Back to Journals » International Journal of Nanomedicine » Volume 21
From Biomedical Mechanisms to Clinical Applications: Research Progress in Plant-Derived Vesicles for Cancer Therapy
Authors Xu JY, Xiao YL ![]() , Yu ZL
, Yu ZL ![]()
Received 9 October 2025
Accepted for publication 25 December 2025
Published 9 January 2026 Volume 2026:21 573022
DOI https://doi.org/10.2147/IJN.S573022
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sachin Mali
Jing-Yi Xu,1 Yi-Lei Xiao,2 Zi-Li Yu1,3
1State Key Laboratory of Oral & Maxillofacial Reconstruction and Regeneration, Key Laboratory of Oral Biomedicine Ministry of Education, Hubei Key Laboratory of Stomatology, School & Hospital of Stomatology Wuhan University, Wuhan, Hubei, People’s Republic of China; 2Department of Neurosurgery, Shandong First Medical University Affiliated Liaocheng People’s Hospital, Liaocheng, Shandong, People’s Republic of China; 3Department of Oral and Maxillofacial Surgery, School & Hospital of Stomatology Wuhan University, Wuhan, Hubei, People’s Republic of China
Correspondence: Yi-Lei Xiao, Email [email protected] Zi-Li Yu, Email [email protected]
Abstract: Extracellular vesicles (EVs) are nanoscale membranous structures secreted by cells, which carry bioactive molecules (eg, lipids, proteins, miRNAs) and facilitate intercellular communication. Recently, EVs have emerged as natural drug delivery systems. While early research focused on mammalian or bacterial EVs, concerns regarding safety, ethics, and cost limit their clinical translation. Plant-derived vesicles (PDVs), isolated from fruits, vegetables, or medicinal herbs, overcome these issues due to their abundant sources, cost-effectiveness, and favorable safety profile. PDVs from plants like citrus, ginger, and ginseng exhibit inherent anticancer effects by inhibiting proliferation and inducing apoptosis. However, PDV research remains nascent and faces major challenges: (1) Scalable production is inefficient, with current isolation methods yielding impurities and batch variations. (2) Unified markers and classification criteria are lacking, hindering data standardization. (3) High heterogeneity and the absence of systematic databases impede matching PDV sources to specific diseases. (4) Safety assessment frameworks are urgently needed, including contraindications and pharmacokinetic studies. This review summarizes the preparation methods, physicochemical properties, anticancer mechanisms, and drug delivery applications of PDVs, while addressing these challenges and future prospects.
Keywords: extracellular vesicles, plant-derived vesicles, nanodrug delivery system, cancer therapy
Background
EVs are nanoscale membranous structures secreted by cells. They are ubiquitously present in various bodily fluids, including cerebrospinal fluid, blood, saliva, and urine. EVs carry a rich cargo of bioactive molecules, such as proteins, miRNAs, and metabolites, derived from their parent cells. These EVs can deliver these biomolecules to recipient cells, thereby modulating their physiological functions. Consequently, EVs play pivotal roles in a wide array of pathological and physiological processes, including immune response, antigen presentation, cell migration, cell differentiation, and tumor invasion.1–3 Since their initial description and study in 1964,4 the secretory mechanisms, morphology, and functions of mammalian-derived EVs (MDEVs) have been progressively elucidated. Their potential value in disease diagnosis and therapy has also been gradually uncovered.5–8 Early research primarily focused on MDEVs,9 while the study of PDVs remained in its infancy. Although multivesicular bodies (MVBs) were observed in carrots by Halperin et al as early as 1967,10 it was not until 2009 that Regente et al successfully isolated PDVs from the extracellular fluid of sunflower seeds.11 Subsequently, PDVs have been isolated from a variety of edible plants by different research groups, including ginseng, ginger, broccoli, bitter melon, oat, grapefruit, lemon, blueberry, orange, sunflower seeds, and Arabidopsis thaliana.12 Over the past decade, research on PDVs has advanced significantly, with initial efforts concentrating on elucidating their fundamental biological characteristics and functions.
Owing to their inherent role in information transfer, EVs are regarded as natural drug delivery vehicles, and substantial research has validated their advantages for this application. However, EVs derived from mammalian cells or bacteria raise concerns regarding safety, ethics, and cost-effectiveness during clinical translation.
Malignant tumors represent a global public health challenge, accounting for millions of deaths annually.13 According to the latest data from the Global Cancer Observatory, there were approximately 19.3 million new cancer cases and nearly 10 million cancer-related deaths worldwide in 2020.14 Chemotherapy remains a cornerstone of cancer treatment and has played a pivotal role in the fight against tumors.15 It functions by inhibiting the proliferation of cancer cells and inducing apoptosis, thereby effectively reducing tumor volume and suppressing metastasis. However, Traditional chemotherapy faces fundamental challenges in clinical application due to its lack of tumor-specific recognition capability. This non-selective distribution leads to severe systemic toxicity toward normal tissues, while the inability to effectively penetrate tumor tissue barriers results in insufficient drug concentrations at lesion sites. More critically, long-term medication tends to induce multidrug resistance, significantly diminishing therapeutic efficacy.16 Although synthetic delivery systems such as liposomes and polymer nanoparticles have been developed to overcome these limitations, they present new technical challenges in clinical translation. Current Nano-drug delivery systems primarily rely on the Enhanced Permeability and Retention (EPR) effect for passive targeting, but this approach exhibits substantial individual variability and tumor heterogeneity.17 Active targeting strategies,18–20 meanwhile, face practical limitations including low modification efficiency and stability issues. Stability concerns remain prominent, with liposomes prone to drug leakage and phospholipid oxidation, while polymer nanoparticles face potential biocompatibility limitations of carrier materials; both systems are susceptible to rapid clearance by the mononuclear phagocyte system. Most existing nanomedicine delivery systems are limited to intravenous administration, with oral bioavailability typically being low,21 severely impacting patient convenience and long-term treatment compliance. From an industrial perspective, synthetic nanocarriers also encounter challenges in batch-to-batch consistency, complex production processes, and difficulties in ensuring sterility, leading to high production costs and quality control issues. Consequently, there is a pressing need within cancer research to develop highly efficient and targeted delivery systems for chemotherapeutic drugs. PDVs exhibit unique biological properties that make them promising drug delivery platforms.13 Their core advantages manifest in five key aspects: exceptional biocompatibility, with natural origins from edible plants conferring significantly lower immunogenicity than mammalian exosomes;14 natural nanocarrier structure, featuring lipid bilayers capable of co-loading hydrophilic and hydrophobic drug molecules; scalable production potential, where plant cultivation convenience and simplified extraction processes substantially reduce costs;15–18 batch-to-batch consistency, with plant-based sources ensuring stability absent the individual variations seen in mammalian cell culture; and dual therapeutic mechanisms, functioning both as drug carriers and synergistic therapeutic agents through their inherent active components. These characteristics endow PDVs with distinct advantages in safety, cost-effectiveness, and treatment efficacy, offering new directions for drug delivery system development. These properties enable PDVs to serve as safe and ethically non-contentious delivery vehicles, achieving targeted delivery of exogenous drug molecules19 and enhancing therapeutic outcomes through synergistic effects.20 Therefore, PDVs are considered a promising nanomedical tool,13 demonstrating potential efficacy in treating various diseases, including malignant tumors.5,21–25 Moreover, evidence suggests that vesicles derived from medicinal herbs inherently possess therapeutic effects analogous to those of the herbs themselves. Thus, herbal medicine-derived vesicles also present an opportunity to modernize traditional Chinese medicine (TCM).22 However, despite this bright outlook, research into PDVs for drug delivery and therapeutic applications continues to confront several formidable challenges. These include the lack of technologies and equipment for large-scale automated production of PDVs, the absence of unified classification criteria and marker identification standards for PDVs, and the uncertainty regarding the optimal PDV source for specific diseases.
A systematic approach was implemented to identify relevant studies on PDVs in cancer therapy. Comprehensive searches were conducted across seven electronic databases (PubMed, Web of Science, Scopus, Embase, Cochrane Library, CNKI, and Wanfang Data) using structured search syntax combining controlled vocabulary and free-text terms. The search strategy incorporated key concepts including: (“plant-derived extracellular vesicles” OR “plant exosomes” OR “PDEVs”) AND (“cancer” OR “tumor”), supplemented with specific plant names (eg, “grapefruit”, “ginger”, “ginseng”, “bitter melon”). The search timeframe spanned January 2010 to November 2025, covering publications in English and Chinese.
In summary, this review comprehensively summarizes recent advances in PDV research and their applications in cancer therapy, highlighting existing challenges and future research directions to facilitate their further development toward clinical translation.
Overview of PDVs
Biogenesis of PDVs
PDVs are nanoscale membranous structures secreted by plant cells. Analogous to vesicles secreted by mammalian cells, PDVs exhibit high heterogeneity. Their size and composition are influenced by the source of the plant cells, their state (eg, fresh or dried plant material), and environmental conditions.23 The precise mechanisms underlying PDV biogenesis are not yet fully elucidated, with three primary pathways currently proposed (Figure 1).
 |
Figure 1 Biogenesis of PDVs. (A) MVB pathway. (B) Vacuolar pathway. (C) EXPO pathway. |
First, and most commonly described, is the multivesicular body (MVB) pathway. In this pathway, MVBs fuse with the plasma membrane, releasing their intraluminal vesicles into the extracellular space.24 This mechanism was initially observed via transmission electron microscopy in carrots in the 1960s,10 and similar phenomena have been reported by other researchers in Arabidopsis thaliana infected with Turnip mosaic virus or the barley powdery mildew fungus Blumeria graminis f. sp. hordei.25–27 Furthermore, studies indicate that plant-microbe symbiosis can facilitate MVB fusion with the plasma membrane, leading to the release of PDVs into the periarbuscular space between plants and fungi.28 Second is the EXPO (exocyst-positive organelle) pathway. The EXPO pathway mediates vesicle fusion with the plasma membrane via the exocyst protein complex on EXPOs. This plant-specific unconventional secretory pathway was first identified in 2010 in suspension-cultured Arabidopsis and tobacco BY-2 cells.29 PDVs generated via this pathway typically range from 200 to 500 nm in diameter.30 Third is the vacuolar pathway, which was proposed in 2009. Research demonstrates that in Arabidopsis infected with Pseudomonas syringae, vacuoles can fuse with the plasma membrane to release vacuolar proteins extracellularly, representing a plant defense mechanism against intercellular bacterial pathogens.31 It is important to note that these three pathways are not mutually exclusive, which may work in concert for PDV secretion.28 Consequently, the exact biogenesis mechanisms of PDVs remain incompletely defined and constitute an active area of research focus within the PDV field.
Classification and Nomenclature of PDVs
PDVs can be classified according to various criteria, with each subclass exhibiting significant differences in protein markers, vesicle size, and potential functions.32 Based on botanical origin, PDVs can be categorized as derived from fruits, vegetables, or medicinal plants. Classification by biogenesis process yields MVB-derived, EXPO-derived, and vacuolar-derived PDVs. According to biochemical composition and size, PDVs are often classified into exosomes (50–150 nm), microvesicles (150–1000 nm), and apoptotic bodies (1000–5000 nm).33 Furthermore, based on the presence of artificial modification, PDVs are divided into natural PDVs and engineered PDVs.
In addition to the four aforementioned classification systems, the most commonly used approach in current research is based on the extraction method, categorizing PDVs into plant extracellular vesicles (PEVs), plant extracellular vesicle-like nanovesicles (PELNs), and plant lipid-derived nanovesicles (PLNVs). PEVs are typically isolated via tissue infiltration from the plant extracellular environment, such as the leaf apoplast or root exudates. PELNs, also referred to as plant vesicle-like particles (PVLNs), are obtained from disrupted plant tissue (eg, freshly squeezed juice) using high-speed blenders. These particles may encompass intracellular vesicles and vesicles formed by the fusion of damaged cell membranes.34 The lipid fraction further extracted from PELNs is termed PLNVs.24
Different subclasses of PDVs have distinct emphases in scientific research and clinical applications. The biogenesis and secretion pathways of PEVs are relatively well-defined, making them suitable for investigating plant physiology and pathology, such as interkingdom communication and regulation between plants and animals, or microorganisms. In contrast, PELNs/PVLNs are easier to obtain, exhibit higher yield, and are readily characterized and absorbed biologically, rendering them more appropriate for use as biotherapeutic agents and drug delivery carriers.35 Consequently, this review focuses primarily on PELNs/PVLNs and their applications when discussing the use of PDVs in cancer therapy.
Methods for Isolation and Purification of PDVs
Research indicates that PDVs can be isolated from various parts of plants, including fruit juice, pulp, roots, seeds, and dried plant material.5 Consequently, PDVs offer a more diverse source compared to MDEVs. However, the presence of a rigid cell wall in plant cells presents a challenge for studying PDV biogenesis. Furthermore, PDVs tend to adhere to macromolecules such as starch, cellulose, and tannins, complicating the large-scale production of high-purity PDVs.9 Despite their considerable potential, the clinical translation of PDVs is hindered by the lack of standardized isolation and analytical methodologies. Current extraction and purification techniques for PDVs are primarily adapted from existing methods used for MDEVs, relying on differences in size, density, and surface composition for separation (Figure 2 and Table 1).
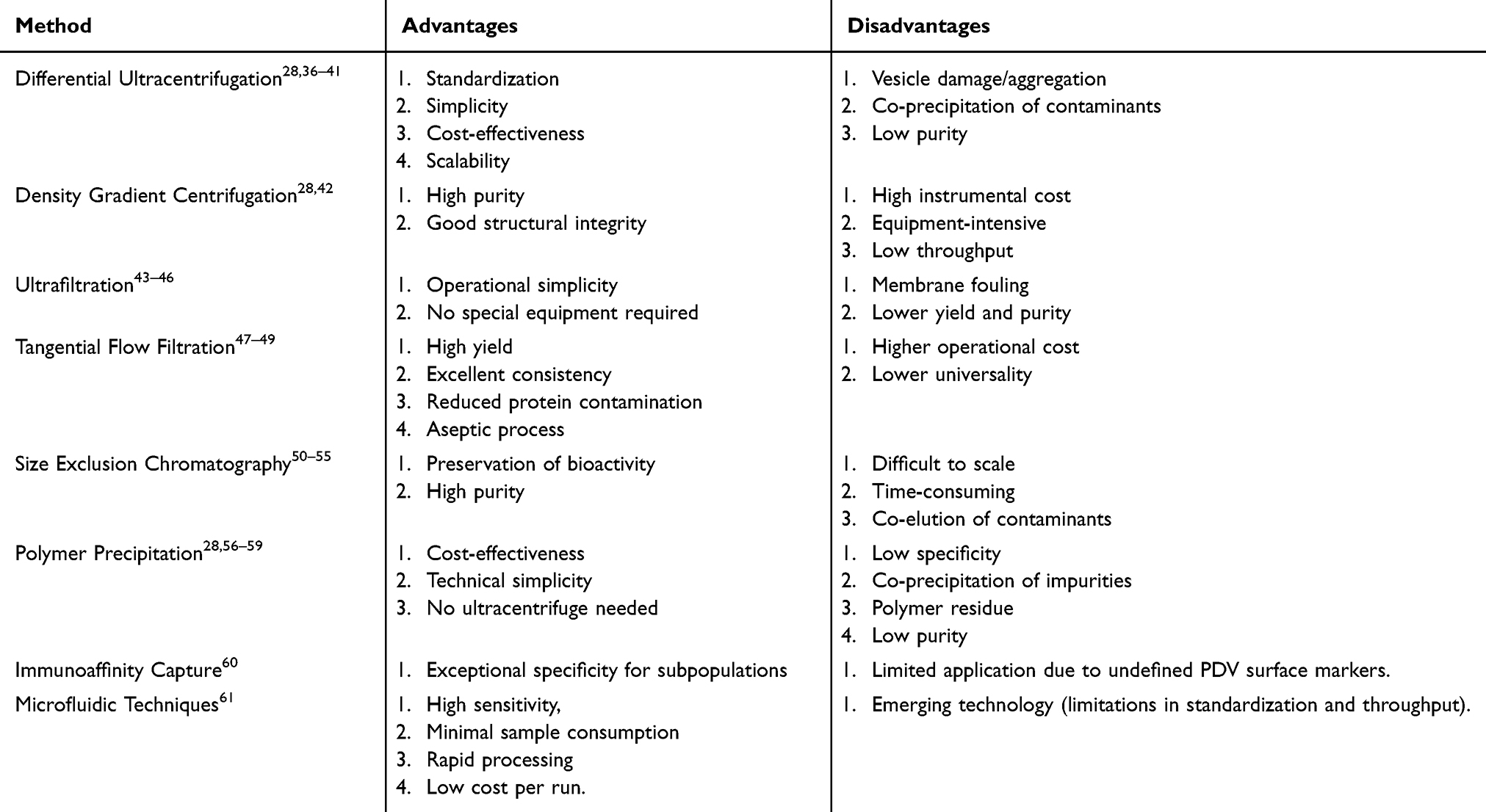 |
Table 1 Comparison of Different PDV Isolation Methods |
 |
Figure 2 Isolation methods and chemical composition of PDVs. (A) Digestion of the plant cell wall using cellulase and pectinase. (B) Acquisition of a PEV-containing solution via vacuum infiltration. (C) Acquisition of a PELN-containing solution via cell wall disruption (eg, extrusion, high-speed centrifugal juicing). (D) Isolation of PDVs by DUC. (E) Isolation by SEC. (F) Isolation by UF. (G) Isolation by TFF. (H) Isolation by PEG precipitation. (I) Isolation by immunoaffinity capture. (J) Isolation by microfluidics. (K) Purification of PDVs by DGC. |
Plant samples require preprocessing before PDV isolation. Obtaining PEVs typically involves vacuum infiltration to extract the plant’s extracellular fluid. Regente et al immersed intact sunflower seeds (after husk removal and a two-hour soak) in an infiltration buffer (50 mM Tris-HCl, pH 7.5, 0.6% NaCl, 0.1% 2-mercaptoethanol) and applied three vacuum pulses (10 seconds each, 30-second intervals) to obtain the sample solution.11 In 2017, Brian et al refined this method by subjecting plant material to 20 seconds of vacuum infiltration followed by centrifugation in a filter (700 × g, 20 min, 4 °C), successfully obtaining apoplastic wash fluid from Arabidopsis rosettes.62 This treatment yields PDVs that more closely adhere to the definition of EVs and exhibit higher purity. However, a significant limitation of this approach is its restricted applicability; due to the underlying mechanism, tissue infiltration is primarily suitable for fresh plant leaves. There are no reported applications of this technique for other plant parts, such as roots, stems, or fruits, severely limiting its scope.
In contrast, obtaining PELNs necessitates cell wall disruption of the plant material. Methods include squeezing, blending, and others, with the specific choice dependent on the plant type. Recommendations from an expert consensus on herbal vesicle research and application suggest that most fresh plants can undergo initial juicing and cell wall disruption for vesicle extraction. Fresh plants with high juice content can be directly squeezed or juiced with disruption. For some juicy plants rich in fibrous components, disruption and juicing may produce excessive flocculent fiber complexes that are difficult to remove during subsequent centrifugation/filtration; in these cases, simple squeezing is a more suitable method for juice collection. Dried products (sun-dried but unprocessed) and plants with low juice content can be juiced with disruption after adding an appropriate amount of pre-cooled phosphate-buffered saline (PBS). Fresh leaves may undergo infiltration to collect apoplastic wash fluid for vesicle acquisition. For fruits, seeds, and other plant parts rich in pectin, initial treatment with cellulase and pectinase can be used to digest the cell wall. Studies show that the specific method employed influences the compositional profile and levels of the resulting PDVs,63 with factors such as the plant source, mixing time and intensity, enzymatic reaction temperature, and duration playing crucial roles. Following the preprocessing of plant samples, the isolation and purification of PDVs can proceed.
Centrifugation-Based Isolation Techniques
Currently, differential centrifugation and sucrose density gradient centrifugation (DGC) remain the predominant isolation methods employed in most studies, capable of yielding PDVs with satisfactory structural integrity and purity.28 Differential ultracentrifugation (DUC) techniques separate different components based on their sequential sedimentation at progressively increasing centrifugal forces. This approach is the most commonly used method for isolating EVs and is widely considered the gold standard.36,37 The technique was initially described by Johnston et al in 1989 for isolating EVs from the culture medium of reticulocyte tissue.38 Subsequently, Théry and colleagues optimized this protocol by introducing an additional centrifugation step, thereby enhancing the separation efficiency.37 A standard DUC protocol typically involves an initial low-speed centrifugation step (<20,000 × g) to remove larger cells, cellular debris, and macromolecular contaminants. This is followed by high-speed or ultracentrifugation (>100,000 × g) to pellet the PDVs. This method is straightforward to implement, cost-effective, and suitable for the large-scale production of PDVs. However, DUC exhibits several significant limitations. During high-speed centrifugation, EVs may experience prolonged exposure to high shear forces, potentially leading to structural damage and aggregation.39,40 The high centrifugal forces can also cause co-precipitation of non-target materials, such as lipoproteins, resulting in contamination of the isolated PDV fraction.41 Furthermore, due to the inherent diversity of plant sources, PDVs isolated via centrifugation from different botanical origins may exhibit variations in particle size, concentration, and purity. Specific parameters of the isolation conditions—including centrifugal force, number of cycles, and temperature—can also influence the final composition and structure of the obtained PDVs.
PDVs isolated by differential centrifugation often possess relatively low purity and typically require further purification. DGC), a derivative ultracentrifugation technique, is commonly employed for this purpose. This method utilizes a centrifuge tube containing a medium (eg, sucrose or iodixanol) forming a continuous or discontinuous density gradient, with density decreasing from the bottom to the top. During ultracentrifugation, vesicles sediment to equilibrium positions within the gradient that correspond to their own buoyant densities. Following ultracentrifugation, most PDVs are typically concentrated within a density range of 1.13–1.19 g/mL, which corresponds to a sucrose concentration gradient of 30%–45%.28 PDVs are usually collected within this specific density band.
A major drawback of DGC is its requirement for expensive, specialized equipment, leading to high instrumental costs. Consequently, this technique is generally employed for PDV isolation in well-equipped laboratories. It is crucial to emphasize that no unified standard currently exists for isolating PDVs using centrifugation-based methods. The specific centrifugation steps can vary significantly depending on experimental conditions, sample source, and particular protocol variations.42 Therefore, isolating PDVs from each plant source necessitates the exploration of a personalized, customized protocol, undoubtedly increasing the complexity and difficulty of the experimental process.
Filtration-Based Separation Techniques
Filtration-based techniques for PDV isolation primarily include ultrafiltration (UF) and size exclusion chromatography (SEC). UF separates PDVs based on their size by filtering a solution under pressure through semi-permeable membranes with specific pore sizes. The protocol typically involves an initial filtration step using membranes with pore sizes of 0.8 μm and 0.45 μm to remove larger particles and cellular debris, resulting in a filtrate enriched with EVs. This is followed by a secondary filtration step using membranes with pore sizes smaller than the target vesicles to isolate the PDVs.43 Although UF is operationally simple, it often yields lower quantities and purity of isolated EVs compared to differential centrifugation.44 A major limitation is membrane fouling, where vesicles and other materials clog the membrane pores, significantly reducing the efficiency, purity, and yield of the UF process.45 This issue is particularly pronounced for plant samples (eg, from mugwort or ginseng) that contain abundant fibrous components post-homogenization, which are prone to clogging the filters. Optimizing filtration parameters—such as flow rate, pressure, and sample concentration, along with regular cleaning to remove accumulated impurities, can mitigate this problem.9 UF can also be combined with other methods, such as medium-speed centrifugation, to enhance sample purity and cost-effectiveness.46
Tangential flow filtration (TFF), a nanofiltration variant, enhances separation efficiency by directing the flow of the solution tangentially across the membrane surface. This hydrodynamic action minimizes pore clogging by preventing particles from accumulating on the membrane. Compared to ultracentrifugation, TFF offers several advantages, including increased PDV yield, reduced co-isolation of protein contaminants, excellent batch-to-batch consistency, and an aseptic operation process, making it a highly efficient method.47 TFF, often in combination with ultracentrifugation, has been successfully employed to isolate PDVs (50–200 nm) from aloe vera peels.48 Additionally, electrophoretic oscillation can be applied to dislodge impurities clogging the pores, thereby improving filtration speed and extending membrane lifespan.49 However, the higher operational costs compared to ultracentrifugation and its relatively lower universality remain notable drawbacks of TFF.47
SEC separates particles based on their hydrodynamic volume by passing the sample through a chromatography column packed with a porous stationary phase. Larger particles, such as PDVs, are unable to enter the pores and are eluted first in the void volume, while smaller molecules, including soluble proteins, penetrate the pores and are retained longer, leading to their separation.50,51 This method preserves the biological activity of PDVs and typically achieves high product purity. Research by Joel et al demonstrated that a combined UF-SEC approach, where plant juice containing PDVs is first concentrated via UF to reduce sample volume before being loaded onto an SEC column, significantly increased EV yield compared to ultracentrifugation alone. Importantly, the protein composition and biophysical properties of the vesicles remained unaltered.52 A comparative study by You et al evaluating Polyethylene glycol (PEG) precipitation, ultracentrifugation, and the UF-SEC method found no significant difference in yield among the techniques, but vesicles isolated via UF-SEC exhibited the highest purity level.53 However, a significant limitation of SEC is its low preparative scale-up potential due to the limited sample loading capacity of the columns and the time-consuming, cumbersome nature of the preparative process.54 Furthermore, vesicles isolated by SEC can be contaminated by lipoproteins and protein aggregates of similar size, which co-elute with the PDVs, thereby reducing the overall separation efficiency.55
Precipitation-Based Isolation Techniques
Precipitation is a technique that induces the sedimentation of PDVs from solution by altering their solubility and dispersion. PEG, a biocompatible polymer approved by the US Food and Drug Administration (FDA), is frequently employed for this purpose. PEG interacts with water molecules, creating a hydrophobic microenvironment around the PDVs that promotes their aggregation and subsequent precipitation. The precipitated vesicles can then be readily pelleted via low-speed centrifugation.56,57 Kalarikkal et al successfully isolated PDVs from ginger roots using PEG 6000. They demonstrated that the PDVs obtained via this method were comparable to those isolated by ultracentrifugation in terms of size, bioactivity, and biochemical composition. Although the recovery rate was approximately 60% of that achieved with ultracentrifugation, the biological activity of the vesicles remained unaffected.58 A significant advantage of the PEG precipitation method is its cost-effectiveness and operational simplicity, as it does not require specialized ultracentrifugation equipment. However, a major drawback of this technique is its propensity for co-precipitation of impurities, including heterogeneous vesicle populations, aggregates, and protein complexes. Furthermore, residual PEG 6000 (typically 4–8%) often remains associated with the isolated PDVs.28,59 Consequently, the purity of the resulting product is generally lower than that achieved with other methods, necessitating additional purification steps.
Immunoaffinity-Based Isolation Techniques
Immunoaffinity-based separation is a highly specific technique that utilizes antibody-conjugated magnetic beads designed to bind specifically to surface antigens on PDVs. The formation of these immuno-complexes allows for the selective isolation of specific extracellular vesicle subpopulations with exceptional specificity.60 However, the application of this technique to PDVs remains limited. This is primarily due to the incomplete characterization of the PDV surfaceome (the complete set of molecules on the cell surface) and the current lack of well-established, universal protein markers for PDVs, which are prerequisites for developing targeted antibodies.
Microfluidic-Based Isolation Techniques
Microfluidic technology enables the precise manipulation and control of fluids within microscale channels using forces such as acoustic, electric, or fluid viscoelastic forces. This capability facilitates the efficient separation and detection of PDVs.61 A key advantage of microfluidic systems is their ability to significantly enhance detection sensitivity while simultaneously minimizing sample and reagent consumption. Attributes such as low sample volume requirements, rapid processing times, and relatively low operational costs position microfluidic technology as an emerging and powerful tool for PDV isolation, holding considerable promise for future applications.
Physical Characterization of PDVs
To validate the efficacy of PDV isolation, a detailed characterization of key parameters, including morphology, particle size, and zeta potential, is essential. The morphology of PDVs is typically assessed using electron microscopy (EM) techniques, such as scanning electron microscopy (SEM), transmission electron microscopy (TEM), cryogenic transmission electron microscopy (cryo-TEM), and atomic force microscopy (AFM). These techniques are widely employed for the characterization and visualization of biological samples.64 Both TEM and SEM can reveal the characteristic cup-shaped morphology of EVs; however, this appearance may be an artifact induced by dehydration and fixation during sample preparation. In contrast, cryo-TEM preserves samples in a vitrified state at liquid nitrogen temperatures, circumventing the need for dehydration or chemical fixation. This approach minimizes structural damage from electron beam radiation and more accurately reveals the near-spherical morphology of vesicles in their native hydrated state.65,66 AFM operates by detecting and recording interactions between a sharp probe tip and the sample surface. A significant advantage of AFM is its ability to analyze samples under near-native conditions without requiring a vacuum environment, providing high-resolution, three-dimensional topographical images. A notable limitation, however, is that the periodic contact of the probe tip can induce vesicle movement, leading to potential signal interference and artifacts.67
The size distribution of PDVs is commonly determined using dynamic light scattering (DLS) and nanoparticle tracking analysis (NTA), with reported diameters typically ranging from 50 to 1000 nm. DLS, also referred to as photon correlation spectroscopy, relies on the scattering of a laser beam by particles in suspension and can measure particles from 1 nm to 6 μm. However, the accuracy of DLS measurements can be compromised in polydisperse suspensions due to their sensitivity to larger particles and its inherently low scattering intensity contrast. NTA tracks the Brownian motion of individual particles in suspension. This method offers rapid and straightforward sample preparation and measurement, enabling the precise sizing of particles as small as 30 nm in diameter. Since NTA analysis is performed directly in liquid suspension, the vesicles can be recovered in their native state post-measurement, ensuring their integrity for subsequent studies. A critical requirement for accurate NTA measurement is the appropriate dilution of the sample to avoid multiple scattering events.68 Owing to its precision and reliability for heterogeneous samples, NTA is currently regarded as the gold standard for extracellular vesicle characterization.66 Zeta potential measurement reflects the effective surface charge of particles in a dispersion and serves as a key indicator of the colloidal stability of the suspension.69 Studies indicate that PDVs isolated from various plant sources generally exhibit a negative zeta potential, with values typically ranging from −49.2mV to −1.5mV.33 It is important to note that the physical characteristics of PDVs, including size and zeta potential, can be influenced by environmental factors such as pH and storage conditions.70
Chemical Composition of PDVs
The composition of PDVs varies significantly depending on their biological source, but typically comprises four major classes of molecules: lipids, proteins, nucleic acids (Figure 2), and secondary metabolites. Each of these components plays a critical role in determining PDV function, stability, and biological activity.30
Lipids
PDVs possess a bilayer lipid membrane structure analogous to that of MDEVs, which effectively protects and delivers their internal cargo of bioactive molecules.71 The lipid composition of mammalian EVs is predominantly characterized by ceramides, cholesterol, and sphingomyelins.72 In contrast, PDVs lack cholesterol and sphingomyelins but are enriched in phospholipids such as phosphatidylcholine (PC), phosphatidic acid (PA), phosphatidylethanolamine (PE), phosphatidylinositol (PI), and phosphatidylglycerol (PG). Among these, PA is of particular interest due to its significant role in intercellular communication, participating in key biological processes including signal transduction, cell proliferation, and differentiation. The saturation level of these lipids influences the specificity of cellular uptake, a factor critical for drug delivery applications.73–76 Variations in lipid composition across different plant sources contribute to differences in the targeting specificity and cellular internalization efficiency of PDVs. Furthermore, the lipid constituents themselves can contribute to the intrinsic therapeutic activity of PDVs. For instance, the abundance of PE and PC in grapefruit-derived PDVs has been implicated in their observed antioxidant and anti-inflammatory processes.70,77
Proteins
Proteomic profiling serves as a vital criterion for verifying the presence of EVs and evaluating their therapeutic potential. While tetraspanins (eg, CD63, CD81, CD9) and TSG101 are well-established protein markers for mammalian EVs, universal protein markers for PDVs remain undefined and require further investigation.73 The protein content within PDVs is generally low. Proteomic analysis, typically conducted via mass spectrometry and liquid chromatography, alongside protein quantification using various assay kits, reveals that PDV proteins primarily include cytosolic proteins (eg, actin and proteases) and membrane proteins functioning as channels and transporters (eg, aquaporins and chloride channels).28 These proteins are postulated to facilitate uptake and internalization into mammalian cells and contribute to membrane stability.78 Research by Pinedo et al identified three commonly occurring protein families in plants: heat shock protein 70 (HSP70), S-adenosyl-homocysteinase, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Their study also proposed syntaxin PEN1, the ABC transporter PEN3, and tetraspanin-8 (TET8) as the most promising candidate protein markers for PDVs.34
Nucleic Acids
Research on PDV nucleic acids has primarily focused on microRNAs (miRNAs). High-throughput small RNA sequencing has confirmed the presence of miRNAs within PDVs. These abundant miRNAs can not only regulate plant biological functions but also target genes within the mammalian genome, thereby modulating mammalian gene expression.79 This suggests that exogenous, plant-derived miRNAs acquired through oral ingestion can be delivered and absorbed via the gastrointestinal tract, potentially influencing physiological and pathological processes in humans. A study by Xiao et al documented the presence of miRNAs in PDVs derived from 11 distinct plant species. These miRNAs were typically 20–22 nucleotides in length, and the variety and abundance of miRNA types varied considerably across the different species.80
Metabolites
Plants produce a vast array of secondary metabolites, which represent an important source of bioactive compounds with potential positive effects on human health.81 PDVs act as natural carriers for these plant metabolites, endowing them with considerable potential for therapeutic applications. It is important to note, however, that not all secondary metabolites produced by a given plant are detectable within its PDVs. For example, Berger et al found that orange juice-derived EVs lacked major active compounds such as vitamin C and naringenin.82
PDVs in Cancer Therapy
The therapeutic potential of PDVs in oncology is primarily manifested through two distinct mechanisms: (1) serving as nanocarriers for the targeted delivery of chemotherapeutic agents to tumor tissues, and (2) exerting direct anticancer effects via their inherent cargo of bioactive molecules. Owing to their innate role in intercellular communication, EVs were rapidly identified as promising candidates for drug delivery systems. Their negatively charged surface confers stability in the circulation, enabling them to efficiently traverse various biological barriers, including tissue, cellular, and intracellular barriers, to achieve targeted delivery and elicit responses in recipient cells. Furthermore, EVs mitigate the toxic side effects typically associated with the introduction of foreign substances into the body.83,84 Historically, research on EV-based delivery systems has predominantly utilized mammalian cell-derived EVs.9 Although promising, these platforms face significant challenges that hinder clinical translation, including limited production scalability, high costs, most notably, sensitive ethical concerns, and potential safety risks associated with the use of mammalian (including human) cell sources.85 In contrast, PDVs not only retain all the advantageous characteristics of mammalian EVs as drug carriers but also offer several unique benefits. As naturally secreted plant nanoparticles, PDVs are devoid of zoonotic or human pathogenic agents and do not cross the placental barrier,86 thereby exhibiting a superior safety profile with lower toxicity and immunogenicity.15 Moreover, the botanical sources for PDVs are abundant and cost-effective through large-scale cultivation. Studies indicate that the yield of PDVs isolated from citrus juice is at least 100-fold higher than that of EVs derived from mammalian cell cultures or bodily fluids.87 Research by Lee et al demonstrated that a standard juicer can process up to 3 liters of ginger juice per hour, a throughput equivalent to the output of 300 cell culture dishes. This approach drastically reduces the costs associated with cell culture media and reagents, as well as the time and labor required for large-scale mammalian cell culture.88 Overall, the cost-yield ratio for PDV production is estimated to be approximately 300 times lower than that for mammalian EVs. Additionally, PDVs can be isolated from in vitro cultured plant cells,89 and their yield can be optimized by modulating cultivation conditions,90 providing a solid foundation for scalable manufacturing. Another significant advantage is the public perception of safety; since the plants commonly used for PDV extraction (eg, fruits, vegetables) are often daily dietary staples and are also integral to TCM in East Asia, both researchers and potential patients harbor significantly fewer safety concerns regarding PDVs compared to their mammalian counterparts. Critically, numerous studies have shown that the bioactive molecules carried by PDVs endow them with intrinsic anticancer properties. A growing body of evidence has documented the anticancer effects of various PDV types (Table 2). Generally, PDVs can effectively inhibit cancer cell proliferation and induce cell death, while exhibiting minimal harmful effects on non-cancerous cells.
 |
Table 2 PDV Research Related to Cancer Therapy |
Citrus-Derived Vesicles
Citrus-derived EVs represent a class of natural nanocarriers that demonstrate significant potential in cancer therapy due to their rich composition of bioactive compounds, including carotenoids, flavonoids, and limonoids. These components possess natural antioxidant, anti-inflammatory, and antitumor activities, enabling PDVs to directly target cancer cells through multiple mechanisms and exert their inherent antitumor capabilities.
Grapefruit-Derived Vesicles
Grapefruit-derived vesicles have been extensively investigated for their dual functionality as both drug carriers and autonomous therapeutic agents. Foundational work by Wang et al established their ability to be efficiently internalized by various cancer cell lines, including GL26 glioma, A549 lung carcinoma, SW620 and CT26 colon carcinomas, and 4T1 breast cancer, without compromising encapsulated cargo activity. This enables the targeted delivery of chemotherapeutic drugs or siRNA while maintaining biological efficacy in vivo. A critical safety advantage is their inability to cross the placental barrier and their absence of toxicity toward major organs, underscoring their biosafety profile.86 Subsequent research has expanded these findings. Zhuang et al confirmed that grapefruit-derived vesicles could deliver miR-17 via intranasal administration to treat mouse brain tumors (GL26),91 while Teng et al further demonstrated their utility in delivering miR-18a, which effectively suppressed the growth of colon cancer (CT26) metastases in the liver.92 In a significant advance in 2021, Niu et al engineered a delivery system by attaching doxorubicin (DOX)-loaded heparin nanoparticles to the surface of grapefruit-derived vesicles. This approach achieved remarkable anti-glioma efficacy in vivo. Their experiments conclusively demonstrated the exceptional ability of these vesicles to cross the blood-brain barrier, their high drug-loading capacity, and their specific targeting of glioma cells. Moreover, grapefruit-derived vesicles significantly prolonged the systemic circulation time of the drug, enhanced its cellular internalization, and potentiated its antitumor effects, highlighting their immense potential as a drug delivery platform for cancer therapy.93
Beyond their role as delivery vehicles, grapefruit-derived vesicles themselves possess intrinsic antitumor activity. These vesicles can inhibit the growth of various cancer cell lines (eg, melanoma A375, human lung cancer A549, and breast cancer MCF-7), reducing cell viability by more than 40% without affecting normal human keratinocytes. In melanoma cells, the vesicles induce cell cycle arrest by downregulating cyclin B1 and B2 and upregulating CDKN1 (cyclin-dependent kinase inhibitor 1). Furthermore, they inhibit the Akt (serine/threonine kinase) and ERK (extracellular signal-regulated kinase) signaling pathways, thereby attenuating cancer cell viability. Grapefruit-derived vesicles also reduce the expression of cathepsins in melanoma cells, diminishing their invasiveness and suggesting a potential role in controlling metastatic progression in cancer therapy.94
Lemon-Derived Vesicles
Lemon-derived EVs represent a promising class of plant-based nanotherapeutics that demonstrate inherent pharmacological activity against cancer cells through multiple mechanistic pathways. Xiao et al established a biomimetic drug delivery system using lemon-derived vesicles that effectively overcomes multi-drug resistance and metastasis in ovarian cancer models, providing crucial evidence for their clinical translation potential.95 These vesicles exhibit selective cytotoxicity against various cancer cell lines, including human lung cancer A549, colorectal adenocarcinoma SW480, and chronic myeloid leukemia LAMA84, while showing minimal effects on normal cells such as bone marrow stromal cells HS5, umbilical vein endothelial cells HUVEC, and peripheral blood mononuclear cells PBMC. The antitumor mechanisms of lemon-derived vesicles involve sophisticated molecular pathways that directly target cancer cell survival. Research indicates that their selective toxicity is mediated through activation of TRAIL-induced apoptosis, which specifically triggers programmed cell death in malignant cells.96 Further mechanistic investigations revealed that these vesicles inhibit lipid metabolism pathways, particularly through downregulation of ACACA, while simultaneously inducing both cell cycle arrest and apoptotic processes.97 Additional studies demonstrate that lemon-derived vesicles cause concentration-dependent S-phase arrest in gastric cancer cell lines AGS, BGC-823, and SGC-7901, accompanied by increased reactive oxygen species (ROS) generation and upregulation of GADD45a, ultimately leading to apoptotic cell death.98
Ginger-Derived Vesicles
Ginger-derived EVs represent a significant class of plant-based nanotherapeutics that harness the inherent bioactivity of traditional medicinal compounds. These vesicles naturally contain bioactive components, including gingerols, shogaols, and zingerone, which demonstrate potent anti-inflammatory and antioxidant properties with established anticancer and antimutagenic activities.122–124 The pharmacological significance of these compounds is particularly evident in cancer prevention and treatment, where zingerone has been shown to enhance T cell-mediated antibody responses in breast cancer models,99 while 6-shogaol exerts antiproliferative effects through induction of oxidative stress, pro-apoptotic signaling, and DNA damage in breast cancer cells, highlighting their potential in cancer immunotherapy.100 Additionally, ginger-derived vesicles themselves have been shown to effectively ameliorate acute colitis by attenuating inflammatory mediators while promoting healing processes, thereby enhancing intestinal repair and preventing the progression to chronic colitis and colitis-associated cancer (CAC).101
To improve tumor-specific delivery, several innovative engineering approaches have been developed for ginger-derived vesicles. Zhang et al implemented a folic acid functionalization strategy, capitalizing on the overexpression of folate receptors on cancer cells. This approach enabled precise tumor targeting and pH-dependent release of DOX payloads, resulting in enhanced chemotherapeutic efficacy against colon cancer while sparing normal tissues.102 In another approach, Lee et al engineered ginger-derived vesicles by conjugating cell-targeting ligands onto their surface using arrowtail RNA nanoparticles. This targeted delivery system exhibited significant tumor suppression in a xenograft model using KB cells (human oral epidermal carcinoma cells).88 Furthermore, Zhang et al developed a sophisticated layer-by-layer assembly technique to functionalize ginger-derived vesicles with fucoidan and polylysine. This design specifically targeted P-selectin (CD62P), which serves as a specific delivery target for colon cancer therapy. The resulting engineered vesicles successfully delivered their payload to vascularized colon tumors. Results demonstrated that these functionalized vesicles significantly inhibited the growth of human colon cancer cells (Luc-HT-29 and HCT-116) and showed potential in preventing the development of drug resistance.103
The clinical relevance of ginger-derived vesicles is particularly emphasized by their oral administration potential. Long et al demonstrated that the oral administration of M13 bacteriophage-loaded ginger-derived vesicles is both safe and effective for treating mouse CAC, offering a promising non-invasive route for clinical translation.104
The collective evidence positions ginger-derived EVs as versatile platforms for cancer therapy, combining inherent bioactive properties with engineerable targeting capabilities. Their dual functionality as both therapeutic agents and drug delivery vehicles, coupled with their excellent safety profile and oral bioavailability, suggests substantial potential for clinical development in cancer prevention and treatment strategies.
Ginseng-Derived Vesicles
Ginseng-derived EVs (GDVs) exhibit a unique convergence of traditional pharmacological knowledge and contemporary nanomedicine applications. In TCM, ginseng has long been utilized as a common supplement to enhance bodily functions. Its active constituents, particularly ginsenosides, exert antitumor effects through multiple mechanisms, including cell cycle inhibition, induction of cell death, modulation of angiogenesis, and suppression of tumor metastasis.125 Beyond ginsenosides, a bioactive fraction isolated from total ginseng saponins, termed PDS-C, has been shown to alleviate chemotherapy-induced myelosuppression and pancytopenia caused by cyclophosphamide (CTX).126 As research progresses, the antitumor properties of other ginseng components are being uncovered. For instance, gintonin can inhibit the metastasis of B16/F10 melanoma cells by inducing the activity of autotaxin (ATX), a key enzyme responsible for the production of extracellular lysophosphatidic acid.127
Recent studies have revealed GDVs’ immunomodulatory capabilities in cancer therapy. Research demonstrates that GDVs induce polarization of tumor-associated macrophages (TAMs) toward the M1 phenotype via TLR4/MyD88 signaling, thereby inhibiting melanoma growth.105 Further investigation revealed that GDVs reprogram TAMs, leading to increased secretion of the chemokines CCL5 and CXCL9. This chemokine gradient promotes the recruitment of CD8⁺ T cells into the tumor microenvironment (TME) and synergizes with PD-1 monoclonal antibody therapy to enhance the antitumor efficacy of tumor-infiltrating T lymphocytes. This combined approach induced robust and sustained systemic antitumor immunity across multiple mouse tumor models.106 Subsequent research elucidates that GDVs modulate the mTOR-T-bet axis to reprogram macrophages, resulting in downregulation of arginase-1 expression, which promotes T cell proliferation and activation while ameliorating T cell exhaustion.107
GDVs exhibit therapeutic effects across various cancer types. In lung cancer, they inhibit cancer cell migration and proliferation by suppressing the pentose phosphate pathway.108 Additionally, GDVs demonstrate blood-brain barrier penetration capability for targeted glioma therapy. Their antitumor action is mediated through the regulation of cell proliferation and modulation of the TME.109
Furthermore, Wang et al developed a novel hybrid membrane cancer vaccine by fusing GDVs with membranes derived from resected autologous tumors. This vaccine enhances adaptive immune responses and suppresses tumor recurrence and metastasis through enhanced antigen phagocytosis and dendritic cell maturation via TLR4 signaling.110
The accumulated evidence positions GDVs as promising therapeutic agents that leverage natural bioactive components to target cancer cells while modulating the tumor immune microenvironment. Their compatibility with existing therapies and demonstrated efficacy across multiple cancer models highlight their potential for clinical translation in cancer treatment.
Bitter Melon-Derived Vesicles
Bitter melon (Momordica charantia) is a plant recognized for its anticancer and anti-inflammatory activities. Extracts derived from bitter melon have demonstrated significant antitumor efficacy against various cancer types, including lymphocytic leukemia, lymphoma, melanoma, and breast cancer.111 The therapeutic applications of bitter melon are further enhanced through its EVs.
Research by Yang et al indicated that bitter melon-derived vesicles not only display intrinsic antitumor activity against oral squamous cell carcinoma (OSCC) but also significantly enhance the therapeutic effect of conventional chemotherapy. Specifically, when combined with 5-fluorouracil (5-FU), bitter melon-derived vesicles demonstrate a synergistic effect that potentiates the drug’s efficacy while simultaneously reducing the development of drug resistance. This combination approach represents a promising strategy for improving treatment outcomes in oral cancer management, particularly in cases where drug resistance poses a significant clinical challenge.112 Furthermore, studies by Wang et al revealed that bitter melon-derived vesicles exert anti-glioma effects by modulating the PI3K/AKT signaling pathway, effectively inhibiting glioma growth and metastasis both in vitro and in vivo.111
The multifaceted nature of bitter melon-derived vesicles extends beyond direct antitumor effects. Their natural origin provides advantages in terms of biocompatibility and reduced immunogenicity, while their ability to target multiple aspects of cancer progression positions them as promising candidates for developing innovative combination therapies against various malignancies, particularly those with limited treatment options and high propensity for drug resistance development.
Garlic-Derived Vesicles
Research by Özkan et al demonstrates that garlic-derived EVs possess inherent antitumor activity by directly inducing caspase-mediated apoptosis in human renal carcinoma (A498) and lung cancer (A549) cells, significantly suppressing their proliferation. Critically, these vesicles exhibit selective cytotoxicity by targeting malignant cells without harming healthy human dermal fibroblasts, underscoring their biosafety and natural targeting specificity.113 In summary, garlic-derived vesicles demonstrate that PDVs can selectively induce apoptosis in cancer cells through natural bioactive compounds, offering a novel approach for developing low-toxicity targeted anticancer drugs.
Tea Flower/Leaf-Derived Vesicles
Tea-derived nanovesicles demonstrate remarkable broad-spectrum antitumor efficacy through multiple synergistic mechanisms, positioning them as versatile candidates for cancer therapy. Chen et al confirmed that natural nanovesicles from tea flowers exhibit potent broad-spectrum antitumor activity in vitro, including inhibitory effects on human breast cancer cell line MCF-7, mouse breast cancer cell line 4T1, human lung cancer cell line A549, and human cervical cancer cell line HeLa. In mouse models, these vesicles suppressed the growth of mammary tumors (MCF-7) and pulmonary metastasis by promoting ROS generation and modulating microbiota specificity.114 The research team further developed tea leaf-derived exosome-like nanoparticles that achieve over 80% cellular internalization in breast cancer models, inducing mitochondrial damage and cell cycle arrest through ROS-mediated pathways. Importantly, these nanoparticles demonstrate exceptional oral bioavailability, being efficiently absorbed through the small intestine and modulating TME without detectable toxicity or immune activation, offering a non-invasive therapeutic approach with significant clinical translation potential.115
Hemp-Derived Vesicles
Hemp-derived EVs demonstrate significant antitumor efficacy through well-defined mechanistic pathways. Research by Tajik et al established that these vesicles reduce the viability of hepatocellular carcinoma cell lines (HepG2 and Huh-7) in a dose- and time-dependent manner, primarily by inducing G0/G1 phase cell cycle arrest and activating the mitochondrial-dependent apoptotic pathway. This targeted approach highlights their capacity to directly interfere with cancer cell proliferation mechanisms while leveraging natural bioactive components.116 The study provides compelling evidence for the specific anticancer activity of hemp-derived vesicles, positioning them as promising candidates for further development in liver cancer therapeutics.
Asparagus-Derived Vesicles
Asparagus cochinchinensis is a renowned medicinal plant known for its various therapeutic effects, including anticancer, antioxidant, and anti-infective properties.128 Research by Zhang et al demonstrated that Asparagus cochinchinensis-derived EVs (ACNVs) significantly inhibit the growth of human hepatocellular carcinoma (HCC) cell lines (HepG2, Hep3B, and SMMC-7721) in vitro, induce apoptosis, and upregulate apoptosis-related factors, while showing no significant toxicity towards normal liver cell lines. In vivo experiments in HepG2 tumor-bearing mice confirmed that ACNVs inhibit tumor growth without producing apparent side effects, underscoring their translational potential.117 The combination of direct anticancer activity and excellent safety profile positions these vesicles as promising candidates for liver cancer therapy.
Artemisia-Derived Vesicles
Artemisia-derived vesicles (ADNVs) exhibit remarkable immunomodulatory capabilities through sophisticated macrophage reprogramming mechanisms. Research by Liu et al confirmed that ADNVs can reprogram TAMs from a pro-tumor phenotype to a pro-inflammatory phenotype, demonstrating potent antitumor activity. Mitochondrial DNA carried by ADNVs is acquired by TAMs, inducing the cGAS-STING pathway to reprogram macrophages, driving the transition from pro-tumor to anti-tumor macrophages, thereby enhancing the cytotoxic T cell response to promote tumor regression. The study further demonstrated that ADNVs can enhance the efficacy of αPD-L1-mediated immunotherapy by activating the STING-driven pathway.118 This ability to convert immunosuppressive macrophages into antitumor effectors highlights the immunotherapeutic potential of these PDVs.
Morus nigra L. Leaf-Derived Vesicles
Morus nigra L. leaves are a natural herb with antioxidant, hypoglycemic, antibacterial, and anti-inflammatory properties. Research by Gao et al indicated that EVs derived from Morus nigra L. Leaves (MLNPs), as a novel oral nanomedicine, exhibit multifaceted functions and potential in hepatocellular carcinoma therapy. MLNPs are specifically taken up by liver tumor cells via galactose receptor-mediated endocytosis, subsequently triggering a surge in intracellular ROS levels and mitochondrial damage, leading to G0/G1 phase cell cycle arrest and apoptosis in tumor cells, effectively inhibiting cell proliferation, migration, and invasion. Furthermore, orally administered MLNPs maintain high stability in simulated gastrointestinal environments and, in vivo, not only directly kill tumor cells but also regulate the balance of gut microbiota. MLNPs demonstrated significant liver enrichment capacity and tumor growth inhibition in a mouse model of liver cancer, without inducing apparent immunogenicity or toxic side effects.119 This comprehensive approach positions them as promising candidates for safe and efficient oral drug development.
Brucea javanica-Derived Vesicles
Brucea javanica (L). Merr. is a traditional Chinese herbal medicine. Oil extracted from its seeds possesses various pharmacological activities, demonstrating efficacy against liver, lung, breast, gastric, and colon cancers, while also reducing adverse reactions caused by chemotherapy and radiotherapy.129 Brucea javanica-derived EVs demonstrate versatile antitumor capabilities through dual targeting mechanisms. Yan et al proved that these extracellular vesicle-like nanoparticles regulate the biological functions of endothelial cells in the TME and inhibit VEGF-mediated angiogenesis. Simultaneously, BF-EVs can transfer active miRNAs into tumor cells, intervening through the oncogenic key pathway PI3K/Akt/mTOR and inhibiting tumor growth, representing an effective nano-delivery platform.120 The ability to concurrently address both tumor vasculature and cancer cell proliferation provides a comprehensive approach to cancer therapy, particularly for aggressive cancers requiring multi-targeted interventions.
Lonicera japonica-Derived Vesicles
Lonicera japonica is a traditional Chinese medicinal herb. Research by Chi et al demonstrated that the active ingredient miR2911 is present in extracellular vesicle-like nanoparticles extracted from honeysuckle. This miRNA inhibits the proliferation of cervical cancer cells and induces apoptosis by targeting the E6/E7 genes of HPV16/18, holding significant importance for developing new therapeutic strategies against HPV-associated cervical cancer.121 These natural miRNA packaging and delivery system offer a targeted approach for treating virus-associated malignancies while leveraging the inherent biocompatibility of PDVs.
Current Challenges and Future Perspectives
As summarized in the preceding sections, substantial progress has been made in the isolation, characterization, and application of PDVs in oncology. Leveraging their inherent advantages as natural nanocarriers, including a distinctive bilayer lipid structure, the capacity for cross-species communication, a payload of endogenous bioactive molecules, and generally high public acceptance, PDVs have demonstrated multifaceted value in cancer therapy. Specifically, PDVs derived from sources such as citrus, ginger, and ginseng can elicit direct antitumor effects by inducing apoptosis, modulating the immune microenvironment, and suppressing metastasis. Concurrently, their utility as drug delivery vehicles enables the efficient transport of chemotherapeutic agents and nucleic acids, significantly enhancing targeting precision while reducing systemic toxicity. Notwithstanding this considerable promise, the clinical translation of PDVs is impeded by four fundamental challenges: the absence of scalable and standardized isolation methods, the lack of a unified classification system and consensus markers, insufficient systematic databases for functional annotation and matching to diseases, and inadequate safety assessment frameworks based on compositional and functional understanding (Figure 3).
 |
Figure 3 Current Challenges and Future Perspectives in PDV Research. (A) Limitations of current plant extracellular vesicle isolation methodologies and future development directions. (B) The requirement for unified marker systems and classification standards for PDVs. (C) Construction of a systematic PDV database for disease matching. (D) Establishment of adverse effect profiles for various PDVs to ensure clinical safety. |
Scalable, Standardized, and Automated Isolation: A Prerequisite for Translation
The development of large-scale, standardized, and automated isolation technologies is a critical prerequisite for the translational application of PDVs. Current methodologies rely predominantly on ultracentrifugation. Although this technique is regarded as the “gold standard” for laboratory-scale isolation of EVs (particularly those from mammalian sources), it exhibits several formidable limitations for industrial-scale production. First, processing efficiency is severely constrained; the typical sample volume per batch is limited to < 500 mL, and the process is prohibitively time-consuming (often requiring several hours), rendering it inadequate for clinical-grade manufacturing demands. Second, the manual, labor-intensive nature of ultracentrifugation, coupled with its limited sample-processing capacity, introduces significant batch-to-batch variability, thereby compromising product consistency and reproducibility, key requirements for clinical standardization. Third, the structural integrity of vesicles is often compromised; the high shear forces generated during ultracentrifugation can cause vesicle rupture or irreversible aggregation, directly impairing their biological functionality. Fourth, co-precipitation of contaminants presents a major issue; plant matrices are rich in impurities such as polysaccharides (eg, starch) and tannins, which readily co-sediment with PDVs, substantially reducing product purity. Fifth, the approach is economically challenging; ultracentrifugation equipment and ancillary systems require substantial capital investment (often reaching millions of monetary units) and incur high maintenance costs. Alternative techniques offer limited solutions. SEC can improve purity but suffers from even lower throughput. Polyethylene glycol (PEG) precipitation, while operationally simple, often leaves residual polymer contaminants that are difficult to eliminate, raising potential clinical safety concerns. Furthermore, PDV yield is subject to variation even when using identical protocols, due to factors such as plant source, harvest season, freshness, and specific isolation parameters.
In response to these challenges, recent innovations have focused on developing efficient, gentle, and cost-effective automated isolation platforms. Notably, a fully automated system (termed FACTORY) integrating continuous-flow centrifugation with TFF has been reported to achieve standardized, automated, and large-scale isolation of EVs from liquid samples.130 Although initially designed for MDEVs, this platform can be adapted for the large-scale production of PDVs from pre-processed botanical sources (eg, garlic). The resulting vesicles are sterile, mycoplasma-free, and exhibit low endotoxin levels, thereby laying a solid foundation for the clinical translation of PDVs.
Based on this, we propose control standards for PDV isolation. Morphological criteria require TEM to demonstrate that prepared PDVs exhibit a characteristic cup-shaped morphology. NTA confirms that the particle size distribution of EVs is concentrated between 50200 nm, consistent with the size range of natural EVs.
However, it is crucial to emphasize that the efficient isolation of PDVs using this system remains contingent on the pre-processing of raw plant material. Divergences in pre-processing protocols significantly impact the yield, purity, and compositional profile of the isolated vesicles. Consequently, future research must prioritize the standardization and automation of upstream plant processing workflows. Ensuring batch-to-batch consistency of initial crude extracts and establishing integrated, end-to-end automated production lines, encompassing plant washing, disruption, vesicle isolation, and quality control, are essential to provide a stable and controllable source of raw material for clinical-grade PDV applications.
Unified Marker Systems and Classification Standards: A Theoretical Imperative
Research on PDVs is constrained by the dual challenges of an incomplete marker system and the absence of universally accepted classification criteria. On one hand, canonical protein markers of mammalian exosomes (eg, tetraspanins CD63, CD81, and CD9) lack conservation in plants, and proposed plant-specific candidates (eg, TET8 in Arabidopsis thaliana) have not garnered cross-species consensus. On the other hand, the International Society for Extracellular Vesicles (ISEV) has not yet established definitive nomenclature rules for PDVs, leading to a proliferation of classification and naming conventions that hinder cross-study comparisons. The compositional profile of PDVs is influenced by the plant source, biogenetic pathway, and biochemical isolation methods. Therefore, establishing a standardized pipeline for the identification and validation of PDVs is imperative. Implementing multi-omics analyses (eg, lipidomics, proteomics, nucleic acid profiling, and metabolomics) to conduct multi-parametric characterization of PDV composition, coupled with the creation of a comprehensive, open-access database, will be instrumental in harmonizing research efforts and providing a robust data foundation for future applications.
Systematic PDV Database Construction: A Core Pathway to Clinical Application
Mirroring the heterogeneity of MDEVs, PDVs from different plant sources exhibit substantial diversity in their cargo and consequent biological activities. This heterogeneity suggests that distinct PDVs may be optimally suited for treating different diseases. Therefore, the construction of a systematic PDV database, incorporating detailed compositional analyses (proteins, lipids, nucleic acids, metabolites) and functional annotations, represents a cornerstone of their clinical translation. However, current research remains fragmented; PDVs from a given plant are typically investigated against only one or a few disease models, leaving their broader therapeutic potential underexplored. Moreover, a robust screening framework for identifying the optimal PDV source for a specific medical indication is presently lacking. Addressing this gap requires the application of high-throughput technologies to comprehensively catalog the biomolecular cargo of diverse PDVs. Building a centralized database and developing AI-driven functional prediction models based on this information will ultimately enable the precise matching of PDV sources to specific disease pathologies. This constitutes a critical research frontier and an essential piece of infrastructure that must be established to facilitate the rational development of PDV-based therapeutics.
Safety Assessment: The Fundamental Guarantee for Clinical Translation
Comprehensive safety profiling constitutes an indispensable foundation for the clinical deployment of PDVs, requiring systematic evaluation of both general toxicological parameters and plant-specific risk factors. The assessment framework must incorporate specific endpoints, including acute toxicity, immunogenicity, and organ-specific toxicity, while addressing unique risks associated with different plant sources, such as potential toxic metabolites in medicinal herb-derived PDVs. Qualified PDVs must have endotoxin levels below 0.25 EU/mL (FDA standard for injectable water), test negative for mycoplasma, and show no contamination in bacterial cultures after 7 days. Following tail vein injection of EVs (1×10^11 particles/dose) in mice, no abnormalities were observed in body weight, blood counts, biochemical parameters, or histology of major organs.130
The necessity for rigorous safety assessment is underscored by the well-established dietary restrictions for patients with specific pathophysiological conditions. Diabetic patients require avoidance of high-glycemic index foods (eg, potatoes, taro, sweet potatoes); renal impairment patients need low-potassium and low-sodium diets (avoiding spinach, celery, amaranth); gout patients must reduce high-purine food intake (eg, mushrooms, legumes, seafood); and thyroid disorder patients should limit high-iodine and goitrogenic foods (eg, kelp, nori, cabbage). These dietary considerations directly inform the safety assessment priorities for PDVs derived from corresponding plant sources.
Given that PDVs inherit both bioactive components and fundamental biological properties from their parent plants, each PDV possesses a unique therapeutic profile requiring a clear definition of both indicated applications and contraindications. Consequently, the establishment of a rigorous safety assessment framework must precisely define critical pharmacological parameters, including in vivo pharmacokinetics, biodistribution patterns, and maximum tolerated dose, while incorporating specific evaluation of immunogenic potential and plant metabolite-related toxicity.
The development of this framework relies on integrating reliable vesicle labeling strategies with advanced tracking methodologies. These technologies are essential for delineating the precise biological fate of PDVs post-administration, particularly for assessing biodistribution and accumulation patterns in non-target tissues. Current research remains predominantly focused on therapeutic benefits, with investigations into potential adverse effects, contraindications, or long-term safety implications still limited.
To address these gaps, operable guidance should include: (1) standardized protocols for acute toxicity testing using relevant animal models; (2) comprehensive immunogenicity assessment, including cytokine profiling and immune cell activation assays; (3) specific screening for plant-derived toxic metabolites; and (4) long-term toxicity studies evaluating cumulative effects. These measures will establish essential risk assessment profiles necessary before clinical deployment, contingent upon acquiring a deep systematic understanding of PDV composition and biological functions.
In summary, we draw a clinical translational roadmap for PDVs. 1) Preclinical Exploration Phase: Focused on resolving fundamental challenges in PDV research, including the establishment of standardized isolation protocols, identification of PDV-specific biomarkers, and clarification of bioactive mechanisms. 2) Process Development Phase: Addressing scalable production technologies (eg, FACTORY), quality control standards, and functional characterization of PDVs from diverse plant sources. 3) Safety and Efficacy Validation Phase: Acknowledge the need for systematic safety assessment frameworks, including pharmacokinetic studies, immunogenicity profiling, and plant-specific risk evaluations (eg, metabolite screening and allergen detection). 4) Clinical and Regulatory Integration Phase: Proposing pathways for clinical trial design, regulatory alignment, and commercialization strategies tailored to PDV-based therapeutics.
Conclusions
PDVs exhibit significant potential in cancer therapeutics, serving as dual-function agents that combine efficient drug delivery with inherent bioactivity. The diversity of botanical sources offers a platform for tailored treatments, yet clinical translation faces challenges in standardization, scalable production, and safety validation. Future efforts should focus on multidisciplinary collaborations that integrate advanced analytics and computational methods to address these gaps. By overcoming these hurdles, PDVs could bridge traditional and modern medicine, paving the way for accessible cancer therapies.
Abbreviations
ACACA, Acetyl-CoA carboxylase alpha; ADNVs, Artemisia-derived vesicles; AFM, Atomic force microscopy; AKT, Serine/threonine kinase (Protein kinase B); ARG1, Arginase-1; ATCC, American Type Culture Collection; ATX, Autotaxin; ACNVs, Asparagus cochinchinensis-derived EVs; BF-EVs, Brucea javanica-derived EVs; CAC, Colitis-associated cancer; cGAS, Cyclic GMP-AMP synthase; CTX, Cyclophosphamide; cryo-TEM, Cryogenic transmission electron microscopy; CXCL9, Chemokine (C-X-C motif) ligand 9; DGC, Density gradient centrifugation; DLS, Dynamic light scattering; DUC, Differential ultracentrifugation; DOX, Doxorubicin; EM, Electron microscopy; ERK, Extracellular signal-regulated kinase; EXPO, Exocyst-positive organelle; EVs, Extracellular vesicles; FDA, US Food and Drug Administration; GADD45a, Growth arrest and DNA damage-inducible protein 45 alpha; GAPDH, Glyceraldehyde-3-phosphate dehydrogenase; GDVs, Ginseng-derived vesicles; HCC, Human hepatocellular carcinoma; HSP70, Heat shock protein 70; HS5, Human bone marrow stromal cells (cell line); HUVEC, Human umbilical vein endothelial cells; HPV, Human papillomavirus; ISEV, International Society for Extracellular Vesicles; M13, 5-glutathione-6-shogaol; MLNPs, Natural exosome-like lipid nanoparticles (from Morus nigra L. leaves); mTOR, Mammalian target of rapamycin; MVBs, Multivesicular bodies; miRNAs, microRNAs; MyD88, Myeloid differentiation primary response 88; NTA, Nanoparticle tracking analysis; OSCC, Oral squamous cell carcinoma; αPD-L1, Anti-programmed death-ligand 1; PBS, Phosphate-buffered saline; PC, Phosphatidylcholine; PA, Phosphatidic acid; PE, Phosphatidylethanolamine; PI, Phosphatidylinositol; PG, Phosphatidylglycerol; PEG, Polyethylene glycol; PDVs, Plant-derived vesicles; PELNs, Plant extracellular vesicle-like nanovesicles; PEVs, Plant extracellular vesicles; PLNVs, Plant lipid-derived nanovesicles; PVLNs, Plant vesicle-like particles; PEN1, Syntaxin PEN1; PEN3, ABC transporter PEN3; PI3K, Phosphatidylinositol 3-kinase; ROS, Reactive oxygen species; SEC, Size exclusion chromatography; SEM, Scanning electron microscopy; STING, Stimulator of interferon genes; TCM, Traditional Chinese medicine; TAMs, Tumor-associated macrophages; TME, Tumor microenvironment; TRAIL, TNF-related apoptosis-inducing ligand; TSG101, Tumor susceptibility gene 101; TET8, Tetraspanin-8; TFF, Tangential flow filtration; TEM, Transmission electron microscopy; TLR4, Toll-like receptor 4; UF, Ultrafiltration; Tris-HCl, Tris(hydroxymethyl)aminomethane-hydrochloric acid; PBMC, Peripheral blood mononuclear cells; 5-FU, 5-fluorouracil.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Taishan Scholar Project of Shandong Province of China (tsqn202103200) and Traditional Chinese Medicine science and Technology Project of Shandong Province (Z20241405).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–25. doi:10.1038/nrm.2017.125
2. Jeppesen DK, Zhang Q, Franklin JL, Coffey RJ. Extracellular vesicles and nanoparticles: emerging complexities. Trends Cell Biol. 2023;33(8):667–681. doi:10.1016/j.tcb.2023.01.002
3. Dixson AC, Dawson TR, Di Vizio D, Weaver AM. Context-specific regulation of extracellular vesicle biogenesis and cargo selection. Nat Rev Mol Cell Biol. 2023;24(7):454–476. doi:10.1038/s41580-023-00576-0
4. Chargaff E, West R. The biological significance of the thromboplastic protein of blood. J Biol Chem. 1946;166(1):189–197. doi:10.1016/S0021-9258(17)34997-9
5. Nemati M, Singh B, Mir RA, et al. Plant-derived extracellular vesicles: a novel nanomedicine approach with advantages and challenges. Cell Commun Signaling. 2022;20(1):69. doi:10.1186/s12964-022-00889-1
6. Buzas EI. The roles of extracellular vesicles in the immune system. Nat Rev Immunol. 2023;23(4):236–250. doi:10.1038/s41577-022-00763-8
7. Cheng L, Hill AF. Therapeutically harnessing extracellular vesicles. Nat Rev Drug Discov. 2022;21(5):379–399. doi:10.1038/s41573-022-00410-w
8. Kalluri R, McAndrews KM. The role of extracellular vesicles in cancer. Cell. 2023;186(8):1610–1626. doi:10.1016/j.cell.2023.03.010
9. Zhao Y, Tan H, Zhang J, et al. Plant-derived vesicles: a new era for anti-cancer drug delivery and cancer treatment. Int J Nanomedicine. 2023;18:6847–6868. doi:10.2147/IJN.S432279
10. Halperin W, Jensen WA. Ultrastructural changes during growth and embryogenesis in carrot cell cultures. J Ultrastruct Res. 1967;18(3–4):428–443. doi:10.1016/S0022-5320(67)80128-X
11. Regente M, Corti-Monzon G, Maldonado AM, Pinedo M, Jorrin J, de la Canal L. Vesicular fractions of sunflower apoplastic fluids are associated with potential exosome marker proteins. FEBS Lett. 2009;583(20):3363–3366. doi:10.1016/j.febslet.2009.09.041
12. Feng J, Xiu Q, Huang Y, Troyer Z, Li B, Zheng L. Plant-derived vesicle-like nanoparticles as promising biotherapeutic tools: present and future. Adv Mater. 2023;35:e2207826.
13. Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. 2023;73(1):17–48. doi:10.3322/caac.21763
14. Qi J, Li M, Wang L, et al. National and subnational trends in cancer burden in China, 2005-20: an analysis of national mortality surveillance data. Lancet Public Health. 2023;8(12):e943–e55. doi:10.1016/S2468-2667(23)00211-6
15. Zhu Y, Zhou X, Yao Z. A mini-review: advances in plant-derived extracellular vesicles as nano-delivery systems for tumour therapy. Front Bioengine Biotechnol. 2022;10:1076348. doi:10.3389/fbioe.2022.1076348
16. Basak D, Arrighi S, Darwiche Y, Deb S. Comparison of anticancer drug toxicities: paradigm shift in adverse effect profile. Life. 2021;12(1):12. doi:10.3390/life12010012
17. de Abreu RC, Fernandes H, da Costa Martins PA, Sahoo S, Emanueli C, Ferreira L. Native and bioengineered extracellular vesicles for cardiovascular therapeutics. Nat Rev Cardiol. 2020;17(11):685–697. doi:10.1038/s41569-020-0389-5
18. Chiang CL, Ma Y, Hou YC, et al. Dual targeted extracellular vesicles regulate oncogenic genes in advanced pancreatic cancer. Nat Commun. 2023;14(1):6692. doi:10.1038/s41467-023-42402-3
19. Kooijmans SA, Aleza CG, Roffler SR, van Solinge WW, Vader P, Schiffelers RM. Display of GPI-anchored anti-EGFR nanobodies on extracellular vesicles promotes tumour cell targeting. J Extracell Vesicles. 2016;5(1):31053. doi:10.3402/jev.v5.31053
20. Jia G, Han Y, An Y, et al. NRP-1 targeted and cargo-loaded exosomes facilitate simultaneous imaging and therapy of glioma in vitro and in vivo. Biomaterials. 2018;178:302–316. doi:10.1016/j.biomaterials.2018.06.029
21. Xia B, Xu F, Chen J, et al. Site-specific adaptive nanovesicles for oral insulin delivery. Sci Adv. 2025;11(39):eady6386. doi:10.1126/sciadv.ady6386
22. MMS ZY, Zhang JY, et al. Analysis of modern research characteristics of plant-derived extracellular vesicles based on literature analysis. Pharmacol Clin Chinese Materia Medica. 2025;1–29.
23. Li D, Yao X, Yue J, et al. Advances in bioactivity of MicroRNAs of plant-derived exosome-like nanoparticles and milk-derived extracellular vesicles. J Agricult Food Chem. 2022;70(21):6285–6299. doi:10.1021/acs.jafc.2c00631
24. Ly NP, Han HS, Kim M, Park JH, Choi KY. Plant-derived nanovesicles: current understanding and applications for cancer therapy. Bioactive Mater. 2023;22:365–383. doi:10.1016/j.bioactmat.2022.10.005
25. Movahed N, Cabanillas DG, Wan J, Vali H, Laliberte JF, Zheng H. Turnip mosaic virus components are released into the extracellular space by vesicles in infected leaves. Plant Physiol. 2019;180(3):1375–1388. doi:10.1104/pp.19.00381
26. An Q, Huckelhoven R, Kogel KH, van Bel AJ. Multivesicular bodies participate in a cell wall-associated defence response in barley leaves attacked by the pathogenic powdery mildew fungus. Cell Microbiol. 2006;8(6):1009–1019. doi:10.1111/j.1462-5822.2006.00683.x
27. Cai Q, He B, Jin H. A safe ride in extracellular vesicles - small RNA trafficking between plant hosts and pathogens. Curr Opin Plant Biol. 2019;52:140–148. doi:10.1016/j.pbi.2019.09.001
28. Wu P, Wu W, Zhang S, et al. Therapeutic potential and pharmacological significance of extracellular vesicles derived from traditional medicinal plants. Front Pharmacol. 2023;14:1272241. doi:10.3389/fphar.2023.1272241
29. Wang J, Ding Y, Wang J, et al. EXPO, an exocyst-positive organelle distinct from multivesicular endosomes and autophagosomes, mediates cytosol to cell wall exocytosis in arabidopsis and tobacco cells. Plant Cell. 2010;22(12):4009–4030. doi:10.1105/tpc.110.080697
30. Cai Q, He B, Wang S, et al. Message in a bubble: shuttling small RNAs and proteins between cells and interacting organisms using extracellular vesicles. Annu Rev Plant Biol. 2021;72(1):497–524. doi:10.1146/annurev-arplant-081720-010616
31. Hatsugai N, Iwasaki S, Tamura K, et al. A novel membrane fusion-mediated plant immunity against bacterial pathogens. Genes Dev. 2009;23(21):2496–2506. doi:10.1101/gad.1825209
32. Chen A, He B, Jin H. Isolation of extracellular vesicles from arabidopsis. Curr Protoc. 2022;2(1):e352. doi:10.1002/cpz1.352
33. Lian MQ, Chng WH, Liang J, et al. Plant-derived extracellular vesicles: recent advancements and current challenges on their use for biomedical applications. J Extracell Vesicles. 2022;11(12):e12283. doi:10.1002/jev2.12283
34. Pinedo M, de la Canal L, de Marcos Lousa C. A call for Rigor and standardization in plant extracellular vesicle research. J Extracell Vesicles. 2021;10(6):e12048. doi:10.1002/jev2.12048
35. Cong M, Tan S, Li S, et al. Technology insight: plant-derived vesicles-How far from the clinical biotherapeutics and therapeutic drug carriers? Adv Drug Delivery Rev. 2022;182:114108.
36. Iwai K, Minamisawa T, Suga K, Yajima Y, Shiba K. Isolation of human salivary extracellular vesicles by iodixanol density gradient ultracentrifugation and their characterizations. J Extracell Vesicles. 2016;5(1):30829. doi:10.3402/jev.v5.30829
37. Thery C, Amigorena S, Raposo G, Clayton A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol. 2006;30(1). doi:10.1002/0471143030.cb0322s30
38. Johnstone RM, Bianchini A, Teng K. Reticulocyte maturation and exosome release: transferrin receptor containing exosomes shows multiple plasma membrane functions. Blood. 1989;74(5):1844–1851. doi:10.1182/blood.V74.5.1844.1844
39. Mol EA, Goumans MJ, Doevendans PA, Sluijter JPG, Vader P. Higher functionality of extracellular vesicles isolated using size-exclusion chromatography compared to ultracentrifugation. Nanomedicine. 2017;13(6):2061–2065. doi:10.1016/j.nano.2017.03.011
40. Konoshenko MY, Lekchnov EA, Vlassov AV, Laktionov PP. Isolation of extracellular vesicles: general methodologies and latest trends. Biomed Res Int. 2018;2018:8545347. doi:10.1155/2018/8545347
41. Raimondo S, Giavaresi G, Lorico A, Alessandro R. Extracellular vesicles as biological shuttles for targeted therapies. Int J Molecular Sci. 2019;21(1):20. doi:10.3390/ijms21010020
42. Alfieri M, Leone A, Ambrosone A. Plant-derived nano and microvesicles for human health and therapeutic potential in nanomedicine. Pharmaceutics. 2021;14(1):13. doi:10.3390/pharmaceutics14010013
43. Sidhom K, Obi PO, Saleem A. A review of exosomal isolation methods: is size exclusion chromatography the best option? Int J Mole Sci. 2020;22(1):21. doi:10.3390/ijms22010021
44. Alvarez ML, Khosroheidari M, Kanchi Ravi R, DiStefano JK. Comparison of protein, microRNA, and mRNA yields using different methods of urinary exosome isolation for the discovery of kidney disease biomarkers. Kidney Int. 2012;82(9):1024–1032. doi:10.1038/ki.2012.256
45. Abramowicz A, Widlak P, Pietrowska M. Proteomic analysis of exosomal cargo: the challenge of high purity vesicle isolation. Mol Biosyst. 2016;12(5):1407–1419. doi:10.1039/C6MB00082G
46. Lee R, Ko HJ, Kim K, et al. Anti-melanogenic effects of extracellular vesicles derived from plant leaves and stems in mouse melanoma cells and human healthy skin. J Extracell Vesicles. 2020;9:1703480.
47. Busatto S, Vilanilam G, Ticer T, et al. Tangential flow filtration for highly efficient concentration of extracellular vesicles from large volumes of fluid. Cells. 2018;7.
48. Kim MK, Choi YC, Cho SH, Choi JS, Cho YW. The antioxidant effect of small extracellular vesicles derived from aloe vera peels for wound healing. Tissue Eng Regen Med. 2021;18(4):561–571. doi:10.1007/s13770-021-00367-8
49. Ko M, Kim HJ, Park J, et al. Isolation of bovine milk exosome using electrophoretic oscillation assisted tangential flow filtration with antifouling of micro-ultrafiltration membrane filters. ACS Appl Mater Interfaces. 2023;15(21):26069–26080. doi:10.1021/acsami.3c00446
50. Boing AN, van der Pol E, Grootemaat AE, Coumans FA, Sturk A, Nieuwland R. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J Extracell Vesicles. 2014;3.
51. Baranyai T, Herczeg K, Onodi Z, et al. Isolation of exosomes from blood plasma: qualitative and quantitative comparison of ultracentrifugation and size exclusion chromatography methods. PLoS One. 2015;10(12):e0145686. doi:10.1371/journal.pone.0145686
52. Nordin JZ, Lee Y, Vader P, et al. Ultrafiltration with size-exclusion liquid chromatography for high yield isolation of extracellular vesicles preserving intact biophysical and functional properties. Nanomedicine. 2015;11(4):879–883. doi:10.1016/j.nano.2015.01.003
53. You JY, Kang SJ, Rhee WJ. Isolation of cabbage exosome-like nanovesicles and investigation of their biological activities in human cells. Bioactive Materials. 2021;6(12):4321–4332. doi:10.1016/j.bioactmat.2021.04.023
54. Wang Y, Wang J, Ma J, Zhou Y, Lu R. Focusing on future applications and current challenges of plant derived extracellular vesicles. Pharmaceuticals. 2022;15.
55. Ruysschaert T, Marque A, Duteyrat JL, Lesieur S, Winterhalter M, Fournier D. Liposome retention in size exclusion chromatography. BMC Biotechnol. 2005;5(1):11. doi:10.1186/1472-6750-5-11
56. Kim M, Jang H, Kim W, Kim D, Park JH. Therapeutic Applications of plant-derived extracellular vesicles as antioxidants for oxidative stress-related diseases. Antioxidants. 2023;12.
57. Garcia-Romero N, Madurga R, Rackov G, et al. Polyethylene glycol improves current methods for circulating extracellular vesicle-derived DNA isolation. J Transl Med. 2019;17(1):75. doi:10.1186/s12967-019-1825-3
58. Kalarikkal SP, Prasad D, Kasiappan R, Chaudhari SR, Sundaram GM. A cost-effective polyethylene glycol-based method for the isolation of functional edible nanoparticles from ginger rhizomes. Sci Rep. 2020;10(1):4456. doi:10.1038/s41598-020-61358-8
59. Gamez-Valero A, Monguio-Tortajada M, Carreras-Planella L, Franquesa M, Beyer K, Borras FE. Size-exclusion chromatography-based isolation minimally alters extracellular vesicles’ characteristics compared to precipitating agents. Sci Rep. 2016;6(1):33641. doi:10.1038/srep33641
60. Yu ZL, Liu XC, Wu M, et al. Untouched isolation enables targeted functional analysis of tumour-cell-derived extracellular vesicles from tumour tissues. J Extracell Vesicles. 2022;11(4):e12214. doi:10.1002/jev2.12214
61. Lin B, Lei Y, Wang J, et al. Microfluidic-based exosome analysis for liquid biopsy. Small Methods. 2021;5(3):e2001131. doi:10.1002/smtd.202001131
62. Rutter BD, Rutter KL, Innes RW. Isolation and quantification of plant extracellular vesicles. Bio Protoc. 2017;7(17):e2533. doi:10.21769/BioProtoc.2533
63. Wang JY, Jayaprakasha GK, Patil BS. Untargeted chemometrics evaluation of the effect of juicing technique on phytochemical profiles and antioxidant activities in common vegetables. Acs Food Sci Technol. 2021;1(1):77–87. doi:10.1021/acsfoodscitech.0c00013
64. Mu N, Li J, Zeng L, et al. Plant-derived exosome-like nanovesicles: current progress and prospects. Int J Nanomedicine. 2023;18:4987–5009. doi:10.2147/IJN.S420748
65. Jung MK, Mun JY. Sample preparation and imaging of exosomes by transmission electron microscopy. J Vis Exp. 2018;(131). doi:10.3791/56482-v
66. Szatanek R, Baj-Krzyworzeka M, Zimoch J, Lekka M, Siedlar M, Baran J. The methods of choice for extracellular vesicles (EVs) characterization. Int J Mole Sci. 2017;19(1):18. doi:10.3390/ijms19010018
67. Ridolfi A, Conti L, Brucale M, et al. Particle profiling of EV-lipoprotein mixtures by AFM nanomechanical imaging. J Extracell Vesicles. 2023;12(10):e12349. doi:10.1002/jev2.12349
68. Bachurski D, Schuldner M, Nguyen PH, et al. Extracellular vesicle measurements with nanoparticle tracking analysis - an accuracy and repeatability comparison between NanoSight NS300 and Zetaview. J Extracell Vesicles. 2019;8(1):1596016. doi:10.1080/20013078.2019.1596016
69. Midekessa G, Godakumara K, Ord J, et al. Zeta potential of extracellular vesicles: toward understanding the attributes that determine colloidal stability. ACS omega. 2020;5(27):16701–16710. doi:10.1021/acsomega.0c01582
70. Wang B, Zhuang X, Deng ZB, et al. Targeted drug delivery to intestinal macrophages by bioactive nanovesicles released from grapefruit. Molecular Therapy. 2014;22(3):522–534. doi:10.1038/mt.2013.190
71. Shao M, Jin X, Chen S, Yang N, Feng G. Plant-derived extracellular vesicles -a novel clinical anti-inflammatory drug carrier worthy of investigation. Biomed Pharmacotherap. 2023;169:115904. doi:10.1016/j.biopha.2023.115904
72. Zhang M, Viennois E, Xu C, Merlin D. Plant derived edible nanoparticles as a new therapeutic approach against diseases. Tissue Barriers. 2016;4(2):e1134415. doi:10.1080/21688370.2015.1134415
73. Karamanidou T, Tsouknidas A. Plant-derived extracellular vesicles as therapeutic nanocarriers. Int J Mole Sci. 2021;23(1):191. doi:10.3390/ijms23010191
74. Suharta S, Barlian A, Hidajah AC, et al. Plant-derived exosome-like nanoparticles: a concise review on its extraction methods, content, bioactivities, and potential as functional food ingredient. J Food Sci. 2021;86(7):2838–2850. doi:10.1111/1750-3841.15787
75. Rome S. Biological properties of plant-derived extracellular vesicles. Food Function. 2019;10(2):529–538. doi:10.1039/C8FO02295J
76. Zhou H, Huo Y, Yang N, Wei T. Phosphatidic acid: from biophysical properties to diverse functions. FEBS J. 2023.
77. Treede I, Braun A, Sparla R, et al. Anti-inflammatory effects of phosphatidylcholine. J Biol Chem. 2007;282(37):27155–27164. doi:10.1074/jbc.M704408200
78. Song H, Canup BSB, Ngo VL, Denning TL, Garg P, Laroui H. Internalization of garlic-derived nanovesicles on liver cells is triggered by interaction with CD98. ACS Omega. 2020;5(36):23118–23128. doi:10.1021/acsomega.0c02893
79. Zhang L, Hou D, Chen X, et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNA. Cell Res. 2012;22(1):107–126. doi:10.1038/cr.2011.158
80. Xiao J, Feng S, Wang X, et al. Identification of exosome-like nanoparticle-derived microRNAs from 11 edible fruits and vegetables. PeerJ. 2018;6:e5186. doi:10.7717/peerj.5186
81. Russell W, Duthie G. Plant secondary metabolites and gut health: the case for phenolic acids. Proc Nutr Soc. 2011;70(3):389–396. doi:10.1017/S0029665111000152
82. Berger E, Colosetti P, Jalabert A, et al. Use of nanovesicles from orange juice to reverse diet-induced gut modifications in diet-induced obese mice. Mol Ther Methods Clin Dev. 2020;18:880–892. doi:10.1016/j.omtm.2020.08.009
83. Elsharkasy OM, Nordin JZ, Hagey DW, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Delivery Rev. 2020;159:332–343. doi:10.1016/j.addr.2020.04.004
84. Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJ. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29(4):341–345. doi:10.1038/nbt.1807
85. Schillaci O, Fontana S, Monteleone F, et al. Exosomes from metastatic cancer cells transfer amoeboid phenotype to non-metastatic cells and increase endothelial permeability: their emerging role in tumor heterogeneity. Scientific Reports. 2017;7(1):4711. doi:10.1038/s41598-017-05002-y
86. Wang Q, Zhuang X, Mu J, et al. Delivery of therapeutic agents by nanoparticles made of grapefruit-derived lipids. Nat Commun. 2013;4(1):1867. doi:10.1038/ncomms2886
87. Stanly C, Moubarak M, Fiume I, Turiц║k L, Pocsfalvi G. Membrane transporters in citrus clementina fruit juice-derived nanovesicles. Int J Mole Sci. 2019;21(1):20.
88. Li Z, Wang H, Yin H, Bennett C, Zhang HG, Guo P. Arrowtail RNA for ligand display on ginger exosome-like nanovesicles to systemic deliver siRNA for cancer suppression. Sci Rep. 2018;8(1):14644. doi:10.1038/s41598-018-32953-7
89. Woith E, Guerriero G, Hausman JF, et al. Plant extracellular vesicles and nanovesicles: focus on secondary metabolites, proteins and lipids with perspectives on their potential and sources. Int J Mole Sci. 2021;23(1):22. doi:10.3390/ijms23010022
90. Logozzi M, Di Raimo R, Mizzoni D, Fais S. The potentiality of plant-derived nanovesicles in human health-a comparison with human exosomes and artificial nanoparticles. Int J Mole Sci. 2022;23(9):4919. doi:10.3390/ijms23094919
91. Zhuang X, Teng Y, Samykutty A, et al. Grapefruit-derived nanovectors delivering therapeutic miR17 through an intranasal route inhibit brain tumor progression. Molecular Therapy. 2016;24(1):96–105. doi:10.1038/mt.2015.188
92. Teng Y, Mu J, Hu X, et al. Grapefruit-derived nanovectors deliver miR-18a for treatment of liver metastasis of colon cancer by induction of M1 macrophages. Oncotarget. 2016;7(18):25683–25697. doi:10.18632/oncotarget.8361
93. Niu W, Xiao Q, Wang X, et al. A biomimetic drug delivery system by integrating grapefruit extracellular vesicles and doxorubicin-loaded heparin-based nanoparticles for glioma therapy. Nano Letters. 2021;21(3):1484–1492. doi:10.1021/acs.nanolett.0c04753
94. Stanly C, Alfieri M, Ambrosone A, Leone A, Fiume I, Pocsfalvi G. Grapefruit-derived micro and nanovesicles show distinct metabolome profiles and anticancer activities in the A375 human melanoma cell line. Cells. 2020;10(1):9. doi:10.3390/cells10010009
95. Xiao Q, Zhao W, Wu C, et al. Lemon-derived extracellular vesicles nanodrugs enable to efficiently overcome cancer multidrug resistance by endocytosis-triggered energy dissipation and energy production reduction. Adv Sci. 2022;9(20):e2105274. doi:10.1002/advs.202105274
96. Raimondo S, Naselli F, Fontana S, et al. Citrus limon -derived nanovesicles inhibit cancer cell proliferation and suppress CML xenograft growth by inducing TRAIL-mediated cell death. Oncotarget. 2015;6(23):19514–19527. doi:10.18632/oncotarget.4004
97. Raimondo S, Saieva L, Cristaldi M, Monteleone F, Fontana S, Alessandro R. Label-free quantitative proteomic profiling of colon cancer cells identifies acetyl-CoA carboxylase alpha as antitumor target of citrus limon-derived nanovesicles. J Proteomics. 2018;173:1–11. doi:10.1016/j.jprot.2017.11.017
98. Yang M, Liu X, Luo Q, Xu L, Chen F. An efficient method to isolate lemon derived extracellular vesicles for gastric cancer therapy. J Nanobiotechnol. 2020;18(1):100. doi:10.1186/s12951-020-00656-9
99. Kazemi M, Jafarzadeh A, Nemati M, et al. Zingerone improves the immune responses in an animal model of breast cancer. J Complement Integr Med. 2021;18(2):303–310. doi:10.1515/jcim-2019-0135
100. Chen CY, Chen YN, Shiau JP, Tang JY, Hou MF, Chang HW. Ginger-derived 3HDT exerts antiproliferative effects on breast cancer cells by apoptosis and DNA damage. Int J Mole Sci. 2023;25(1):24. doi:10.3390/ijms25010024
101. Zhang M, Viennois E, Prasad M, et al. Edible ginger-derived nanoparticles: a novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials. 2016;101:321–340. doi:10.1016/j.biomaterials.2016.06.018
102. Zhang M, Xiao B, Wang H, et al. Edible ginger-derived nano-lipids loaded with doxorubicin as a novel drug-delivery approach for colon cancer therapy. Molecular Therapy. 2016;24(10):1783–1796. doi:10.1038/mt.2016.159
103. Zhang M, Yang C, Yan X, Sung J, Garg P, Merlin D. Highly biocompatible functionalized layer-by-layer ginger lipid nano vectors targeting p-selectin for delivery of doxorubicin to treat colon cancer. Adv Ther. 2019;2.
104. Long D, Alghoul Z, Sung J, Yang C, Merlin D. Oral administration of M13-loaded nanoliposomes is safe and effective to treat colitis-associated cancer in mice. Expert Opin Drug Deliv. 2023;20(10):1443–1462. doi:10.1080/17425247.2023.2231345
105. Cao M, Yan H, Han X, et al. Ginseng-derived nanoparticles alter macrophage polarization to inhibit melanoma growth. J Immunotherap Cancer. 2019;7(1):326. doi:10.1186/s40425-019-0817-4
106. Han X, Wei Q, Lv Y, et al. Ginseng-derived nanoparticles potentiate immune checkpoint antibody efficacy by reprogramming the cold tumor microenvironment. Molecular Therapy. 2022;30(1):327–340. doi:10.1016/j.ymthe.2021.08.028
107. Lv Y, Li M, Weng L, et al. Ginseng-derived nanoparticles reprogram macrophages to regulate arginase-1 release for ameliorating T cell exhaustion in tumor microenvironment. J Exp Clin Cancer Res. 2023;42(1):322. doi:10.1186/s13046-023-02888-7
108. Yang L, Jin WQ, Tang XL, et al. Ginseng-derived nanoparticles inhibit lung cancer cell epithelial mesenchymal transition by repressing pentose phosphate pathway activity. Front Oncol. 2022;12:942020. doi:10.3389/fonc.2022.942020
109. Kim J, Zhu Y, Chen S, et al. Anti-glioma effect of ginseng-derived exosomes-like nanoparticles by active blood-brain-barrier penetration and tumor microenvironment modulation. J nanobiotechnol. 2023;21(1):253. doi:10.1186/s12951-023-02006-x
110. Wang H, Mu J, Chen Y, et al. Hybrid ginseng-derived extracellular vesicles-like particles with autologous tumor cell membrane for personalized vaccination to inhibit tumor recurrence and metastasis. Adv Sci. 2024;11(17):e2308235. doi:10.1002/advs.202308235
111. Bin Wang X-JG, Cai H, Zhu Y-H. Momordica charantia-derived extracellular vesicles-like nanovesicles inhibited glioma proliferation, migration, and invasion by regulating the PI3K/AKT signaling pathway. Functional Foods. 2022;90.
112. Yang M, Luo Q, Chen X, Chen F. Bitter melon derived extracellular vesicles enhance the therapeutic effects and reduce the drug resistance of 5-fluorouracil on oral squamous cell carcinoma. J Nanobiotechnol. 2021;19(1):259. doi:10.1186/s12951-021-00995-1
113. Özkan PK İ, Yıldırım M, Ünsal N. Hazal Yılmaz, Dilek Telci, Fikrettin Şahin Garlic (Allium sativum)-derived SEVs inhibit cancer cell proliferation and induce caspase mediated apoptosis. Scientific Reports. 2021;11(1):14773. doi:10.1038/s41598-021-93876-4
114. Chen Q, Li Q, Liang Y, et al. Natural exosome-like nanovesicles from edible tea flowers suppress metastatic breast cancer via ROS generation and microbiota modulation. Acta Pharmaceutica Sinica B. 2022;12(2):907–923. doi:10.1016/j.apsb.2021.08.016
115. Chen Q, Zu M, Gong H, et al. Tea leaf-derived exosome-like nanotherapeutics retard breast tumor growth by pro-apoptosis and microbiota modulation. J Nanobiotechnol. 2023;21(1):6. doi:10.1186/s12951-022-01755-5
116. Tajik T, Baghaei K, Moghadam VE, Farrokhi N, Salami SA. Extracellular vesicles of cannabis with high CBD content induce anticancer signaling in human hepatocellular carcinoma. Biomed Pharmacotherap. 2022;152:113209. doi:10.1016/j.biopha.2022.113209
117. Zhang L, He F, Gao L, et al. Engineering exosome-like nanovesicles derived from asparagus cochinchinensis can inhibit the proliferation of hepatocellular carcinoma cells with better safety profile. Int J Nanomedicine. 2021;16:1575–1586. doi:10.2147/IJN.S293067
118. Liu J, Xiang J, Jin C, et al. Medicinal plant-derived mtDNA via nanovesicles induces the cGAS-STING pathway to remold tumor-associated macrophages for tumor regression. J Nanobiotechnol. 2023;21(1):78. doi:10.1186/s12951-023-01835-0
119. Gao Q, Chen N, Li B, et al. Natural lipid nanoparticles extracted from Morus nigra L. leaves for targeted treatment of hepatocellular carcinoma via the oral route. J Nanobiotechnol. 2024;22(1):4. doi:10.1186/s12951-023-02286-3
120. Yan G, Xiao Q, Zhao J, et al. Brucea javanica derived exosome-like nanovesicles deliver miRNAs for cancer therapy. J Controlled Release. 2024;367:425–440. doi:10.1016/j.jconrel.2024.01.060
121. Chi Y, Shi L, Lu S, et al. Inhibitory effect of Lonicera japonica-derived exosomal miR2911 on human papilloma virus. J Ethnopharmacol. 2024;318:116969. doi:10.1016/j.jep.2023.116969
122. Mashhadi NS, Ghiasvand R, Askari G, Hariri M, Darvishi L, Mofid MR. Anti-oxidative and anti-inflammatory effects of ginger in health and physical activity: review of current evidence. Int J Prev Med. 2013;4(Suppl 1):S36–42.
123. Shukla Y, Singh M. Cancer preventive properties of ginger: a brief review. Food Chem Toxicol. 2007;45(5):683–690. doi:10.1016/j.fct.2006.11.002
124. Kundu JK, Na HK, Surh YJ. Ginger-derived phenolic substances with cancer preventive and therapeutic potential. Forum Nutr. 2009;61:182–192.
125. Yue PY, Mak NK, Cheng YK, et al. Pharmacogenomics and the Yin/Yang actions of ginseng: anti-tumor, angiomodulating and steroid-like activities of ginsenosides. Chin Med. 2007;2(1):6. doi:10.1186/1749-8546-2-6
126. Sun X, Zhao YN, Qian S, et al. Ginseng-derived panaxadiol saponins promote hematopoiesis recovery in cyclophosphamide-induced myelosuppressive mice: potential novel treatment of chemotherapy-induced cytopenias. Chin J Integr Med. 2018;24(3):200–206. doi:10.1007/s11655-017-2754-8
127. Hwang SH, Lee BH, Kim HJ, et al. Suppression of metastasis of intravenously-inoculated B16/F10 melanoma cells by the novel ginseng-derived ingredient, gintonin: involvement of autotaxin inhibition. Int J Oncol. 2013;42(1):317–326. doi:10.3892/ijo.2012.1709
128. Cathcart P, Stebbing J. Aloe vera, a natural cancer soother? Lancet Oncol. 2016;17(4):421. doi:10.1016/S1470-2045(16)00161-3
129. Li KW, Liang YY, Wang Q, et al. Brucea javanica: a review on anticancer of its pharmacological properties and clinical researches. Phytomedicine. 2021;86:153560. doi:10.1016/j.phymed.2021.153560
130. Liu XC, Tian JW, Xu JY, et al. Extracellular vesicle manufacture via FACTORY: fully automated collection technology and optimum machinery for clinical translational applications. Trends Biotechnol. 2025;43(11):2947–2969. doi:10.1016/j.tibtech.2025.06.020
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.