Back to Journals » International Journal of Nanomedicine » Volume 17
Extracellular Vesicles: The Next Generation Theranostic Nanomedicine for Inflammatory Bowel Disease
Authors Li DF, Yang MF, Xu J, Xu HM ![]() , Zhu MZ
, Zhu MZ ![]() , Liang YJ
, Liang YJ ![]() , Zhang Y
, Zhang Y ![]() , Tian CM, Nie YQ
, Tian CM, Nie YQ ![]() , Shi RY, Wang LS
, Shi RY, Wang LS ![]() , Yao J
, Yao J ![]()
Received 14 April 2022
Accepted for publication 25 August 2022
Published 5 September 2022 Volume 2022:17 Pages 3893—3911
DOI https://doi.org/10.2147/IJN.S370784
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Dongwoo Khang
De-feng Li,1,* Mei-feng Yang,2,* Jing Xu,3 Hao-ming Xu,3 Min-zheng Zhu,3 Yu-jie Liang,4 Yuan Zhang,5 Cheng-mei Tian,6 Yu-qiang Nie,3 Rui-yue Shi,1 Li-sheng Wang,1 Jun Yao1
1Department of Gastroenterology, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; The First Affiliated Hospital, Southern University of Science and Technology), Shenzhen, People’s Republic of China; 2Department of Hematology, Yantian District People’s Hospital, Shenzhen, People’s Republic of China; 3Department of Gastroenterology and Hepatology, Guangzhou Digestive Disease Center, Guangzhou First People’s Hospital (School of Medicine of South China University of Technology), Guangzhou, People’s Republic of China; 4Department of Child and Adolescent Psychiatry, Shenzhen Kangning Hospital, Shenzhen, People’s Republic of China; 5Department of Medical Administration, Huizhou Institute of Occupational Diseases Control and Prevention, Huizhou, People’s Republic of China; 6Department of Emergency, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; The First Affiliated Hospital, Southern University of Science and Technology), Shenzhen, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Li-sheng Wang; Jun Yao, Department of Gastroenterology, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; The First Affiliated Hospital, Southern University of Science and Technology), No. 1017, Dongmen North Road, Luohu District, Shenzhen, 518020, People’s Republic of China, Tel +86 755 25533018, Email [email protected]; [email protected]
Abstract: The recent rapid development in the field of extracellular vesicles (EVs) based nanotechnology has provided unprecedented opportunities for nanomedicine platforms. As natural nanocarriers, EVs such as exosomes, exosome-like nanoparticles and outer membrane vesicles (OMVs), have unique structure/composition/morphology characteristics, and show excellent physical and chemical/biochemical properties, making them a new generation of theranostic nanomedicine. Here, we reviewed the characteristics of EVs from the perspective of their formation and biological function in inflammatory bowel disease (IBD). Moreover, EVs can crucially participate in the interaction and communication of intestinal epithelial cells (IECs)-immune cells-gut microbiota to regulate immune response, intestinal inflammation and intestinal homeostasis. Interestingly, based on current representative examples in the field of exosomes and exosome-like nanoparticles for IBD treatment, it is shown that plant, milk, and cells-derived exosomes and exosome-like nanoparticles can exert a therapeutic effect through their components, such as proteins, nucleic acid, and lipids. Moreover, several drug loading methods and target modification of exosomes are used to improve their therapeutic capability. We also discussed the application of exosomes and exosome-like nanoparticles in the treatment of IBD. In this review, we aim to better and more clearly clarify the underlying mechanisms of the EVs in the pathogenesis of IBD, and provide directions of exosomes and exosome-like nanoparticles mediated for IBD treatment.
Keywords: extracellular vesicles, exosome, exosome-like nanoparticles, inflammatory bowel disease, theranostic
Introduction
Inflammatory bowel diseases (IBD) is a term of chronic autoimmune diseases of gastrointestinal tract, typically categorized into one of two subtypes: ulcerative colitis and Crohn’s disease.1 IBD is mainly characterized by intermittent recurrence and quiescence inflammatory remission, and it is apparently associated with increased colorectal cancer risk, high morbidity, and decreased life quality.2–4 Ulcerative colitis is confined to the colon, extend from the rectum to the cecum in a contiguous manner, shows superficial mucosal inflammation, and results in ulceration, severe bleeding, toxic megacolon, and fulminant colitis.5,6 In contrast, Crohn’s disease affects any part of the gastrointestinal tract in a discontinuous manner, has transmural inflammation, and contributes to fibrotic stricture, fistulas, and abscesses.7,8 The incidence of IBD is approximately 0.5–24.5 cases per 100,000 person-years for ulcerative colitis and 0.1–16 cases per 100,000 person-years for Crohn’s disease.9,10 Indeed, more than 1.8 million patients in the United States and approximately 3.5 million patients worldwide are suffering from IBD.9,11 Unfortunately, the incidence of IBD is dramatically increased worldwide paralleled by the rise of industrial development, imposing heavy economic burdens both on their families and public healthcare.12
The current knowledge of IBD indicates that it is linked to the interaction of multiple factors, including environmental, genetic, intestinal microbiota, and immune response factors.13–16 It is known that the underlying mechanism of IBD is an inflammatory response caused by the abnormal response of the intestinal mucosal immune system, and the release of a large amount of pro-inflammatory factors.17,18 However, the intestinal immune system is an intricate signaling network, that can protect dietary antigens or beneficial substances, defend against harmful substances, and maintain immune homeostasis.19–21 The intestinal immune system consists of a single layer of epithelial cells linked by tight junctions and intercalated with immune cells. Intestinal epithelial cells (IECs) mainly act as a physical barrier and separate from gastrointestinal lumen contents. The immune cells are composed of the innate immune cells (granulocytes, macrophages, and dendritic cells (DCs)) and adaptive immune cell (B cells and T cells).1 Cell trafficking can prevent the invasion of pathogens and maintain immune homeostasis. When the intestinal immune system is dysregulated, it elicits an abnormally intestinal inflammatory response and IBD. Although the etiology of IBD has been extensively studied, the pathogenesis of IBD is still ambiguous.
The IBD armamentarium mainly depends on non-targeted therapies, such as 5-aminosalicylates, glucocorticoid, and immunosuppressive agents, as well as targeted biologic therapies, such as anti-TNF antibodies (Infliximab (IFX), Adalimumab (ADL), Golimumab (GOLI), and Certolizumab pegol (CZP)), against the p40 subunit of IL-12 and IL-23 (Ustekinumab), and JAK signaling pathway inhibitors (Tofacitinib).22,23 Despite the beneficial effects of these medications, up to 30% of patients do not have a response to initial treatment, and in up to 50% of patients, the response is lost over time.22 Moreover, their side effects can not be ignored, such as allergic reactions and toxicities to healthy organs. Therefore, it is urgent to develop new medications with preferable therapeutic efficacy and limited side effects.
Recently, studies have reported that the extracellular vesicles (EVs), including exosomes, outer membrane vesicles (OMVs), and plant-derived exosome-like nanoparticles (PDENs), released by different types of cells in gastrointestinal tract, gastrointestinal microbiota, and edible plants, play an essential role in the pathogenesis and treatment of IBD.24–26 EVs have been shown to form a novel model of cell trafficking, implicating in cellular signaling to regulate biological processes, such as immunomodulation and regeneration.27–29 Moreover, it has been demonstrated that EVs are critical actors in intercellular communication either at a paracrine level or at a distance. In addition, their cargoes, including lipids, proteins, and RNA, play critical roles in immune system modulation, and possess the ideal delivery system in the treatment of IBD.30–33 Compared with synthetic drug carriers, exosomes and exosomes-like nanoparticles originating from their own cells or diets, such as milk, edible vegetables, and fruits, have considerable advantages of biocompatibility, immune tolerance, and non-toxicity.34–36 Therefore, exosomes and exosome-like nanoparticle-mediated delivery systems have gained considerable attention in treating IBD.
In view of the vital role of EVs in IBD, herein we comprehensively reviewed recent advances in exosomes, PDENs, and OMVs in the pathophysiology of IBD, summarized exosomes or exosome-like nanoparticle-mediated delivery system in the treatment of IBD, and predicted future directions for research efforts of IBD.
Biogenesis, Composition, Uptake and General Functions of Exosomes
Exosomes are bilayer vesicular nanoparticles with heterogeneity and nano-size (30–150 nm), representing a component of a broader class of EVs released by cells due to environmental stimulation or self-activation (Figure 1).37–39 In the beginning, exosomes are simply regarded as inconsequential “garbage” via a process of the cells eliminating unwanted cellular components into the extracellular environment.40 However, more recent studies have found that exosomes play important roles in the physiological and pathological processes, as well as essential mediators of cell-cell communication.40–42 In this section, we briefly discussed the characterization of exosomes.
 |
Figure 1 Biogenesis and composition of extracellular vesicles. (A) Exosomes are formed by the inward budding of endosomes into multivesicular bodies (MVBs) then formation of intraluminal vesicles (ILVs). MVB either fuses directly with lysosomes, or fuses with autophagosomes during autophagy, or which can secrete into the extracellular space as exosomes through fusion with cellular plasma membrane. RAB family members, such as Rab11, Rab27 and Rab35, have vital role in the exosome secretion through regulating MVBs trafficking and docking at the plasma membrane. In addition, soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) protein can induce the exosome secretion by recruiting MVBs docking at the plasma membrane and initiating membrane fusion. (B) The contents of released exosomes include proteins, DNA, mRNA, and miRNA. |
The Biogenesis of Exosomes
Exosomes are derived from endosomal structures that take place via an endocytosis of invaginated endosomes from the plasma membrane43 (Figure 1). Moreover, the formation and release of exosomes depend on an endosomal sorting complex required for transport (ESCRT)-dependent and ESCRT-independent machinery44 (Figure 1). RAB family members, such as Rab11, Rab27 and Rab35, play a vital role in the exosome secretion through regulating multivesicular bodies (MVBs) trafficking and docking at the plasma membrane.45 In addition, soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) protein can induce the exosome secretion by recruiting MVBs docking at the plasma membrane and initiating membrane fusion.46 However, the processes underlying exosome biogenesis are intricate, and still not fully understood.43,47
The Composition of Exosomes
Exosomes have been regarded as an evolutionarily conserved set of particles from parental cells, and are composed of various components, mainly including proteins, lipids, and nucleic acids43,48 (Figure 1). To date, a large variety of constitutive components have been identified in exosomes from different types of cells, and approximately 9769 proteins, 1116 lipids, and 6246 nucleic acids are obtained in the ExoCarta exosomes database (http://www.exocarta.org), indicating their complexity and potential functional diversities. However, the cargoes of exosomes are not completely unraveled, which needs to be understood in further studies.
The Uptake and Function of Exosomes
Exosome uptake may involve three mechanisms: interacting receptor-ligand, fusing with the plasma membrane of the target cell, and getting internalized by target cells.49–51 Scientists have recently discovered that exosomes play a crucial role in cell differentiation, programmed cell death, disease development and recovery, immune response, and inflammation by transporting cargoes of proteins, nucleic acids, and lipids (Figure 1).52–54 In brief, exosomes can communicate with target cells by the above-mentioned mechanisms, fuse with the plasma membrane, and release bioactive cargoes directly into the cytoplasm, leading to activation of the relevant signaling pathway and regulation of the specific gene expression. Nevertheless, the underlying mechanisms of exosomes communicating with target cells remain largely undetermined.
Advantages of Exosomes as Drug Delivery Systems
Exosomes are continuously released into our bodily fluids as part of an intercellular communication network. They are, therefore, attracting increasing attention to improve the treatment of human diseases that are difficult to cure using conventional medicines or methods.55,56 The advantages of exosome-based drug delivery systems are largely realized: (1) The exosome membrane consists of two compartments, an aqueous core and a lipophilic shell formed by a lipid bilayer, making it possible to compartmentalize and solubilize both hydrophilic and hydrophobic materials.40,55 (2) Exosomes are coated by various cell membranes from our own body’s cells, such as red blood cells (RBCs), platelets, white blood cells (WBCs) or cancer cells, and have displayed good biocompatibility, prolonged circulation, and tissue-targeting capacity.57 (3) They are small enough and capable of crossing physiological barriers, such as the gastrointestinal barrier and blood-brain barrier, thus making it possible for targeted drug deliver.56 Currently, a lot of exosome-based drug delivery systems have been completed in early clinical trials, most of which are concentrated in the field of cancer, inflammation, and autoimmunity disease.58–60 Nonetheless, the exosome-based drug delivery systems are challenged their widespread application, such as undetermined toxicity, undefined component, and commercial pharmaceutic productions.61–63
Sources of Exosomes
The exosomes are ubiquitously present.64 Therefore, this section will start with a brief description of the sources of exosomes involved in the pathogenesis and treatment of IBD.
Mesenchymal Stem Cell (MSCs)-Derived Exosomes
MSCs have unique biomedical properties due to their stemness, including differentiation, self-renewal, and colony formation. Moreover, MSCs are multipotent progenitor cells that can be isolated from bone marrow, umbilical cord tissue, amniotic fluid, adipose tissue, and dental pulp.65,66 Besides, MSCs express low levels of major histocompatibility complex (MHC) class I molecules and MHC class II molecules, which empower them to have low immunogenicity.67 Importantly, MSCs have diverse functions, such as anti-inflammatory, anti-apoptotic, and neuroprotective actions, when producing and releasing a broad range of bioactive molecules (Figure 2).68,69,70,71 In addition, MSCs are amongst the largest cellular procedures of exosomes, which have amounts of cargoes, including 850 gene products and 150 miRNAs described, and play a vital role in the regulation of inflammation, metabolic disorders, and cell damages.72–74 Accumulating evidence has demonstrated that MSC-derived exosomes are considered to be a potential option for the treatment of human disease, such as Alzheimer’s disease (AD), osteoarthritis, and autoimmune disease.75–77 Although MSC-derived exosomes show increasing potential for human disease, there is still a lack of evidence. Further research should focus on their functions and sorting mechanisms. Moreover, future clinical trials should be performed to verify the efficacy and safety of MSC-derived exosomes and establish a comprehensive theoretical basis for the clinical implementation.
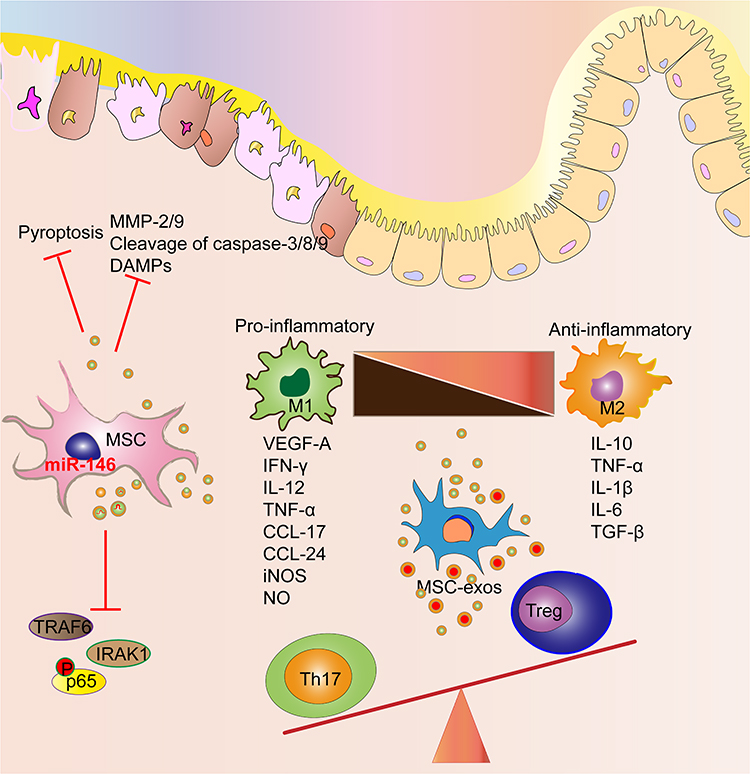 |
Figure 2 MSCs-Exo regulate the intestinal barrier, immune response and inflammatory responses, which provide an effective therapeutic strategy for IBD. |
Plant-Derived Exosome-Like Nanoparticles (PDENs)
Despite a growing appreciation of the importance of exosomes in mammals, PDENs are detected earlier than mammal cell-derived exosomes.78 Exosome-like nanoparticles of plant cells can be released through multiple pathways. For example, the fusion of MVBs with the plasma membrane (PM) can release intraluminal vesicles (ILVs) as exosomes; the vacuole fuses with the PM as a vacuolar pathway to release exosomes; and exocyst-positive organelle (EXPO) can also direct release exosomes (Figure 3). PDENs are mostly isolated from edible plants, such as ginger, broccoli, Citrus sinensis, lemon, and grapefruit (Figure 3).79–83 Recent evidence has demonstrated that PDENs can be delivered to other organs through blood flow and function distantly in the recipient cells. Moreover, PDENs have gained huge attention due to their potential to regulate physiological and pathological processes, and develop therapeutic vehicles.84,85 Although clinical trials are performed to investigate the efficacy and safety of PDENs for the treatment of human diseases, the number of studies is limited (such as NCT01294072, NCT01668849, and NCT03493984). Of note, there is no standard method for isolation, purification, and production of PDENs in clinical practice. Therefore, further research should formulate the protocols for isolating PDENs and perform significant clinical trials.
 |
Figure 3 Biogenesis of plant-derived exosome-like nanoparticles and therapeutics application for IBD. (A) Vacuolar pathway; (B) MVBs pathway; (C) Exocytosis pathway by exocyst-positive organelle (EXPO). |
Milk-Derived Exosomes
Milk is a natural drink, has a large content of protein, fat, vitamins, and other nutrients, and can be used to supplement nutrition in daily life.86,87 Meanwhile, milk also plays an essential role in organism development and immune response.88 Milk-derived exosomes are vital components of milk, released from mammary gland epithelial cells of all mammals including human and dairy cows.89,90 Accumulating evidence indicates that milk-derived exosomes play a crucial role in the appropriate maturation of the intestine and development of the gut microbiome, while deficiency of milk-derived exosomes contributes to metabolic and immunological disease in the newborn infants.91–93 Moreover, increasing studies show that milk-derived exosomes, as drug delivery systems, are conducted for the treatments of various diseases, including IBD.94,95 The details of milk-derived exosomes in the treatment of IBD will be discussed in the section about the therapeutic effects of exosomes on IBD.
Immune Cell-Derived Exosomes
The immune cells consist of innate cells (including granulocytes, macrophages, and DCs) and adaptive immune cells (including B cells and T cells).96 Accumulating evidence has implicated that the immune cells secret exosomes, and then engage in the transfer of information.97 A great deal of attention has been paid to the role of immune cell-derived exosomes both in healthy physiological and pathological processes.97–99 For example, B cell-derived exosomes bind with MHC class II, and induce antigen-specific T cell responses.100 DC-derived exosomes can trigger regulatory T cells (Treg) activation while inhibiting T helper type 1 (Th1) cells, which modulate immune responses in IBD (Figure 4).31 The function of immune cell-derived exosomes in the IBD will be discussed in the section on exosomes and IBD.
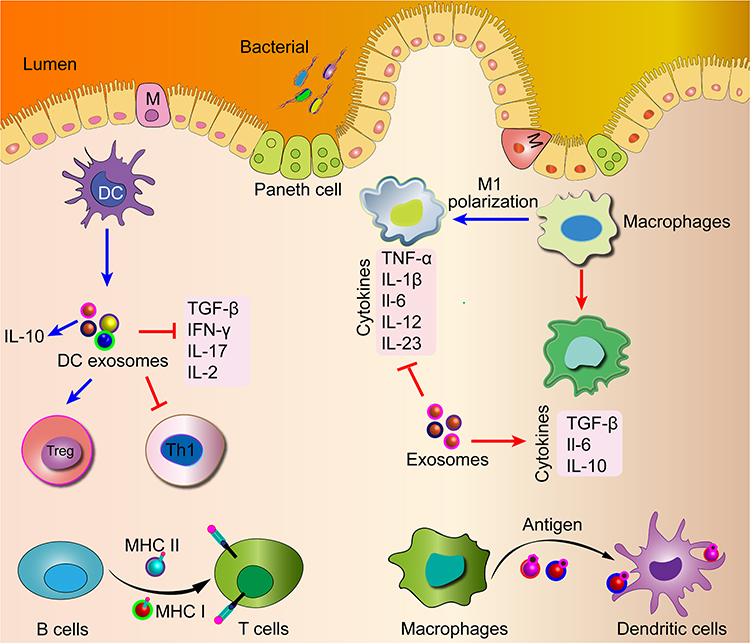 |
Figure 4 Exosomes derived from immune cells can modulate immune responses in the pathogenesis of IBD and also have been used in vaccine and drug delivery research. |
Exosomes in Body Fluids
Exosomes have been isolated from various physiological fluids, such as blood,101 urine,102 saliva,103 amniotic fluid,104 and breast milk.105 Recently, exosomes can be used as biomarkers and indicators in diagnosing diseases, including IBD, due to their extensive existence in the above-mentioned biological fluids.30,106–108 Shao et al have reported that a serum exosome, pregnancy zone protein (PZP), is up-regulated in the IBD patients, and can be regarded as a promising biomarker of serological detection for IBD diagnosis.108 Zhang et al have indicated that a salivary exosome, proteasome subunit alpha type 7 (PSMA7), is present at high levels in IBD patients and may be a promising indicator for IBD diagnosis.109 Unfortunately, there are not many studies of body fluid-derived exosomes until now. Therefore, additional research on the body fluid-derived exosomes in IBD patients will be beneficial for a better understanding.
EVs in the Pathogenesis of IBD
EVs play crucial roles in the pathogenesis of IBD. Their cargoes not only facilitate or alleviate the pathogenesis of IBD, but also can be used as diagnostic and therapeutic markers.41,108–110 In this section, we will discuss the EVs derived from IECs, immune cells, and gut microbiota function in the pathogenesis of IBD.
Intestinal Epithelial Cells (IECs)-Derived Exosomes in IBD
IECs release exosomes to communicate with host immune cells and gut microbiota, and help maintain a homeostatic environment in the gut. The target and function of IECs-derived exosomes are to stimulate DCs, regulatory T cells (Treg), and macrophage maturation with tolerogenic properties by immunoregulatory pathways, and maintain intestinal homeostasis (Figure 5).24,25,31 For instance, an IECs-derived exosome, TGF-β1, can alleviate colonic inflammation in experimental mouse colitis through stimulating immunosuppressive DCs and Treg cells.111 Interestingly, an IEC-derived exosome, αβ6, activates LTGFβ in intestinal tolerogenic DCs, and Treg cells, and produces TGF-β1.112 In addition, an IEC-derived exosome, ANXA1, can activate the wound repair circuit, and promote the repair of intestinal mucosal wound in experimental mouse colitis.113 Although IECs are not primarily antigen-presenting cells (APCs), the IEC-derived exosomes can participate in antigen presentation within the intestinal mucosa and immunoregulatory process due to their expression of major histocompatibility complex I (MHC-1), MHC-II, and human leukocyte antigen-DM (HLA-DM).114 For example, MHC-II can activate B cell development, consolidate B cell maturation, and initiate adaptive immune.115
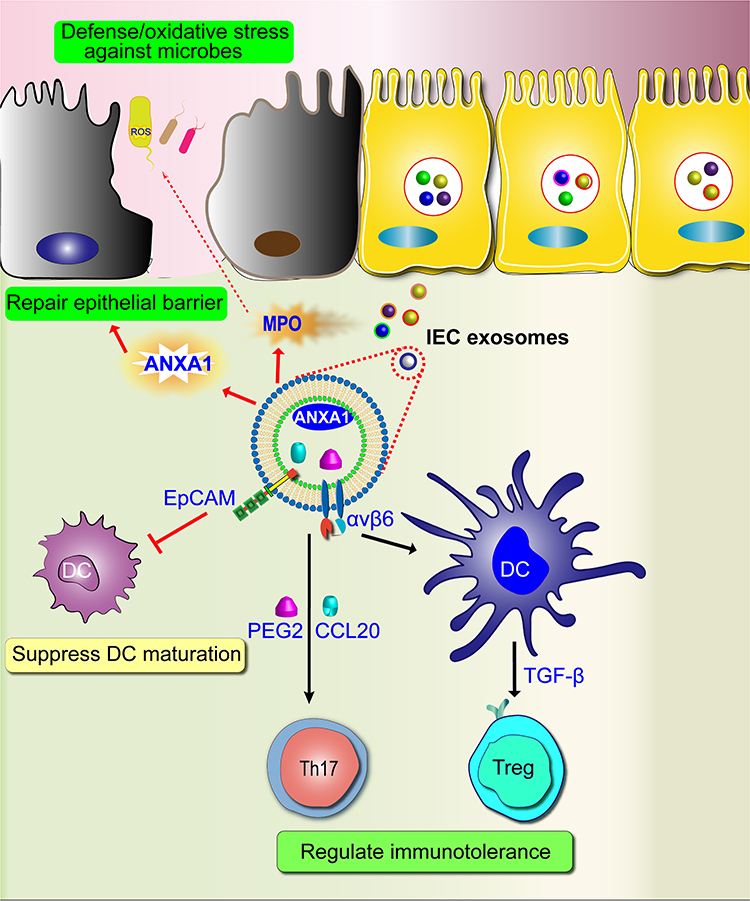 |
Figure 5 Function of IECs-derived exosomes in IBD. IECs-derived exosomes can promote antigen presentation, increases intestinal immunotolerance, repair epithelia barrier. |
In the pathogenesis of IBD, an IECs-derived exosome, epithelial cell adhesion molecular (EpCAM), is greatly elevated, induces DC apoptosis, suppresses DC maturation, and inhibits the functions of DCs (Figure 5).116 IEC-derived exosomes, such as GSDMD, IL-1β, and caspase-8, induce IECs pyroptosis, produce pro-inflammatory cytokines, and promote intestinal inflammation in experimental colitis model.117 However, myeloperoxidase (MPO), derived from IEC exosomes in IBD patients, as a defense protein, can induce the oxidation reaction by producing reactive oxidants.118 Exosomes isolated from IECs infected with adherent-invasive Escherichia coli can activate nuclear factor-κB (NF-κB) and MAPK pathways in naïve macrophages, resulting in an increased secretion of pro-inflammatory cytokines IL-6 and TNF-α.119 Of note, the important role of IEC-derived exosomes is to regulate the pro- and anti-inflammatory immune balance. Once the balance is disrupted in the intestinal immune system, it will lead to intestinal inflammation. Therefore, they may be promising targets for the treatment of IBD.
Immune Cells-Derived Exosomes in IBD
Accumulating studies have indicated that both the innate and adaptive immune responses participate greatly in the IBD pathogenesis.120,121 The innate immune response is quicker to trigger the phagocytic responses and antigen presentation, accompanied by the initiation of the adaptive immune system.121 This pertains to most immune cells, such as macrophages, DCs, neutrophils, and monocytes (Figure 4).121
Macrophages play a pivotal role in the maintenance of intestinal homeostasis and the pathogenesis of IBD (Figure 4).122–124 Macrophages are classified into two subsets with seeming different functions: M1 macrophages secrete higher levels of pro-inflammatory cytokines including TNF-α, IL-1α, IL-12, and IL-6. On the contrary, M2 macrophages produce higher levels of anti-inflammatory cytokine of IL-10.122–124 Macrophage-derived exosomes are involved in the pathogenesis of IBD. An M1 macrophages-derived exosome, miR-21a-5p, can suppress the expression of E-cadherin, then promote ILC2 activation, and result in the destruction of the intestinal mucosal epithelium in experimental mouse colitis.125 Yang et al have reported that M2 macrophage-derived exosomes can suppress the expressions of pro-inflammatory cytokines including IL-1β, IL-6, and IL-17A, and exert protective effects on DSS-induced colitis.126 Moreover, an M2 macrophage-derived exosome, miR-590-3p, can inhibit the secretion of pro-inflammatory cytokines, such as IL-1β, IL-6, and TNF-α, promote epithelial wound repair, and attenuate DSS-induced colitis.127 In addition, MSC-derived exosomes can attenuate colitis by suppressing the release of colon M1 macrophages and pro-inflammatory cytokines, and promoting the polarization of M2 macrophages.128 Therefore, the balance of M1/M2 macrophages may be an effective treatment strategy for IBD.
DCs are the most effective and professional APCs of the colonic immune system, which can capture antigens, and initiate an immune response, and DCs-derived exosomes may possess immune-stimulatory or suppressive effects (Figure 4).124 DC-derived exosomes, CD80 and CD86, as costimulatory markers, can trigger naïve CD4+ T activation, thus amplifying the initiation of primary adaptive immune responses.129 Moreover, a DC-derived exosome, TGF-β1, possesses strong immunosuppressive activity by inducing Treg cells in IBD mouse model.130,131 In addition, DC-derived exosomes treated with IL-10 can attenuate 2,4,6-trinitrobenesulfonic acid (TNBS)-induced mouse colitis by inhibiting the expressions of TNF-α, IL-2, and IFN-γ and promoting the expressions of IL-10 and Treg cells.132 It is conceivable that DC-derived may be a promising target for the therapeutic intervention of IBD.
Other immune cell-derived exosomes can also influence IBD progression in different ways (Figure 4). Neutrophil-derived exosomes can contribute to the decrease of epithelial cadherins and promote neutrophil recruitment in colitis.133 Treg cell-derived exosomes can promote IEC proliferation, inhibit IEC apoptosis, and protect the intestinal barrier in IBD mouse model.134,135 Granulocytic myeloid-derived exosomes can alleviate IBD by diminishing the proportion of Th1 cells, and promoting the expansion of Treg cells.136 These immune cell-derived exosomes may be the new therapeutic intervention for IBD in the future.
Gut Microbiota-Derived OMVs in IBD
Gut microbiota-derived EVs also play a crucial role in the immunomodulation and intestinal barrier integrity of IBD.137,138 The gut microbiota commonly includes bacteria, fungi, and viruses.139 In the present review, we will mainly focus on the bacterial extracellular vesicles (BEVs) in the IBD. Typical Gram-negative bacteria are released when the outer membrane lipid asymmetry leads to blebbing, resulting in the production of BEVs known as outer membrane vesicles (OMVs). BEVs are rich in various outer membrane proteins, such as DNA, RNA, virulence factors, membrane-bound proteins and polysaccharides, and peptidoglycans (Figure 6). Under physiological conditions, the gut microbiota interacts with the host, protects the intestinal barrier, and maintains intestinal immune homeostasis. For instance, Lactobacillus rhamnosus GG-derived EVs can protect the colonic tissue damage and alleviate intestinal inflammation via inhibiting the activation of TLR4-NF-κB-NLRP3 axis.140 Akkermansia muciniphila-derived EVs can have a protective function on colitis, and ameliorate intestinal inflammation in the DSS-induced colitis model.141 However, the microbial diversity is prominently changed in the pathogenesis of IBD, indicating that the proportion of bacteria with anti-inflammatory properties is lower, whereas the proportion of bacteria with pro-inflammatory properties is relatively higher.142 Subsequently, alterations in the composition of intestinal bacteria can influence intestinal homeostasis and affect the intestinal barrier by these bacteria-derived exosomes mediating cross-talk between microbiota, intestinal epithelia, intestinal immune cells, and mucosal immunity. For example, EVs are derived from Fusobacterium nucleatum (Fn), a Gram-negative specific anaerobe, that can aggravate intestinal barrier disruption by promoting the differentiation of pro-inflammatory macrophages and accelerating IEC necroptosis.143 Escherichia coli (E. coli)-derived OMVs, peptidoglycan, can increase the expressions of pro-inflammatory cytokines including NF-κB, IL-6, and IL-8, via activating nucleotide-binding oligomerization domain containing 1 (NOD1) signaling pathway in IECs.144 Notably, exploring the function of probiotics and pathogenic bacteria, and balancing probiotics and pathogenic bacteria will be helpful in clarifying the relationship between gut microbiota and IBD, and identifying new therapeutic targets for IBD patients.
 |
Figure 6 Schematic illustrate origin and composition of bacterial extracellular vesicles. The immunomodulatory effects and Inflammatory pathway network elicited by microbiota‐derived BEVs in the gut were summarized. |
Therapeutic Effects of EVs on IBD
It is clear that EVs are involved in intestinal intercellular communication and the pathogenesis of IBD suggesting that EVs can be promising therapeutic options for IBD. The interest in EV research also propels the development of artificial nanoparticles, exosome-like nanoparticles, which are employed in the treatment of IBD. The application of IECs, intestinal immune cells, gut microbiota-derived exosomes, and OMVs in the treatment of IBD has been discussed in EVs in the Pathogenesis of IBD. Therefore, in this section, we will discuss MSC-, plant-, and milk-derived exosomes and exosome-like nanoparticles in the treatment of IBD.
Methods for Loading Drugs into EVs
Efficient loading of therapeutic cargo packaging into exosomes and exosome-like nanoparticles is a very critical step.145,146 Currently, different methods are commonly utilized for loading biological cargoes into exosomes and exosome-like nanoparticles including co-incubation, electroporation, and sonication (Table 1).147 Co-incubation is a passive cargo loading technique, and the drugs move through a concentration gradient into the extracted exosomes and exosome-like nanoparticles.148 In addition to co-incubation, the electroporation method uses an electric field to form temporary hydrophilic pores on the phospholipid membrane of the exosomes and exosome-like nanoparticles to load the biological cargoes.149 Besides, sonication is an alternative method of loading drug into exosomes and exosome-like nanoparticles through drugs adhering to the surface of exosomes and exosome-like nanoparticles.94,150 However, there is a lack of horizontal comparison of the advantages and disadvantages of these loading methods in the current research.
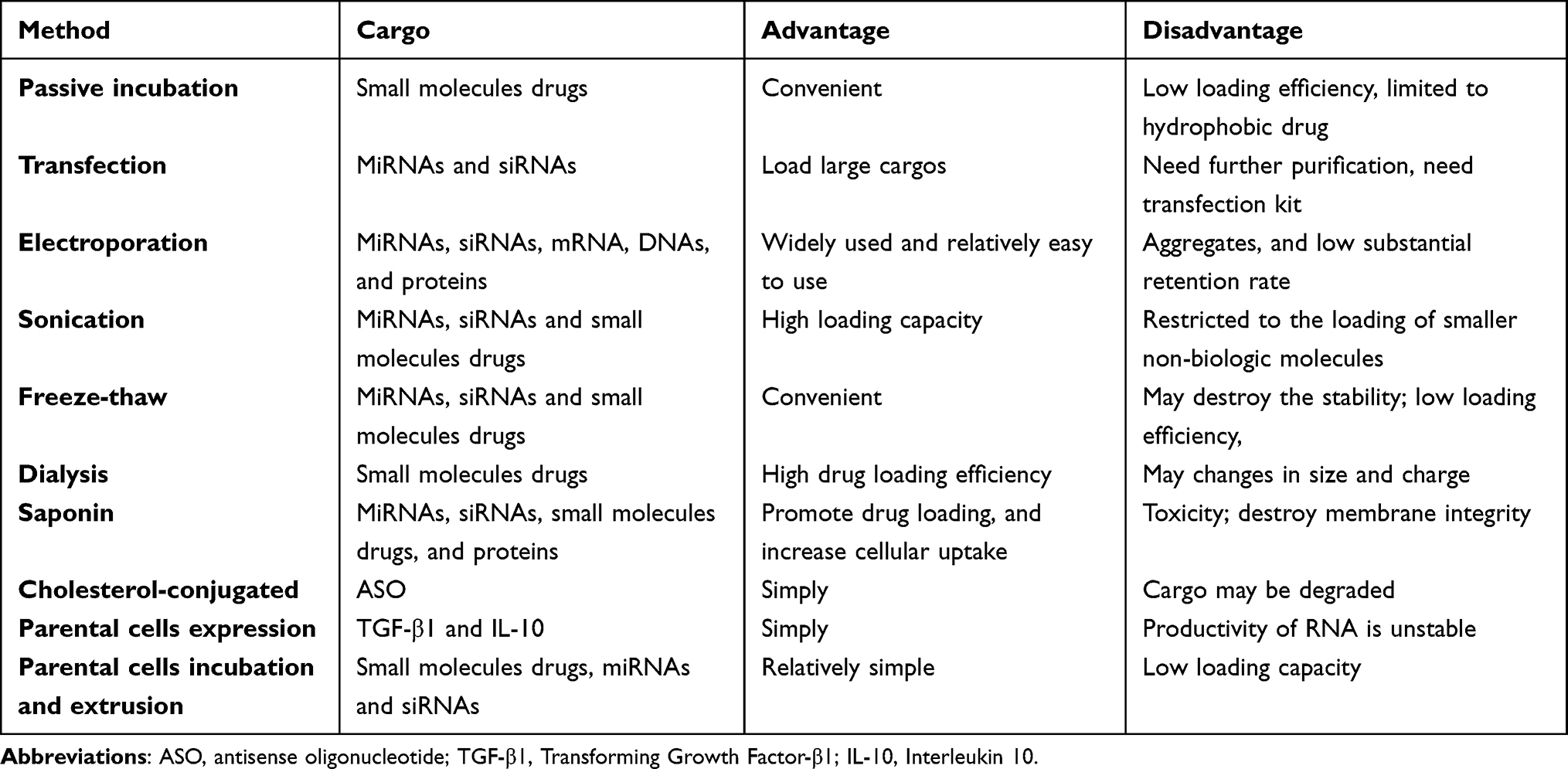 |
Table 1 Exosome Cargo Loading Method |
Therapeutic Effects of MSC-Derived Exosomes Against IBD
Currently, MSC-derived exosomes have exhibited great potential in the treatment of IBD (Table 2). For example, human umbilical cord-derived MSC (HucMSC)-derived exosomes can promote the expressions of anti-inflammatory cytokine, such as IL-10, inhibit the levels of pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6, and alleviate DSS-induced colitis.151 Moreover, human bone marrow MSC (hBM‐MSC)-derived exosomes can upregulate the expression of anti-inflammatory factors, such as IL-10, IL-10, and TGF-β, decline the levels of pro-inflammatory factors, VEGF-A, IFN-γ, IL-12, TNF-α, CCL-24, and CCL-17, and ameliorate DSS-induced colitis by promoting the polarization of M2 macrophages.152 In addition, human adipose MSC (hADSC)-derived exosomes can protect the intestinal barrier integrity, promote the IEC proliferation, and resist the intestinal inflammatory injury in DSS-induced colitis.153 Moreover, MicroRNA-146, is a well-known anti-inflammatory miRNA, and exosomes obtained from MSCs overexpressing miR-146, can regulate NF-κB p65 phosphorylation and inhibit the expressions of TNF receptor-associated factor 6 (TRAF6) and IL −1 receptor-associated kinase 1 (IRAK1), thereby inhibiting the release of inflammatory factors in macrophages, and reducing colonic inflammation29 (Figure 2). Collectively, MSC-derived exosomes can protect the colitis mice model by restoring mucosal barrier repair and maintaining intestinal immune homeostasis.154–157 Excitedly, a randomized, double-blind controlled trial has demonstrated that MSCs-derived exosomes are an effective and safe option to treat complex perianal fistulas in patients with Crohn’s disease who do not respond to conventional or biological treatments, or both.158 Subsequently, MSC-derived exosomes are considered an effective therapeutic approach for IBD patients in clinical practice.159,160 Therefore, MSC-derived exosomes may provide an effective therapeutic strategy for IBD treatment.
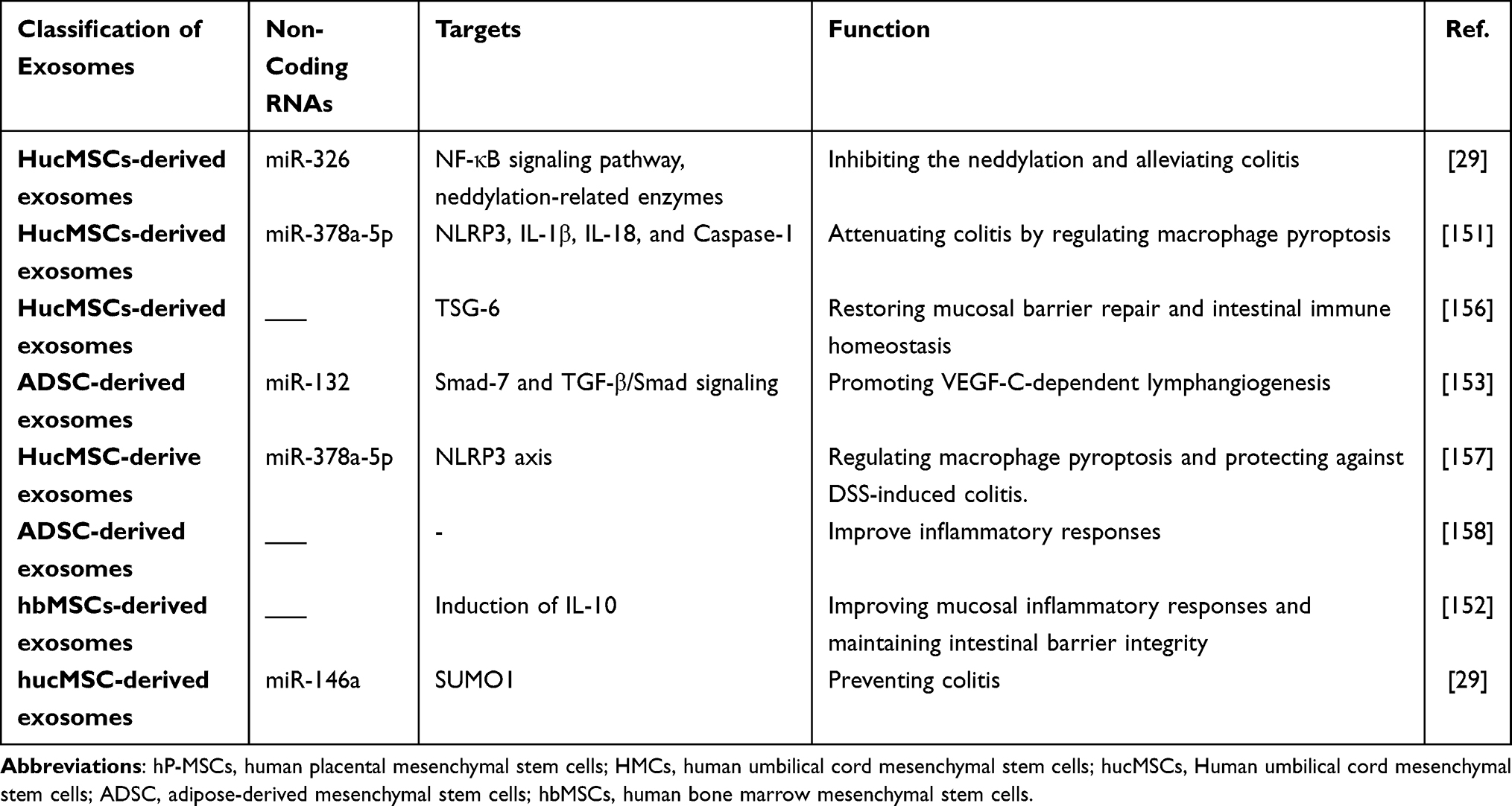 |
Table 2 MSCs-Derived Exosomes for IBD Therapy |
Therapeutic Activity of PEDNs Against IBD
The edible PEDNs can also relieve intestinal inflammation and maintain intestinal immune homeostasis (Table 3). For instance, oral administration of tea leaf-derived exosome-like nanoparticles can decrease the levels of pro-inflammatory cytokines (TNF-α, IL-6, and IL-12), increase the amount of anti-inflammatory cytokine IL-10, restore disrupted intestinal barriers, and alleviate experimental mouse colitis.34 Moreover, grape exosome-like nanoparticles can mediate impaired intestinal tissue remodeling, and protect against DSS-induced colitis by strongly promoting the proliferation of Lgr5hi intestinal stem cells.161 In addition, broccoli-derived exosome-like nanoparticles can not only inhibit activation of intestinal DCs, but also induce tolerogenic DCs via activating the AMPK signaling pathway, contributing to prevent DSS-induced colitis in mice.80 Furthermore, Zhang et al have discovered that oral administration of ginger-derived exosome-like nanoparticles can reduce the susceptibility of mice to DSS-induced colitis by increasing the survival and proliferation of IECs and promoting pro-healing factors including interleukin (IL)-10 and IL-22.162 Recently, Sriwastva et al have found that mulberry bark-derived exosome-nanoparticles can activate aryl hydrocarbon receptor (AhR) signaling and increase the amount of anti-microbial peptides (AMPs) in IECs, inhibiting intestinal inflammation, and altering gut microbiota composition in experimental mouse colitis.163 Additionally, oral administration of turmeric-derived exosome-like nanovesicles can ameliorate mouse colitis and accelerate colitis resolution via decreasing the expressions of pro-inflammatory cytokines, including TNF-α, IL-6, and IL-1β, and increasing the levels of the antioxidant gene, heme oxygenase-1 (HO-1) in mice colitis models.164 Unfortunately, clinical trials are limited to the efficacy and safety of PDENs for IBD patients.
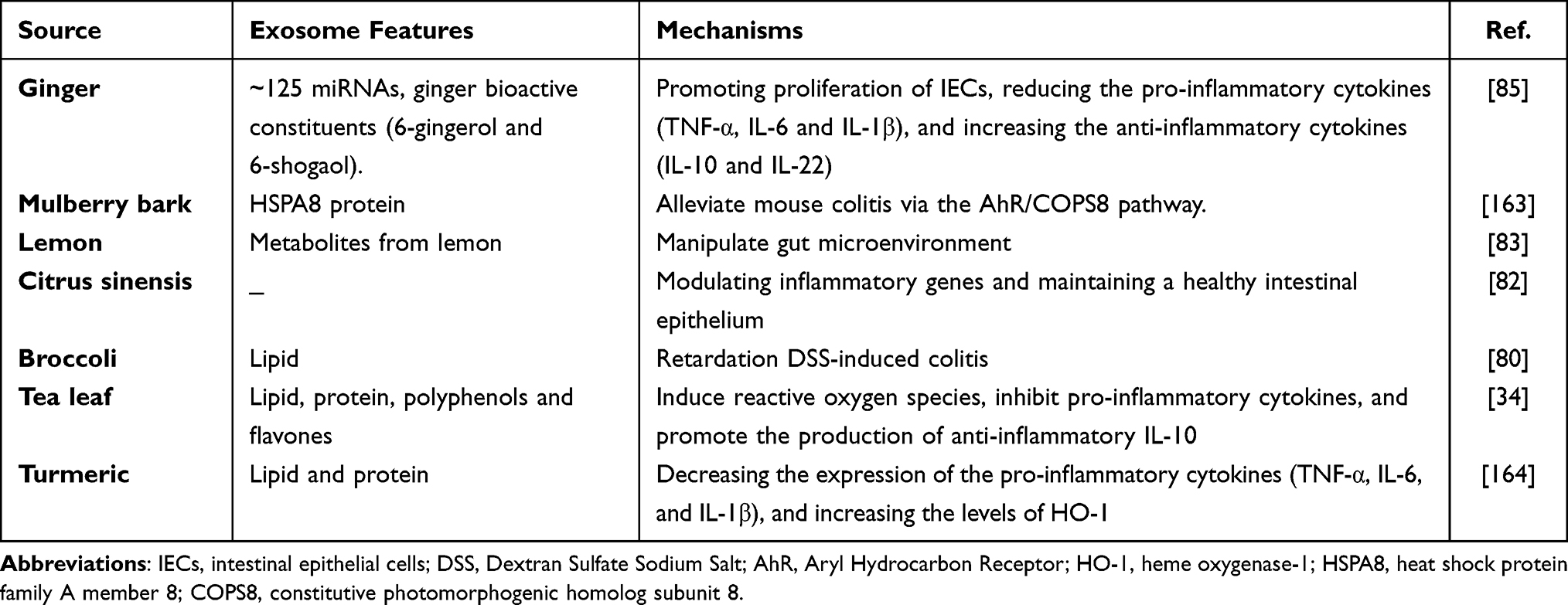 |
Table 3 Features of Plant-Derived Exosome-Like Nanoparticles Regulate Intestinal Microenvironment |
The Therapeutic Effect of Milk-Derived Exosomes on IBD
Milk-derived exosomes play an important role in the development of the digestive tract, which regulate intercellular signaling, inflammation and immune response, thus protecting against stress and various disease conditions including IBD.88,165,166 Milk-derived exosomes contain lipids, nucleic acid, and proteins, exerting biological function,167 and they can be used as an oral natural drug delivery system in the treatment of various diseases including IBD.94 MiR-148a, a milk-derived exosome cargo, plays a key role in promoting intestinal maturation, sustaining barrier function, and suppressing the activity of NF-κB.88 Moreover, insulin-like growth factor-1 (IGF-1), as an important milk-derived exosome cargo, can promote IEC proliferation, improve intestinal barrier function, and protect IEC from intestinal injury.168,169 In addition, milk-derived exosomes can restore the expression of the protein zonula occludens 1 (ZO-1), and decrease the intestinal permeability barrier in the DSS-induced mouse colitis.170 Recently, Reif et al have reported that oral administration of cow and human milk-derived exosomes can attenuate the severity of DSS-induce mouse colitis by down-regulating pro-inflammatory cytokines TNF-α and IL-6.171 We have previously demonstrated that the NF-κB signaling pathway is activated, and link with the pathogenesis of ulcerative colitis.172 In a recent study, oral administration of milk-derived exosomes can reduce intestinal epithelium disruption, inhibit infiltration of inflammatory cells and alleviate colonic inflammation by inactivating NF-κB signaling pathway in the ulcerative colitis mice model.35 On the other side, it has been demonstrated that milk-derived exosomes can deliver both hydrophilic and lipophilic small molecules including chemo drugs.173 For instance, milk-derived exosomes, as a drug delivery system for loading curcumin can resist degradation by human digestive enzymes and possess enhanced intestinal permeability in vitro.174 Although milk-derived exosomes have several advantages, such as no adverse immune, no inflammatory response, and considerable oral bioavailability, how to maintain their stability in the gastrointestinal fluid has not yet been fully understood. Indeed, the technique of scalability and stability of milk-derived exosomes has witnessed rapid progress over the past few years,175 and they may provide a promising treatment strategy for IBD in the future.
Conclusions and Future Perspective
Accumulating evidence suggests that EVs play a pivotal role in the pathogenesis of IBD, and a better understanding of them can clarify the underlying mechanism in IBD, providing new insights into the therapeutic strategies of IBD. In the present review, we systematically discussed the current knowledge about EVs in the pathogenesis of IBD, and the feasibility of the application of exosomes and exosome-like nanoparticles for IBD treatment. Under normal condition, IEC-, intestinal immune cell- and gut microbiota-derived EVs primarily function to defend against pathogens, modulate the immune response, and maintain the intestinal hemostasis. Nevertheless, under pathological conditions, IECs, intestinal immune cells, and gut microbiota also secret EVs and intervene in the pathological mechanisms of IBD, such as EpCAM, CD80, CD86, and so on. In addition, EVs crucially participate in the interaction and communication of IEC-immune cells-gut microbiota in the intestinal hemostasis. Therefore, once the imbalance is disrupted, it may contribute to immune response, intestinal inflammation and intestinal dysbacteriosis. However, further studies are required to identify the chemical contents of the EVs, and help clarify the pathogenesis of IBD and provide new therapeutic targets.
The use of exosomes and exosome-like nanoparticles as alternative modalities for IBD treatment is deemed to be safe, available, and cost-effective. The exosomes and exosome-like nanoparticles isolated from the edible plants, MSCs, milk and other cells can protect the intestinal barrier, maintain gut microbiota, and attenuate experimental mouse colitis. However, most current knowledge on exosomes and exosome-like nanoparticles for IBD treatment mainly comes from in vitro experiments and animal models of DSS-induced colitis. Although exosomes and exosome-like nanoparticles can be used as oral drug delivery vehicles due to their good biodistribution and inherent biocompatibility, how to target deliver the drug encapsulated by exosomes and exosome-like nanoparticles to the lesion site is still challenging. Moreover, clinical trials of exosomes and exosome-like nanoparticles for the treatment of IBD are still limited, and their administration route, dosage, and adverse events should be investigated before entering the clinical application.
Abbreviations
IBD, Inflammatory bowel diseases; EVs, extracellular vesicles; PDENs, plant-derived exosome-like nanoparticles; OMVs, outer membrane vesicle; PM, plasma membrane; TGF-β, transforming growth factor β; IL-10, interleukin-10; IFX, Infliximab; ADL, Adalimumab; GOLI, Golimumab; CZP, Certolizumab pegol; EV, extracellular vesicle; ILVs, intraluminal vesicles; ESCRT, Endosomal Sorting Complex Required for Transport; MVBs, multivesicular bodies; SNARE, soluble N-ethylmaleimide-sensitive factor attachment protein receptor; PS, phatidylserine; RBCs, red blood cells; WBCs, platelets, white blood cells; MSCs, mesenchymal stem cells; MHC, major histocompatibility complex; AD, Alzheimer’s disease; IECs, intestinal epithelial cells; GDM, gestational diabetes mellitus; TDEs, tumor-derived exosomes; ANXA1, annexin A1; IGF-1, insulin-like growth factor-1; NF-κB, nuclear factor-κB; ZO-1, zonula occludens 1; PZP, pregnancy zone protein; PSMA7, proteasome subunit alpha type 7; TME, tumor microenvironment; DCs, dendritic cells; Treg, regulatory T cells; APCs, antigen-presenting cells; MHC-1, major histocompatibility complex I; HLA-DM, human leukocyte antigen-DM; EpCAM, epithelial cell adhesion molecular; MPO, myeloperoxidase; IL, interleukin; TNBS, trinitrobenesulfonic acid; NOD1, nucleotide-binding oligomerization domain containing 1; HucMSCs, human umbilical cord-derived mesenchymal stem cells; hBM‐MSCs, human bone marrow‐derived mesenchymal stem cells; hADSCs, human adipose tissue derived MSCs; GELNs, grape exosome-like nanoparticles; AhR, aryl hydrocarbon receptor; AMPs, anti-microbial peptides; MDEs, milk-derived exosomes.
Acknowledgments
This work was supported by Guangdong Basic and Applied Basic Research Foundation (No. 2018A0303100024), Three Engineering Training Funds in Shenzhen (No. SYLY201718, No. SYJY201714 and No. SYLY201801), Science and Technology Innovation Committee of Shenzhen (No. JCYJ20150403101028164, No. JCYC20170307100911479, No. JCYJ20190807145617113 and JCYJ20210324113802006), National Natural Science Foundation of China (No. 81800489).
Author Contributions
Rui-yue Shi, Li-sheng Wang and Jun Yao share co-corresponding authorship. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflict of interest.
References
1. Chang JT. Pathophysiology of inflammatory bowel diseases. N Engl J Med. 2020;383:2652–2664. doi:10.1056/NEJMra2002697
2. Francescone R, Hou V, Grivennikov SI. Cytokines, IBD, and colitis-associated cancer. Inflamm Bowel Dis. 2015;21:409–418. doi:10.1097/MIB.0000000000000236
3. Stormont JM, Shah AN, Sharma AK, et al. Colorectal cancer in IBD patients. Am J Gastroenterol. 2013;108:1535. doi:10.1038/ajg.2013.203
4. Hoie O, Wolters F, Riis L, et al. Ulcerative colitis: patient characteristics may predict 10-yr disease recurrence in a European-wide population-based cohort. Am J Gastroenterol. 2007;102:1692–1701. doi:10.1111/j.1572-0241.2007.01265.x
5. Nakase H, Sato N, Mizuno N, et al. The influence of cytokines on the complex pathology of ulcerative colitis. Autoimmun Rev. 2021;21:103017. doi:10.1016/j.autrev.2021.103017
6. Venkateswaran N, Weismiller S, Clarke K. Indeterminate colitis - update on treatment options. J Inflamm Res. 2021;14:6383–6395. doi:10.2147/JIR.S268262
7. Shen B, Kochhar GS, Rubin DT, et al. Treatment of pouchitis, Crohn’s disease, cuffitis, and other inflammatory disorders of the pouch: consensus guidelines from the International Ileal Pouch Consortium. Lancet Gastroenterol Hepatol. 2022;7:69–95. doi:10.1016/S2468-1253(21)00214-4
8. Nayar S, Cho JH. From single-target to cellular niche targeting in Crohn’s disease: intercepting bad communications. EBioMedicine. 2021;74:103690. doi:10.1016/j.ebiom.2021.103690
9. Ng SC, Shi HY, Hamidi N, et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies. Lancet. 2017;390:2769–2778. doi:10.1016/S0140-6736(17)32448-0
10. Bernstein CN, Burchill C, Targownik LE, et al. Events within the first year of life, but not the neonatal period, affect risk for later development of inflammatory bowel diseases. Gastroenterology. 2019;156:2190–2197 e10. doi:10.1053/j.gastro.2019.02.004
11. Nguyen GC, Chong CA, Chong RY. National estimates of the burden of inflammatory bowel disease among racial and ethnic groups in the United States. J Crohns Colitis. 2014;8:288–295. doi:10.1016/j.crohns.2013.09.001
12. Kaplan GG. The global burden of IBD: from 2015 to 2025. Nat Rev Gastroenterol Hepatol. 2015;12:720–727. doi:10.1038/nrgastro.2015.150
13. Kaplan GG. IBD: global variations in environmental risk factors for IBD. Nat Rev Gastroenterol Hepatol. 2014;11:708–709.
14. Putignani L, Del Chierico F, Vernocchi P, et al. Gut microbiota dysbiosis as risk and premorbid factors of ibd and ibs along the childhood-adulthood transition. Inflamm Bowel Dis. 2016;22:487–504. doi:10.1097/MIB.0000000000000602
15. Ocansey DKW, Wang L, Wang J, et al. Mesenchymal stem cell-gut microbiota interaction in the repair of inflammatory bowel disease: an enhanced therapeutic effect. Clin Transl Med. 2019;8:31. doi:10.1186/s40169-019-0251-8
16. Schirmer M, Garner A, Vlamakis H, et al. Microbial genes and pathways in inflammatory bowel disease. Nat Rev Microbiol. 2019;17:497–511. doi:10.1038/s41579-019-0213-6
17. Witkowski M, Witkowski M, Gagliani N, et al. Recipe for IBD: can we use food to control inflammatory bowel disease? Semin Immunopathol. 2018;40:145–156. doi:10.1007/s00281-017-0658-5
18. Nishida A, Inoue R, Inatomi O, et al. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin J Gastroenterol. 2018;11:1–10. doi:10.1007/s12328-017-0813-5
19. Mahapatro M, Erkert L, Becker C. Cytokine-mediated crosstalk between immune cells and epithelial cells in the gut. Cells. 2021;10:111. doi:10.3390/cells10010111
20. Ala M, Ala M. Metformin for cardiovascular protection, inflammatory bowel disease, osteoporosis, periodontitis, polycystic ovarian syndrome, neurodegeneration, cancer, inflammation and senescence: what is next? ACS Pharmacol Transl Sci. 2021;4:1747–1770. doi:10.1021/acsptsci.1c00167
21. Zimmerman NP, Vongsa RA, Wendt MK, et al. Chemokines and chemokine receptors in mucosal homeostasis at the intestinal epithelial barrier in inflammatory bowel disease. Inflamm Bowel Dis. 2008;14:1000–1011. doi:10.1002/ibd.20480
22. Baumgart DC, Le Berre C. Newer biologic and small-molecule therapies for inflammatory bowel disease. N Engl J Med. 2021;385:1302–1315. doi:10.1056/NEJMra1907607
23. Lu Q, Yang MF, Liang YJ, et al. Immunology of inflammatory bowel disease: molecular mechanisms and therapeutics. J Inflamm Res. 2022;15:1825–1844. doi:10.2147/JIR.S353038
24. Zhang H, Wang L, Li C, et al. Exosome-induced regulation in inflammatory bowel disease. Front Immunol. 2019;10:1464. doi:10.3389/fimmu.2019.01464
25. Ayyar KK, Moss AC. Exosomes in Intestinal Inflammation. Front Pharmacol. 2021;12:658505. doi:10.3389/fphar.2021.658505
26. Wang H, Ye C, Wu Y, et al. Exosomes in inflammatory bowel disease: what have we learned so far? Curr Drug Targets. 2020;21:1448–1455. doi:10.2174/1389450121666200428102330
27. Thakur A, Parra DC, Motallebnejad P, et al. Exosomes: small vesicles with big roles in cancer, vaccine development, and therapeutics. Bioact Mater. 2022;10:281–294. doi:10.1016/j.bioactmat.2021.08.029
28. Saad MH, Badierah R, Redwan EM, et al. A comprehensive insight into the role of exosomes in viral infection: dual faces bearing different functions. Pharmaceutics. 2021;13. doi:10.3390/pharmaceutics13091405
29. Shen Z, Huang W, Liu J, et al. Effects of mesenchymal stem cell-derived exosomes on autoimmune diseases. Front Immunol. 2021;12:749192. doi:10.3389/fimmu.2021.749192
30. Wang X, Zhou G, Zhou W, et al. Exosomes as a new delivery vehicle in inflammatory bowel disease. Pharmaceutics. 2021;13:1644. doi:10.3390/pharmaceutics13101644
31. Ocansey DKW, Zhang L, Wang Y, et al. Exosome-mediated effects and applications in inflammatory bowel disease. Biol Rev Camb Philos Soc. 2020;95:1287–1307. doi:10.1111/brv.12608
32. Yang C, Merlin D. Nanoparticle-mediated drug delivery systems for the treatment of IBD: current perspectives. Int J Nanomedicine. 2019;14:8875–8889. doi:10.2147/IJN.S210315
33. Liang Y, Duan L, Lu J, et al. Engineering exosomes for targeted drug delivery. Theranostics. 2021;11:3183–3195. doi:10.7150/thno.52570
34. Zu M, Xie D, Canup BSB, et al. “Green” nanotherapeutics from tea leaves for orally targeted prevention and alleviation of colon diseases. Biomaterials. 2021;279:121178. doi:10.1016/j.biomaterials.2021.121178
35. Tong L, Hao H, Zhang Z, et al. Milk-derived extracellular vesicles alleviate ulcerative colitis by regulating the gut immunity and reshaping the gut microbiota. Theranostics. 2021;11:8570–8586. doi:10.7150/thno.62046
36. Xu XH, Yuan TJ, Dad HA, et al. Plant exosomes as novel nanoplatforms for MicroRNA transfer stimulate neural differentiation of stem cells in vitro and in vivo. Nano Lett. 2021;21:8151–8159. doi:10.1021/acs.nanolett.1c02530
37. Kalluri R. The biology and function of exosomes in cancer. J Clin Invest. 2016;126:1208–1215. doi:10.1172/JCI81135
38. Tkach M, Thery C. Communication by extracellular vesicles: where we are and where we need to go. Cell. 2016;164:1226–1232. doi:10.1016/j.cell.2016.01.043
39. Xie W, Deng WW, Zan M, et al. Cancer cell membrane camouflaged nanoparticles to realize starvation therapy together with checkpoint blockades for enhancing cancer therapy. ACS Nano. 2019;13:2849–2857. doi:10.1021/acsnano.8b03788
40. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367. doi:10.1126/science.aau6977
41. Valter M, Verstockt S, Finalet Ferreiro JA, et al. Extracellular vesicles in inflammatory bowel disease: small particles, big players. J Crohns Colitis. 2021;15:499–510. doi:10.1093/ecco-jcc/jjaa179
42. Shetty AK, Upadhya R. Extracellular vesicles in health and disease. Aging Dis. 2021;12:1358–1362. doi:10.14336/AD.2021.0827
43. Zhang Y, Liu Y, Liu H, et al. Exosomes: biogenesis, biologic function and clinical potential. Cell Biosci. 2019;9:19. doi:10.1186/s13578-019-0282-2
44. Babst M. MVB vesicle formation: ESCRT-dependent, ESCRT-independent and everything in between. Curr Opin Cell Biol. 2011;23:452–457. doi:10.1016/j.ceb.2011.04.008
45. Ciardiello C, Cavallini L, Spinelli C, et al. Focus on extracellular vesicles: new frontiers of cell-to-cell communication in cancer. Int J Mol Sci. 2016;17:175. doi:10.3390/ijms17020175
46. Bebelman MP, Smit MJ, Pegtel DM, et al. Biogenesis and function of extracellular vesicles in cancer. Pharmacol Ther. 2018;188:1–11. doi:10.1016/j.pharmthera.2018.02.013
47. Blanc L, Vidal M. New insights into the function of Rab GTPases in the context of exosomal secretion. Small GTPases. 2018;9:95–106. doi:10.1080/21541248.2016.1264352
48. Jeppesen DK, Fenix AM, Franklin JL, et al. Reassessment of exosome composition. Cell. 2019;177:428–445 e18. doi:10.1016/j.cell.2019.02.029
49. Munich S, Sobo-Vujanovic A, Buchser WJ, et al. Dendritic cell exosomes directly kill tumor cells and activate natural killer cells via TNF superfamily ligands. Oncoimmunology. 2012;1:1074–1083. doi:10.4161/onci.20897
50. Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles. 2014;3:24641. doi:10.3402/jev.v3.24641
51. Tian T, Zhu YL, Hu FH, et al. Dynamics of exosome internalization and trafficking. J Cell Physiol. 2013;228:1487–1495. doi:10.1002/jcp.24304
52. Dhondt B, Geeurickx E, Tulkens J, et al. Unravelling the proteomic landscape of extracellular vesicles in prostate cancer by density-based fractionation of urine. J Extracell Vesicles. 2020;9:1736935. doi:10.1080/20013078.2020.1736935
53. Maacha S, Bhat AA, Jimenez L, et al. Extracellular vesicles-mediated intercellular communication: roles in the tumor microenvironment and anti-cancer drug resistance. Mol Cancer. 2019;18:55. doi:10.1186/s12943-019-0965-7
54. Urbanelli L, Buratta S, Tancini B, et al. The role of extracellular vesicles in viral infection and transmission. Vaccines. 2019;7. doi:10.3390/vaccines7030102
55. Ren J, He W, Zheng L, et al. From structures to functions: insights into exosomes as promising drug delivery vehicles. Biomater Sci. 2016;4:910–921. doi:10.1039/C5BM00583C
56. Liao W, Du Y, Zhang C, et al. Exosomes: the next generation of endogenous nanomaterials for advanced drug delivery and therapy. Acta Biomater. 2019;86:1–14. doi:10.1016/j.actbio.2018.12.045
57. Yong T, Zhang X, Bie N, et al. Tumor exosome-based nanoparticles are efficient drug carriers for chemotherapy. Nat Commun. 2019;10:3838. doi:10.1038/s41467-019-11718-4
58. Lopez de Las Hazas MC, Gil-Zamorano J, Cofan M, et al. One-year dietary supplementation with walnuts modifies exosomal miRNA in elderly subjects. Eur J Nutr. 2021;60:1999–2011. doi:10.1007/s00394-020-02390-2
59. Mustapic M, Tran J, Craft S, et al. Extracellular vesicle biomarkers track cognitive changes following intranasal insulin in Alzheimer’s disease. J Alzheimers Dis. 2019;69:489–498. doi:10.3233/JAD-180578
60. Athauda D, Gulyani S, Karnati HK, et al. Utility of neuronal-derived exosomes to examine molecular mechanisms that affect motor function in patients with parkinson disease: a secondary analysis of the Exenatide-PD trial. JAMA Neurol. 2019;76:420–429. doi:10.1001/jamaneurol.2018.4304
61. Perocheau D, Touramanidou L, Gurung S, et al. Clinical applications for exosomes: are we there yet? Br J Pharmacol. 2021;178:2375–2392. doi:10.1111/bph.15432
62. Ciferri MC, Quarto R, Tasso R. Extracellular vesicles as biomarkers and therapeutic tools: from pre-clinical to clinical applications. Biology. 2021;10. doi:10.3390/biology10050359
63. Song Y, Kim Y, Ha S, et al. The emerging role of exosomes as novel therapeutics: biology, technologies, clinical applications, and the next. Am J Reprod Immunol. 2021;85:e13329. doi:10.1111/aji.13329
64. Akers JC, Gonda D, Kim R, et al. Biogenesis of extracellular vesicles (EV): exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol. 2013;113:1–11. doi:10.1007/s11060-013-1084-8
65. Teixeira FG, Carvalho MM, Sousa N, et al. Mesenchymal stem cells secretome: a new paradigm for central nervous system regeneration? Cell Mol Life Sci. 2013;70:3871–3882. doi:10.1007/s00018-013-1290-8
66. Lai RC, Yeo RW, Lim SK. Mesenchymal stem cell exosomes. Semin Cell Dev Biol. 2015;40:82–88. doi:10.1016/j.semcdb.2015.03.001
67. Lee M, Jeong SY, Ha J, et al. Low immunogenicity of allogeneic human umbilical cord blood-derived mesenchymal stem cells in vitro and in vivo. Biochem Biophys Res Commun. 2014;446:983–989. doi:10.1016/j.bbrc.2014.03.051
68. Le Blanc K, Davies LC. Mesenchymal stromal cells and the innate immune response. Immunol Lett. 2015;168:140–146. doi:10.1016/j.imlet.2015.05.004
69. Forbes GM, Sturm MJ, Leong RW, et al. A Phase 2 study of allogeneic mesenchymal stromal cells for luminal Crohn’s disease refractory to biologic therapy. Clin Gastroenterol Hepatol. 2014;12:64–71. doi:10.1016/j.cgh.2013.06.021
70. Nammian P, Asadi-Yousefabad SL, Daneshi S, et al. Comparative analysis of mouse bone marrow and adipose tissue mesenchymal stem cells for critical limb ischemia cell therapy. Stem Cell Res Ther. 2021;12:58. doi:10.1186/s13287-020-02110-x
71. Ryu DJ, Jeon YS, Park JS, et al. Comparison of bone marrow aspirate concentrate and allogenic human umbilical cord blood derived mesenchymal stem cell implantation on chondral defect of knee: assessment of clinical and magnetic resonance imaging outcomes at 2-Year follow-up. Cell Transplant. 2020;29:963689720943581. doi:10.1177/0963689720943581
72. Wang X, Thomsen P. Mesenchymal stem cell-derived small extracellular vesicles and bone regeneration. Basic Clin Pharmacol Toxicol. 2021;128:18–36. doi:10.1111/bcpt.13478
73. Kim GB, Shon OJ, Seo MS, et al. Mesenchymal stem cell-derived exosomes and their therapeutic potential for osteoarthritis. Biology. 2021;10. doi:10.3390/biology10040285
74. Xia C, Dai Z, Jin Y, et al. Emerging antioxidant paradigm of mesenchymal stem cell-derived exosome therapy. Front Endocrinol (Lausanne). 2021;12:727272. doi:10.3389/fendo.2021.727272
75. Guo M, Yin Z, Chen F, et al. Mesenchymal stem cell-derived exosome: a promising alternative in the therapy of Alzheimer’s disease. Alzheimers Res Ther. 2020;12:109. doi:10.1186/s13195-020-00670-x
76. Aneesh A, Liu A, Moss HE, et al. Emerging concepts in the treatment of optic neuritis: mesenchymal stem cell-derived extracellular vesicles. Stem Cell Res Ther. 2021;12:594. doi:10.1186/s13287-021-02645-7
77. Kim GU, Sung SE, Kang KK, et al. Therapeutic potential of mesenchymal stem cells (MSCs) and MSC-derived extracellular vesicles for the treatment of spinal cord injury. Int J Mol Sci. 2021;22:13672. doi:10.3390/ijms222413672
78. Lange H, Gagliardi D. Plant exosomes and cofactors. Enzymes. 2012;31:31–52.
79. Mao Y, Han M, Chen C, et al. A biomimetic nanocomposite made of a ginger-derived exosome and an inorganic framework for high-performance delivery of oral antibodies. Nanoscale. 2021;13:20157–20169. doi:10.1039/D1NR06015E
80. Deng Z, Rong Y, Teng Y, et al. Broccoli-derived nanoparticle inhibits mouse colitis by activating dendritic cell AMP-Activated Protein Kinase. Mol Ther. 2017;25:1641–1654. doi:10.1016/j.ymthe.2017.01.025
81. Wang QL, Zhuang X, Sriwastva MK, et al. Blood exosomes regulate the tissue distribution of grapefruit-derived nanovector via CD36 and IGFR1 pathways. Theranostics. 2018;8:4912–4924. doi:10.7150/thno.27608
82. Bruno SP, Paolini A, D’Oria V, et al. Extracellular vesicles derived from citrus sinensis modulate inflammatory genes and tight junctions in a human model of intestinal epithelium. Front Nutr. 2021;8:778998. doi:10.3389/fnut.2021.778998
83. Lei C, Teng Y, He L, et al. Lemon exosome-like nanoparticles enhance stress survival of gut bacteria by RNase P-mediated specific tRNA decay. iScience. 2021;24:102511. doi:10.1016/j.isci.2021.102511
84. Diez-Sainz E, Lorente-Cebrian S, Aranaz P, et al. Potential mechanisms linking food-derived MicroRNAs, gut microbiota and intestinal barrier functions in the context of nutrition and human health. Front Nutr. 2021;8:586564. doi:10.3389/fnut.2021.586564
85. Sundaram K, Miller DP, Kumar A, et al. Plant-derived exosomal nanoparticles inhibit pathogenicity of porphyromonas gingivalis. iScience. 2019;21:308–327. doi:10.1016/j.isci.2019.10.032
86. Haug A, Hostmark AT, Harstad OM. Bovine milk in human nutrition–a review. Lipids Health Dis. 2007;6:25. doi:10.1186/1476-511X-6-25
87. Zhang B, Yang Y, Xiang L, et al. Adipose-derived exosomes: a novel adipokine in obesity-associated diabetes. J Cell Physiol. 2019;234:16692–16702. doi:10.1002/jcp.28354
88. Melnik BC, Stremmel W, Weiskirchen R, et al. Exosome-derived MicroRNAs of human milk and their effects on infant health and development. Biomolecules. 2021;11:851. doi:10.3390/biom11060851
89. Alsaweed M, Hartmann PE, Geddes DT, et al. MicroRNAs in breastmilk and the lactating breast: potential immunoprotectors and developmental regulators for the infant and the mother. Int J Environ Res Public Health. 2015;12:13981–14020. doi:10.3390/ijerph121113981
90. Zempleni J, Aguilar-Lozano A, Sadri M, et al. Biological activities of extracellular vesicles and their cargos from bovine and human milk in humans and implications for infants. J Nutr. 2017;147:3–10. doi:10.3945/jn.116.238949
91. Admyre C, Johansson SM, Qazi KR, et al. Exosomes with immune modulatory features are present in human breast milk. J Immunol. 2007;179:1969–1978. doi:10.4049/jimmunol.179.3.1969
92. Zhou Q, Li M, Wang X, et al. Immune-related microRNAs are abundant in breast milk exosomes. Int J Biol Sci. 2012;8:118–123. doi:10.7150/ijbs.8.118
93. de la Torre Gomez C, Goreham RV, Bech Serra JJ, et al. “Exosomics”-A review of biophysics, biology and biochemistry of exosomes with a focus on human breast milk. Front Genet. 2018;9:92. doi:10.3389/fgene.2018.00092
94. Zhong J, Xia B, Shan S, et al. High-quality milk exosomes as oral drug delivery system. Biomaterials. 2021;277:121126. doi:10.1016/j.biomaterials.2021.121126
95. Sedykh S, Kuleshova A, Nevinsky G. Milk exosomes: perspective agents for anticancer drug delivery. Int J Mol Sci. 2020;21:6646. doi:10.3390/ijms21186646
96. Neurath MF. Targeting immune cell circuits and trafficking in inflammatory bowel disease. Nat Immunol. 2019;20:970–979. doi:10.1038/s41590-019-0415-0
97. Tavasolian F, Hosseini AZ, Rashidi M, et al. The impact of immune cell-derived exosomes on immune response initiation and immune system function. Curr Pharm Des. 2021;27:197–205. doi:10.2174/1381612826666201207221819
98. La Salvia S, Musante L, Lannigan J, et al. T cell-derived extracellular vesicles are elevated in essential HTN. Am J Physiol Renal Physiol. 2020;319:F868–F875. doi:10.1152/ajprenal.00433.2020
99. Elashiry M, Elashiry MM, Elsayed R, et al. Dendritic cell derived exosomes loaded with immunoregulatory cargo reprogram local immune responses and inhibit degenerative bone disease in vivo. J Extracell Vesicles. 2020;9:1795362. doi:10.1080/20013078.2020.1795362
100. Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183:1161–1172. doi:10.1084/jem.183.3.1161
101. Baranyai T, Herczeg K, Onodi Z, et al. Isolation of exosomes from blood plasma: qualitative and quantitative comparison of ultracentrifugation and size exclusion chromatography methods. PLoS One. 2015;10:e0145686. doi:10.1371/journal.pone.0145686
102. Street JM, Koritzinsky EH, Glispie DM, et al. Urine exosome isolation and characterization. Methods Mol Biol. 2017;1641:413–423.
103. Cheng J, Nonaka T, Wong DTW. Salivary exosomes as nanocarriers for cancer biomarker delivery. Materials. 2019;12(4):654. doi:10.3390/ma12040654
104. Dixon CL, Sheller-Miller S, Saade GR, et al. Amniotic fluid exosome proteomic profile exhibits unique pathways of term and preterm labor. Endocrinology. 2018;159:2229–2240. doi:10.1210/en.2018-00073
105. Miyake H, Lee C, Chusilp S, et al. Human breast milk exosomes attenuate intestinal damage. Pediatr Surg Int. 2020;36:155–163. doi:10.1007/s00383-019-04599-7
106. Sun Y, Li L, Wang L, et al. Body fluids-derived exosomes: paving the novel road to lung cancer diagnosis and therapy. Anticancer Agents Med Chem. 2017;17:1734–1740. doi:10.2174/1871520617666170327112815
107. Cappello F, Logozzi M, Campanella C, et al. Exosome levels in human body fluids: a tumor marker by themselves? Eur J Pharm Sci. 2017;96:93–98. doi:10.1016/j.ejps.2016.09.010
108. Shao J, Jin Y, Shao C, et al. Serum exosomal pregnancy zone protein as a promising biomarker in inflammatory bowel disease. Cell Mol Biol Lett. 2021;26:36. doi:10.1186/s11658-021-00280-x
109. Zheng X, Chen F, Zhang Q, et al. Salivary exosomal PSMA7: a promising biomarker of inflammatory bowel disease. Protein Cell. 2017;8:686–695. doi:10.1007/s13238-017-0413-7
110. Larabi A, Barnich N, Nguyen HTT. Emerging role of exosomes in diagnosis and treatment of infectious and inflammatory bowel diseases. Cells. 2020;9:1111. doi:10.3390/cells9051111
111. Jiang L, Shen Y, Guo D, et al. EpCAM-dependent extracellular vesicles from intestinal epithelial cells maintain intestinal tract immune balance. Nat Commun. 2016;7:13045. doi:10.1038/ncomms13045
112. Chen X, Song CH, Feng BS, et al. Intestinal epithelial cell-derived integrin alphabeta6 plays an important role in the induction of regulatory T cells and inhibits an antigen-specific Th2 response. J Leukoc Biol. 2011;90:751–759. doi:10.1189/jlb.1210696
113. Leoni G, Neumann PA, Kamaly N, et al. Annexin A1-containing extracellular vesicles and polymeric nanoparticles promote epithelial wound repair. J Clin Invest. 2015;125:1215–1227. doi:10.1172/JCI76693
114. Lin XP, Almqvist N, Telemo E. Human small intestinal epithelial cells constitutively express the key elements for antigen processing and the production of exosomes. Blood Cells Mol Dis. 2005;35:122–128. doi:10.1016/j.bcmd.2005.05.011
115. Merkenschlager J, Eksmond U, Danelli L, et al. MHC class II cell-autonomously regulates self-renewal and differentiation of normal and malignant B cells. Blood. 2019;133:1108–1118. doi:10.1182/blood-2018-11-885467
116. Kojima M, Costantini TW, Eliceiri BP, et al. Gut epithelial cell-derived exosomes trigger posttrauma immune dysfunction. J Trauma Acute Care Surg. 2018;84:257–264. doi:10.1097/TA.0000000000001748
117. Bulek K, Zhao J, Liao Y, et al. Epithelial-derived gasdermin D mediates nonlytic IL-1beta release during experimental colitis. J Clin Invest. 2020;130:4218–4234. doi:10.1172/JCI138103
118. Slater TW, Finkielsztein A, Mascarenhas LA, et al. Neutrophil microparticles deliver active myeloperoxidase to injured mucosa to inhibit epithelial wound healing. J Immunol. 2017;198(7):2886–2897. doi:10.4049/jimmunol.1601810
119. Carriere J, Bretin A, Darfeuille-Michaud A, et al. Exosomes released from cells infected with Crohnʼs disease–associated adherent-invasive escherichia coli activate host innate immune responses and enhance bacterial intracellular replication. Inflamm Bowel Dis. 2016;22(3):516–528. doi:10.1097/MIB.0000000000000635
120. Corridoni D, Chapman T, Ambrose T, et al. Emerging mechanisms of innate immunity and their translational potential in inflammatory bowel disease. Front Med. 2018;5:32. doi:10.3389/fmed.2018.00032
121. Geremia A, Biancheri P, Allan P, et al. Innate and adaptive immunity in inflammatory bowel disease. Autoimmun Rev. 2014;13:3–10. doi:10.1016/j.autrev.2013.06.004
122. Han X, Ding S, Jiang H, et al. Roles of macrophages in the development and treatment of gut inflammation. Front Cell Dev Biol. 2021;9:625423. doi:10.3389/fcell.2021.625423
123. Ruder B, Becker C. At the forefront of the mucosal barrier: the role of macrophages in the intestine. Cells. 2020;9(10):2162. doi:10.3390/cells9102162
124. Wang J, Chen WD, Wang YD. The relationship between gut microbiota and inflammatory diseases: the role of macrophages. Front Microbiol. 2020;11:1065. doi:10.3389/fmicb.2020.01065
125. Lu J, Liu D, Tan Y, et al. M1 Macrophage exosomes MiR-21a-5p aggravates inflammatory bowel disease through decreasing E-cadherin and subsequent ILC2 activation. J Cell Mol Med. 2021;25:3041–3050. doi:10.1111/jcmm.16348
126. Yang R, Liao Y, Wang L, et al. Exosomes derived from M2b macrophages attenuate DSS-induced colitis. Front Immunol. 2019;10:2346. doi:10.3389/fimmu.2019.02346
127. Deng F, Yan J, Lu J, et al. M2 macrophage-derived exosomal miR-590-3p attenuates DSS-induced mucosal damage and promotes epithelial repair via the LATS1/YAP/ beta-catenin signalling axis. J Crohns Colitis. 2021;15:665–677. doi:10.1093/ecco-jcc/jjaa214
128. Arabpour M, Saghazadeh A, Rezaei N. Anti-inflammatory and M2 macrophage polarization-promoting effect of mesenchymal stem cell-derived exosomes. Int Immunopharmacol. 2021;97:107823. doi:10.1016/j.intimp.2021.107823
129. Thery C, Duban L, Segura E, et al. Indirect activation of naive CD4+ T cells by dendritic cell-derived exosomes. Nat Immunol. 2002;3:1156–1162. doi:10.1038/ni854
130. Shahir M, Mahmoud Hashemi S, Asadirad A, et al. Effect of mesenchymal stem cell-derived exosomes on the induction of mouse tolerogenic dendritic cells. J Cell Physiol. 2020;235:7043–7055. doi:10.1002/jcp.29601
131. Wei G, Jie Y, Haibo L, et al. Dendritic cells derived exosomes migration to spleen and induction of inflammation are regulated by CCR7. Sci Rep. 2017;7:42996. doi:10.1038/srep42996
132. Yang X, Meng S, Jiang H, et al. Exosomes derived from interleukin-10-treated dendritic cells can inhibit trinitrobenzene sulfonic acid-induced rat colitis. Scand J Gastroenterol. 2010;45:1168–1177. doi:10.3109/00365521.2010.490596
133. Butin-Israeli V, Houser MC, Feng M, et al. Deposition of microparticles by neutrophils onto inflamed epithelium: a new mechanism to disrupt epithelial intercellular adhesions and promote transepithelial migration. FASEB J. 2016;30:4007–4020. doi:10.1096/fj.201600734R
134. Liao F, Lu X, Dong W. Exosomes derived from T regulatory cells relieve inflammatory bowel disease by transferring miR-195a-3p. IUBMB Life. 2020;72:2591–2600.
135. Figliuolo da Paz VR, Jamwal DR, Kiela PR. Intestinal regulatory T cells. Adv Exp Med Biol. 2021;1278:141–190. doi:10.1007/978-981-15-6407-9_9
136. Wang Y, Tian J, Tang X, et al. Exosomes released by granulocytic myeloid-derived suppressor cells attenuate DSS-induced colitis in mice. Oncotarget. 2016;7:15356–15368. doi:10.18632/oncotarget.7324
137. Chang X, Wang SL, Zhao SB, et al. Extracellular vesicles with possible roles in gut intestinal tract homeostasis and IBD. Mediators Inflamm. 2020;2020:1945832. doi:10.1155/2020/1945832
138. Liu Y, Defourny KAY, Smid EJ, et al. Gram-positive bacterial extracellular vesicles and their impact on health and disease. Front Microbiol. 2018;9:1502. doi:10.3389/fmicb.2018.01502
139. Simoes CD, Maganinho M, Sousa AS. FODMAPs, inflammatory bowel disease and gut microbiota: updated overview on the current evidence. Eur J Nutr. 2022;61(3):1187–1198. doi:10.1007/s00394-021-02755-1
140. Tong L, Zhang X, Hao H, et al. Lactobacillus rhamnosus GG derived extracellular vesicles modulate gut microbiota and attenuate inflammatory in DSS-induced colitis mice. Nutrients. 2021;13:3319. doi:10.3390/nu13103319
141. Kang CS, Ban M, Choi EJ, et al. Extracellular vesicles derived from gut microbiota, especially Akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS One. 2013;8:e76520. doi:10.1371/journal.pone.0076520
142. Qi Y, Wu HM, Yang Z, et al. New insights into the role of oral microbiota dysbiosis in the pathogenesis of inflammatory bowel disease. Dig Dis Sci. 2022;67:42–55. doi:10.1007/s10620-021-06837-2
143. Liu L, Liang L, Yang C, et al. Extracellular vesicles of Fusobacterium nucleatum compromise intestinal barrier through targeting RIPK1-mediated cell death pathway. Gut Microbes. 2021;13:1–20. doi:10.1080/19490976.2021.1902718
144. Canas MA, Fabrega MJ, Gimenez R, et al. Outer membrane vesicles from probiotic and commensal escherichia coli activate NOD1-mediated immune responses in intestinal epithelial cells. Front Microbiol. 2018;9:498. doi:10.3389/fmicb.2018.00498
145. Sun Y, Liu G, Zhang K, et al. Mesenchymal stem cells-derived exosomes for drug delivery. Stem Cell Res Ther. 2021;12:561. doi:10.1186/s13287-021-02629-7
146. Herrmann IK, Wood MJA, Fuhrmann G. Extracellular vesicles as a next-generation drug delivery platform. Nat Nanotechnol. 2021;16:748–759. doi:10.1038/s41565-021-00931-2
147. Xu M, Yang Q, Sun X, et al. Recent advancements in the loading and modification of therapeutic exosomes. Front Bioeng Biotechnol. 2020;8:586130. doi:10.3389/fbioe.2020.586130
148. Gyorgy B, Hung ME, Breakefield XO, et al. Therapeutic applications of extracellular vesicles: clinical promise and open questions. Annu Rev Pharmacol Toxicol. 2015;55:439–464. doi:10.1146/annurev-pharmtox-010814-124630
149. Familtseva A, Jeremic N, Tyagi SC. Exosomes: cell-created drug delivery systems. Mol Cell Biochem. 2019;459:1–6. doi:10.1007/s11010-019-03545-4
150. Zhang M, Xiao B, Wang H, et al. Edible ginger-derived nano-lipids loaded with doxorubicin as a novel drug-delivery approach for colon cancer therapy. Mol Ther. 2016;24:1783–1796. doi:10.1038/mt.2016.159
151. Mao F, Wu Y, Tang X, et al. Exosomes derived from human umbilical cord mesenchymal stem cells relieve inflammatory bowel disease in mice. Biomed Res Int. 2017;2017:5356760. doi:10.1155/2017/5356760
152. Cao L, Xu H, Wang G, et al. Extracellular vesicles derived from bone marrow mesenchymal stem cells attenuate dextran sodium sulfate-induced ulcerative colitis by promoting M2 macrophage polarization. Int Immunopharmacol. 2019;72:264–274. doi:10.1016/j.intimp.2019.04.020
153. Yu H, Yang X, Xiao X, et al. Human adipose mesenchymal stem cell-derived exosomes protect mice from DSS-induced inflammatory bowel disease by promoting intestinal-stem-cell and epithelial regeneration. Aging Dis. 2021;12:1423–1437. doi:10.14336/AD.2021.0601
154. Chen Q, Duan X, Xu M, et al. BMSC-EVs regulate Th17 cell differentiation in UC via H3K27me3. Mol Immunol. 2020;118:191–200. doi:10.1016/j.molimm.2019.12.019
155. Yu T, Chu S, Liu X, et al. Extracellular vesicles derived from EphB2-overexpressing bone marrow mesenchymal stem cells ameliorate DSS-induced colitis by modulating immune balance. Stem Cell Res Ther. 2021;12:181. doi:10.1186/s13287-021-02232-w
156. Yang S, Liang X, Song J, et al. A novel therapeutic approach for inflammatory bowel disease by exosomes derived from human umbilical cord mesenchymal stem cells to repair intestinal barrier via TSG-6. Stem Cell Res Ther. 2021;12(1):315. doi:10.1186/s13287-021-02404-8
157. Cai X, Zhang ZY, Yuan JT, et al. hucMSC-derived exosomes attenuate colitis by regulating macrophage pyroptosis via the miR-378a-5p/NLRP3 axis. Stem Cell Res Ther. 2021;12:416. doi:10.1186/s13287-021-02492-6
158. Panes J, Garcia-Olmo D, Van Assche G, et al. Expanded allogeneic adipose-derived mesenchymal stem cells (Cx601) for complex perianal fistulas in Crohn’s disease: a Phase 3 randomised, double-blind controlled trial. Lancet. 2016;388:1281–1290. doi:10.1016/S0140-6736(16)31203-X
159. Panes J, Garcia-Olmo D, Van Assche G, et al. Long-term efficacy and safety of stem cell therapy (Cx601) for complex perianal fistulas in patients with Crohn’s disease. Gastroenterology. 2018;154:1334–1342 e4. doi:10.1053/j.gastro.2017.12.020
160. Barnhoorn MC, Wasser M, Roelofs H, et al. Long-term evaluation of allogeneic bone marrow-derived mesenchymal stromal cell therapy for Crohn’s disease perianal fistulas. J Crohns Colitis. 2020;14:64–70. doi:10.1093/ecco-jcc/jjz116
161. Ju S, Mu J, Dokland T, et al. Grape exosome-like nanoparticles induce intestinal stem cells and protect mice from DSS-induced colitis. Mol Ther. 2013;21:1345–1357. doi:10.1038/mt.2013.64
162. Zhang M, Viennois E, Prasad M, et al. Edible ginger-derived nanoparticles: a novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials. 2016;101:321–340. doi:10.1016/j.biomaterials.2016.06.018
163. Sriwastva MK, Deng ZB, Wang B, et al. Exosome-like nanoparticles from Mulberry bark prevent DSS-induced colitis via the AhR/COPS8 pathway. EMBO Rep. 2022;23:e53365. doi:10.15252/embr.202153365
164. Liu C, Yan X, Zhang Y, et al. Oral administration of turmeric-derived exosome-like nanovesicles with anti-inflammatory and pro-resolving bioactions for murine colitis therapy. J Nanobiotechnology. 2022;20:206. doi:10.1186/s12951-022-01421-w
165. Colitti M, Sgorlon S, Stefanon B. Exosome cargo in milk as a potential marker of cow health. J Dairy Res. 2020;87:79–83. doi:10.1017/S0022029920000485
166. Zempleni J, Sukreet S, Zhou F, et al. Milk-derived exosomes and metabolic regulation. Annu Rev Anim Biosci. 2019;7:245–262. doi:10.1146/annurev-animal-020518-115300
167. Feng X, Chen X, Zheng X, et al. Latest trend of milk derived exosomes: cargos, functions, and applications. Front Nutr. 2021;8:747294. doi:10.3389/fnut.2021.747294
168. Shelby RD, Cromeens B, Rager TM, et al. Influence of growth factors on the development of necrotizing enterocolitis. Clin Perinatol. 2019;46:51–64. doi:10.1016/j.clp.2018.10.005
169. Jeschke MG, Bolder U, Chung DH, et al. Gut mucosal homeostasis and cellular mediators after severe thermal trauma and the effect of insulin-like growth factor-I in combination with insulin-like growth factor binding protein-3. Endocrinology. 2007;148:354–362.
170. Benmoussa A, Diallo I, Salem M, et al. Concentrates of two subsets of extracellular vesicles from cow’s milk modulate symptoms and inflammation in experimental colitis. Sci Rep. 2019;9:14661. doi:10.1038/s41598-019-51092-1
171. Reif S, Elbaum-Shiff Y, Koroukhov N, et al. Cow and human milk-derived exosomes ameliorate colitis in DSS murine model. Nutrients. 2020;12:2589. doi:10.3390/nu12092589
172. Li DF, Chang X, Zhao JL, et al. Colonic epithelial PHLPP2 deficiency promotes colonic epithelial pyroptosis by activating the NF-kappaB signaling pathway. Oxid Med Cell Longev. 2021;2021:5570731. doi:10.1155/2021/5570731
173. Munagala R, Aqil F, Jeyabalan J, et al. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016;371:48–61. doi:10.1016/j.canlet.2015.10.020
174. Vashisht M, Rani P, Onteru SK, et al. Curcumin encapsulated in milk exosomes resists human digestion and possesses enhanced intestinal permeability in vitro. Appl Biochem Biotechnol. 2017;183:993–1007. doi:10.1007/s12010-017-2478-4
175. Kandimalla R, Aqil F, Tyagi N, et al. Milk exosomes: a biogenic nanocarrier for small molecules and macromolecules to combat cancer. Am J Reprod Immunol. 2021;85:e13349. doi:10.1111/aji.13349
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.