Back to Journals » Drug Design, Development and Therapy » Volume 18
Exploiting the Zebrafish Model for Sepsis Research: Insights into Pathophysiology and Therapeutic Potentials
Authors He J, Xu P, Chen R, Chen M, Wang B ![]() , Xie Y, Yang Q, Sun D
, Xie Y, Yang Q, Sun D ![]() , Ji M
, Ji M
Received 11 October 2024
Accepted for publication 8 November 2024
Published 22 November 2024 Volume 2024:18 Pages 5333—5349
DOI https://doi.org/10.2147/DDDT.S500276
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Jiaxuan He,1,* Peiye Xu,1,* Rongbing Chen,2 Mengyan Chen,3 Beier Wang,4 Yilun Xie,4 Qinsi Yang,5 Da Sun,1 Mingxia Ji3
1Institute of Life Sciences & Biomedical Collaborative Innovation Center of Zhejiang Province, Wenzhou University, Wenzhou, 325035, People’s Republic of China; 2Department of Biomedical Engineering, City University of Hong Kong, Hong Kong, SAR 999077, People’s Republic of China; 3Department of Critical Care Medicine, Yiwu Central Hospital, the Affiliated Yiwu Hospital of Wenzhou Medical University, Yiwu, 322000, People’s Republic of China; 4Department of Hepatobiliary-Pancreatic Surgery, the Second Affiliated Hospital of Wenzhou Medical University, Wenzhou, 325000, People’s Republic of China; 5Wenzhou Institute, University of Chinese Academy of Sciences, Wenzhou, 325000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Mingxia Ji; Da Sun, Email [email protected]; [email protected]
Abstract: Sepsis, a severe condition instigated by infections, continues to be a primary global cause of death, typified by systemic inflammation and advancing immune dysfunction. Comprehending the complex pathological processes that underlie sepsis is integral to the creation of efficacious treatments. Despite the inability of animal models to entirely reproduce the clinical intricacies related to sepsis, they are invaluable instruments for the exploration and development of therapeutic approaches. Within this context, the zebrafish model is particularly noteworthy due to its genetic tractability, transparency, and appropriateness for high-throughput screening of genetic mutants and therapeutic compounds. This scholarly review emphasizes the crucial role that the zebrafish disease model plays in enhancing our comprehension of sepsis, by exploring its applications in deciphering immune and inflammatory responses, evaluating the consequences of genetic alterations, and examining novel therapeutic agents. The Insights derived from zebrafish research not only augment our understanding of the underlying mechanisms of sepsis, but also possess considerable potential for the transference of these discoveries into clinical therapies, thus potentially transforming the approach to sepsis management. The objective of this scholarly article is to underscore the importance of zebrafish in the realm of biomedical research pertaining to sepsis, and to delineate forthcoming opportunities for utilizing this model in clinical applications.
Keywords: Sepsis, Zebrafish models, Inflammation, Immunology, Drug development
Graphical Abstract:

Introduction
Sepsis represents a significant global health concern, accounting for approximately 20% of all deaths worldwide.1 Annually, it affects approximately 48.9 million individuals, resulting in approximately 11 million deaths. The burden is particularly pronounced in low- and middle-income countries, which account for approximately 85% of sepsis cases and related fatalities. Children are particularly susceptible to sepsis, with nearly half of all global cases occurring among this group, resulting in 2.9 million deaths each year.2 In light of these statistics, there is a pressing need for more scientific research to be conducted on sepsis, as well as the development of more scientific and reliable animal models for the investigation of mechanisms and the testing of drugs.
Traditional rodent models have been instrumental in sepsis research, yet they present significant translational challenges to human sepsis due to species-specific immune responses and progression of the disease.3 These models often use lethal doses of pathogens that are not typically reflective of human sepsis, which generally evolves from localised to systemic infections.4 Furthermore, rodent models tend to produce a more uniform and acute sepsis response compared to the variable and sometimes chronic nature observed in humans.5 Additionally, replicating human-standard care, such as ventilatory support and intensive monitoring, is challenging in these models, leading to a simplified understanding of sepsis and its management.6
Due to the limitations of traditional animal models, zebrafish models have become an invaluable tool in the study of sepsis, Previous studies have shown that zebrafish has certain consistency with mouse models in terms of immune response, cytokine secretion, inflammatory response, and organ dysfunction. For example, Zebrafish and mice show similar upregulation patterns of pro-inflammatory cytokines such as Tumor necrosis factor-alpha (TNF-α) and Interleukin—1β (L-1β) through LPS-induced inflammatory response.7 In addition to its physiological and genetic similarities with humans, it also exhibits unique characteristics such as optical transparency and genetic modification,8,9 enable detailed studies of host-pathogen interactions and the development of sepsis in real-time. By using this model, we are able to analyze inflammatory and immune responses at a cellular and molecular level in depth,10,11 facilitating high-throughput screening of potential therapies and accelerating the discovery of new treatments.12 In addition, advanced imaging and genetic tools in zebrafish can improve understanding of sepsis-related organ dysfunction and recovery,13 thereby advancing the development of more effective sepsis treatments.
The use of zebrafish as a model organism in sepsis research has the potential to make a significant contribution to improving current treatment strategies and discovering new therapeutic pathways. A review of sepsis research will be presented in this review, with special emphasis on zebrafish models as well as the integration and significance of zebrafish models. An overview of the approaches to the construction and application of these models will be provided in order to advance understanding of this deadly and complex disease (Figure 1).
 |
Figure 1 As an emerging sepsis model animal, zebrafish has the advantages of high throughput and easy screening, especially in: (A) Simulating the infection process in human body. (B) Sepsis is a disease with high mortality and morbidity. (C) Having high homology with human genes, unique optical transparency and genetic modifiability. |
The Pathogenesis of Sepsis
Immune Response Mechanisms in Sepsis
Sepsis is a complex syndrome characterized by a dysregulated host response to infection, leading to widespread inflammation, immune dysregulation, and multiple organ dysfunction.14 Understanding the underlying immune mechanisms is crucial for developing effective therapies. The inflammatory cascade in sepsis involves the innate immune system recognizing pathogen-associated molecular patterns (PAMPs) through pattern recognition receptors (PRRs) like Toll-like receptors (TLRs).15 This recognition triggers immune cell activation, leading to the release of cytokines and chemokines such as TNF-α, IL-1, Interleukin-6 (IL-6), and interferons.16,17 These pro-inflammatory cytokines recruit additional immune cells to the infection site, amplifying the inflammatory response in a process known as a “cytokine storm”.18 While essential for combating infections, this cytokine storm can cause severe inflammation, resulting in tissue damage and organ dysfunction if not regulated.19 Overproduction of cytokines increases vascular permeability, leading to fluid leakage into tissues, hypotension, and disseminated intravascular coagulation, which are hallmarks of severe sepsis.20
In sepsis, the immune system shifts from a hyper-inflammatory state to a phase of immune suppression, termed immunoparalysis, significantly compromising the host’s ability to combat infections and coordinate effective immune responses.21 This shift involves complex interactions between the innate and adaptive immune systems.22 Initially, the innate response activates to eliminate pathogens, involving macrophages and dendritic cells that phagocytize pathogens and secrete cytokines to stimulate other immune cells. Excessive or uncontrolled responses lead to tissue damage and systemic inflammation. As sepsis progresses, lymphocyte function and numbers decrease, including T cells, B cells, and natural killer cells.23 This lymphocyte apoptosis is a hallmark of sepsis-induced immunosuppression, diminishing the immune system’s ability to respond to new infections. Impaired phagocytosis by neutrophils and macrophages exacerbates this condition, allowing secondary infections to take hold more readily.24 Additionally, alterations in cytokine profiles occur, with increased levels of anti-inflammatory cytokines inhibiting various immune cells and suppressing the immune response.25 These cytokine shifts reflect broader dysregulation of immune signaling mechanisms, disrupting the balance between pro-inflammatory and anti-inflammatory actions during sepsis.
Sepsis is often characterized by an outbreak of inflammation and multiple organ failure throughout the body, which is often seen in basic research studies (Figure 2).
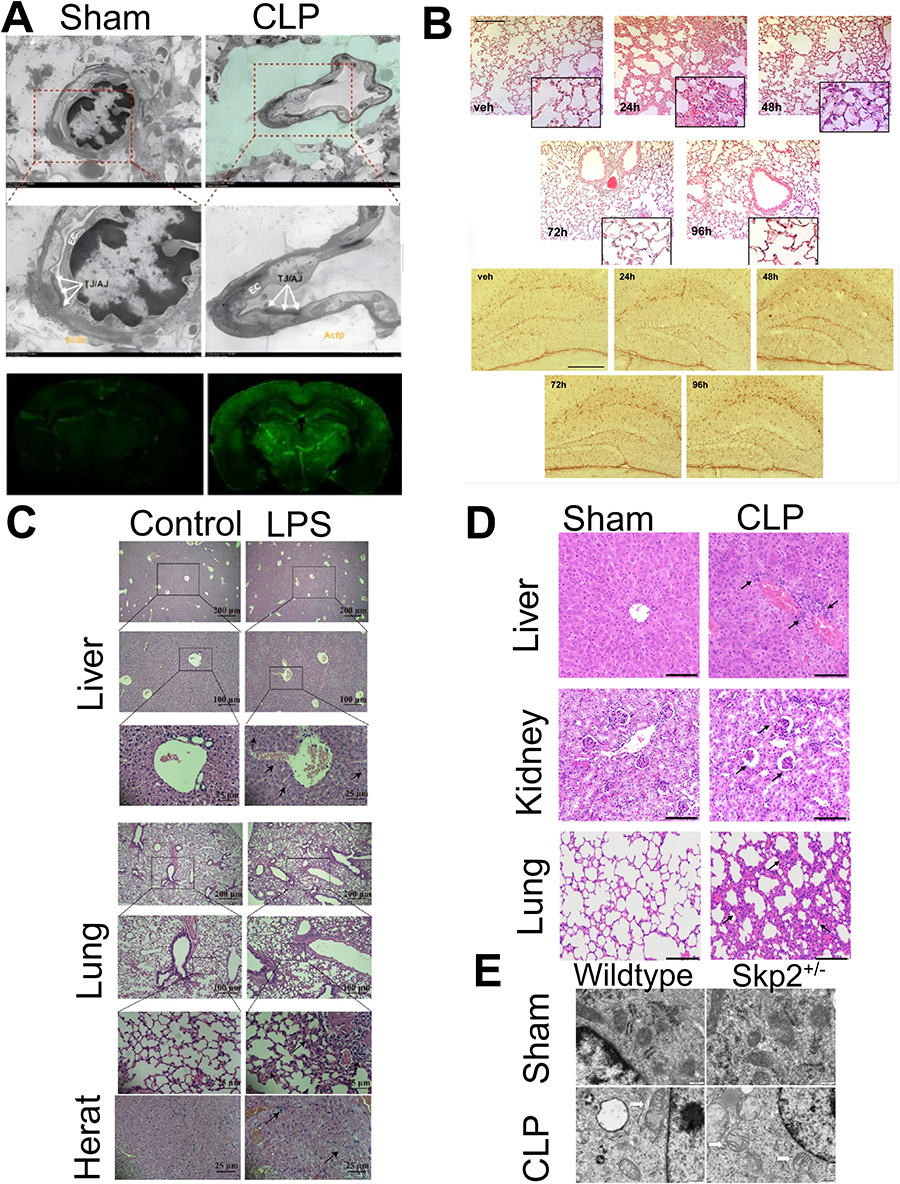 |
Figure 2 (A) The brain of mice with sepsis develops edema and the blood-brain barrier permeability is damaged. Reprinted from Guo J, Kong Z, Yang S, et al. Therapeutic Effects of Orexin-A in Sepsis-Associated Encephalopathy in Mice. J Neuroinflammation. 2024;21:131. Creative Commons.26 (B) Both lung and brain inflammation occurred in mice with aerosolized LPS-induced sepsis. Reprinted from Ritter K, Rissel R, Renz M, Ziebart A, Schäfer MKE, Kamuf J. Nebulized Lipopolysaccharide Causes Delayed Cortical Neuroinflammation in a Murine Model of Acute Lung Injury. Int J Mol Sci. 2024;25(10117). Creative Commons.27 (C) The liver, lungs, and heart of mice with LPS-induced sepsis were damaged by inflammation. Reprinted from Yan C, Kuang W, Jin L, et al. Carvacrol Protects Mice against LPS-Induced Sepsis and Attenuates Inflammatory Response in Macrophages by Modulating the ERK1/2 Pathway. Sci Rep. 2023;13:12809. Creative Commons.28 (D) The liver, kidney, and lung of CLP-induced sepsis mouse models showed inflammatory infiltration. Reprinted from Wang S, Jiang D, Huang F, et al. Therapeutic Effect of Echinococcus Granulosus Cyst Fluid on Bacterial Sepsis in Mice. Parasit Vectors. 2023;16:450. Creative Commons.29 (E) Iron death and mitochondrial damage in alveolar epithelial cells of Skp2±CLP sepsis mice. Reprinted from Chen Z, Zhang J, Gao S, et al. Suppression of Skp2 Contributes to Sepsis-Induced Acute Lung Injury by Enhancing Ferroptosis through the Ubiquitination of SLC3A2. Cell Mol Life Sci. 2024;81:325. Creative Commons.30 Copyright © 2024 Springer. |
Organ dysfunction and recovery
Organ dysfunction in sepsis is a direct consequence of the systemic inflammatory response and the dysregulated immune activity triggered by the infection.31 This multifaceted syndrome impacts several vital organs, causing conditions such as acute kidney injury, acute respiratory distress syndrome (ARDS), hepatic dysfunction, and cardiovascular instability. Developing therapeutic strategies aimed at enhancing recovery and reducing sepsis-related mortality will be greatly enhanced by understanding the underlying mechanisms responsible for organ dysfunction.
The pathophysiology of organ dysfunction in sepsis involves several interconnected mechanisms.32 Firstly, the overwhelming inflammatory response can lead to widespread endothelial damage, which disrupts the microvascular blood flow. This disruption impairs oxygen and nutrient delivery and waste removal, which are critical for cellular function. Secondly, the excessive production of cytokines during sepsis can have direct cytotoxic effects and can initiate apoptosis in various organ cells.33 Additionally, the activation of the coagulation cascade can lead to microthrombi formation, further impairing blood flow and exacerbating organ damage.34
The recovery from organ dysfunction in sepsis involves the stabilization of hemodynamics, the restoration of impaired organ function, and the mitigation of ongoing inflammation. Clinical management focuses on supporting the affected organs through interventions such as renal replacement therapy for kidney failure, mechanical ventilation for ARDS, and pharmacological support for cardiovascular and hepatic functions.35 Emerging treatments that aim to modulate the immune response or enhance cellular repair processes are also being explored.
Key Factors Influencing the Development of Sepsis
Microbial Factors
The virulence of a pathogen in sepsis is largely determined by its ability to evade the host’s immune defenses and establish an infection.36 Many bacteria possess specialized virulence factors that enhance their pathogenicity. For example, LPS, a major component of the outer membrane of Gram-negative bacteria, is recognized as one of the most potent inducers of the inflammatory response in sepsis.37 In the event that it is not promptly controlled, it can trigger a wide array of immune reactions that will result in tissue damage and organ failure.
In addition to LPS, bacterial toxins are of particular significance in the progression of sepsis, as they can cause direct cellular injury and modulate immune responses.38 Toxins such as exotoxins and enterotoxins disrupt cell membranes and interfere with cellular signaling pathways, leading to cell death and tissue necrosis.39 These actions not only facilitate the spread of the infection but also trigger severe systemic inflammation that is characteristic of sepsis.
Recent studies have also highlighted the role of genetic and metabolic adaptations of pathogens in influencing their virulence during sepsis.40 Pathogens that can rapidly alter their gene expression in response to the host environment are more likely to evade immune responses and sustain infections that lead to sepsis. Moreover, the ability of pathogens to metabolize host-derived nutrients and resist antimicrobial agents significantly affects their survival and virulence.
Host Response
A hallmark of sepsis is the dysregulation of the immune response, which is characterized by an initial hyperinflammatory state followed by a period of immunosuppression.41 During the hyperinflammatory phase, cytokine storm can lead to widespread tissue damage. This is followed by a compensatory anti-inflammatory response syndrome (CARS), during which the immune system’s responsiveness is markedly diminished, increasing vulnerability to secondary infections and affecting recovery.42
The genetic makeup of an individual significantly influences the variability in immune response to infections leading to sepsis.43 Certain genetic polymorphisms affect cytokine production, immune cell activation, and even the effectiveness of microbial clearance.44 These genetic factors can explain why some individuals develop severe sepsis and organ failure while others with similar infections do not.
Comorbidities such as diabetes, chronic kidney disease, or chronic respiratory conditions can impair the immune system’s effectiveness and alter the host’s response to infection.45 These conditions often exacerbate the severity of sepsis by compromising the body’s ability to mount an adequate immune response. Furthermore, they can influence the pharmacokinetics and pharmacodynamics of drugs employed in the management of sepsis, thereby complicating treatment approaches.
Environmental Factors
It is evident that environmental factors play a significant role in the progression and management of sepsis, particularly within healthcare settings. The incidence of hospital-acquired infections (HAIs) and the prevalence of antimicrobial resistance are two critical aspects that can dramatically affect sepsis outcomes.46,47
Hospitals, especially intensive care units (ICUs), are high-risk environments for the acquisition of infections due to the concentration of vulnerable patients with various invasive devices like catheters and ventilators.48 These devices provide a route for pathogens to enter the body and cause infections that may lead to sepsis. The pathogens responsible for HAIs often include multidrug-resistant organisms, which complicate treatment options and increase mortality rates. For instance, methicillin-resistant Staphylococcus aureus (MRSA) and multidrug-resistant Gram-negative bacteria are common culprits in these environments.49
The extensive use of antibiotics in hospitals has led to an increase in antimicrobial resistance, which is a significant factor in the complexity and severity of sepsis cases.50 A resistant strain of bacteria poses a greater challenge to treat and can spread rapidly within a hospital setting, complicating sepsis management.51 Effective antibiotic stewardship and infection control practices are essential to mitigate this issue, emphasizing the need for judicious use of antibiotics and adherence to strict hygiene protocols.
The Establishment and Application of Zebrafish Sepsis Model
Clinical Relevance of Zebrafish Sepsis Model: Validation of Pathological Similarity to Human Sepsis
The zebrafish model is considered a powerful tool for studying human sepsis due to its genetic and immune response similarities.52 To validate its clinical relevance, we focused on examining the pathological states in zebrafish that closely mimic those observed in human sepsis. This section highlights key histological and molecular biology features to confirm the model’s validity.
Histological Validation
Acute kidney injury (AKI) in zebrafish is characterized by tubular necrosis and hepatocellular degeneration, which are similar to the acute organ damage seen in human sepsis.53 Hematoxylin-Eosin (H&E) staining and immunohistochemistry were employed to highlight critical pathological features such as cellular integrity loss, immune cell infiltration, and tissue hemorrhage, aligning with human sepsis characteristics.54,55
Molecular Biology Features
To further validate the zebrafish sepsis model, we analyzed the molecular responses using quantitative PCR (qPCR),56 Western blotting,57 and enzyme-linked immunosorbent assays (ELISA). These techniques allowed us to measure the expression levels of key cytokines and signaling molecules involved in the inflammatory response,58 such as TNF-α, IL-1β, NF-κB, and MAPK pathways. The patterns observed were compared with human data to assess the translational potential of the zebrafish model.
The Establishment and Application of Zebrafish Sepsis Model
Establishment of Infection Experiment
Microbial infection plays a pivotal role in initiating inflammation in patients with sepsis, marking the onset of immune response and the release of inflammatory mediators.59 As sepsis progresses, the presence of pathogens and their toxins can significantly amplify inflammatory responses, potentially leading to systemic inflammation, organ damage, and multiple organ failure.21
(1) Escherichia coli (E. coli)
E. coli is a versatile species with a wide range of ecological niches and interactions with hosts, including mutualistic and pathogenic relationships.60 Specifically, septicemia-associated E. coli (SEPEC) is strongly linked to sepsis.61 Uropathogenic E. coli (UPEC), commonly used in sepsis model infections, exemplifies the pathogenic potential to induce systemic infections that can escalate to sepsis.62
Barber et al63 successfully employed zebrafish embryo infection models to simulate and study these interactions. The method involved injecting zebrafish embryos with ExPEC strains, resulting in significant inflammatory responses, including cytokine storm and symptoms such as rapid heart rate, endothelial leakage, and edema.
UPEC, a major pathogen causing urinary tract infections, can ascend from the ureter to the kidney, leading to acute pyelonephritis and potentially progressing to sepsis.64 Tran et al65 injected UPEC into zebrafish’s abdominal cavity to simulate human infection. The results showed that the systemic infection model of zebrafish inoculated with UPEC was successfully established, with decreased survival rates and increased UPEC load after infection.
(2) Streptococcus pneumoniae (S. pneumoniae)
S. pneumoniae causes a range of respiratory infections, including otitis media, sinusitis, pneumonia, and sepsis.66 In sepsis caused by S. pneumoniae, the bacteria colonize the respiratory tract and release toxins and cell wall components that initiate an inflammatory response. This triggers the release of inflammatory mediators and activates the immune system, leading to systemic inflammatory response syndrome (SIRS).
Rounioja S. et al67 demonstrated this by injecting the pathogenic pneumococcal strain TIGR4 (T4) into zebrafish embryos. Signs of severe infection, such as lack of movement and reduced heart rate, were observed at 100 CFU/injection. Post-infection, the expression of TNF-α and IL-1β was highly induced, and systemic immune reactions occurred.
(3) Streptococcus agalactiae (Group B Streptococcus, GBS)
GBS is a gram-positive capsular bacterium and a major cause of neonatal pneumonia, septicemia and meningitis.68,69 Studies have found that injection of highly virulence clinical isolate COH1 of GBS into zebrafish led to up-regulation of pro-inflammatory cytokines and chemokines, increased mortality, and destruction of blood vessels compared with control group.70
(4) Pseudomonas aeruginosa (P. aeruginosa)
P. aeruginosa is a ubiquitous gram-negative bacterium that can infect various plants and animals. It is an important opportunistic pathogen in humans, often causing severe local or systemic infections.71 P. aeruginosa is mainly used in the study of corneal infection, burn wound infection, acute pneumonia,72 and sepsis in mouse models.73
When two laboratory strains of P. aeruginosa (PAO1 and PAK) expressing green fluorescent protein (GFP) were microinjected into zebrafish embryo tail veins, immediate bacterial accumulation near the injection site was observed. These aggregates contained red blood cells and infected phagocytes, which blocked blood vessels completely. The immune response of the zebrafish was activated, releasing a large number of neutrophils and phagocytes.74
(5) Staphylococcus aureus (S. aureus)
S. aureus is a common bacterium found on the skin and mucosal surfaces of the human body, capable of causing various infections. S. aureus infection can lead to local epidermal infections such as impetigo, boils, and cellulitis, as well as more serious infections like sepsis, endocarditis, and pneumonia.75
In zebrafish larvae infection with a 2.5×104 CFUs strain of S. aureus fluorescein (Cowan I pCN56_GFPmut2) 48 hours post-fertilization showed systemic inflammatory infection (increased serum concentration of TNF-α) within 3.5 hours of infection, observed using fluorescence microscopy.76 The infection ultimately led to the death of the zebrafish.
(6) Klebsiella pneumoniae (cKP)
cKP is a non-motility gram-negative bacterium with cysts that are commonly found in the environment. It is often associated with alcohol use disorders and community-acquired pneumonia in diabetics. As a result of multidrug-resistant cKP (MDR), pneumonia, urinary tract infections, and bloodstream infections are now common hospital-acquired infections.77
Galvez-Silva et al collected juvenile fish two days after fertilization (dpf) and injected wild-type highly virulent Klebsiella pneumoniae SGH10 strain and its ΔwcaJ mutant derivative into juvenile fish through the ear sac. By resulting in fluorescence microscopy results, the cKP group showed more aggregation of neutral particles.78
Chemical Infection
Chemicals agents, such as LPS, are employed in zebrafish models to simulate bacterial infections and elicit immune and inflammatory responses that are analogous to those observed in sepsis. LPS, which is found in the outer membrane of Gram-negative bacteria, including E. coli and P aeruginosa, binds to toll-like receptor 4 (TLR4) on innate immune cells, initiating a cascade of immune reactions.79
Mechanism of LPS-induced sepsis in zebrafish: Upon LPS exposure, zebrafish activate their immune system similarly to mammals, releasing a suite of cytokines, chemokines, and interferons.80 This response leads to an inflammatory imbalance, precipitating systemic inflammation—a hallmark of sepsis. In experimental setups, zebrafish larvae at three days post-fertilization are exposed to varying concentrations of LPS to observe dose-dependent effects on immune activation and physiological stress responses.81
The larvae are typically immersed in a solution containing different concentrations of LPS (ranging from 25 to 200 µg/mL) for periods spanning 2 to 24 hours to induce inflammatory responses. Phenotypic assessments are conducted to evaluate inflammation-related symptoms such as fin edema, decreased microvascular circulation, and overall morbidity and mortality rates. Notable findings include increased production of reactive oxygen species (ROS) and inflammatory mediators, as well as observed behavioral changes such as reduced movement and increased mortality rates.
Following LPS exposure, there is a notable increase in pro-inflammatory cytokines, including TNF-α and IL-1β, along with elevated HMGB1 expression, indicating a robust immune response.82 The zebrafish model also exhibits a reduction in neutrophil counts and disrupted brain capillary circulation, which closely resembles the clinical symptoms of sepsis.83 This provides a comprehensive system to study the progression and mitigation of systemic inflammation.
Activation of downstream signaling molecules, such as NFκB and MyD88, and the resulting release of pro-inflammatory cytokines are conserved reactions between teleosts and mammals.84 LPS induces the downstream release of a series of pro-inflammatory factors by up-regulating the expression of NF-κB and then MyD88, and generally increases the vascular permeability accompanied by a decrease in the expression of tight link protein closure proteins A and B, claudin 5a and b, and their intracellular companion ZO-1. By affecting these proteins, LPS leads to the loss of vascular integrity and the classic septic pathological state of exudative edema.82
Application of Gene Editing Technology
With the advent of gene editing, particularly the use of CRISPR/Cas9 technology, models of sepsis in zebrafish have been revolutionized. The technology enables precise genetic modifications at the genetic level, allowing DNA sequences to be knocked out, replaced, or inserted.85,86 These modifications are crucial for studying the roles of specific genes in immune response and inflammation during sepsis.
CRISPR/Cas9 functions by creating specific incisions at predetermined locations within the genome, thereby enabling researchers to investigate the consequences of disrupting or modifying genes associated with the immune system, inflammatory regulation, and antibacterial defense.87 For instance, the elimination of genes linked to the immune response can diminish the organism’s capacity to resist infections, increasing its vulnerability to sepsis. This is accomplished by compromising the host’s capacity to mount an efficacious defense against invading pathogens, thereby facilitating the progression to systemic inflammation and sepsis.
In zebrafish models, CRISPR/Cas9 has been employed to ablate genes that are essential for immune system functionality, including those encoding cytokines and their receptors. The ablation of these genes can result in heightened susceptibility to infection and an elevated propensity for the development of sepsis. For instance, zebrafish deficient in the crp genes, which are involved in the immune response, exhibit increased vulnerability to pneumococcal infections. In infectious disease studies, zebrafish are infected with pathogens (such as Edwardsiella piscicida) by removing genes involved in the immune response (such as TNF-α),56 and CRISPR/Cas9 has been used to study the pathways of anti-inflammatory agents (primarily glucocorticoids) and to explore the mechanisms of pathogen infection and immune response.88 This vulnerability is further compounded when additional immune-related genes are knocked out, mirroring the complex interactions that contribute to the pathophysiology of sepsis in humans.89
The above description shows a variety of methods for modeling sepsis, including infecting model animals with different pathogens and chemicals, and using gene editing technology to study the role of specific genes in sepsis, and gives solutions to the challenges encountered by different substances (Table 1). This will help us better understand the pathophysiological mechanisms of sepsis and provide an important tool for developing new treatment options for clinical sepsis.
 |
Table 1 Methods and Challenges in Establishing Zebrafish Sepsis Models |
Treatment Application Based on Zebrafish Sepsis Model
Due to their high-throughput screening capabilities, transparency, and similarity to human immune systems, zebrafish models have become important tools for studying sepsis and anti-inflammatory drugs.98 The introduction of pathogenic bacteria or inflammatory inducers into zebrafish can be used to evaluate drug candidates’ anti-inflammatory and antisepsis properties, thereby speeding up drug development.99 The transparency of zebrafish larvae allows real-time observation of the effects of drugs on inflammation and sepsis, especially the dynamic changes in immune cells, Primarily by studying the mechanism of resolution, by characterizing real-time neutrophil migration patterns. The high reproduction rate and low breeding cost make it suitable for large-scale high-throughput screening experiments.100 To evaluate its effect on inflammation markers, a candidate drug may be dissolved in water or administered directly as part of the screening process, followed by microscopic observation and biochemical analysis. In order to develop drug screening strategies for anti-inflammatory models of zebrafish sepsis, additional research is necessary to understand the mediators and receptors that turn inflammation into resolution in zebrafish. A further advantage of developing selective antibodies will be to characterize inflammatory responses to zebrafish at the protein level in depth. By using genetic tools, such as TALENS and CRISPR-Cas9, it is possible to generate new mutant strains of zebrafish to assess the likelihood of biological effects, such as anti-TNF strategies, on zebrafish.101 Accordingly, a comprehensive utilization of zebrafish model advantages can contribute to the development of effective anti-sepsis and anti-inflammatory drugs, as well as enhance patients’ therapeutic outcome and prognosis.
Novel Antibiotics and Anti-Inflammatory Agents
Peptidoglycan hydrolase (PGH) targets specific bonds within the bacterial peptidoglycan (PG) network and has demonstrated efficacy in disrupting biofilms. Due to their high lytic activity, PGHs are considered effective antistaphylococcal agents.102 MEndoB, a variant selected from a chimeric PGH library, has shown an exceptional ability to eliminate staphylococcus in human serum. In zebrafish models, treatment with MEndoB significantly reduced infection rates and yielded improvements in survival and inflammation comparable to those observed with the positive control, vancomycin.
Synthetic Molecules and Immune Modulation
Poly(I:C) is a synthetic double-stranded RNA (dsRNA) molecule commonly used as a viral genomic RNA mimic.103 It is primarily employed in immunology research to activate the immune system response. Poly(I:C) activates specific immune cell and molecular pathways, such as pattern recognition receptors like TLR3, MDA5, and RIG-I, by simulating viral infection. This activation triggers an antiviral response, including the production of interferons.104,105 In a study, poly(I:C) was administered as an adjuvant in combination with the inactivated viral hemorrhagic septicemia virus (VHSV) (CSpIC+iV vaccine) to adult zebrafish. The CSpIC+iV formulation provided significant protection against VHSV-induced mortality106.
Natural Compounds and Their Roles
Flavonoids possess various biological activities, including anti-inflammatory, antioxidant, antibacterial, and anti-allergic effects107. Urushetin, a naturally occurring flavonoid, has been utilized to mitigate LPS-induced inflammation and endotoxic shock in zebrafish models. Studies have demonstrated that the urushetin treatment group exhibits reduced mortality, abnormalities, and heart rate reduction in zebrafish larvae injected with LPS. Additionally, urushetin inhibits the expression of LPS-induced pro-inflammatory gene and decreases the recruitment of macrophages and neutrophils to the site of inflammation in zebrafish larvae108.
Glucosamine and Its Anti-Inflammatory Effects
Glucosamine (GlcN) is an amino derivative of glucose, known for its anti-inflammatory and antioxidant roles, particularly in cartilage and joints109. In zebrafish models, GlcN preconditioning has been shown to inhibit the mRNA expression of LPS-induced inflammatory genes in visceral tissues. It also improves the levels of O-GlcNAcylation and decreases OGA expression in these tissues. Furthermore, GlcN treatment alleviates cell infiltration and pulmonary edema associated with pneumonia in zebrafish110.
Combating Antibiotic Resistance
Antibiotics are commonly used to treat sepsis, but the emergence of drug-resistant bacteria in patients with sepsis has significantly reduced their efficacy, posing substantial risks to clinical treatment111. Recent studies have explored the combination of the antibiotic ceftriaxone (CTX) and Cpl-711, a phage-derived endolysin, to test their synergistic effect on multidrug-resistant pneumococcal strains. Treatment results in zebrafish models demonstrated higher survival rates and reduced bacterial loads compared to monotherapy with either CTX or Cpl-711 alone112.
The primary function of mainstream anti-sepsis drugs, such as antibiotics, inactivated viruses, and antioxidants, is to boost the immune system, release anti-inflammatory factors, reduce systemic inflammation, and slow the infection process (Table 2). Due to the high optical transparency of zebrafish and the availability of various transgenic lines, the pathological processes of sepsis can be observed in greater detail. This makes zebrafish a valuable model for future drug screening and the development of new therapeutic strategies.
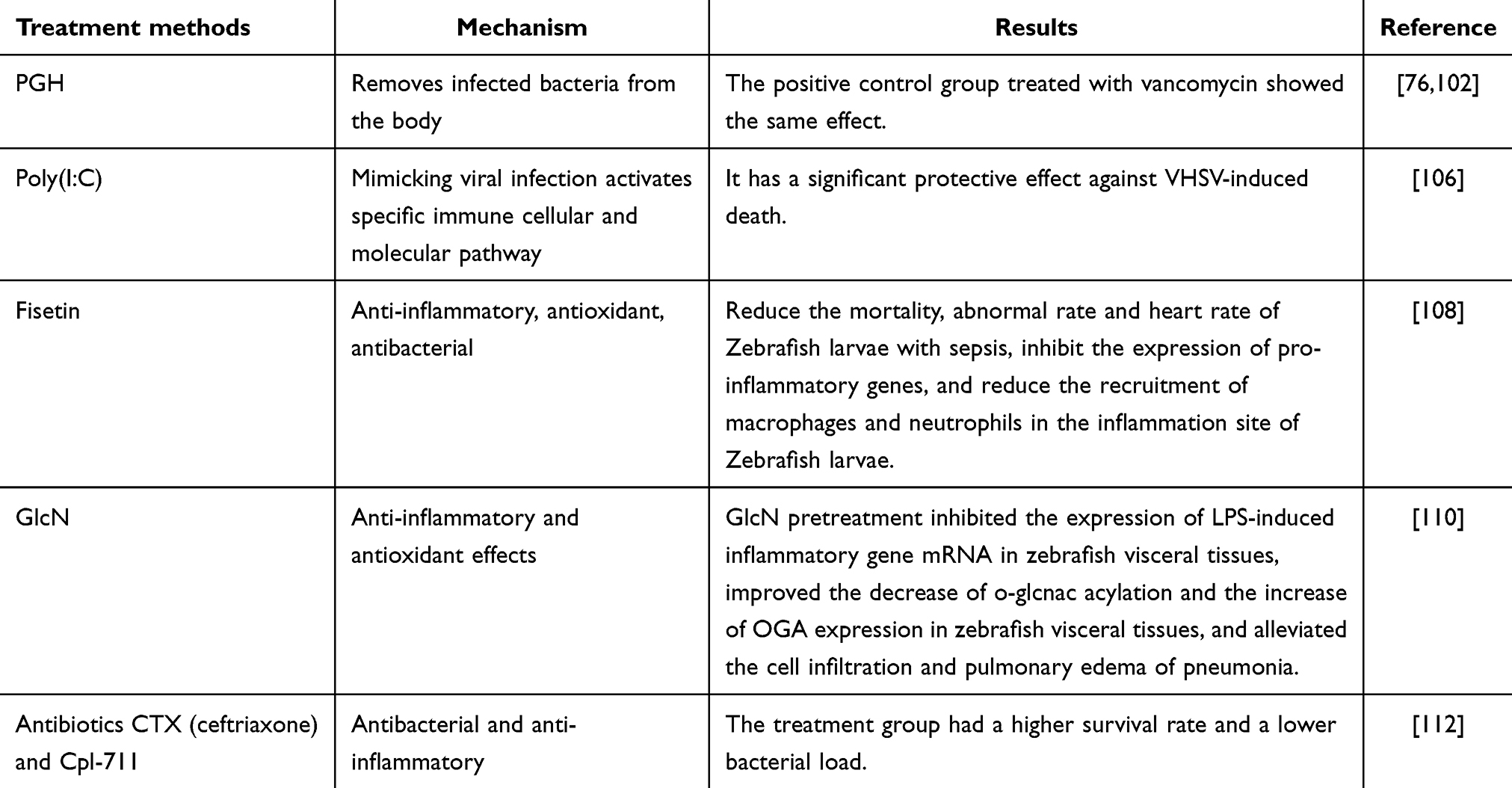 |
Table 2 Treatment of Zebrafish Sepsis |
Challenges and Perspectives of the Zebrafish Model in Sepsis Research
As the utilisation of zebrafish in sepsis research continues to expand, it is crucial to recognise and address the inherent challenges, while exploring the potential advancements, in order to enhance the translational value of this model. These challenges include ensuring precise control of infection dose, managing the variability in immune response caused by genetic diversity among strains of zebrafish, and accurately replicating the complex interactions between the various immune cells observed in human sepsis patients113.
To address these issues, researchers are developing more refined techniques for infection delivery and monitoring in zebrafish. These include microinjection and real-time imaging, which allow for more controlled and consistent experimental conditions89,114. Further advances in genetic engineering, such as conditional regulation of CRISPR/Cas9, enable the creation of genetically modified zebrafish that more closely mimic human immune responses or genetically predisposed conditions causing sepsis115.
Furthermore, the zebrafish model has the potential to significantly advance sepsis research, particularly through the integration of cutting-edge technologies: (1) Gene editing technologies allow researchers to manipulate the zebrafish genome to introduce or disrupt genes known to be involved in the immune response to sepsis82. This can lead to a better understanding of the genetic basis of sepsis and the identification of new therapeutic targets. (2) Zebrafish models can be employed in personalized medicine by testing patient-specific bacterial strains or therapies within a controlled environment116. This approach can facilitate the tailoring of treatments based on individual genetic and immunological profiles. (3) The high-throughput screening capabilities of zebrafish models render them optimal for testing combination therapies98. Multiple pathways involved in sepsis can be simultaneously targeted by these therapies, potentially resulting in more effective treatments with fewer side effects (Figure 3).
 |
Figure 3 The zebrafish model has the potential to significantly advance the investigation of sepsis, especially by integrating cutting-edge technologies: (A) Gene editing technology allows researchers to manipulate the zebrafish genome to introduce or disrupt bases known to be involved in the immune response to sepsis. (B) The zebrafish model can be used for personalized medicine to accelerate the transformation of the basis to the clinic. (C) High throughput screening this as well as transparency properties, sepsis development, and mechanisms are further in-depth. |
Conclusion
The utilization of zebrafish in sepsis research has substantiated its compatibility with human pathological processes, particularly in elucidating the mechanisms of immune dysregulation and organ dysfunction. This model has effectively bridged translational gaps by highlighting significant similarities in immune responses and tissue pathology between zebrafish and humans, supported by rigorous histological and molecular validations.
Advancements in conditional gene editing within the zebrafish model are enhancing our genetic understanding of sepsis, leading to potential gene-specific therapies. Moreover, the model’s suitability for high-throughput drug screening is instrumental in accelerating the development of combination therapies. These technological and methodological enhancements in zebrafish research are poised to revolutionize sepsis treatment strategies, moving towards more personalized and efficacious approaches. Overall, the zebrafish model continues to be a pivotal tool in sepsis research, promising novel insights and therapeutic breakthroughs that are expected to significantly impact clinical outcomes in sepsis management.
Abbreviation
PRRs, pattern recognition receptors; PAMPs, Pathogen-associated molecular patterns; TLRs, Toll-like receptors; IL-1β, Interleukin—1β; TNF-α, Tumor necrosis factor-alpha; IL-6, Interleukin-6; ARDS, Acute respiratory distress syndrome; LPS, Lipopolysaccharides; CARS, Compensatory anti-inflammatory response syndrome; HAIs, Hospital-acquired infections; ICU, Intensive care units; MRSA: Methicillin-resistant Staphylococcus aureus; Qpcr, Quantitative PCR; ELISA, Enzyme-linked immunosorbent assays; AKI, Acute kidney injury; E. coli, Escherichia coli; S. pneumoniae, Streptococcus pneumoniae; S, Agalactiae: Streptococcus agalactiae; SIRS, Systemic inflammatory response syndrome; P. Aeruginosa: Pseudomonas aeruginosa; cKPK, Klebsiella pneumoniae; dpf, days after fertilization; TLR4, Toll-like receptor 4; PGH, Peptidoglycan hydrolase; PG, Peptidoglycan; dsRNA, Double-stranded RNA; GlcN, Glucosamine; CTX, Ceftriaxone.
Compliance with Ethics Requirement
This article does not address ethical requirements.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Natural Science Foundation of Zhejiang Province (LGF21H150001), and the Graduate Scientific Research Foundation of Wenzhou University (3162024003056 and 3162024004102).
Disclosure
The authors have declared no conflicts of interest in this work.
References
1. Rudd KE, Johnson SC, Agesa KM, et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: analysis for the Global Burden of Disease Study. Lancet. 2020;395(10219):200–211. doi:10.1016/S0140-6736(19)32989-7
2. Fleischmann-Struzek C, Goldfarb DM, Schlattmann P, Schlapbach LJ, Reinhart K, Kissoon N. The Global Burden of Paediatric and Neonatal Sepsis: a Systematic Review. Lancet Respir Med. 2018;6:223–230. doi:10.1016/S2213-2600(18)30063-8
3. Nandi M, Jackson SK, Macrae D, Shankar-Hari M, Tremoleda JL, Lilley E. Rethinking Animal Models of Sepsis - Working towards Improved Clinical Translation Whilst Integrating the 3Rs. Clin Sci. 2020;134:1715–1734. doi:10.1042/CS20200679
4. Cai L, Rodgers E, Schoenmann N, Raju RP. Advances in Rodent Experimental Models of Sepsis. Int J Mol Sci. 2023;24(9578). doi:10.3390/ijms24119578
5. Stortz JA, Raymond SL, Mira JC, Moldawer LL, Mohr AM, Efron PA. Murine Models of Sepsis and Trauma: can We Bridge the Gap? ILAR J. 2017;58:90–105. doi:10.1093/ilar/ilx007
6. Guillon A, Preau S, Aboab J, et al. Preclinical Septic Shock Research: why We Need an Animal ICU. Ann Intensive Care. 2019;9:66. doi:10.1186/s13613-019-0543-6
7. Yang -L-L, Wang G-Q, Yang L-M, Huang Z-B, Zhang W-Q, Yu L-Z. Endotoxin Molecule Lipopolysaccharide-Induced Zebrafish Inflammation Model: a Novel Screening Method for Anti-Inflammatory Drugs. Molecules. 2014;19:2390–2409. doi:10.3390/molecules19022390
8. Qu J, Yan M, Fang Y, et al. Zebrafish in Dermatology: a Comprehensive Review of Their Role in Investigating Abnormal Skin Pigmentation Mechanisms. Front Physiol. 2023;14:1296046. doi:10.3389/fphys.2023.1296046
9. Boopathi S, Haridevamuthu B, Mendonca E, et al. Combined Effects of a High-Fat Diet and Polyethylene Microplastic Exposure Induce Impaired Lipid Metabolism and Locomotor Behavior in Larvae and Adult Zebrafish. Scie Total Enviro. 2023;902:165988. doi:10.1016/j.scitotenv.2023.165988
10. Wang L, Ma J, Wu W, et al. Effect of Aerobic Exercise as A Treatment on Type 2 Diabetes Mellitus with Depression-like Behavior Zebrafish. Life Sci. 2022;300:120578. doi:10.1016/j.lfs.2022.120578
11. Guru A, Manjunathan T, Sudhakaran G, Juliet A, Gopinath P, Arockiaraj J. 6-Gingerdione Reduces Apoptotic Conditions in HepG2 Cells and Inhibits Inflammatory Cytokine Gene Expression in Alcoholic Liver Injured Zebrafish Larvae. Chem. Biodivers. 2023;20:e202200959. doi:10.1002/cbdv.202200959
12. Lubin A, Otterstrom J, Hoade Y, et al. Automated and High-Throughput Drug Screening Platform for Zebrafish Embryos. Biol Open. 2021;10:bio058513. doi:10.1242/bio.058513
13. Leiba J, Özbilgiç R, Hernández L, et al. Molecular Actors of Inflammation and Their Signaling Pathways: mechanistic Insights from Zebrafish. Biology. 2023;12:153. doi:10.3390/biology12020153
14. van der Poll T, Shankar-Hari M, Wiersinga WJ. The Immunology of Sepsis. Immunity. 2021;54:2450–2464. doi:10.1016/j.immuni.2021.10.012
15. Wiersinga WJ, Leopold SJ, Cranendonk DR, van der Poll T. Host Innate Immune Responses to Sepsis. Virulence. 2014;5:36–44. doi:10.4161/viru.25436
16. Mahapatra S, Heffner AC. Septic Shock. StatPearls; StatPearls Publishing: Treasure Island (FL). 2024.
17. Chan L, Karimi N, Morovati S, et al. The Roles of Neutrophils in Cytokine Storms. Viruses. 2021;13:2318. doi:10.3390/v13112318
18. Chousterman BG, Swirski FK, Weber GF. Cytokine Storm and Sepsis Disease Pathogenesis. Semin Immunopathol. 2017;39:517–528. doi:10.1007/s00281-017-0639-8
19. Gierlikowska B, Stachura A, Gierlikowski W, Demkow U. The Impact of Cytokines on Neutrophils’ Phagocytosis and NET Formation during Sepsis-A Review. Int J Mol Sci. 2022;23(5076). doi:10.3390/ijms23095076
20. T I, M L, Jh L. Sepsis-Induced Coagulopathy and Disseminated Intravascular Coagulation. Semin Thromb Hemost. 2020;46. doi:10.1055/s-0039-1694995
21. Cao M, Wang G, Xie J. Immune Dysregulation in Sepsis: experiences, Lessons and Perspectives. Cell Death Discov. 2023;9:1–11. doi:10.1038/s41420-023-01766-7
22. Malakooti M, Kelleher M, Wald E. Immune Dysregulation in Sepsis. Clin Pediatr Emergency Med. 2014;15:163–168. doi:10.1016/j.cpem.2014.04.002
23. Tu X, Huang H, Xu S, Li C, Luo S. Single-Cell Transcriptomics Reveals Immune Infiltrate in Sepsis. Front Pharmacol. 2023;14:1133145. doi:10.3389/fphar.2023.1133145
24. Zhou M, Aziz M, Yen H-T, Ma G, Murao A, Wang P. Extracellular CIRP Dysregulates Macrophage Bacterial Phagocytosis in Sepsis. Cell Mol Immunol. 2023;20:80–93. doi:10.1038/s41423-022-00961-3
25. Venet F, Monneret G. Advances in the Understanding and Treatment of Sepsis-Induced Immunosuppression. Nat Rev Nephrol. 2018;14:121–137. doi:10.1038/nrneph.2017.165
26. Guo J, Kong Z, Yang S, et al. Therapeutic Effects of Orexin-A in Sepsis-Associated Encephalopathy in Mice. J Neuroinflammation. 2024;21:131. doi:10.1186/s12974-024-03111-w
27. Ritter K, Rissel R, Renz M, Ziebart A, Schäfer MKE, Kamuf J. Nebulized Lipopolysaccharide Causes Delayed Cortical Neuroinflammation in a Murine Model of Acute Lung Injury. Int J Mol Sci. 2024;25(10117). doi:10.3390/ijms251810117
28. Yan C, Kuang W, Jin L, et al. Carvacrol Protects Mice against LPS-Induced Sepsis and Attenuates Inflammatory Response in Macrophages by Modulating the ERK1/2 Pathway. Sci Rep. 2023;13:12809. doi:10.1038/s41598-023-39665-7
29. Wang S, Jiang D, Huang F, et al. Therapeutic Effect of Echinococcus Granulosus Cyst Fluid on Bacterial Sepsis in Mice. Parasit Vectors. 2023;16:450. doi:10.1186/s13071-023-06021-7
30. Chen Z, Zhang J, Gao S, et al. Suppression of Skp2 Contributes to Sepsis-Induced Acute Lung Injury by Enhancing Ferroptosis through the Ubiquitination of SLC3A2. Cell Mol Life Sci. 2024;81:325. doi:10.1007/s00018-024-05348-3
31. Fujishima S. Organ Dysfunction as a New Standard for Defining Sepsis. Inflamm Regen. 2016;36:24. doi:10.1186/s41232-016-0029-y
32. Pool R, Gomez H, Kellum JA. Mechanisms of Organ Dysfunction in Sepsis. Crit Care Clin. 2018;34:63–80. doi:10.1016/j.ccc.2017.08.003
33. Wen X, Xie B, Yuan S, Zhang J. The “Self-Sacrifice” of ImmuneCells in Sepsis. Front Immunol. 2022;13:833479. doi:10.3389/fimmu.2022.833479
34. Maneta E, Aivalioti E, Tual-Chalot S, et al. Endothelial Dysfunction and Immunothrombosis in Sepsis. Front Immunol. 2023;14:1144229. doi:10.3389/fimmu.2023.1144229
35. Lelubre C, Vincent J-L. Mechanisms and Treatment of Organ Failure in Sepsis. Nat Rev Nephrol. 2018;14:417–427. doi:10.1038/s41581-018-0005-7
36. Ramachandran G. Gram-Positive and Gram-Negative Bacterial Toxins in Sepsis: a Brief Review. Virulence. 2014;5:213–218. doi:10.4161/viru.27024
37. Sondhi P, Adeniji T, Lingden D, Stine KJ. Advances in Endotoxin Analysis. Adv Clin Chem. 2024;118:1–34. doi:10.1016/bs.acc.2023.11.001
38. Joffre J, Hellman J, Ince C, Ait-Oufella H. Endothelial Responses in Sepsis. Am J Respir Crit Care Med. 2020;202:361–370. doi:10.1164/rccm.201910-1911TR
39. Qu L, Wang L, Ji H, et al. Toxic Mechanism and Biological Detoxification of Fumonisins. Toxins. 2022;14(182). doi:10.3390/toxins14030182
40. Chen H, Zhang X, Su H, et al. Immune Dysregulation and RNA N6-Methyladenosine Modification in Sepsis. Wiley Interdiscip Rev RNA. 2023;14:e1764. doi:10.1002/wrna.1764
41. Bauer M, Coldewey SM, Leitner M, Löffler B, Weis S, Wetzker R. Deterioration of Organ Function As a Hallmark in Sepsis: the Cellular Perspective. Front Immunol. 2018;9:1460. doi:10.3389/fimmu.2018.01460
42. Buerfent BC, Gondorf F, Wohlleber D, Schumak B, Hoerauf A, Hübner MP. Escherichia Coli-Induced Immune Paralysis Is Not Exacerbated during Chronic Filarial Infection. Immunology. 2015;145:150–160. doi:10.1111/imm.12435
43. Wang W, Liu C-F. Sepsis Heterogeneity. World J Pediatr. 2023;19:919–927. doi:10.1007/s12519-023-00689-8
44. Davenport EE, Burnham KL, Radhakrishnan J, et al. Genomic Landscape of the Individual Host Response and Outcomes in Sepsis: a Prospective Cohort Study. Lancet Respir Med. 2016;4:259–271. doi:10.1016/S2213-2600(16)00046-1
45. Yende S, Kellum JA, Talisa VB, et al. Long-Term Host Immune Response Trajectories Among Hospitalized Patients With Sepsis. JAMA Network Open. 2019;2:e198686. doi:10.1001/jamanetworkopen.2019.8686
46. Beggs C, Knibbs LD, Johnson GR, Morawska L. Environmental Contamination and Hospital-Acquired Infection: factors That Are Easily Overlooked. Indoor Air. 2015;25:462–474. doi:10.1111/ina.12170
47. Ter Horst R, Jaeger M, Smeekens SP, et al. Host and Environmental Factors Influencing Individual Human Cytokine Responses. Cell. 2016;167:1111–1124.e13. doi:10.1016/j.cell.2016.10.018
48. Djuric O, Markovic-Denic L, Jovanovic B, Bumbasirevic V. High Incidence of Multiresistant Bacterial Isolates from Bloodstream Infections in Trauma Emergency Department and Intensive Care Unit in Serbia. Acta Microbiol Immunol Hung. 2019;66:307–325. doi:10.1556/030.66.2019.007
49. Kim H, Kim ES, Lee SC, et al. Decreased Incidence of Methicillin-Resistant Staphylococcus aureus bacteremia in Intensive Care Units: a 10-Year Clinical, Microbiological, and Genotypic Analysis in a Tertiary Hospital. Antimicrob Agents Chemother. 2020;64:e01082–20. doi:10.1128/AAC.01082-20
50. Barrasa-Villar JI, Aibar-Remón C, Prieto-Andrés P, Mareca-Doñate R, Moliner-Lahoz J. Impact on Morbidity, Mortality, and Length of Stay of Hospital-Acquired Infections by Resistant Microorganisms. Clin Infect Dis. 2017;65:644–652. doi:10.1093/cid/cix411
51. Ohnuma T, Chihara S, Costin B, et al. Resistance Profiles, and Outcomes of Bloodstream Infections in Community-Onset Sepsis in the United States. Crit Care Med. 2023;51:1148–1158. doi:10.1097/CCM.0000000000005870
52. Guru A, Arockiaraj J. Exposure to Environmental Pollutant Bisphenol A Causes Oxidative Damage and Lipid Accumulation in Zebrafish Larvae: protective Role of WL15 Peptide Derived from Cysteine and Glycine-Rich Protein 2. J Biochem Moler Toxico. 2023;37:e23223. doi:10.1002/jbt.23223
53. Wen X, Cui L, Morrisroe S, et al. A Zebrafish Model of Infection-Associated Acute Kidney Injury. Am J Physiol Renal Physiol. 2018;315. doi:10.1152/ajprenal.00328.2017
54. Shi Q, Zhang X, Liu X, Yan C, Lu S. Visualization of PFOA Accumulation and Its Effects on Phospholipid in Zebrafish Liver by MALDI Imaging. Anal Bioanal Chem. 2024;416:2493–2501. doi:10.1007/s00216-024-05214-y
55. Huang L, Jia K, Xiong H, et al. Oxyfluorfen Exposure Can Cause Acute Kidney Injury by Promoting ROS-Induced Oxidative Stress and Inflammation in Zebrafish. J Hazard Mater. 2022;440:129823. doi:10.1016/j.jhazmat.2022.129823
56. Shanaka KASN, Jung S, Madushani KP, et al. Generation of Viperin-Knockout Zebrafish by CRISPR/Cas9-Mediated Genome Engineering and the Effect of This Mutation under VHSV Infection. Fish Shellfish Immunol. 2022;131:672–681. doi:10.1016/j.fsi.2022.10.040
57. Sudhakaran G, Rajesh R, Guru A, et al. Deacetylated Nimbin Analog N2 Fortifies Alloxan-Induced Pancreatic β-Cell Damage in Insulin-Resistant Zebrafish Larvae by Upregulating Phosphoenolpyruvate Carboxykinase (PEPCK) and Insulin Levels. Toxicol Appl Pharmacol. 2022;454:116229. doi:10.1016/j.taap.2022.116229
58. Li S, Jiang C, Chen H, et al. Pre-Injection of Zebrafish (Danio Rerio) Tnfb Polyclonal Antibody Decreases the Mortality of Vibrio vulnificus Infected Zebrafish. Front Vet Sci. 2021;8:741242. doi:10.3389/fvets.2021.741242
59. Lonsdale DO, Shah RV, Lipman J. Infection, Sepsis and the Inflammatory Response: mechanisms and Therapy. Front Med Lausanne. 2020;7:588863. doi:10.3389/fmed.2020.588863
60. Quaglio AEV, Grillo TG, De Oliveira ECS, Di Stasi LC, Sassaki LYGM. Inflammatory Bowel Disease and Colorectal Cancer. World J Gastroenterol. 2022;28:4053–4060. doi:10.3748/wjg.v28.i30.4053
61. Priyanka P, Meena PR, Raj D, et al. Urinary Tract Infection and Sepsis Causing Potential of Multidrug-Resistant Extraintestinal Pathogenic E. Coli Isolated from Plant-Origin Foods. Int J Food Microbiol. 2023;386:110048. doi:10.1016/j.ijfoodmicro.2022.110048
62. Shea AE, Forsyth VS, Stocki JA, et al. Emerging Roles for ABC Transporters as Virulence Factors in Uropathogenic Escherichia coli. Proc Natl Acad Sci U S A. 2024;121:e2310693121. doi:10.1073/pnas.2310693121
63. Barber AE, Fleming BA, Mulvey MA. Similarly Lethal Strains of Extraintestinal Pathogenic Escherichia coli Trigger Markedly Diverse Host Responses in a Zebrafish Model of Sepsis. mSphere. 2016;1:e00062–16. doi:10.1128/mSphere.00062-16
64. Whelan S, Lucey B, Finn K. Uropathogenic Escherichia coli (UPEC)-Associated Urinary Tract Infections: the Molecular Basis for Challenges to Effective Treatment. Microorganisms. 2023;11:2169. doi:10.3390/microorganisms11092169
65. Tran VLT, Hortle E, Britton WJ, Oehlers SH. Common Anti-Haemostatic Medications Increase the Severity of Systemic Infection by Uropathogenic Escherichia coli. Microbiol Res. 2022;254:126918. doi:10.1016/j.micres.2021.126918
66. van der Poll T, Opal SM. Pathogenesis, Treatment, and Prevention of Pneumococcal Pneumonia. Lancet. 2009;374:1543–1556. doi:10.1016/S0140-6736(09)61114-4
67. Rounioja S, Saralahti A, Rantala L, et al. Defense of Zebrafish Embryos against Streptococcus pneumoniae Infection Is Dependent on the Phagocytic Activity of Leukocytes. Dev Comp Immunol. 2012;36:342–348. doi:10.1016/j.dci.2011.05.008
68. Johri AK, Paoletti LC, Glaser P, et al. Group B Streptococcus: global Incidence and Vaccine Development. Nat Rev Microbiol. 2006;4:932–942. doi:10.1038/nrmicro1552
69. van Sorge NM, Doran KS. Defense at the Border: the Blood-Brain Barrier versus Bacterial Foreigners. Future Microbiol. 2012;7:383–394. doi:10.2217/fmb.12.1
70. Kim BJ, Hancock BM, Cid ND, Bermudez A, Traver D, Doran KS. Streptococcus agalactiae Infection in Zebrafish Larvae. Microb Pathogenesis. 2015;79. doi:10.1016/j.micpath.2015.01.007
71. Qin S, Xiao W, Zhou C, et al. Pseudomonas aeruginosa: pathogenesis, Virulence Factors, Antibiotic Resistance, Interaction with Host, Technology Advances and Emerging Therapeutics. Signal Transduct Target Ther. 2022;7:199. doi:10.1038/s41392-022-01056-1
72. Sathe N, Beech P, Croft L, Suphioglu C, Kapat A, Athan E. Pseudomonas aeruginosa: infections and Novel Approaches to Treatment “Knowing the Enemy” the Threat of Pseudomonas aeruginosa and Exploring Novel Approaches to Treatment. Infectious Med. 2023;2:178–194. doi:10.1016/j.imj.2023.05.003
73. Singh VK, Almpani M, Maura D, et al. Tackling Recalcitrant Pseudomonas aeruginosa Infections in Critical Illness via Anti-Virulence Monotherapy. Nat Commun. 2022;13:5103. doi:10.1038/s41467-022-32833-9
74. Brannon MK, Davis JM, Mathias JR, et al. Pseudomonas aeruginosa Type III Secretion System Interacts with Phagocytes to Modulate Systemic Infection of Zebrafish Embryos. Cell Microbiol. 2009;11:755–768. doi:10.1111/j.1462-5822.2009.01288.x
75. Howden BP, Giulieri SG, Wong Fok Lung T, et al. Staphylococcus aureus Host Interactions and Adaptation. Nat Rev Microbiol. 2023;21:380–395. doi:10.1038/s41579-023-00852-y
76. Roehrig C, Huemer M, Lorgé D, et al. MEndoB, a Chimeric Lysin Featuring a Novel Domain Architecture and Superior Activity for the Treatment of Staphylococcal Infections. mBio. 15:e02540–23. doi:10.1128/mbio.02540-23
77. Stojowska-Swędrzyńska K, Łupkowska A, Kuczyńska-Wiśnik D, Laskowska E. Antibiotic Heteroresistance in Klebsiella pneumoniae. Int J Mol Sci. 2021;23:449. doi:10.3390/ijms23010449
78. Gálvez-Silva M, Varas MA, Allende ML, Chávez FP, Marcoleta AE. Zebrafish Larvae Microinjection and Automated Fluorescence Microscopy for Studying klebsiella Pneumoniae Infection and the Host Immune Response. In: Foodborne Bacterial Pathogens: Methods and Protocols. New York, NY: Springer US; 2025:171–179.
79. A B, A G, M G, et al. Propofol Ameliorates Exaggerated Human Neutrophil Activation in a LPS Sepsis Model. J Inflamm Res. 2021;14. doi:10.2147/JIR.S314192
80. Zheng D, Liwinski T, Elinav E. Inflammasome Activation and Regulation: toward a Better Understanding of Complex Mechanisms. Cell Discov. 2020;6:1–22. doi:10.1038/s41421-020-0167-x
81. Zhang Y, Ning B. Signaling Pathways and Intervention Therapies in Sepsis. Sig Transduct Target Ther. 2021;6:1–36. doi:10.1038/s41392-021-00816-9
82. Philip AM, Wang Y, Mauro A, et al. Development of a Zebrafish Sepsis Model for High-Throughput Drug Discovery. Mol Med. 2017;23:134–148. doi:10.2119/molmed.2016.00188
83. Liu D, Huang S-Y, Sun J-H, et al. Sepsis-Induced Immunosuppression: mechanisms, Diagnosis and Current Treatment Options. Mil Med Res. 2022;9:56. doi:10.1186/s40779-022-00422-y
84. Philip AM, Vijayan MM. Stress-Immune-Growth Interactions: cortisol Modulates Suppressors of Cytokine Signaling and JAK/STAT Pathway in Rainbow Trout Liver. PLoS One. 2015;10:e0129299. doi:10.1371/journal.pone.0129299
85. Li T, Yang Y, Qi H, et al. CRISPR/Cas9 Therapeutics: progress and Prospects. Sig Transduct Target Ther. 2023;8:1–23. doi:10.1038/s41392-023-01309-7
86. Adli M. The CRISPR Tool Kit for Genome Editing and Beyond. Nat Commun. 2018;9. doi:10.1038/s41467-018-04252-2
87. Chen C, Zhang Z, Liu C, Sun P, Liu P, Li X. ABCG2 Is an Itaconate Exporter That Limits Antibacterial Innate Immunity by Alleviating TFEB-Dependent Lysosomal Biogenesis. Cell Metab. 2024;36:498–510.e11. doi:10.1016/j.cmet.2023.12.015
88. Jiang Y, Lu Y, Jiang X, et al. Glucocorticoids Induce Osteoporosis Mediated by Glucocorticoid Receptor-Dependent and -Independent Pathways. Biomed Pharm. 2020;125:109979. doi:10.1016/j.biopha.2020.109979
89. Saralahti AK, Harjula S-KE, Rantapero T, et al. Characterization of the Innate Immune Response to Streptococcus pneumoniae Infection in Zebrafish. PLoS Genet. 2023;19:e1010586. doi:10.1371/journal.pgen.1010586
90. Bonten M, Johnson JR, van den Biggelaar AHJ, et al. Epidemiology of Escherichia coli Bacteremia: a Systematic Literature Review. Clin Infect Dis. 2021;72:1211–1219. doi:10.1093/cid/ciaa210
91. Kralicek SE, Sitaraman LM, Kuprys PV, et al. Clinical Manifestations and Stool Load of Atypical Enteropathogenic Escherichia coli Infections in United States Children and Adults. Gastroenterology. 2022;163:1321–1333. doi:10.1053/j.gastro.2022.07.064
92. Weiser JN, Ferreira DM, Paton JC. Streptococcus pneumoniae: transmission, Colonization and Invasion. Nat Rev Microbiol. 2018;16:355–367. doi:10.1038/s41579-018-0001-8
93. Sironi M, Cagliani R, Forni D, Clerici M. Evolutionary Insights into Host–Pathogen Interactions from Mammalian Sequence Data. Nat Rev Genet. 2015;16:224–236. doi:10.1038/nrg3905
94. Lohrmann F, Hufnagel M, Kunze M, et al. Neonatal Invasive Disease Caused by Streptococcus agalactiae in Europe: the DEVANI Multi-Center Study. Infection. 1. doi:10.1007/s15010-022-01965-x
95. Grandy S, Scur M, Dolan K, Nickerson R, Cheng Z. Using Model Systems to Unravel Host–Pseudomonas aeruginosa Interactions. Environ Microbiol. 2023;25:1765–1784. doi:10.1111/1462-2920.16440
96. Santos RA, Cardoso C, Pedrosa N, et al. LPS-Induced Mortality in Zebrafish: preliminary Characterisation of Common Fish Pathogens. Microorganisms. 2023;11:2205. doi:10.3390/microorganisms11092205
97. Doudna JA. The Promise and Challenge of Therapeutic Genome Editing. Nature. 2020;578:229–236. doi:10.1038/s41586-020-1978-5
98. Patton EE, Zon LI, Langenau DM. Zebrafish Disease Models in Drug Discovery: from Preclinical Modelling to Clinical Trials. Nat Rev Drug Discov. 2021;20:611–628. doi:10.1038/s41573-021-00210-8
99. Neto S, Vieira A, Oliveira H, Espiña B. Assessing Acinetobacter Baumannii Virulence and Treatment with a Bacteriophage Using Zebrafish Embryos. THE FASEB Journal. 2023;37:e23013. doi:10.1096/fj.202300385R
100. Choi T-Y, Choi T-I, Lee Y-R, Choe S-K, Kim C-H. Zebrafish as an Animal Model for Biomedical Research. Exp Mol Med. 2021;53(3):310–317. doi:10.1038/s12276-021-00571-5
101. Zanandrea R, Bonan CD, Campos MM. Zebrafish as a Model for Inflammation and Drug Discovery. Drug Discovery Today. 2020;25(12):2201–2211. doi:10.1016/j.drudis.2020.09.036
102. Szweda P, Schielmann M, Kotlowski R, Gorczyca G, Zalewska M, Milewski S. Peptidoglycan Hydrolases-Potential Weapons against Staphylococcus aureus. Appl Microbiol Biotechnol. 2012;96(5):1157–1174. doi:10.1007/s00253-012-4484-3
103. Chera JS, Kumar S, Bairagi AK, et al. Poly(I:C), a double stranded RNA analog, activates the anti-viral DNA sensors in Buffalo fibroblasts. Veterinary Vaccine. 2023;2(1):100016. doi:10.1016/j.vetvac.2023.100016
104. Palchetti S, Starace D, De Cesaris P, Filippini A, Ziparo E, Riccioli A. Transfected Poly(I:C) Activates Different dsRNA Receptors, Leading to Apoptosis or Immunoadjuvant Response in Androgen-Independent Prostate Cancer Cells. J Biol Chem. 2015;290(9):5470–5483. doi:10.1074/jbc.M114.601625
105. Komal A, Noreen M, El-Kott AF. TLR3 Agonists: RGC100, ARNAX, and Poly-IC: a Comparative Review. Immunol Res. 2021;69(4):312–322. doi:10.1007/s12026-021-09203-6
106. Kavaliauskis A, Arnemo M, Kim S-H, et al. Use of Poly(I:C) Stabilized with Chitosan As a Vaccine-Adjuvant Against Viral Hemorrhagic Septicemia Virus Infection in Zebrafish. Zebrafish. 2015;12(6):421–431. doi:10.1089/zeb.2015.1126
107. Urati A, Angati A, Singh Gautam A, Dey M, Pandey SK, Singh RK. Neuroprotective Responses of Quercetin in Regulation of Biochemical, Structural, and Neurobehavioral Effects in 28-Day Oral Exposure of Iron in Rats. Toxicol Mech Methods. 2024;34(1):57–71. doi:10.1080/15376516.2023.2256840
108. Molagoda IMN, Jayasingha JACC, Choi YH, Jayasooriya RGPT, Kang C-H, Kim G-Y. Fisetin Inhibits Lipopolysaccharide-Induced Inflammatory Response by Activating β-Catenin, Leading to a Decrease in Endotoxic Shock. Sci Rep. 2021;11(1):8377. doi:10.1038/s41598-021-87257-0
109. Al-Saadi HM, Pang K-L, Ima-Nirwana S, Chin K-Y. Multifaceted Protective Role of Glucosamine against Osteoarthritis: review of Its Molecular Mechanisms. Scientia Pharmaceutica. 2019;87(34). doi:10.3390/scipharm87040034
110. Hwang J-S, Kim K-H, Park J, et al. Glucosamine Improves Survival in a Mouse Model of Sepsis and Attenuates Sepsis-Induced Lung Injury and Inflammation. J Biol Chem. 2019;294:608–622. doi:10.1074/jbc.RA118.004638
111. Niederman MS, Baron RM, Bouadma L, et al. Initial Antimicrobial Management of Sepsis. Crit Care. 2021;25:307. doi:10.1186/s13054-021-03736-w
112. Letrado P, Corsini B, Díez-Martínez R, Bustamante N, Yuste JE, García P. Bactericidal Synergism between Antibiotics and Phage Endolysin Cpl-711 to Kill Multidrug-Resistant Pneumococcus. Future Microbiol. 2018;13:1215–1223. doi:10.2217/fmb-2018-0077
113. Larijani B, Hamidpour SK, Tayanloo-Beik A, et al. An Overview of Zebrafish Modeling Methods in Drug Discovery and Development. Adv Exp Med Biol. 2022;1387:145–169. doi:10.1007/5584_2021_684
114. Zhang X, Song J, Klymov A, et al. Monitoring Local Delivery of Vancomycin from Gelatin Nanospheres in Zebrafish Larvae. Int J Nanomed. 2018;13:5377–5394. doi:10.2147/IJN.S168959
115. Singh Angom R, Wang Y, Wang E, Dutta SK, Mukhopadhyay D. Conditional, Tissue-Specific CRISPR/Cas9 Vector System in Zebrafish Reveals the Role of Nrp1b in Heart Regeneration. Arterioscler Thromb Vasc Biol. 2023;43:1921–1934. doi:10.1161/ATVBAHA.123.319189
116. Baxendale S, van Eeden F, Wilkinson R. The Power of Zebrafish in Personalised Medicine. Adv Exp Med Biol. 2017;1007:179–197. doi:10.1007/978-3-319-60733-7_10
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.