scavenging but was also implicated in the regulation of enzyme transcription or translation. However, most studies have not evaluated the activation of signal transducer and activator of transcription 1 (STAT1) or the MAPK pathway, which also contribute to iNOS expression; therefore, the mechanism remains partially undefined.
Enzyme-Targeted Anti-Inflammatory Activity
Several compounds from P. crocatum also target the enzymes associated with inflammatory signaling. Erigeside II (8) showed a 92.9% inhibition of soluble epoxide hydrolase (sEH) at 100 µM (IC50 58.5 µM).25 sEH is an attractive target as it regulates the degradation of anti-inflammatory fatty acid epoxides into pro-inflammatory diols. Although the percentage of inhibition is high, the IC50 value in the tens of micromolar range represents moderate potency compared to the latest generation of synthetic sEH inhibitors (typically in the nanomolar range). Thus, erigeside II could be viewed as a more reasonable initial scaffold than a direct candidate.
In silico studies further support this anti-inflammatory potential (Table 7). Docking simulation showed that apigenin had an affinity of −9.9 kcal/mol for Cyclooxygenase-2 (COX-2), higher than kaempferitrin and rutin (−8.7 kcal/mol).72,73 Structurally, apigenin is a flavonoid aglycone with a lower molecular weight and more balanced lipophilicity than the glycoside forms. Glycosylated flavonoids (rutin and kaempferitrin) show high affinity; however, their ADMET profiles do not support their oral use, likely because of their large molecular size and high polarity. These results indicate that flavonoid aglycones are feasible COX-2 modulator candidates.
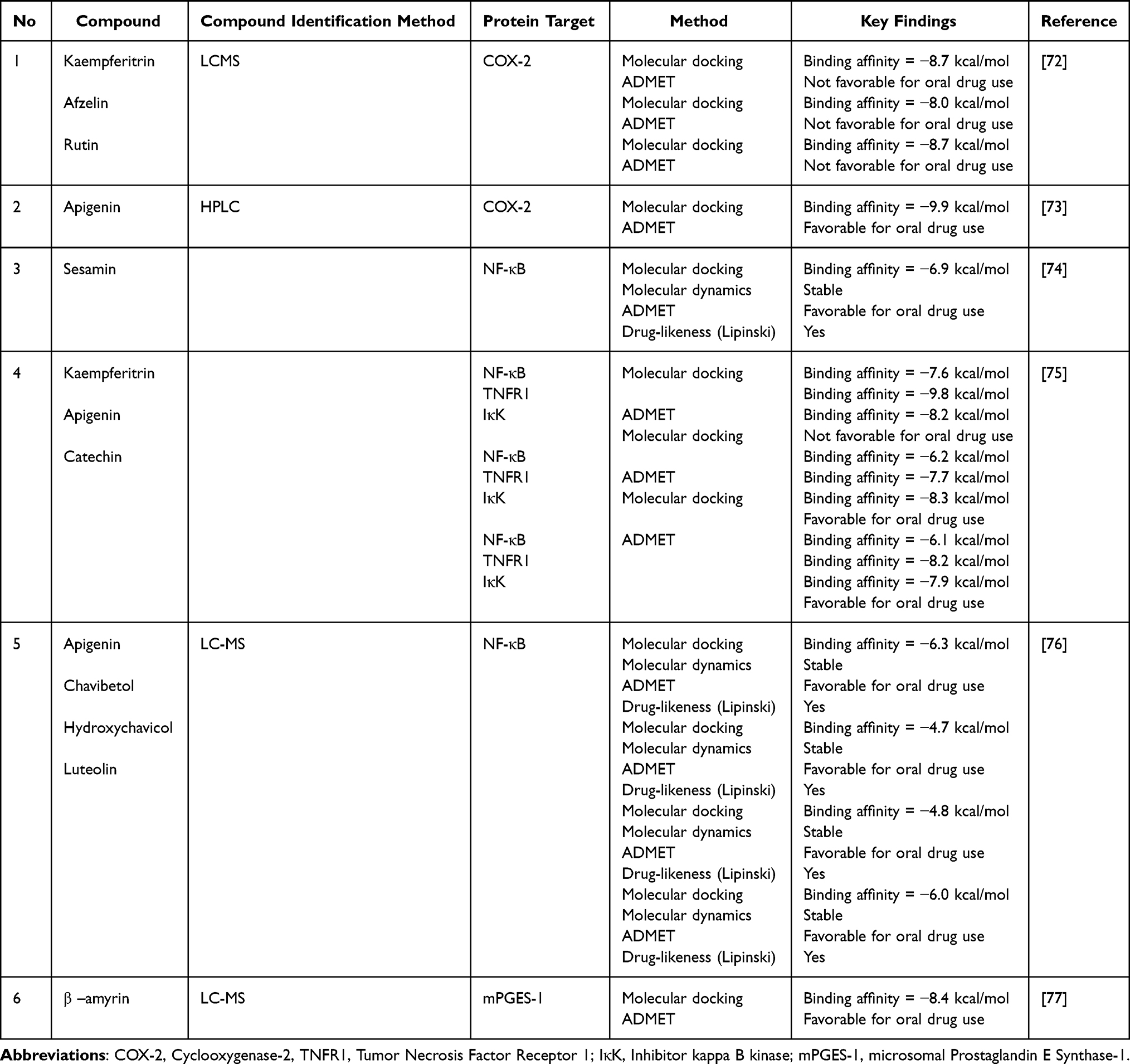 |
Table 7 In silico Study of P. crocatum Compounds and Protein-Related Anti-Inflammation Activity |
Docking simulations were performed for NF-κB, TNFR1, and IKK. The results showed that kaempferitrin had a strong affinity for TNFR1 (−9.8 kcal/mol), while apigenin and catechin showed values of −7 to −8 kcal/mol with better ADMET profiles.74–76 Sesamin displayed complex stability in molecular dynamics simulations and met Lipinski’s criteria, rendering it a more drug-like candidate than the glycosylated flavonoids. However, docking does not always correspond directly to biological activity in living cells, and thus requires further biochemical validation.
In vivo Evidence and Immunomodulation
In vivo studies provide additional evidence of systemic anti-inflammatory effects. In a rat atherosclerosis model, the n-butanol fraction reduced TNF-α levels by 45.6% at 150 mg/kg BW, indicating systemic activity.69 Other studies have demonstrated that concentrated extracts reduced edema in a rheumatoid arthritis model, while 70% ethanol extract at 200 mg/kg reduced CD4+ and CD8+ lymphocyte populations by approximately 60%.68 These findings suggest that P. crocatum may influence both the innate and adaptive immune responses. However, the relatively high doses used and absence of pharmacokinetic data raise questions regarding the clinical feasibility and potential risk of non-selective immunosuppression.
Conceptual Integration: Immune Homeostasis Regulation
Although the immunostimulatory and anti-inflammatory effects initially seem contradictory, they are likely to be derived from the same modulation of the macrophage signaling networks. During microbial infection, moderately activated macrophage pathways strengthen phagocytosis and pathogen elimination via NO production. Conversely, overstimulation of the same pathways under sterile inflammatory conditions induces the excessive production of cytokines and inflammatory mediators. Thus, the pharmacological profile of P. crocatum indicates dependent regulation of macrophage signaling, rather than simple immune stimulation or suppression. Both activities accumulate on the TLR4–NF-κB–iNOS signaling axis. Activation of this pathway promotes host defense in infection models, whereas suppression of downstream mediators, such as TNF-α, IL-6, and NO, limits inflammatory damage in LPS-induced systems. The dual physiological role of NO—as an antimicrobial agent and inflammatory mediator illustrates this regulatory balance. A dual modulation model was proposed to conceptualize context-dependent immunological behavior (Figure 9). This model emphasizes that P. crocatum does not act as a simple immunostimulant or immunosuppressant but rather as a context-dependent regulator of immune homeostasis. Such dual functionality is particularly relevant for therapeutic strategies targeting infections accompanied by uncontrolled inflammation.
 |
Figure 9 Dual immunological modulation of P. crocatum in infection and inflammatory contexts. |
Overall, our findings indicate that P. crocatum functions as a bifunctional regulator of immune homeostasis, enhancing innate immune responses during infection while suppressing excessive inflammatory signaling. However, the existing studies have largely relied on a limited number of experimental models that commonly use higher concentrations or doses. Further research integrating molecular pathway analysis, pharmacokinetic characterization, and multi-model validation is required to clarify the potential of P. crocatum as a source of immunoregulatory agents.
Anticancer Activity
Extract-Level Cytotoxicity in Breast and Cervical Cancer Models
The cytotoxic profile of P. crocatum extract showed significant variation between the cancer subtypes and extraction solvents (Table 8). The most prominent activity was observed in MCF-7 breast cancer cells, where the 96% ethanol extract reached an IC50 of 6.7 μg/mL, indicating strong growth inhibition.8 This potency contrasts significantly with the activity of the methanol extract on 4T1 cells (IC50 = 120 μg/mL) and HeLa cells (LC50 up to 0.8 mg/mL), highlighting a striking difference in selectivity.78,79 In T47D cells, the methanol extract suppressed proliferation (IC50 = 44.25 μg/mL) but led to minimal apoptosis, indicating a cytostatic rather than cytotoxic effect.80 Inhibition of extracellular signal-regulated kinase 1/2 (ERK1/2) phosphorylation suggests disruption of the MAPK pathway, a key regulator of proliferation and viability in breast cancer. Further suppression of insulin-induced ERK activation implies the upstream regulation of receptor-mediated signaling. These results indicate that P. crocatum extract may act primarily through the blockade of proliferation signaling, rather than through the direct induction of apoptosis. However, the lack of mechanistic analysis beyond ERK phosphorylation limits the pathway identification. Additionally, the IC50 values of the extracts above 40 μg/mL (except for MCF-7) represent a barrier to translation unless isolated and enriched active constituents are identified.
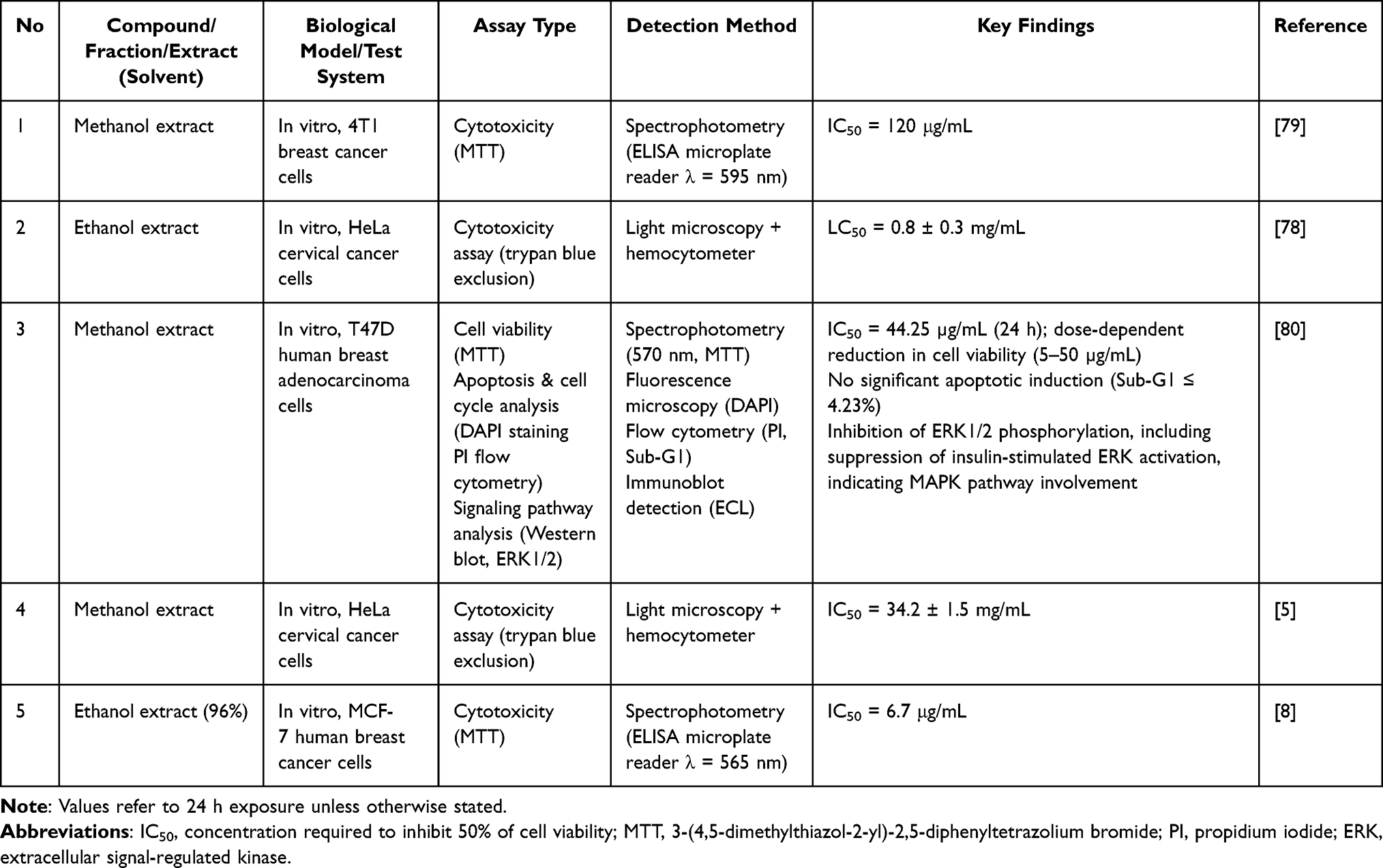 |
Table 8 Anticancer Activity of P. crocatum Extracts, Fractions, and Isolated Compounds |
In silico Targeting of Oncogenic Proteins
Docking analysis showed that β-asarone, methylpiperonyl ketone, and p-coumaric acid target human epidermal growth factor receptor 2 (HER-2), matrix metalloproteinase-9 (MMP-9) and TNF-α, which play key roles in tumor progression, invasion, and modulation of the inflammatory microenvironment. Among these, β-asarone consistently exhibited strong binding scores, particularly to MMP-9 (−216.1 kcal/mol), indicating potential anti-metastatic activity through the inhibition of extracellular matrix degradation.81 p-Coumaric acid showed the strongest predicted MMP-9 interaction (−220.9 kcal/mol), despite its relatively simple phenylpropanoid structure. Importantly, all three compounds met Lipinski’s criteria and exhibited favorable ADMET predictions, positioning them as viable small-molecule candidates for oral administration.
Antidiabetic and Antihyperuricemic Activities
Cellular and Enzymatic in vitro Evidence
Two lignans (Pipcroside B and C) from the ethyl acetate fraction showed moderate potential in cellular pyruvate dehydrogenase (PDH) assays (IC50 ≈ 80–100 μM), indicating the potential for enhanced mitochondrial glucose oxidation (Table 9).27 However, these micromolar concentrations represent a moderate intrinsic potential compared with clinically relevant metabolic modulators. On the other hand, a few extracts showed significant α-glucosidase inhibition at low μg/mL ranges (ethyl acetate IC50 = 5.3 μg/mL),65 suggesting that semi-polar phytochemicals may act as competitive enzyme inhibitors. Marked differences between mg/mL and μg/mL IC50 reports among studies emphasize the need for standardization and methodological consistency.
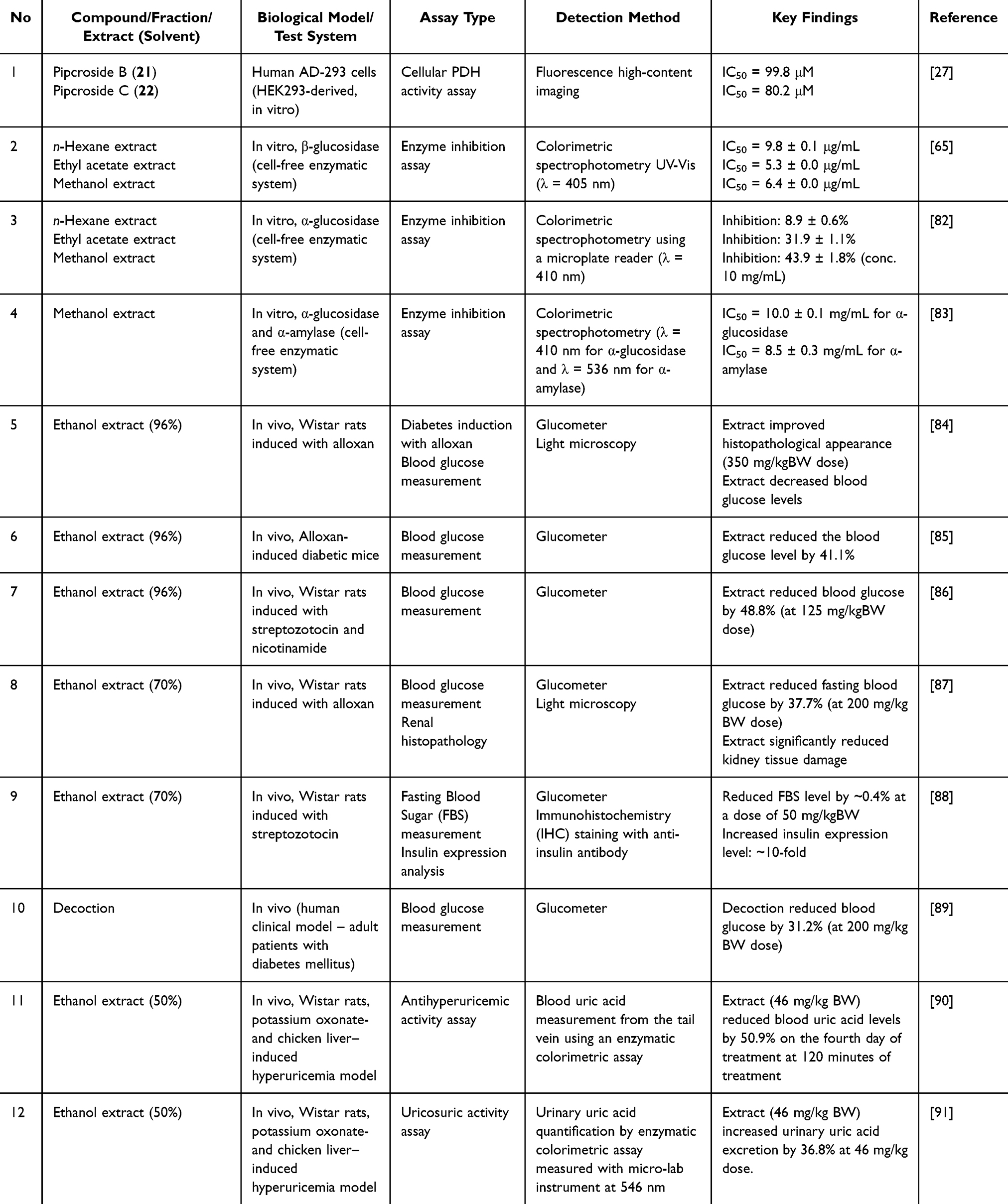 |
Table 9 Antidiabetic Activity of P. crocatum |
In vivo Antidiabetic Activity
Studies in rats have consistently shown glucose-lowering effects (37–49%) in streptozotocin (STZ)- and alloxan-induced models.85–87 Interestingly, an approximately ten-fold increase in insulin expression was observed in rats treated with STZ and exposed to 70% ethanol extract, suggesting a potential protective or regenerative effect on β-cells. The improved renal histopathology in diabetic rats further supports the protective effect against diabetic nephropathy. Collectively, these findings suggest multitarget modulation, including maintenance of pancreatic β-cells, reduction of oxidative stress, improved peripheral glucose utilization, and suppression of intestinal glucose absorption. Clinical studies reporting a 31.2% decrease in glucose levels reinforce translational relevance,89 although controlled and randomized clinical trials are required.
Antihyperuricemic Activity
The ethanol extract of P. crocatum also demonstrated antihyperuricemic activity by reducing uric acid levels by 50.9% and increasing uricosuria by 36.8% in rats.90,91 This indicates the presence of additive metabolic regulatory effects, which likely occur through the inhibition of xanthine oxidase or the modulation of renal urate transporters. It is worth noting that this dual antihyperglycemic–antihyperuricemic profile is highly relevant regarding the strong clinical correlation between type 2 diabetes and hyperuricemia.
Hepatoprotective Effect
The potential hepatoprotective effects of P. crocatum extract were assessed in HepG2 human hepatocellular carcinoma cells exposed to acetaminophen (APAP)-induced damage (Table 10). APAP-induced liver damage is primarily mediated through the CYP2E1-dependent bioactivation of N-acetyl-p-benzoquinone imine (NAPQI), resulting in decreased glutathione levels, oxidative stress in the mitochondria, and secretion of inflammatory cytokines. In HepG2 cells, P. crocatum extract exhibited significant hepatoprotective effects at 25 µg/mL, with reductions in lactate dehydrogenase (LDH) (45–50%), AST (35–40%), and ALT (20–25%) levels.92 Interestingly, reactive oxygen species (ROS) levels declined by up to 70% and TNF-α levels were reduced by approximately 50%.93 These findings suggest that hepatoprotection is mediated by attenuation of oxidative stress, inhibition of inflammatory signaling, and reduction of necrotic cell death. However, mechanistic investigations into nuclear factor erythroid 2-related factor 2 (Nrf2) induction, mitochondrial membrane potential, and caspase involvement have not been conducted, thus limiting the identification of pathways.
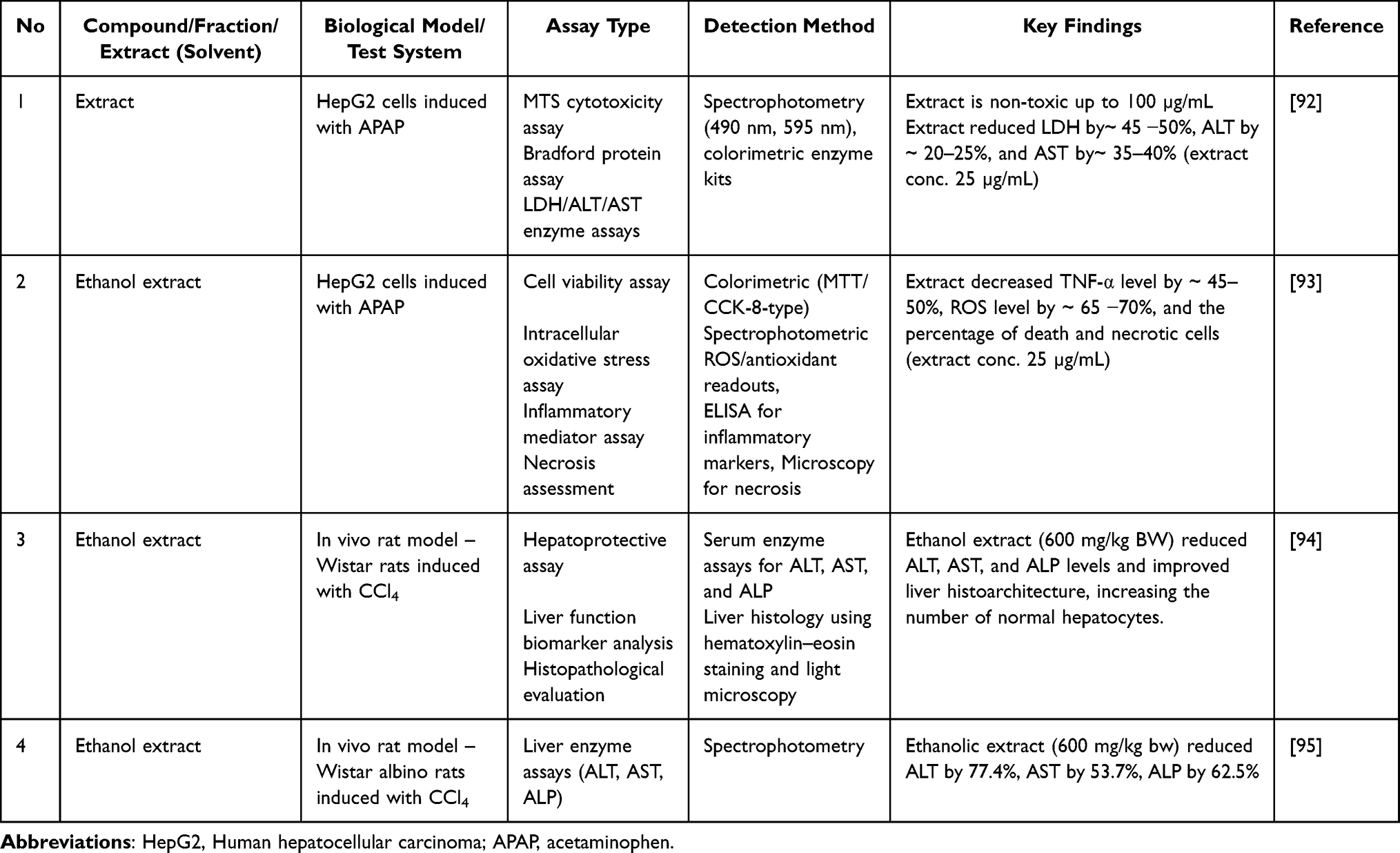 |
Table 10 Hepatoprotective Effect of P. crocatum |
The ethanol extract (600 mg/kg body weight) significantly reduced liver enzyme levels (ALT, AST, and alkaline phosphatase [ALP]) and restored liver structure in a CCl4-induced rat model. The reduction in ALT levels (77.4%) indicated substantial attenuation of liver cell leakage. Histological improvements further imply functional recovery rather than limited biochemical modulation.94,95 However, the high dose required (600 mg/kg) prompts translation-related issues regarding equivalent human dosing, bioavailability of active compounds, and long-term safety profiles.
Docking analysis revealed allyl pyrocatechol and hydroxychavicol as compounds with the potential to bind CYP2E1 (−7.3 kcal/mol).96 Considering the role of CYP2E1 in the bioactivation of APAP and CCl4, partial inhibition may reduce the production of reactive intermediates. However, no enzyme inhibition assays confirmed the suppression of CYP2E1 activity, the docking affinity indicated moderate interaction, and selectivity toward other CYP isoforms was not evaluated. Therefore, CYP2E1 inhibition remains hypothetical.
Other Activities
Adulticidal and Larvicidal Activity
As shown in Table 11, the ethanol extract exhibited adult mosquito-killing effects against female Aedes aegypti mosquitoes with moderate toxicity (a maximum mortality rate of 43.3% and an LC50 value of 9901 ppm).97 Notably, the lack of statistically significant differences among the various concentrations (ANOVA, p = 0.541) indicates a weak dose-response relationship. From a developmental perspective, a mortality rate of less than 50% in adult mosquitoes at practical concentrations represents suboptimal potential. Adults have a thicker cuticle and more mature detoxification enzymes (eg, cytochrome P450 monooxygenase), which may reduce the availability of the extract compared to larvae. Therefore, the crude ethanol extract is not suitable as a standalone adulticide without further purification or formulation optimization.
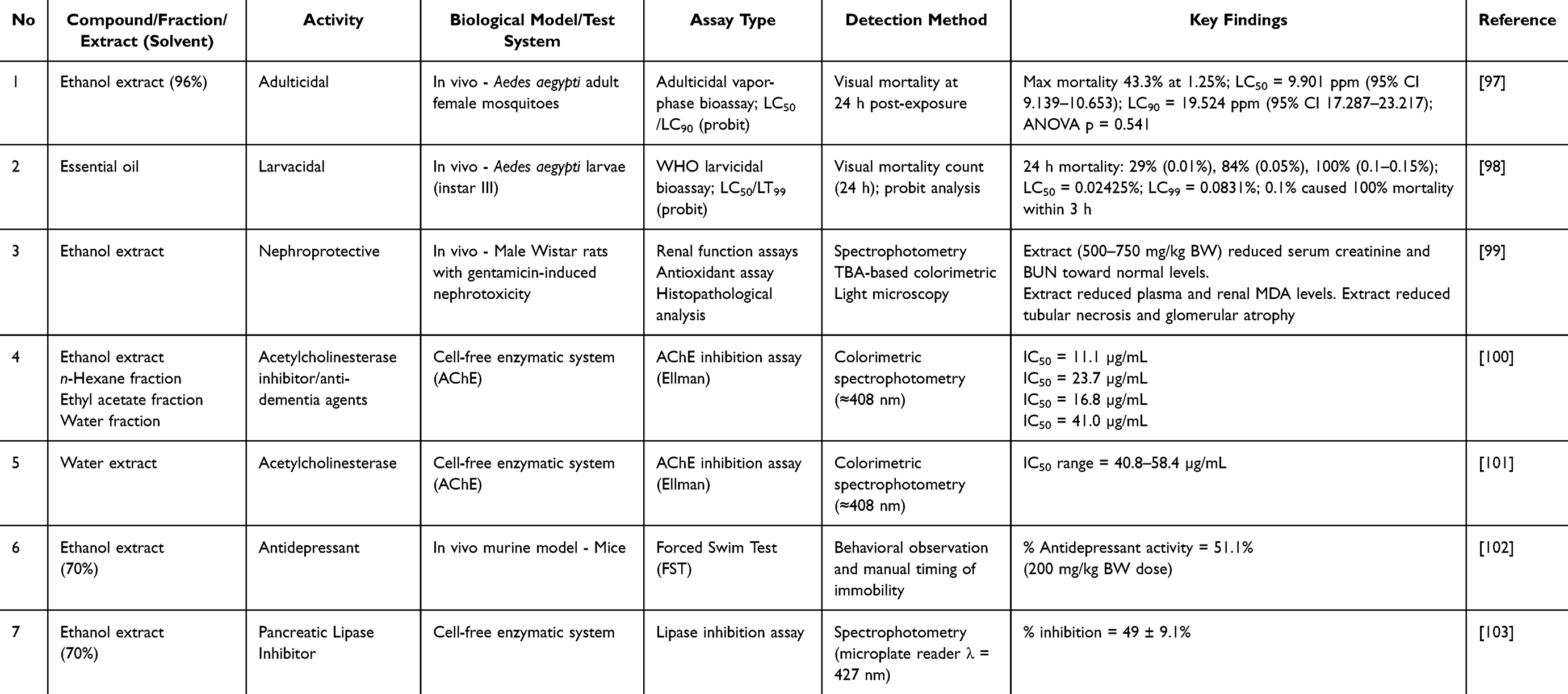 |
Table 11 Other Activities of Piper crocatum |
In contrast, P. crocatum essential oil exhibited significant larvicidal activity with 100% mortality at concentrations ≥0.1% within 3 h. The LC50 and LC99 values were 0.02425% and 0.0831%, respectively, suggesting promising larvicidal potential according to the World Health Organization standard test.98 The rapidly lethal effect points to possible neurotoxicity or disruption of membrane integrity, in line with other Piper species reported to contain essential oils rich in monoterpenoids and phenylpropanoids. These include acetylcholinesterase inhibition, octopamine receptor interference, mitochondrial membrane depolarization, and respiratory failure due to cuticle penetration. Unfortunately, no molecular or enzymatic assays have been performed to verify its mechanism of action.
Nephroprotective Activity
Gentamicin-induced nephrotoxicity involves the formation of reactive oxygen species and disruption of tubular epithelium. Treatment with ethanol extract (500–750 mg/kg body weight) improved renal function biomarkers (creatinine and blood urea nitrogen [BUN]), reduced MDA levels, and restored histological architecture.99 The reduction in lipid peroxidation indicates antioxidant-driven protection. However, no mechanistic investigations on Nrf2 activation, mitigation of mitochondrial dysfunction, or regulation of inflammatory cytokines have been conducted. The high effective dose poses translation-related considerations regarding bioavailability and determination of an equivalent dose in humans.
Acetylcholinesterase (AChE) Inhibition
P. crocatum also shows potential as an anti-dementia agent for the treatment of Alzheimer’s disease through AChE inhibition. Among the fractions tested, the ethanol extract exhibited the strongest AChE inhibition (IC50 = 11.1 µg/mL), approaching moderate potency when compared to known plant-derived cholinesterase inhibitors.100 Docking results (Table 12) indicate that flemiphilippinin A (−11.2 kcal/mol) and columbin (−9.2 kcal/mol) interact with the active site via sufficiently strong binding energies.101 However, these results indicate that the anti-dementia potential is still preliminary because of the absence of kinetic inhibition studies (competitive/non-competitive), the prediction of blood–brain barrier (BBB) penetration has not been experimentally verified, and the selectivity toward butyrylcholinesterase has not been evaluated.
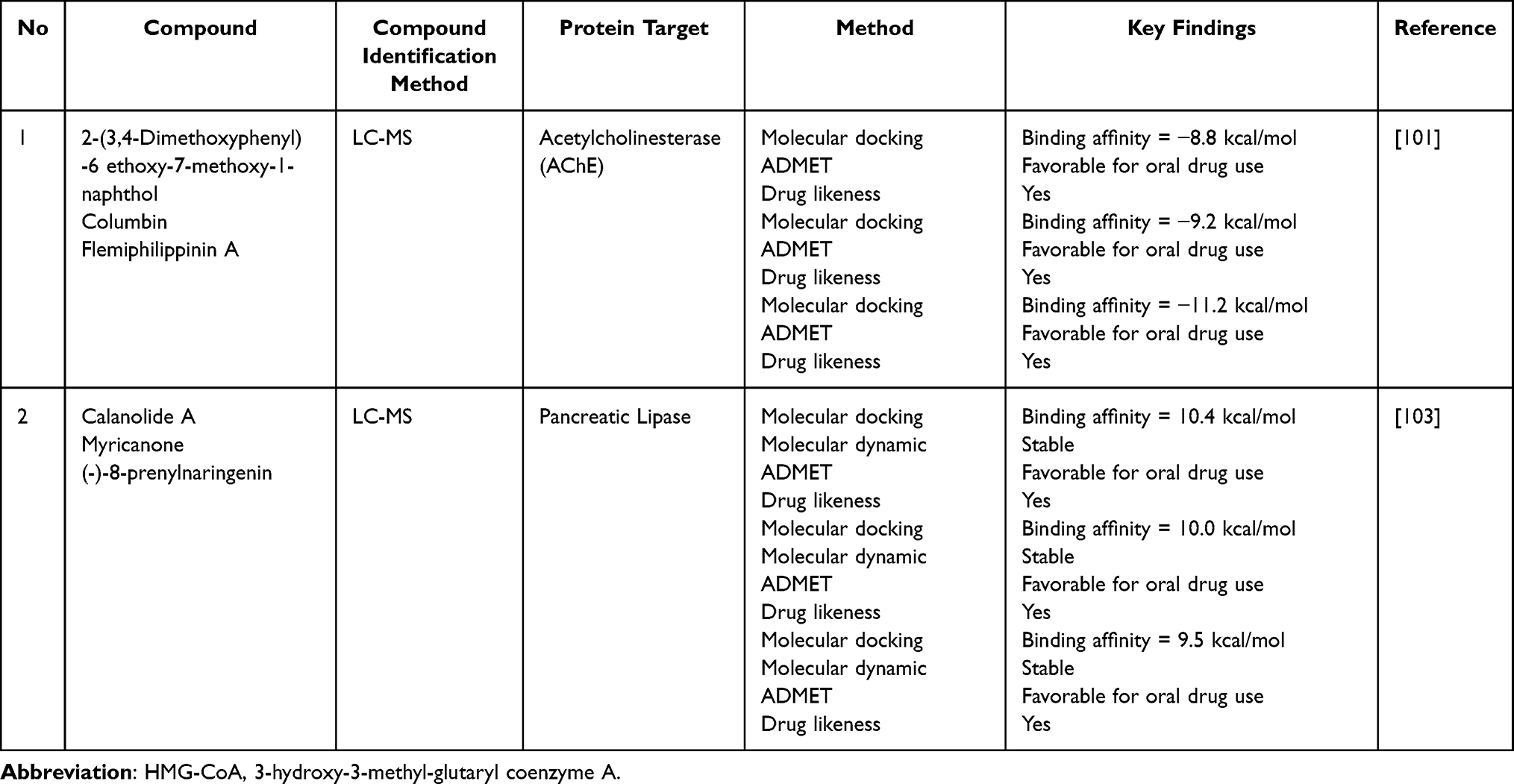 |
Table 12 In silico Study of P. crocatum Compounds and Protein-Related Other Activities |
Antidepressant Activity
In a study on antidepressant activity, the ethanol extract of P. crocatum reduced the immobility time by 51.1% in the forced swim test (FST), indicating central nervous system activity.102 However, mechanistic attribution remains speculative because of the lack of investigation into monoaminergic pathways; the absence of measurements of serotonin, dopamine, or brain-derived neurotrophic factor (BDNF); and the possibility that confounding effects from sedatives cannot be ruled out.
Anti-Obesity: Pancreatic Lipase Inhibition
A 70% ethanol extract of P. crocatum inhibited pancreatic lipase activity by 49 ± 9.1% in a cell-free enzymatic assay, implying moderate potential to block the digestion of dietary lipids.103 Pancreatic lipase is the main enzyme involved in the hydrolysis of triglycerides in the digestive tract and its inhibition is a well-established pharmacological strategy for reducing intestinal fat absorption. The reported inhibitory activity demonstrated that P. crocatum may promote lipid metabolism regulation and weight management. This mechanism is highly relevant in the context of metabolic disorders, where overconsumption and accumulation of lipids are closely related to insulin resistance caused by obesity and the development of type 2 diabetes mellitus. In line with these metabolic relationships, several studies have reported the antidiabetic effects of P. crocatum extract, including improvements in glycemic parameters and markers of oxidative stress. Therefore, the inhibition of pancreatic lipase may represent one of the complementary mechanisms through which P. crocatum mediates its metabolic regulatory effects, linking its anti-obesity potential to broader antidiabetic activity.
Systems-Level Therapeutic Integration of P. crocatum
The pharmacological activities of P. crocatum appear to be interconnected rather than isolated. Across antimicrobial, antioxidant, anti-inflammatory, immunomodulatory, metabolic, and hepatoprotective studies, several recurring pathways are evident. Multiple constituents appear to converge on shared biological processes, supporting the view that P. crocatum acts through a multitarget pharmacological network rather than through a single mechanism (Figure 10).
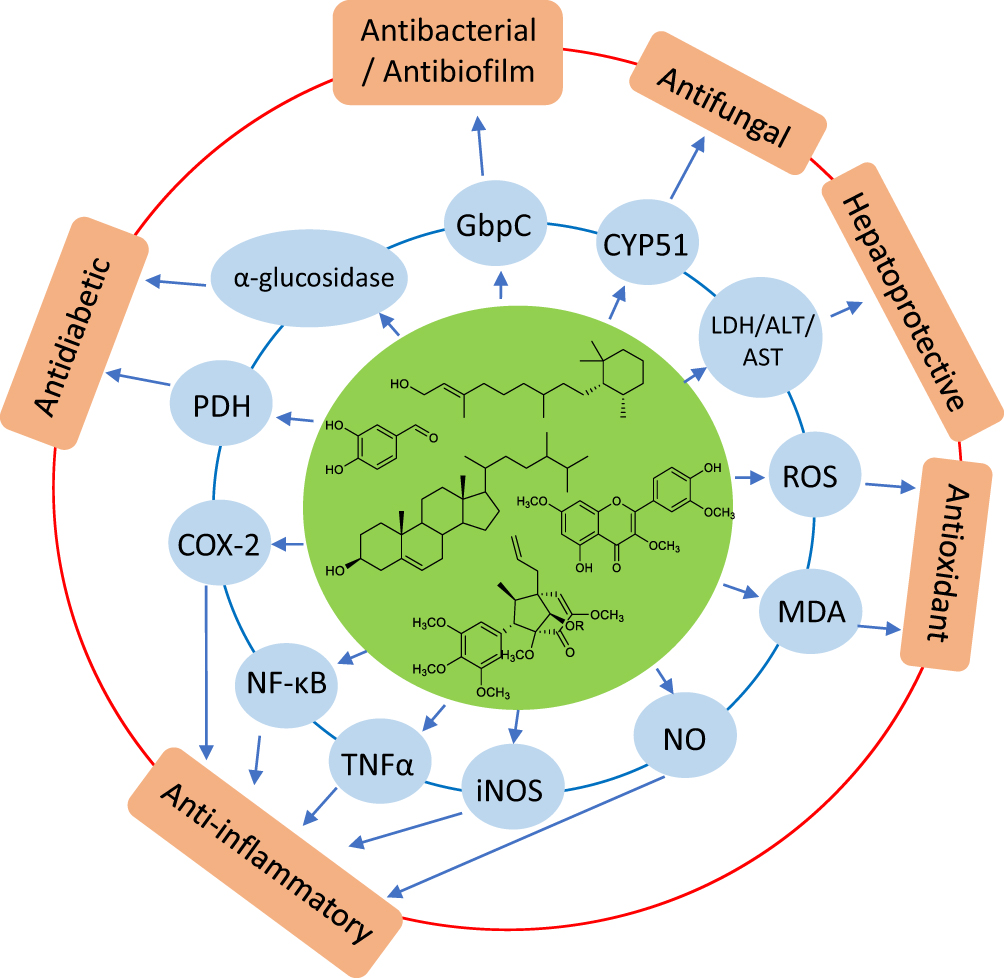 |
Figure 10 Multi-target pharmacological network of P. crocatum. |
Integrated evidence further indicated that the pharmacological effects of P. crocatum are organized around a defined set of shared molecular targets, as shown in Figure 10. Multiple bioactive constituents converge on key nodes, including microbial-associated proteins (GbpC), sterol biosynthesis enzymes (CYP51), inflammatory regulators (NF-κB, COX-2, iNOS), redox mediators (ROS, MDA, NO), metabolic enzymes (α-glucosidase, PDH), and hepatic biomarkers (LDH, ALT, AST). This convergence suggests that the observed biological activities arise from the coordinated modulation of interconnected pathways, thus providing a mechanistic basis for the system-level pharmacological profile of P. crocatum.
Infection–Inflammation Axis
One of the most consistent and mechanistically coherent domains is the infection–inflammation axis, which integrates antibacterial, antifungal, and antibiofilm activities with immunomodulatory and anti-inflammatory responses. Extracts and isolated compounds exhibit activity against a broad spectrum of pathogens and interfere with biofilm-associated targets, such as GbpC.40,54 Antifungal activity is primarily associated with disruption of ergosterol biosynthesis via CYP51 inhibition, leading to impaired membrane integrity.20,45 These antimicrobial mechanisms are functionally linked to host immune responses in which macrophage activation enhances NO-mediated pathogen clearance. However, the excessive activation of these pathways may lead to inflammatory damage. In this context, P. crocatum demonstrates the ability to suppress downstream inflammatory mediators, including TNF-α and NO, primarily through modulation of the NF-κB–iNOS signaling axis. This dual activity indicates that P. crocatum functions not only as an antimicrobial or anti-inflammatory agent but also as a regulator of infection-associated inflammation, maintaining a balance between host defense and tissue protection.
Redox–Inflammation Axis
Oxidative stress is a central node that is associated with multiple pharmacological effects. Antioxidant studies have demonstrated that extracts and flavonoids such as pachypodol effectively scavenge ROS and inhibit lipid peroxidation, as reflected by reduced MDA levels.29,67 Importantly, redox modulation is closely associated with inflammatory signaling. The observed suppression of ROS was accompanied by the downregulation of iNOS expression and decreased production of pro-inflammatory mediators, suggesting coordinated control of redox-sensitive pathways. Given that ROS contribute to NF-κB activation, its attenuation may indirectly limit the transcription of inflammatory cytokines such as TNF-α. Therefore, the antioxidant activity of P. crocatum extends beyond chemical radical scavenging and plays a functional role in regulating inflammation, reinforcing its role as a redox modulator of inflammation.
Metabolic Regulation Axis
The metabolic effects of P. crocatum integrate its antidiabetic and antiobesity activities into a unified framework. Inhibition of α-glucosidase suggests reduced intestinal glucose absorption, whereas modulation of PDH indicates potential regulation of mitochondrial energy metabolism. In parallel, inhibition of pancreatic lipase supports reduced lipid digestion and absorption, linking metabolic control to obesity-related pathways. These mechanisms are supported by in vivo evidence of improved glycemic regulation and metabolic parameters.88,90 Collectively, these findings suggest that P. crocatum modulates interconnected metabolic pathways rather than single enzymatic targets, which is consistent with a systems-level approach to metabolic diseases.
Hepatic Protection and Detoxification Axis
The hepatoprotective activity of P. crocatum is closely associated with modulation of oxidative stress and inflammatory responses. Experimental studies have demonstrated reductions in key liver damage biomarkers including LDH, ALT, and AST, accompanied by decreased ROS levels.94 These effects suggest attenuation of hepatocellular injury and improved cellular integrity. Although the molecular targets remain partially defined, the integration of redox regulation, inflammatory suppression, and metabolic modulation indicates that hepatoprotection arises from multipathway interactions rather than a single mechanism. This reinforces the position of P. crocatum as a system-level modulator of liver-associated disorders.
Conceptual Framework: Multitarget Homeostatic Regulation
Collectively, these interconnected axes support a unified conceptual model, in which P. crocatum functions as a regulator of physiological homeostasis. Rather than producing isolated pharmacological effects, its bioactive constituents modulate shared signaling pathways, particularly those involving NF-κB, ROS, metabolic enzymes, and membrane-associated targets. This system-level behavior reflects a form of context-dependent pharmacology, where immune activation is enhanced under infectious conditions, whereas excessive inflammation is suppressed in chronic states. This bidirectional modulation is consistent with the emerging drug discovery strategies that emphasize network regulation over single-target inhibition. However, despite this promising profile, its translation into clinical application remains limited by several factors, including high effective doses in vivo, lack of pharmacokinetic and bioavailability data, variability in the extract composition, and insufficient molecular target validation. Addressing these limitations is essential to advance P. crocatum from a pharmacological resource to a viable scaffold for drug development.
Lead Compound Prioritization
To facilitate the application of the research findings, the bioactive compounds identified from P. crocatum were systematically prioritized based on their pharmacological potential, mechanism of action, safety profile, and drug development suitability. This integrated assessment revealed varying levels of developmental relevance ranging from high-priority drug candidates to preliminary scaffolds (Table 13).
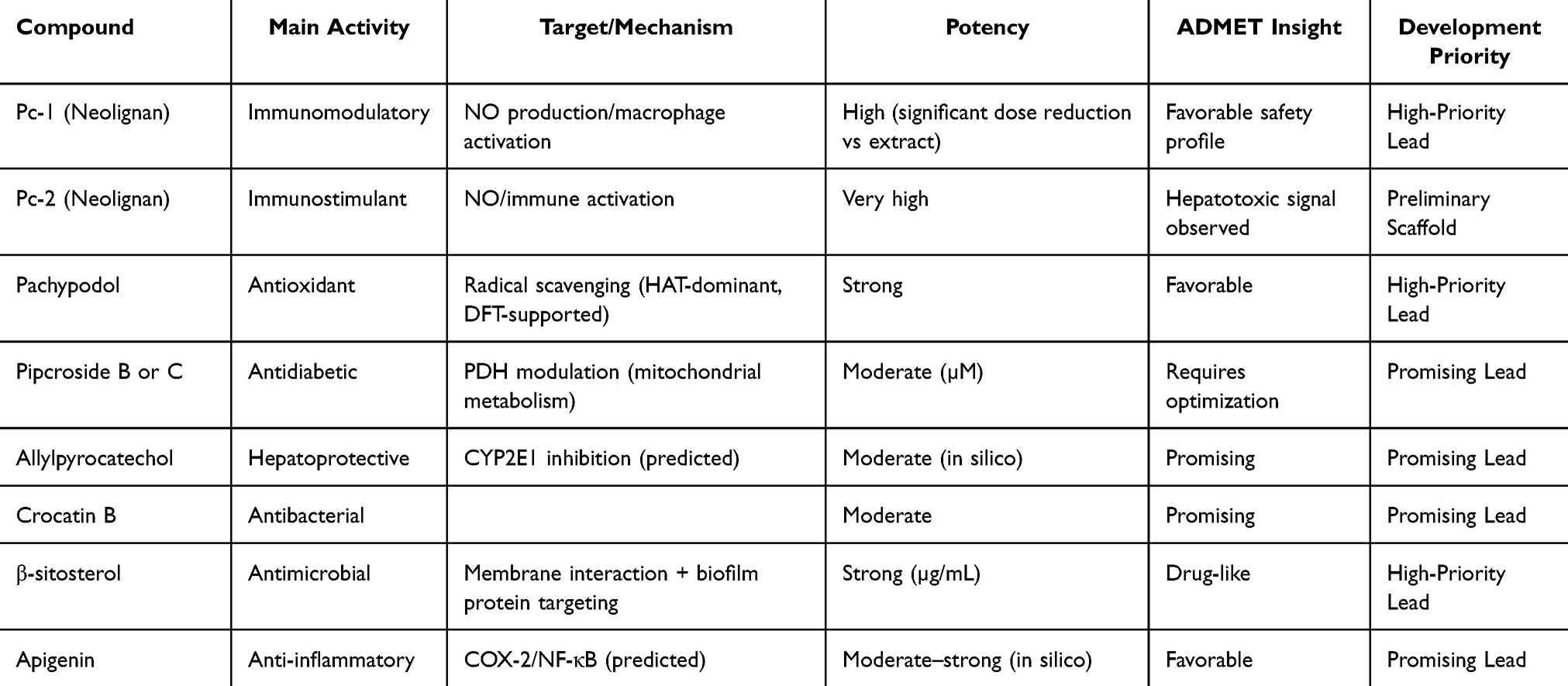 |
Table 13 Lead Compound Prioritization for Drug Development |
Among the evaluated compounds, Pc-1 (neolignan) and pachypodol have been identifed as the most promising high-priority candidates. Pc-1 demonstrates a good balance between immunomodulatory efficacy and safety, particularly when compared with Pc-2, which, despite its higher potency, exhibits signs of hepatotoxicity.28 This highlights a key principle in drug development: potency alone does not determine translational value. In parallel, pachypodol exhibited strong antioxidant activity, as supported by experimental tests and DFT analysis, which confirmed a thermodynamically favorable HAT mechanism. Its consistent activity across various redox-related assays, along with a favorable ADMET profile, further reinforces its potential as a redox-active pharmacological framework.29 β-Sitosterol also qualifies as a high-priority lead compound because of its relatively strong antimicrobial potential at the µg/mL level and its multitarget mode of action, which involves membrane interactions and disruption of biofilm-associated proteins.30,41,44 The reported drug-like characteristics of these prioritized compounds further support their suitability for drug development.
Compounds such as pipcroside B or C, allyl pyrocatechol, and apigenin have been identified as promising candidates. Pipcroside derivatives exhibit moderate activity in modulating pyruvate dehydrogenase, suggesting a role in mitochondrial glucose metabolism. However, their potency remains in the micromolar range and requires further optimization. Allylpyrocatechol showed potential hepatoprotective activity via predicted CYP2E1 interactions; however, this mechanism has not yet been experimentally validated. Similarly, apigenin exhibits strong binding affinity for inflammatory targets, such as COX-2 and NF-κB, in silico, supported by favorable ADMET properties and a relatively drug-like physicochemical profile. However, there has been no direct validation in a P. crocatum-specific model.
In contrast, Pc-2 is more appropriately classified as a modifiable scaffold rather than a direct lead candidate, despite its strong immunostimulatory potential, because of its hepatotoxic effects. This emphasizes the importance of safety profiles in the early-stage prioritization process and suggests that Pc-2 may serve as a structural template for further modification rather than as a direct development candidate.
Overall, this prioritization confirms that the therapeutic potential of P. crocatum lies not only in its chemical diversity but also in the varying translational potential of its constituents. The identification of high-priority compounds and modifiable scaffolds provides a rational basis for drug development. However, this prioritization also highlights an important gap: several structurally characterized metabolites remain poorly explored or have not yet been pharmacologically evaluated.
Underexplored Compounds and Research Gaps
Several compounds isolated from P. crocatum have not yet been studied in sufficient pharmacological detail. This gap is important because some of these metabolites share structural features with compounds that have already shown biological activity. For example, 4-allyl-2,6-dimethoxy-3-hydroxy-1-D-glucopyranoside (7) and erigeside II (8) are structurally related to syringin (3), as they contain a phenylpropanoid glycoside framework with hydroxylated and methoxylated aromatic groups. Erigeside II has been reported to inhibit soluble epoxide hydrolase, whereas compound 7 remains less explored. This structural relationship suggests that compound 7 may warrant further evaluation in assays related to inflammatory enzyme modulation and redox-associated pathways.
A similar consideration applies to the pipcroside group. Pipcroside A (20), pipcroside B (21), and pipcroside C (22) share structural similarities with nectamazin A (27), particularly in their lignan-derived glycosidic framework and methoxylated aromatic substituents. Nectamazin A has shown antibacterial, antifungal, and multitarget in silico activities against proteins involved in microbial growth and biofilm formation. Therefore, the pipcroside derivatives may also have related biological potential, although this assumption still requires direct experimental validation. The reported pyruvate dehydrogenase-modulating activity of pipcroside B and C further suggests that this subgroup may have pharmacological relevance beyond metabolic regulation.
These examples indicate that the pharmacological profile of P. crocatum remains incomplete. Several structurally identified but less-explored metabolites may contribute to the overall activity of this plant or provide additional scaffolds for lead optimization. Future studies should prioritize bioassay-guided evaluation of these compounds, followed by target-based assays, pharmacokinetic assessment, and structure–activity relationship analysis to clarify their relevance in drug discovery.
Limitations, Regulatory Considerations, and Future Directions
Despite the growing pharmacological evidence for P. crocatum, several limitations remain. Most studies are still based on crude extracts, fractions, or preliminary in vitro and in silico models. In vivo validation, pharmacokinetic characterization, and standardized toxicity evaluation are still limited. Differences in extraction methods, plant origin, dosage, and chemical standardization also make direct comparison among studies difficult. These limitations need to be addressed before P. crocatum-derived compounds can be considered reliable drug candidates.
From a regulatory perspective, further development of P. crocatum-based products requires plant authentication, raw material standardization, quality control of marker compounds, reproducible extraction procedures, and comprehensive safety evaluation. For isolated compounds intended for drug development, pharmacokinetic profiling, toxicological assessment, mechanism validation, and efficacy testing in relevant preclinical models are also required.
Although several metabolites identified from P. crocatum show pharmacological potential, their commercial development remains limited. Some widely distributed plant metabolites, including phytosterols and selected flavonoids, are already used as ingredients in nutraceutical or cosmetic products from other botanical sources. However, there is currently limited evidence that compounds specifically isolated from P. crocatum have been developed into standardized commercial drug products. This highlights the need for further formulation development and regulatory assessment before P. crocatum-derived metabolites can be translated into commercial therapeutic products.
Based on the available evidence, oral and local routes appear to be the most relevant routes of administration for P. crocatum-based preparations. Oral administration in the form of decoctions or extracts may be suitable for systemic applications, including metabolic, inflammatory, and hepatoprotective effects. In contrast, local dosage forms, such as mouthwash, gel, and root canal irrigant solutions, may be more appropriate for oral infections, periodontal conditions, and antibiofilm applications because they allow direct contact with the target site. For isolated compounds with limited solubility or oral bioavailability, formulation strategies such as nanoformulations, topical delivery systems, or prodrug approaches may be required. However, the selection of an appropriate dosage form should be supported by pharmacokinetic, toxicity, and target-site validation studies.
Future research should therefore move beyond descriptive bioactivity screening toward mechanism-driven studies, target validation, and pharmacokinetic evaluation. Several pathways repeatedly appear in the available studies, including the TLR4–NF-κB–iNOS pathway in inflammation, CYP2E1-mediated bioactivation in hepatotoxicity, and mitochondrial enzymes in metabolic regulation. These pathways provide a rational basis for focused validation using enzyme assays, gene expression analysis, and pathway-specific reporter systems.
From a medicinal chemistry perspective, several compounds identified in this review may serve as starting points for structural optimization. Neolignans such as Pc-1 show a favorable balance between efficacy and safety in immunomodulatory models, whereas pachypodol provides a redox-active scaffold with a clearer antioxidant mechanism. In contrast, Pc-2 highlights the need for structural modification to reduce toxicity despite its potent immunostimulatory activity. Strategies such as functional group modification, scaffold simplification, hybrid molecule development, prodrug design, nanoformulation, and delivery system modification may help improve selectivity, potency, bioavailability, and pharmacokinetic properties. However, comprehensive ADMET profiling and in vivo pharmacokinetic studies are still needed to connect in vitro activity with clinical feasibility.
The multitarget pharmacological profile of P. crocatum suggests that its therapeutic value may not rely on a single target. Instead, its constituents may act through multiple pathways that are relevant to complex conditions such as metabolic syndrome, chronic inflammation, and infection-related disorders. Future studies using systems biology approaches, multiparametric analysis, and tissue-level models may help clarify these interactions. Overall, development of P. crocatum as a source of drug candidates will require standardized materials, validated mechanisms, improved pharmacokinetic data, and careful safety evaluation.
Conclusion
Current scientific evidence highlights the potential of P. crocatum as a valuable source of bioactive compounds for drug discovery and therapeutic development. Phytochemical studies have identified a variety of secondary metabolites, including phenolics, neolignans, flavonoids, terpenoids, and phytosterols, several of which exhibit meaningful biological activities. Pharmacological studies further show that P. crocatum has antimicrobial, antibiofilm, antioxidant, anti-inflammatory, antidiabetic, hepatoprotective, anticancer, and immunomodulatory effects. These activities appear to involve the modulation of several molecular pathways, including oxidative stress regulation, inflammatory signaling, microbial membrane disruption, and inhibition of key metabolic enzymes. This multi-target pharmacological profile supports the traditional use of this plant and highlights its potential relevance for developing therapeutic agents targeting infectious diseases, metabolic disorders, and inflammatory conditions.
Despite these promising findings, several limitations remain. Most studies still rely on crude extracts evaluated at relatively high concentrations, whereas pharmacokinetic data, toxicity evaluation, and detailed mechanism-of-action studies are limited. Inconsistencies in extraction methods, experimental models, and compound standardization also complicate direct comparisons among studies. Future studies should prioritize bioassay-guided isolation, structural optimization of key compounds, molecular-level mechanism validation, and comprehensive pharmacokinetic evaluation. Overall, P. crocatum represents a chemically diverse and pharmacologically relevant natural source, but its development as a source of drug candidates will require interdisciplinary research integrating natural product chemistry, pharmacology, medicinal chemistry, and translational validation.
Acknowledgments
The authors are grateful to Universitas Padjadjaran for supporting this work through the Beasiswa Program Doktor Padjadjaran (BPDP) 2024 (grant number 1727/UN6.3.1/PT.00/2024), Penelitian Disertasi Doktor 2026 (grant number 1548/UN6.3.1/PT.00/2026), and Penelitian Fundamental Reguler (PFR) Grant (grant number 1549/UN6.3.1/PT.00/2025).
This publication charge is funded by Universitas Padjadjaran through the Indonesian Endowment Fund for Education (LPDP) on behalf of the Indonesian Ministry of Higher Education, Science, and Technology and managed under the EQUITY Program (Contract No. 4303/B3/DT.03.08/2025 and 3927/UN6. RKT/HK.07.00/2025).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Atanasov AG, Waltenberger B, Pferschy-Wenzig E-M, et al. Discovery and resupply of pharmacologically active plant-derived natural products: a review. Biotechnol Adv. 2015;33(8):1582–35. doi:10.1016/j.biotechadv.2015.08.001
2. Newman DJ, Cragg GM. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J Nat Prod. 2020;83(3):770–803. doi:10.1021/acs.jnatprod.9b01285
3. Yanti E. The effect of using red betel leaves (Piper crocatum) for vaginal discharge among fertile age women (FAW). Malays J Nurs. 2017;8(3):21–26.
4. Henri H, Sari DP, Hakim L. Medicinal plants for traditional treatment used by the Malays in South Bangka Regency, Indonesia. Biosaintifika. 2022;14(1):125–134. doi:10.15294/biosaintifika.v14i1.34455
5. Rosyadi A, Faizah RN, Nuri N, Puspitasari E. Anticancer properties of methanolic extract of Piper crocatum leaf using BST and cytotoxicity on HeLa cell lines. Ann Trop Med Public Health. 2020;23(3):3–11. doi:10.36295/ASRO.2020.2331
6. Yaturramadhan H, Futri CL. Formulation of natural handsanitizer from red betle (Piper crocatum) leaves and aloe vera (Aloe vera). J Public Health Pharm. 2021;1(2):43–45. doi:10.56338/jphp.v1i2.3718
7. Herryawan H. Effect of topical application red betel (Piper crocatum) leaf gel on clinical attachment level repair patient with chronic periodontitis. J Dentomaxillofac Sci. 2017;2(3):160–163.
8. Assidqi K, Sianipar N, Tarigan R. Bioactive compounds of Indonesian red betel (Piper crocatum) extract and its inhibitory activity in MCF-7 cell line. Rasayan J Chem. 2022;15(1):232–238. doi:10.31788/RJC.2022.1516468
9. Lister INE, Ginting CN, Girsang E, et al. Antioxidant properties of red betel (Piper crocatum) leaf extract and its compounds. J Nat Remedies. 2019;19(4):198–205. doi:10.18311/jnr/2019/23633
10. Fauziyah PN, Sukandar EY, Ayuningtyas DK. Combination effect of antituberculosis drugs and ethanolic extract of selected medicinal plants against multi-drug resistant Mycobacterium tuberculosis isolates. Sci Pharm. 2017;85(1):14. doi:10.3390/scipharm85010014
11. Yuliana T, Meliani D, Kurnia D. Potential antibacterial of leaf sirih merah against Enterococcus Faecalis ATCC 29212 bacteria. Comb Chem High Throughput Screen. 2025;28(18):3260–3272. doi:10.2174/0113862073344642241120041947
12. Hasibuan MS, Yasni S, Bintang M, Ranti AS. Antihyperglycemic activity of Piper crocatum leaves and Cinnamomum burmannii bark mixture extract in streptozotocin-induced diabetic rats. J Math Fund Sci. 2016;48(2):178–191. doi:10.5614/j.math.fund.sci.2016.48.2.8
13. Kamaruzaman SRS, Kasim KF, Jaafar MN. The effect of harvesting time on the antioxidant and antidiabetic activity of Piper crocatum (sirih merah) extract. IOP Conf Ser Mater Sci Eng. 2020;864(1):012211. doi:10.1088/1757-899X/864/1/012211
14. Lely N, Arifin H, Aldi Y, Sri Wahyuni F. Anti-inflammatory effects of methanol extract, hexane, ethyl acetate, and butanol fraction of Piper crocatum Ruiz & Pav leaves on lipopolysaccharide-induced RAW 264.7 cells. Pharmacogn J. 2021;13(6):1341–1346. doi:10.5530/pj.2021.13.169
15. Haddaway NR, Page MJ, Pritchard CC, McGuinness LA. PRISMA2020: an R package and Shiny app for producing PRISMA 2020-compliant flow diagrams, with interactivity for optimised digital transparency and Open Synthesis. Campbell Syst Rev. 2022;18(2):e1230. doi:10.1002/cl2.1230
16. Byrd A. Tropica Color Cyclopedia of Exotic Plants and Trees.
17. Chaveerach A, Mokkamul P, Sudmoon R, Tanee T. Ethnobotany of the genus Piper (Piperaceae) in Thailand. Ethnobot Res Appl. 2006;4:223–231. doi:10.17348/era.4.0.223-231
18. Puspita PJ, Safithri M, Sugiharti NP. Antibacterial activities of sirih merah (Piper crocatum) leaf extracts. Curr Biochem. 2018;5(3):1–10. doi:10.29244/cb.5.3.1-10
19. Li HX, Widowati W, Azis R, Yang SY, Kim YH, Li W. Chemical constituents of the Piper crocatum leaves and their chemotaxonomic significance. Biochem Syst Ecol. 2019;86:103905. doi:10.1016/j.bse.2019.05.013
20. Siswina T, Rustama MM, Sumiarsa D, Apriyanti E, Dohi H, Kurnia D. Antifungal constituents of Piper crocatum and their activities as ergosterol biosynthesis inhibitors discovered via in silico study using ADMET and drug-likeness analysis. Molecules. 2023;28(23):7705. doi:10.3390/molecules28237705
21. Hartini YS, Nugroho L. The accumulation of two neolignan in the leaves, stems, and flower of red betel (Piper crocatum Ruiz & Pav.). J Phys Conf Ser. 2017;835(1):012017. doi:10.1088/1742-6596/835/1/012017
22. Cui Q, Du R, Liu M, Rong L. Lignans and their derivatives from plants as antivirals. Molecules. 2020;25(1):183. doi:10.3390/molecules25010183
23. Pilkington LI. Lignans: a chemometric analysis. Molecules. 2018;23(7):1666. doi:10.3390/molecules23071666
24. Fernando A, Yuliandari R, Rullah K, et al. Cytotoxic activities of fractions and two isolated compounds from sirih merah (Indonesian red betel), Piper crocatum Ruiz & Pav. Procedia Chem. 2014;13:79–84. doi:10.1016/j.proche.2014.12.009
25. Li HX, Yang SY, Kim YH, Li W. Isolation of two new compounds and other constituents from leaves of Piper crocatum and study of their soluble epoxide hydrolase activities. Molecules. 2019;24(3):489. doi:10.3390/molecules24030489
26. Arbain D, Syafni N, Ismed F, Yousuf S, Choudhary MI. Bicyclo[3.2.1]octanoid neolignans from Indonesian red betle leaves (Piper crocatum Ruiz & Pav.). Phytochem Lett. 2018;24:163–166. doi:10.1016/j.phytol.2018.02.006
27. Chai YJ, Go Y, Zhou HQ, et al. Unusual bicyclo [3.2. 1] octanoid neolignans from leaves of Piper crocatum and their effect on pyruvate dehydrogenase activity. Plants. 2021;10(9):1855. doi:10.3390/plants10091855
28. Hartini YS, Widyarini S, Nugroho LH. The comparison of two neolignans isolated from red betel leaf and its extract against macrophage phagocytic activity, the level of AST, and histopathological features of the liver in mice. Orient Pharm Exp Med. 2018;18(4):325–333. doi:10.1007/s13596-018-0326-x
29. Apriyanti E, Al-Anshori J, Takaya A, Kai K, Rustaman, Kurnia D. Flavonoids from Piper crocatum Ruiz & Pav.: isolation, structural characterization, antioxidant properties, and density functional theory insights. Results Chem. 2026;23:103152. doi:10.1016/j.rechem.2026.103152
30. Tristyaningrum NA, Herlina T, Heliawati L, Nurdin D, Evangelina IA, Kurnia D. Cassipourol isolated from red betel (Piper crocatum): molecular docking and ADMET analysis against Candida albicans. Results Chem. 2025;18:102716. doi:10.1016/j.rechem.2025.102716
31. Dagne A, Degu S, Abebe A, Bisrat D. Antibacterial activity of a phenylpropanoid from the root extract of carduus leptacanthus fresen. J Trop Med. 2023;2023(1):4983608. doi:10.1155/2023/4983608
32. Merkl R, HRádkoVá I, FIlIp V, Šmidrkal J. Antimicrobial and antioxidant properties of phenolic acids alkyl esters. Czech J Food Sci. 2010;28(4):275–279. doi:10.17221/132/2010-CJFS
33. Syafni N, Putra DP, Arbain D. 3, 4-dihydroxybenzoic acid and 3, 4-dihydroxybenzaldehyde from the fern Trichomanes chinense L.; isolation, antimicrobial and antioxidant properties. Indones J Chem. 2012;12(3):273–278. doi:10.22146/ijc.21342
34. Maisuria VB, Hosseinidoust Z, Tufenkji N. Polyphenolic extract from maple syrup potentiates antibiotic susceptibility and reduces biofilm formation of pathogenic bacteria. Appl Environ Microbiol. 2015;81(11):3782–3792. doi:10.1128/AEM.00239-15
35. Friedman M, Henika PR, Mandrell RE. Antibacterial activities of phenolic benzaldehydes and benzoic acids against Campylobacter jejuni, Escherichia coli, Listeria monocytogenes, and Salmonella enterica. J Food Prot. 2003;66(10):1811–1821. doi:10.4315/0362-028X-66.10.1811
36. Heliawati L, Lestari S, Imani Y, Aminingsih T, Wulandari A, Dikdik K. Potential of neolignan from Piper crocatum as antimicrobial against pathogenic oral microbes and its prospects as an oral medication. Trends Sci. 2025;23:11608. doi:10.48048/tis.2026.11608
37. Salsabila DAN, Herlina T, Kurnia D. Nectamazin A from Piper crocatum as a bioactive compound against oral and throat diseases: in vitro and in silico study. Results Chem. 2026;22:103103. doi:10.1016/j.rechem.2026.103103
38. Adamczak A, Ożarowski M, Karpiński TM. Antibacterial activity of some flavonoids and organic acids widely distributed in plants. J Clin Med. 2019;9(1):109. doi:10.3390/jcm9010109
39. Elkhayat E. Cytotoxic and antibacterial constituents from the roots of Sonchus oleraceus L. growing in Egypt. Pharmacogn Mag. 2009;5(20):324–328. doi:10.4103/0973-1296.58154
40. Zajdel SM, Graikou K, Głowniak K, Chinou I. Chemical analysis of Penstemon campanulatus (Cav.) Willd.—Antimicrobial activities. Fitoterapia. 2012;83(2):373–376. doi:10.1016/j.fitote.2011.11.021
41. Anwar R, Sukmasari S, Aisyah S, Ilfani D. Antimicrobial activity of β-sitosterol isolated from Kalanchoe tomentosa leaves against Staphylococcus aureus and Klebsiella pneumonia. Pak J Biol Sci. 2022;25(7):602–607. doi:10.3923/pjbs.2022.602.607
42. Evangelina IA, Herdiyati Y, Laviana A, et al. Bio-mechanism inhibitory prediction of β-sitosterol from Kemangi (Ocimum basilicum L.) as an inhibitor of MurA enzyme of oral bacteria: in vitro and in silico study. Adv Appl Bioinform Chem. 2021;14:103–115. doi:10.2147/AABC.S301488
43. Luhata LP, Usuki T. Antibacterial activity of β-sitosterol isolated from the leaves of Odontonema strictum (Acanthaceae). Bioorg Med Chem Lett. 2021;48:128248. doi:10.1016/j.bmcl.2021.128248
44. Tao R, Wang CZ, Kong ZW. Antibacterial/antifungal activity and synergistic interactions between polyprenols and other lipids isolated from Ginkgo biloba L. leaves. Molecules. 2013;18(2):2166–2182. doi:10.3390/molecules18022166
45. Tristyaningrum NA, Herlina T, Kurnia D. β-Sitosterol from Piper crocatum: a dual-action antifungal and antibacterial agent for oral infections. Trends Sci. 2025;22(10):10420. doi:10.48048/tis.2025.10420
46. Lestari S, Kurnia D, Mayanti T, Heliawati L. Antimicrobial activities of stigmasterol from Piper crocatum: in vitro and in silico. J Chem. 2024;2024(1):2935516. doi:10.1155/2024/2935516
47. Rachmawaty F, Julianto T, Tamhid H. Isolation and identification of active compounds from ethanol extract Piper crocatum Leaves as antituberculosis candidate. Bangladesh J Med Sci. 2025;24:778–784. doi:10.3329/bjms.v24i3.82928
48. Kusuma S, Zuhrotun A, Meidina F. Antibacterial spectrum of ethanol extract of Indonesian red piper betel leaf (Piper crocatum Ruiz & Pav) against Staphylococcus species. Int J Pharma Sci Res. 2016;7(11):448–452.
49. Rachmawaty FJ, Julianto TS, Tamhid HA. Preliminary study on fractions’ activities of red betel vine (Piper crocatum Ruiz & Pav) leaves ethanol extract toward Mycobacterium tuberculosis. AIP Conf Proc. 2018;1954(1):030004.
50. Kusuma S, Tjitraresmi A, Susanti G. Antibacterial effect of red piper betle leaf (Piper crocatum Ruiz and Pav.) ethanolic extracts to Lactobacillus acidophilus and Lactobacillus bifidus growth inhibition. Asian J Pharm Clin Res. 2017;10(14):65–68. doi:10.22159/ajpcr.2017.v10s2.19490
51. Meliani D, Yuliana T, Kurnia D. Bioactive fraction of Piper crocatum as a potential antibacterial agent: LC-MS identification, docking study, molecular dynamics, and ADMET prediction. Curr Res Biotechnol. 2026;11:100364. doi:10.1016/j.crbiot.2025.100364
52. Kusuma S, Manan WS, Budiman F. Inhibitory effect of red piper betel leaf ethanol extract (Piper crocatum Ruiz and Pav.) against Trichomonas vaginalis trophozoites in vitro. Asian J Pharm Clin Res. 2017;10:311–314. doi:10.22159/ajpcr.2017.v10i11.20778
53. Meidarlina I, Damayanti L, Rikmasari R. Efficacy of red betel leaf (Piper crocatum) against Candida albicans over acrylic denture surface: an in vitro study. J Int Oral Health. 2021;13(3):281–287. doi:10.4103/jioh.jioh_359_20
54. Kusuma SAF, Hendriani R, Genta A. Antimicrobial spectrum of red Piper betel leaf extract (Piper crocatum Ruiz & Pav) as natural antiseptics against airborne pathogens. J Pharm Sci Res. 2017;9(5):583.
55. Hartini YS, Nugroho LH. Antibacterial effect of red betel (Piper crocatum) extract in combination with vancomycin against Staphylococcus aureus. Biodiversitas. 2020;21(7):3271–3274. doi:10.13057/biodiv/d210750
56. Pangabdian F, Soetanto S, Suardita K. The effective concentration of red betel leaf (Piper crocatum) infusion as root canal irrigant solution. Dent J. 2012;45(1):12–16. doi:10.20473/j.djmkg.v45.i1.p12-16
57. Rachman SN, Skania PC, Ayustina S. The effect red betel leaves (Piper crocatum) on vaginal discharge (Fluor albus) in teenagers. Science Midwifery. 2024;12(3):1254–1260.
58. Siregar R, Sondang S, Aritonang NJ. Effectiveness of red betel (Piper crocatum) leaf ethanol extract mouthwash in reducing dental plaque index. Asian J Pharm Res Dev. 2021;9(1):5–7. doi:10.22270/ajprd.v9i1.902
59. Rachman SN, Fathiyati F, Anwar A. The effect red betel leaves (Piper crocatum) on pregnancy gingivitis. Jur Penelit Pendidik IPA. 2024;10(11):8977–8984.
60. Mulyanti S, Karo MB, Sari M R. Effectiveness of red betel leaf (Piper crocatum) in reducing symptoms of Fluor albus in women of reproductive age. Siklus J Res Midwifery Politek Tegal. 2025;14(2):25–29. doi:10.30591/siklus.v14i02.7448
61. Alfarabi M, Bintang M, Suryani, Safithri M. The comparative ability of antioxidant activity of Piper crocatum in inhibiting fatty acid oxidation and free radical scavenging. HAYATI J Biosci. 2010;17(4):201–204. doi:10.4308/hjb.17.4.201
62. Fatmawaty AN, Fadhil N, Prasasty V, Prasasty VD. Potential in vitro and in vivo antioxidant activities from Piper crocatum and Persea americana leaf extracts. Biomed Pharmacol J. 2019;12(2):661–667. doi:10.13005/bpj/1686
63. Safithri M, Andrianto D, Arda AG, et al. The effect of red betel (Piper crocatum) water fraction as tyrosinase inhibitors: in vitro, molecular docking, and molecular dynamics studies. J King Saud Univ Sci. 2023;35(10):102933. doi:10.1016/j.jksus.2023.102933
64. Safithri M, Nur Faridah D, Ramadani F, Pratama R. Antioxidant activity of ethanol extract and fractions of Piper crocatum with Rancimat and cuprac methods. Turk J Biochem. 2022;47(6):795–801. doi:10.1515/tjb-2021-0300
65. Irawan C, Tambunan JA, Rachmy S, Putri ID. Phytochemical analysis of red betel (Piper crocatum Ruiz & Pav) stem extracts and its antioxidant and alpha-glucosidase inhibitory potentials. Trop J Nat Prod Res. 2024;8(4):7042–7048.
66. Lely N, Aldi Y, Wahyuni FS. Anti-inflammatory effect of isolate compounds from ethyl acetate fraction of Piper crocatum Ruiz and Pav leaves on lipopolysaccharide-induced RAW 264.7 cells. HAYATI J Biosci. 2023;30(1):28–34. doi:10.4308/hjb.30.1.28-34
67. Gong Y, Li HX, Guo RH, et al. Anti-allergic inflammatory components from the leaves of Piper crocatum Ruiz & Pav. Biol Pharm Bull. 2021;44(2):245–250. doi:10.1248/bpb.b20-00726
68. Maslikah SI, Wulandari N, Lestari SR, Arifah SN. The effects of red betel (Piper crocatum) leaf extract to decreasing profile population of CD4+ and CD8+ cells in rheumatoid arthritis-induced mice. IOP Conf Ser Earth Environ Sci. 2019;276(1):012044. doi:10.1088/1755-1315/276/1/012044
69. Oktavia NS, Mustika A, Hidayati AN. Mechanism of the effect of Piper crocatum extract on wound healing of Wistar rats post-excision mammary tumor based on IL-10 level, TGF-β1 expression, VEGF expression, Collagen density, and clinical features. Open Vet J. 2025;15(3):1264. doi:10.5455/OVJ.2025.v15.i3.18
70. Wahjuni S, Wita I, Astawa INM. Anti-inflammatory effect of red Piper crocatum leaves extract decrease TNF-α and IL-6 levels in wistar rat with atherosclerosis. Bali Med J. 2016;5(2):51–56. doi:10.15562/bmj.v5i2.210
71. Savitri L, Maslikah SI, Susilowati. Effect of red betel leaf extract (Piper crocatum) against interleukin-1 beta (IL-1β) levels and thickness of feet oedema in Mus musculus (Swiss strain) rheumatoid arthritis model. AIP Conf Proc. 2020;2231(1):040012.
72. Amalia A, Maslikah SI, Lestari SR. Virtual screening flavonoid compounds from red betel (Piper crocatum Ruiz & Pav.) as inhibitor of cyclooxygenase-2 (COX-2). AIP Conf Proc. 2020;2231(1):040043.
73. Maslikah SI, Amalia A, Afifah S. Red betel apigenin compound (Piper crocatum Ruiz & Pav.) as an anti-inflammatory rheumatoid arthritis agent through virtual screening. AIP Conf Proc. 2019;2120(1):080003.
74. Djati MS, Fatchiyah F, Widyananda MH, Rifa`i M, Warsito W, Djati MS. Molecular docking and dynamic simulation of sesamin from Piper crocatum Ruiz & Pav as an inhibitor of canonical NF-κB: potential anti-inflammatory mechanism. J Pharm Pharmacogn Res. 2025;13(3):848–856. doi:10.56499/jppres24.2070_13.3.848
75. Maslikah S, Lestari S, Handayani N, et al. The anti-inflammatory potential of red betel (Piper crocatum) leaves through inhibitory mechanism on Nfκb signaling pathway: drug-like candidate study. Nat Life Sci Commun. 2023;22(1):e2023005.
76. Maslikah SI, Putra WE. Molecular dynamics simulation of various bioactive compounds of red betel (Piper crocatum) as anti-inflammatory drug-like candidates in rheumatoid arthritis treatment. Trends Sci. 2024;21(3):7334. doi:10.48048/tis.2024.7334
77. Afifah S, Lukiati B, Maslikah SI. The potentials of red betel (Piper crocatum Ruiz & Pav) terpenoid compounds as microsomal Prostaglandin E Synthase-1 (mPGES-1) enzyme inhibitor of rheumatoid arthritis through virtual screening. AIP Conf Proc. 2020;2231(1):040023.
78. Anugrahwati M, Purwaningsih T, Manggalarini J, Alnavis N, Wulandari D, Pranowo H. Extraction of ethanolic extract of red betel leaves and its cytotoxicity test on HeLa cells. Procedia Eng. 2016;148:1402–1407. doi:10.1016/j.proeng.2016.06.569
79. Zulharini M, Sutejo IR, Fadliyah H, Jenie RI. Methanolic extract of red betel leaves (Piper crocatum Ruiz & Pav) perform cytotoxic effect and antimigration activity toward metastatic breast cancer. Indones J Cancer Chemoprevent. 2017;8(3):94–100. doi:10.14499/indonesianjcanchemoprev8iss3pp94-100
80. Wicaksono BD, Handoko YA, Arung ET, et al. Antiproliferative effect of the methanol extract of Piper crocatum Ruiz & Pav leaves on human breast (T47D) cells in-vitro. Trop J Pharm Res. 2009;8(4):345–352.
81. Abkar AH, Djati MS, Widodo W. In silico study to predict the potential of beta asarone, methyl piperonylketone, coumaric acid in Piper crocatum as anticancer agents. J Exp Life Sci. 2021;11(3):89–99. doi:10.21776/ub.jels.2021.011.03.04
82. Abdullah W, Seno DSH, Safithri M. Total phenolics, flavonoids, and α-glucosidase inhibitory activity of red betel (Piper crocatum) extract in various solvents. Indones J Appl Res. 2024;5(2):112–123.
83. Hartini YS, Setyaningsih D. The potency of red betel (Piper crocatum Ruiz & Pav.) methanolic extract as α-amylase and α-glucosidase inhibitor. Adv Biol Sci Res. 2022;22:209–212.
84. Damanik EB, Manafe DR, Kareri DG. The effect of red betel leaf (Piper crocatum) ethanol extract on the histopathological eye image of alloxan-induced diabetic rat (Rattus norvegicus). Trop J Nat Prod Res. 2024;8(4):6771–6774.
85. Johari A, Mailuddillahi V, Naswir MM. The potential of red betel leaf as a hypoglycemic agent in alloxan-induced mice. Open Access Res J Biol Pharm. 2025;13(2):001–006. doi:10.53022/oarjbp.2025.13.2.0023
86. Navirius FJ, Pamudji G, Herowati R. Effect of red betel leaf extract (Piper crocatum) on blood glucose levels and kidney histopathology in streptozotosin-nicotinamide induced streptozotosin-nicotinamide rats. J La Medihealtico. 2023;4(3):144–156. doi:10.37899/journallamedihealtico.v4i3.896
87. Solfiane R, Rahmadhani D. Effect of Piper crocatum extract on kidney histopathology in alloxan-induced wistar white rats (Rattus norvegicus). Vet Biomed Clin J. 2024;6(1):8–15. doi:10.21776/ub.VetBioClinJ.2024.006.01.2
88. Sudiana IK, Purwanto B. The effect of red betel leaves (Piper crocatum Ruiz and Pav.) on blood sugar and insulin expression levels in rat (Rattus norvegicus domesticus) models of diabetes mellitus. Drug Invention Today. 2019;11(3):553–557.
89. Pane HW, Suryantara B, Kristiarini JJ. Comparison of red betel leaf decoction and Aloe vera decoction on decreased sugar levels in patients with diabetes mellitus at Setia Janji Health Center. Contagion. 2023;5(2):637–651. doi:10.30829/contagion.v5i2.15007
90. Wahyuningsih S, Noviana RN, Sholihah PSD, Vikasari SN. Antihyperuricemia effect of Piper crocatum Ruiz & Pav leave ethanol extract in male wistar rats. IOP Conf Ser Earth Environ Sci. 2022;1104(1):012023. doi:10.1088/1755-1315/1104/1/012023
91. Hermanto F, Wahyuningsih S. In vivo uricosuric effect of red betel leaves (Piper crocatum Ruiz & Pav) ethanol extract. J Sains Kes. 2024;6(2):269–273. doi:10.25026/jsk.v6i2.2135
92. Lister I, Ginting CN, Girsang E, et al. Piper crocatum ameliorates APAP-induced hepatotoxicity through antioxidant and anti-inflammatory mechanisms. J Math Fundam Sci. 2022;54(1):109–120. doi:10.5614/j.math.fund.sci.2022.54.1.6
93. Ginting CN, Lister INE, Girsang E, et al. Hepatotoxicity prevention in Acetaminophen-induced HepG2 cells by red betel (Piper crocatum Ruiz and Pav) extract from Indonesia via antioxidant, anti-inflammatory, and anti-necrotic. Heliyon. 2021;7(1):e05620. doi:10.1016/j.heliyon.2020.e05620
94. Novilia L, Harahap U, Hsb PAZ. Effect of red betel (Piper crocatum Ruiz and Pav.) ethanol extract against carbon tetrachloride induce hepatic injury in rats. Asian J Pharm Res Dev. 2017;5(5):1–8.
95. Novilia L, Harahap U, Hasibuan PAZ. Evaluation of hepatoprotective effect of ethanolic extract from red betel (Piper crocatum Ruiz and Pav.) leaves. Asian J Pharm Clin Res. 2018;11(7):248. doi:10.22159/ajpcr.2018.v11i7.25542
96. Lister I, Ginting CN, Girsang E, Widowati W, Sholihah IA. In silico approach for hepatoprotective activity of Piper crocatum leaf toward cytochrome P450 2E1 protein. Thai J Pharm Sci. 2021;45(4):273–276. doi:10.56808/3027-7922.2502
97. Aziz A, Rulen BN, Fitria E. Test of the extract effectiveness of red Betel Leaf (Piper crocatum Ruiz and PAV) as electricmate method (Plate) to Aedes aegypti mosquito. Ecol Environ Conserv Pap. 2021;27:S80–87.
98. Setiawan B, Zarqya I, Putro S, Khasanah F. The effect of red betel leaf’s essential oil (Piper crocatum Ruiz & Pav.) against third Instar Aedes aegypti larvae. Pak J Med Health Sci. 2019;13(4):1162–1165.
99. Dewi IP, Fajrin FA, Yumna NK, Rahmawati HN, Christanty FM, Holidah D. Protective effects of Piper crocatum ethanol extract on Gentamicin-induced nephrotoxicity in rats. Pharm Educ. 2023;23(4):32–36. doi:10.46542/pe.2023.234.3236
100. Nurinsani EYY, Andrianto D, Safithri M. Acetylcholinesterase inhibition activity and phytochemical screening of red betel leaf (Piper crocatum Ruiz & Pav) as anti-dementia agents. BIO Web Conf. 2024;123:02009. doi:10.1051/bioconf/202412302009
101. Safithri M, Koendhori EB, Andrianto D, et al. Analysis of bioactive compounds Piper crocatum as inhibitors of acetylcholinesterase in silico and in vitro. Trends Sci. 2025;22(4):9437. doi:10.48048/tis.2025.9437
102. Sentat T, Apriliana A, Sari SLR, et al. Antidepressant activity test of red betel leaf ethanol extract (Piper crocatum) on male white mice (Mus musculus) using the forced swim test method. Al Ulum J Sains Teknol. 2025;11(2):47–52. doi:10.31602/jst.v11i2.18542
103. Gholam GM, Andrianto D, Septaningsih DA, Safithri M. Computational and in vitro investigation of Piper crocatum bioactive compounds as pancreatic lipase inhibitors. Karbala Int J Mod Sci. 2025;11(3):17. doi:10.33640/2405-609X.3423
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.