Back to Journals » Drug Design, Development and Therapy » Volume 20
Extracellular Vesicles in Osteosarcoma: Mechanisms, Diagnostics and Therapeutic Applications
Authors Yin C, Wu Y, Qu X, Wu W, Zhang B, Chen J ![]()
Received 3 September 2025
Accepted for publication 29 November 2025
Published 6 January 2026 Volume 2026:20 565059
DOI https://doi.org/10.2147/DDDT.S565059
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Georgios Panos
Chaoyi Yin,* Yiqi Wu,* Xiliang Qu, Wenren Wu, Binshan Zhang, Jinshui Chen
Dongguan Hospital of Guangzhou University of Chinese Medicine, Dongguan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Binshan Zhang, Dongguan Hospital of Guangzhou University of Chinese Medicine, Dongguan, People’s Republic of China, Email [email protected] Jinshui Chen, Dongguan Hospital of Guangzhou University of Chinese Medicine, Dongguan, People’s Republic of China, Email [email protected]
Abstract: Osteosarcoma is a primary bone malignancy of adolescents and young adults with marked heterogeneity and a high metastatic propensity. Five-year survival exceeds 70% in localized disease but falls to about 20% with pulmonary metastasis or chemoresistance, and overall outcomes have plateaued for decades. Extracellular vesicles (EVs) have emerged as critical mediators of osteosarcoma progression and metastasis. EVs remodel the tumor microenvironment (TME) by promoting immune evasion, extracellular matrix reprogramming, and angiogenesis, while also facilitating invasion, epithelial–mesenchymal transition (EMT)–like plasticity, and formation of lung pre-metastatic niches through organotropic integrins and glycoproteins. Their cargo, including proteins, lipids, and nucleic acids, drives intercellular communication that sustains proliferation, migration, and therapy resistance under metabolic or hypoxic stress. Clinically, the stability of EVs in body fluids and their tumor-specific molecular signatures highlight their promise as liquid-biopsy biomarkers for early diagnosis, prognosis, and treatment monitoring. Therapeutically, EVs are being engineered as delivery vehicles for drugs or RNA therapeutics, and interventions targeting their biogenesis, cargo sorting, or uptake are under exploration. Future research should integrate single-EV multi-omics, longitudinal cohort validation, and causal perturbation models to delineate functional mechanisms. Rational strategies that modulate EV dynamics and incorporate standardized analytic pipelines may transform EVs into actionable biomarkers and therapeutic targets, offering new avenues to overcome resistance and improve clinical outcomes in osteosarcoma.
Keywords: EVs, osteosarcoma, ncRNAs, TME, mechanisms, therapeutic strategies
Introduction
Osteosarcoma is a primary malignant bone tumor that predominantly occurs in adolescents and young adults, characterized by high heterogeneity and a propensity for metastasis.1 Through postoperative combination chemotherapy or neoadjuvant therapy, the 5-year survival rate for patients with localized osteosarcoma can exceed 70%. However, once pulmonary metastasis or drug resistance occurs, the 5-year survival rate significantly drops to around 20%.2,3 The high metastatic potential and drug resistance of osteosarcoma are closely related to its TME. The TME includes various immune cells, osteoblasts, osteoblast-like stromal cells, vascular endothelial cells, etc. These components interact with tumor cells through secreted factors and EVs, collectively determining the tumor’s biological behavior and treatment response.4 The TME in osteosarcoma is predominantly immunosuppressive and rich in abundant cancer-associated fibroblasts (CAFs). These CAFs form dense barriers, preventing drug penetration and leading to low response to various treatments.5
EVs are phospholipid bilayer vesicles secreted by cells, with a diameter ranging from approximately 30–1000 nm, and are present in various body fluids, including blood. They carry cargo such as ncRNAs and lipids, transmitting information between donor and recipient cells.6 Non coding RNAs (ncRNAs) represent a broad class of transcripts that do not encode proteins yet exert pervasive regulatory functions at the transcriptional and post transcriptional levels. Among them, microRNAs (miRNAs), long non coding RNAs (lncRNAs) and circular RNAs (circRNAs) are particularly relevant in cancer, where they modulate oncogenic and tumour suppressive pathways, epithelial mesenchymal plasticity, angiogenesis and immune responses.7,8 Increasing evidence indicates that ncRNAs are not only dysregulated within tumour cells but are also selectively packaged into EVs.
EV associated ncRNAs exhibit distinct patterns of enrichment compared with their cellular counterparts, reflecting active loading mechanisms rather than passive leakage.9 Sorting into EVs involves components of the endosomal sorting complex required for transport, ceramide dependent pathways and several RNA binding proteins that recognise specific sequence or structural motifs. Once enclosed within the lipid bilayer, ncRNAs are protected from extracellular RNases and can be detected in a variety of body fluids, providing a stable source of information on tumor biology.10 Functionally, EV mediated transfer of ncRNAs reprogrammes recipient cells within the tumor microenvironment, including stromal, endothelial and immune populations, thereby influencing proliferation, invasion, angiogenesis, immune escape and therapy resistance. In osteosarcoma, these properties position EV carried ncRNAs as both active drivers of disease progression and accessible liquid biopsy biomarkers, which will be the focus of the following sections.
In osteosarcoma, both malignant cells and surrounding stromal and immune cells release EVs that are taken up locally and at distant sites, thereby reshaping cell phenotype and gene expression.11,12 EVs promote proliferation, invasion, angiogenesis, and extracellular matrix remodeling, foster immunosuppression, and condition premetastatic niches, especially in the lung where EV integrin and glycoprotein patterns can enhance vascular permeability and recruit pro-metastatic stromal elements in osteosarcoma.13 Clinically, EVs are attractive as liquid-biopsy biomarkers because of their stability in circulation and tumor-specific molecular signatures, enabling applications in diagnosis, risk stratification, minimal residual disease monitoring, and response assessment.14 At the same time, progress depends on rigorous pre-analytical control and standardized isolation, normalization, and analytical pipelines to ensure reproducibility across cohorts. A deeper mechanistic understanding of EV biogenesis, cargo sorting, organotropism, and uptake in osteosarcoma will inform the development of ncRNA-based diagnostic panels and therapeutic strategies that either exploit EVs as delivery vehicles or disrupt pathological EV communication networks.15,16
This review synthesizes the roles of EVs in the osteosarcoma tumor microenvironment, detailing how they shape immunosuppression, promote angiogenesis, and remodel the extracellular matrix. It further examines EV-driven programs that enhance tumor-cell proliferation, EMT, and premetastatic niche formation. We then summarize advances in EV-derived noncoding RNAs as diagnostic and prognostic biomarkers for osteosarcoma. Finally, we discuss therapeutic strategies targeting ncRNAs and recent progress in exploiting EVs as drug carriers, and we outline future directions for translating EV-based approaches into clinical benefit.
Biogenesis Mechanisms and Classification of EVs
EVs Detection Methods
Research on EVs depends on reproducible preprocessing and stringent quality control. Per MISEV2023, methods should report particle size, concentration, surface charge, and purity, include procedural and transport blanks, and use positive reference materials for inter-batch calibration. For low-yield specimens, assess co-isolation of lipoproteins, protein aggregates, and organelle debris, and confirm findings with at least two orthogonal techniques to reduce single-assay bias.17
Morphological and biophysical characterization provides the first layer of heterogeneity. Transmission electron microscopy verifies bilayer membranes and intravesicular structures, with immunogold for nanoscale antigen localization. Scanning electron microscopy offers broader topographic context.18 Cryo-electron microscopy preserves hydrated, near-native morphology. Atomic force microscopy supplies nanoscale topography and mechanics in liquid. Population metrics come from dynamic light scattering, nanoparticle tracking analysis, and tunable resistive pulse sensing; each requires standardized settings and calibration, with TRPS additionally monitoring ionic current stability.19,20 Small-particle or nano-flow cytometry resolves single EV events near 100 nm with fluorescence, and should report lower detection limits, gating strategy, sheath-fluid background, and dye controls.21
Molecular profiling links physical subtypes to function. Western blotting verifies transmembrane, cytosolic, and non-EV co-isolates to gauge integrity and purity. ELISA supports high-throughput quantification and can be coupled to digital arrays. Mass-spectrometry proteomics and lipidomics provide unbiased molecular signatures across fractions.22 At the nucleic-acid level, qPCR with strand-displacement, rolling-circle, or CRISPR-Cas amplification detects low-abundance EV RNAs. Total internal reflection fluorescence enables real-time single-vesicle readouts, and super-resolution imaging resolves nanoscale organization of surface markers.23
For rapid subpopulation detection and liquid biopsy, nano-flow cytometry enables single-vesicle phenotyping, Simoa quantifies targeted subpopulations in minimal plasma, horseradish-peroxidase colorimetric or fluorescent assays give minute-scale readouts, electrochemical stripping voltammetry with quantum-dot amplification increases sensitivity in microliter volumes, surface plasmon resonance provides label-free kinetics, and surface-enhanced Raman scattering enhances specificity in complex matrices, with microfluidic integration improving throughput.24 A practical workflow quantifies size and concentration with NTA or TRPS, confirms morphology by TEM or cryo-EM, verifies markers by Western blotting or ELISA, and maps composition with mass spectrometry or nucleic-acid assays. Transparent reporting of controls, calibration, and inter-batch normalization is essential for comparability.
Isolation and Characterization of EVs
For osteosarcoma cohorts, plasma is generally preferred over serum to limit platelet activation–derived vesicles; EDTA anticoagulation, rapid double-spin platelet removal, temperature control, and hemolysis scoring are critical preanalytical steps that influence EV yield and purity.25 Current clinical-leaning isolation workflows employ size-exclusion chromatography to separate vesicles from soluble proteins and lipoproteins, often coupled with tangential-flow filtration for scalable concentration and buffer exchange; differential ultracentrifugation remains widely used in discovery settings but can co-pellet contaminants if not combined with density or chromatographic steps. Immunoaffinity enrichment directed at tetraspanins or tumor-associated antigens can increase analytical sensitivity for low-abundance osteosarcoma EVs at the cost of narrower coverage.26 Across methods, quality control should pair particle counting and sizing with biochemical purity metrics and functional recovery of marker cargo to mitigate platform-dependent discrepancies.
Quantification and analytical readouts require equal attention. Nanoparticle tracking analysis, tunable resistive pulse sensing, and nano-flow cytometry yield non-interchangeable counts unless calibrated with traceable reference materials and harmonized thresholds; reporting according to MIFlowCyt-EV and related single-particle guidelines facilitates inter-study comparison.27 For osteosarcoma applications, candidate clinical endpoints include diagnostic discrimination, risk stratification, and treatment monitoring using EV-associated proteins and DNAs in addition to ncRNAs. Published studies have linked circulating EV markers such as PD-L1 and N-cadherin with pulmonary metastasis risk, while EV-associated repetitive-element DNA has shown high diagnostic accuracy in receiver-operating-characteristic analyses. These data motivate fit-for-purpose assay development but also underscore the need to control for confounders such as lipoprotein co-isolation, platelet-derived vesicle carryover, and batch variability, and to validate stability during storage and freeze–thaw.
Classification of EVs
Based on their biosynthetic mechanisms and size, EVs can be classified into three major categories.28 Among these, exosomes are small EVs derived from the endosomal pathway, with a diameter of approximately 30–100 nm.29 During late endocytosis, the endosomal membrane invaginates to form intraluminal vesicles (ILVs).30 Exosomes are released when multivesicular bodies (MVBs) fuse with the plasma membrane, discharging their ILVs extracellularly. Microvesicles are vesicles formed by the direct outward budding and pinching off of the plasma membrane, typically ranging from 100–1000 nm in diameter.31 The formation of microvesicles is associated with cytoskeletal rearrangement and altered phospholipid asymmetry; typical markers include phosphatidylserine externalization (eg, Annexin V binding). Apoptotic bodies are larger vesicles produced during cell apoptosis as the cell disintegrates, with diameters ranging up to 500 nm–5 μm.32 Apoptotic bodies often contain nuclear fragments and organelle components and are part of the apoptotic cell clearance mechanism. As research deepens, EV classification continues to expand, with recent studies reporting various types such as autophagic EVs, stress-induced EVs, and matrix vesicles.33
Biogenesis Mechanisms of EVs
EV biogenesis involves complex cellular mechanisms. The formation of ILVs within MVBs can be mediated by the ESCRT pathway or occur via ESCRT-independent pathways mediated by lipids and tetraspanins.34 MVBs can fuse with lysosomes for degradation or, under the action of RAB GTPases and SNARE proteins, fuse with the plasma membrane to release their internal vesicles as exosomes.35 In contrast, microvesicle budding is regulated by cytoskeletal proteins and membrane phospholipases, while apoptotic bodies accompany the activation of apoptotic signals. Within the donor cell, the endocytic pathway forms early and late endosomes.36 The membrane of late endosomes buds inward to form ILVs within MVBs. MVBs can selectively fuse with lysosomes for degradation or fuse with the plasma membrane to release the contained ILVs as exosomes.37 The unique aspect of EVs as intercellular communication carriers lies in their membrane-enclosed structure, which protects internal nucleic acids from degradation by nucleases in body fluids. Through fusion with the recipient cell membrane or endocytosis by the recipient cell, EVs achieve efficient cargo delivery. Consequently, ncRNAs inside EVs can be stably delivered to target cells and exert regulatory functions. Various cells within osteosarcoma and its TME can secrete EVs. These EVs are taken up by cells in local and distant tissues, participating in remodeling the TME and regulating tumor metastasis pathways. The classification of EVs and their generation mechanisms are extremely complex, and further research is still needed in the future (Figure 1).
 |
Figure 1 Biogenesis, classification, release, and uptake routes of EVs. EVs comprise exosomes, microvesicles, and apoptotic bodies that differ by cellular origin, size, and molecular composition. Exosomes (≈30–150 nm) arise within the endosomal system when intraluminal vesicles form in multivesicular bodies (MVBs). Their biogenesis is governed by ESCRT-0/-I/-II/-III complexes with accessory factors such as ALIX and TSG101, and can also proceed through ESCRT-independent routes involving ceramide generation and tetraspanin-enriched microdomains. MVB–plasma membrane fusion and vesicle release are regulated by Rab GTPases and SNARE machinery, yielding vesicles enriched for tetraspanins and syntenin. Microvesicles (≈100–1000 nm) bud outward from the plasma membrane following calcium influx, phospholipid scrambling with phosphatidylserine exposure, and actomyosin contraction driven by ARF6 and the RhoA–ROCK axis. Their membranes often retain parent-cell receptors and integrins that influence tropism. Apoptotic bodies (>1 µm) originate during programmed cell death through caspase-dependent blebbing and membrane fragmentation; they can encapsulate organelles, chromatin, and histones. Cargo selection checkpoints are indicated at endosomal and plasma-membrane microdomains, where ESCRT components, tetraspanins, and RNA-binding or lipid-sorting factors contribute to protein, lipid, and RNA loading. Principal uptake routes depicted include clathrin- or caveolin-mediated endocytosis, macropinocytosis, phagocytosis, and direct membrane fusion at recipient cells. This Figure created with BioRender. |
EVs Induced Immune Escape in Osteosarcoma
Within osteosarcoma, EVs facilitate immune evasion through convergent effects on lymphoid, myeloid, and stromal compartments. They dampen CD8⁺ T-cell and NK-cell cytotoxicity by transferring checkpoint ligands and regulatory cargos that reduce interferon-γ output and promote exhaustion.38 They reprogram myeloid cells toward M2-like macrophages and expand MDSCs, elevating IL-10, TGF-β, and arginase to suppress antigen-specific T-cell proliferation. They impair dendritic-cell maturation and limit effective HLA class I presentation, which constrains cross-priming. In parallel, they remodel matrix and metabolism by increasing ECM deposition and stiffness, altering endothelial adhesion, and enriching adenosine and lactate, thereby restricting effector trafficking. EVs also condition lung pre-metastatic niches by educating resident myeloid and stromal cells toward tolerogenic states. Framed by these effects, EVs operate as system-level coordinators of immune escape and reveal intervention points at cargo loading, biogenesis, release, and uptake (Figure 2).
 |
Figure 2 EVs mediate immune escape in osteosarcoma. EVs derived from osteosarcoma and its microenvironment mediate immune escape by suppressing cytotoxic lymphocytes, expanding regulatory and suppressive cell populations, and impairing antigen presentation. Through the transfer of noncoding RNAs and checkpoint ligands, EVs polarize macrophages toward the M2 phenotype, promote MDSC and Treg expansion, inhibit DC maturation, and suppress NK and CD8⁺ T-cell activity, collectively establishing a tolerogenic tumor microenvironment that supports metastasis and immune evasion. This figure was created with BioRender.com. |
EV-Induced M2-Like Macrophage Polarization and Immunoregulatory Cascades
Exosomes secreted by osteosarcoma can influence macrophage polarization, skewing them towards the pro-tumor M2 phenotype. Research by Chen et al found that exosomes secreted by osteosarcoma cells, carrying Tim-3, drive macrophage polarization towards the M2 type while releasing cytokines such as IL-10 and VEGF, thereby remodeling the immune microenvironment.39 This cascade further induces EMT in osteosarcoma cells, significantly enhancing their migration, invasion, and lung metastasis potential. Studies by Wang et al discovered that exosomes secreted by osteosarcoma cells are enriched with lncRNA ELFN1-AS1. After macrophages phagocytose these exosomes, lncRNA ELFN1-AS1 enters the macrophages and acts as a sponge for miR-138-5p and miR-1291, upregulating the level of the transcription factor CREB1, thereby inducing macrophage polarization to the M2 type.40 Polarized M2 tumor-associated macrophages (TAMs) suppress T cell-mediated anti-tumor immunity and promote tumor growth and metastasis. Small exosomes released by osteosarcoma cells enriched with miR-487a can be taken up by macrophages, directly inhibiting Notch2 and activating GATA3 signaling, thereby reprogramming macrophages to an immunosuppressive M2 phenotype.41 M2 macrophages subsequently drive osteosarcoma cell proliferation, migration, invasion, and EMT through the secretion of various pro-tumor factors, ultimately forming a malignant positive feedback loop that promotes lung metastasis.
Conversely, ncRNAs carried by EVs secreted by M2 macrophages also act on tumor cells, further strengthening the immunosuppressive cycle. Zhang et al found that the long non-coding RNA LIFR-AS1 in exosomes derived from M2 macrophages acts as a competing endogenous RNA “sponge” to adsorb miR-29a, relieving its inhibition of the transcription factor NFIA, thereby upregulating NFIA and driving osteosarcoma cell proliferation, migration, invasion, and inhibiting apoptosis.42 Knocking down exosomal LIFR-AS1 attenuated the tumor-promoting effect of macrophages, while inhibiting miR-29a or compensatory knockdown of NFIA could respectively reverse or restore this inhibitory effect. Liu et al reported that exosomes released by M2-type macrophages carry highly expressed miR-221-3p.43 When osteosarcoma cells take up these exosomes, miR-221-3p can target and inhibit the expression of SOCS3 within the tumor cells, relieving the inhibition of the JAK2/STAT3 pathway, thereby persistently activating STAT3 signaling. This series of pathway changes promotes tumor cell proliferation, migration, and invasion.
EV-Driven Expansion and Activation of Myeloid-Derived Suppressor Cells
MDSCs are a class of immature myeloid cells derived from the bone marrow that accumulate extensively in the TME. They are mainly phenotypically divided into PMN-MDSCs and monocytic MDSCs. Within the TME, chronic inflammatory signals can drive biased differentiation of bone marrow hematopoiesis towards MDSCs and block their further differentiation into mature dendritic cells or macrophages, forming a persistent immunosuppressive microenvironment. Research by Guo et al found that exosomes secreted by tumor cells are enriched with miR-29a and miR-92a.44 These two miRNAs are delivered to MDSCs, downregulating Hbp1 and Prkar1a respectively, thereby driving rapid expansion of MDSCs and conferring immunosuppressive functions. Enhanced MDSCs further shape the tumor “cold” microenvironment, helping tumor cells evade immune surveillance. miR-21 carried by exosomes secreted by CAFs drives the differentiation of monocytes into immunosuppressive M-MDSCs by activating the STAT3 signaling axis in the TME.45 M-MDSCs further significantly enhance cancer cell resistance to cisplatin through immunosuppressive and inflammatory cytokine network amplification effects. Research by Deng et al found that exosomes released by osteosarcoma cells carry the protein S100A11, which can bind to USP9X and activate the JAK2-STAT3 pathway in the pre-metastatic niche of the lung, causing lung interstitial macrophages to secrete large amounts of the chemokine CXCL2. This recruits granulocytic MDSCs (gMDSCs), creating a strongly immunosuppressive pre-metastatic niche.46
EV-Facilitated Induction and Stabilization of Regulatory T Cells
Tregs in the TME are typically recruited in large numbers or induced locally by chemokines secreted by tumor cells and associated stroma. They highly express checkpoint molecules such as CTLA-4 and PD-1, secrete IL-10 and IL-35, release CD39/CD73 to generate adenosine, competitively deplete IL-2, and synergistically suppress the functions of effector CD8⁺ T cells and antigen-presenting cells. They can also induce M2-type macrophage and MDSC expansion, thereby forming an immunosuppressive “protective shield” that promotes tumor immune evasion and drug resistance. After acquiring oxaliplatin resistance, tumor cells secrete large amounts of miR-208b via exosomes and deliver it to CD4⁺ T cells, inhibiting its target gene PDCD4, thereby inducing Treg expansion and exacerbating immunosuppression.47 Increased Tregs weaken anti-tumor immunity and form a drug-resistant microenvironment, reducing tumor response to FOLFOX therapy. Plasma exosomal miR-208b levels are closely associated with drug resistance and can serve as a non-invasive biomarker for predicting oxaliplatin sensitivity. The lung is prone to forming a pre-metastatic niche, and immunosuppressive Tregs are key shapers within it. Studies found that exosomes released by tumor cells can reprogram lung fibroblasts, causing them to secrete large amounts of the chemokine CCL1, which then drives CD4⁺ T cell differentiation into Tregs via CCR8 signaling activation, constructing a tolerant microenvironment and “paving the way” for tumor colonization.48 Tumor metabolic reprogramming leads to lactate enrichment in the microenvironment, and tumor-infiltrating Tregs rely on highly expressed monocarboxylate transporter MCT1 to uptake lactate to maintain immunosuppression. Exosomes released by bone marrow mesenchymal stem cells carry miR-124, which can directly target and downregulate MCT1, limiting Treg lactate uptake and impairing their suppressive function, thereby slowing tumor growth and significantly enhancing PD-1 blockade efficacy.49
EV-Impaired Dendritic-Cell Maturation and Antigen Presentation
As a key component of anti-tumor immune cells, dendritic cells (DCs) primarily function in antigen presentation. Studies found that in the TME, the maturation and activation of DCs are significantly inhibited, rendering them unable to exert anti-tumor functions. Research by Gao et al found that osteosarcoma cell-derived exosomes carry miR-133a, whose elevated expression can directly target and downregulate the transcription factor RBP-J, inhibiting DC maturation and activation, thereby weakening DC-mediated anti-tumor immunity.50 A miR-133a inhibitor could restore RBP-J levels and enhance DC transition to a mature phenotype, improving immune clearance capability against osteosarcoma. Exosomes secreted by tumor cells are enriched with the long non-coding RNA MALAT1, which can be taken up by DCs and activate the AKT/mTOR pathway, enhancing DC autophagy while inhibiting their phagocytosis and co-stimulatory molecule expression, thereby impairing T cell proliferation and anti-tumor immunity.51 Inhibiting exosomal-MALAT1 blocks AKT/mTOR signaling, reduces autophagy levels, restores DC function, and enhances T cell responses, significantly suppressing tumor growth. Additionally, studies found that exosomes released by tumor cells are enriched with fatty acids, which can force tumor-infiltrating DCs to shift their metabolism toward oxidative phosphorylation and accumulate large amounts of lipids. Exosomes further induce mature DCs to undergo lipid peroxidation via the NRF2/GPX4 signaling pathway and succumb to ferroptosis, impairing antigen presentation and creating an immunosuppressive microenvironment that promotes tumor growth.52 Downregulating Rab27a to inhibit exosome secretion significantly reduces DC lipid load and peroxidation levels, restoring their immune activity and suppressing tumor progression.
EV-Mediated NK-Cell Dysfunction and Checkpoint Signaling
NK cells are a subset of lymphocytes in the innate immune system that can directly recognize and eliminate virally infected or malignantly transformed tumor cells. NK cells exert anti-tumor effects by releasing perforin/granzymes, FasL- and TRAIL-mediated apoptosis pathways, and producing cytokines. They can also shape adaptive immune responses. Exosomal circUHRF1 derived from cancer cells can be taken up by NK cells and act as a competitive endogenous RNA, sponging miR-449c-5p. This relieves the inhibition of the immunosuppressive receptor TIM-3, leading to TIM-3 upregulation and suppression of IFN-γ secretion by NK cells. NK cell dysfunction and reduced infiltration further create an immunosuppressive microenvironment, promoting tumor immune escape and weakening the efficacy of anti-PD-1 immunotherapy. Elevated plasma exosomal circUHRF1 levels are associated with poor prognosis.53 Furthermore, research by Wang et al found that exosomes released by tumor cells are enriched with PD-L1, which can bind to PD-1 on the surface of NK cells, significantly inhibiting their activation and cytotoxicity, thereby providing a barrier for tumor immune escape.54 Blocking the PD-1/PD-L1 axis (using inhibitors or antagonists) can reverse the inhibitory effect of exosomal PD-L1 on NK cells and restore anti-leukemia immune function. Research by Tang et al found that miR-552-5p is significantly elevated in patient plasma exosomes. It can be taken up by NK cells, downregulating activating receptors such as NKp30, NKp46, and NKG2D, and inhibiting the secretion of perforin, granzymes, and IFN-γ.55 This attenuates NK cell killing function and is clinically associated with poorer survival prognosis. Exosomes released by tumor cells are enriched with lncRNA NEAT1, which can recruit the histone methyltransferase EZH2 to silence the transcription factor PBX1. This suppresses NKG2D expression on NK cells and impairs their ability to secrete TNF-α and IFN-γ, leading to a significant decline in NK cell killing activity. Impaired NK function not only contributes to tumor immune escape but is also accompanied by upregulation of the tumor proliferation marker Ki-67 and PD-L1, accelerating myeloma growth and progression.56 Silencing NEAT1 or blocking the NEAT1-EZH2-PBX1 pathway can restore NK cell effector function and reduce tumor burden.
Tumor EVs display checkpoint ligands and carry immunoregulatory factors that dampen T-cell activation, induce regulatory T-cell expansion, and skew macrophages toward an M2-like phenotype. They impair dendritic-cell maturation and antigen presentation, while fostering the accumulation of myeloid-derived suppressor cells, thereby blunting antitumor immunity. The net effect is a tolerant tumor immune microenvironment less responsive to immune surveillance and checkpoint blockade. Such EV-mediated immune editing correlates with disease progression and poor therapeutic control.
EV-Mediated Angiogenesis in Osteosarcoma
Angiogenesis provides essential support for solid tumor growth and metastasis. Osteosarcoma grows rapidly and has a high demand for blood supply. EV-derived ncRNAs play an indispensable role in promoting tumor angiogenesis. Osteosarcoma cells transmit pro-angiogenic signals to vascular endothelial cells via exosomes, inducing the latter’s proliferation, migration, and capillary structure formation.
ncRNAs carried by EVs can also transfer between tumor cells, regulating their ability to secrete pro-angiogenic factors. Research by Li et al found that exosomes secreted by osteosarcoma cells are highly enriched with lncRNA OIP5-AS1, which competitively binds to miR-153, relieving miR-153’s inhibition of the autophagy key molecule ATG5. Suppressed autophagy causes a slight increase in intracellular ROS and activates HIF-1α, which transcriptionally upregulates VEGFA and ANGPT2.57 Concurrently, excessive ATG5 indirectly promotes phosphorylation of the FAK-Src-paxillin complex, enhancing tumor cell secretion of MMP-9 and soluble VEGFR1. These changes lead to a significant increase in local VEGF expression and accelerated matrix degradation, thereby markedly promoting angiogenesis and perfusion capacity.
In the TME, EVs released by bone marrow mesenchymal stem cells carry lncRNA NORAD, which can “sponge” miR-877-3p in the cytoplasm of osteosarcoma cells, upregulating the transcriptional coactivator CREBBP.58 CREBBP binds to and acetylates HIF-1α, enhancing its stability and recruitment efficiency to the promoters of VEGF-A and PDGF-B, while synergizing with p300 to promote transcription of ANGPT2 and IL-8, thereby promoting angiogenesis through multiple pathways. Furthermore, in osteosarcoma cells with loss of RB1 function, uncontrolled E2F drives high-level expression of UHRF1.59 UHRF1 can inhibit AMPK activation and downregulate the anti-angiogenic factor SEMA3E, lifting the natural constraint on endothelial cell migration and angiogenesis. Additionally, UHRF1 also prompts tumor cells to secrete exosomes enriched with urokinase-type plasminogen activator (uPA). uPA-carrying exosomes, upon entering the stroma, initiate the plasmin-MMP cascade, rapidly degrading the matrix and releasing stored pro-angiogenic ligands such as VEGF and FGF, while stimulating endothelial cells to express uPAR and VEGFR2. This series of mechanisms ultimately establishes a high-density microvascular network in distant organs like the lung, providing pathways for tumor dissemination. Similarly, research by Tao et al found that osteosarcoma cells package large amounts of lncRNA EWSAT1 into exosomes and transport them to vascular endothelial cells.60 EWSAT1 directly binds to the 3’UTR region of VEGFR2, promoting its translation and membrane localization. This process significantly enhances endothelial cell sensitivity to ligands like VEGF-A and ANGPT2. Meanwhile, within the tumor itself, EWSAT1 acts as a ceRNA to sponge inhibitors miR-24/miR-330-5p, relieving translational repression of VEGF-A, IL-8, and MMP-9, thereby substantially increasing pro-angiogenic ligand concentration. This superimposed effect rapidly induces the AKT-eNOS and MAPK-ERK signaling cascades, promoting vascular branching, generation, and tumor growth. Interestingly, osteosarcoma cells actively package miR-199a-5p into exosomes and deliver them to human umbilical vein endothelial cells (HUVECs). miR-199a-5p targets and silences VEGFA mRNA, inhibiting VEGF-A/VEGFR2 signaling expression. This leads to simultaneous suppression of the PI3K/Akt/eNOS pathway and the MAPK/ERK branch. These changes manifest as endothelial cell cycle arrest, impaired pseudopod formation, reduced secretion of basement membrane proteases, and a significant decline in lumen formation efficiency.61
Additionally, studies found that MIAT enriched in serum exosomes from osteosarcoma patients enters osteosarcoma cells and competitively binds with miR-613, relieving its inhibition of GPR158 mRNA.62 GPR158, as a GPCR-like membrane receptor, upregulates the AKT/GSK3β/β-catenin pathway and promotes transcription of VEGF-A and ANGPTL4, reinforcing its own pro-angiogenic secretory profile. Bevacizumab not only directly neutralizes VEGF-A but also significantly inhibits tumor tubule formation and in vivo microvessel density by reducing exosomal-MIAT load, elevating miR-613 levels, and downregulating GPR158 expression.
EVs released by osteosarcoma cells and stromal components carry pro-angiogenic cargo. Endothelial uptake of EVs promotes migration, tube formation, and permeability, while diminishing pericyte coverage to stabilize nascent vessels. Hypoxia not only increases EV biogenesis but also enriches their pro-angiogenic payload, amplifying vascular sprouting in the tumor bed. These changes collectively support nutrient delivery and metastatic dissemination.
EV-Driven ECM in Osteosarcoma
The dynamic remodeling of the ECM plays a critical role in local tumor infiltration and metastatic spread. Osteosarcoma originates in bone tissue, where the surrounding matrix primarily consists of bone matrix (collagen, calcified deposits, etc.) along with cartilage and fibrous tissue.38 Osteosarcoma cells secrete enzymes to degrade the bone matrix and promote bone resorption, thereby creating space for their own growth and invasion. Non-coding RNAs (ncRNAs) carried by osteosarcoma-derived EVs effectively facilitate ECM remodeling and degradation.
In the local bone TME, osteosarcoma EVs disrupt the balance of bone remodeling. Garimella et al first reported the role of osteosarcoma exosomes in the bone microenvironment. Their study detected multiple bone-remodeling factors such as matrix metalloproteinases (MMPs), RANK ligand, and TGF-β in exosomes secreted by osteosarcoma cells. These exosomes act on osteoclasts and osteoblasts, disrupting bone remodeling homeostasis and driving bone resorption and dissolution.
Exosomes released by osteosarcoma cells are rapidly taken up by bone marrow-derived macrophages (BMDMs), significantly promoting their differentiation into osteoclasts with high tartrate-resistant acid phosphatase (TRAP) activity. Specifically, exosome-enriched miR-501-3p enters BMDMs, inhibits PTEN, and thereby releases the brake on the PI3K/Akt cascade.63 This activates core osteoresorptive signaling networks (eg, c-Fos/NFATc1) and upregulates effector molecules like TRAP and Cathepsin K. In vitro experiments showed that miR-501-3p-enriched exosomes not only accelerate osteoclast maturation but also cause significant bone loss and destruction. Similarly, sEVs released by osteosarcoma cells carry highly expressed miR-19a-3p, which, upon uptake by RAW264.7 cells and BMMs, enhances pre-osteoclast migration, maturation, and matrix resorption, leading to severe osteoporosis in mouse models. miR-19a-3p also directly suppresses PTEN to activate the PI3K/Akt cascade and accelerate osteoclastogenesis.64 Conversely, miR-146a-5p in osteosarcoma-derived sEVs inhibits NF-κB signaling in pre-osteoclasts, blocking differentiation and drastically reducing mature osteoclast numbers.65 Osteoclast deficiency weakens the bone microenvironment’s barrier against tumor dissemination, facilitating distant metastasis. Patient biopsies with fewer osteoclasts often indicate higher malignancy. By suppressing osteoclasts via exosomes, osteosarcoma maintains moderate bone destruction to preserve a tumor-friendly “space” and growth factor-rich environment.
Regarding soft tissue ECM, osteosarcoma EVs also alter matrix components and stromal cells. Exosomes from osteosarcoma cells are taken up by neighboring fibroblasts, reprogramming them into CAFs. CAFs exhibit pro-tumor phenotypes and secrete matrix-degrading enzymes and invasion promoting molecules. Mazumdar et al demonstrated that TGF-β1 in osteosarcoma exosomes activates the SMAD2 pathway in lung fibroblasts, driving their differentiation into CAFs with myofibroblast features.66 These CAFs produce more MMPs, pro-inflammatory cytokines, etc., altering lung ECM composition and structure to favor tumor cell adhesion and growth.67 Chang et al further identified a specific pathway. Osteosarcoma-secreted exosomal lncRNA linc00881 is taken up by normal lung fibroblasts, where it acts as a ceRNA to sequester miR-29c-3p, thereby relieving inhibition of MMP2.68 This triggers fibroblasts to express high levels of MMP2 and activate the NF-κB pathway. Subsequently, stimulated fibroblasts secrete pro-inflammatory and matrix-remodeling factors (eg, IL-1β, IL-8), transitioning to a CAF phenotype. These changes disrupt lung ECM integrity and induce targeted metastasis of osteosarcoma cells. Wang et al reported that exosomes from adipose-derived mesenchymal stem cells (ADSCs) enter osteosarcoma cells and upregulate collagen β-galactosyltransferase 2 (COLGALT2).69 COLGALT2 upregulation coincides with increased vimentin (an intermediate filament protein linked to cell migration and epithelial-mesenchymal transition) and MMP2/9 (key ECM-degrading enzymes) in tumor cells. Collectively, these changes strongly promote osteosarcoma growth and metastasis.
EVs remodel the ECM locally and distally through multiple pathways. In the bone environment, they promote bone destruction while maintaining a moderately structured niche for tumor survival. In soft tissues and distant organs, they induce fibroblast-to-CAF transformation, driving the secretion of enzymes and cytokines that degrade normal ECM and generate a metastasis-permissive stromal environment.
EV-Mediated Promotion of Proliferation and Invasion in Osteosarcoma
Beyond influencing the microenvironment, ncRNAs carried by EVs can directly act on osteosarcoma cells to promote malignant progression. Jiang et al found that miR-144-3p expression was significantly downregulated in osteosarcoma cell-derived exosomes, while ZEB1 was highly expressed.70 Further research revealed that highly expressed miR-144-3p directly inhibits ZEB1, leading to intracellular Fe2⁺ and ROS accumulation, decreased GSH/GSSG ratio, reduced GPX4 and xCT, and increased ACSL4, thereby triggering ferroptosis and ultimately impairing osteosarcoma cell migration and invasion. Conversely, low expression of miR-144-3p in exosomes suppresses ferroptosis by downregulating ZEB1 expression, promoting tumor cell invasion and migration. Additionally, studies showed that circular RNA circ-LMO7 is significantly downregulated in osteosarcoma tissues and cell lines.71 circ-LMO7 acts as a molecular “sponge” for miR-21-5p, weakening its inhibitory effect on the downstream tumor suppressor gene ARHGAP24. This elevates ARHGAP24 protein levels, blocking the Rho GTPase-related cytoskeleton remodeling pathway and suppressing malignant phenotypes. circ-LMO7 is markedly downregulated in osteosarcoma cell-derived exosomes, promoting tumor cell invasion and migration by impairing tumor suppressor function. Furthermore, exosomes released by cancer-associated fibroblasts (CAFs) are enriched with lncRNA SNHG17, which can be taken up by osteosarcoma cells to promote their migration.72 SNHG17 acts as a molecular sponge to adsorb miR-2861, relieving miR-2861’s inhibition of MMP2, thereby enhancing matrix degradation activity and driving tumor invasion. Knockdown of SNHG17 in vitro and in vivo significantly slowed tumor growth and increased apoptosis, indicating that blocking the SNHG17/miR-2861/MMP2 axis may be a novel therapeutic strategy for osteosarcoma. Liu et al discovered that MSC derived exosomes transfer abundant miR-769-5p to osteosarcoma cells, elevating this microRNA in tumor tissues and cell lines versus normal controls.73 Upregulated miR-769-5p directly inhibits DUSP16, releasing its phosphatase-mediated suppression of the JNK/p38 MAPK cascade. This sustains stress-proliferation signaling activation, enhancing osteosarcoma cell migration, while accelerating lung metastasis in mice. Similarly, Qi et al found that exosomes from BMSCs carry miR-208a, which is taken up by osteosarcoma cells. miR-208a directly binds and downregulates the tumor suppressor PDCD4, relieving its inhibition of the ERK1/2 signaling cascade.74 This increases ERK1/2 phosphorylation and activates downstream pro-malignant transcriptional programs. Inhibiting miR-208a restores PDCD4 levels and suppresses ERK1/2 activation, thereby impeding osteosarcoma cell invasion and metastasis.
Interestingly, MSC-derived exosomes can be phagocytosed by osteosarcoma cells, substantially increasing endogenous miR-22 levels.75 miR-22 overexpression inhibits Twist1 expression, suppressing osteosarcoma cell clonogenicity, DNA synthesis, migration, and invasion. These phenotypes are partially rescued by Twist1 complementation, confirming direct binding between miR-22 and the Twist1 3’UTR. miR-22-loaded MSC-derived exosomes more potently inhibit malignant behaviors, suggesting they act as delivery vehicles to suppress osteosarcoma progression by downregulating Twist1 and modulating CADM1-related pathways.
EVs drive osteosarcoma proliferation and motility by delivering ncRNAs, and oncogenic proteins that activate MAPK/ERK and Wnt/β-cateninpathways. Their cargo induces EMT-like programs, cytoskeletal remodeling mediated by Rho GTPases, and upregulation of MMP2 and MMP9, which promotes invadopodia formation and efficient ECM degradation.
EVs Orchestrate EMT in Osteosarcoma
EMT is a biological process through which epithelial-like tumor cells acquire mesenchymal characteristics and serves as a prerequisite for metastasis. Although osteosarcoma originates from mesenchymal tissue, its cells can exhibit either “epithelial-like” or “mesenchymal-like” heterogeneity. EV-derived ncRNAs enhance proliferative signaling and induce EMT by regulating multiple signaling pathways within tumor cells, thereby increasing the metastatic potential of osteosarcoma. Research by Xiao et al revealed that circ_001569 is significantly elevated in exosomes derived from osteosarcoma cells. By acting as a molecular “sponge” for miR-185-5p, circ_001569 relieves miR-185-5p’s suppression of its target gene FLOT2, leading to FLOT2 upregulation and activation of the EMT-associated protein network. The circ_001569/miR-185-5p/FLOT2 axis is a key driver of EMT in osteosarcoma.76 Additionally, TAZ in osteosarcoma cells directly activates the transcription of miR-135b. In turn, miR-135b suppresses three key negative regulators of the TAZ inhibitory pathway: LATS2, APC, and GSK3β, thereby forming a self-reinforcing positive feedback loop.77 This sustains TAZ signaling activation, driving EMT and proliferation. Further studies showed that TAZ knockdown suppresses cell growth and EMT, while simultaneous miR-135b overexpression partially restores these phenotypes, synergistically amplifying malignant progression. Ma et al found that exosomes released by tumor cells are enriched with miR-20b-5p, which can be taken up and accumulate in the cytoplasm of homologous or neighboring cells. miR-20b-5p directly targets and downregulates TGFBR2, inhibiting TGF-β.78 Reduced TGFBR2 levels activate EMT-related transcriptional programs, resulting in decreased E-cadherin expression, increased vimentin, and significantly enhanced cell migration and invasion. Injection of miR-20b-5p-enriched exosomes induced pronounced pathological damage at tissue sites, accompanied by marked alterations in EMT markers. Similarly, Hua et al demonstrated that under hypoxic conditions, HIF-1α overexpression polarizes macrophages toward the M2-like TAM phenotype. These TAMs transfer circ_0003137-enriched EVs to tumor cells, significantly elevating circ_0003137 levels in the latter.79 This circular RNA directly binds and recruits PTBP1, stabilizing PLOD3 mRNA and upregulating PLOD3 protein. This activates the EMT program and enhances tumor cell migration and invasion. In vivo knockdown of circ_0003137 expression significantly suppressed EMT and tumor growth while prolonging survival in mice.
During osteosarcoma development and progression, EV-derived ncRNAs contribute to tumor immune evasion by inducing an immunosuppressive microenvironment. They promote blood vessel formation to supply the tumor and remodel the local ECM, impacting osteosarcoma proliferation, migration, and invasion (Figure 3).
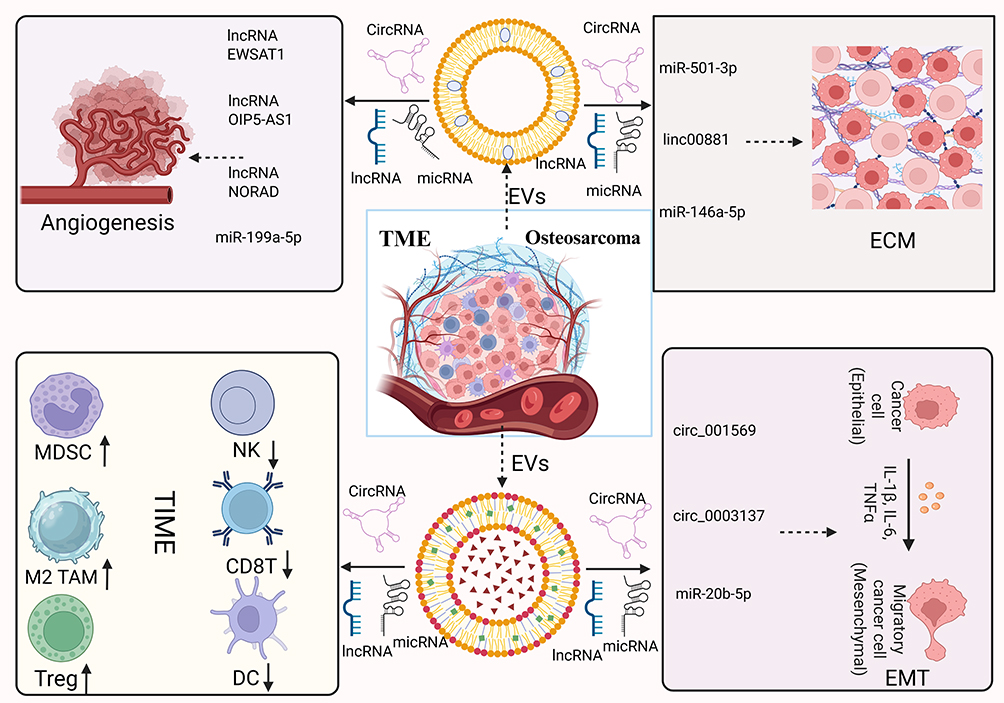 |
Figure 3 The multifaceted roles of EVs in the osteosarcoma microenvironment. Osteosarcoma cells and their diverse EV sources, along with their derived ncRNAs, exhibit multiple biological functions within the TM). EV-carried molecules such as miR-21, miR-143, circ\_103801, TGF-β1, VEGF-A, and MMP-2/9 can drive immune suppression, promote ECM remodeling and invasion, induce angiogenesis, activate EMT, and ultimately remodel the TME. This Figure was created by biorender. |
Pre-Metastatic Niche Formation Driven by EVs
Before metastasizing to distant sites, solid tumors often establish a microenvironment in target organs that favors tumor cell survival and growth, known as the pre-metastatic niche (PMN). The PMN encompasses a series of changes in the metastatic organ, such as increased vascular permeability, matrix remodeling, immune cell recruitment and polarization, “pre-adapting” the organ to the colonization needs of circulating tumor cells. Mounting evidence indicates that exosomes secreted by primary tumors are key drivers of PMN formation.
Initially, Silva et al demonstrated that exosomes from cancer cells are taken up by hepatic Kupffer cells, inducing TGF-β release, which activates hepatic stellate cells to overproduce fibronectin, creating a fibrotic matrix.80 This remodeled microenvironment further recruits bone marrow-derived macrophages, establishing an immunosuppressive, tumor-colonization-friendly PMN in the liver and significantly increasing subsequent hepatic metastatic burden. The MIF highly expressed in exosomes is a key driver of these changes. Blocking MIF prevents PMN formation and drastically reduces liver metastasis. In osteosarcoma, EVs released by tumor cells are enriched with S100A11, which binds and activates the deubiquitinase USP9X in lung interstitial macrophages, triggering JAK2/STAT3 signaling and inducing high secretion of CXCL2.46 This chemokine drives massive infiltration of granulocytic MDSCs, shaping an immunosuppressive PMN and paving the way for tumor cell lung colonization. Furthermore, studies found that high levels of the transcription factor IRF5 in tumors correlate with prolonged survival in osteosarcoma patients, primarily by inhibiting the production and targeted delivery of tumor-derived EVs.81 IRF5 retains its own transcripts within vesicles, altering vesicle protein/nucleic acid composition and reducing their delivery efficiency to metastasis-prone sites like the lungs, thereby blocking PMN construction. Conversely, EVs secreted by IRF5-deficient tumor cells show significantly increased quantity and chemotactic capacity, forming immune-evasive PMNs in organs like the lungs, promoting colonization and metastasis of orthotopically or intravenously injected tumor cells. This reveals a novel function of IRF5 in remodeling tumor-derived EV (t-dEV) programming to restrict metastasis, providing a potential target for PMN intervention. Exosomes released by cancer cells carry high levels of miR-25-3p and are taken up by vascular endothelial cells, downregulating transcription factors KLF2 and KLF4.82 This further modulates VEGFR2 and the expression of tight junction proteins ZO-1 and Claudin5. These changes weaken the endothelial barrier and accelerate neovascularization, significantly increasing vascular permeability. The vascular leakage induced by exosomal miR-25-3p creates a favorable microenvironment for heterogeneous tumor cell colonization in the liver and lungs, markedly enhancing distant metastasis. Circulating exosomal miR-25-3p levels are significantly higher in metastatic patients than in non-metastatic patients, suggesting its role as a key driver of PMN formation. Hypoxia enhances the enrichment of miR-4508 in tumor cell exosomes. These hypoxia-derived exosomes (H-TDEs), upon uptake by lung fibroblasts, downregulate the transcription factor RFX1.83 Reduced RFX1 releases transcriptional inhibition of IL-17A, promoting IL-17A secretion and activating the p38 MAPK-NF-κB cascade. This drives fibroblasts toward a pro-inflammatory, pro-ECM remodeling phenotype, establishing an immunosuppressive and matrix-remodeling PMN that significantly enhances tumor cell colonization and growth in the lungs. Similarly, exosomes secreted by tumor cells under hypoxia are enriched with circular RNA Circ-ZNF609.84 Upon uptake by HUVECs, these exosomes simultaneously activate angiogenesis and disrupt tight junctions. Circ-ZNF609 acts as a “sponge” for miR-150-5p, relieving its inhibition of VEGFA and promoting endothelial proliferation and neovascularization. Additionally, Circ-ZNF609 competes with the RNA-binding protein HuR, blocking HuR’s binding to Claudin-1, reducing the translation of these barrier proteins and increasing vascular permeability. These synergistic effects facilitate PMN formation in lung tissue, significantly enhancing tumor cell distant metastasis.
Osteosarcoma remodels the microenvironment of distant metastatic organs by releasing EVs and their ncRNAs (Table 1). At PMN sites, EVs and their ncRNA derivatives remodel normal stroma into an environment rich in degradative enzymes and inflammatory factors through pro-inflammatory and pro-fibrotic actions. Second, they construct an immunosuppressive microenvironment, inducing the inactivation of local anti-tumor immune cells. Meanwhile, they promote vascular alterations that facilitate tumor cell entry. Together, these mechanisms vastly increase the probability of circulating osteosarcoma cells successfully colonizing metastatic sites and growing into metastatic lesions.
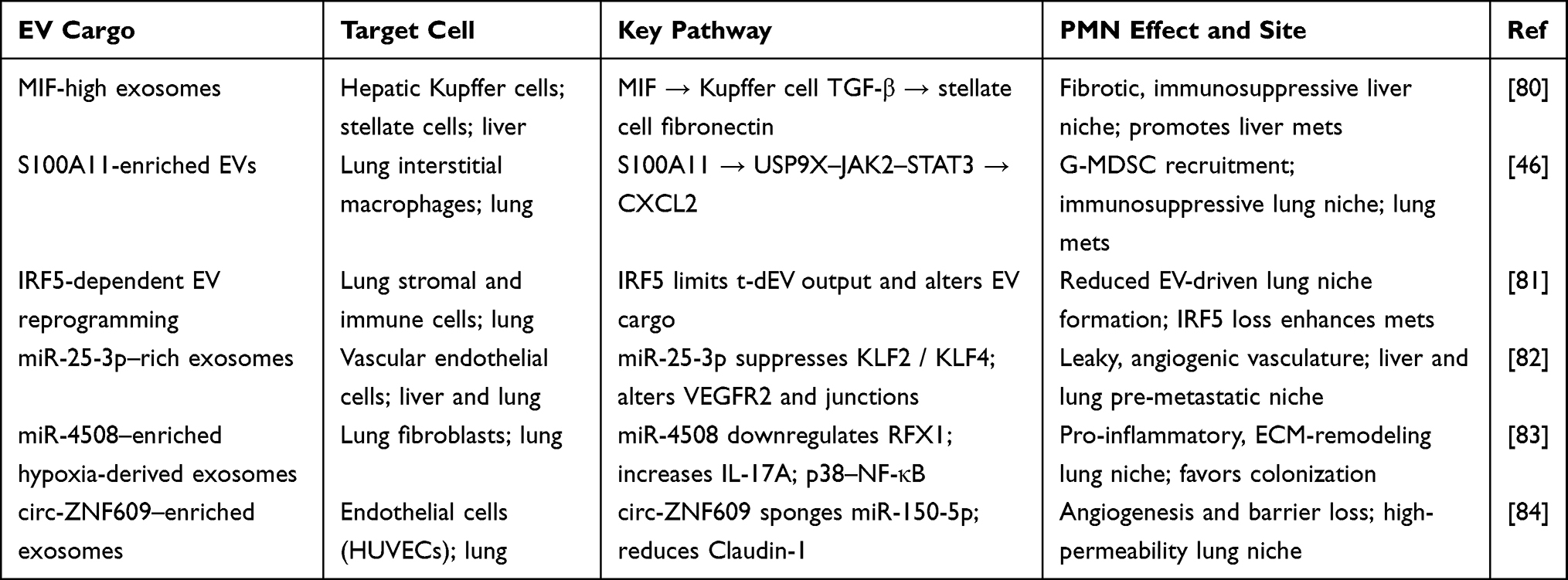 |
Table 1 EV-Driven Mechanisms of Pre-Metastatic Niche Formation |
EVs from osteosarcoma preferentially accumulate in the lung, where they prime stromal and endothelial cells to increase vascular leakiness and inflammatory signaling. They activate resident fibroblasts, remodel ECM architecture, and recruit bone-marrow–derived cells through chemokine and alarmin pathways, establishing a receptive soil for incoming tumor cells. EV integrin and glycoprotein patterns may encode organotropism, guiding niche formation at specific sites. This conditioned niche accelerates seeding efficiency and outgrowth of disseminated osteosarcoma cells.
Mechanisms of EVs-Driven Resistance in Osteosarcoma
EVs and their derived ncRNAs play a crucial role in chemoresistance in osteosarcoma. Exosomes from drug-resistant cells can transfer resistance-associated miRNAs or lncRNAs to sensitive cells, conferring a drug-tolerant phenotype.
lncRNA ANCR was significantly elevated in exosomes from chemoresistant osteosarcoma cells. When taken up by sensitive cells, ANCR rapidly enhanced their in vitro proliferation and tolerance to doxorubicin, while in vivo it promoted tumor growth and reduced mouse survival. ANCR knockdown via siRNA attenuated proliferation in resistant cells and restored drug sensitivity, confirming ANCR as a key molecule maintaining the resistant phenotype. Analysis of patient tumor samples revealed that exosomal ANCR levels negatively correlated with doxorubicin resistance and overall survival, highlighting its prognostic value.85 Additionally, exosomes from chemoresistant osteosarcoma cells were enriched with miR-331-3p, which, upon uptake by sensitive cells, significantly boosted their proliferative capacity and conferred chemoresistance. Further studies showed that miR-331-3p upregulated autophagy-related pathways, sustaining metabolic adaptation under drug stress to counteract chemotherapy toxicity.86 Exosomes released by cisplatin-resistant osteosarcoma cells transported high levels of the transcription factor CTCF into sensitive cells, markedly enhancing their cisplatin tolerance. Zhan et al discovered that CTCF promoted transcription of lncRNA IGF2-AS in recipient cells. IGF2-AS competitively bound miR-579-3p, relieving its suppression of MSH6. This elevated MSH6 expression and initiated autophagy programs. Enhanced autophagy provided metabolic buffering against drug stress, driving tumor cell proliferation, tumorigenicity, and sustained chemoresistance.87 Exosomes from osteosarcoma cells delivered miR-143-3p to recipient cells. This exogenous miR-143-3p activated downstream resistance networks in recipient cells, increasing P-glycoprotein (P-gp) levels and promoting drug efflux, thereby significantly diminishing doxorubicin cytotoxicity. Inhibiting miR-143-3p reversed the exosome-conferred resistant phenotype.88 Furthermore, Pan et al demonstrated that hsa_circ_103801, enriched in exosomes from cisplatin-resistant osteosarcoma cells, substantially upregulated MRP1 and P-gp expression, suppressed apoptosis, and reduced cisplatin cytotoxicity, driving acquired resistance.89 Clinical analysis linked high exosomal hsa_circ_103801 levels to shorter patient survival, indicating its role as a poor prognostic marker. Silencing hsa_circ_103801 or blocking its exosomal transfer restored drug sensitivity, confirming its function as a key driver of cisplatin resistance.
EVs serve as critical carriers for information exchange between tumor cells and their microenvironment, driving chemoresistance through multiple mechanisms. Here, we summarize clinical trials related to EVs and drug resistance in osteosarcoma. (Table 2). Ongoing and completed clinical trials further underscore the translational potential of EV-based biomarkers and novel systemic therapies in osteosarcoma and related malignancies. NCT05101655 is designed to develop and clinically validate a microfluidics-based exosome detection platform for minimally invasive monitoring in osteosarcoma, aiming to enable earlier detection of lung metastasis and dynamic assessment of treatment response.90 NCT03108677 is an observational study profiling circulating exosomal RNA in patients with high-grade osteosarcoma, comparing those with and without pulmonary metastasis to evaluate EV-derived RNA signatures as biomarkers of metastatic risk.91 NCT04690231 is an interventional trial in relapsed or refractory osteosarcoma that compares apatinib plus ifosfamide/etoposide with ifosfamide/etoposide alone, thereby assessing whether VEGFR2 inhibition combined with conventional chemotherapy can improve outcomes in this setting.92 In parallel, NCT03678883 (Actuate 1801) is a phase Ib/II study investigating the safety and preliminary antitumor activity of the GSK-3β inhibitor elraglusib (9-ING-41), administered alone or in combination with standard cytotoxic agents in patients with advanced refractory solid tumors and sarcomas, providing early clinical evidence for pathway-targeted strategies that may also be relevant to osteosarcoma.93
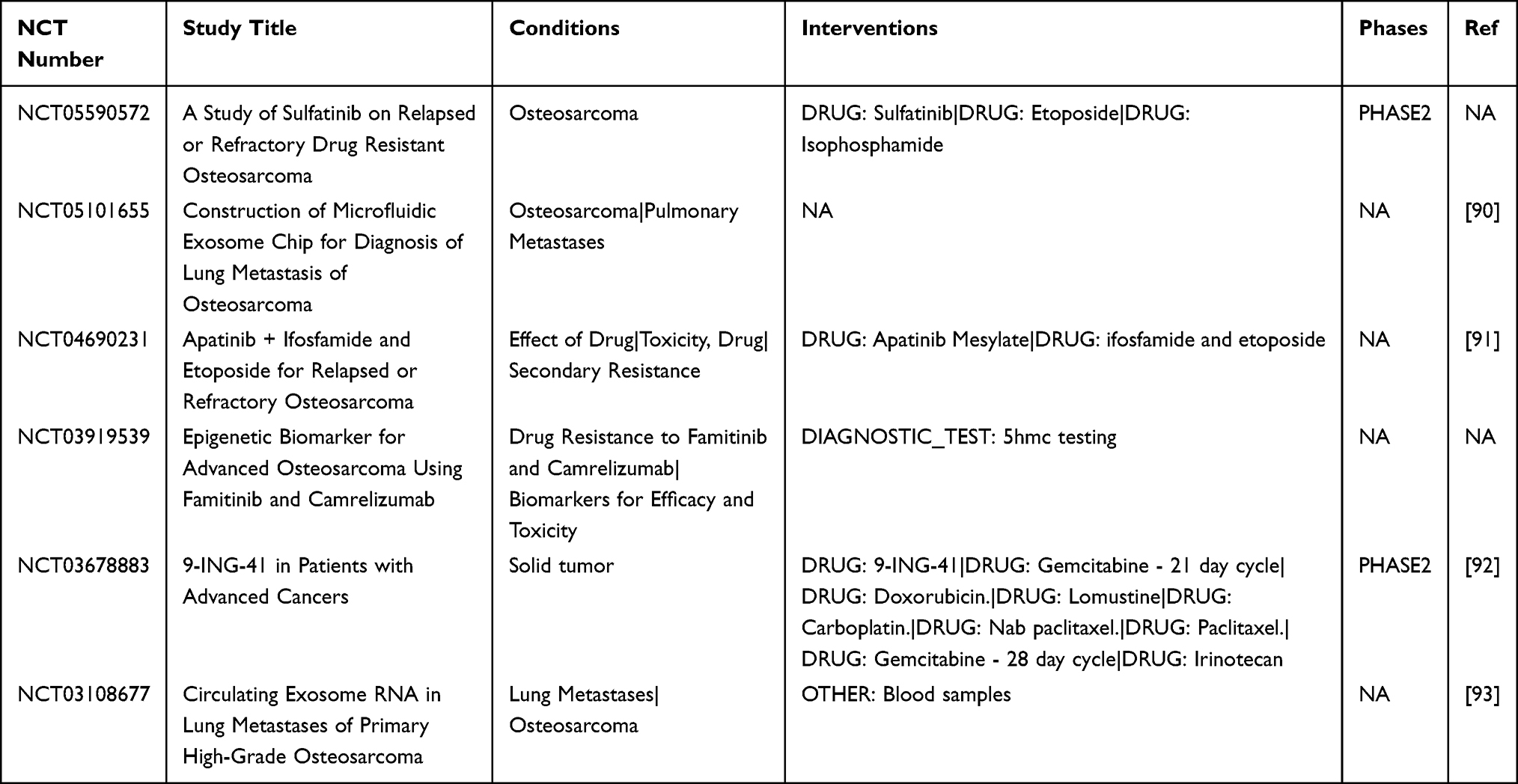 |
Table 2 In Osteosarcoma, Clinical Trials Based on EVs and Their Derived ncRNAs and Drug Resistance |
EV Biomarkers for Diagnosis and Prognosis of Osteosarcoma
EVs and their ncRNAs stably exist in body fluids, and EV-derived ncRNAs show significant differences between healthy individuals and osteosarcoma patients, making them valuable for diagnosis and prognostic evaluation (Table 3). Liquid biopsy, a technique that detects tumor-derived components in blood or other body fluids to obtain tumor information offers advantages over traditional tissue biopsy, including minimal invasiveness and repeatable sampling. Liquid biopsy leverages analytes shed into body fluids to non-invasively capture tumor dynamics, including genomic, proteomic, and vesicular signals.94 Among these analytes, EVs are attractive because their lipid bilayer protects proteins and nucleic acids from enzymatic degradation, enabling longitudinal sampling and improved preanalytical stability compared with many free biomarkers. Consensus guidance from the International Society for EVs codifies minimal reporting items across preanalytics, isolation, and characterization, which is essential for reproducibility and clinical comparability.95 These recommendations emphasize disclosure of particle size and concentration, purity indices, and appropriate positive and negative controls, together with orthogonal readouts to reduce method-specific bias. EVs, capable of stably carrying ncRNAs reflecting tumor characteristics, are considered ideal liquid biopsy biomarker carriers. In recent years, multiple studies have identified potential exosomal ncRNA biomarkers in serum, plasma, and other samples from osteosarcoma patients.96
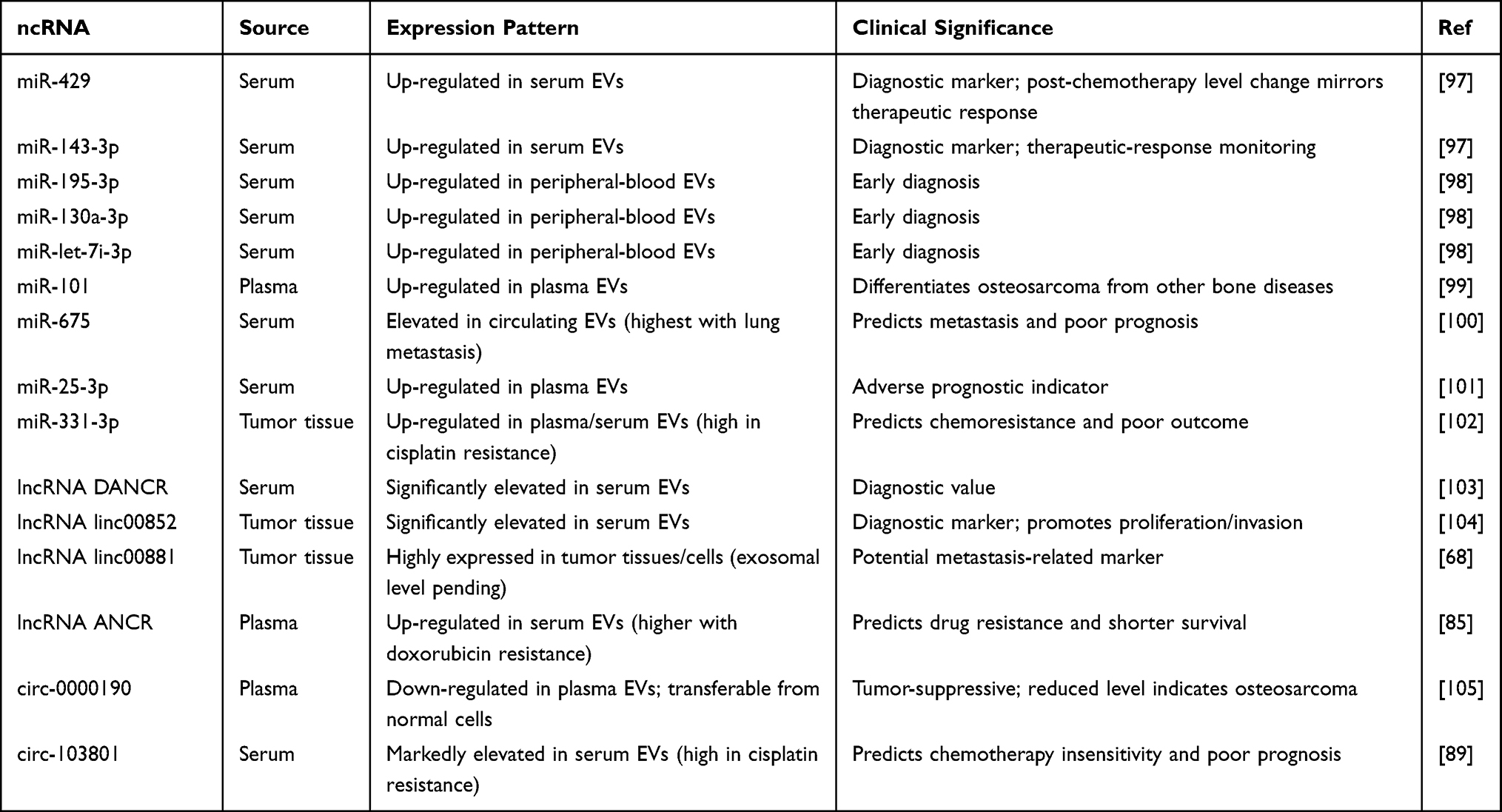 |
Table 3 EV-Derived ncRNA Biomarkers for Osteosarcoma Diagnosis, Prognosis |
EV-Based Liquid Biopsy for Diagnosis
EVs provide a minimally invasive platform for early diagnosis of OS by virtue of their stability in body fluids and encapsulation of tumor-derived molecular cargo.
EV-Derived ncRNAs as Diagnostic Biomarkers
Early studies revealed dysregulated miRNAs in peripheral blood EVs of osteosarcoma patients for disease detection. Yang et al reported that serum exosomal miR-429 and miR-143-3p were significantly elevated in osteosarcoma patients compared to healthy controls, serving as diagnostic markers.97 Ye et al further identified significantly upregulated miR-195-3p, miR-130a-3p, and let-7i-3p in EVs from osteosarcoma patients, supporting their role as early diagnostic biomarkers.98 Zhang et al detected elevated plasma exosomal miR-101 levels in osteosarcoma patients, distinguishing it from other bone diseases.99
EV-derived lncRNAs also show significant value in osteosarcoma identification and diagnosis. Jiang et al found exosomal lncRNA DANCR levels substantially increased in osteosarcoma patients versus healthy individuals, with P<0.05.103 Li et al reported significantly elevated serum exosomal lncRNA linc00852 in osteosarcoma patients. This lncRNA—previously shown to promote proliferation and invasion in osteosarcoma cells stably exists in blood exosomes, suggesting its potential as a diagnostic marker.104
Regarding circRNAs, Li et al reported decreased plasma exosomal circ-0000190 levels in osteosarcoma patients versus normal controls. Normal donor cells could transfer circ-0000190 to osteosarcoma cells via exosomes, inhibiting tumor progression.105 Thus, circ-0000190 acts as a tumor-suppressive circRNA, and its reduced plasma exosomal levels may indicate osteosarcoma presence, supporting its role as an auxiliary diagnostic marker.
EV-Associated Proteins and Other Cargos for Early Detection
In the protein domain, elevated circulating EV-associated TGF-β and other differentially expressed EV proteins have been reported in OS patients compared to healthy controls, enabling discrimination of disease presence. At the nucleic-acid level, EV-associated repetitive-element DNA—including HSATI, LINE1-P1, and Charlie3—is markedly enriched in the serum of patients with osteosarcoma and yields ROC AUCs ≥0.90, indicating strong diagnostic potential.106 Although lipid-only EV biomarker studies in OS are fewer, recent work using Raman or mass-spectrometric lipid/protein fingerprinting of plasma EVs suggests that lipid composition differences may further augment diagnostic classification. Together, EV proteins, EV-DNA repetitive sequences, and EV lipidomic profiles constitute a promising multi-modal biomarker repertoire for OS diagnosis. However, large-scale validation with standardized EV isolation, proper negative controls, and BLANK calibration remains imperative for clinical translation.
EV-Integrated Multi-Analyte Liquid Biopsy Panels
Evidence for EV-based diagnostics in osteosarcoma is moving from descriptive observations to quantitative readouts. Serum exosomal proteins involved in adhesion and immune modulation show discriminative potential. A combined assay of exosomal PD-L1 and N-cadherin associates with increased risk of pulmonary metastasis, supporting their use as adjuncts to imaging for early risk stratification. Proteomics adds breadth to single-analyte testing. In EVs harvested from osteosarcoma tissue explants, twenty-plus proteins are upregulated relative to normal bone–derived vesicles, arguing for multi-marker panels that may outperform single proteins and generalize across cohorts.93 Beyond ncRNAs, EV-associated DNA has emerged as a complementary signal, with exploratory studies reporting enrichment of repetitive elements in patient plasma and high ROC performance. These findings remain preliminary and require independent validation under standardized isolation and quantification protocols. Together, protein-panel approaches, potentially augmented by EV-DNA readouts, outline a feasible path for low-volume, rapid-turnaround diagnostics, provided that thresholds are calibrated prospectively within harmonized analytical pipelines.107
EV-Derived Biomarkers for Osteosarcoma Prognosis
Beyond diagnosis, EV-derived molecular signatures are increasingly recognized as prognostic indicators in OS.
EV-Derived ncRNA Signatures Predicting Outcome
Ev-derived micRNA plays a crucial role in the prognosis of osteosarcoma. For predicting osteosarcoma prognosis, Gong et al observed that circulating exosomal miR-675 levels were significantly higher in patients with lung metastasis than in metastasis-free patients.100 The same study noted that post-chemotherapy changes in miR-429 and miR-143-3p levels correlated with treatment response, suggesting their utility for efficacy monitoring. miR-675 levels inversely correlated with expression of its target gene CALN1 (a potential tumor suppressor) in tumor tissues, suggesting that high miR-675 promotes metastasis and correlates with poor prognosis. Similarly, Yoshida et al linked elevated plasma exosomal miR-25-3p to adverse outcomes in osteosarcoma patients.101 Another study showed that EV-derived miR-331-3p was closely associated with cisplatin resistance, where high expression indicated worse prognosis.102 These findings expand the exosomal miRNA biomarker repertoire for osteosarcoma and provide a basis for developing multi-miRNA assays.
In addition, there are also some related studies on EV-derived lncRNAs. The lncRNA linc00881, identified by Chang et al and highly expressed in osteosarcoma tissues/cells, warrants further evaluation of its exosomal blood levels in relation to metastasis.68 EV-derived lncRNA ANCR expression strongly correlates with doxorubicin resistance, and its high levels predict lower overall survival in patients.85
Regarding circRNAs, circRNA, hsa_circ-103801, was markedly elevated in cisplatin-resistant osteosarcoma cells and patient serum exosomes. Pan et al confirmed that circ-103801 promotes chemoresistance, and serum exosomal circ-103801 levels can predict patient prognosis.89 High circ-103801 indicates chemotherapy insensitivity and poor prognosis, establishing it as an adverse prognostic biomarker.
EV-Associated Proteins and Other Cargos as Prognostic Biomarkers
Elevated serum EV-associated PD-L1 and N-cadherin levels correlate with pulmonary metastasis and advanced disease stage, suggesting potential for risk stratification of metastatic progression.108 Additionally, the abundance of EV-associated SENP1 protein in plasma has been linked to larger tumour size, increased necrosis, lung metastases and poorer disease-free and overall survival in OS patients.109 While EV-DNA repetitive element enrichment has been primarily studied for diagnosis, its dynamic changes may serve as a surrogate for tumour burden or residual disease. Emerging lipid/protein fingerprint studies show that EV lipidomic/biophysical signatures alter with disease progression, indicating that EV composition evolves alongside tumour aggressiveness. Together, these findings position EV proteins, EV-DNA and EV lipidomic profiles as candidate prognostic biomarkers in OS. Prospective multi-centre validation, standardised pre-analytical workflows, and integration with clinical prognostic models are essential next steps.
Multi-Marker EV-Based Liquid Biopsy Panels for Integrated Prognostic Prediction
For prognosis and treatment monitoring, circulating EV proteins involved in immune checkpoint signaling and cellular stress correlate with clinical outcomes. A prospective clinical study demonstrated that baseline serum exosomal PD-L1 stratifies disease-free and overall survival in osteosarcoma, with prespecified concentration thresholds and an association with pulmonary metastasis, supporting its use for baseline risk assessment and on-treatment kinetics.110 In a spontaneous canine osteosarcoma model, mass spectrometry showed that exosomal proteomic signatures correlate with carboplatin response and resistance, indicating that vesicle-number–normalized protein fingerprints can serve as follow-up indicators. Beyond proteins, individual reports link EV-borne SENP1 with tumor burden, necrosis rate, stage, and inferior survival, nominating the de-SUMOylation axis as a potential liquid readout that warrants larger cohorts and mechanistic confirmation.109 Collectively, EV-based panels capturing checkpoint, stress, and inflammatory pathways provide a biologically grounded framework for minimal residual disease surveillance and early progression alerts.
Therapeutic Applications of EVs in Osteosarcoma
EVs and the ncRNAs they carry not only promote osteosarcoma development; some ncRNAs can also inhibit tumor progression. Based on these ncRNAs, extensive research is attempting to develop therapies targeting ncRNAs to block pro-tumor EV communication. Simultaneously, nanotechnology utilizing EVs as drug carriers is rapidly advancing, showing promise in osteosarcoma treatment (Figure 4).
 |
Figure 4 Process of autologous EV therapy for osteosarcoma. The key steps in autologous EV therapy from patient to reinfusion. From raw material collection to engineering modification to therapeutic cargo loading, and finally to administration into the patient. Each step requires careful design and strict control. This Figure was created by biorender. |
Therapeutic Strategies Targeting ncRNAs
Given that multiple EV-carried miRNAs and lncRNAs exhibit pro-tumor effects in osteosarcoma, inhibiting these oncogenic ncRNAs or upregulating tumor-suppressive ncRNAs represents a targeted therapeutic approach. Strategies such as using antisense oligonucleotides (ASOs) or small interfering RNA (siRNA) sequences to knock down oncogenic ncRNAs, employing miRNA mimics to supplement tumor-suppressive miRNAs, or blocking ncRNA packaging into EVs via upstream regulation have gradually become key ideas for treating osteosarcoma.111,112 EVs can be collected and used precisely for the precise treatment of patients (Figure 3).
Zhang et al found that in vitro downregulation of CASC15 expression inhibits osteosarcoma cell migration by releasing miR-338-3p and reducing RAB14 levels.113 Thus, designing ASOs or siRNAs targeting CASC15 may replicate this inhibitory effect in vivo. Similarly, the pro-angiogenic and pro-metastatic lncRNA EWSAT1 is an ideal target.114 Tao et al developed an antisense oligonucleotide (EWSAT1-KD) that precisely targets EWSAT1, effectively reducing EWSAT1 levels in osteosarcoma models and achieving suppression of tumor growth and angiogenesis.60 These results demonstrate that intervening in ncRNAs can reverse their tumorigenic effects. For oncogenic miRNAs like miR-21, “anti-miR” (antagomir) oligonucleotides can be used to specifically bind and functionally block them.115 Similar studies indicate that upregulating tumor-suppressive lncRNAs GAS5 can overcome chemotherapy resistance by competitively binding miR-221-3p, thereby relieving inhibition of DKK2.116 Therefore, altering the ncRNA network balance to disrupt the malignant cycle of tumor cells is feasible.
miR-143, miR-206, miR-29c, etc., exert inhibitory effects in osteosarcoma and are often downregulated in tumor tissues. miR-206 acts as a tumor suppressor in osteosarcoma. Zhang et al found that BMSC-derived exosomes transport miR-206 into osteosarcoma cells and inhibit tumors by targeting the TRA2B gene.117 This suggests that nanocarriers or engineered exosomes could be designed to deliver miR-206 analogs to tumors, mimicking the anti-tumor effects of BMSC exosomes. miR-101 has also been confirmed to inhibit lung metastasis in osteosarcoma. Increasing miR-101 levels in tumor tissue may serve as a strategy to curb metastasis.99 Additionally, another study found that upregulating AGAP1 protein can block the effect of exosomal miR-1307 on tumor cells.118 AGAP1 competitively binds the target mRNA of miR-1307, counteracting its pro-tumor function. This indirectly provides a method for supplementing tumor-suppressive factors.
Engineered EVs for Targeted Drug Delivery
The engineering of EVs has emerged as a pivotal strategy to enhance their therapeutic potential and overcome the inherent limitations of native vesicles in biodistribution, targeting efficiency, and loading capacity. EVs possess intrinsic advantages as delivery vectors due to their nanoscale size, biocompatibility, and ability to traverse biological barriers such as the blood–brain and tumor–stromal interfaces.119 However, native EVs are prone to rapid clearance by the mononuclear phagocyte system and exhibit poor pharmacokinetic control. Thus, rational engineering has been introduced to improve their circulation half-life, tumor selectivity, and therapeutic payload stability.
Genetic and Molecular Engineering
Genetic modification of donor cells enables the generation of EVs enriched with desired ligands, membrane proteins, or therapeutic molecules. Commonly used scaffolds such as LAMP2B and PDGFR have been fused with tumor-homing peptides (eg, RGD, TLYP-1) to confer receptor-specific tropism toward αvβ3- or NRP1-positive tumor vasculature.120 Engineered EVs displaying these ligands exhibit enhanced adhesion to osteosarcoma endothelium and increased intratumoral accumulation. In addition, genetic engineering can direct EVs to carry therapeutic biomolecules including cytokines, siRNAs, or CRISPR–Cas components for gene modulation. Recent studies have also introduced immune checkpoint regulators such as PD-1 or CTLA-4 into EV membranes to enhance tumor immune engagement or suppress immunosuppressive circuits, establishing an interface between EV engineering and immunotherapy.
Chemical and Surface Functionalization
Beyond genetic editing, chemical modification approaches have been developed to fine-tune the physicochemical and targeting properties of EVs. Click chemistry enables precise, bio-orthogonal conjugation of peptides, aptamers, or antibodies to EV membranes without affecting vesicle integrity.14 PEGylation reduces opsonization and prolongs circulation time, while metabolic glycoengineering introduces artificial sugar residues that improve stealth characteristics and ligand density.121 Aptamer- and antibody-functionalized EVs—targeting molecules such as HER2, EpCAM—achieve highly specific tumor recognition and facilitate endocytic uptake. These approaches are particularly valuable in osteosarcoma, where integrin-rich vasculature and dense ECM hinder drug penetration.
Enhanced Cargo Loading and Controlled Release
Efficient therapeutic delivery hinges on achieving high encapsulation efficiency together with spatiotemporally controlled release, without compromising vesicle integrity or bioactivity. Contemporary loading strategies such as electroporation, sonication, extrusion, and cellular nanoporation can increase intraluminal payloads of RNAs, proteins, and small-molecule chemotherapeutics, provided that process parameters are tightly controlled to limit membrane disruption, cargo aggregation, or loss of surface markers.122 In practice, electroporation benefits from low-conductivity buffers and pulse optimization to reduce RNA precipitation. Sonication requires duty-cycle tuning to avoid irreversible bilayer damage. Extrusion can equalize size distributions but may alter surface charge; cellular nanoporation boosts vesicle yield and loading by stimulating vesicle biogenesis at the producer-cell level, which can improve scalability for difficult cargos.123 Across methods, rigorous orthogonal quality control is essential, including nanoparticle tracking analysis for size–concentration baselines, transmission or cryo-electron microscopy for morphology, Western blot or immunoassay for canonical markers, and release assays that quantify bioactive cargo rather than total mass alone.
Regulating EV Production or Uptake
Beyond directly targeting ncRNAs themselves, intervening in EV generation and release can reduce the delivery of harmful ncRNAs. Commonly used experimental EV inhibitors like GW4869 (a sphingomyelinase inhibitor) significantly reduce exosome production.124 Studies in tumor models have verified that applying GW4869 attenuates tumor metastasis and immunosuppressive effects because tumor cells cannot efficiently transmit pro-tumor signals externally.125 However, the specificity and in vivo safety of such drugs require consideration. Additionally, blocking EV uptake by recipient cells. For example, using competitive antibodies to occlude key receptor-ligand molecules (eg, integrins, tetraspanins) on EV membranes can reduce EV-ncRNA entry into target cells. Nevertheless, systemic application of this strategy may disrupt normal intercellular communication and warrants cautious evaluation. Interestingly, some drugs can alter the quantity or cargo of tumor EVs, enabling them to exert anti-tumor effects. Notaro et al reported that the cannabinoid receptor agonist WIN significantly increases exosome secretion by osteosarcoma cells, but exosomes released by WIN-treated cells exhibit anti-migratory effects, inhibiting the motility and invasion of untreated tumor cells.126,127 Although the specific mechanism remains unclear, this finding suggests that certain drugs may achieve anti-tumor effects by influencing EV pathways, offering innovative therapeutic approaches.
Combined Immunotherapy Strategies
Given the role of EV-ncRNAs in forming immunosuppressive microenvironments, targeting these ncRNAs may synergize with immunotherapy. Blocking the release or function of PD-L1 in osteosarcoma EVs could enhance the efficacy of ICIs. Similarly, inhibiting EV-induced M2 polarization (eg, targeting ELFN1-AS1 or miR-221-3p) would help alter macrophage composition in the TME, boosting anti-tumor immunity.128 This indicates that in the era of immunotherapy, therapies targeting EV-ncRNAs could serve as adjuncts to convert “cold” osteosarcoma into “hot” tumors, improving the immune system’s response to tumors. Currently, most ncRNA-targeting therapies remain in preclinical stages, but encouraging evidence exists. For instance, the success of EWSAT1-KD and the effects of CASC15 knockdown provide models for developing nucleic acid-based drugs for osteosarcoma. With advances in nucleic acid delivery technologies (eg, lipid nanoparticles, viral vectors), efficient delivery of these ncRNA inhibitors to osteosarcoma lesions may become feasible.129,130 Future strategies may combine anti-ncRNA therapies with traditional surgery, chemotherapy, and emerging immunotherapies to build multi-pronged comprehensive treatment approaches to overcome metastasis and drug resistance in osteosarcoma.
EVs as Drug Carriers
EVs, especially exosomes, are viewed as highly promising nanocarriers for drug delivery due to their low immunogenicity, excellent biocompatibility, and natural transmembrane transport capacity. Compared to synthetic nanoparticles, cell-derived EVs more easily evade immune clearance and efficiently fuse with target cells. For osteosarcoma, a tumor requiring systemic therapy yet constrained by vascular barriers using EVs as drug carriers may increase drug concentration at tumor sites while reducing systemic toxicity.
Chemotherapy Drug Delivery
Wei et al reported a remarkable achievement: they loaded the chemotherapeutic drug doxorubicin (Dox) into MSC-derived exosomes to prepare an “Exo-Dox” nanoformulation. In in vitro and in vivo experiments, Exo-Dox showed stronger osteosarcoma cell-killing effects and lower cardiotoxicity than free doxorubicin.131 The reason is that exosomes specifically enrich and deliver doxorubicin into tumor cells while prolonging the drug’s half-life locally. This study provides evidence for using EVs to enhance chemotherapy efficacy in osteosarcoma. Currently, teams are attempting to load doxorubicin or cisplatin into patient-derived MSC exosomes for treating refractory osteosarcoma patients.132
Gene and ncRNA Drug Delivery
Beyond traditional small-molecule drugs, EVs can deliver gene-editing tools (eg, siRNA, mRNA, CRISPR-Cas9) or ncRNA mimics.133 The aforementioned miRNA mimics could be encapsulated into EVs, whose surfaces are modified with osteosarcoma-targeting peptides for directed tumor delivery. This method theoretically enables efficient delivery of tumor-suppressive miRNAs while reducing degradation and immune reactions against free RNA in the blood. Progress has been made in some tumor models (eg, exosome-delivered let-7 miRNA suppressing lung cancer), but further exploration is needed in osteosarcoma regarding loading efficiency and tissue tropism of EV carriers. Another concept involves using tumor-tropic EV carriers to deliver CRISPR-Cas9 complexes for editing and knocking out specific oncogenes or fusion genes in osteosarcoma cells.134 This requires solving challenges in packaging Cas9 into EVs and ensuring efficient release, but success could open new paths for precision therapy.
Immune Drug and Vaccine Carriers
EVs can also serve as carriers for anti-tumor immune agents. Since osteosarcoma is less responsive to immunotherapy, an improved strategy is to deliver immune stimulants directly to tumor sites. EVs may carry tumor antigen peptides, adjuvants, or RNA vaccines into antigen-presenting cells to enhance immunogenicity. Loading antigens extracted from osteosarcoma cells onto autologous dendritic cell (DC)-derived exosomes and reinfusing them into patients can activate T cells to generate specific immune responses.135 Such “exosome vaccines” have been reported in Phase I trials for melanoma, inducing anti-tumor immunity. For osteosarcoma patients, especially those with minimal residual disease or high relapse risk, exosome vaccines may offer new adjuvant immunotherapy options.136
Cross-Barrier Delivery
Osteosarcoma sometimes metastasizes extraosseously (eg, brain metastases), requiring therapeutic drugs to cross the blood-brain barrier (BBB).137 Studies show EVs can traverse the BBB to deliver drugs into the brain. Thus, EV-based drug delivery may also be useful for rare brain metastases or spinal osteosarcoma causing leptomeningeal infiltration. Similarly, EVs can potentially penetrate dense bone tissue, delivering drugs deep into the bone marrow microenvironment to eliminate residual tumor cells more effectively than free drugs.138
Translating EVs into clinical therapy faces challenges. For instance, large-scale production of high-purity EVs, consistency and efficiency in drug loading, and post-administration biodistribution and clearance require systematic study and standardization. Currently, exosomes are mostly produced from MSCs or allogeneic cell lines due to ease of culture and extraction. Safety-wise, EVs themselves are cell-derived with low immunogenicity; preliminary studies show they do not cause significant adverse immune reactions as carriers. However, potential harmful components in EVs must be excluded, a risk reducible by using engineered producer cells.
EV–Synthetic Carrier Hybrids for Drug Delivery
Hybridizing EVs with synthetic nanocarriers has emerged as a rational strategy to couple the biological recognition of EV membranes with the high payload capacity and tunable release of man-made systems. Early studies established polyethylene glycol mediated membrane fusion between EVs and liposomes as a tractable route to generate chimeric vesicles while preserving key surface proteins, with fusion efficiency verified by Förster resonance energy transfer together with size and morphology readouts by nanoparticle tracking, dynamic light scattering and electron or atomic force microscopy.139 These hybrid vesicles improved intracellular delivery and stability compared with free drug and conventional liposomes, providing a methodological foundation for subsequent applications in oncology. Beyond liposomes, several architectures extend the concept of EV–synthetic integration. EV membranes can be fused or interdigitated with polymeric nanoparticles to form shells that enhance cellular uptake, facilitate endosomal escape and enable stimuli responsive release through pH, redox or enzyme sensitive components in the synthetic layer. Orthogonal chemical strategies, including click chemistry and metabolic glycoengineering, allow post-insertion of targeting ligands and polyethylene glycol to modulate circulation and macrophage uptake, while preserving antigenic motifs required for EV homing. Reviews of hybrid membrane nanovesicles highlight these design spaces and summarize how composition governs pharmacokinetics, biodistribution and immunogenicity across tumor models.
Methodologically, the construction of EV–synthetic hybrids relies on controlled fusion or assembly processes. Polyethylene glycol driven fusion, freeze–thaw cycling, gentle sonication and extrusion can mix lipid bilayers and co-load hydrophobic or hydrophilic cargos without excessive membrane damage when parameters are optimized. Quality control benefits from paired orthogonal assays that quantify fusion (FRET), vesicle number and size (NTA or TRPS), morphology (TEM or Cryo-EM) and marker integrity alongside ligand density and cargo bioavailability.140 Emerging reports also describe polymer brush growth or membrane cloaking around EVs to increase stability and reduce protein corona formation, broadening the engineering toolkit for complex payloads such as nucleic acids and protein therapeutics.
Evidence in solid tumors indicates that EV–liposome hybrids and EV–polymer composites can co-deliver chemotherapeutics with RNA payloads, prolong circulation, and increase intratumoral accumulation relative to either component alone.141 Although osteosarcoma specific datasets remain limited, cross-tumor findings together with generalizable targeting motifs such as RGD family peptides and folate suggest a plausible path to bone-seeking delivery and metastasis-site conditioning.142 Recent translational reviews outline key standardization needs that are directly applicable to hybrids, including definition of critical quality attributes, alignment with MISEV-informed marker panels and adoption of functional potency assays normalized by vesicle number rather than bulk mass. Harmonized release specifications and GMP-compatible purification by size-exclusion or tangential-flow filtration will be essential for batch reproducibility and regulatory evaluation.
EV–synthetic carrier hybrids offer a coherent framework to integrate biological tropism with programmable materials properties. Liposomal fusion provides an experimentally accessible entry point, while polymeric and chemically defined variants expand control over targeting and release. Consolidating construction protocols, orthogonal analytics and stability criteria is likely to accelerate clinical translation and will position these hybrids as credible candidates for precision drug delivery in osteosarcoma and related sarcomas.
EV Standardization Challenges in Clinical Translation
Clinical translation of EV analytics and therapeutics requires a unified framework spanning preanalytics, isolation, identity and potency assessment, quantification, stability, study design, and regulation. In osteosarcoma, where lesions are deep-seated and pulmonary dissemination is common, these constraints are amplified by low circulating signal, lipoprotein-rich matrices, and center-to-center variability.143
Preanalytical control is the first bottleneck. Anticoagulant selection, venipuncture technique, processing delays, platelet removal, storage temperature, and hemolysis alter vesicle yield and phenotype. Without locked standard operating procedures and embedded internal quality controls, downstream differences cannot be attributed to biology.144 Osteosarcoma studies should define acceptable windows from draw to processing, specify double-spin parameters for platelet-poor plasma, and document exclusion criteria for inflammatory flares or strenuous exercise that inflate lipoproteins and confound particle readouts.145
Isolation and purity standardization remain unresolved. Differential ultracentrifugation, size-exclusion chromatography, and tangential-flow filtration enrich overlapping but nonidentical particle populations and frequently co-isolate lipoproteins, protein aggregates, and other non-vesicular nanoparticles. In the absence of reference materials and explicit purity thresholds, batch consistency and potency readouts are difficult to interpret. Reporting should combine orthogonal purity metrics, including particle-to-protein and particle-to-lipid ratios, apolipoprotein depletion indices, and nuclease or protease resistance assays that discriminate membrane-protected cargo from surface adsorption.146 In low-yield osteosarcoma cohorts, targeted immunoenrichment can increase analytical sensitivity but narrows biological breadth, which must be acknowledged when comparing studies.
Metrological traceability limits comparability across platforms. Counts derived from nanoparticle tracking analysis, tunable resistive pulse sensing, and nano-flow cytometry diverge when calibration beads, thresholding, and gating differ. Traceable standards, cross-platform commutability studies, and explicit uncertainty budgets are needed before particle numbers can serve as primary endpoints. Given modest effect sizes in osteosarcoma liquid biopsy, lack of traceability risks false negatives and unstable cut points.147
Identity and potency assays are inconsistently defined. Core markers such as CD81, syntenin, and TSG101 are unevenly adopted and rarely complemented by ligand density, receptor binding competence, or measures of bioavailable rather than total cargo. Potency is often inferred from surrogate readouts rather than mechanism-linked bioassays.148 Osteosarcoma applications should link assays to intended clinical function, for example checkpoint signaling in lymphocytes, macrophage polarization indices, endothelial permeability, or matrix remodeling, and normalize effects to vesicle number or membrane area rather than bulk protein mass.
Stability and biobanking introduce additional uncertainty. Freeze–thaw cycles, buffer composition, cryoprotectants, and lyophilization can alter membrane integrity, aggregation state, and cargo activity.149 Fit-for-purpose stability studies should define acceptance criteria for particle recovery, size distribution, surface markers, and functional potency over storage intervals that mirror clinical workflows. This is particularly important for longitudinal surveillance, where comparability depends on stability-indicating assays.
Clinical validity and utility require prospective, preregistered designs. Assays should be locked before validation with prespecified endpoints such as diagnostic discrimination, metastasis risk, minimal residual disease, or treatment response. Models must adjust for clinical covariates and undergo external, multicenter verification to demonstrate generalizability.150 For osteosarcoma, pulmonary metastasis risk and response to neoadjuvant chemotherapy are natural endpoints that can anchor utility claims.
Regulatory alignment is still emerging. Jurisdictions variably classify EVs as biologics, gene therapy vectors, or advanced therapy medicinal products. Each designation entails distinct expectations for manufacturing controls, release specifications, potency testing, and pharmacovigilance. Early dialogue with regulators should agree on product definitions, critical quality attributes, lot-release criteria, and GMP-compatible analytics.151 When EVs serve as analytes rather than therapeutics, laboratories should still adopt proficiency testing, shared reference EVs, and transparent reporting to facilitate cross-center pooling.
Osteosarcoma-specific considerations include low circulating abundance, interference from lipoproteins and acute-phase proteins in adolescent patients, and the need to separate signals from primary bone lesions and lung metastases. Strategies that enrich disease-relevant subpopulations through immunocapture, single-particle phenotyping by nano-flow cytometry, and paired cargo–function readouts can mitigate these constraints, provided that capture bias is quantified and reported. Integrating these elements into a harmonized workflow will convert EV assays from descriptive tools into clinically reliable diagnostics and will de-risk the path for EV-based therapeutics.
Conclusion
EVs drive osteosarcoma pathogenesis through multidimensional mechanisms. EVs create a permissive TME for local growth and proliferation by inducing immunosuppression, promoting angiogenesis, and remodeling the ECM. Additionally, they fuel metastasis by enhancing tumor cell invasion, migration, and EMT. Critically, EVs establish PMNs in distant organs, laying the groundwork for metastatic colonization. Clinically, EV-derived ncRNAs serve as novel biomarkers, enabling early diagnosis and precise prognostic prediction for osteosarcoma. Targeting these key ncRNAs holds promise for suppressing tumor progression, while leveraging EVs as drug delivery vehicles could significantly enhance therapeutic efficacy. In immunotherapy, modulating EV-carried ncRNAs may reverse the “cold” tumor status of osteosarcoma, improving response to immune-based treatments. Future advancements in nucleic acid therapeutics and EV-based nanomedicines may undergo clinical trials, ultimately benefiting patients with drug-resistant or metastatic osteosarcoma. However, rigorous preclinical and clinical validation is essential to confirm the safety and efficacy of these strategies. In conclusion, EVs and their ncRNA cargo unveil new perspectives on osteosarcoma development and treatment, offering renewed hope for improving patient outcomes.
Consent for Publication
Consent for publication was obtained from all individual participants included in the study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors declare that they have no conflict of interest.
References
1. Yu S, Yao X. Advances on immunotherapy for osteosarcoma. Mol Cancer. 2024;23(1):192. doi:10.1186/s12943-024-02105-9
2. Beird HC, Bielack SS, Flanagan AM, et al. Gorlick R: osteosarcoma. Nat Rev Dis Primers. 2022;8(1):77. doi:10.1038/s41572-022-00409-y
3. Wen Y, Tang F, Tu C, Hornicek F, Duan Z, Min L. Immune checkpoints in osteosarcoma: recent advances and therapeutic potential. Cancer Lett. 2022;547:215887. doi:10.1016/j.canlet.2022.215887
4. Nirala BK, Yamamichi T, Petrescu DI, Shafin TN, Yustein JT. Decoding the impact of tumor microenvironment in osteosarcoma progression and metastasis. Cancers. 2023;15(20):5108. doi:10.3390/cancers15205108
5. Yu L, Zhang J, Li Y. Effects of microenvironment in osteosarcoma on chemoresistance and the promise of immunotherapy as an osteosarcoma therapeutic modality. Front Immunol. 2022;13:871076. doi:10.3389/fimmu.2022.871076
6. Jeppesen DK, Zhang Q, Franklin JL, Coffey RJ. Extracellular vesicles and nanoparticles: emerging complexities. Trends Cell Biol. 2023;33(8):667–28. doi:10.1016/j.tcb.2023.01.002
7. Toden S, Zumwalt TJ, Goel A. Non-coding RNAs and potential therapeutic targeting in cancer. Biochim Biophy Acta Rev Cancer. 2021;1875(1):188491. doi:10.1016/j.bbcan.2020.188491
8. Nemeth K, Bayraktar R, Ferracin M, Calin GA. Non-coding RNAs in disease: from mechanisms to therapeutics. Nat Rev Genet. 2024;25(3):211–232. doi:10.1038/s41576-023-00662-1
9. Sha G, Zhang W, Jiang Z, Zhao Q, Wang D, Tang D. Exosomal non-coding RNA: a new frontier in diagnosing and treating pancreatic cancer: a review. Int J Biol Macromol. 2024;263(Pt 1):130149. doi:10.1016/j.ijbiomac.2024.130149
10. Zhang W, Jiang Z, Tang D. The value of exosome-derived noncoding RNAs in colorectal cancer proliferation, metastasis, and clinical applications. Clin Transl Oncol. 2022;24(12):2305–2318. doi:10.1007/s12094-022-02908-6
11. Marar C, Starich B, Wirtz D. Extracellular vesicles in immunomodulation and tumor progression. Nat Immunol. 2021;22(5):560–570. doi:10.1038/s41590-021-00899-0
12. Zhang X, Zhang H, Gu J, et al. Engineered extracellular vesicles for cancer therapy. Adv Mat. 2021;33(14):e2005709. doi:10.1002/adma.202005709
13. Kumar MA, Baba SK, Sadida HQ, et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduction Targeted Ther. 2024;9(1):27. doi:10.1038/s41392-024-01735-1
14. Rädler J, Gupta D, Zickler A, Andaloussi SE. Exploiting the biogenesis of extracellular vesicles for bioengineering and therapeutic cargo loading. Mol Ther. 2023;31(5):1231–1250. doi:10.1016/j.ymthe.2023.02.013
15. Mehta MJ, Shin D, Park HS, et al. Exosome-based theranostic for gastrointestinal cancer: advances in biomarker discovery and therapeutic engineering. Small Methods. 2025;9(7):e2402058. doi:10.1002/smtd.202402058
16. Perut F, Roncuzzi L, Baldini N. The Emerging Roles of Extracellular Vesicles in Osteosarcoma. Front Oncol. 2019;9:1342. doi:10.3389/fonc.2019.01342
17. Sonbhadra S, Mehak Pandey LM, Pandey LM. Biogenesis, isolation, and detection of exosomes and their potential in therapeutics and diagnostics. Biosensors. 2023;13(8):802. doi:10.3390/bios13080802
18. Yang Q, Liu J, Wu B, Wang X, Jiang Y, Zhu D. Role of extracellular vesicles in osteosarcoma. Int J Med Sci. 2022;19(8):1216–1226. doi:10.7150/ijms.74137
19. Théry C, Amigorena S, Raposo G, Clayton A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protocols Cell Biol. 2006;30(1). Chapter 3:Unit 3.22. doi:10.1002/0471143030.cb0322s30
20. Tatischeff I, Larquet E, Falcón-Pérez JM, Turpin PY, Kruglik SG. Fast characterisation of cell-derived extracellular vesicles by nanoparticles tracking analysis, cryo-electron microscopy, and Raman tweezers microspectroscopy. J Extracell Vesicles. 2012;1(1). doi:10.3402/jev.v1i0.19179
21. Dragovic RA, Gardiner C, Brooks AS, et al. Sizing and phenotyping of cellular vesicles using Nanoparticle Tracking Analysis. Nanomedicine. 2011;7(6):780–788. doi:10.1016/j.nano.2011.04.003
22. Gardiner C, Ferreira YJ, Dragovic RA, Redman CW, Sargent IL. Extracellular vesicle sizing and enumeration by nanoparticle tracking analysis. J Extracell Vesicles. 2013;2(1). doi:10.3402/jev.v2i0.19671
23. van der Pol E, van Gemert MJ, Sturk A, Nieuwland R, Van Leeuwen TG. Single vs. swarm detection of microparticles and exosomes by flow cytometry. J Thrombosis Haemostasis. 2012;10(5):919–930. doi:10.1111/j.1538-7836.2012.04683.x
24. Shami-Shah A, Travis BG, Walt DR. Advances in extracellular vesicle isolation methods: a path towards cell-type specific EV isolation. Extracellular Vesicles Circulating Nucleic Acids. 2023;4(3):447–460. doi:10.20517/evcna.2023.14
25. Kowal EJK, Ter-Ovanesyan D, Regev A, Church GM. Extracellular vesicle isolation and analysis by western blotting. Methods Mol Biol. 2017;1660:143–152.
26. Karimi N, Dalirfardouei R, Dias T, Lötvall J, Lässer C. Tetraspanins distinguish separate extracellular vesicle subpopulations in human serum and plasma - Contributions of platelet extracellular vesicles in plasma samples. J Extracell Vesicles. 2022;11(5):e12213. doi:10.1002/jev2.12213
27. Nolan JP, Chiu DT, Welsh JA. Rigor and reproducibility: status and challenges for single vesicle analysis. Extracellular Vesicles Circulating Nucleic Acids. 2022;3(3):244–248. doi:10.20517/evcna.2022.28
28. Vergani E, Daveri E, Vallacchi V, et al. Extracellular vesicles in anti-tumor immunity. Semi Cancer Biol. 2022;86(Pt 1):64–79. doi:10.1016/j.semcancer.2021.09.004
29. Peinado H, Alečković M, Lavotshkin S, et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nature Med. 2012;18(6):883–891. doi:10.1038/nm.2753
30. Liu C, Liu D, Wang S, Gan L, Yang X, Ma C. Identification of the SNARE complex that mediates the fusion of multivesicular bodies with the plasma membrane in exosome secretion. J Extracell Vesicles. 2023;12(9):e12356. doi:10.1002/jev2.12356
31. Yang S, Zheng B, Raza F, et al. Tumor-derived microvesicles for cancer therapy. Biomater Sci. 2024;12(5):1131–1150. doi:10.1039/D3BM01980B
32. Zhou M, Li YJ, Tang YC, et al. Apoptotic bodies for advanced drug delivery and therapy. J Controlled Release. 2022;351:394–406. doi:10.1016/j.jconrel.2022.09.045
33. Wu Y, Ai H, Xi Y, et al. Osteoclast-derived apoptotic bodies inhibit naive CD8(+) T cell activation via Siglec15, promoting breast cancer secondary metastasis. Cell Rep Med. 2023;4(9):101165. doi:10.1016/j.xcrm.2023.101165
34. Wei D, Zhan W, Gao Y, et al. RAB31 marks and controls an ESCRT-independent exosome pathway. Cell Res. 2021;31(2):157–177. doi:10.1038/s41422-020-00409-1
35. Ostrowski M, Carmo NB, Krumeich S, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol. 2010;12(1):19–30. sup pp 11-13. doi:10.1038/ncb2000
36. Larios J, Mercier V, Roux A, Gruenberg J. ALIX- and ESCRT-III-dependent sorting of tetraspanins to exosomes. J Cell Biol. 2020;219(3). doi:10.1083/jcb.201904113
37. Ghossoub R, Lembo F, Rubio A, et al. Syntenin-ALIX exosome biogenesis and budding into multivesicular bodies are controlled by ARF6 and PLD2. Nat Commun. 2014;5(1):3477. doi:10.1038/ncomms4477
38. Huang X, Wang L, Guo H, Zhang W, Shao Z. Single-cell transcriptomics reveals the regulative roles of cancer associated fibroblasts in tumor immune microenvironment of recurrent osteosarcoma. Theranostics. 2022;12(13):5877–5887. doi:10.7150/thno.73714
39. Cheng Z, Wang L, Wu C, Huang L, Ruan Y, Xue W. Tumor-derived exosomes induced M2 macrophage polarization and promoted the metastasis of osteosarcoma cells through Tim-3. Archiv Med Res. 2021;52(2):200–210. doi:10.1016/j.arcmed.2020.10.018
40. Wang B, Wang X, Li P, et al. Osteosarcoma cell-derived exosomal ELFN1-AS1 mediates macrophage M2 polarization via sponging miR-138-5p and miR-1291 to promote the tumorgenesis of osteosarcoma. Front Oncol. 2022;12:881022. doi:10.3389/fonc.2022.881022
41. Wang P, Yang L, Dong J, et al. The sEVs miR-487a/Notch2/GATA3 axis promotes osteosarcoma lung metastasis by inducing macrophage polarization toward the M2-subtype. Can Cell Inter. 2024;24(1):301. doi:10.1186/s12935-024-03488-x
42. Zhang H, Yu Y, Wang J, et al. Macrophages-derived exosomal lncRNA LIFR-AS1 promotes osteosarcoma cell progression via miR-29a/NFIA axis. Can Cell Inter. 2021;21(1):192. doi:10.1186/s12935-021-01893-0
43. Liu W, Long Q, Zhang W, Zeng D, Hu B, Liu S. Chen L: miRNA-221-3p derived from M2-polarized tumor-associated macrophage exosomes aggravates the growth and metastasis of osteosarcoma through SOCS3/JAK2/STAT3 axis. Aging. 2021;13(15):19760–19775. doi:10.18632/aging.203388
44. Guo X, Qiu W, Wang J, et al. Glioma exosomes mediate the expansion and function of myeloid-derived suppressor cells through microRNA-29a/Hbp1 and microRNA-92a/Prkar1a pathways. Int J Cancer. 2019;144(12):3111–3126. doi:10.1002/ijc.32052
45. Zhao Q, Huang L, Qin G, et al. Cancer-associated fibroblasts induce monocytic myeloid-derived suppressor cell generation via IL-6/exosomal miR-21-activated STAT3 signaling to promote cisplatin resistance in esophageal squamous cell carcinoma. Cancer Lett. 2021;518:35–48. doi:10.1016/j.canlet.2021.06.009
46. Deng C, Xu Y, Chen H, et al. Extracellular-vesicle-packaged S100A11 from osteosarcoma cells mediates lung premetastatic niche formation by recruiting gMDSCs. Cell Rep. 2024;43(2):113751. doi:10.1016/j.celrep.2024.113751
47. Ning T, Li J, He Y, et al. Exosomal miR-208b related with oxaliplatin resistance promotes Treg expansion in colorectal cancer. Mol Ther. 2021;29(9):2723–2736. doi:10.1016/j.ymthe.2021.04.028
48. Wang M, Qin Z, Wan J, et al. Tumor-derived exosomes drive pre-metastatic niche formation in lung via modulating CCL1(+) fibroblast and CCR8(+) Treg cell interactions. Cancer Immunol Immunother. 2022;71(11):2717–2730. doi:10.1007/s00262-022-03196-3
49. Gao T, Lin YQ, Ye HY, Lin W-M. Lin WM: miR-124 delivered by BM-MSCs-derived exosomes targets MCT1 of tumor-infiltrating Treg cells and improves ovarian cancer immunotherapy. Neoplasma. 2023;70(6):713–721. doi:10.4149/neo_2023_230711N362
50. Gao X, Han D, Fan W. Down-regulation of RBP-J mediated by microRNA-133a suppresses dendritic cells and functions as a potential tumor suppressor in osteosarcoma. Exp Cell Res. 2016;349(2):264–272. doi:10.1016/j.yexcr.2016.10.019
51. Liu Y, Yin Z, Lu P, et al. Lung carcinoma cells secrete exosomal MALAT1 to inhibit dendritic cell phagocytosis, inflammatory response, costimulatory molecule expression and promote dendritic cell autophagy via AKT/mTOR pathway. Onco Targets Ther. 2020;13:10693–10705. doi:10.2147/OTT.S256669
52. Yang J, Zhang M, Zhang X, et al. Glioblastoma-derived exosomes promote lipid accumulation and induce ferroptosis in dendritic cells via the NRF2/GPX4 pathway. Front Immunol. 2024;15:1439191. doi:10.3389/fimmu.2024.1439191
53. Zhang PF, Gao C, Huang XY, et al. Cancer cell-derived exosomal circUHRF1 induces natural killer cell exhaustion and may cause resistance to anti-PD1 therapy in hepatocellular carcinoma. Mol Cancer. 2020;19(1):110. doi:10.1186/s12943-020-01222-5
54. Wang D, Zhou F, He L, et al. AML cell-derived exosomes suppress the activation and cytotoxicity of NK cells in AML via PD-1/PD-L1 pathway. Cell Biol Int. 2024;48(10):1588–1598. doi:10.1002/cbin.12225
55. Tang CW, Yang JH, Qin JW, et al. Regulation of the PD-1/PD-L1 axis and NK cell dysfunction by exosomal miR-552-5p in gastric cancer. Dig Dis Sci. 2024;69(9):3276–3289. doi:10.1007/s10620-024-08536-0
56. Wang QM, Lian GY, Sheng SM, et al. Exosomal lncRNA NEAT1 inhibits NK-cell activity to promote multiple myeloma cell immune escape via an EZH2/PBX1 axis. Molecular Cancer Res. 2024;22(2):125–136. doi:10.1158/1541-7786.MCR-23-0282
57. Li Y, Lin S, Xie X, Zhu H, Fan T, Wang S. Highly enriched exosomal lncRNA OIP5-AS1 regulates osteosarcoma tumor angiogenesis and autophagy through miR-153 and ATG5. Am J Transl Res. 2021;13(5):4211–4223.
58. Feng D, Li Z, Yang L, et al. BMSC-EV-derived lncRNA NORAD facilitates migration, invasion, and angiogenesis in osteosarcoma cells by regulating CREBBP via delivery of miR-877-3p. Oxid Med Cell Longev. 2022;2022(1):8825784. doi:10.1155/2022/8825784
59. Wu SC, Kim A, Gu Y, et al. UHRF1 overexpression promotes osteosarcoma metastasis through altered exosome production and AMPK/SEMA3E suppression. Oncogenesis. 2022;11(1):51. doi:10.1038/s41389-022-00430-6
60. Tao SC, Huang JY, Wei ZY, Li ZX, Guo SC. EWSAT1 acts in concert with exosomes in osteosarcoma progression and tumor-induced angiogenesis: the “double stacking effect”. Adv Biosyst. 2020;4(9):e2000152. doi:10.1002/adbi.202000152
61. Zhang L, Cao H, Gu G, et al. Exosomal MiR-199a-5p inhibits tumorigenesis and angiogenesis by targeting VEGFA in osteosarcoma. Front Oncol. 2022;12:884559. doi:10.3389/fonc.2022.884559
62. Wang BD, Yu XJ, Hou JC, et al. Bevacizumab attenuates osteosarcoma angiogenesis by suppressing MIAT encapsulated by serum-derived extracellular vesicles and facilitating miR-613-mediated GPR158 inhibition. Cell Death Dis. 2022;13(3):272. doi:10.1038/s41419-022-04620-3
63. Lin L, Wang H, Guo W, He E, Huang K, Zhao Q. Osteosarcoma-derived exosomal miR-501-3p promotes osteoclastogenesis and aggravates bone loss. Cell Signalling. 2021;82:109935. doi:10.1016/j.cellsig.2021.109935
64. Luo T, Zhou X, Jiang E, Wang L, Ji Y, Shang Z. Osteosarcoma cell-derived small extracellular vesicles enhance osteoclastogenesis and bone resorption through transferring MicroRNA-19a-3p. Front Oncol. 2021;11:618662. doi:10.3389/fonc.2021.618662
65. Araki Y, Aiba H, Yoshida T, et al. Osteosarcoma-derived small extracellular vesicles enhance tumor metastasis and suppress osteoclastogenesis by miR-146a-5p. Front Oncol. 2021;11:667109. doi:10.3389/fonc.2021.667109
66. Mazumdar A, Urdinez J, Boro A, et al. Exploring the role of osteosarcoma-derived extracellular vesicles in pre-metastatic niche formation and metastasis in the 143-B xenograft mouse osteosarcoma model. Cancers. 2020;12(11):3457. doi:10.3390/cancers12113457
67. Mazumdar A, Urdinez J, Boro A, et al. Gvozdenovic A: osteosarcoma-derived extracellular vesicles induce lung fibroblast reprogramming. Int J Mol Sci. 2020;21(15):5451. doi:10.3390/ijms21155451
68. Chang X, Tan Q, Xu J, et al. Tumor-derived exosomal linc00881 induces lung fibroblast activation and promotes osteosarcoma lung migration. Can Cell Inter. 2023;23(1):287. doi:10.1186/s12935-023-03121-3
69. Wang Y, Chu Y, Li K, et al. Exosomes secreted by adipose-derived mesenchymal stem cells foster metastasis and osteosarcoma proliferation by increasing COLGALT2 expression. Front Cell Develop Biol. 2020;8:353. doi:10.3389/fcell.2020.00353
70. Jiang M, Jike Y, Liu K, et al. Exosome-mediated miR-144-3p promotes ferroptosis to inhibit osteosarcoma proliferation, migration, and invasion through regulating ZEB1. Mol Cancer. 2023;22(1):113. doi:10.1186/s12943-023-01804-z
71. Luo A, Liu H, Huang C, Wei S. Exosome-transmitted circular RNA circ-LMO7 facilitates the progression of osteosarcoma by regulating miR-21-5p/ARHGAP24 axis. Cancer Biol Ther. 2024;25(1):2343450. doi:10.1080/15384047.2024.2343450
72. Zhao A, Zhao Z, Liu W, et al. Carcinoma-associated fibroblasts promote the proliferation and metastasis of osteosarcoma by transferring exosomal LncRNA SNHG17. Am J Transl Res. 2021;13(9):10094–10111.
73. Liu W, Wang B, Duan A, et al. Exosomal transfer of miR-769-5p promotes osteosarcoma proliferation and metastasis by targeting DUSP16. Can Cell Inter. 2021;21(1):541. doi:10.1186/s12935-021-02257-4
74. Qin F, Tang H, Zhang Y, Zhang Z, Huang P, Zhu J. Bone marrow-derived mesenchymal stem cell-derived exosomal microRNA-208a promotes osteosarcoma cell proliferation, migration, and invasion. J Cell Physiol. 2020;235(5):4734–4745. doi:10.1002/jcp.29351
75. Ruan Q, Wang C, Wu Y, Zhu Q. Exosome microRNA-22 inhibiting proliferation, migration and invasion through regulating Twist1/CADM1 axis in osteosarcoma. Sci Rep. 2024;14(1):761. doi:10.1038/s41598-023-50612-4
76. Xiao B, Zhang X, Li X, Zhao Z. Circ_001569 regulates FLOT2 expression to promote the proliferation, migration, invasion and EMT of osteosarcoma cells through sponging miR-185-5p. Open Life Sci. 2020;15(1):476–487. doi:10.1515/biol-2020-0050
77. Shen S, Huang K, Wu Y, et al. A miR-135b-TAZ positive feedback loop promotes epithelial-mesenchymal transition (EMT) and tumorigenesis in osteosarcoma. Cancer Lett. 2017;407:32–44. doi:10.1016/j.canlet.2017.08.005
78. Ma H, Jiang B, Ren Q, et al. Exosomal miR-20b-5p induces EMT and enhances progression in non-small cell lung cancer via TGFBR2 downregulation. J Biochem Mol Toxicol. 2024;38(12):e70080. doi:10.1002/jbt.70080
79. Wang CW, Muzakky H, Firdi NP, et al. Deep learning to assess microsatellite instability directly from histopathological whole slide images in endometrial cancer. Npj Digital Med. 2024;7(1):143. doi:10.1038/s41746-024-01131-7
80. Costa-Silva B, Aiello NM, Ocean AJ, et al. Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver. Nat Cell Biol. 2015;17(6):816–826. doi:10.1038/ncb3169
81. Roberts BK, Li DI, Somerville C, et al. IRF5 suppresses metastasis through the regulation of tumor-derived extracellular vesicles and pre-metastatic niche formation. Sci Rep. 2024;14(1):15557. doi:10.1038/s41598-024-66168-w
82. Zeng Z, Li Y, Pan Y, et al. Cancer-derived exosomal miR-25-3p promotes pre-metastatic niche formation by inducing vascular permeability and angiogenesis. Nat Commun. 2018;9(1):5395. doi:10.1038/s41467-018-07810-w
83. Jia W, Liang S, Lin W, et al. Hypoxia-induced exosomes facilitate lung pre-metastatic niche formation in hepatocellular carcinoma through the miR-4508-RFX1-IL17A-p38 MAPK-NF-κB pathway. Int J Bio Sci. 2023;19(15):4744–4762. doi:10.7150/ijbs.86767
84. Mao Y, Wang J, Wang Y, Fu Z, Dong L, Liu J. Hypoxia induced exosomal Circ-ZNF609 promotes pre-metastatic niche formation and cancer progression via miR-150-5p/VEGFA and HuR/ZO-1 axes in esophageal squamous cell carcinoma. Cell Death Discovery. 2024;10(1):133. doi:10.1038/s41420-024-01905-8
85. Hu X, Wen Y, Tan LY, et al. Exosomal long non-coding RNA ANCR mediates drug resistance in osteosarcoma. Front Oncol. 2021;11:735254. doi:10.3389/fonc.2021.735254
86. Meng C, Yang Y, Feng W, Ma P, Bai R. Exosomal miR-331-3p derived from chemoresistant osteosarcoma cells induces chemoresistance through autophagy. J Orthopaedic Surg Res. 2023;18(1):892. doi:10.1186/s13018-023-04338-8
87. Zhan H, Xiao J, Wang P, et al. Exosomal CTCF confers cisplatin resistance in osteosarcoma by promoting autophagy via the IGF2-AS/miR-579-3p/MSH6 axis. J Oncol. 2022;2022:9390611. doi:10.1155/2022/9390611
88. Cai T, Zhang C, Zhan T. Transfer of exosomal microRNAs confers doxorubicin resistance in osteosarcoma cells. Mol Med Reports. 2023;27(4). doi:10.3892/mmr.2023.12973
89. Pan Y, Lin Y, Mi C. Cisplatin-resistant osteosarcoma cell-derived exosomes confer cisplatin resistance to recipient cells in an exosomal circ_103801-dependent manner. Cell Biol Int. 2021;45(4):858–868. doi:10.1002/cbin.11532
90. Kumar K, Kim E, Alhammadi M, et al. Recent advances in microfluidic approaches for the isolation and detection of exosomes. TrAC Trends Analytical Chemistr. 2023;159:116912. doi:10.1016/j.trac.2022.116912
91. Xie L, Xu J, Sun X, et al. Apatinib plus ifosfamide and etoposide for relapsed or refractory osteosarcoma: a retrospective study in two centres. Oncol Lett. 2021;22(1):552. doi:10.3892/ol.2021.12813
92. Mahalingam D, Shroff RT, Carneiro BA, et al. Preliminary results from the randomized Phase 2 study (1801 part 3B) of elraglusib in combination with gemcitabine/nab-paclitaxel (GnP) versus GnP alone in patients (pts) with previously untreated metastatic pancreatic ductal adenocarcinoma (mPDAC). J Clin Oncol. 2025;43(16_suppl):4006. doi:10.1200/JCO.2025.43.16_suppl.4006
93. Bao Q, Gong L, Wang J, Wen J, Shen Y, Zhang W. Extracellular vesicle RNA sequencing reveals dramatic transcriptomic alterations between metastatic and primary osteosarcoma in a liquid biopsy approach. Ann Surg Oncol. 2018;25(9):2642–2651. doi:10.1245/s10434-018-6642-z
94. Yu D, Li Y, Wang M, et al. Exosomes as a new frontier of cancer liquid biopsy. Mol Cancer. 2022;21(1):56. doi:10.1186/s12943-022-01509-9
95. Guo S, Wang X, Shan D, et al. The detection, biological function, and liquid biopsy application of extracellular vesicle-associated DNA. Biomarker Res. 2024;12(1):123. doi:10.1186/s40364-024-00661-2
96. Gao X, Gao B, Li S. Extracellular vesicles: a new diagnostic biomarker and targeted drug in osteosarcoma. Front Immunol. 2022;13:1002742. doi:10.3389/fimmu.2022.1002742
97. Yang L, Li H, Huang A. MiR-429 and MiR-143-3p function as diagnostic and prognostic markers for osteosarcoma. Clin Lab. 2020;66(10). doi:10.7754/Clin.Lab.2020.191237
98. Ye Z, Zheng Z, Peng L. MicroRNA profiling of serum exosomes in patients with osteosarcoma by high-throughput sequencing. J Investigative Med. 2020;68(4):893–901. doi:10.1136/jim-2019-001196
99. Zhang K, Dong C, Chen M, et al. Extracellular vesicle-mediated delivery of miR-101 inhibits lung metastasis in osteosarcoma. Theranostics. 2020;10(1):411–425. doi:10.7150/thno.33482
100. Gong L, Bao Q, Hu C, et al. Exosomal miR-675 from metastatic osteosarcoma promotes cell migration and invasion by targeting CALN1. Biochem Biophys Res Commun. 2018;500(2):170–176. doi:10.1016/j.bbrc.2018.04.016
101. Yoshida A, Fujiwara T, Uotani K, et al. Clinical and functional significance of intracellular and extracellular microRNA-25-3p in osteosarcoma. Acta Medica Okayama. 2018;72(2):165–174. doi:10.18926/AMO/55857
102. Zu D, Dong Q, Chen S, et al. Wu B: miRNA-331-3p affects the proliferation, metastasis, and invasion of osteosarcoma through SOCS1/JAK2/STAT3. J Oncol. 2022;2022:6459029. doi:10.1155/2022/6459029
103. Jiang N, Wang X, Xie X, et al. lncRNA DANCR promotes tumor progression and cancer stemness features in osteosarcoma by upregulating AXL via miR-33a-5p inhibition. Cancer Lett. 2017;405:46–55. doi:10.1016/j.canlet.2017.06.009
104. Li Q, Wang X, Jiang N, et al. Exosome-transmitted linc00852 associated with receptor tyrosine kinase AXL dysregulates the proliferation and invasion of osteosarcoma. Cancer Med. 2020;9(17):6354–6366. doi:10.1002/cam4.3303
105. Li S, Pei Y, Wang W, Liu F, Zheng K, Zhang X. Extracellular nanovesicles-transmitted circular RNA has_circ_0000190 suppresses osteosarcoma progression. J Cell Mol Med. 2020;24(3):2202–2214. doi:10.1111/jcmm.14877
106. Cambier L, Stachelek K, Triska M, et al. Extracellular vesicle-associated repetitive element DNAs as candidate osteosarcoma biomarkers. Sci Rep. 2021;11(1):94. doi:10.1038/s41598-020-77398-z
107. Zhai C, Xu J, Yang Y, et al. Heterogeneous analysis of extracellular vesicles for osteosarcoma diagnosis. Anal Chem. 2024;96(23):9486–9492. doi:10.1021/acs.analchem.4c00941
108. Wang J, Zhang H, Sun X, et al. Exosomal PD-L1 and N-cadherin predict pulmonary metastasis progression for osteosarcoma patients. J Nanobiotechnol. 2020;18(1):151. doi:10.1186/s12951-020-00710-6
109. Wang L, Wu J, Song S, et al. Plasma exosome-derived sentrin SUMO-specific protease 1: a prognostic biomarker in patients with osteosarcoma. Front Oncol. 2021;11:625109. doi:10.3389/fonc.2021.625109
110. Lyskjær I, Kara N, De Noon S, et al. Osteosarcoma: novel prognostic biomarkers using circulating and cell-free tumour DNA. European J Cancer. 2022;168:1–11. doi:10.1016/j.ejca.2022.03.002
111. Kok VC, Yu CC. Cancer-derived exosomes: their role in cancer biology and biomarker development. Int J Nanomed. 2020;15:8019–8036. doi:10.2147/IJN.S272378
112. Zhu T, Han J, Yang L, et al. Immune microenvironment in osteosarcoma: components, therapeutic strategies and clinical applications. Front Immunol. 2022;13:907550. doi:10.3389/fimmu.2022.907550
113. Zhang H, Wang J, Ren T, et al. LncRNA CASC15 is upregulated in osteosarcoma plasma exosomes and CASC15 knockdown inhibits osteosarcoma progression by regulating miR-338-3p/RAB14 axis. Onco Targets Ther. 2020;13:12055–12066. doi:10.2147/OTT.S282053
114. Shen D, Liu Y, Liu Y, et al. Long non-coding RNA EWSAT1 promoted metastasis and actin cytoskeleton changes via miR-24-3p sponging in osteosarcoma. J Cell Mol Med. 2021;25(2):716–728. doi:10.1111/jcmm.16121
115. Qi J, Zhang R, Wang Y. Exosomal miR-21-5p derived from bone marrow mesenchymal stem cells promote osteosarcoma cell proliferation and invasion by targeting PIK3R1. J Cell Mol Med. 2021;25(23):11016–11030. doi:10.1111/jcmm.17024
116. Li G, Yan X. Long non-coding RNA GAS5 promotes cisplatin-chemosensitivity of osteosarcoma cells via microRNA-26b-5p/TP53INP1 axis. J Orthopaedic Surg Res. 2023;18(1):890. doi:10.1186/s13018-023-04387-z
117. Zhang H, Wang J, Ren T, et al. Bone marrow mesenchymal stem cell-derived exosomal miR-206 inhibits osteosarcoma progression by targeting TRA2B. Cancer Lett. 2020;490:54–65. doi:10.1016/j.canlet.2020.07.008
118. Han F, Pu P, Wang C, et al. Osteosarcoma cell-derived exosomal miR-1307 promotes tumorgenesis via targeting AGAP1. Biomed Res Int. 2021;2021(1):7358153. doi:10.1155/2021/7358153
119. Rakshit T, Pal S. Extracellular vesicles for drug delivery and theranostics in vivo. JACS Au. 2024;4(2):318–327. doi:10.1021/jacsau.3c00611
120. Liang X, Niu Z, Galli V, et al. Extracellular vesicles engineered to bind albumin demonstrate extended circulation time and lymph node accumulation in mouse models. J Extracell Vesicles. 2022;11(7):e12248. doi:10.1002/jev2.12248
121. Sulthana S, Shrestha D, Aryal S. Maximizing liposome tumor delivery by hybridizing with tumor-derived extracellular vesicles. Nanoscale. 2024;16(35):16652–16663. doi:10.1039/D4NR02191F
122. Krivitsky V, Krivitsky A, Mantella V, et al. Ultrafast and controlled capturing, loading, and release of extracellular vesicles by a portable microstructured electrochemical fluidic device. Adv Mat. 2023;35(44):e2212000. doi:10.1002/adma.202212000
123. Driedonks TAP, Ressel S, Tran Ngoc Minh T, Buck AH, Nolte-’t Hoen ENM. Intracellular localisation and extracellular release of Y RNA and Y RNA binding proteins. J Extracellular Biol. 2024;3(1):e123. doi:10.1002/jex2.123
124. Peng Y, Zhao M, Hu Y, et al. Blockade of exosome generation by GW4869 inhibits the education of M2 macrophages in prostate cancer. BMC Immunol. 2022;23(1):37. doi:10.1186/s12865-022-00514-3
125. Wan X, Fang Y, Du J, Cai S, Dong H. GW4869 can inhibit epithelial-mesenchymal transition and extracellular HSP90α in gefitinib-sensitive NSCLC cells. Onco Targets Ther. 2023;16:913–922. doi:10.2147/OTT.S428707
126. Notaro A, Sabella S, Pellerito O, et al. Involvement of PAR-4 in cannabinoid-dependent sensitization of osteosarcoma cells to TRAIL-induced apoptosis. Int J Bio Sci. 2014;10(5):466–478. doi:10.7150/ijbs.8337
127. Notaro A, Sabella S, Pellerito O, Vento R, Calvaruso G, Giuliano M. The secreted protein acidic and rich in cysteine is a critical mediator of cell death program induced by WIN/TRAIL combined treatment in osteosarcoma cells. Int J Oncol. 2016;48(3):1039–1044. doi:10.3892/ijo.2015.3307
128. He Y, Zhou H, Xu H, You H, Cheng H. Construction of an immune-related lncRNA signature that predicts prognosis and immune microenvironment in osteosarcoma patients. Front Oncol. 2022;12:769202. doi:10.3389/fonc.2022.769202
129. Chen C, Xie L, Ren T, Huang Y, Xu J, Guo W. Immunotherapy for osteosarcoma: fundamental mechanism, rationale, and recent breakthroughs. Cancer Lett. 2021;500:1–10.
130. Orrapin S, Moonmuang S, Udomruk S, Yongpitakwattana P, Pruksakorn D, Chaiyawat P. Unlocking the tumor-immune microenvironment in osteosarcoma: insights into the immune landscape and mechanisms. Front Immunol. 2024;15:1394284. doi:10.3389/fimmu.2024.1394284
131. Xie X, Lian S, Zhou Y, et al. Tumor-derived exosomes can specifically prevent cancer metastatic organotropism. J Controlled Release. 2021;331:404–415. doi:10.1016/j.jconrel.2021.01.030
132. Lotfy A, AboQuella NM, Wang H. Mesenchymal stromal/stem cell (MSC)-derived exosomes in clinical trials. Stem Cell Res Ther. 2023;14(1):66. doi:10.1186/s13287-023-03287-7
133. Zhao J, Cui X, Zhan Q, et al. CRISPR-Cas9 library screening combined with an exosome-targeted delivery system addresses tumorigenesis/TMZ resistance in the mesenchymal subtype of glioblastoma. Theranostics. 2024;14(7):2835–2855. doi:10.7150/thno.92703
134. Lin Y, Wu J, Gu W, et al. Exosome-liposome hybrid nanoparticles deliver CRISPR/Cas9 system in MSCs. Adv Sci. 2018;5(4):1700611. doi:10.1002/advs.201700611
135. Popowski KD, Moatti A, Scull G, et al. Inhalable dry powder mRNA vaccines based on extracellular vesicles. Matter. 2022;5(9):2960–2974. doi:10.1016/j.matt.2022.06.012
136. Luo X, McAndrews KM, Arian KA, et al. Development of an engineered extracellular vesicles-based vaccine platform for combined delivery of mRNA and protein to induce functional immunity. J Controlled Release. 2024;374:550–562. doi:10.1016/j.jconrel.2024.08.017
137. Terstappen GC, Meyer AH, Bell RD, Zhang W. Strategies for delivering therapeutics across the blood-brain barrier. Nat Rev Drug Discov. 2021;20(5):362–383. doi:10.1038/s41573-021-00139-y
138. Kim J, Zhu Y, Chen S, et al. Anti-glioma effect of ginseng-derived exosomes-like nanoparticles by active blood-brain-barrier penetration and tumor microenvironment modulation. J Nanobiotechnol. 2023;21(1):253. doi:10.1186/s12951-023-02006-x
139. Piffoux M, Silva AKA, Wilhelm C, Gazeau F, Tareste D. Modification of extracellular vesicles by fusion with liposomes for the design of personalized biogenic drug delivery systems. ACS Nano. 2018;12(7):6830–6842. doi:10.1021/acsnano.8b02053
140. Mukherjee D, Paul D, Sarker S, et al. Polyethylene glycol-mediated fusion of extracellular vesicles with cationic liposomes for the design of hybrid delivery systems. ACS Appl Bio Mater. 2021;4(12):8259–8266. doi:10.1021/acsabm.1c00804
141. Jhan YY, Palou zuniga G, Singh KA, Gaharwar AK, Alge DL, Bishop CJ. Polymer-coated extracellular vesicles for selective codelivery of chemotherapeutics and siRNA to cancer cells. ACS Appl Bio Mater. 2021;4(2):1294–1306. doi:10.1021/acsabm.0c01153
142. Geng L, Matsumoto M, Yao F, et al. Microfluidic post-insertion of polyethylene glycol lipids and KK or RGD high functionality and quality lipids in milk-derived extracellular vesicles. European J Pharmaceutical Sci. 2024;203:106929. doi:10.1016/j.ejps.2024.106929
143. Welsh JA, Van Der Pol E, Arkesteijn GJA, et al. MIFlowCyt-EV: a framework for standardized reporting of extracellular vesicle flow cytometry experiments. J Extracell Vesicles. 2020;9(1):1713526. doi:10.1080/20013078.2020.1713526
144. Witwer KW, Buzás EI, Bemis LT, et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J Extracell Vesicles. 2013;2(1). doi:10.3402/jev.v2i0.20360
145. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404.
146. Crescitelli R, Lässer C, Lötvall J. Isolation and characterization of extracellular vesicle subpopulations from tissues. Nat Protocols. 2021;16(3):1548–1580. doi:10.1038/s41596-020-00466-1
147. Carney RP, Mizenko RR, Bozkurt BT, et al. Harnessing extracellular vesicle heterogeneity for diagnostic and therapeutic applications. Nature Nanotechnol. 2025;20(1):14–25. doi:10.1038/s41565-024-01774-3
148. Wu M, Wang M, Jia H, Wu P. Extracellular vesicles: emerging anti-cancer drugs and advanced functionalization platforms for cancer therapy. Drug Delivery. 2022;29(1):2513–2538. doi:10.1080/10717544.2022.2104404
149. Görgens A, Corso G, Hagey DW, et al. Identification of storage conditions stabilizing extracellular vesicles preparations. J Extracell Vesicles. 2022;11(6):e12238. doi:10.1002/jev2.12238
150. Zeng H, Guo S, Ren X, Wu Z, Liu S, Yao X. Current strategies for exosome cargo loading and targeting delivery. Cells. 2023;12(10):1416. doi:10.3390/cells12101416
151. Shekari F, Alibhai FJ, Baharvand H, et al. Cell culture-derived extracellular vesicles: considerations for reporting cell culturing parameters. J Extracellular Biol. 2023;2(10):e115. doi:10.1002/jex2.115
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Harnessing EVs-ncRNA for Lung Cancer: From Oncogenic Pathways to Novel Diagnostic and Therapeutic Strategies
Shi Y, Chen H, Li W, An S, Li L, Gao H
International Journal of Nanomedicine 2025, 20:9031-9054
Published Date: 14 July 2025