Back to Journals » International Journal of Nanomedicine » Volume 21
Exosomes as Cellular Communicators and Therapeutic Agents in Orthopedic Diseases: From Mechanisms to Intervention
Authors Zhang J, Zhang M ![]() , Ren X
, Ren X ![]() , Li M, Zhu Y
, Li M, Zhu Y ![]() , Shou D, Zhang Y
, Shou D, Zhang Y ![]()
Received 17 November 2025
Accepted for publication 18 February 2026
Published 13 March 2026 Volume 2026:21 582197
DOI https://doi.org/10.2147/IJN.S582197
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Eng San Thian
Jianwen Zhang,1,2 Minjie Zhang,2 Xiaoru Ren,2 Mengying Li,2 Yiwen Zhu,1,3 Dan Shou,2 Yang Zhang1,2
1Institute of Orthopaedics and Traumatology, The First Affiliated Hospital of Zhejiang Chinese Medical University (Zhejiang Provincial Hospital of Chinese Medicine), Hangzhou, People’s Republic of China; 2School of Pharmaceutical Sciences, Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China; 3The First School of Clinical Medicine, Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China
Correspondence: Yang Zhang; Dan Shou, Email [email protected]; [email protected]
Abstract: Exosome-mediated intercellular communication has become a critical mechanism in the pathogenesis, progression, and regenerative repair of orthopedic diseases. By delivering bioactive molecules, exosomes dynamically regulate bone remodeling, cartilage homeostasis, and inflammatory responses—processes that are commonly disrupted in conditions such as osteoporosis, osteoarthritis, and bone non-union. Current therapeutic approaches often fail to achieve complete tissue repair or reverse disease progression, representing a major clinical challenge in orthopedics. This systematic review examines how exosome secretion, cargo loading, and cellular uptake are modulated by physical, chemical, biological, and pharmacological factors, thereby influencing disease progression and tissue repair. Furthermore, we evaluate the translational potential of engineered exosomes as targeted therapeutic strategies and analyze the dual dilemmas currently faced in exosome research and clinical translation: on one hand, exosomes themselves encounter technical bottlenecks such as standardization of isolation, drug-loading efficiency, large-scale production, and targeted delivery; on the other hand, their clinical application remains limited by unclear in vivo metabolic mechanisms, lack of efficacy evaluation systems, and insufficient clinical validation. Overcoming these challenges will be essential to advancing the real-world clinical application of exosomes in orthopedics.
Keywords: exosomes, orthopedic diseases, intercellular communication
Introduction
Orthopedic diseases pose a significant threat to global health, with rising incidences in recent years. They are primarily classified according to the affected anatomical region: joint-related disorders such as osteoarthritis (OA) and rheumatoid arthritis (RA); bone-related diseases including osteoporosis (OP), osteonecrosis of the femoral head (ONFH), osteonecrosis (OS), and fractures; and spinal pathologies such as intervertebral disc degeneration (IVDD).
While the clinical manifestations of orthopedic diseases vary, these conditions undoubtedly have a significant impact on patients’ daily lives.1 Patients commonly experience pain, restricted mobility, and, in severe cases, varying degrees of functional loss.2 Epidemiological data indicates that over the past three decades, not only osteoarthritis and rheumatoid arthritis but also other orthopedic diseases have shown a continuous upward trend in incidence.3–5 Although the pathogenic mechanisms differ among these diseases, they generally involve the following key pathological processes: immune-inflammatory responses, imbalance between osteogenesis and osteoclastogenesis, abnormal angiogenesis, and involvement of the tumor microenvironment.6–11
In addressing the adverse impacts of orthopedic diseases, current treatment approaches primarily rely on two modalities: pharmacological therapy and surgical intervention. Pharmacological treatment mainly involves the use of nonsteroidal anti-inflammatory drugs (NSAIDs), analgesics, glucocorticoids, disease-modifying antirheumatic drugs (DMARDs), anti-osteoporotic agents, and biologics. However, these medications are primarily employed to alleviate pain and slow disease progression, rather than to target the underlying pathological mechanisms of the disease. In contrast, surgical intervention is highly effective in addressing structural issues caused by orthopedic conditions, such as local mechanical disorders, structural damage, trauma, deformities, severe degeneration, and tumors. Nonetheless, it also carries the risk of more significant adverse effects, including unavoidable tissue damage, nonunion of bones, and difficulties in cartilage repair.12–14
In recent years, extracellular vesicles (EVs) have become a forefront direction in orthopedic research. These membrane-bound structures are released by cells and carry bioactive molecules such as proteins, nucleic acids, and lipids. They facilitate intercellular communication and regulate key processes essential for maintaining tissue homeostasis in orthopedics, including osteogenesis, cartilage protection, angiogenesis, and immunomodulation.
EVs exhibit significant heterogeneity, primarily comprising three subtypes: exosomes, microvesicles, and apoptotic bodies. They differ fundamentally in origin, size, and function: exosomes (30–150 nm) are generated via the endocytic pathway and participate in precise signal transduction under homeostatic conditions; microvesicles (100–1000 nm) are released through plasma membrane budding and often mediate prominent signaling events during cellular stress; while apoptotic bodies (500–5000 nm) are products of apoptosis, responsible for clearing cellular debris.15–17 Given that current research on orthopedic diseases primarily focuses on exosomes and microvesicles among EVs, this article will provide an overview of EVs and then focus specifically on recent advances in the study of exosomes and microvesicles.
However, the clinical translation of natural exosomes faces inherent limitations, including low yield, cargo heterogeneity, lack of tissue-specific targeting, and rapid in vivo clearance. These challenges necessitate the development of strategies to enhance exosome production, engineer their cargo, and improve delivery efficiency.
The integration of biomaterials science and exosome engineering has opened new avenues for tailored exosome-based therapies.18 Approaches such as cytokine preconditioning, incorporation of metallic ions, and combination with bioactive materials are being explored to optimize exosome function for orthopedic applications.19
This review systematically examines the role of exosome-mediated intercellular communication in orthopedic diseases, with emphasis on how modulation strategies can overcome existing limitations and enhance therapeutic potential. Current challenges and future directions for clinical translation are also discussed.
Biological Characterization of Exosomes
Exosomes, nanoscale EVs with diameters ranging from 40 to 160 nm (average ~100 nm), are formed through the endosomal pathway.20 Their biogenesis involves four key stages: (1) endocytic internalization of molecular cargo; (2) early endosomal sorting; (3) maturation into multivesicular bodies (MVBs); (4) Rab-GTPase-dependent fusion with the plasma membrane for extracellular release.21 During this process, cargo designated for recycling traffics to the Golgi apparatus or plasma membrane via recycling endosomes22 (Figure 1).
 |
Figure 1 Processes of exosome formation and secretion. The process of exosome-mediated intercellular communication includes: (a) Formation of early sorting endosome; (b)The intraluminal vesicle (ILV) of an MVB are released into the extracellular space through exocytosis, forming exosomes; (c) Uptake of exosomes by recipient cells. (d) The contents of exosomes mainly include proteins, lipids, nucleic acids, as well as carbohydrates, metabolites, etc. |
Exosomal cargos comprise three principal biomolecular classes: proteins, nucleic acids, and lipids.23 The proteins in exosomes can be mainly categorized as responsible for cell adhesion and targeting, membrane fusion, endosomal sorting complexes required for transport proteins (ESCRT), transport binding and other proteins. The tetraspanins (CD9, CD37, CD53, CD63, CD81, CD82) and integrin proteins are mainly used for cell adhesion; Ras-associated binding (Rab-GTPase), membrane associated proteins, and heat shock proteins (HSP) are used for membrane fusion; tumor susceptibility gene 101 (TSG101), and vacuolar protein sorting 4 (VPS4) are responsible for cellular transport.24 The nucleic acid repertoire predominantly includes messenger RNAs (mRNAs) and non-coding RNAs, particularly microRNAs (miRNAs). Emerging evidence demonstrates that exosomal miRNAs regulate critical cellular processes such as osteogenic differentiation, angiogenesis, metastatic niche formation, and disease pathogenesis through cross-species gene silencing.25
Mechanisms and Roles of Exosome-Mediated Intercellular Communication in Orthopedic Diseases
Skeletal homeostasis is not a static state but a dynamic equilibrium maintained through the coordinated interplay of multiple tissues—including bone, cartilage, synovium, immune cells, and vasculature. This balance is fundamentally regulated by a complex and orderly network of intercellular communication, where cells exchange information via direct contact, soluble factors, and extracellular vesicles such as exosomes.26–28 These interactions govern core processes like bone remodeling, cartilage integrity, injury repair, and immune surveillance. Disruption of this communicative network shifts the microenvironment from a state of orderly repair to a pathological trajectory of chronic destruction and functional loss.
Exosomes have emerged as central mediators in this context, carrying bioactive cargoes (miRNAs, proteins, lipids) that can reprogram cellular behavior.29,30 Their therapeutic potential lies in their ability to restore disrupted communication circuits in various orthopedic diseases, which can be broadly categorized into the following pathophysiological themes (Figure 2). In the context of the pathogenic mechanisms underlying orthopedic diseases, exosomes primarily exert therapeutic effects across different types of orthopedic conditions by targeting immune-inflammatory dysregulation, restoring osteoblast-osteoclast balance, promoting tissue regeneration and matrix repair, and modulating the tumor microenvironment. These processes exhibit both overlap and distinctiveness.
 |
Figure 2 Strategies for treating orthopedic diseases with exosomes. Exosomes and their cargoes primarily exert therapeutic effects on orthopedic diseases by promoting bone formation, enhancing anti-inflammatory capabilities, facilitating angiogenesis, and inducing tumor cell apoptosis. In the figures, upward arrows indicate a promoting or enhancing effect, while downward arrow represent an inhibitory or suppressive effect. The plus sign (+) denotes an increase in secretion, and the minus sign (-) signifies a decrease in production. |
Targeting Immune-Inflammatory Dysregulation
Bone serves not only as the structural support of the human body but also as a crucial immune organ, maintaining the stability of the bone microenvironment through the secretion of various inflammatory factors. In fact, many key factors leading to orthopedic diseases, such as aging, metabolic dysfunction, and cellular genetic mutations, originate from an imbalance in immune-inflammatory interactions.31
In the pathogenesis of OA, common triggers like mechanical stress, injury, and metabolic dysregulation disrupt intercellular homeostasis. This dysregulation sends signals recognized by macrophages, leading to their activation and the release of a series of inflammatory factors, such as interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α).32,33 These cytokines establish a destructive positive feedback loop with chondrocytes and synovial fibroblasts: they not only directly inhibit the synthesis of type II collagen and proteoglycans by chondrocytes and activate matrix metalloproteinases (MMPs) to accelerate matrix degradation, but also further induce chondrocytes to secrete more inflammatory factors, promoting synovial hyperplasia and pannus formation.34 Consequently, a local “low-grade inflammatory” state becomes a self-perpetuating chronic destructive process. A strikingly similar mechanism of chronic low-grade inflammation dominates the progression of intervertebral disc degeneration. Nutrient deprivation and abnormal mechanical stress cause cellular stress in nucleus pulposus cells35. This, in turn, activates pathways such as NF-κB, leading to the secretion of inflammatory factors and matrix-degrading enzymes. This process creates a self-perpetuating catabolic environment, ultimately resulting in dehydration of the nucleus pulposus and rupture of the annulus fibrosus.
The inflammatory dialogue in RA begins with the “erroneous recognition” by the autoimmune system. Triggered by autoantigens, adaptive immunity is activated, involving interactions between T/B cells and macrophages. Fibroblast-like synoviocytes (FLSs) are activated and secrete large amounts of inflammatory factors such as IL-6, IL-17, and RANKL, exacerbating chondrocyte damage.36
Furthermore, the relationship between immunity and inflammation also influences fracture healing, and this relationship is dynamic. In the early stages of fracture healing, a moderate inflammatory response helps clear necrotic tissue at the fracture site and promotes angiogenesis. At this stage, released TNF-α and IL-6 primarily assist in recruiting cells necessary for tissue regeneration, such as bone mesenchymal stem cells, to the fracture area. However, excessive inflammatory cytokines can disrupt the bone repair process, potentially leading to bacterial infection at the fracture site, delayed union, or non-union.9 However, in diseases such as osteonecrosis of the femoral head, the intense inflammatory storm induced by ischemia (eg, TNF-α, IL-1β) is destructive. It not only suppresses osteoblast function and stimulates excessive osteoclast activation but also alters the fate of bone marrow mesenchymal stem cells, promoting their differentiation toward adipocytes rather than osteoblasts. Together, these effects disrupt the coupling of “bone formation and bone resorption”, which constitutes the core mechanism leading to structural collapse of the femoral head.
For orthopedic diseases triggered by immune-inflammatory responses, commonly used treatments include NSAIDs. These medications broadly inhibit inflammatory factors and alleviate disease-related pain. However, their drawbacks are evident: due to their broad suppression of inflammatory factors, they cannot precisely target the specific sites requiring inflammation resolution and may hinder some beneficial inflammatory responses.37 For severe damage or functional loss caused by orthopedic diseases, joint replacement surgery is also a common therapeutic approach. However, studies indicate that after joint replacement, the prosthesis inevitably creates friction with surrounding tissues. This mechanical damage can further exacerbate the inflammatory response, potentially leading to more severe prosthetic synovitis.38,39
Exosomes demonstrate core regulatory potential in the immunoinflammatory treatment of orthopedic diseases. Their essential role lies in serving as key carriers of intercellular communication, delivering bioactive substances such as miRNAs and proteins to reshape the disordered immune microenvironment, thereby halting the “inflammatory storm” and initiating repair programs.40 Despite differences in species of cellular origin, the potent immunosuppressive properties of mesenchymal stem cell (MSC)-derived exosomes enable them to effectively inhibit inflammation and promote bone regeneration across various orthopedic conditions. For instance, exosomes regulate synovial immune responses via the targeted delivery of anti-inflammatory cytokines and non-coding RNAs, promoting cartilage matrix regeneration.41 Additionally, M2 macrophage-derived exosomes modified with oligo-lysine and MMP-cleavable polyethylene glycol (PEG) exhibit enhanced enrichment within inflammatory regions due to their membrane structure, enabling targeted exosome therapy for rheumatoid arthritis.42 To address periprosthetic osteolysis induced by prostheses, existing research has utilized bone marrow mesenchymal stem cell (BMSC)-derived exosomes encapsulated within M2 macrophage membranes to inhibit inflammation at the joint site.43
Restoring Osteogenic-Osteoclastic Balance
The homeostasis and health of the skeletal system fundamentally rely on the dynamic balance between bone formation, led by osteoblasts, and bone resorption, carried out by osteoclasts. This intricate process is known as “bone remodeling”. These two cell types are tightly coupled through a complex intercellular communication network: osteoblasts and their precursor cells express RANKL, which binds to the RANK receptor on osteoclast precursors, driving their differentiation and activation. Simultaneously, osteoblasts secrete osteoprotegerin (OPG), which acts as a “decoy” receptor, competitively inhibiting RANKL and thus negatively regulating bone resorption. Additionally, signaling pathways such as Wnt/β-catenin are deeply involved in modulating osteoblast activity.44 Under physiological conditions, the activities of these two cell types are highly coordinated in time and space, ensuring stable bone mass and microstructural renewal.45
However, this core balance is disrupted in various orthopedic diseases. In postmenopausal osteoporosis, estrogen deficiency leads to an increase in pro-inflammatory cytokines, causing an imbalance in the RANKL/OPG ratio. This results in excessive enhancement of osteoclast activity and progressive bone loss.46 In rheumatoid arthritis, inflamed synovial tissue produces large amounts of RANKL and inflammatory factors, activating osteoclasts and leading to periarticular bone erosion.47
The pivotal role of exosomes in maintaining skeletal homeostasis is most prominently reflected in their exquisite regulation of the “osteogenic-osteoclastic balance”—the core unit of bone remodeling. Exosomes, particularly those derived from mesenchymal stem cells (MSCs), can bidirectionally correct this imbalance through the specific “cargo” they carry. On one hand, they can deliver pro-osteogenic miRNAs (such as miR-29a) to activate osteogenic signaling pathways like Wnt/β-catenin, thereby promoting the osteogenic differentiation of bone marrow mesenchymal stem cells and enhancing osteoblast activity.44,48 On the other hand, by transferring molecules such as miR-223 or mimicking the function of OPG, they can inhibit the RANKL/RANK signaling pathway, effectively curbing the differentiation and activation of osteoclasts.49,50 This dual-faceted regulation, which simultaneously targets both bone formation and bone resorption in a “promote formation while curbing resorption” manner, positions exosomes as a highly promising intelligent therapeutic tool. They possess the potential to fundamentally restore the dynamic balance of bone metabolism at the level of cellular communication, offering novel strategies for treating diseases characterized by bone loss, such as osteoporosis and periprosthetic osteolysis.
Promoting Tissue Regeneration and Matrix Repair
The pathogenesis of many degenerative and traumatic orthopedic conditions, such as OA, critical-sized bone defects, and intervertebral disc degeneration, is characterized by a failure of intrinsic tissue repair. This failure stems from a hostile microenvironment that disrupts the normal anabolic-catabolic balance.51 In OA, for instance, chronic low-grade inflammation drives chondrocytes and synovial cells to produce excessive matrix-degrading enzymes (eg, MMP-13, ADAMTS-5) while suppressing the synthesis of essential matrix components like type II collagen and aggrecan.52 In non-healing fractures or large bone defects, impaired angiogenesis, insufficient stem cell recruitment, and persistent inflammation create a regenerative deficit.53 Similarly, in disc degeneration, nutrient deprivation and inflammatory stress lead to nucleus pulposus cell senescence and apoptosis, resulting in progressive loss of proteoglycans and disc height.54
Exosomes offer a powerful, cell-free strategy to overcome these regenerative barriers by reprogramming the pathological microenvironment towards a pro-repair state. They function as multifaceted signaling hubs that deliver a synchronized combination of bioactive cargoes—including pro-angiogenic miRNAs (eg, miR-126), anabolic growth factors (eg, TGF-β), and survival-promoting miRNAs (eg, miR-21)—directly to resident and progenitor cells.55 In OA, stem cell-derived exosomes can suppress catabolic signaling pathways in chondrocytes, reduce the expression of MMPs, and simultaneously upregulate the production of collagen and aggrecan, thereby shifting the balance from degradation to matrix synthesis.56 In disc repair, exosomes can inhibit nucleus pulposus cell apoptosis, mitigate inflammation, and stimulate the synthesis of extracellular matrix components to restore disc hydration and mechanical function.57 By integrating these coordinated pro-regenerative signals, exosomes effectively convert a stagnant or catabolic tissue niche into one conducive to structured and functional tissue restoration.
Modulating the Tumor Microenvironment
The progression of tumors within the skeletal system is far from being merely the proliferation of isolated cancer cells; rather, it is a “vicious dialogue” meticulously orchestrated by tumor cells and co-opted by the normal bone microenvironment. Bone tumor cells establish a complex pro-tumorigenic communication network with osteoclasts, osteoblasts, immune cells, and stromal cells by secreting various signaling molecules (such as RANKL, PTHrP, CCL5, etc).58 The core function of this network is to construct a self-reinforcing “malignant niche”: tumor signals continuously activate osteoclasts, leading to osteolytic destruction while simultaneously releasing growth factors stored within the bone matrix, which in turn further nourish tumor growth.59
In response to this mechanism, modern therapeutic strategies are shifting from “killing tumor cells” to “disrupting their communication alliance”. For example, emerging strategies use exosomal miRNAs, such as miR-144, to suppress the proliferation and metastasis of OS.60 Furthermore, integrin-modified exosomes can be used to deliver lncRNA MEG3, demonstrating tumor-targeting efficacy.61
Targeting and Modulating Exosomal Intercellular Communication in Orthopedic Diseases
Exosome-mediated intercellular communication constitutes a sophisticated biological cascade comprising three core phases: (1) vesicle biogenesis and secretion, (2) cargo composition and functional programming, (3) recipient cell recognition and signal transduction. Modulation of these regulatory nodes enables exosomes to exert control over orthopedic disease progression.
For instance, preconditioning infrapatellar fat pad-derived mesenchymal stem cells (IPFP-MSCs) with TNF-α significantly enhances exosome secretion. Mechanistic studies reveal that TNF-α-primed IPFP-MSC exosomes exhibit enrichment of low-density lipoprotein receptor-related protein 1 (LRP1)—a key mediator of proteoglycan aggregation in articular cartilage—which potentiates extracellular matrix (ECM) homeostasis and mitigates OA-induced cartilage degeneration.62
The intercellular communication mediated by exosomes is a precisely orchestrated process, which amplifies the influence of external factors. These factors can modulate exosomal communication—and thereby influence the progression of orthopedic diseases—by regulating exosome secretion, biogenesis, cargo composition, and recipient cell uptake.
Mechanism of Exosome Secretion
The secretion of bone-associated exosomes involves multiple links, and in this process, many external factors can affect the secretion of exosomes. Some of these factors can regulate the secretion of exosomes by affecting the sorting-related proteins in exosomes, such as the Rab family, which is the main protein group responsible for intracellular localization and the regulation of intracellular endocytosis and extracellular transport63 which can regulate exosome secretion and have an impact on disease.64,65 Some factors, including icariin, regulate exosome secretion by promoting the production of its precursors. For instance, icariin enhances exosome production in osteoclasts by increasing the cytoplasmic formation of MVBs, which in turn improves bone healing in defect areas.66
Regulation of Exosomal Precursor MVBs (or ILVs)
The biogenesis of MVBs and ILVs is classified into ESCRT-dependent and ESCRT-independent pathways. The ESCRT system comprises a cascade of protein complexes that cooperatively mediate the sorting of ubiquitinated proteins and lipids into ILVs. Within this system, ESCRT-0 and ESCRT-I recognize and sequester ubiquitinated cargo,67,68 while ESCRT-II and ESCRT-III drive membrane curvature and scission, ultimately facilitating MVB maturation.69 TSG101, a core component of ESCRT-I, synergizes with ESCRT-0 and ESCRT-II to sort endosomal membrane components into ILVs through inward budding. Genetic ablation of TSG101 significantly impairs MVB biogenesis and subsequent exosome secretion, highlighting its essential role in this pathway.70
In contrast, the ESCRT-independent pathway bypasses canonical protein complexes and instead relies on lipid-driven mechanisms. This pathway utilizes ceramide-enriched microdomains, tetraspanin family proteins (eg, CD63 and CD9), and HSP (eg, HSP70) to promote spontaneous membrane invagination and cargo packaging into ILVs.71 For instance, pharmacological modulation of neutral sphingomyelinase (nSMase) activity directly regulates ceramide biosynthesis, thereby controlling the efficiency of MVB membrane budding.72
Biological Factors
The use of the nSMase inhibitor GW4869 reduces ceramide production and decreases the production of MVB and exosomes. In addition, phospholipase D2 has the ability to break down the lipid lecithin on exosome membranes into phosphatidic acid, which is similar to ceramide and promotes inward outgrowth of MVB membranes, which in turn promotes MVB production.70
Pharmaceutical Factors
Some herbal components can also regulate the amount of MVBs and ILVs and influence osteoblast function. For example, Icariin increased the presence of MVB in the cytoplasm, led to an increase in exosomes secreted by osteoclasts, and enhanced bone healing in areas of bone defects.66
Regulating the Intracellular Transport of the Exosomal Precursor MVB
The intracellular transport of MVBs requires the synergistic action of the cytoskeleton, kinesins, and Rab proteins. In the presence of kinesins, MVBs move along the cytoskeleton, while Rab proteins regulate their localization and transport direction, determining whether MVBs are degraded in lysosomes or fuse with the plasma membrane to secrete exosomes.73 Rab proteins play critical roles in this process. For example, Rab27a and Rab27b are key regulators of exosome secretion by facilitating MVB trafficking to the cell periphery and their docking at the plasma membrane. Knockdown of Rab27b reduces MVB size, whereas knockdown of Rab27a increases it.65
Rab39 regulates MVB localization through interaction with the BLOC-One Related Complex (BORC), which promotes basolateral exosome release.74 Additionally, ubiquitination—a common post-translational modification—regulates protein degradation and exosome secretion. The ESCRT complex recognizes ubiquitinated proteins and sorts them intoILVs of MVBs, which are subsequently released as exosomes. Recently, ISGylation, a ubiquitination-like modification, was found to inhibit exosome secretion by promoting MVB-lysosome fusion and ILV degradation.75
MVB acidification critically determines their fate: low-pH MVBs are targeted for lysosomal degradation, whereas neutral-pH MVBs migrate to the cell periphery for exosome release. This acidification is mediated by vacuolar-type H⁺-ATPase (V-ATPase), a proton pump that transports protons into the MVB lumen via ATP hydrolysis. nSMase2 activity inhibits V-ATPase assembly, reducing MVB acidification and thereby promoting exosome secretion. Calcium ion concentration also modulates exosome secretion, with elevated Ca2⁺ levels enhancing release.76 Furthermore, SNARE proteins regulate MVB trafficking by generating mechanical forces that drive MVB-plasma membrane fusion.77
Biological Factors
The inflammatory factor TNF-α activates nSMase2 on endosomes, which counteracts V-ATPase activity on MVB membranes, reducing endosomal acidification and promoting small exosome secretion.78 Furthermore, pretreatment of infrapatellar fat pad-derived IPFP-MSCs with TNF-α enhances exosome secretion, thereby alleviating OA-induced cartilage damage.62
Autophagy-Mediated Exosome Secretion
Autophagy is an intracellular degradation method, which mainly involves wrapping the cytoplasm, proteins, and organelles to form an autophagosome through a bilayer membrane and transporting damaged organelles, cytoplasmic contents, and proteins to lysosomes to complete decomposition and recycling.79 Autophagy can be categorized into macroautophagy, microautophagy, and molecular chaperone-mediated autophagy. Among them, macroautophagy mainly involves the formation of autophagosomes; microautophagy refers to the fact that lysosomes can directly phagocytose cytoplasmic components without autophagosomes; and molecular chaperone-mediated autophagy mainly leads proteins with specific amino acid sequences to lysosomes for degradation and recycling by selectively targeting them.80
Current studies have found that autophagy and exosome secretion share the same molecular mechanism, for example, autophagosomes generated by autophagy fuse with MVB formed by exosome endocytosis to form amphiphilic bodies, a structure that integrates autophagic degradation and exosome secretion; in addition to this, autophagy and exosome secretion both depend on lysosomal function; autophagy requires the autophagosome to interact with the lysosome to expel waste products, and exosome secretion is affected by the lysosome, exosome secretion increases when acidification of lysosomes is interrupted with the pro-lysosomal agents chloroquine and bafilomycin.81
After plasma membrane invagination to form ILVs, MVBs can either bind to the plasma membrane to deliver exosomes to the extracellular compartment or to the lysosome to degrade the contents, and in this way the amount of exosomes secreted is regulated.82 Thus autophagy-related proteins as well as signaling pathways can regulate exosome secretion and maturation. Common autophagy-related proteins that regulate exosome secretion mainly include Autophagy-related gene 5 (ATG5), Microtubule-associated protein light chain 3 (LC3), and it was found that knockdown of ATG5 on MVBs can reduce exosome secretion, on the other hand, LC3 mainly regulates MVB membrane growth and exosome sorting by binding to ESCRT complexes such as TSG101.83 Common signaling pathways mainly include AMP-activated protein kinase pathway (AMPK) and Mechanistic Target of Rapamycin Complex 1 signaling pathway (mTORC1), and AMPK activation promotes autophagy by inhibiting mTORC1, while enhancing exocytosis. Autophagy while enhancing exosome secretion to transmit stress signals to neighboring cells. For example, mTORC1 is a key negative regulator of autophagy, and activation of mTORC 1 inhibits exosome secretion, which is enhanced after mTORC1 inhibition by rapamycin.84
Pharmaceutical Factors
The Scutellarin primarily activates autophagy in nucleus pulposus cells (NPCs) by inhibiting the PI3K/PTEN/Akt pathway. Additionally, it upregulates Rab8a expression, which promotes the release of exosomes from rat NPCs. These findings suggest a potential mechanism for utilizing exosomes in the treatment of IVDD.85
Physical Factors
Low-intensity pulses promoted the release of exosomes from MSCs by activating autophagy and enhanced the positive effects of MSCs on cartilage.86 Under 2% hypoxic conditions, the upregulation of HIF-1α modulates mitophagy to maintain mitochondrial function, promotes the secretion of exosomes, and alleviates disc degeneration.57
Exosomes derived from plasma of cold-stressed mice decrease autophagy markers (LC3-II/LC3-I ratio) and increase P62/SQSTM1 accumulation in BMSCs through the action of their cargo miR-25-3p, which leads to cellular senescence and bone loss. Importantly, pharmacological inhibition of exosome release using GW4869 alleviates cold-induced bone loss.87
Regulation of Exosome Secretion Numbers
Fine-tuning the regulation of exosome secretion quantity is a crucial strategy for optimizing their biological functions and therapeutic potential. The level of secretion directly determines the intensity of intercellular signaling mediated by exosomes, thereby influencing downstream physiological or pathological processes. Current research indicates that their secretion can be modulated at multiple levels: first, by regulating key nodes in their biogenesis pathway, such as the formation of MVBs, the activity of ESCRT complexes, or Rab protein-mediated transport; second, by directly stimulating parental cells with physical, chemical, or biological factors (eg, hypoxia, mechanical stress, cytokine preconditioning, or specific drugs) to upregulate exosome production and release; and third, by employing engineering approaches (eg, gene editing or pharmacological intervention) to modify parental cells to enhance their basal secretion efficiency. These regulatory strategies aim to dynamically and controllably increase the yield of therapeutically active exosomes according to the demands of specific disease microenvironments, thereby laying the foundation for their effective application in regenerative medicine and disease treatment.
Chemical Factors (Including Pharmaceutical Factors)
It was found that pretreatment of Synovium-Derived Mesenchymal Stem Cells (SMSCs) with the metallic element strontium (Sr) resulted in an increase in the number of Sr-SMSC-Exo secretion, inhibition of chondrocyte iron apoptosis, and enhancement of the protection of bone microstructure, which enhanced the therapeutic efficacy of the exosomes in OA.88
Treatment of human MSC exosomes with GuiLu-ErXian Glue extract (GLEXG) was found to promote MSC-induced cartilage formation in vitro by increasing the release of exosomes and to play a role in delaying aging.89 Treatment of fibroblast-like synoviocytes (FLSs) with the phytoestrogen genistein (GEN) alleviated RA by reducing exosome release from these cells.90 In addition, alendronate (ALN), a commonly used therapeutic agent in orthopedics, has been shown to regulate the osteogenic-osteoclastic balance and promote bone formation by inhibiting the generation of osteoclast-derived exosomes.91
Physical Factors
Studies have shown that mechanical stimulation of bone cells promotes the production and release of exosomes containing bone-regulating proteins. These bone-regulating protein-enriched exosomes can effectively enhance bone formation. By comparing Human Umbilical Cord Mesenchymal Stem Cells (HucMSCs) cultured under normoxic and hypoxic conditions, researchers found that hypoxic conditions effectively promote exocytosis, enhance exosome secretion, and thereby facilitate fracture healing.92
Biological Factors
The current study found Cytokines play an important role in promoting exosome secretion, and some cytokines can regulate exosome secretion by activating specific signaling pathways. Certain pro-inflammatory factors can not only enhance the anti-inflammatory efficacy of exosomes but also effectively boost their production.
For example, pretreatment with TNF-α can activate the PI3K/AKT signaling pathway in IPFP-MSCs, which in turn promotes exosome secretion, and improves OA therapeutic efficacy.62,93 IL-1β can also increase exosome secretion and induce the expression of genes related to OA in articular cartilage.94
Tropoelastin (TE), a lipolytic protein with a high molecular weight, pretreatment with proelastin increased the ability of Adipose-Derived Stem Cells (ADSCs) to secrete exosomes. Furthermore, compared to untreated ADSC-Exo, Tropoelastin-Pretreated ADSC-Exo demonstrated enhanced therapeutic efficacy in an OA model.95
Pretreatment of MSC-derived exosomes with tauroursodeoxycholic acid (TUDCA)—an amphiphilic molecule that helps maintain cholesterol homeostasis—was found to enhance exosome production by upregulating exocytosis-related proteins. Additionally, it promoted the release of more anti-inflammatory factors and osteogenic proteins from the exosomes. As a result, researchers believe that TUDCA-preprocessed MSC-exosomes represent a promising material for bone regeneration therapy in the future.96 Furthermore, the adenosine A2A receptor (A2AR) has been found to regulate exosome secretion. The A2AR agonist CGS21680 effectively promotes the release of macrophage-derived exosomes, thereby enhancing vascular formation within the fracture microenvironment. In contrast, the A2AR antagonist ZM241385 not only inhibits the production of macrophage-derived exosomes but may also contribute to non-union of bone (Table 1).97
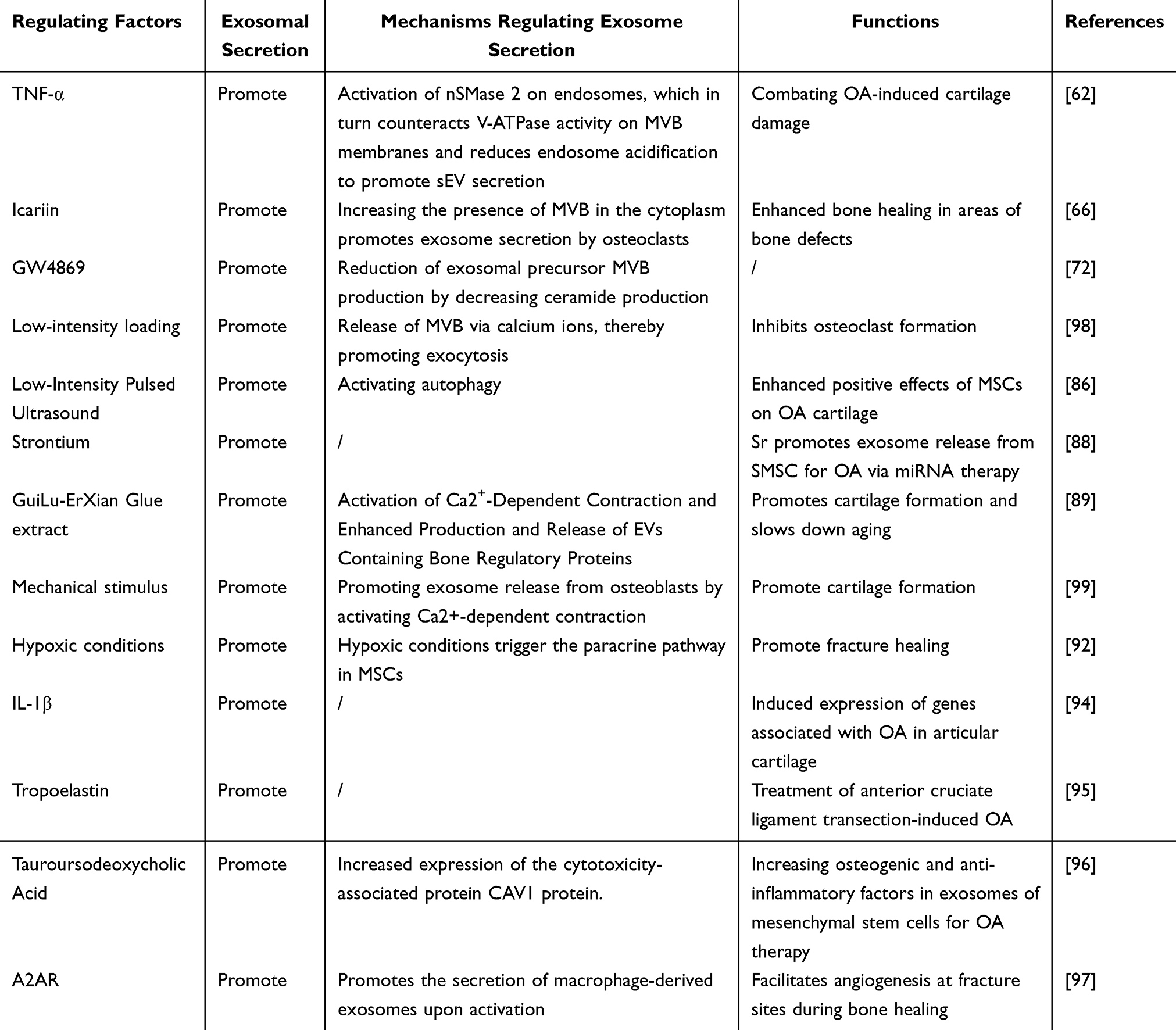 |
Table 1 Factors Regulating Exosome Secretion |
Regulation of Substances Transported in Exosomes
Exosomes contain various bioactive components, including proteins, nucleic acids, and lipids. Nucleic acids encompass miRNA, mRNA, long non-coding RNA, and circular RNA, while proteins such as CD9, CD81, TSG101, and ALIX facilitate exosome-cell fusion and signal transduction. Studies have shown that exosomal bioactive components demonstrate efficacy in regulating bone metabolism, promoting bone regeneration, and accelerating fracture healing.
Leveraging these characteristics, recent experimental work has increasingly shifted toward investigating the impact of external factors on exosomal cargo delivery. This encompasses the regulation of exosomal cargo loading (miRNAs, proteins) and the subsequent uptake processes by recipient cells.
Altering miRNAs in Exosomes
Current studies demonstrate that exosomal miRNAs from diverse tissue sources exhibit remarkable therapeutic potential in orthopedic diseases. Modifying the composition of miRNAs carried by exosomes is one of the core strategies for precisely regulating their biological functions and therapeutic specificity. As key components of exosomal signaling “cargo”, miRNAs can regulate gene expression at the post-transcriptional level by targeting specific mRNAs in recipient cells, thereby influencing critical physiological and pathological processes such as cell proliferation, differentiation, metabolism, and inflammatory responses. Currently, researchers primarily alter the miRNA profile of exosomes by intervening in the state of parental cells or engineering the exosomes themselves. Common approaches include treating parental cells with physical stimuli (eg, hypoxia, mechanical stress), chemical factors (eg, metal ions, specific drugs), or biological factors (eg, cytokine preconditioning) to upregulate or downregulate the enrichment of specific miRNAs in exosomes. Alternatively, synthetic or selected therapeutic miRNAs can be directly loaded into isolated exosomes via methods such as electroporation, sonication, or transfection. In the field of orthopedic diseases, engineered exosomes loaded with osteogenic miRNAs, anti-inflammatory miRNAs, or pro-angiogenic miRNAs can more precisely modulate immune-inflammatory responses, osteoblast-osteoclast balance, and angiogenesis within the skeletal microenvironment. This offers targeted and efficient novel therapeutic avenues for conditions such as osteoarthritis, osteoporosis, and bone defects.
Pharmaceutical Factors
Numerous drugs for orthopedic diseases are known to regulate exosomal miRNAs, including both herbal and chemical agents. Herbal compounds such as psoralen (a key active ingredient in traditional OP treatments) downregulate hsa-miR-125b-5p in periodontal ligament stem cell exosomes.100,101 Similarly, fucoidan (a seaweed-derived sulfated polysaccharide) exerts anti-osteoporotic effects by suppressing macrophage M1 polarization and osteoclast differentiation. Pretreatment of MSC-derived exosomes with fucoidan upregulates miR-146b-5p.102,103 Other herbal components, including icariin (from Epimedium) and quercetin (a polyhydroxy flavonoid), elevate miR-331-3p in osteoclast exosomes and miR-124-3p in BMSC exosomes, respectively.66,104 Notably, cinobufagin (a toad venom compound) upregulates miR-3102-5p in RAW-Exos, demonstrating antitumor and anti-inflammatory properties.105 Additionally, Morinda officinalis polysaccharide (MOP), traditionally used for bone strengthening, enhances exosomal miR-101-3p, providing insights into glucocorticoid-induced OP management.106
Furthermore, numerous traditional Chinese herbal formulations modulate exosomal miRNAs. For instance, Wen-Shen-Tong-Luo-Zhi-Tong Decoction (WSTLZT), a widely prescribed formula for OP, upregulates miR-122-5p in adipose-derived exosomes.107 Similarly, Zhuanggu Busui Formula (composed of Epimedium, Astragalus, Cortex Eucommiae, Panax ginseng, and Salvia miltiorrhiza) enhances osteogenic differentiation and suppresses bone loss in orthopedic diseases.108 Further studies demonstrate that this formula elevates miR-5100, miR-450b-5p, and miR-126a-3p in myogenic exosomes while reducing miR-669a-5p levels, highlighting its regulatory role in bone metabolism.109
In addition to traditional Chinese medicine, chemical agents can modulate exosomal miRNA expression. Digoxin (DIG), a cardiac glycoside, exhibits novel therapeutic potential in orthopedic diseases by downregulating miR-146b-5p in M1 macrophage-derived exosomes.110 Chondroitin sulfate (CS), a common OA therapeutic administered via intra-articular injection, faces efficacy limitations due to variability in purity and sourcing. To address this, researchers semi-synthesized CS derivatives (CS-semi) with controlled molecular weights using CS-A, which enhance miR-122-5p levels in serum exosomes.111 Pretreatment of BMSC-exosomes with thyroid hormone effectively promotes the enrichment of let-7a-5p in exosomes and significantly inhibits the expression of MMPs, leading to increased ECM expression and ultimately delaying the progression of osteoarthritis.112
Chemical Factors
Multiple metallic elements regulate exosomal miRNA expression, influencing bone disease therapeutics. Magnesium ions suppress miR-381 in macrophage-derived exosomes via autophagy,113 while high zinc concentrations (eg, 50 μM) elevate miR-146b-5p in senescent BMSC exosomes.114 Sr, a trace element, enhances bone regeneration through Sr-substituted calcium silicate, which upregulates miR-146a in BMSC exosomes.115 Furthermore, Sr pretreatment of SMSCs optimizes exosomal miRNA profiles by increasing miR-143-3p and suppressing miR-146a and miR-26a-5p, thereby ameliorating chondrocyte dysfunction.88
Metallic biomaterials similarly modulate miRNAs. Lithium-containing biomaterials upregulate miR-455-3p in BMSC exosomes,116 while the ionic products of 45S5 Bioglass® downregulate miR-342-5p and upregulate miR-1290 in MSC-derived exosomes.117
Physical Factors
Hypoxic pulp stem cell exosomes show elevated miR-210-3p.118 Hypoxic preconditioning of human umbilical cord MSCs (HucMSCs) enhances exosomal miR-126 production.92 Temperature also modulates miRNAs: cold exposure increases miR-25-3p in cold temperature-induced exosomes (CT-Exo).87 Mechanical stimuli, such as strain on osteoblasts or synovial joints, elevate miR-3110-5p and miR-3058-3p to promote bone repair,119 while reducing miR-214-3p in bone marrow mesenchymal stem cell (BMMSC)-Exo to improve bone vascularization and density.120 Additionally, Fe3O4 nanoparticles combined with a static magnetic field (SMF) enhance osteogenesis and angiogenesis via exosomal miR-1260a upregulation.121
Biological Factors
Inflammatory cytokines regulate exosomal miRNAs. IL-1β treatment increases miR-147b in MSC-Exo,122 while TGF-β1 stimulation elevates miR-135b.123 Conversely, pathological factors like RAS activation in breast cancer cells upregulate exosomal miR-494-3p, driving osteolytic bone metastasis.124 Muscle growth inhibitors suppress osteoblast exosomal miR-218, impairing osteogenic differentiation,125 and glucocorticoids (GCs) elevate miR-1a-3p in M1 macrophage exosomes, exacerbating femoral head OS.126
Tropoelastin approaches modulate miRNAs in adipose-derived stem cell exosomes (eg, upregulated miR-451-5p for cartilage regeneration95 (Table 2).
 |
Table 2 Factors Affecting miRNAs in Bone-Associated Exosomes |
Altering the Proteins in Exosomes
Exosomes are nanosized extracellular vesicles secreted by cells, characterized by a phospholipid bilayer membrane enriched with specific proteins such as tetraspanins (eg, CD9, CD81), integrins, and immunomodulatory molecules. These surface proteins facilitate exosome targeting, cellular adhesion, immune modulation, and intercellular communication. Strategies to enhance exosome therapeutic efficacy include modifying their protein composition to improve targeting and cellular uptake. For example, engineered exosomes encapsulating Bone Morphogenetic Protein-2 (BMP-2) within hydrogels exhibit prolonged BMP-2 stability and significantly enhanced bone regeneration capacity.127
Pharmaceutical Factors
Methotrexate (MTX) and sulfasalazine (SASP), both antifolate antirheumatic drugs, are frequently used in combination therapy. Studies reveal that MTX and SASP synergistically modify the protein profile of exosomes derived from the human synovial sarcoma cell line SW982. These modified exosomes show increased levels of Rab7B, a protein associated with anti-inflammatory macrophage activation, and reduced oxidative stress markers, offering a potential therapeutic strategy for RA.128–130
Chemical Factors
Hydrogen sulfide (H2S) plays a protective role in OA by mitigating chondrocyte damage and promoting cartilage integrity.131 Recent research demonstrates that H2S induces M2 macrophage polarization and alters exosomal protein composition, particularly enriching moesin. This moesin-enriched exosomal cargo enhances MSC endocytosis and bone regeneration via immune microenvironment remodeling.132 Additionally, lithium (Li) enhances Wnt10a secretion by facilitating its transport through Rab11a and Rab11 FIP1 complexes, thereby augmenting exosome-mediated osteogenesis (Table 3).133
 |
Table 3 Factors Affecting Proteins in Bone-Associated Exosomes |
Regulation of Receptor Cells by Exosomes
Exosome uptake by recipient cells occurs through three primary mechanisms: (1) Ligand-receptor interaction: Ligands on the exosomal membrane bind to cell surface receptors, activating receptor-mediated signaling pathways. This triggers endocytosis, allowing the internalization of exosomal cargo into the recipient cell; (2) Direct endocytosis: Recipient cells engulf exosomes via endocytic pathways. Following internalization, exosomal components (eg, proteins, nucleic acids) are released into the cytoplasm, with some participating in the biogenesis of new MVB (3) Membrane fusion: The exosomal membrane fuses or undergoes hemi-fusion with the plasma membrane, directly releasing proteins, mRNAs, miRNAs, and other cargo into the cytosol.134
Exosomes Regulate Their Ability to Bind to Receptor Cells
The binding capacity of exosomes to recipient cells is governed by multiple factors, including membrane fusion-associated proteins, interactions between exosomal surface proteins and cellular receptors, and bioactive cargo components (proteins, nucleic acids, lipids) that modulate recipient cell phenotypes and functions.135 Notably, nucleic acids such as miRNAs exert regulatory effects by post-transcriptionally silencing target genes in recipient cells. Current strategies to enhance exosome-recipient cell binding primarily utilize genetic engineering approaches. For example, studies have genetically modified exosomal glycosylation patterns to upregulate E-selectin ligands (eg, CD44), improving their affinity for E-selectin receptors on bone endothelial cells. Additionally, chemical conjugation of bone-targeting peptides (eg, (DSS)6) to exosome membranes enhances skeletal tropism, effectively mitigating bone loss in murine models of smoking-associated OP. However, these genetic modifications predominantly induce transient effects, limiting their long-term therapeutic efficacy.136
Exosomes Alter Receptor Cell Function by Delivering miRNAs
Different types of miRNAs in exosomes have distinct mechanisms to alter the function of recipient cells. Common mechanisms primarily include (1) regulating gene expression, where exosomal miRNAs affect cellular biological functions by modulating key signaling pathways in recipient cells; (2) influencing cell function through promoting proliferation, migration, or inducing apoptosis; and (3) immunomodulation, in which exosomal miRNAs suppress inflammatory responses by inhibiting inflammatory factors and key genes within inflammatory signaling pathways. For example, mechanical stress-induced exosomal miR-92a-3p promotes the proliferation and differentiation of BMSCs via targeted delivery. This pathway demonstrates significant potential as a therapeutic target for treating glucocorticoid-induced OP.137
Pharmaceutical Factors
In the treatment of OA, fucoidan reduces chondrocyte inflammation by upregulating exosomal miR-146b-5p and targeting tumor necrosis factor receptor-associated factor 6 (TRAF6), thereby inhibiting the PI3K/AKT/mTOR pathway. This enhances autophagy activation in chondrocytes, demonstrating significant therapeutic effects on OA.103 Compound CS-semi 5 inhibits p38 and MMP expression through upregulation of serum exosomal miR-122-5p, which binds specifically to the p38 3’ UTR region, promoting cartilage matrix synthesis and homeostasis to exert therapeutic effects in OA.111 Digoxin (DIG) downregulates miR-146b-5p in M1 macrophage exosomes and directly targets the 3’ UTR of USP3 (ubiquitin-specific protease 3). USP3 knockdown activates the NF-κB signaling pathway, thereby regulating the pro-inflammatory and catabolic state of chondrocytes and improving the osteoarthritic microenvironment.110 By inhibiting the delivery of exosomal miR-155 from M1 macrophages to CD4⁺ T cells, berberine (BBR) modulates CD4⁺ T cell immunometabolism and differentiation, thereby exerting anti-arthritic effects.138
In the treatment of OP, Wen Shen Tong Luo Zhi Tong Tang (WSTLZT) upregulates miR-122-5p in adipose-derived exosomes, regulates the SPRY2 protein-mediated MAPK signaling pathway, enhances osteogenic differentiation, inhibits adipogenic differentiation, delays bone necrosis, and exerts therapeutic effects on OP.107 Cinobufagin upregulates miR-3102-5p in exosomes derived from RAW264.7 cells, targeting the 3’ UTR of the lipid-oxidizing protein ALOX15 to suppress its expression and reduce lipid peroxidation in osteoblasts of osteoporotic mice.105 Icariin plays a critical role in treating bone defects by inducing miRNA-331-3p upregulation in osteoclast-derived exosomes (OC-exo), which targets fibroblast growth factor 23 (FGF23), reduces Dickkopf-related protein 1 (DKK1), and subsequently upregulates alkaline phosphatase (ALP) to stimulate osteoclast activity and enhance osteoblast function.66 Treatment with MQEF in a mouse model of hormone-induced ischemic OS of the femoral head significantly altered microRNA expression in bone tissue-derived exosomes. Specifically, miR-185-3p (7.2-fold increase) and miR-1b-5p were upregulated, while miR-129b-5p (2.2-fold decrease) and miR-223-5p were downregulated.139 Triptolide, a key active component of thunder god vine (Tripterygium wilfordii), demonstrates potent anti-inflammatory and immunomodulatory effects against RA. Furthermore, treatment of synoviocytes with triptolide inhibits the secretion of exosomes containing miR-221.140
Biological Factors
Biological factors such as myostatin promote osteoblast differentiation and bone formation by upregulating miR-218 in osteoblast-derived exosomes.125 Muscle-derived exosomal miR-27a-3p regulates osteoblast differentiation by targeting adenomatous polyposis coli (APC), activating the β-catenin pathway to mediate bone homeostasis.141 Inflammatory factor-treated exosomes alter miRNA profiles; for instance, TGF-β1-treated exosomes significantly increase miR-135b expression, which downregulates specificity protein 1 (SP1), an inhibitor of chondrocyte proliferation, thereby promoting chondrocyte proliferation and mitigating OA-induced cartilage damage.123 Pretreatment of ADSC-Exos with TE increases miR-451-5p expression and enhances exosome release capacity.95 Glucocorticoid-stimulated M1 macrophage exosomes elevate miR-1a-3p levels, promoting adipogenic differentiation of BMSCs and inducing glucocorticoid-associated OS of the femoral head,126 suggesting a novel therapeutic target for glucocorticoid-related bone diseases.
Chemical Factors
Lithium-substituted bioactive glass-ceramics (Li-BGC) promote cartilage repair by modulating BMSC-derived exosomes. Li-BGC upregulates exosomal miR-455-3p, which inhibits histone deacetylase 2 (HDAC2) to enhance histone H3 acetylation and gene transcription.116 Sr-substituted calcium silicate upregulates miR-146a in BMSC-derived exosomes, inhibits SMAD4 and NF2 translocation, enhances angiogenesis in human umbilical vein endothelial cells (HUVECs), and accelerates fracture healing.115
Physical Factors
Mechanical stress increases exosomal miR-92a-3p expression, which is internalized by MSCs to inhibit phosphatase and tensin homolog (PTEN) activity, activating the PI3K/AKT pathway and promoting osteogenic gene expression for bone regeneration.137 Temperature and oxygen also modulate exosomal miRNAs. Cold exposure elevates miR-25-3p in chondrocyte-derived exosomes (CT-exo), inhibiting BMSC autophagy and osteogenic differentiation by downregulating SATB2, leading to bone loss.87 Hypoxia preconditioning enhances exosomal miR-126 through HIF-1α, boosting HUVEC proliferation, angiogenesis, and migration for fracture healing.92 HIF-1α induced exosomal miR-21-5p promotes adipose stem cell differentiation and ectopic osteogenesis. Low-dose Fe3O4 nanoparticles combined with SMF upregulate exosomal miR-1260a, targeting HDAC7 inhibition for bone growth and COL4A2 inhibition for angiogenesis.121 Hypoxia elevates the level of the epigenetic factor miR-221-3p in MSC-exos and facilitates its transfer to senescent NPCs, where it regulates the DDIT4/NF-κB signaling pathway to delay NPC senescence142 (Figure 3).
 |
Figure 3 Regulation of exosomal miRNAs by external factors. Several external factors—such as fucoidan, digoxin (DIG), TGF-β1, hypoxia, strontium-substituted calcium silicate ceramics (Sr-CS), lithium-substituted bioactive glass-ceramics (Li-BGC), and Wen Shen Tong Luo Zhi Tong Tang (WSTLZT)—influence the uptake of exosomes by recipient cells through the modulation of exosomal miRNAs. In the figure, blue arrows represent negative effects, while red arrows indicate positive effects. |
Exosomes Regulate Recipient Cells Through Modulation of Proteins
Exosomes can modulate recipient cell function through multiple mechanisms after delivery. These include: (1) regulating intracellular signaling pathways to alter cellular behavior; (2) influencing exosome uptake efficiency by modifying the endocytic capacity and cytoskeletal dynamics of recipient cells. Recent studies revealed that Qiangguyin (QGY)-treated osteoclast-derived exosomes activate β-catenin signaling in BMSCs, thereby counteracting the inhibitory effects of osteoclast exosomes on BMSC osteogenic differentiation and ameliorating OP in rat models.143
Chemical Factors (Including Pharmaceutical Factors)
Moesin, a protein significantly enriched in H2S-induced exosomes, promotes exosome uptake by MSCs, thereby activating the β-catenin signaling pathway and enhancing osteogenic differentiation.132 H2S facilitates the enrichment of membrane synaptic proteins in BMMSC-derived exosomes, which increases exosomal uptake by MSCs and promotes bone regeneration via β-catenin signaling. Extracellular signal-regulated kinase 1 (ERK1), a mitogen-activated protein kinase (MAPK), is present in exosomes and induces osteoclast differentiation. Guizhi Fuling Decoction (GZFL) reduces bone destruction, modulates the bone tumor microenvironment, and inhibits myeloma progression by suppressing ERK1 activity in myeloma cell-derived exosomes.144
Biological Factors
Irisin, an exercise-induced myokine, exerts bone-protective effects by being transported to osteoblasts via exosomes. It binds to caveolin-1 (Cav1) on osteoblast membranes to promote exosome endocytosis, subsequently forming a FNDC5/irisin-Cav1-AMPKα complex that activates the AMPK pathway. This pathway reduces ferroptosis, enhances osteoblast proliferation, and rescues bone loss Irisin, an exercise-induced myokine, exerts bone-protective effects by being transported to osteoblasts via exosomes. It binds to Cav1 on osteoblast membranes to promote exosome endocytosis, subsequently forming a FNDC5/irisin-Cav1-AMPKα complex that activates the AMPK pathway. This pathway reduces ferroptosis, enhances osteoblast proliferation, and rescues bone loss.145 TNF-α enhances exosomal secretion by upregulating Wnt-3a in adipose-derived stem cell (ASC) exosomes. Wnt-3a synergizes with other TNF-α induced factors (eg, BMP-2) to activate osteogenic genes (eg, type I collagen) in human osteoblasts (HOBs) via exosomal delivery, promoting bone regeneration.93
Physical Factors
Cationic exosomes generated by anchoring cations and anti-biotin proteins to lipid bilayers exhibit enhanced uptake by deep-layer chondrocytes compared to anionic exosomes. Although the precise mechanism remains unclear, this strategy demonstrates potential for targeted exosome therapies in OA.146 Hypoxia alters exosomal protein composition; hypoxia-stimulated BMSCs release EVs enriched with BCL2/adenovirus E1B 19 kDa interacting protein 3 (BNIP3). BNIP3 interacts with annexin A2 (ANXA2), displacing transcription factor EB (TFEB) from the ANXA2-TFEB complex. This promotes TFEB nuclear translocation, activates autophagy/lysosomal genes, mitigates mitochondrial autophagic flux impairment, and attenuates IVDD57 (Figure 4).
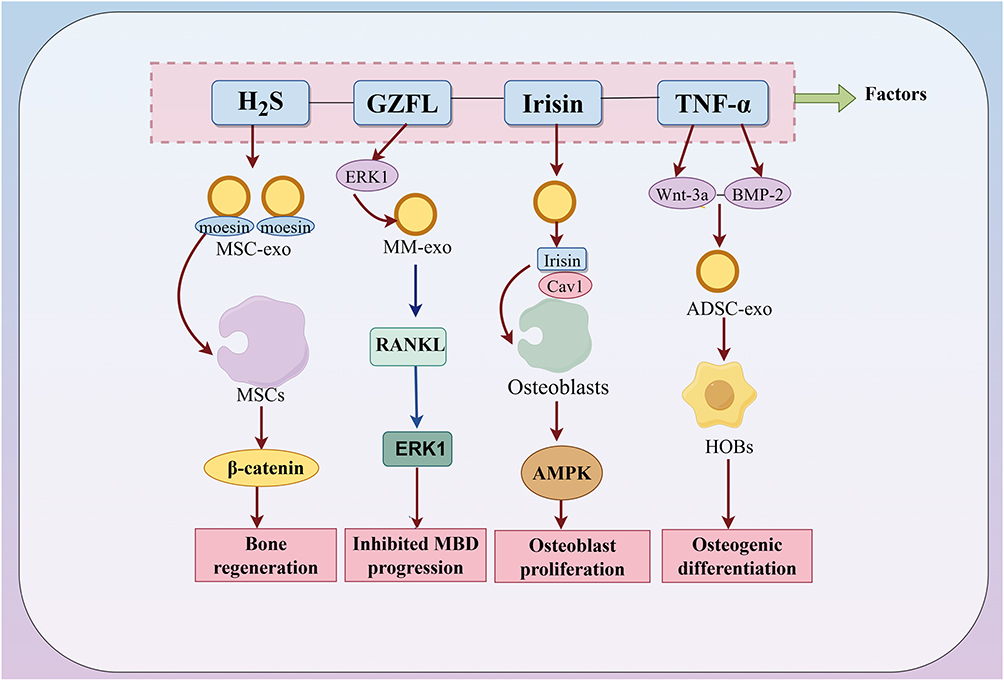 |
Figure 4 Regulation of exosomal proteins by external factors. External factors such as hydrogen sulfide (H2S), Guizhi Fuling Decoction (GZFL), irisin, and TNF-α influence bone metabolism through the modulation of exosomal proteins. In the figure, blue arrows represent negative effects, while red arrows indicate positive effects. |
Discussion
This article provides a systematic examination of the roles of exosomes in orthopedic diseases. We specifically focus on how exosomes-mediated intercellular communication modulates pathological processes in conditions such as osteoarthritis (OA), osteoporosis (OP), osteosarcoma (OS), rheumatoid arthritis (RA), intervertebral disc degeneration (IVDD), osteonecrosis of the femoral head (ONFH), and fractures—diseases that collectively represent a major source of global morbidity. The limitations of conventional surgical and pharmacological treatments, including severe systemic toxicity, adverse side effects, and inconsistent therapeutic efficacy, underscore the urgent need for novel therapeutic strategies.147
Exosomes have emerged as a highly promising avenue in this context. This review synthesizes their therapeutic potential by analyzing how they restore tissue homeostasis through the targeted regulation of four core pathological themes: (1) modulating immune-inflammatory dysregulation, (2) restoring osteogenic-osteoclastic balance, (3) promoting tissue regeneration and matrix repair, and (4) reshaping the tumor microenvironment. Clinical instances of exosome application are incorporated to highlight their translational promise.148–150
Furthermore, acknowledging the inherent challenges in using natural exosomes—such as low yield, heterogeneity, and lack of targeting—this review dedicates significant analysis to strategies that enhance their therapeutic efficacy. We detail how external physical, chemical, biological, and pharmacological factors can be harnessed to precisely modulate the exosome lifecycle. This includes boosting their biogenesis and secretion (eg, via cytokine preconditioning or mechanical stimuli) and engineering their cargo composition (eg, enriching specific miRNAs or proteins), thereby amplifying their therapeutic impact in orthopedic diseases.151
The biogenesis of exosomes is tightly regulated by multifactorial inputs that dictate their pathophysiological roles. External stimuli—including pharmacological agents (eg, epimedium glycosides), inflammatory cytokines (eg, TNF-α), and physical triggers (eg, low-intensity pulsed ultrasound, hypoxia)—alter exosome secretion by modulating multivesicular body (MVB) dynamics.66,78,98,125 For instance, TNF-α enhances exosome release via endosomal acidification inhibition and MVB trafficking regulation,78 while mechanical or hypoxic stimuli directly augment exosome production.86,92,99 These findings highlight strategies to optimize exosome yield for therapeutic applications.
At the pharmacological level, beyond natural compounds, synthetic small molecules and the repurposing of already approved drugs demonstrate significant potential. For instance, anti-rheumatic drugs such as Methotrexate (MTX) and Sulfasalazine (SASP) can alter the proteome of exosomes derived from synovial cells, enhancing their anti-inflammatory properties. Digoxin has been found to improve the arthritic microenvironment by modulating miRNAs within macrophage-derived exosomes. In the future, systematically screening FDA-approved drug libraries to discover their effects on exosome biology represents a promising shortcut to accelerating clinical translation.128–130
In previous studies, safety investigations have been conducted on BMSC-derived EVs for the treatment of combat-related post-traumatic osteoarthritis in humans. Despite limitations such as the small number of participants and the lack of randomized control groups, it cannot be denied that the injection of BMSC extracellular vesicle isolate product effectively alleviated osteoarthritis-related pain with only mild side effects, thereby providing direction for the clinical application of exosomes.152 Although the specific role of exosomes has not been detailed, it still provides confidence in future.
However, uncontrolled biodistribution potentially leading to off-target effects, along with safety concerns arising from the heterogeneity of exosomal cargo—including immunogenicity and unintended pro-inflammatory signaling (eg, amplification of TNF-α in osteoarthritis)—remain critical challenges limiting the clinical translation of exosome-based therapies.92 Engineering approaches, such as surface modification with targeting ligands (eg, magnetic nanoparticles) or cargo engineering (eg, miRNA/Rab-7b enrichment), are being developed to enhance tissue specificity and therapeutic efficacy.103,130,153
Notably, exosome-mediated effects often exhibit biphasic responses: while moderate mechanical loading promotes osteogenesis, excessive stress accelerates cartilage degradation via MMP-13 upregulation, underscoring the need for disease-stage-specific dosing119.
Exosome cargo modulation represents another critical regulatory layer. For example, hydrogen sulfide (H2S) enhances MSC exosome uptake via membrane protein enrichment, promoting bone regeneration.132 However, miRNA pleiotropy complicates their clinical application, necessitating single-cell omics approaches to resolve context-dependent sorting mechanisms and identify disease-specific signatures.154
At the receptor-cell interface, exosome internalization efficiency depends on surface marker recognition (eg, CD44 downregulation in OA chondrocytes may impair targeting). Strategies to improve specificity include magnetic field-guided delivery of iron oxide-labeled exosomes or exploiting receptor-mediated endocytosis pathways (eg, arginine-rich CPPs for membrane fusion enhancement).153 Nevertheless, the spatiotemporal dynamics of exosome-cell interactions remain poorly defined, particularly in heterogeneous patient populations.
In summary, exosome-based therapies require precise control over biosynthesis, cargo composition, and delivery mechanisms. Future research should prioritize resolving single-cell exosome heterogeneity, optimizing engineered exosomal targeting, and establishing standardized protocols for clinical translation.
Conclusion
As key mediators orchestrating intercellular communication within the skeletal system, exosomes have emerged as central players in elucidating pathological mechanisms and advancing therapeutic strategies in orthopedic diseases. Current research into exosome-mediated therapy primarily focuses on restoring tissue homeostasis and promoting repair by strategically modulating pathological cell-cell dialogues. This is achieved through three principal approaches targeting the communication process itself: regulating exosome biogenesis and secretion to influence signal output, enhancing their targeting specificity to ensure precise signal delivery, or engineering their cargo to carry corrective instructions (eg, proteins, miRNAs, mRNAs). The entire lifecycle of an exosome—from its biogenesis and secretion by donor cells to its uptake by recipient cells—constitutes a complete cycle of intercellular signaling. This cycle is susceptible to regulation by various physical, chemical, and biological factors, which can alter the “message” being sent. Therefore, the advancement of exosome-based therapies critically depends on gaining precise control over this communicative process—specifically, by programming the information content (cargo), optimizing the signal release (secretion), and ensuring accurate reception (targeting and uptake)—to achieve the desired therapeutic outcome.
This review has synthesized the role of exosomes as communication vehicles in orthopedics. Their innate biological features—low immunogenicity, biocompatibility, andtropism—make them ideal for delivering intercellular signals in a therapeutic context. However, several barriers related to their nature as communication tools hinder clinical translation: (1) the lack of standardized methods for obtaining consistent “signal carriers” (isolation/production); (2) the need for more sophisticated “address codes” for tissue-specific delivery (targeting); and (3) the inherent “message variation” depending on the donor cell’s state (functional heterogeneity). Furthermore, while research has extensively decoded miRNA-based “messages”, the roles of other molecular languages within exosomes, such as lipids, mRNAs, and other non-coding RNAs, remain poorly translated, representing a significant gap in fully understanding this communication system.
Overcoming these challenges is fundamental to evolving exosomes from promising biological messengers into reliable therapeutic agents. Future research must prioritize: establishing robust manufacturing protocols to ensure batch-to-batch consistency of the communication vehicle; developing next-generation engineering techniques to install precise targeting directives and load optimized cargo; and implementing integrated multi-omics analyses to fully decipher the complex language of exosomal communication. By systematically addressing these bottlenecks, we can transition from merely observing intercellular crosstalk to actively and precisely reprogramming it, ultimately harnessing the power of exosome-mediated communication to develop transformative treatments for orthopedic diseases.
Data Sharing Statement
No data was used for the research described in the article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Science Foundation of China (grant number 82474240); Zhejiang Provincial Medical and Health Science and Technology (grant number 2024KY1223); Research Project of Zhejiang Chinese Medical University (grant number 2023JKZKTS34).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Laucis NC, Hays RD, Bhattacharyya T. Scoring the SF-36 in orthopaedics: a brief guide. J Bone Joint Surg Am. 2015;97(19):1628–25. doi:10.2106/jbjs.O.00030
2. Jiang F, Lu C, Zeng Z, Sun Z, Qiu Y. Global burden of disease for musculoskeletal disorders in all age groups, from 2024 to 2050, and a bibliometric-based survey of the status of research in geriatrics, geriatric orthopedics, and geriatric orthopedic diseases. J Orthop Surg Res. 2025;20(1):179. doi:10.1186/s13018-025-05580-y
3. Steinmetz JD, Culbreth GT, Haile LM, et al. Global, regional, and national burden of osteoarthritis, 1990-2020 and projections to 2050: a systematic analysis for the Global Burden of Disease Study 2021. Lancet Rheumatol. 2023;5(9):e508–e522. doi:10.1016/s2665-9913(23)00163-7
4. Long H, Liu Q, Yin H, et al. Prevalence trends of site-specific osteoarthritis from 1990 to 2019: findings from the global burden of disease study 2019. Arthritis Rheumatol. 2022;74(7):1172–1183. doi:10.1002/art.42089
5. Zhang Z, Gao X, Liu S, et al. Global, regional, and national epidemiology of rheumatoid arthritis among people aged 20-54 years from 1990 to 2021. Sci Rep. 2025;15(1):10736. doi:10.1038/s41598-025-92150-1
6. Raisz LG. Pathogenesis of osteoporosis: concepts, conflicts, and prospects. J Clin Invest. 2005;115(12):3318–3325. doi:10.1172/jci27071
7. Petek D, Hannouche D, Suva D. Osteonecrosis of the femoral head: pathophysiology and current concepts of treatment. EFORT Open Rev. 2019;4(3):85–97. doi:10.1302/2058-5241.4.180036
8. Su Z, Fang X, Duan H. The paradoxical role of stem cells in osteosarcoma: from pathogenesis to therapeutic breakthroughs. Front Oncol. 2025;15:1643491. doi:10.3389/fonc.2025.1643491
9. Einhorn TA, Gerstenfeld LC. Fracture healing: mechanisms and interventions. Nat Rev Rheumatol. 2015;11(1):45–54. doi:10.1038/nrrheum.2014.164
10. Song C, Zhou Y, Cheng K, et al. Cellular senescence - Molecular mechanisms of intervertebral disc degeneration from an immune perspective. Biomed Pharmacother. 2023;162:114711. doi:10.1016/j.biopha.2023.114711
11. Wang Y, Cheng H, Wang T, Zhang K, Zhang Y, Kang X. Oxidative stress in intervertebral disc degeneration: molecular mechanisms, pathogenesis and treatment. Cell Prolif. 2023;56(9):e13448. doi:10.1111/cpr.13448
12. Gau YC, Yeh TJ, Hsu CM, Hsiao SY, Hsiao HH. Pathogenesis and treatment of myeloma-related bone disease. Int J Mol Sci. 2022;23(6). doi:10.3390/ijms23063112
13. Eguia A, Bagán-Debón L, Cardona F. Review and update on drugs related to the development of osteonecrosis of the jaw. Med Oral Patol Oral Cir Bucal. 2020;25(1):e71–e83. doi:10.4317/medoral.23191
14. Carrle D, Bielack SS. Current strategies of chemotherapy in osteosarcoma. Int Orthop. 2006;30(6):445–451. doi:10.1007/s00264-006-0192-x
15. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200(4):373–383. doi:10.1083/jcb.201211138
16. Kowal J, Arras G, Colombo M, et al. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc Natl Acad Sci U S A. 2016;113(8):E968–77. doi:10.1073/pnas.1521230113
17. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi:10.1080/20013078.2018.1535750
18. Fu YJ, Jian J, Liu C, et al. Application of stem cell-derived exosomes in bone and joint diseases: recent advances enabled by diverse carrier technologies. World J Stem Cells. 2025;17(11):113631. doi:10.4252/wjsc.v17.i11.113631
19. Chu CH, Lee RP, Wu WT, Chen IH, Yeh KT, Wang CC. Advancing osteoarthritis treatment: the therapeutic potential of mesenchymal stem cell-derived exosomes and biomaterial integration. Biomedicines. 2024;12(11). doi:10.3390/biomedicines12112478
20. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478). doi:10.1126/science.aau6977
21. Krylova SV, Feng D. The machinery of exosomes: biogenesis, release, and uptake. Int J Mol Sci. 2023;24(2). doi:10.3390/ijms24021337
22. Han QF, Li WJ, Hu KS, et al. Exosome biogenesis: machinery, regulation, and therapeutic implications in cancer. Mol Cancer. 2022;21(1):207. doi:10.1186/s12943-022-01671-0
23. Tian J, Han Z, Song D, et al. Engineered exosome for drug delivery: recent development and clinical applications. Int J Nanomed. 2023;18:7923–7940. doi:10.2147/ijn.S444582
24. He C, Zheng S, Luo Y, Wang B. Exosome theranostics: biology and translational medicine. Theranostics. 2018;8(1):237–255. doi:10.7150/thno.21945
25. Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: trafficking, sorting, and function. Genomics Proteomics Bioinf. 2015;13(1):17–24. doi:10.1016/j.gpb.2015.02.001
26. Xu K, Huang RQ, Wen R, Yang Y, Cheng Y, Chang B. The role of Clec11a in bone construction and remodeling. Front Endocrinol. 2024;15:1429567. doi:10.3389/fendo.2024.1429567
27. Stegen S, Carmeliet G. Metabolic regulation of skeletal cell fate and function. Nat Rev Endocrinol. 2024;20(7):399–413. doi:10.1038/s41574-024-00969-x
28. Kanda K, Asawa Y, Inaki R, Fujihara Y, Hoshi K, Hikita A. Requirement of direct contact between chondrocytes and macrophages for the maturation of regenerative cartilage. Sci Rep. 2021;11(1):22476. doi:10.1038/s41598-021-01437-6
29. Ye QY, Cui Y, Wang HY, et al. Exosomal communication: a pivotal regulator of bone homeostasis and a potential therapeutic target. Front Pharmacol. 2024;15:1516125. doi:10.3389/fphar.2024.1516125
30. Wu Y, Li J, Zeng Y, et al. Exosomes rewire the cartilage microenvironment in osteoarthritis: from intercellular communication to therapeutic strategies. Int J Oral Sci. 2022;14(1):40. doi:10.1038/s41368-022-00187-z
31. Tsukasaki M, Takayanagi H. Osteoimmunology: evolving concepts in bone-immune interactions in health and disease. Nat Rev Immunol. 2019;19(10):626–642. doi:10.1038/s41577-019-0178-8
32. Hu K, Song M, Song T, Jia X, Song Y. Osteoimmunology in osteoarthritis: unraveling the interplay of immunity, inflammation, and joint degeneration. J Inflamm Res. 2025;18:4121–4142. doi:10.2147/jir.S514002
33. Zhang K, Wang Z, He J, et al. Mechanisms of synovial macrophage polarization in osteoarthritis pathogenesis and their therapeutic implications. Front Immunol. 2025;16:1637731. doi:10.3389/fimmu.2025.1637731
34. Wang L, He C. Nrf2-mediated anti-inflammatory polarization of macrophages as therapeutic targets for osteoarthritis. Front Immunol. 2022;13:967193. doi:10.3389/fimmu.2022.967193
35. Semenistaja S, Skuja S, Kadisa A, Groma V. Healthy and osteoarthritis-affected joints facing the cellular crosstalk. Int J Mol Sci. 2023;24(4). doi:10.3390/ijms24044120
36. Zheng Y, Wei K, Jiang P, et al. Macrophage polarization in rheumatoid arthritis: signaling pathways, metabolic reprogramming, and crosstalk with synovial fibroblasts. Front Immunol. 2024;15:1394108. doi:10.3389/fimmu.2024.1394108
37. Torres HM, Arnold KM, Oviedo M, Westendorf JJ, Weaver SR. Inflammatory processes affecting bone health and repair. Curr Osteoporos Rep. 2023;21(6):842–853. doi:10.1007/s11914-023-00824-4
38. Vasconcelos DM, Ribeiro-da-Silva M, Mateus A, et al. Immune response and innervation signatures in aseptic hip implant loosening. J Transl Med. 2016;14(1):205. doi:10.1186/s12967-016-0950-5
39. Morrison TA, Figgie M, Miller AO, Goodman SM. Periprosthetic joint infection in patients with inflammatory joint disease: a review of risk factors and current approaches to diagnosis and management. Hss J. 2013;9(2):183–194. doi:10.1007/s11420-013-9338-8
40. Lan S, Zhang C. Roles of exosomes in immune regulation of osteoarthritis and their applications in inflammation repair. Front Immunol. 2025;16:1611718. doi:10.3389/fimmu.2025.1611718
41. Zhang J, Rong Y, Luo C, Cui W. Bone marrow mesenchymal stem cell-derived exosomes prevent osteoarthritis by regulating synovial macrophage polarization. Aging. 2020;12(24):25138–25152. doi:10.18632/aging.104110
42. Wang Z, Zhang C, Meng J, et al. A targeted exosome therapeutic confers both CfDNA scavenging and macrophage polarization for ameliorating rheumatoid arthritis. Adv Mater. 2023;35(48):e2302503. doi:10.1002/adma.202302503
43. Zhang Z, Ma T, Liu Q, et al. Exosomes derived from bone marrow mesenchymal stem cells encapsulated in M2 macrophage cell membrane targeted to inhibit joint periprosthetic inflammation. ACS Appl Mater Interfaces. 2025;17(15):22279–22292. doi:10.1021/acsami.4c22304
44. Hu L, Chen W, Qian A, Li YP. Wnt/β-catenin signaling components and mechanisms in bone formation, homeostasis, and disease. Bone Res. 2024;12(1):39. doi:10.1038/s41413-024-00342-8
45. Yahara Y, Nguyen T, Ishikawa K, Kamei K, Alman BA. The origins and roles of osteoclasts in bone development, homeostasis and repair. Development. 2022;149(8). doi:10.1242/dev.199908
46. Rosen CJ, Kessenich CR. The pathophysiology and treatment of postmenopausal osteoporosis. An evidence-based approach to estrogen replacement therapy. Endocrinol Metab Clin North Am. 1997;26(2):295–311. doi:10.1016/s0889-8529(05)70248-6
47. Wang C, Zhang H, Wang X, Ma X, Zhang J, Zhang Y. Targeting BRD4 to attenuate RANKL-induced osteoclast activation and bone erosion in rheumatoid arthritis. Mol Cell Biochem. 2025;480(3):1669–1684. doi:10.1007/s11010-024-05073-2
48. Zhang L, Jiao G, Ren S, et al. Exosomes from bone marrow mesenchymal stem cells enhance fracture healing through the promotion of osteogenesis and angiogenesis in a rat model of nonunion. Stem Cell Res Ther. 2020;11(1):38. doi:10.1186/s13287-020-1562-9
49. Mi B, Chen L, Xiong Y, et al. Osteoblast/osteoclast and immune cocktail therapy of an exosome/drug delivery multifunctional hydrogel accelerates fracture repair. ACS Nano. 2022;16(1):771–782. doi:10.1021/acsnano.1c08284
50. Yang JG, Sun B, Wang Z, et al. Exosome-targeted delivery of METTL14 regulates NFATc1 m6A methylation levels to correct osteoclast-induced bone resorption. Cell Death Dis. 2023;14(11):738. doi:10.1038/s41419-023-06263-4
51. Steltzer SS, Abraham AC, Killian ML. Interfacial tissue regeneration with bone. Curr Osteoporos Rep. 2024;22(2):290–298. doi:10.1007/s11914-024-00859-1
52. Cardoneanu A, Macovei LA, Burlui AM, et al. Temporomandibular joint osteoarthritis: pathogenic mechanisms involving the cartilage and subchondral bone, and potential therapeutic strategies for joint regeneration. Int J Mol Sci. 2022;24(1). doi:10.3390/ijms24010171
53. Henle P, Zimmermann G, Weiss S. Matrix metalloproteinases and failed fracture healing. Bone. 2005;37(6):791–798. doi:10.1016/j.bone.2005.06.015
54. Zhao DW, Zhang J, Chen C, et al. Rejuvenation modulation of nucleus pulposus progenitor cells reverses senescence-associated intervertebral disc degeneration. Adv Mater. 2025;37(7):e2409979. doi:10.1002/adma.202409979
55. Zhong T, Gao N, Niu H, et al. Targeted delivery of engineered extracellular vesicles to simultaneously promote vascularization and muscle regeneration in ischemic limbs. J Control Release. 2025;384:113938. doi:10.1016/j.jconrel.2025.113938
56. Tao SC, Yuan T, Zhang YL, Yin WJ, Guo SC, Zhang CQ. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics. 2017;7(1):180–195. doi:10.7150/thno.17133
57. Jin Y, Wu O, Chen Q, et al. Hypoxia-preconditioned BMSC-derived exosomes induce mitophagy via the BNIP3-ANAX2 axis to alleviate intervertebral disc degeneration. Adv Sci. 2024;11(34):e2404275. doi:10.1002/advs.202404275
58. Song C, Tong T, Dai B, et al. Osteoimmunology in bone malignancies: a symphony with evil. J Natl Cancer Cent. 2024;4(4):354–368. doi:10.1016/j.jncc.2024.09.001
59. Esposito M, Kang Y. Targeting tumor-stromal interactions in bone metastasis. Pharmacol Ther. 2014;141(2):222–233. doi:10.1016/j.pharmthera.2013.10.006
60. Zhao M, Huang J, Gui K, et al. The downregulation of miR-144 is associated with the growth and invasion of osteosarcoma cells through the regulation of TAGLN expression. Int J Mol Med. 2014;34(6):1565–1572. doi:10.3892/ijmm.2014.1963
61. Huang X, Wu W, Jing D, et al. Engineered exosome as targeted lncRNA MEG3 delivery vehicles for osteosarcoma therapy. J Control Release. 2022;343:107–117. doi:10.1016/j.jconrel.2022.01.026
62. Wu J, Wu J, Xiang W, et al. Engineering exosomes derived from TNF-α preconditioned IPFP-MSCs enhance both yield and therapeutic efficacy for osteoarthritis. J Nanobiotechnol. 2024;22(1):555. doi:10.1186/s12951-024-02795-9
63. Blanc L, Vidal M. New insights into the function of Rab GTPases in the context of exosomal secretion. Small GTPases. 2018;9(1–2):95–106. doi:10.1080/21541248.2016.1264352
64. Xu S, Cao B, Xuan G, et al. Function and regulation of Rab GTPases in cancers. Cell Biol Toxicol. 2024;40(1):28. doi:10.1007/s10565-024-09866-5
65. Ostrowski M, Carmo NB, Krumeich S, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol. 2010;12(1):19–30; suppp1–13. doi:10.1038/ncb2000
66. Zhang Y, Zhang M, Li M, Miao M, Shou D, Tong P. Icariin-enhanced osteoclast-derived exosomes promote repair of infected bone defects by regulating osteoclast and osteoblast communication. Int J Nanomed. 2024;19:12389–12407. doi:10.2147/ijn.S483621
67. Mosesso N, Nagel MK, Isono E. Ubiquitin recognition in endocytic trafficking - with or without ESCRT-0. J Cell Sci. 2019;132(16). doi:10.1242/jcs.232868
68. Kostelansky MS, Sun J, Lee S, et al. Structural and functional organization of the ESCRT-I trafficking complex. Cell. 2006;125(1):113–126. doi:10.1016/j.cell.2006.01.049
69. Hierro A, Sun J, Rusnak AS, et al. Structure of the ESCRT-II endosomal trafficking complex. Nature. 2004;431(7005):221–225. doi:10.1038/nature02914
70. Ghossoub R, Lembo F, Rubio A, et al. Syntenin-ALIX exosome biogenesis and budding into multivesicular bodies are controlled by ARF6 and PLD2. Nat Commun. 2014;5:3477. doi:10.1038/ncomms4477
71. Kenific CM, Zhang H, Lyden D. An exosome pathway without an ESCRT. Cell Res. 2021;31(2):105–106. doi:10.1038/s41422-020-00418-0
72. Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319(5867):1244–1247. doi:10.1126/science.1153124
73. Cai H, Reinisch K, Ferro-Novick S. Coats, tethers, Rabs, and SNAREs work together to mediate the intracellular destination of a transport vesicle. Dev Cell. 2007;12(5):671–682. doi:10.1016/j.devcel.2007.04.005
74. Matsui T, Sakamaki Y, Nakashima S, Fukuda M. Rab39 and its effector UACA regulate basolateral exosome release from polarized epithelial cells. Cell Rep. 2022;39(9):110875. doi:10.1016/j.celrep.2022.110875
75. Villarroya-Beltri C, Baixauli F, Mittelbrunn M, et al. ISGylation controls exosome secretion by promoting lysosomal degradation of MVB proteins. Nat Commun. 2016;7:13588. doi:10.1038/ncomms13588
76. Savina A, Furlán M, Vidal M, Colombo MI. Exosome release is regulated by a calcium-dependent mechanism in K562 cells. J Biol Chem. 2003;278(22):20083–20090. doi:10.1074/jbc.M301642200
77. Jahn R, Cafiso DC, Tamm LK. Mechanisms of SNARE proteins in membrane fusion. Nat Rev Mol Cell Biol. 2024;25(2):101–118. doi:10.1038/s41580-023-00668-x
78. Choezom D, Gross JC. Neutral sphingomyelinase 2 controls exosome secretion by counteracting V-ATPase-mediated endosome acidification. J Cell Sci. 2022;135(5). doi:10.1242/jcs.259324
79. Duraes FV, Niven J, Dubrot J, Hugues S, Gannagé M. Macroautophagy in endogenous processing of self- and pathogen-derived antigens for MHC Class II presentation. Front Immunol. 2015;6:459. doi:10.3389/fimmu.2015.00459
80. Galluzzi L, Baehrecke EH, Ballabio A, et al. Molecular definitions of autophagy and related processes. EMBO J. 2017;36(13):1811–1836. doi:10.15252/embj.201796697
81. Shelke GV, Williamson CD, Jarnik M, Bonifacino JS. Inhibition of endolysosome fusion increases exosome secretion. J Cell Biol. 2023;222(6). doi:10.1083/jcb.202209084
82. Baixauli F, López-Otín C, Mittelbrunn M. Exosomes and autophagy: coordinated mechanisms for the maintenance of cellular fitness. Front Immunol. 2014;5:403. doi:10.3389/fimmu.2014.00403
83. de Medina P, Bunay J, Poirot M, Record M, Silvente-Poirot S. Targeting NR1H/liver X receptor with dendrogenin A differentiates tumor cells to activate a new secretory pathway releasing immunogenic anti-tumor vesicles enriched in LC3-II-associated exosomes. Autophagy. 2023;19(3):1036–1038. doi:10.1080/15548627.2022.2116175
84. Zou W, Lai M, Zhang Y, et al. Exosome release is regulated by mTORC1. Adv Sci. 2019;6(3):1801313. doi:10.1002/advs.201801313
85. Hu SQ, Zou YP, Jiang YQ, et al. Scutellarin-mediated autophagy activates exosome release of rat nucleus pulposus cells by positively regulating Rab8a via the PI3K/PTEN/Akt pathway. Cell Biol Int. 2022;46(10):1588–1603. doi:10.1002/cbin.11838
86. Xia P, Wang Q, Song J, et al. Low-intensity pulsed ultrasound enhances the efficacy of bone marrow-derived MSCs in osteoarthritis cartilage repair by regulating autophagy-mediated exosome release. Cartilage. 2022;13(2):19476035221093060. doi:10.1177/19476035221093060
87. Lei LM, Li FX, Lin X, et al. Cold exposure-induced plasma exosomes impair bone mass by inhibiting autophagy. J Nanobiotechnol. 2024;22(1):361. doi:10.1186/s12951-024-02640-z
88. Yuan W, Liu J, Zhang Z, et al. Strontium-Alix interaction enhances exosomal miRNA selectively loading in synovial MSCs for temporomandibular joint osteoarthritis treatment. Int J Oral Sci. 2025;17(1):6. doi:10.1038/s41368-024-00329-5
89. Yang YH, Wen CS, Kuo YL, et al. GuiLu-ErXian Glue extract promotes mesenchymal stem cells (MSC)-Induced chondrogenesis via exosomes release and delays aging in the MSC senescence process. J Ethnopharmacol. 2023;317:116784. doi:10.1016/j.jep.2023.116784
90. Liu J, Shi J, Niu S, et al. Genistein alleviates rheumatoid arthritis by inhibiting fibroblast-like synovial exosome secretion regulated by the Rab27/nSMase2/Mfge8 pathway. Food Funct. 2025;16(4):1407–1422. doi:10.1039/d4fo05730a
91. Faria LP, Arana-Chavez VE, Holliday LS. Alendronate alters the release of EVs by Raw 264.7 osteoclasts. J Histochem Cytochem. 2025;73(9–10):371–377. doi:10.1369/00221554251379540
92. Liu W, Li L, Rong Y, et al. Hypoxic mesenchymal stem cell-derived exosomes promote bone fracture healing by the transfer of miR-126. Acta Biomater. 2020;103:196–212. doi:10.1016/j.actbio.2019.12.020
93. Lu Z, Chen Y, Dunstan C, Roohani-Esfahani S, Zreiqat H. Priming adipose stem cells with tumor necrosis factor-alpha preconditioning potentiates their exosome efficacy for bone regeneration. Tissue Eng Part A. 2017;23(21–22):1212–1220. doi:10.1089/ten.tea.2016.0548
94. Kato T, Miyaki S, Ishitobi H, et al. Exosomes from IL-1β stimulated synovial fibroblasts induce osteoarthritic changes in articular chondrocytes. Arthritis Res Ther. 2014;16(4):R163. doi:10.1186/ar4679
95. Meng S, Tang C, Deng M, et al. Tropoelastin-pretreated exosomes from adipose-derived stem cells improve the synthesis of cartilage matrix and alleviate osteoarthritis. J Funct Biomater. 2023;14(4). doi:10.3390/jfb14040203
96. Cha KY, Cho W, Park S, et al. Generation of bioactive MSC-EVs for bone tissue regeneration by tauroursodeoxycholic acid treatment. J Control Release. 2023;354:45–56. doi:10.1016/j.jconrel.2022.12.053
97. Wang D, Wang J, Zhou J, Zheng X. The role of adenosine receptor A2A in the regulation of macrophage exosomes and vascular endothelial cells during bone healing. J Inflamm Res. 2021;14:4001–4017. doi:10.2147/jir.S324232
98. Bratengeier C, Johansson L, Liszka A, Bakker AD, Hallbeck M, Fahlgren A. Mechanical loading intensities affect the release of extracellular vesicles from mouse bone marrow-derived hematopoietic progenitor cells and change their osteoclast-modulating effect. FASEB J. 2024;38(1):e23323. doi:10.1096/fj.202301520R
99. Morrell AE, Brown GN, Robinson ST, et al. Mechanically induced Ca(2+) oscillations in osteocytes release extracellular vesicles and enhance bone formation. Bone Res. 2018;6:6. doi:10.1038/s41413-018-0007-x
100. Lin Z, Zheng J, Chen J, Chen M, Dong S. Antiosteoporosis effect and possible mechanisms of the ingredients of fructus psoraleae in animal models of osteoporosis: a preclinical systematic review and meta-analysis. Oxid Med Cell Longev. 2021;2021:2098820. doi:10.1155/2021/2098820
101. Yu J, Wu X, Zhang W, et al. Effect of psoralen on the regulation of osteogenic differentiation induced by periodontal stem cell-derived exosomes. Hum Cell. 2023;36(4):1389–1402. doi:10.1007/s13577-023-00918-2
102. Jin X, Zhu L, Li X, et al. Low‑molecular weight fucoidan inhibits the differentiation of osteoclasts and reduces osteoporosis in ovariectomized rats. Mol Med Rep. 2017;15(2):890–898. doi:10.3892/mmr.2016.6062
103. Lou C, Jiang H, Lin Z, et al. MiR-146b-5p enriched bioinspired exosomes derived from fucoidan-directed induction mesenchymal stem cells protect chondrocytes in osteoarthritis by targeting TRAF6. J Nanobiotechnol. 2023;21(1):486. doi:10.1186/s12951-023-02264-9
104. Dong S, Xu G, Li X, et al. Exosomes derived from quercetin-treated bone marrow derived mesenchymal stem cells inhibit the progression of osteoarthritis through delivering miR-124-3p to chondrocytes. DNA Cell Biol. 2024;43(2):85–94. doi:10.1089/dna.2023.0341
105. Geng Z, Sun T, Yu J, et al. Cinobufagin suppresses lipid peroxidation and inflammation in osteoporotic mice by promoting the delivery of miR-3102-5p by macrophage-derived exosomes. Int J Nanomed. 2024;19:10497–10512. doi:10.2147/ijn.S483849
106. Wu P, Jiao F, Huang H, et al. Morinda officinalis polysaccharide enable suppression of osteoclastic differentiation by exosomes derived from rat mesenchymal stem cells. Pharm Biol. 2022;60(1):1303–1316. doi:10.1080/13880209.2022.2093385
107. Wang L, Pan Y, Liu M, et al. Wen-Shen-Tong-Luo-Zhi-Tong Decoction regulates bone-fat balance in osteoporosis by adipocyte-derived exosomes. Pharm Biol. 2023;61(1):568–580. doi:10.1080/13880209.2023.2190773
108. Zhang H, Zhou C, Zhang Z, et al. Integration of network pharmacology and experimental validation to explore the pharmacological mechanisms of zhuanggu busui formula against osteoporosis. Front Endocrinol. 2021;12:841668. doi:10.3389/fendo.2021.841668
109. Chen W, Zheng H, Liao Q, et al. Zhuang-Gu-Fang promotes osteoblast differentiation via myoblasts and myoblast-derived exosomal miRNAs: miR-5100, miR-126a-3p, miR-450b-5p, and miR-669a-5p. Phytomedicine. 2024;130:155718. doi:10.1016/j.phymed.2024.155718
110. Jia H, Duan L, Yu P, Zhou Y, Liu R, Wang H. Digoxin ameliorates joint inflammatory microenvironment by downregulating synovial macrophage M1-like-polarization and its-derived exosomal miR-146b-5p/Usp3&Sox5 axis. Int Immunopharmacol. 2022;111:109135. doi:10.1016/j.intimp.2022.109135
111. Li X, Zhou Y, Chen X, et al. Semi-synthetic chondroitin sulfate CS-semi5 upregulates miR-122-5p, conferring a therapeutic effect on osteoarthritis via the p38/MMP13 pathway. Acta Pharm Sin B. 2024;14(8):3528–3542. doi:10.1016/j.apsb.2024.05.016
112. Shao L, Ding L, Li W, et al. Let-7a-5p derived from parathyroid hormone (1-34)-preconditioned BMSCs exosomes delays the progression of osteoarthritis by promoting chondrocyte proliferation and migration. Stem Cell Res Ther. 2025;16(1):299. doi:10.1186/s13287-025-04416-0
113. Zhu Y, Zhao S, Cheng L, et al. Mg(2+) -mediated autophagy-dependent polarization of macrophages mediates the osteogenesis of bone marrow stromal stem cells by interfering with macrophage-derived exosomes containing miR-381. J Orthop Res. 2022;40(7):1563–1576. doi:10.1002/jor.25189
114. Yin S, Lin S, Xu J, Yang G, Chen H, Jiang X. Dominoes with interlocking consequences triggered by zinc: involvement of microelement-stimulated MSC-derived exosomes in senile osteogenesis and osteoclast dialogue. J Nanobiotechnol. 2023;21(1):346. doi:10.1186/s12951-023-02085-w
115. Liu L, Yu F, Li L, et al. Bone marrow stromal cells stimulated by strontium-substituted calcium silicate ceramics: release of exosomal miR-146a regulates osteogenesis and angiogenesis. Acta Biomater. 2021;119:444–457. doi:10.1016/j.actbio.2020.10.038
116. Liu L, Yu F, Chen L, Xia L, Wu C, Fang B. Lithium-containing biomaterials stimulate cartilage repair through bone marrow stromal cells-derived exosomal miR-455-3p and Histone H3 acetylation. Adv Healthc Mater. 2023;12(11):e2202390. doi:10.1002/adhm.202202390
117. Wu Z, He D, Li H. Bioglass enhances the production of exosomes and improves their capability of promoting vascularization. Bioact Mater. 2021;6(3):823–835. doi:10.1016/j.bioactmat.2020.09.011
118. Tian J, Chen W, Xiong Y, et al. Small extracellular vesicles derived from hypoxic preconditioned dental pulp stem cells ameliorate inflammatory osteolysis by modulating macrophage polarization and osteoclastogenesis. Bioact Mater. 2023;22:326–342. doi:10.1016/j.bioactmat.2022.10.001
119. Zhu Y, Li Y, Cao Z, et al. Mechanically strained osteocyte-derived exosomes contained miR-3110-5p and miR-3058-3p and promoted osteoblastic differentiation. Biomed Eng Online. 2024;23(1):44. doi:10.1186/s12938-024-01237-9
120. Wang X, Li X, Li J, et al. Mechanical loading stimulates bone angiogenesis through enhancing type H vessel formation and downregulating exosomal miR-214-3p from bone marrow-derived mesenchymal stem cells. FASEB J. 2021;35(1):e21150. doi:10.1096/fj.202001080RR
121. Wu D, Chang X, Tian J, et al. Bone mesenchymal stem cells stimulation by magnetic nanoparticles and a static magnetic field: release of exosomal miR-1260a improves osteogenesis and angiogenesis. J Nanobiotechnol. 2021;19(1):209. doi:10.1186/s12951-021-00958-6
122. Kim M, Shin DI, Choi BH, Min BH. Exosomes from IL-1β-Primed mesenchymal stem cells Inhibited IL-1β- and TNF-α-Mediated inflammatory responses in osteoarthritic SW982 cells. Tissue Eng Regen Med. 2021;18(4):525–536. doi:10.1007/s13770-020-00324-x
123. Wang R, Xu B, Xu H. TGF-β1 promoted chondrocyte proliferation by regulating Sp1 through MSC-exosomes derived miR-135b. Cell Cycle. 2018;17(24):2756–2765. doi:10.1080/15384101.2018.1556063
124. Kim O, Tran PT, Gal M, et al. RAS‑stimulated release of exosomal miR‑494‑3p promotes the osteolytic bone metastasis of breast cancer cells. Int J Mol Med. 2023;52(3). doi:10.3892/ijmm.2023.5287
125. Qin Y, Peng Y, Zhao W, et al. Myostatin inhibits osteoblastic differentiation by suppressing osteocyte-derived exosomal microRNA-218: a novel mechanism in muscle-bone communication. J Biol Chem. 2017;292(26):11021–11033. doi:10.1074/jbc.M116.770941
126. Duan P, Yu YL, Cheng YN, et al. Exosomal miR-1a-3p derived from glucocorticoid-stimulated M1 macrophages promotes the adipogenic differentiation of BMSCs in glucocorticoid-associated osteonecrosis of the femoral head by targeting Cebpz. J Nanobiotechnol. 2024;22(1):648. doi:10.1186/s12951-024-02923-5
127. Yang Z, Li X, Gan X, et al. Hydrogel armed with Bmp2 mRNA-enriched exosomes enhances bone regeneration. J Nanobiotechnol. 2023;21(1):119. doi:10.1186/s12951-023-01871-w
128. Friedman B, Cronstein B. Methotrexate mechanism in treatment of rheumatoid arthritis. Joint Bone Spine. 2019;86(3):301–307. doi:10.1016/j.jbspin.2018.07.004
129. Plosker GL, Croom KF. Sulfasalazine: a review of its use in the management of rheumatoid arthritis. Drugs. 2005;65(13):1825–1849. doi:10.2165/00003495-200565130-00008
130. Tsuno H, Suematsu N, Sato T, et al. Effects of methotrexate and salazosulfapyridine on protein profiles of exosomes derived from a human synovial sarcoma cell line of SW982. Proteomics Clin Appl. 2016;10(2):164–171. doi:10.1002/prca.201500064
131. Song Y, Wu S, Zhang R, Zhong Q, Zhang X, Sun X. Therapeutic potential of hydrogen sulfide in osteoarthritis development. Front Pharmacol. 2024;15:1336693. doi:10.3389/fphar.2024.1336693
132. Zhou YK, Han CS, Zhu ZL, et al. M2 exosomes modified by hydrogen sulfide promoted bone regeneration by moesin mediated endocytosis. Bioact Mater. 2024;31:192–205. doi:10.1016/j.bioactmat.2023.08.006
133. Chen C, Wang B, Zhao X, et al. Lithium promotes osteogenesis via Rab11a-facilitated exosomal Wnt10a secretion and β-Catenin signaling activation. ACS Appl Mater Interfaces. 2024;16(24):30793–30809. doi:10.1021/acsami.4c04199
134. Aswani BS, Hegde M, Vishwa R, et al. Tackling exosome and nuclear receptor interaction: an emerging paradigm in the treatment of chronic diseases. Mil Med Res. 2024;11(1):67. doi:10.1186/s40779-024-00564-1
135. Wang Y, Xiao T, Zhao C, Li G. The regulation of exosome generation and function in physiological and pathological processes. Int J Mol Sci. 2023;25(1). doi:10.3390/ijms25010255
136. Wang Y, Sun L, Dong Z, et al. Targeted inhibition of ferroptosis in bone marrow mesenchymal stem cells by engineered exosomes alleviates bone loss in smoking-related osteoporosis. Mater Today Bio. 2025;31:101501. doi:10.1016/j.mtbio.2025.101501
137. Xu N, Cui G, Zhao S, et al. Therapeutic effects of mechanical stress-induced C2C12-Derived exosomes on glucocorticoid-induced osteoporosis through miR-92a-3p/PTEN/AKT signaling pathway. Int J Nanomed. 2023;18:7583–7603. doi:10.2147/ijn.S435301
138. Cai WW, Gao Y, Cheng JW, et al. Berberine modulates the immunometabolism and differentiation of CD4(+) T cells alleviating experimental arthritis by suppression of M1-exo-miR155. Phytomedicine. 2024;124:155255. doi:10.1016/j.phymed.2023.155255
139. Zhu W, Zhang F, Lu J, et al. The analysis of Modified Qing’ E Formula on the differential expression of exosomal miRNAs in the femoral head bone tissue of mice with steroid-induced ischemic necrosis of femoral head. Front Endocrinol. 2022;13:954778. doi:10.3389/fendo.2022.954778
140. Li N, Chen Z, Feng W, et al. Triptolide improves chondrocyte proliferation and secretion via down-regulation of miR-221 in synovial cell exosomes. Phytomedicine. 2022;107:154479. doi:10.1016/j.phymed.2022.154479
141. Xu Q, Cui Y, Luan J, Zhou X, Li H, Han J. Exosomes from C2C12 myoblasts enhance osteogenic differentiation of MC3T3-E1pre-osteoblasts by delivering miR-27a-3p. Biochem Biophys Res Commun. 2018;498(1):32–37. doi:10.1016/j.bbrc.2018.02.144
142. Zhao Y, Chen L, Jiang S, et al. Exosomes derived from MSCs exposed to hypoxic and inflammatory environments slow intervertebral disc degeneration by alleviating the senescence of nucleus pulposus cells through epigenetic modifications. Bioact Mater. 2025;49:515–530. doi:10.1016/j.bioactmat.2025.02.046
143. Zhang X, Meng S, Xu Y, et al. New Qiangguyin activates Wnt/β-catenin pathway by down-regulating Notum to improve osteoporosis in rats. J Ethnopharmacol. 2025;347:119745. doi:10.1016/j.jep.2025.119745
144. Yu M, Zhang J, Fu J, Li S, Cui X. Guizhi Fuling decoction protects against bone destruction via suppressing exosomal ERK1 in multiple myeloma. Phytomedicine. 2025;140:156627. doi:10.1016/j.phymed.2025.156627
145. Tao L, Wang J, Wang K, et al. Exerkine FNDC5/irisin-enriched exosomes promote proliferation and inhibit ferroptosis of osteoblasts through interaction with Caveolin-1. Aging Cell. 2024;23(8):e14181. doi:10.1111/acel.14181
146. Zhang C, Pathrikar TV, Baby HM, et al. Charge-reversed exosomes for targeted gene delivery to cartilage for osteoarthritis treatment. Small Meth. 2024;8(9):e2301443. doi:10.1002/smtd.202301443
147. Moseng T, Vliet Vlieland TPM, Battista S, et al. EULAR recommendations for the non-pharmacological core management of Hip and knee osteoarthritis: 2023 update. Ann Rheum Dis. 2024;83(6):730–740. doi:10.1136/ard-2023-225041
148. He L, He T, Xing J, et al. Bone marrow mesenchymal stem cell-derived exosomes protect cartilage damage and relieve knee osteoarthritis pain in a rat model of osteoarthritis. Stem Cell Res Ther. 2020;11(1):276. doi:10.1186/s13287-020-01781-w
149. Yu H, Huang Y, Yang L. Research progress in the use of mesenchymal stem cells and their derived exosomes in the treatment of osteoarthritis. Ageing Res Rev. 2022;80:101684. doi:10.1016/j.arr.2022.101684
150. Zhu Y, Wang Y, Zhao B, et al. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res Ther. 2017;8(1):64. doi:10.1186/s13287-017-0510-9
151. Yang Z, Zhang W, Ren X, Tu C, Li Z. Exosomes: a friend or foe for osteoporotic fracture? Front Endocrinol. 2021;12:679914. doi:10.3389/fendo.2021.679914
152. Dordevic M, East J. Pilot safety study of an extracellular vesicle isolate product for treatment of osteoarthritis in combat-related injuries: one year follow up. J Stem Cell Res. 2021;2(2). doi:10.52793/jscr.2021.2(2)-21
153. Nakase I, Noguchi K, Aoki A, Takatani-Nakase T, Fujii I, Futaki S. Arginine-rich cell-penetrating peptide-modified extracellular vesicles for active macropinocytosis induction and efficient intracellular delivery. Sci Rep. 2017;7(1):1991. doi:10.1038/s41598-017-02014-6
154. Ferguson S, Yang KS, Weissleder R. Single extracellular vesicle analysis for early cancer detection. Trends Mol Med. 2022;28(8):681–692. doi:10.1016/j.molmed.2022.05.003
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.