Back to Journals » Drug Design, Development and Therapy » Volume 20
Fermentation-Induced Changes in Phytochemical Composition and Pharmacological Activities of Zingiberaceae Plants: Insight from in vitro and in vivo Studies
Authors Sammulia SF, Suhaera S, Prayoga DK, Pitriani P, Ramadhania ZM ![]() , Fristiohady A, Milanda T
, Fristiohady A, Milanda T
Received 17 February 2026
Accepted for publication 9 April 2026
Published 21 April 2026 Volume 2026:20 604038
DOI https://doi.org/10.2147/DDDT.S604038
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Suci Fitriani Sammulia,1– 3,* Suhaera Suhaera,1,3,* Deshanda Kurniawan Prayoga,1,4,* Pitriani Pitriani,5,* Zelika Mega Ramadhania,3,* Adryan Fristiohady,6,* Tiana Milanda3,*
1Doctoral Program in Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 2Department of Pharmacy, Faculty of Health Sciences, Institut Kesehatan Mitra Bunda, Batam, Riau Islands, Indonesia; 3Department of Biological Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, Indonesia; 4Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 5Departement of Pharmaceutics and Pharmaceutical Technology, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 6Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Halu Oleo, Kendari, Southeast Sulawesi, Indonesia
*These authors contributed equally to this work
Correspondence: Tiana Milanda, Department of Biological Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, West Java, 46363, Indonesia, Tel +6222-84288888 ext 3510, Email [email protected]
Abstract: The Zingiberaceae family has long been used in traditional medicine due to its rich array of secondary metabolites. However, its low bioavailability, limited stability in its native form, degradation during digestion, and poor solubility in water all restrict its absorption in the human body. Fermentation represents an effective biotechnological method for modifying the phytochemical composition and potentially enhancing its pharmacological effects. This study aims to explore the impact of fermentation on Zingiberaceae, focusing on the alteration of phytochemical profiles and the enhancement of pharmacological activities. Articles were sourced from the Scopus and PubMed databases and filtered for publications between 2015 and 2025; there were 2 articles that were electronically removed before screening due to duplication, yielding 62 articles. These articles were then further screened based on titles, abstracts, and full texts, resulting in five relevant studies. Fermentation was found to improve the phytochemical profile, influenced by the microbial strains used and the physicochemical properties of the phytochemicals. The fermentation process enhanced the stability of compounds, such as converting 6-gingerol to 6-shogaol and transforming glycosides into aglycones, which are more easily absorbed by the body. Additionally, fermentation increased phenolic and flavonoid content, accompanied by enhanced antioxidant and anti-inflammatory activities. Pharmacologically, in vitro studies showed that fermented extracts modulate cytokine signaling pathways in immune cells while enhancing anti-aging properties and skin barrier protection. Meanwhile, in vivo studies demonstrated improvements in metabolic regulation and neuroprotective effects in cognitive disorders. Further mechanistic investigations are needed to clarify the pathways through which fermentation influences the behavior of phytoconstituents and their pharmacological performance. This review provides an overview of preclinical fermentation studies on Zingiberaceae plants, both in vitro and in vivo, with a focus on their phytochemical composition and effectiveness in enhancing pharmacological activity.
Keywords: fermentation, fermented plant-based, phytochemicals, pharmacology activity, Zingiberaceae
Introduction
The use of medicinal plants for the management of the various diseases has been practiced for centuries and is widely known as phytotherapy.1 The therapeutic potential of medicinal plants is primarily attributed to their rich content of secondary metabolites, commonly referred to as phytochemicals, which are naturally synthesized and continuously replenished in plant tissues.2–5 These bioactive compounds were abundantly present in different plant parts, including seeds, barks, roots, leaves, flowers, and fruits. In plants, phytoconstituents also function as protective agents against environmental stressors and pathogenic microorganisms.5,6
Among medicinal plant groups, the Zingiberaceae family is recognized as one of the most important sources of pharmacologically active compounds. Zingiberaceae is a monocotyledonous plant family predominantly distributed across South and Southeast Asia, including Indonesia, Singapore, Malaysia, comprising approximately 53 genera and more than 1300 species.7,8 Some well-known plants in the Zingiberaceae family, such as Zingiber officinale, Alpinia galanga, Curcuma longa, Kaempferia galanga, Curcuma xanthorrhiza, and Amomum villosum, have potential pharmacological effects. Plants of this family are extensively used in the food industry, particularly as flavoring agents and spices in culinary applications. Beyond their role in food, Zingiberaceae plants have attracted considerable attention due to their medicinal properties.9–11 Numerous studies have reported their diverse biological activities, including antioxidant,12–14 anti-inflammatory,15–17 anti-proliferative,18,19 antiplatelet,20,21 anti-ulcer,22,23 anticonvulsive,24,25 and analgesic effects.26 As a result, these plants are widely incorporated into traditional medicine systems as well as pharmaceutical products. Currently, there have been several studies in comprehensive metabolomics profiling of the chemical composition and metabolite variability of Zingiberaceae plants. For instance, metabolite profiling studies across multiple Zingiberaceae species have identified a broad spectrum of compounds, including curcumin, gingerol, galangin, organic acids, flavonoids, and diarylheptanoids.27–29
Although Zingiberaceae plants exhibit considerable pharmacological potential, the biological effectiveness of their phytoconstituents is constrained by several inherent limitations. These include low bioavailability and limited stability in their native forms, degradation during digestion, and poor aqueous solubility, all of which restrict efficient absorption in the human body.30–32 Many key bioactive compounds from Zingiberaceae, such as curcumin and 6-gingerol, display poor water solubility and are susceptible to degradation when exposed to heat, oxygen, or light.33–35 In addition, conventional extraction techniques frequently result in partial degradation or incomplete liberation of bioactive compounds from plant cell walls.36–38 Even when successfully extracted, these compounds often exhibit low intestinal absorption and minimal systemic availability. For example, the oral bioavailability of curcumin in humans has been reported to be less than 1%.39–42 These limitations highlight the need for processing strategies that can modify phytochemical composition while preserving or enhancing biological activity.
Fermentation represents a viable approach to addressing these limitations; it is a biotechnological process that employs microorganisms classified as Generally Recognized as Safe (GRAS) to modify the chemical structure of natural compounds into forms with enhanced biological activity, thereby potentially improving pharmacological effects.39–42 Fermentation processes in plants can enhance the pharmacological activity of bioactive compounds through enzymatic mechanisms involving β-glucosidase. This enzyme plays a critical role in hydrolyzing glycoside bonds between sugar moieties and aglycones in plant glycosides. During fermentation, microorganisms such as Aspergillus spp., Saccharomyces spp., and lactic acid bacteria produce β-glucosidase, which catalyzes the conversion of glycosides into free aglycones and simple sugars. This biotransformation leads to an increase in the lipophilicity of the resulting compounds, thereby facilitating their cellular uptake and improving their bioavailability. Consequently, the release of aglycone forms is associated with enhanced pharmacological activities, including antioxidant, antimicrobial, anti-inflammatory, and immunomodulatory effects. Through microbial and enzymatic transformations, fermentation can overcome key challenges in the development of plant-based therapeutics, particularly by converting glycosylated compounds with low bioavailability and poor absorption into aglycone forms that are more readily absorbed by the human body.43–45 In addition, fermentation has been shown to enhance the absorption and systemic distribution of bioactive compounds. Evidence supporting these effects has been reported for members of the Zingiberaceae family. For example, fermentation of Curcuma longa using Lactobacillus rhamnosus FN7 resulted in a significant increasing in phenolic and flavonoid contents, accompanied by enhanced antioxidant and anti-inflammatory activities.46–48 Although research on fermented Zingiberaceae has increased in recent years, existing reviews largely emphasize fermentation techniques and microbial strain selection, with limited integration of phytoconstituents changes and functional bioactivity outcomes.
Taking this all into consideration, this review aims to provide an examine the effects of fermentation on phytochemical composition and associated pharmacological activities of plants belonging to the Zingiberaceae family. The review synthesizes current evidence on fermentation-driven qualitative and quantitiative changes in bioactive compounds and evaluates how these compositional modifications are associated with biological compounds based on preclinical evidence, in vitro and in vivo studies. Overall, this review provides a structured and critical synthesis of existing findings and highlight research gaps relevant to the development of fermented Zingiberaceae-based plants, with particular emphasis on underlying mechanisms, phytoconstituent alterations, cellular-level evaluations using cell lines or assay kits, and pharmacological assesments in animal models.
Methods
Search Strategy
The search strategy was initially concentrated on articles published between 2015 and 2025. References were obtained from Scopus and PubMed Databases using keywords such as “Zingiberaceae,” “Fermentation,” and “Pharmacological Activity.” The related terms and alternative phrases pertinent to the research topics were also included. Throughout the search process, synonyms and related terminology were employed interchangeably. Boolean operators like “AND” and “OR” were utilized to refine the search results, facilitating the inclusion or exclusion of specific terms. The search continued until August 2025, following a combination of medical subject headings (MeSH terms). The documents selected for review adhered to the inclusion criteria, focusing specifically on the fermentation of Zingiberaceae and its enhancement of pharmacological activity, rather than on general biological effects. In total, 5 articles were chosen for in-depth examination.
Data Selection and Collection Process
The study selection process was conducted systematically in accordance with the PRISMA 2020 guidelines to ensure transparency and reproducibility. Initially, a total of 64 records were identified through a comprehensive search of electronic database (Pubmed and Scopus). After removing 2 duplicate records, 62 articles remained for the initial screening stage. During the initial screening stage, titles and abstracts were screened using the Rayyan.ai (https://new.rayyan.ai/) to identify potentially relevant articles. Studies that met the initial criteria proceeded to full-text evaluation.
To be eligible, articles needed to be reported about the fermentation methods and their influence in phytoconstituents, with a specific focus on fermentation methods, including the microorganisms employed, fermentation conditions, and quantitative analysis of phytoconstituents, as well as report preclinical findings derived from cell-line experiments, assay-based evaluations, and animal models was considered. Articles were excluded if they were reviews, commentaries, or did not primarily focus on the specific species of interest. To evaluate the quality of the included studies and address variations in evidence strength, a structured quality assessment was performed. Studies were evaluated based on the transparency of their fermentation parameters. Data from the selected studies were manually retrieved and systematically extracted, covering a variety of study designs. These included analyses of phytoconstituent profiles based on these microorganisms used and fermentation conditions, with comparisons of phytochemical levels before and after fermentation. For in vitro investigations, extracted information included the type of cell lines employed, induction methods, and exposure duration. For in vivo studies, relevant data such as animal models, induction protocols, use of negative and positive controls, and study duration in weeks were recorded. Based on these inclusion criteria, a total of 5 articles were selected for detailed analysis (Figure 1).
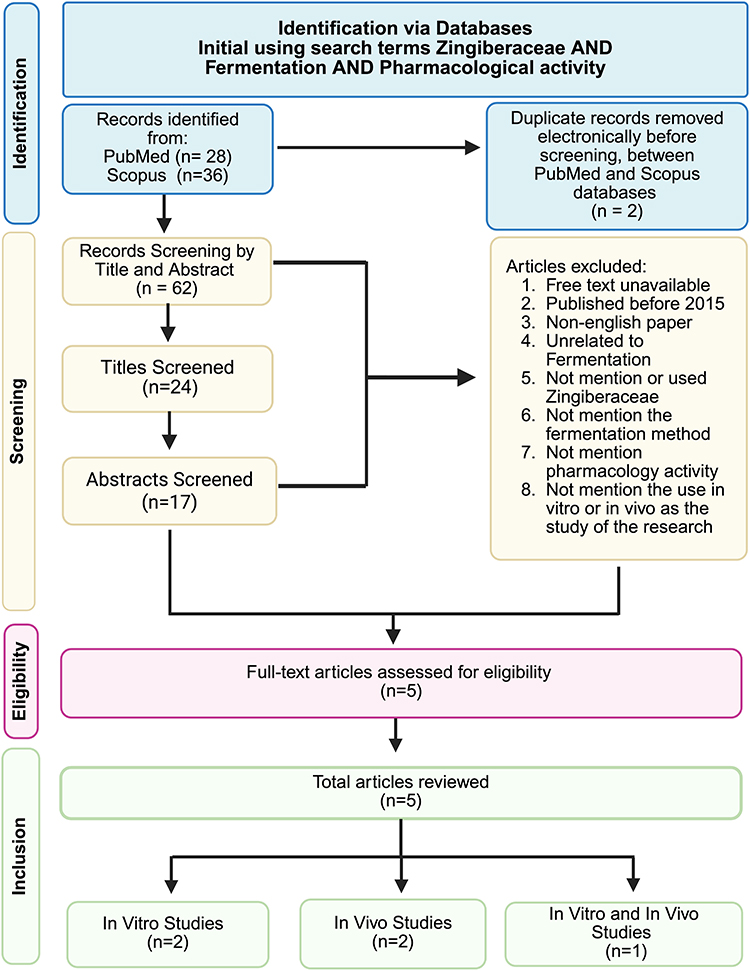 |
Figure 1 The study design of the article review. |
Data Synthesis
Given the considerable variability across the studies and the differing outcome measures, a systematic synthesis of the evidence was performed. The findings were summarized qualitatively and organized thematically based on the reported results. To provide comprehensive context and address any gaps in the literature, this synthesis includes detailed sections on the species-specificity of microbial species that modulate phytoconstituents during fermentation. This thorough approach serves as a valuable resource for researchers, integrating biological context with an extensive review of the latest evidence.
Characteristics of Included Studies
The literature search conducted across two selected databases identified a total of 64 records. Duplicate records (n=2) were removed electronically, leaving 62 articles for screening. Titles and abstracts were screened for relevance using Rayyan.ai, resulting in 17 articles selected for full-text assessment. After detailed assessment, 5 studies met the eligibility criteria and were included in the final analysis. The study selection workflow is illustrated in Figure 1, which outlines the stages of identification, screening, eligibility, assessment, and final inclusion in accordance with the Standard PRISMA-based selection process. Key characteristics of included studies were systematically extracted, with particular attention to fermented plant-based interventions and their mechanisms influencing the upregulation or downregulation of phytoconstituent levels. The selected studies were categorized into two main groups, in vitro and in vivo investigations, to allow structured evaluation of evidence across experimental and translational contexts. For each study, descriptive information was recorded, including study design, biological model or population, intervention and induction methods, primary outcomes assessed. These outcomes included biomarkers such as pro-inflammatory gene expression levels, including Interleukin-6 (IL-6), Tumor Necrosis Factor–alpha (TNF-α). Additional literature was consulted to support contextual interpretation and discussion, particularly regarding mechanisms affecting phytoconstituent levels and associated pharmacological enhancement. These supplementary sources were used solely for contextual support and were not included in the PRISMA flow diagram.
Results
Fermentation functions as an effective bioprocess capable of modifying the phytochemical composition of medicinal plants, particularly those belonging to the Zingiberaceae family, with consequent effects on their pharmacological activity. The collective evidence analyzed indicates that fermentation using specific microorganisms produce both qualitative and quantitative alterations in phytoconstituents of Zingiberaceae plants. These compositional changes are consistently associated with measurable effects on bioactivity, bioavailability, and therapeutic applicability, as observed across in vitro and in vivo experimental models. However, the direction of these changes varies considerable depending on the plant species, the microbial strains employed, and the specific fermentation conditions applied, highlighting the importance of controlled and species fermentation strategies. Across the literature studies, 5 articles were included. However, 2 article discussed in vitro studies, 2 articles discussed in vivo studies, and 1 article discussed both preclinical studies, including in vitro and in vivo. Across the included studies, fermentation was found to modulate phytoconstituent levels in Zingiberaceae plants, resulting in both upregulation and downregulation-specific compounds while maintaining or enhancing anti-inflammatory pharmacological activity.
Fermentation-Induced Modulation of Phytoconstituents in Zingiberaceae Plants
Fermentation is a biotechnological process used to modify the content and structure of bioactive compounds in natural materials, including flavonoids. This process involves the use of specific microorganisms, which enhances the bioavailability and pharmacological activity of flavonoid compounds.49–52 Fermentation can convert glycosides into aglycones, which possess higher bioavailability. Several studies have demonstrated that flavonoid compounds are more stable and bioavailable after fermentation compared to their pre-fermentation state.53–55
Fermentation can quantitatively alter the phytoconstituents in the Zingiberaceae family, influenced by the type of fermentation and the microorganisms used during fermentation, as well as the fermentation conditions, such as temperature, pH, duration, and media composition.53,56,57 Solid-state fermentation (SSF) using Lactobacillus plantarum (KCTC 3108, FLP8) on Zingiberaceae officinale showed that 6-gingerol and 6-shogaol remained unchanged after 48 hours of fermentation, indicating that both compounds are stable under these fermentation conditions. However, when a different strain of Lactobacillus plantarum (KCL005, FLP9) was used with a shorter fermentation duration of 24 hours, a significant increase in 6-gingerol and 6-shogaol was observed before and after fermentation.58–61 This could be attributed to SSF providing a controlled enzymatic environment with low air activity, leading to the degradation of sensitive phenolic compounds while supporting partial biotransformation reactions that can modify the structure of specific compounds without damaging their core molecular structure.62–64 The difference in strains has a greater impact on biotransformation compared to the plant species itself. Therefore, it can be concluded that SSF is suitable for modifying target bioactive compounds while maintaining the stability of their core structure, thereby yielding more effective and safer compounds for clinical applications.65
Other studies have demonstrated that Curcuma longa, containing the bioactive compound curcumin, exhibits antioxidant and anti-inflammatory activities. This study utilized yeast extract and ethanol for fermentation at 37°C for 24 hours, after which the curcumin content in the fermented Curcuma longa was measured.65 The curcumin content after fermentation showed a decrease, which may be due to the degradation of curcumin during fermentation. This suggests that microbial activity can break down the compound into other bioactive metabolites, a phenomenon also observed with the compound demethoxycurcumin.66 However, no significant changes were observed in bisdemethoxycurcumin after fermentation, indicating that this compound is stable during the fermentation process. The fermentation in this study also released compounds bound to the cellular matrix, as well as non-chromophoric precursors, which, initially undetectable by analytical methods, underwent changes that allowed them to be detected using more sensitive analyses.66 Therefore, it can be concluded that fermentation can facilitate chemical changes and enhance the bioactivity of compounds in Curcuma longa, thereby strengthening its therapeutic potential and making it suitable for drug development applications (Figure 2).
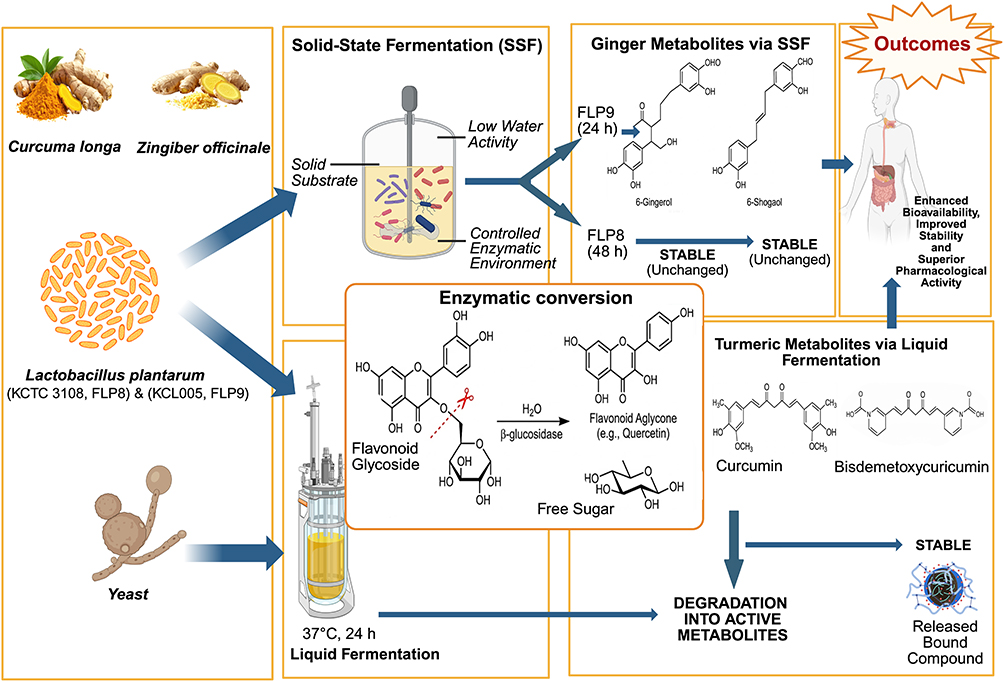 |
Figure 2 Mechanisms of fermentation-induced phytochemical transformation in Zingiberaceae plants. Notes: Aglycone: the non-sugar compound remaining after replacement of the glycosyl group from a glycoside.6-shogaol: a potent dehydrated derivative of 6-gingerol produced during fermentation. FLP8, Lactobacillus plantarum strain KCTC 3108. FLP9, Lactobacillus plantarum strain KCL005. |
Overall, the reviewed evidence demonstrates that fermentation acts as a decisive modulator of phytoconstituents profiles in Zingiberaceae plants by altering both compound abundance and structural form (Table 1). Although the underlying molecular mechanisms were not directly investigated in most studies, the observed-changes in phytochemical profiles and biological activity suggest that fermentation-related biotransformation plays an important role in improving the functional properties of Zingiberaceae-derived compounds. Further mechanistic investigations are required to clarify the pathways through which fermentation influences phytoconstituent behavior and pharmacological performance.
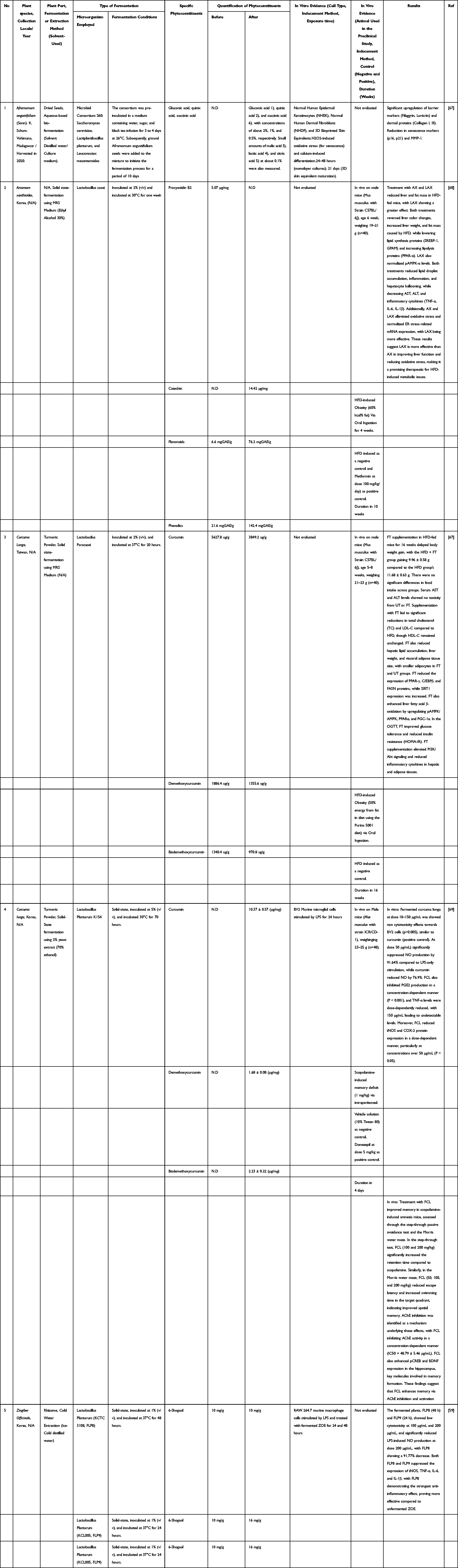 |
Table 1 Phytochemical Profile and Biological Potential of Fermented Medicinal Plants |
Species-Specific Microbial Modulation of Phytoconstituents
The fermentation of plants within the Zingiberaceae family is increasingly recognized as a biochemical process governed by microbial species-specificity. Each microbial species introduces a distinct enzymatic and metabolic capacity that shapes the conversion, upregulation and stabilization of plant-derived bioactive compounds.70–72 These phytoconstituents, primarily curcuminoids, gingerols, shogaols, and phenolic derivatives, undergo targeted transformations depending on the metabolic repertoire of the fermenting microbe, resulting in enhanced bioactivity and functional diversity.
Studies combining microbiome and metabolomic analyses reveal that microbial species interact synergistically to drive phytochemical enhancement during fermentation. For example, Bacillus coagulans and lactic acid bacteria such as Leuconostoc and Pediococcus were shown to upregulate β-glucosidase activity, hydrolyzing glycosidic bonds in flavonoid conjugates and thereby increasing free aglycone levels such as quercetin and caffeic acid.73–75 This enzymatic hydrolysis not only liberates bound phenolics but also enhances their solubility and antioxidant capacity, contributing to the improved biological activity observed in fermented Zingiberaceae extracts. Concurrently, fungal species such as Rhizopus spp. and Aspergillus spp. were consistently detected in fermentation systems exhibiting extensive substrate degradation. These fungi produced amylolytic and proteolytic enzymes that preceded bacterial metabolism and coincided with increased bioavailaibility of low-molecular-weight metabolites.64,76–78
Microbial species also differed in the metabolite classes enriched during fermentation, Lactobacillus plantarum was associated with increased amino acid and polyphenol levels, whereas Lactobacillus buchneri was linked to elevated volatile fatty acid production.79–82 In addition, fermentation using Lactobacillus brevis and Weissela cibaria was associated with selective release of bound ferulic acid and lignans.83–85 Overall, microbial species identity influenced both the direction and magnitude of phytochemical modulation during Zingiberaceae fermentation, resulting in distinct bioactive compound profile across fermentation systems.
In vitro Evidence
In vitro experimental studies commonly employ various cell-line models to investigate fermented natural products, including the murine macrophage cell line RAW 264.7, the murine microglial cell line BV-2, microbial consortia, and skin models such as Normal Human Epidermal Keratinocytes (NHEK), Normal Human Dermal Fibroblasts (NHDF), and 3D bioprinted skin equivalents.58,69,86,87 These cell-line models are typically stimulated with lipopolysaccharide (LPS) to activate inflammatory responses, including the induction of inducible nitric oxide synthase (iNOS), tumor necrosis factor (TNF-α), interleukin (IL-6), and IL-1β.67,88–90 In contrast, biological models of skin protection are often subjected to oxidative stress (H2O2 exposure and calcium-induced differentiation) to simulate cellular aging/senescence and disruption of the skin barrier.87,91,92
Recent studies on fermentation have demonstrated its potential to enhance the pharmacological activity of phytoconstituents compared with their non-fermented counterparts.93,94 Kim et al58 evaluated the anti-inflammatory activity of Zingiber officinale extract using LPS-stimulated murine RAW 264.7 macrophages. Fermented Zingiber officinale with Lactobacillus plantarum KCTC 3108 significantly reduced nitric oxide production by 91.77%, to 0.87 ± 0.12 µM, and fermentation with L. plantarum KCL005 resulted in a greater suppression of nitric oxide compared with the non-fermented extract. Gene-expression analyses further revealed that fermented Zingiber officinale markedly downregulated iNOS, TNF-α, IL-6, and IL-1β, while simultaneously increasing the concentrations of bioactive compounds, namely 6-gingerol and 6-shogaol. These findings are consistent with those reported by Eun et al,86 in which fermented Curcuma longa exhibited superior neuroprotective activity compared with curcumin controls by suppressing NO production, inhibiting PGE2 synthesis, and reducing iNOS and COX-2 expression in a dose-dependent manner Beyond its anti-inflammatory effects, fermented Aframomum angustifolium extract demonstrated significant tissue-repair mechanisms in skin cell models, showing substantial upregulation of key skin-barrier markers (filaggrin and loricrin), increased synthesis of dermal matrix proteins (collagen types I and III), and mitigation of aging through reductions in senescence markers (p16, p21) and Matrix Metalloproteinase-1 (MMP-1)87 (Figure 3).
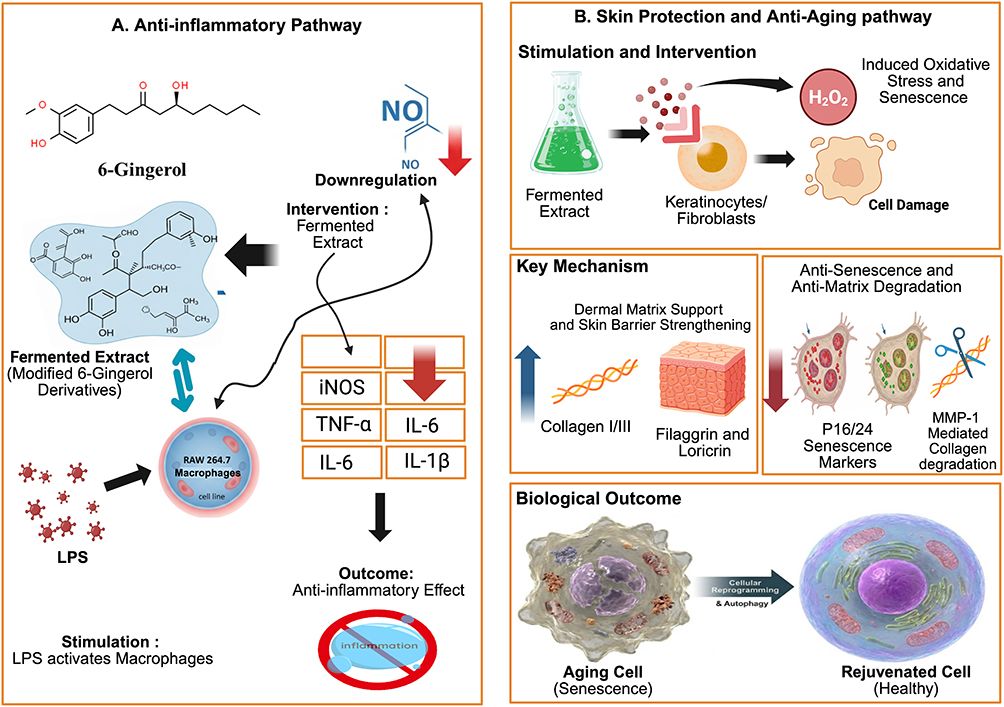 |
Figure 3 In vitro pharmacological mechanisms of fermented Zingiberaceae extracts. (A) Anti-inflammatory In LPS-stimulated RAW 264.7. (B) Skin protection and anti-aging pathway in Keratinocytes/Fibroblasts. Abbreviations: LPS, lipopolysaccharide; NO, nitric oxide; iNOS, inducible nitric oxide synthase; TNF-α, tumor necrosis factor-alpha; IL, interleukin; H2O2, hydrogen peroxide; MMP-1, matrix metalloproteinase-1. Notes: Blue upward arrows (↑) indicate stimulation, Red downward arrows (↓) indicate reductions or downregulation of inflammatory/aging markers. |
Overall, fermentation of plant extracts consistently enhances the pharmacological performance of phytoconstituents by modulating cytokine-signaling pathways in immune cells, while also improving anti-aging properties and skin-barrier protection. These observations indicate that the data presented in Table 1 provide a clear overview of in vitro studies demonstrating that fermented extracts modulate oxidative stress and inflammatory pathways, regulate nitric oxide signaling, and thus highlight their potential for the development of therapeutic agents for inflammation management and cosmetic applications.
In vivo Evidence
In vivo evidence indicates that fermentation consistently enhanced the pharmacological effects of Zingiberaceae phytoconstituents in metabolic, inflammatory, and cognitive impairment models. Fermented plant preparations suppressed pro-inflammatory gene expression and reduced oxidative stress in high-fat diet-induced models, while also improving cognitive performance in scopolamine-induced amnesia mice.86,95,96 These findings indicate that fermentation-associated phytochemical modifications are functionally relevant in vivo, resulting in measurable biological outcomes.
Several studies demonstrated that microbial fermentation improved metabolic regulation by modulating lipid metabolism-related pathways. In high-fat diet-induced obese mice, fermented turmeric prepared using Lactobacillus paracasei showed greater efficacy than their unfermented counterparts, as evidenced by reduced expression of adipogenesis and lipogenesis markers such as Sterol Regulatory Element-Binding Protein 1c (SREBP-1c) and Fatty Acid Synthase (FASN), alongside increased Sirtuin 1 (SIRT1) expression in visceral adipose tissue and liver.95–97 In parallel, fermented preparations enhanced hepatic fatty acid β-oxidation, reflected by increased levels of phosphorylated AMPK (pAMPK)/ AMP-activated protein kinase (AMPK), Peroxisome Proliferator Activated Receptor-Alpha (PPARα), and PPAR Gamma Coactivator-1 alpha (PGC-1α), suggesting protective effects against hepatic steatosis.98–100 These effects were associated with activation of AMPK signaling and suppression of adipocyte differentiation-related transcription factors. Specific phytoconstituents generated or enriched during fermentation, including Calebin-A and bisacurone, were implicated in regulating lipogenic protein expression and promoting phosphorylation pathways linked to metabolic improvement.101–104
Furthermore, fermentation effects in cognitive impairment models also demonstrated neuroprotective effects. In scopolamine-induced amnesia mice, fermented extracts significantly increased hippocampal expression of phosphorylated CREB (pCREB) and Brain Derived Neurotropic Factor (BDNF), key molecular markers involved in memory formation.86,105,106 pCREB and BDNF is a crucia; transcription factor that turns on specific genes involved in memory, learning, and cell survival by being activated by signals like Cyclic adenosine monophosphate (cAMP), calcium, and stress. Its presence indicates active gene expression, and its levels are studies in relation to brain disorders where it’s often reduced in Alzheimer's disease.107,108 Furthermore, Immunohistochemical and Western blot analyses revealed that fermented treatments reversed scopolamine-induced reduction in pCREB and BDNF-positive cells in the hippocampus. Behavioral assessments, including step-through passive avoidance and Morris water maze tests, further confirmed that fermented plant treatments significantly improved memory retention compared with scopolamine-treated controls and showed comparable effects to donezepil used as a positive control (Figure 4). 107,108
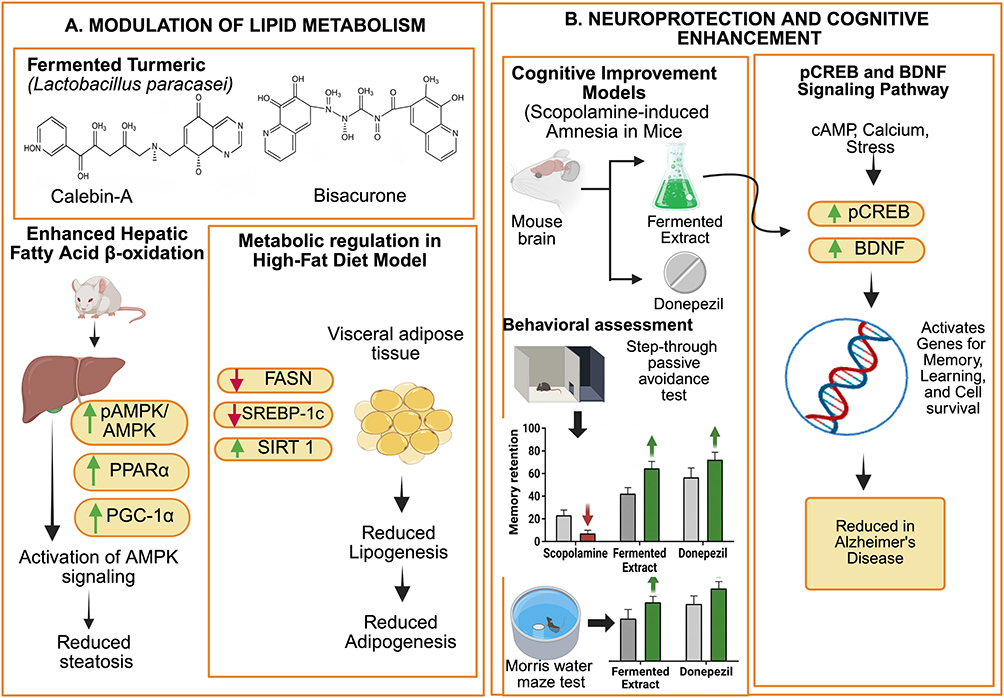 |
Figure 4 In vivo pharmacological effects of fermented Zingiberaceae extracts on metabolic and cognitive functions. (A) Mechanism; Modulation of Lipid Metabolism. (B) Mechanism: Neuroprotection and Cognitive Enhancement. Abbreviations: AMPK, AMP-activated protein kinase; PPARα, peroxisome proliferator-activated receptor alpha; PGC-1α, PPAR Gamma Coactivator-1 alpha; FASN, fatty acid synthase; SREBP-1c, sterol regulatory element-binding protein 1c; SIRT1, sirtuin 1; pCREB, phospho-cAMP response element-binding protein; BDNF, brain-derived neurotrophic factor. Notes: Green arrows upward (↑) indicate an increase or activation of the pathway. Red arrows downward (↓) indicate a decrease or inhibition of metabolic markers. |
Overall, the in vivo evidence indicates (Table 1) that fermentation of Zingiberaceae plants consistently enhances pharmacological activity through modulation of inflammatory responses, metabolic regulation and cognitive function. These findings support the relevance of fermentation as a bioprocess that improves the biological performance of plant-derived phytoconstituents and strengthens their translational potential in preclinical therapeutic applications.
Discussion
Fermentation has emerged as a biotechnological strategy capable of reshaping the phytochemical composition of medicinal plants, including members of Zingiberaceae family, with downstream effects on pharmacological activity.46,109 Across the reviewed studies, fermentation consistently altered the qualitative and quantitative profiles of bioactive constituents, which was frequently accompanied by enhanced biological effects in both preclinical studies (in vitro and in vivo models).110–112 Importantly, these outcomes were not uniform, but were strongly influenced by plant species, microbial strains, and fermentation parameters, indicating that fermentation acts as a context-dependent modulator rather than a universally enhancing process.41,113–115
Several studies reported an apparent contradiction, in which a quantitative reduction in certain phytoconstituents coincided with enhanced pharmacological activity. This phenomenon frequently ascribed to microbial enzyme-mediated biotransformation, specifically the hydrolysis of inactive glycosidic forms into more biologically active aglycones.116–118 Fermentation might also help break down complex phytochemicals into smaller molecules, which could make it easier for cells to take them up and for tissues to absorb them.68,119,120 From a conventional phytochemical perspective, reductions in total apparent compounds may be interpreted as a loss of therapeutic potential. However, the reviewed evidence suggests that biological activity is not solely determined by total compound concentration, but also by chemical form, molecular size and functional availability.121,122
Similar compositional shifts were observed in the fermentation of A. xanthioides using Lactobacillus casei, which resulted in increased total flavonoid and phenolic content alongside the loss of specific compounds such as procyanidin B2.49,123,124 The increase in total phenolics was proposed to arise from the release of matrix-bound compounds or the generation of low molecular weight phenolics during microbial metabolism.125–128 These compositional changes were associated with significant hepatoprotective effects in a rat model of non-alcoholic fatty liver disease. Although direct causal mechanisms were not fully elucidated, the findings indicate that fermentation can enhance the functional availability of bioactive constituents, thereby influencing systemic biological responses.129–132
Another study demonstrated that ginger (Z. officinale) fermented with L. plantarum bacteria leads to increased stability of 6-gingerol and 6-shogaol after the fermentation process. The bacteria produce enzymes that can remove a single water molecule (a dehydration process) from 6-gingerol, converting it into 6-shogaol. The compound 6-shogaol is more stable and exhibits significantly higher antioxidant activity compared to regular gingerol. It has been shown to enhance the immune cell (RAW 264.7 macrophage) ability to combat inflammation in vitro.58,133–135 This study explains that ginger can act as an immunomodulator through the pro-inflammatory signaling pathways. Fermented ginger was found to inhibit Nitric Oxide (NO) production by more than 90% (a very strong effect) and reduce cytokine expression such as Tumor Necrosis Factor-α (TNF-α), IL-6, and IL-1β.136–139 The study further indicates that the increased concentration of 6-shogaol works through dual inhibition of the Mitogen-Activated Protein Kinase (MAPK) pathway, specifically p44/c-Jun NH2-terminal kinase (JNK), and Nuclear factor-κB (NF-κB), by preventing phosphorylation of p65 and the degradation of Inhibitor of κB (IκB), which in turn modulates macrophage chemotaxis at relatively low concentrations (100–200 μg/mL).140,141 Thus, it could be applied in the treatment of chronic inflammation, such as rheumatoid arthritis and atherosclerosis, where macrophage infiltration into target tissues is a key pathological factor.
From a bioavailability and ADME perspective, ginger-derived phenolics are known to undergo rapid Phase II metabolism, resulting in low systemic exposure following oral administration. Fermentation may partially mitigate this limitation by increasing the relative abundance of more lipophilic metabolites, such as 6-shogaol, which may enhance intestinal permeability.142–145 In addition, fermentation generated diverse secondary metabolites that may act synergistically potentially amplifying biological effects despite low concentrations of individual compounds.95,146 Nevertheless, direct ADME investigations of fermented Zingiberaceae products remain scarce, and further studies are required to confirm whether fermentation meaningfully alters absorption, distribution, metabolism or excretion profiles in vivo.
In vivo investigations further indicate that fermented turmeric products may influence gut microbiota composition and metabolic health.95,147–149 High-fat diet models consistently showed reduced microbial diversity, whereas dietary administration of fermented turmeric partially restored species richness and increased the relative abundance of Akkermansia muciniphila, a bacterium negatively associated with metabolic disorders. Proposed explanations include antioxidant-mediated support of anaerobic bacterial growth and increased mucin availability stimulated by bacterial components such as lipoteichoic acid.150,151 Additionally, fermentation-derived oligosaccharides released from polysaccharide degradation may serve as substrates for microbial fermentation in the gut.152–154 Although direct mechanistic pathways were not systematically investigated, the temporal association between altered phytochemicals composition and biological outcomes suggest that fermentation may enhance functional exposure to bioactive constituents in vivo.
These findings highlight the fermentation effects toward phytoconstituents and their pharmacological activities. Fermentation helps break down large plant molecules into much smaller particles with higher bioavailability, making it easier to penetrate the dermal layers through the stratum corneum.155–158 The results of this study show an increase in compounds (organic acids, amino acids, and polyphenols) after fermentation. The enhanced antioxidant activity, as tested by DPPH and FRAP assays, is attributed to the formation of new metabolites resulting from microbial bio decomposition, which stimulate the production of Collagen I and II, as well as protect elastin from enzymatic degradation in the Extracellular Matrix.87,159–161 Furthermore, fermentation strengthens the epidermal barrier through the synthesis of key proteins, such as Loricrin and Filaggrin.162–164 These proteins are critical in clinical practice for treating dry and sensitive skin, as their anti-inflammatory activity can suppress IL-8.165–167 Additionally, toxicity testing revealed that FAA does not induce cytotoxicity in human skin cells, even at high concentrations, indicating it is safe for long-term use.87 However, clinical validation remains necessary before therapeutic claims can be established.
Fermentation has emerged as a pivotal biotechnological strategy for modulating phytochemical profiles to enhance the pharmacological efficacy of medicinal plants. In the Zingiberaceae family, the primary transformation mechanisms focus on specific dehydration processes and the bioconversion of phenolic constituents. For instance, the fermentation of ginger (Zingiber officinale) by lactic acid bacteria triggers the conversion of 6-gingerol into 6-shogaol via the removal of water molecules.58,133–135 This transformation is highly significant, as 6-shogaol exhibits superior chemical stability and more potent antioxidant and anti-inflammatory properties compared to its native form. This phenomenon is distinct when compared to the Fabaceae family (eg., soybean), where the dominant mechanism is not dehydration but rather enzymatic hydrolysis mediated by β-glucosidase. In Fabaceae, fermentation aims to cleave glycosidic bonds in isoflavone glycosides to release isoflavone aglycones (daidzein and genistein) from their sugar moieties.68,75,168 Although both processes share the common goal of increasing lipophilicity and bioavailability, Zingiberaceae exhibits more complex internal structural alterations. These changes directly modulate pro-inflammatory cytokine signaling pathways, specifically the Mitogen-Activated Protein Kinase (MAPK) and Nuclear Factor-kappa B (NF-kB) pathways, at the cellular level.
Furthermore, when compared to the Theaceae family (as in the black tea production process), the mechanisms occurring in the Zingiberaceae family are reductive and selective, resulting in the production of small molecules.169,170 In contrast, in Theaceae, fermentation via enzymatic oxidation often involves the polymerization of catechins into larger molecules such as teaflavins and tearubigins. Although these polymers provide unique sensory characteristics, these large molecules sometimes have limitations in penetrating biological layers. On the other hand, fermentation in the Zingiberaceae family produces much smaller particles that can penetrate the stratum corneum and dermal layers more effectively. This provides therapeutic advantages for the Zingiberaceae family in dermatological applications, such as the stimulation of type I and II collagen and the protection of elastin, which are not significantly observed in the fermentation products of the other plant families that produce large polyphenols.87,171
Differences in mechanisms are also clearly evident when comparing the Zingiberaceae family with the Asteraceae family. In the Asteraceae, fermentation is often focused on degrading the rigid lignocellulosic matrix of the cell wall to release bound phenolic.172–174 Although this process increases the quantitative yield of active compounds, it also creates new secondary metabolites through microbial biological decomposition that possess a better safety (toxicity) profile.175 Overall, other plant families use fermentation primarily to increase the concentration or solubility of compounds with low bioavailability into stable, lipophilic pharmacological agents with high affinity for biological targets in both in vitro and in vivo models.
Limitations of the Study
Although the present review provides comprehensive insights into the fermentation effects toward Zingiberaceae plants and phytoconstituents. This review has several limitations that should be acknowledged. Firstly, although enhance biological activity following fermentation was consistently reported, most studies relied on associative evidence, with limited direct validation of underlying molecular mechanism, such as enzyme-specific biotransformation pathways or metabolite-target interactions. Second, phytoconstituent characterization varied substantially across studies, with many relying on total phenolic or flavonoid content rather that comprehensive metabolic profiling, limiting direct comparability and synthesis. Furthermore, the absence of clinical validation and standardized outcomes measures limits translational interpretation. In addition, many in vitro and in vivo studies employed exposure levels that may not reflect achievable concentrations in humans, particularly given known ADME constraints of Zingiberaceae-derived compounds. These limitations highlight the need for future studies integrating controlled fermentation parameters, targeted metabolomics, and translation models to strengthen causal inference and applicability.
Conclusion
This review elaborates an overview of the complex and multifaceted fermentation as biologically relevant and methodologically adaptable strategy to enhance the phytochemical profile and pharmacological potential of Zingiberaceae plants. These transformations are strongly influenced by fermentation parameters, particularly microbial strain selection, substrate composition, fermentation time, temperature, and oxygen availability, which together determine the direction of phytochemical changes. A key insight emerging from this review is the pronounced species specificity with Zingiberaceae family. Fermentation outcomes differ substantially among genera and species such as Aframomum angustifolium, Amomum xanthioides, Curcuma longa and Zingiber officinale. These specificities explain why similar fermentation approaches may enhance anti-inflammatory or antioxidant activity in one species while primarily improving antidiabetic effects in another. From a pharmacological perspective, converging in vitro and in vivo evidence demonstrates that fermentation frequently enhances biological activity compared to non-fermented extracts. Overall, fermentation represents a promising approach to unlock the therapeutic potential of Zingiberaceae phytoconstituents through controlled biotransformation. Future perspective should prioritize standardized fermentation protocols, comparative multi species studies, and mechanistic investigations linking specific microbial enzymes to pharmacological outcomes.
Acknowledgments
The authors gratefully acknowledge (1) the Indonesian Education Scholarship (Beasiswa Pendidikan Indonesia), the Center for Higher Education Funding and Assessment (Pusat Pembiayaan dan Asesmen Pendidikan Tinggi), the Ministry of Higher Education, Science, and Technology of Republic Indonesia, and the Indonesia Endowment Fund for Education (LPDP) for supporting the tuition fee and the framework of the dissertation project of the first author and (2) the Rector of Universitas Padjadjaran via the Directorate of Research, Downstream, and Community Engagement for facilitating the APC. The APC is funded by Universitas Padjadjaran through the Indonesian Endowment Fund for Education (LPDP) on behalf of the Indonesian Ministry of Higher Education, Science and Technology, and managed under the EQUITY Program. (Contract No. 4303/B3/ DT.03.08/2025 and 3927/UN6.RKT/HK.07.00/2025). The present study was conducted in the framework of the dissertation project of the first author at the Doctoral Program in Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, West Java, Indonesia.
Disclosure
The authors declared that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Kuna L, Jakab J, Smolic R. et al. Peptic ulcer disease: a brief review of conventional therapy and herbal treatment options. J Clin Med. 2019;8(2):179. doi:10.3390/jcm8020179
2. Choi W, Choi CH, Kim YR, et al. HerDing: herb recommendation system to treat diseases using genes and chemicals. Database. 2016;2016:baw011. doi:10.1093/database/baw011
3. Lin LT, Hsu WC, Lin CC. Antiviral Natural Products and Herbal Medicines. J Tradit Complement Med. 2014;4(1):24–25. doi:10.4103/2225-4110.124335
4. Gupta M, Kapoor B, Gupta R, et al. Plants and phytochemicals for treatment of peptic ulcer: an overview. S Afr J Bot. 2021;138:105–114. doi:10.1016/j.sajb.2020.11.030
5. Zhang L, Zhuang H, Zhang Y, et al. Plants for health: an ethnobotanical 25-year repeat survey of traditional medicine sold in a major marketplace in North-west Yunnan, China. J Ethnopharmacol. 2018;224:119–125. doi:10.1016/j.jep.2018.05.029
6. Lalitha A. Phytochemicals and health benefits. Ann Geriatric Educ Med Sci. 2024;11(1):29–31. doi:10.18231/j.agems.2024.007
7. Debnath S, Vijayan D. Diversity, phytogeographical distribution, endemism and conservation status of Zingiberaceae in India. Plant Sci Today. 2024. doi:10.14719/pst.2708
8. Nikalje GC, Chonde A, Srivastava S, et al. Wild Vegetables of the Family Zingiberaceae. In: Wild Vegetables: Morphology, Phytochemistry and Utility - (Part 2). Vol. 2025. Bentham Science Publishers; 2025:171–173. doi:10.2174/9798898810009125010032
9. Chen S, Lin R, Lu H, et al. Effects of phenolic acids on free radical scavenging and heavy metal bioavailability in kandelia obovata under cadmium and zinc stress. Chemosphere. 2020;249:126341. doi:10.1016/j.chemosphere.2020.126341
10. Sharifi-Rad M, Varoni EM, Salehi B, et al. Plants of the Genus Zingiber as a Source of Bioactive Phytochemicals: from Tradition to Pharmacy. Molecules. 2017;22(12):2145. doi:10.3390/molecules22122145
11. Ashutosh P, Aabhash S, Ayush T, et al. An Overview of Zingiber officinale as an Essence of Life and Therapeutic Applications. J Sci Innov Nat Earth. 2024;4(4):1–7. doi:10.59436/jsiane.267.2583-2093
12. Romero A, Forero M, Sequeda-Castañeda LG, et al. Effect of ginger extract on membrane potential changes and AKT activation on a peroxide-induced oxidative stress cell model. J King Saud Univ Sci. 2018;30(2):263–269. doi:10.1016/j.jksus.2017.09.015
13. Tanvir EM, Hossen MS, Hossain MF, et al. Antioxidant Properties of Popular Turmeric (Curcuma longa) Varieties from Bangladesh. J Food Qual. 2017;2017:1–8. doi:10.1155/2017/8471785
14. Mao QQ, Xu XY, Cao SY, et al. Bioactive Compounds and Bioactivities of Ginger (Zingiber officinale Roscoe). Foods. 2019;8(6):185. doi:10.3390/foods8060185
15. Kazemi S, Yaghooblou F, Siassi F, et al. Cardamom supplementation improves inflammatory and oxidative stress biomarkers in hyperlipidemic, overweight, and obese pre-diabetic women: a randomized double-blind clinical trial. J Sci Food Agric. 2017;97(15):5296–5301. doi:10.1002/jsfa.8414
16. Lin X, Bai D, Wei Z, et al. Curcumin attenuates oxidative stress in RAW264.7 cells by increasing the activity of antioxidant enzymes and activating the Nrf2-Keap1 pathway. PLoS One. 2019;14(5):e0216711. doi:10.1371/journal.pone.0216711
17. Hsiang CY, Lo HY, Huang HC, et al. Ginger extract and zingerone ameliorated trinitrobenzene sulphonic acid-induced colitis in mice via modulation of nuclear factor-κB activity and interleukin-1β signalling pathway. Food Chem. 2013;136(1):170–177. doi:10.1016/j.foodchem.2012.07.124
18. Qiblawi S, Dhanarasu S, Faris MAI. Chemopreventive Effect of Cardamom (Elettaria cardamomum L.) Against Benzo(α)Pyrene-Induced Forestomach Papillomagenesis in Swiss Albino Mice. J Environ Pathol Toxicol Oncol. 2015;34(2):95–104. doi:10.1615/JEnvironPatholToxicolOncol.2015010838
19. Meiyanto E, Putri H, Arum Larasati Y, et al. Anti-proliferative and Anti-metastatic Potential of Curcumin Analogue, Pentagamavunon-1 (PGV-1), Toward Highly Metastatic Breast Cancer Cells in Correlation with ROS Generation. Adv Pharm Bull. 2019;9(3):445–452. doi:10.15171/apb.2019.053
20. Nielsen M, Witt MR, Thørgersen H. [3H] diazepam specific binding to rat cortex in vitro is enhanced by oleic, arachidonic and docosahexenoic acid isolated from pig brain. Eur J Pharmacol. 1988;146(2–3):349–353. doi:10.1016/0014-2999(88)90315-9
21. Liao YR, Leu YL, Chan YY, et al. Anti-Platelet Aggregation and Vasorelaxing Effects of the Constituents of the Rhizomes of Zingiber officinale. Molecules. 2012;17(8):8928–8937. doi:10.3390/molecules17088928
22. Shin JK, Park JH, Kim KS, et al. Antiulcer Activity of Steamed Ginger Extract against Ethanol/HCl-Induced Gastric Mucosal Injury in Rats. Molecules. 2020;25(20):4663. doi:10.3390/molecules25204663
23. Prayoga D, Aulifa D, Budiman A, et al. Etlingera Elatior Inflorescence Extract Mitigates Acute Gastric Ulcers by Suppressing the Expression of Inducible Nitric Oxide Synthase in Ethanol-Induced Wistar Rats. J Exp Pharmacol. 2025;17:343–357. doi:10.2147/JEP.S524517
24. Gawel K, Kukula-Koch W, Banono NS, et al. 6-Gingerol, a Major Constituent of Zingiber officinale Rhizoma, Exerts Anticonvulsant Activity in the Pentylenetetrazole-Induced Seizure Model in Larval Zebrafish. Int J Mol Sci. 2021;22(14):7745. doi:10.3390/ijms22147745
25. Hosseini A, Mirazi N. Acute administration of ginger (Zingiber officinale rhizomes) extract on timed intravenous pentylenetetrazol infusion seizure model in mice. Epilepsy Res. 2014;108(3):411–419. doi:10.1016/j.eplepsyres.2014.01.008
26. Ojewole JAO. Analgesic, antiinflammatory and hypoglycaemic effects of ethanol extract of Zingiber officinale (roscoe) rhizomes (Zingiberaceae) in mice and rats. Phytother Res. 2006;20(9):764–772. doi:10.1002/ptr.1952
27. Chumroenphat T, Somboonwatthanakul I, Saensouk S, et al. The Diversity of Biologically Active Compounds in the Rhizomes of Recently Discovered Zingiberaceae Plants Native to North Eastern Thailand. Pharmacogn J. 2019;11(5):1014–1022. doi:10.5530/pj.2019.11.160
28. Zhang Y, Su R, Yuan H, et al. Widely Targeted Volatilomics and Metabolomics Analysis Reveal the Metabolic Composition and Diversity of Zingiberaceae Plants. Metabolites. 2023;13(6):700. doi:10.3390/metabo13060700
29. Mohd RNZN, Ab Ghani N, Rasol NE. Metabolites Profiling and Fingerprinting of the Ethanolic Extract of Zingiber zerumbet (L.) Roscoe ex Sm. J Sci Math Lett. 2022;10(1):97–107. doi:10.37134/jsml.vol10.1.10.2022
30. Sheshrao K, Chandile MA, Khule SB, et al. A Brief Review on Herbal Curcuma Longa. Int J Pharma Res Appl. 2024;09(06):660–668. doi:10.35629/4494-0906660668
31. Thanawala S, Shah R, Alluri KV, et al. Comparative bioavailability of curcuminoids from a water-dispersible high curcuminoid turmeric extract against a generic turmeric extract: a randomized, cross-over, comparative, pharmacokinetic study. J Pharm Pharmacol. 2021;73(6):816–823. doi:10.1093/jpp/rgab028
32. Telange DR, Pandhare PV, Sawarkar KR, et al. Extraction, Isolation, Characterization, and Development of Phospholipids Complex Nanocarrier for Improved Solubility, Antiasthmatic, and Pharmacokinetic Potential of Curcuminoids. Curr Pharm Biotechnol. 2025;26(15):2474–2491. doi:10.2174/0113892010326636240919094339
33. Yit KH, Zainal AZ. Antimicrobial Potential of Natural Compounds of Zingiberaceae Plants and their Synthetic Analogues: a Scoping Review of In vitro and In silico Approaches. Curr Top Med Chem. 2024;24(13):1158–1184. doi:10.2174/0115680266294573240328050629
34. Zhou M, Li F, Chen J, et al. Research progress on natural bio-based encapsulation system of curcumin and its stabilization mechanism. Food Science and Technology. 2022;42:e78422. doi:10.1590/fst.78422
35. Alshaikh F, Al‐Samydai A, Issa R, et al. Encapsulation of gingerol into nanoliposomes: evaluation of in vitro anti‐inflammatory and anti‐cancer activity. Biomed Chromatogr. 2024;38(8). doi:10.1002/bmc.5899
36. Thandar A, Sreemoy KD. Ultrasonic assisted medicinal plant extraction: the review. World J Adv Res Rev. 2025;27(2):1348–1359. doi:10.30574/wjarr.2025.27.2.2811
37. O EM, Benkirane C, Abid M, et al. Sustainable Extraction of Bioactive Compounds From Plant Materials. Igi Global. 2025;2025:433–464. doi:10.4018/979-8-3693-9826-5.ch018
38. Selim D, Khiralla G, El Behairy U, et al. Bioavailability, Stability and Antioxidant Activity of Ginger and Curcumin Before and After Encapsulation. J Food Dairy Sci. 2023;14(4):81–91. doi:10.21608/jfds.2023.200452.1103
39. Cabanillas BLA, Valdez BO, Gutiérrez GEP, et al. Microbial Production of Bioactive Compounds. In: Microbial Products for Future Industrialization. 2023;Vol. 2023:181–198. doi:10.1007/978-981-99-1737-2_10
40. Li C, Naeem A, Shen J, et al. Advances in the pharmaceutical research of curcumin for oral administration. Open Chem. 2023;21(1). doi:10.1515/chem-2023-0171
41. Luo X, Dong M, Liu J, et al. Fermentation: improvement of pharmacological effects and applications of botanical drugs. Front Pharmacol. 2024;15:1430238. doi:10.3389/fphar.2024.1430238
42. Rustamova N, Huang G, Isokov M, et al. Modification of natural compounds through biotransformation process by microorganisms and their pharmacological properties. Fitoterapia. 2024;179:106227. doi:10.1016/j.fitote.2024.106227
43. Theilmann MC, Goh YJ, Nielsen KF, et al. Lactobacillus acidophilus Metabolizes Dietary Plant Glucosides and Externalizes Their Bioactive Phytochemicals. mBio. 2017;8(6). doi:10.1128/mBio.01421-17
44. Son J, Jeong KJ. Engineering of Leuconostoc citreum for Efficient Bioconversion of Soy Isoflavone Glycosides to Their Aglycone Forms. Int J Mol Sci. 2022;23(17):9568. doi:10.3390/ijms23179568
45. Macedo GA, Caria CRP, Barbosa PM, et al. Bioaccessibility Evaluation of Soymilk Isoflavones with Biotransformation Processing. Foods. 2023;12(18):3401. doi:10.3390/foods12183401
46. Lo KJ, Choudhary S, Ho CT, et al. Exploring the phytochemical composition and pharmacological effects of fermented turmeric using the isolated strain Lactobacillus rhamnosus FN7. J Food Bioactives. 2024; 13–24. doi:10.31665/JFB.2024.18368.
47. Ghosh S. Enhancement of Antioxidant Activity in Plant-Based Foods through Fermentation. J Food Biotechnol. 2025;06(01):44–48. doi:10.51470/FAB.2025.6.1.44
48. Yong C, Yoon Y, Yoo H, et al. Effect of Lactobacillus Fermentation on the Anti-Inflammatory Potential of Turmeric. J Microbiol Biotechnol. 2019;29(10):1561–1569. doi:10.4014/jmb.1906.06032
49. De Montijo PS, Razola DMC, Barbieri F, et al. Impact of Lactic Acid Bacteria Fermentation on Phenolic Compounds and Antioxidant Activity of Avocado Leaf Extracts. Antioxidants. 2023;12(2):298. doi:10.3390/antiox12020298
50. Singh B, Gupta S. Biotransformation based Pharmacophores from Plant Extract of Picrorhiza kurroa Royle Ex Benth. J Res Applied Sci Biotechnology. 2022;1(1):49–58. doi:10.55544/jrasb.1.1.7
51. Dahiya A, Majee C, Mazumder R, et al. Insight into the Glycosylation Methods of the Flavonoids as an Approach to Enhance its Bioavailability and Pharmacological Activities. Ind J Pharm Edu Res. 2023;57(2):354–371. doi:10.5530/ijper.57.2.45
52. König A, Sadova N, Dornmayr M, et al. Combined acid hydrolysis and fermentation improves bioactivity of citrus flavonoids in vitro and in vivo. Commun Biol. 2023;6(1):1083. doi:10.1038/s42003-023-05424-7
53. Yang T, Huang Y, Li Q, et al. Determination of Ten Flavonoids in the Raw and Fermented Fructus Aurantii by Quantitative Analysis of Multicomponents via a Single Marker (QAMS) Based on UPLC. J Anal Methods Chem. 2023;2023:1–11. doi:10.1155/2023/6067647
54. Park CM, Kim GM, Cha GS. Biotransformation of Flavonoids by Newly Isolated and Characterized Lactobacillus pentosus NGI01 Strain from Kimchi. Microorganisms. 2021;9(5):1075. doi:10.3390/microorganisms9051075
55. Johnson JB, Mani JS, Broszczak D, et al. Hitting the sweet spot: a systematic review of the bioactivity and health benefits of phenolic glycosides from medicinally used plants. Phytother Res. 2021;35(7):3484–3508. doi:10.1002/ptr.7042
56. Yang Z, Guo Z, Yan J, et al. Nutritional components, phytochemical compositions, biological properties, and potential food applications of ginger (Zingiber officinale): a comprehensive review. J Food Composit Analysis. 2024;128:106057. doi:10.1016/j.jfca.2024.106057
57. Tan Y, Gao M, Li L, et al. Functional components and antioxidant activity were improved in ginger fermented by Bifidobacterium adolescentis and Monascus purpureus. LWT. 2024;197:115931. doi:10.1016/j.lwt.2024.115931
58. Kim JE, Park KH, Park J, et al. Immunomodulatory Potential of 6-Gingerol and 6-Shogaol in Lactobacillus plantarum-Fermented Zingiber officinale Extract on Murine Macrophages. Int J Mol Sci. 2025;26(5):2159. doi:10.3390/ijms26052159
59. Maghraby YR, Labib RM, Sobeh M, et al. Gingerols and shogaols: a multi-faceted review of their extraction, formulation, and analysis in drugs and biofluids to maximize their nutraceutical and pharmaceutical applications. Food Chem X. 2023;20:100947. doi:10.1016/j.fochx.2023.100947
60. Tóth B, Horváth A, Laczkovich OJ, et al. Storage Conditions Influence the Quality of Ginger–A Stability Study Inspired by Clinical Trials. Planta Med. 2024;90(09):736–740. doi:10.1055/a-2283-8147
61. Zhou Y, Zheng J, Zhu Z, et al. Perspectives on two major bioactives in ginger, 6-gingerol and 6-shogaol, focusing on the in vivo metabolic fate and bioactivities. Crit Rev Food Sci Nutr. 2025:1–22. doi:10.1080/10408398.2025.2550519.
62. Rodríguez-Couto S. Solid-State Fermentation for Laccases Production and Their Applications. In: Current Developments in Biotechnology and Bioengineering. Elsevier; 2018:211–234. doi:10.1016/B978-0-444-63990-5.00011-6
63. Šelo G, Planinić M, Tišma M, et al. Bioconversion of Grape Pomace with Rhizopus oryzae under Solid-State Conditions: changes in the Chemical Composition and Profile of Phenolic Compounds. Microorganisms. 2023;11(4):956. doi:10.3390/microorganisms11040956
64. Verduzco-Oliva R, Gutierrez-Uribe JA. Beyond Enzyme Production: solid State Fermentation (SSF) as an Alternative Approach to Produce Antioxidant Polysaccharides. Sustainability. 2020;12(2):495. doi:10.3390/su12020495
65. Prachi M, Sunita M. Phytochemical Analysis and Biological Activities of Curcumin. Int J Multidisciplinary Res. 2023;5(4):5842. doi:10.36948/ijfmr.2023.v05i04.5842
66. Dhanya CS, Paul W, Victor SP, et al. On improving the physiological stability of curcuminoids: curcumininoid-silver nanoparticle complex as a better and efficient therapeutic agent. Nano-Struct Nano-Objects. 2021;25:100661. doi:10.1016/j.nanoso.2020.100661
67. Kim S, Choi SY, Lee HI, et al. Protective Responses of Green Yuja Peel Extracts to Lipopolysaccharide-Induced Inflammation and Reactive Oxygen Species Production in RAW264.7 Cells. Prev Nutr Food Sci. 2024;29(3):301–310. doi:10.3746/pnf.2024.29.3.301
68. Godse R, Kulkarni R. Nutraceutical Augmentation of Soybean Products Using Microbial β-Glucosidases. Food Biotechnol. 2023;37(3):235–259. doi:10.1080/08905436.2023.2236692
69. Bonafini C, Marzotto M, Olioso D, et al. Biological activity of interferon gamma and lipopolysaccharide on the nitric oxide production in C6 astroglioma cells and some unexpected effects of potentization. Int J High Dilut Res. 2021;14(2):23–27. doi:10.51910/ijhdr.v14i2.779
70. Nurmilah S, Frediansyah A, Cahyana Y, et al. Exploring microbial dynamics and metabolomic profiling of isoflavone transformation in black and yellow soybean Tempe for sustainable functional foods. Food Chem Mol Sci. 2025;11:100279. doi:10.1016/j.fochms.2025.100279
71. Kalebar VU, Chakraborty P. Role of Bioactive Metabolites Produced by Plants and Microbes in Fuel Production. In: Emerging Sustainable Technologies for Biofuel Production. Spinger Nature Link; 2024:427–454. doi:10.1007/978-3-031-52167-6_18
72. M LR, Bruniaux P, Raveschot C, et al. Lactobacillus Use for Plant Fermentation: new Ways for Plant-Based Product Valorization. In: Lactobacillus - A Multifunctional Genus. IntechOpen; 2023. doi:10.5772/intechopen.104958
73. Kang Y, Sun Y, Cui J, et al. Combined microbiome and metabolomics analysis of yupingfeng san fermented by Bacillus coagulans: insights into probiotic and herbal interactions. PeerJ. 2025;13:e19206. doi:10.7717/peerj.19206
74. Tang F, Wei B, Qin C, et al. Enhancing the inhibitory activities of polyphenols in passion fruit peel on α-Amylase and α-Glucosidase via β-Glucosidase-producing Lactobacillus fermentation. Food Biosci. 2024;62:105005. doi:10.1016/j.fbio.2024.105005
75. Paventi G, Di Martino C, Coppola F, et al. β-Glucosidase Activity of Lactiplantibacillus Plantarum: a Key Player in Food Fermentation and Human Health. Preprint. 2025;1:1795. doi:10.20944/preprints202503.1795.v1
76. De Villa R, Roasa J, Mine Y, et al. Impact of solid-state fermentation on factors and mechanisms influencing the bioactive compounds of grains and processing by-products. Crit Rev Food Sci Nutr. 2023;63(21):5388–5413. doi:10.1080/10408398.2021.2018989
77. Zhang P, Zhang J, Li L, et al. The Release of Bound Phenolics to Enhance the Antioxidant Activity of Cornmeal by Liquid Fermentation with Bacillus subtilis. Foods. 2025;14(3):499. doi:10.3390/foods14030499
78. Tang Q, Huang J, Zhang S, et al. Keystone microbes affect the evolution and ecological coexistence of the community via species/strain specificity. J Appl Microbiol. 2022;132(2):1227–1238. doi:10.1111/jam.15255
79. Yan C, Chen X, Liu Q, et al. Effects of Lactiplantibacillus plantarum on Metabolites and Flavors in Synthetic Microbiota During Baijiu Fermentation. Foods. 2024;14(1):31. doi:10.3390/foods14010031
80. Li R, Luo W, Liu Y, et al. The investigation on the characteristic metabolites of Lactobacillus plantarum RLL68 during fermentation of beverage from by-products of black tea manufacture. Curr Res Food Sci. 2022;5:1320–1329. doi:10.1016/j.crfs.2022.07.014
81. Zhang X, Zhang C, Xiao L, et al. Gas chromatography-mass spectrometry and non-targeted metabolomics analysis reveals the flavor and nutritional metabolic differences of cow’s milk fermented by Lactiplantibacillus plantarum with different phenotypic. Food Biosci. 2024;60:104433. doi:10.1016/j.fbio.2024.104433
82. Xu D, Wang N, Rinne M, et al. The bacterial community and metabolome dynamics and their interactions modulate fermentation process of whole crop corn silage prepared with or without inoculants. Microb Biotechnol. 2021;14(2):561–576. doi:10.1111/1751-7915.13623
83. Gaur G, Gänzle MG. Conversion of (poly)phenolic compounds in food fermentations by lactic acid bacteria: novel insights into metabolic pathways and functional metabolites. Curr Res Food Sci. 2023;6:100448. doi:10.1016/j.crfs.2023.100448
84. Zhang Y, Ye Q, Liu B, et al. Fermenting Distiller’s Grains by the Domesticated Microbial Consortium To Release Ferulic Acid. J Agric Food Chem. 2024;72(16):9259–9267. doi:10.1021/acs.jafc.3c08067
85. Koistinen VM, Hanhineva K. Microbial and endogenous metabolic conversions of rye phytochemicals. Mol Nutr Food Res. 2017;61(7). doi:10.1002/mnfr.201600627
86. Eun CS, Lim JS, Lee J, et al. The protective effect of fermented Curcuma longa L. on memory dysfunction in oxidative stress-induced C6 gliomal cells, proinflammatory-activated BV2 microglial cells, and scopolamine-induced amnesia model in mice. BMC Complement Altern Med. 2017;17(1):367. doi:10.1186/s12906-017-1880-3
87. Albouy M, Aubailly S, Jeanneton O, et al. Skin-protective biological activities of bio-fermented Aframomum angustifolium extract by a consortium of microorganisms. Front Pharmacol. 2023;14:1303198. doi:10.3389/fphar.2023.1303198
88. Mohammed INA, Perimal EK, Vidyadaran S, et al. Zerumbone ameliorates neuroinflammation in LPS-induced SH-SY5Y cells, an in vitro model of neuropathic pain: targeting NO, IL-6, and TNF-α. Neurosci Res Notes. 2025;8(3):374. doi:10.31117/neuroscirn.v8i3.374
89. Khanh NLT, Thi HB, Van ATT, et al. Inhibition of pro-inflammatory cytokines by homalolide A and homalomenol A isolated from rhizomes of Homalomena pendula. Zeitschrift für Naturforschung C. 2025;80(7–8):353–359. doi:10.1515/znc-2024-0152
90. Kim M, Yun J, Yamakuni K. Anti‐inflammatory Effects of Zerumbone on Chronic Inflammation Models in Differentiated Human THP‐1 Monocytes Induced by Lipopolysaccharide. FASEB J. 2017;31(S1):5036–5048. doi:10.1096/fasebj.31.1_supplement.lb330
91. Papaccio F, D′Arino A, Caputo S, et al. Focus on the Contribution of Oxidative Stress in Skin Aging. Antioxidants. 2022;11(6):1121. doi:10.3390/antiox11061121
92. Dorf N, Maciejczyk M. Skin senescence—from basic research to clinical practice. Front Med Lausanne. 2024;11:1484345. doi:10.3389/fmed.2024.1484345
93. Massaro S, Sica J, Ghion G, et al. Enhancing Malva sylvestris extract properties through lactic acid bacteria fermentation: impact on phytochemical profile and bioactivity. Food Product Process Nutrit. 2025;7(1):40. doi:10.1186/s43014-025-00315-2
94. Beck F, Pedersen NR, Nielsen DS. Fermented Rapeseed and Soybean Alone and in Combination with Macro Algae Inhibit Human and Pig Pathogenic Bacteria In Vitro. Microorganisms. 2024;12(5):891. doi:10.3390/microorganisms12050891
95. Lin WS, Hwang SE, Koh YC, et al. Modulatory Effects of Lactobacillus paracasei -Fermented Turmeric on Metabolic Dysregulation and Gut Microbiota in High-Fat Diet-Induced Obesity in Mice. J Agric Food Chem. 2024;72(32):17924–17937. doi:10.1021/acs.jafc.4c01501
96. Hwang SJ, Choi YJ, Wang JH, et al. Lactobacillus Casei-fermented Amomum Xanthioides Mitigates non-alcoholic fatty liver disease in a high-fat diet mice model. Biomed. Pharmacother. 2024;172:116250. doi:10.1016/j.biopha.2024.116250
97. Qayyum N, Ismael M, Haoyue H, et al. Dietary supplementation of probiotic Lactobacillus modulates metabolic dysfunction-associated steatotic liver disease and intestinal barrier integrity in obesity-induced mice. J Food Sci. 2024;89(12):10113–10133. doi:10.1111/1750-3841.17439
98. Li Z, Zhao M, Ji X, et al. Alginate oligosaccharides improve high-fat induced hepatic steatosis via PGC-1α-mediated lipophagy and fatty acid β-oxidation pathway. J Funct Foods. 2023;110:105825. doi:10.1016/j.jff.2023.105825
99. Arulkumar R, Jung HJ, Noh SG, et al. Soyasapogenol C from Fermented Soybean (Glycine Max) Acting as a Novel AMPK/PPARα Dual Activator Ameliorates Hepatic Steatosis: a Novel SANDA Methodology. Int J Mol Sci. 2022;23(10):5468. doi:10.3390/ijms23105468
100. Ma K, Sheng W, Song X, et al. Chlorogenic Acid from Burdock Roots Ameliorates Oleic Acid-Induced Steatosis in HepG2 Cells through AMPK/ACC/CPT-1 Pathway. Molecules. 2023;28(21):7257. doi:10.3390/molecules28217257
101. Lai C, Liao S, Tsai M, et al. Calebin‐A inhibits adipogenesis and hepatic steatosis in high‐fat diet‐induced obesity via activation of AMPK signaling. Mol Nutr Food Res. 2015;59(10):1883–1895. doi:10.1002/mnfr.201400809
102. Han SO, Kim G, Shim YY, et al. Anti-Obesity Effects of Abeliophyllum distichum Extracts via the AMPK Signaling Pathway in 3T3-L1 Adipocytes. Prev Nutr Food Sci. 2025;30(4):360–369. doi:10.3746/pnf.2025.30.4.360
103. Lim SH, Lee HS, Han HK, et al. Saikosaponin A and D Inhibit Adipogenesis via the AMPK and MAPK Signaling Pathways in 3T3-L1 Adipocytes. Int J Mol Sci. 2021;22(21):11409. doi:10.3390/ijms222111409
104. Hong JW, Park HY, Kim HA, et al. Inhibition Effect of Adipogenesis and Lipogenesis via Activation of AMPK in Preadipocytes Treated with Canavalia gladiata Extract. Int J Mol Sci. 2023;24(3):2108. doi:10.3390/ijms24032108
105. Lazarova M, Stefanova M, Denev P, et al. Neuroprotective Effect of Marrubium vulgare Extract in Scopolamine-Induced Cognitive Impairment in Rats: behavioral and Biochemical Approaches. Biology. 2024;13(6):426. doi:10.3390/biology13060426
106. Jeon G, Hun Lee S, Kwon YS, et al. Cognitive Enhancing Activity of Fermented Aloe arborescens Extract on Scopolamine-induced Memory Impairment in Mice. Pharmacogn Mag. 2023;19(1):5–12. doi:10.1177/09731296221137408
107. Lee CH, Ko MS, Kim YS, et al. Neuroprotective Effects of Davallia mariesii Roots and Its Active Constituents on Scopolamine-Induced Memory Impairment in In vivo and In Vitro Studies. Pharmaceuticals. 2023;16(11):1606. doi:10.3390/ph16111606
108. Djeuzong E, Kandeda AK, Djiogue S, et al. Antiamnesic and Neuroprotective Effects of an Aqueous Extract of Ziziphus jujuba Mill. (Rhamnaceae) on Scopolamine-Induced Cognitive Impairments in Rats. Evid Based Complement Alternat Med. 2021;2021:1–15. doi:10.1155/2021/5577163
109. Danthanarayana DN, Perera HDSM. Investigating the effect of fermentation by Lactobacillus reuteri on biochemical properties of Ceylon Cinnamon. Int J Pharm Pract. 2024;32(Supplement_2):ii65–ii65. doi:10.1093/ijpp/riae058.077
110. Indira A, Shahar B, Joshi B, Chongtham N. Assessment of bioactive compound variations and in-vitro and in-vivo antioxidant activity in edible fresh and processed Bambusa nutans shoot through FTIR, GC/MS and HPLC analyses. Food Chem. 2024;452:139552. doi:10.1016/j.foodchem.2024.139552
111. Lee JY, Park HM, Kang CH. Antioxidant Effect via Bioconversion of Isoflavonoid in Astragalus membranaceus Fermented by Lactiplantibacillus plantarum MG5276 In Vitro and In vivo. Fermentation. 2022;8(1):34. doi:10.3390/fermentation8010034
112. Ryu KM, Kim H, Woo J, et al. Enhancement of the bioactive compounds and biological activities of maca (Lepidium meyenii) via solid-state fermentation with Rhizopus oligosporus. Food Sci Biotechnol. 2024;33(11):2585–2596. doi:10.1007/s10068-023-01508-6
113. Lai X, Liu S, Zhang Y, et al. Decoding Forage-Driven Microbial–Metabolite Patterns: a Multi-Omics Comparison of Eight Tropical Silage Crops. Fermentation. 2025;11(8):480. doi:10.3390/fermentation11080480
114. Merenkova S, Fatkullin R, Kalinina I. Effect of Fermentation on the Biochemical Parameters Antioxidant Capacity and Dispersed Composition of Plant Beverages Based on Barley and Hemp Seeds. Fermentation. 2022;8(8):384. doi:10.3390/fermentation8080384
115. Li C, Liu T, Li X, et al. Review of quinoa fermentation: product diversity, process optimization, and nutritional enhancement. Front Nutr. 2025;12:1605558. doi:10.3389/fnut.2025.1605558
116. Yan Z, Niu W, Liu Y, et al. Microbial fermentation alters the phytochemical profile of Ginkgo biloba leaves. Future Food. 2025;12:100764. doi:10.1016/j.fufo.2025.100764
117. Zhang J, Xia N, Teng J, et al. Effect of lactic acid bacteria fermentation on extractable and non-extractable polyphenols of soybean milk: influence of β-glucosidase and okara. Food Biosci. 2023;56:103326. doi:10.1016/j.fbio.2023.103326
118. Lee SH, Lee NR, Heun JJ, et al. Evolution-aided enhancement of β-glucosidase activity for improved conversion of isoflavone glucosides to aglycones by Lactobacillus gasseri. Food Chem. 2025;493:146055. doi:10.1016/j.foodchem.2025.146055
119. Vijayakumar V, Variyar EJ, H M. Effect of Fermentation on the Bioactivities of the Aqueous Extract of Viburnum Coriaceum Blume – Positive as Well as Negative Inferences on Experimental Basis. 2022. doi:10.21203/rs.3.rs-2303144/v1
120. Ran J, Su Y, Wang P, et al. Effect of Lactobacillus acidophilus fermentation on bioaccessibility: the relationship between biotransformation and antioxidant activity of apple polyphenols based on metabolomics. LWT. 2023;190:115360. doi:10.1016/j.lwt.2023.115360
121. Ciupei D, Colişar A, Leopold L, et al. Polyphenols: from Classification to Therapeutic Potential and Bioavailability. Foods. 2024;13(24):4131. doi:10.3390/foods13244131
122. Di Lorenzo C, Colombo F, Biella S, et al. Polyphenols and Human Health: the Role of Bioavailability. Nutrients. 2021;13(1):273. doi:10.3390/nu13010273
123. Azkia MN, Cahyanto MN, Mayangsari Y, et al. Enhancement of phenolic profile and antioxidant activity of black glutinous rice (Oryza sativa var. glutinosa) due to tape fermentation. Arab J Chem. 2023;16(11):105275. doi:10.1016/j.arabjc.2023.105275
124. Ayar-Sümer EN, Verheust Y, Özçelik B, et al. Impact of Lactic Acid Bacteria Fermentation Based on Biotransformation of Phenolic Compounds and Antioxidant Capacity of Mushrooms. Foods. 2024;13(11):1616. doi:10.3390/foods13111616
125. Essa HA, El-Metwally AE, Elkady NA. The potential protective effects of fermented extra virgin olive oil on high-fructose diet-induced non-alcoholic steatohepatitis in rats. Food Product Process Nutrit. 2025;7(1):44. doi:10.1186/s43014-025-00320-5
126. Gao Z, Xie M, Zhou R, et al. Ameliorative effects of Monascus-fermented hawthorn extract on a high-fat diet-induced rat model of non-alcoholic fatty liver disease. Heliyon. 2024;10(18):e37354. doi:10.1016/j.heliyon.2024.e37354
127. Pratiwi VN, Anggraini CD, Kardina RN, et al. Snakefruit (Salacca Zalacca (Gaerth.)voss) Vinegar with Mixed Culture Fermentation and Lipid Regulation: A Study on Ccl4-Induced Male Rats. 2025. doi:10.21203/rs.3.rs-6307778/v1
128. Zubaidah E, Susanti I, Sujuti H, et al. The distinctive hepatoprotective activity of turmeric kombucha (Curcuma longa) induced by diethylnitrosamine in Balb/C mice. Food Biosci. 2023;55:103043. doi:10.1016/j.fbio.2023.103043
129. Lee NK, Lee Y, Shin DS, et al. Hepatoprotective Effect of Lactiplantibacillus plantarum DSR330 in Mice with High Fat Diet-Induced Nonalcoholic Fatty Liver Disease. J Microbiol Biotechnol. 2024;34(2):399–406. doi:10.4014/jmb.2310.10026
130. Sabirin F, Lim SM, Neoh CF, et al. Hepatoprotection of Probiotics Against Non-Alcoholic Fatty Liver Disease in vivo: a Systematic Review. Front Nutr. 2022;9:844374. doi:10.3389/fnut.2022.844374
131. Chen TY, Chen YL, Chiu WC, et al. Effects of the Water Extract of Fermented Rice Bran on Liver Damage and Intestinal Injury in Aged Rats with High-Fat Diet Feeding. Plants. 2022;11(5):607. doi:10.3390/plants11050607
132. Park CH, Kang H, Lee SG. Enhanced Anti-Inflammatory and Non-Alcoholic Fatty Liver Disease (NAFLD) Improvement Effects of Bacillus subtilis-Fermented Fagopyrum tataricum Gaertner. Fermentation. 2024;10(3):116. doi:10.3390/fermentation10030116
133. Lgaz H, Messali M, Lee H. Mechanistic Evaluation of Radical Scavenging Pathways in Ginger Phenolics: a DFT Study of 6-Gingerol, 6-Shogaol, and 6-Paradol. Int J Mol Sci. 2025;26(22):11217. doi:10.3390/ijms262211217
134. Bischoff-Kont I, Primke T, Niebergall LS, et al. Ginger Constituent 6-Shogaol Inhibits Inflammation- and Angiogenesis-Related Cell Functions in Primary Human Endothelial Cells. Front Pharmacol. 2022;13:844767. doi:10.3389/fphar.2022.844767
135. Zhou X, Münch G, Wohlmuth H, et al. Synergistic Inhibition of Pro-Inflammatory Pathways by Ginger and Turmeric Extracts in RAW 264.7 Cells. Front Pharmacol. 2022;13:818166. doi:10.3389/fphar.2022.818166
136. Pázmándi K, Ágics B, Szöllősi AG, et al. Ginger-derived bioactive compounds attenuate the Toll-like receptor mediated responses of human dendritic cells. Eur J Pharmacol. 2024;967:176399. doi:10.1016/j.ejphar.2024.176399
137. Attallah KA, El-Dessouki AM, Abd-Elmawla MA, et al. The therapeutic potential of naturally occurring 6-shogaol: an updated comprehensive review. Inflammopharmacology. 2025. doi:10.1007/s10787-025-01812-z
138. Guo X, Qiu J, Qian Y. 6-Shogaol Mitigates Sepsis-Associated Hepatic Injury through Transcriptional Regulation. Nutrients. 2021;13(10):3427. doi:10.3390/nu13103427
139. Zhou X, Al-Khazaleh A, Afzal S, et al. 6-Shogaol and 10-Shogaol Synergize Curcumin in Ameliorating Proinflammatory Mediators via the Modulation of TLR4/TRAF6/MAPK and NFκB Translocation. Biomol Ther. 2023;31(1):27–39. doi:10.4062/biomolther.2022.039
140. Shan L, Tyagi A, Ham HJ, et al. Uncovering the antiinflammatory potential of Lactiplantibacillus Plantarum fermented Cannabis Sativa L seeds. NPJ Sci Food. 2024;8(1):42. doi:10.1038/s41538-024-00285-8
141. Songvut P, Nakareangrit W, Cholpraipimolrat W, et al. Unraveling the interconversion pharmacokinetics and oral bioavailability of the major ginger constituents: [6]-gingerol, [6]-shogaol, and zingerone after single-dose administration in rats. Front Pharmacol. 2024;15:1391019. doi:10.3389/fphar.2024.1391019
142. Gonzalez-Ramirez M, Kazakova J, Garcia-Serrano P, et al. Commercial wine yeast nitrogen requirement influences the production of secondary metabolites (aroma, hydroxytyrosol, melatonin and other bioactives) during alcoholic fermentation. Int J Food Microbiol. 2024;421:110788. doi:10.1016/j.ijfoodmicro.2024.110788
143. Lin YM, Chung YC, Chen PY, et al. Fermentation of Chenopodium formosanum Leaf Extract with Aspergillus oryzae Significantly Enhanced Its Physiological Activities. Appl Sci. 2023;13(5):2917. doi:10.3390/app13052917
144. Koistinen VM, Hedberg M, Shi L, et al. Metabolite Pattern Derived from Lactiplantibacillus plantarum —Fermented Rye Foods and In Vitro Gut Fermentation Synergistically Inhibits Bacterial Growth. Mol Nutr Food Res. 2022;66(21). doi:10.1002/mnfr.202101096
145. Bodade R, Kumar A, Sharma VK, et al. Editorial: recent advances in biotechnological applications of microbial secondary metabolites. Front Microbiol. 2024;15:1525146. doi:10.3389/fmicb.2024.1525146
146. Xiang X, Liu Y, Zeng Z, et al. Exploration of the mechanism of fermented turmeric preventing glucolipid metabolic disorder: insights into signaling of microbiota-SCFAs-brown adipose tissue axis in mice. J Funct Foods. 2025;125:106675. doi:10.1016/j.jff.2025.106675
147. Ghanbari F, Abedpoor N, Peymani M, et al. Effects of the Phytocompound Combination against Dysbiosis Induced by AGE-Rich High-fat Diet in Mice. Int J Med Lab. 2023. doi:10.18502/ijml.v10i2.12948
148. Mamun MAA, Rakib A, Mandal M, et al. Impact of a High-Fat Diet on the Gut Microbiome: a Comprehensive Study of Microbial and Metabolite Shifts During Obesity. Cells. 2025;14(6):463. doi:10.3390/cells14060463
149. Zhang J, Hu B, Deng X, et al. Multiomics analysis investigating the impact of a high-fat diet in female Sprague–Dawley rats: alterations in plasma, intestinal metabolism, and microbial composition. Front Nutr. 2024;11:1359989. doi:10.3389/fnut.2024.1359989
150. Cai Z, Guo Y, Wang S, et al. Lactiplantibacillus plantarum lipoteichoic acid as a key surface molecule in immune enhancement and pathogen defense. Food Sci Human Wellness. 2025;2025:9250669. doi:10.26599/FSHW.2025.9250669
151. Wang S, Ahmadi S, Nagpal R, et al. Lipoteichoic acid from the cell wall of a heat killed Lactobacillus paracasei D3-5 ameliorates aging-related leaky gut, inflammation and improves physical and cognitive functions: from C. elegans to mice. Geroscience. 2020;42(1):333–352. doi:10.1007/s11357-019-00137-4
152. de Oliveira DP, Todorov SD, Fabi JP. Exploring the Prebiotic Potentials of Hydrolyzed Pectins: mechanisms of Action and Gut Microbiota Modulation. Nutrients. 2024;16(21):3689. doi:10.3390/nu16213689
153. Zhang X, Liu Y, Chen XQ, et al. Catabolism of Saccharina japonica polysaccharides and oligosaccharides by human fecal microbiota. LWT. 2020;130:109635. doi:10.1016/j.lwt.2020.109635
154. Wang Q, Chen H, Yin M, et al. In vitro digestion and human gut microbiota fermentation of Bletilla striata polysaccharides and oligosaccharides. Front Cell Infect Microbiol. 2023;13:1105335. doi:10.3389/fcimb.2023.1105335
155. Ziemlewska A, Nizioł-łukaszewska Z, Zagórska-Dziok M, et al. Anti-Aging, Anti-Inflammatory, and Cytoprotective Properties of Lactobacillus- and Kombucha-Fermented C. pepo L. Peel and Pulp Extracts with Prototype Skin Toner Development. Molecules. 2025;30(20):4082. doi:10.3390/molecules30204082
156. Li Q, Zhu H, Hou R, et al. Microbial Fermentation Potentiates the Multifunctional Skin-Care Activities of Polianthes tuberosa L. Flower Extract: antioxidant, Anti-Glycation, and Anti-Melanogenic Effects. Cosmetics. 2025;12(6):243. doi:10.3390/cosmetics12060243
157. Hering A, Ochocka JR, Baranska H, et al. Mangiferin and Hesperidin Transdermal Distribution and Permeability through the Skin from Solutions and Honeybush Extracts (Cyclopia sp.)—A Comparison Ex Vivo Study. Molecules. 2021;26(21):6547. doi:10.3390/molecules26216547
158. Mhlanga P, Elegbeleye J, Sivakumar D. Changes in bioactive compounds and antioxidant activity during fermentation and in vitro digestion of melon and pumpkin leaves smoothies. Int J Food Sci Technol. 2025;60(2). doi:10.1093/ijfood/vvaf156
159. Surya R, Kim JY, Kamal N, et al. Primary perspectives towards kimchi as a beauty food enhancing collagen, elastin, hyaluronic acid, and antioxidant enzymes in skin cells. Discover Food. 2025;5(1):160. doi:10.1007/s44187-025-00466-8
160. Yan X, Yang M, Cai X, et al. Fermented rice bran extract delays skin aging by increasing the synthesis of collagen and elastin. Front Pharmacol. 2025;16. doi:10.3389/fphar.2025.1692491
161. Wang M, Yang W, Chen M, et al. Bioactive components and anti-photoaging activity of Aspergillus oryzae fermented rice bran extract. Food Biosci. 2025;67:106354. doi:10.1016/j.fbio.2025.106354
162. Nakamizo S, Yan X, Kabashima K. Enhancement of skin barrier function and augmentation of epidermal cell-cell interactions by galactomyces ferment filtrate. J Dermatol Sci. 2024;115(2):94–97. doi:10.1016/j.jdermsci.2024.03.011
163. Yan X, Tsuji G, Hashimoto-H A, et al. Galactomyces Ferment Filtrate Potentiates an Anti-Inflammaging System in Keratinocytes. J Clin Med. 2022;11(21):6338. doi:10.3390/jcm11216338
164. Yang F, Hu Y, Wu M, et al. Biologically Active Components and Skincare Benefits of Rice Fermentation Products: a Review. Cosmetics. 2025;12(1):29. doi:10.3390/cosmetics12010029
165. Tsuji G, Hashimoto-H A, Yumine A, et al. PDE4 inhibition by difamilast regulates filaggrin and loricrin expression via keratinocyte proline-rich protein in human keratinocytes. J Dermatol Sci. 2023;110(2):61–68. doi:10.1016/j.jdermsci.2023.04.007
166. Kim Y, Lim KM. Skin barrier dysfunction and filaggrin. Arch Pharm Res. 2021;44(1):36–48. doi:10.1007/s12272-021-01305-x
167. Wang Q, Sun Y, Cheng J, et al. 51888 An Oat-Containing Cream Designed to Boost Loricrin, Claudin 1 and Improve Skin Barrier: observation from In Vitro and Clinical Studies. J Am Acad Dermatol. 2024;91(3):AB121. doi:10.1016/j.jaad.2024.07.487
168. Li C, Xu T, Liu XW, et al. The expression of β-glucosidase during natto fermentation increased the active isoflavone content. Food Biosci. 2021;43:101286. doi:10.1016/j.fbio.2021.101286
169. Ivanović M, Makoter K, Razboršek MI. Comparative study of chemical composition and antioxidant activity of essential oils and crude extracts of four characteristic zingiberaceae herbs. Plants. 2021;10(3):1–20. doi:10.3390/plants10030501
170. Zheng O. Thearubigins: a comprehensive study from formation mechanism to biological activity and application prospects. Theoret Natl Sci. 2025;77(1):24–33. doi:10.54254/2753-8818/2024.LA19832
171. Wichayapreechar P, Charoenjittichai R, Prasansuklab A, et al. Exploring the In Vitro Antioxidant, Anti-Aging, and Cytotoxic Properties of Kaempferia galanga Linn. Rhizome Extracts for Cosmeceutical Formulations. Cosmetics. 2024;11(3):97. doi:10.3390/cosmetics11030097
172. Bing RG, Sulis DB, Carey MJ, et al. Beyond low lignin: identifying the primary barrier to plant biomass conversion by fermentative bacteria. Sci Adv. 2024;10(42). doi:10.1126/sciadv.adq4941
173. Steck J, Junker F, Eichhöfer H, et al. Chemically Different but Often Mistaken Phenolic Polymers of Food Plants: proanthocyanidins and Lignin in Seeds. J Agric Food Chem. 2022;70(37):11704–11714. doi:10.1021/acs.jafc.2c03782
174. Manuela RGG, Rita ML. Cellulases, hemicellulases and ligninolytic enzymes: mechanism of action, optimal processing conditions and obtaining value-added compounds in plant matrices. MOJ Food Process Technol. 2022;10(1):30–37. doi:10.15406/mojfpt.2022.10.00270
175. Chouhan V, Kumar S, Raypa P, et al. Biotechnological Advances in Enhancing Secondary Metabolite Production. Plant Secondary Metabolites. CRC Press; 2025:384–414. doi:10.1201/9781003717454-17
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Fermented Plant-Derived Phytoconstituents in Biocosmetics: Pharmacological Enhancements and Applications
Suhaera S, Sammulia SF, Prayoga DK, Ramadhania ZM, Mita SR, Fristiohady A, Milanda T
Clinical, Cosmetic and Investigational Dermatology 2026, 19:602919
Published Date: 15 April 2026