Back to Journals » Drug Design, Development and Therapy » Volume 18
Effects of Natural Products through Inhibiting Endoplasmic Reticulum Stress on Attenuation of Idiopathic Pulmonary Fibrosis
Authors Deng J ![]() , Liu J, Chen W, Liang Q, He Y, Sun G
, Liu J, Chen W, Liang Q, He Y, Sun G
Received 22 August 2023
Accepted for publication 23 April 2024
Published 17 May 2024 Volume 2024:18 Pages 1627—1650
DOI https://doi.org/10.2147/DDDT.S388920
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
JiuLing Deng,1,* Jing Liu,1,* WanSheng Chen,2,3,* Qing Liang,1 YuQiong He,2,3 GuangChun Sun1
1Department of Pharmacy, Shanghai Fifth People’s Hospital, Fudan University, Shanghai, 200240, People’s Republic of China; 2Institute of Chinese Materia Medica, Shanghai University of Traditional Chinese Medicine, Shanghai, 201203, People’s Republic of China; 3Department of Pharmacy, Changzheng Hospital, Second Military Medical University, Shanghai, 200003, People’s Republic of China
*These authors contributed equally to this work
Correspondence: GuangChun Sun, Department of Pharmacy, Shanghai Fifth People’s Hospital, Fudan University, Shanghai, 200240, People’s Republic of China, Tel +86 21 24289464, Email [email protected] YuQiong He, Institute of Chinese Materia Medica, Shanghai University of Traditional Chinese Medicine, Shanghai, 201203, People’s Republic of China, Tel +86 21 81886182, Email [email protected]
Abstract: With ever-increasing intensive studies of idiopathic pulmonary fibrosis (IPF), significant progresses have been made. Endoplasmic reticulum stress (ERS)/unfolded protein reaction (UPR) is associated with the development and progression of IPF, and targeting ERS/UPR may be beneficial in the treatment of IPF. Natural product is a tremendous source of new drug discovery, and accumulating studies have reported that many natural products show potential therapeutic effects for IPF via modulating one or more branches of the ERS signaling pathway. Therefore, this review focuses on critical roles of ERS in IPF development, and summarizes herbal preparations and bioactive compounds which protect against IPF through regulating ERS.
Keywords: idiopathic pulmonary fibrosis, endoplasmic reticulum stress, natural products, mechanisms
Introduction
Idiopathic pulmonary fibrosis (IPF) is an idiopathic interstitial pneumonia featured by progressive dyspnea, exercise intolerance, hypoxemia, and respiratory failure, occurring primarily in older adults.1 IPF is identified by the presence of patchy areas of fibrotic remodeling in the distal lung parenchyma with fibroblastic foci.2,3 Currently, the epidemiological studies of IPF indicated a morbidity of 2–30 cases/100,000 per annum and a prevalence of 10–60 cases/100,000.4 What’s more, median survival times for suffering from IPF are considered to be from 3 to 5 year.5–9 So far, several evidence shows that the progress of IPF is influenced by various factors including environmental factors,10 gene variants,11 aging alterations,12 epigenetic reprogramming,12 and comorbid diseases.13 However, the etiology is still unclear, the current perspectives on the etiology of IPF is the appearance of extracellular matrix (ECM) and fibrosis caused by continuous local micro-injuries causing DNA damage, imbalanced cell death and anomalous tissue remodeling.4,14–16 Drug development for IPF has been challenging because of poorly understood disease etiology.17 Consequently, it is in great demand to clarify the pathological mechanisms of IPF, and discovery potential drug candidates.
Previously, IPF believed to be a chronic and sustained inflammatory response process,18 however, current evidence suggests that the fibrotic process is primarily driven by abnormal activation of alveolar epithelial cells (AECs). The activated AECs can release mediators that facilitate the proliferation of resident mesenchymal cells, attract circulating fibrocytes, and induce the epithelial to mesenchymal transition (EMT),19 ultimately making for fibrous lesion formation. Consequently, the focus produce excessive much collagen-based ECM, causing scar and lung remodeling.20
The endoplasmic reticulum (ER) as an active intracellular organelle is stemmed from the outer membrane of the nucleus. Under physiological conditions, a cell generates about 4×106 proteins every minute, furthermore, the function of the ER is to preliminary fold and process not less than one-third of those proteins.21 The ER not only harmonizes protein fold, process, assemble and transport, but also degrades the misfolded proteins. The factors, for instance, protein load, cell metabolism, redox balance, and calcium homeostasis, can affect the ER function by promoting ER stress (ERS) as well as the adaptive response – unfolded protein reaction (UPR)21–25 and cell death.26
Recently, cumulative investigates have disclosed that ERS can promote the progression of many diseases, including depression,27 cardiovascular,28,29 neurodegenerative diseases,30 cancer,31 obesity, and diabetes.32–35 These years, numerous investigations have reported that ERS is closely associated with the development of lung diseases such as silicosis,36 asthma,37 non-small cell lung cancer,38 acute lung injury,39 and IPF.40,41 It has been demonstrated that during the progress of IPF, ERS is activated and administration of the ERS inhibitors can alleviate fibroblast proliferation and improve lung function, suggesting an important factor of ERS for the pathogenesis of IPF.42,43
Lately, there has been a growing area of interest in studying natural products for their potential pharmacological activities and mechanisms in treating IPF. All sorts of natural products, for instance, alkaloids, flavonoids, polyphenols, terpenoids, and steroids have been reported to have ability to prevent IPF development owing to inhibiting ERS, inflammatory, apoptotic, and oxidative actions. However, there is currently a lack of comprehensive studies that have summarized the role of natural products in treating IPF through the inhibition of ERS signaling. Therefore, in this article, we underwent a thorough search of databases including PubMed, Web of Science, and CNKI databases for reviews and articles published from 1998 to 2023 (up to June), with search terms (“Pulmonary fibrosis” OR “Lung fibrosis”) AND (“endoplasmic reticulum stress” OR “Unfolded protein reaction”) AND (“bioactive compound” OR “plant extract” OR “herbal preparation”). In summary, this paper concludes the participation of ERS in IPF development, and summarizes the natural products which provide potential benefits in the treatment of IPF through regulating ERS signaling pathway.
Endoplasmic Reticulum Stress and the Unfolded Protein Reaction
As an organelle, the ER is essential to regulate proteostasis, calcium storage, lipid synthesis, and mitochondrial function. Both protein misfolding and subsequent ERS causing the activation of UPR through regulating protein kinase-like endoplasmic reticulum kinase (PERK), activating transcription factor 6 (ATF6), and inositol-requiring enzyme 1α (IRE1α), to maintain its membrane-bound state of non-activation or bind to glucose-reactive protein 78 (GRP78). The UPR is activated to cope with the stress reaction; however, serious situation can ultimately lead to cell death. During the process of UPR, amounts of chaperones, for instance, protein disulfide isomerases (PDIs), cyclophilin B, CaBP1 (calcium-binding protein) and stromal Cell Derived Factor 2 Like Protein 1 (SDF2-L1), are upregulated with the purpose of restoring ER homeostasis.44,45
The ATF6 pathway is activated by unfolded/misfolded proteins and separated from the immunoglobulin heavy-chain binding protein (BIP, namely, GRP78 or heat shock 70 kDa protein 5 (HSPA5)). In resting state, BIP remains stably bound to ATF6, but upon activation, the dissociation of BIP initiates a cascade of signaling pathways.46 However, the activation of IRE1 and PERK remains unclear, despite the presence of compatible sensing domains that facilitate homodimerize. Three different hypotheses have been proposed to explain this phenomenon: direct recognition, indirect recognition, and the hybrid recognition. Direct recognition is defined as that unfolded proteins activate the sensor IRE1 to trigger the UPR in the ER luminal;47 Indirect recognition is defined as that both PERK and IRE1 form a steady complex with BIP;48 Hybrid recognition is defined as that BIP dissociation and unfolded protein binding trigger the UPR signaling pathways.49–51
ATF6
Normally, ATF6 remains inactive in its membrane-bound form; however, ATF6 activation results in its free from the ER and move to the Golgi,52 where it is divided by site-1 proteases into amino terminal (N-terminal) and carboxy terminal domains, then N-terminal fragment is liberated, subsequently, these both new and smaller proteins are transferred into the nucleus to carry out transcriptional activities.53,54 Consequently, several chaperones proteins, including protein disulfide isomerase associated 6 (PDIA6) gene,55 calreticulin,56 and X-box binding protein 1 (XBP1), are activated by IRE1-XBP1 pathway.57,58
IRE1
IRE1 as a kind of type I transmembrane protein kinases existed in the ER transmits stress signals in answer to unfolded protein, which is the first and the most evolutionarily conserved branch of the UPR.59–61 Once ERS are detected, trans-autophosphorylation and dimerization of IRE1 activate its RNase domain. Although endoribonuclease activity plays a significant part in reducing the amount of proteins entering the ER lumen, and regulating the XBP1mRNA transcript.52,62 This splicing causes a change in the C-terminal region of XBP1, and only the XBP1 piece, which is a transcriptional factor in connection with a diversity of UPR target genes,63 including refolding and degrading genes.64
PERK
PERK is also a transmembrane endoplasmic reticulum resident protein. Similar to IRE1, PERK also belongs to Type I transmembrane protein. When feeling ERS, PERK is activated, and then homodimerized and auto-phosphorylated.65 Subsequently, the alpha subunit of eukaryotic translation Initiation factor 2α (eIF2α) at Ser51 is activated and phosphorylated (p-eIF2α), which causes the suppressant of eIF2β and suppression of protein synthesis.66 Therefore, PERK plays an indispensable role in decreasing ERS through reducing the production of nascent proteins. ATF4 is a key gene for ERS-induced autophagy and apoptosis by activating these proteins,67,68 such as C/EBP homologous protein (CHOP),69 parkin,70 and CD36,71 et al. The phosphorylated eIF2α can selectively activate ATF4, resulting in modulating amino acid transport, antioxidant defenses, and the biosynthesis of lipids the transcription factor.52,72 Notably, when the ERS persists continuously, autophagy is defined as the last guardian to restore the homeostasis of ER through engulfing the damaged ER73 (Figure 1).
The Effect of Endoplasmic Reticulum Stress in Different Cell Types on Lung Fibrosis
Based on the currently available data, it has been proved that ERS and UPR are related to lung fibrosis via initiating the induction of alveolar epithelial cells (AECs) apoptosis, differentiation of fibroblasts to myofibroblasts, M2 polarization of macrophage, and Th17 cell differentiation26,74–77(Figure 2).
Epithelial Cells
Recent research has proposed that ERS contributes to the progression of IPF. In animal experiment, BLM has been shown to enhance the induction of ERS in AECs, leading to lung injury and fibrosis.78 A full range of ERS markers including ATF4, ATF6, CHOP, and BIP have been found to have overexpression in AECs in the lung tissues of IPF patients.79,80 Surfactant Protein C (SP-C), a secreted transmembrane protein, is thoroughly found in AEC II. Overexpression of mutant SP-C protein or L188Q SP-C (a mutant form of SP-C) protein will activate the UPR and ER-associated degradation pathways, ultimately resulting in increased AECs apoptosis and lung pathological changes.81,82 Bridges et al also confirmed that the increase of exon 4 deleted SP-C protein could enhance sequential ER accumulation, followed by apoptotic cell death.83
Previous studies by Lawson et al have demonstrated that promoted-expression of L188Q SP-C or tunicamycin treatment can induce fibrotic remodeling and AECs apoptosis, suggesting there is a balance between AEC II and UPR. However, the BLM-treated L188Q SP-C mice possessed higher apoptosis of AECs and more numbers of fibroblasts than BLM-treated WT mice. Simultaneously, higher caspase-12 levels have also been observed in lung tissues in BLM-challenged-L188Q SP-C mice.78 A recent study also confirmed that L188Q SP-C expression impaired AEC II expansion during postnatal alveolarization, giving rise to a significant and perpetual decrease of AEC II numbers in adult mice, besides that, the level of the mutant allele was related to delayed onset of AEC II proliferation.84 Recently, Rodriguez et al have proposed that murine fibrosis models based on SP-C mutations cause activation of the AEC II UPR and ERS.85 Collectively, these reports indicate that the relationship between ERS and AECs apoptosis or survival is unclear. It seems that only when the epithelial cells are damaged, the ERS will be more sensitive to fibrotic remodeling.
As is known to us, CHOP, an ERS-induced transcription factor, contributes to the progress of BLM-induced fibrosis in mice lung.86 Several other researches have demonstrated that AEC II can induce apoptosis and consecutive fibrosis through promoting the induction of CHOP.87,88 Tanaka et al revealed that BLM-induced lung inflammation, apoptosis and fibrosis were attenuated in CHOP gene deficient mice.89 Further, Yang et al identified that inhibition of CHOP gene mitigated lung fibrosis through inhibiting Shh/HH signaling pathway in fibroblasts.90 Then, Yang et al also indicated that CHOP knockdown promotes engraftment and suppresses the myofibroblast change of lung resident mesenchymal/stromal cells during BLM-induced pulmonary fibrosis.91 These findings indicate that targeting CHOP may be a promising way to treat pulmonary fibrosis.
Fibroblasts
Although the repeated epithelial micro-injury is defined as a driver for IPF pathology, more and more studies have pointed that how fibroblasts respond to both nearby cells and the damaged microenvironment is a critical issue to be addressed. And, the proliferation, migration and differentiation of fibroblasts have been found to facilitate the occurrence and progression of lung fibrosis.20,92–95
ERS and the UPR are known to facilitate the development of lung fibrosis by regulating myofibroblast proliferation and differentiation.42,88,96,97 Lately, accumulating researches have pointed out the importance of ERS with regard to fibroblast during lung fibrosis.98 Baek et al provided the first evidence that the activation of UPR could promote differentiation of fibroblasts during fibrosis.99 TGF-β1-induced obviously increased expression of BIP, XBP-1, and ATF6α protein, which were coincided with an up-regulation of α-SMA and collagen I in mice or human fibroblasts. Further, 4-Phenylbutyric acid (4-PBA), as a chemical chaperone, evidently inhibited TGF-β1-induced myofibroblasts differentiation with increasing the activation of UPR, α-SMA and collagen.99,100 Cao et al have indicated that SiO2 exposure promotes the accretion of misfolded protein with triggering UPR in fibroblast, which conduces to the upregulation of ERS-related proteins.101 A similar result reported by Cheng et al found that ERS was activated in L929 and HPF-a cells induced by SiO2, which promoted activation of fibroblasts.102 In addition, PI3K/AKT signaling, upstream of ERS, can regulate fibroblast proliferation and differentiation, resulting in BLM-induced lung fibrosis.42 In fact, ERS can also activate fibroblast proliferation. ERS was activated in the course of myofibroblasts differentiation of human lung fibroblasts treated with cigarette smoke.103 Thioredoxin domain-containing protein 5 (TXNDC5), a resident ER protein, has been observed to participate in fibroblasts activation.104 A study has revealed that TXNDC5 is associated with excessive fibroblast activation, proliferation, and ECM production.1 Chen et al also revealed that IRE1α-XBP-1 signaling pathway was bound up with TXNDC5 in crystalline silica-induced pulmonary fibrogenesis model.104 Lee et al also confirmed that TXNDC5 promoted fibrogenesis by strengthening TGF-β1 signal via direct binding with TGFBR1 in fibroblasts.1 Overall, available data indicate that ERS conduces to a vulnerable fibroblast activation state in lung fibrosis. However, further research on pathological mechanism in lung fibrosis is still needed.
M2 Macrophages
In addition to endothelial injury and myofibroblasts differentiation, ERS can alter the phenotype of immune/inflammatory cells, particularly macrophages. The enhanced M2 macrophages polarization can prompt fibroblast activation through secreting profibrotic mediators (such as TGF-β1, platelet-derived growth factor (PDGF), matrix metalloproteinase 9 (MMP-9), tissue inhibitors of metal proteinase 1 (TIMP1), and CCL18).105–108 Accumulating evidence has revealed that ERS is pivotal in macrophage phenotypes, particularly in M2 macrophage polarization.109–111 Ryan et al have demonstrated that the expression of ERS genes in alveolar macrophages from patients with IPF or from mice with a fibrotic phenotype are up-regulated.112 In alveolar macrophages obtained from mice with fibrosis or allergic airway inflammation, the elevated CHOP expression along with ERS can modulate the generation of M2 macrophages, which then trigger development of lung fibrosis. Similarly, the deficiency of CHOP can attenuate the generation of M2 macrophages.110 Oh et al have demonstrated that the promoted ERS is necessary to the generation of the M2 macrophages through regulating JNK and PPARγ. Similarly, suppression of ERS shifted differentiated M2 macrophages toward an M1 phenotype.111 These literatures reveal that ERS (particularly CHOP) is associated with the M2 macrophage polarization, contributing to progression of fibrosis. However, the precise mechanism of how ERS signaling pathway participates in M2 macrophages still remains elusive.
Nonetheless, the role of ERS in macrophage is protective in several models of lung fibrosis. Apoptosis of macrophage plays an important role in host protection against mycobacterial infections.113 In the present study, ERS mediated macrophages apoptosis is thought to be crucial in host defenses against intracellular pathogens.114 Then, a recent study showed that the weakened PERK-eIF2α-ATF4 signal pathway could reduce THP-1 macrophages apoptosis and promote mycobacteria survival in the infected-macrophages.115 ERS markers (BIP and CHOP)-mediated macrophage apoptosis can protect against BLM-induced fibrosis.116 Hu et al have reported that silica-stimulated ERS is involved in the apoptosis of alveolar macrophages.117 The calcium-induced potassium ion channel KCa3.1 has been explicitly implied as a prospective treatment method to fibrotic diseases, especially IPF.118–120 Importantly, activation of the KCa3.1 ion channel induces cells including fibroblasts, macrophages, and epithelial cells in IPF.118 Perera et al discovered that barricade of the KCa3.1 ion channel mitigates the ERS and apoptosis in AEC II and macrophages.121 These data disclose that ERS in macrophage may act as protective or harmfulness effects, whereas ERS-induced apoptosis of M2 macrophages could be a great force and new treatment strategy to alleviate fibrosis in lung.
Th17 Cell Differentiation
In addition to innate immunity, ERS/UPR signaling is involved in the adaptive immune system. Differentiation of Th17 cells calls for two cytokines, including IL-6 and TGF-β.122 Th17 cells differentiation has been verified to be in connection with lung fibrosis.123,124 Preliminary studies have revealed that the CD147 protein is increased during pulmonary fibrosis.125 Geng et al have shown that CD147 promotes M1 macrophage and stimulates the differentiation of Th17 cells in lung interstitial fibrosis, perhaps through regulating IL-6, IL-1β, IL-12, and IL-23.126 In particulate matter (PM2.5)-induced lung fibrosis model, the activation of IL-17 and Th17 cell differentiation was observed.124 Dong et al have demonstrated that IL-27 mitigates BLM-induced fibrosis through modulating Th17 differentiation and cytokine secretion in lung.127 It has been proposed that inhibition of Th17 cell differentiation can result in attenuation of pulmonary fibrosis.128 Brucklacher et al have demonstrated that ERS-induced by hypoxia or nutrient deprivation can facilitate Th17 cell differentiation via sustained cytoplasmic calcium levels.129 These results revealed that Th17 cells differentiation may be a significant factor for the development of IPF.
Potential Therapeutic Approach
Mechanisms of Potential Target-TXNDC5 or STING
TXNDC5, an ER-localized protein disulfide isomerase, can catalyze the rearrangement of disulfide bonds. TXNDC5 acts as a molecular chaperone to reduce abnormal protein synthesis, promote ECM protein folding, and contribute to ECM protein stability.130 Numerous studies have shown that TXNDC5 is significantly up-regulated in lung and lung fibroblasts of IPF patients or mice by BLM-induced fibrosis. TXNDC5 promotes fibrogenesis by directly binding to TGF-β1 receptors and stabilizing TGF-β1 signaling in lung fibroblasts. In addition, in lung fibroblasts, TXNDC5 is upregulated by TGF-β1 stimulation s through ERS/ATF6-dependent transcriptional control.1 Chen et al also pointed the importance of IRE1α-TXNDC5 signaling to fibroblast activation.104 In addition to pulmonary fibrosis, TXNDC5 also plays a promoting role in other fibrotic diseases. The research group of Chen et al has demonstrated that TXNDC5 is an important promoting factor of cardiac fibrosis through promoting ECM deposition and fibrosis activation by redox-sensitive c-Jun N-terminal kinase signaling. TXNDC5 deletion protects against β agonist-induced fibrosis.130 Besides, Chen et al demonstrated that loss of TXNDC5 in kidney fibroblasts extenuated the progression of established fibrosis, hinting the potential of TXNDC5 for intervening renal fibrosis and chronic kidney diseases131 (Figure 3).
Stimulator of interferon genes (STING), namely Transmembrane protein 173 (TMEM173), is an ER-associated membrane protein activated by cyclic GMP-AMP synthase (cGAS), DEAD-box helicase 41 (DDX41) and interferon-inducible protein 16 (IFI16), in reaction to binding either host- or pathogen-derived cytosolic double-stranded DNA (dsDNA) or cyclic dinucleotides (CDNs).132,133 Increasing studies revealed the relevance and cross-regulation between the ER and STING, where the ERS known as the UPR brings into focus.134 Deng et al indicated that STING-mediated ERS signal pathway was activated in lung fibrosis mice.40 Zhang et al showed that deletion of STING can mitigate the expression of ERS-related proteins PERK, eIF2α and IRE1α.135 In turn, ERS perhaps induce activation of STING in one way or another lacking the mitochondrial intermediary, for example through stabilizing STING oligomerization or shifting STING trafficking, while the definitive mechanism has been uncleared. Since unbalanced cell death processes are involved in lung fibrosis, current evidence indicates that the deletion of STING causes an aggravated fibrosis independently of type I IFN signaling and featured with a prolonged inflammation136 (Figure 4). In addition to pulmonary fibrosis, STING also plays a promoting role in other fibrotic diseases. Xiao et al described that STING-IRF3-NLRP3 signaling promotes hepatocyte pyroptosis and hepatic inflammation in liver fibrosis.137 H-151, a selective inhibitor of the cGAS-STING signaling pathway, can mitigate cardiac fibrosis by preserving myocardial function after myocardial infarction.138 The STING-PERK-eIF2α signaling pathway makes a significant contribution to cellular senescence and organ fibrosis. Targeting the cGAS-STING-PERK signaling pathway mitigated lung and kidney fibrosis.139
These data indicated that targeting TXNDC5 or STING may be a forceful therapeutic strategy to alleviate lung fibrosis, and then improving pulmonary function and medical prognosis in patients with IPF.
Natural Products Alleviate Pulmonary Fibrosis via Regulating Endoplasmic Reticulum Stress
Currently, only the two drugs, pirfenidone and nintedanib, are able to postpone IPF progression, but drugs neither improve or even stabilize lung function and enhance quality of life. More importantly, the two therapeutic drugs have undesirable adverse effects, including gastrointestinal tract (nausea, diarrhea and dyspepsia, et al), skin reactions (rash and photosensitivity, et al), nervous system, diarrhea, and nausea.140–142 Consequently, it is in a popular to develop new drugs for IPF with fewer poisonousness and side reactions. Nowadays, over decades of researches into the mechanism of IPF, ERS has been defined to be crucial. A variety of ERS-related proteins are associated with fibrotic response, and under certain condition, inhibition of these important factors were in connection with improvement of lung fibrosis.50 Then, the natural products which have proven to provide benefits in IPF treatment by regulating ERS are summarized and categorized (Tables 1 and 2) (Figures 5 and 6).
 |
Table 1 Bioactive Compound Ameliorate Pulmonary Fibrosis by Regulating Endoplasmic Reticulum Stress |
 |
Table 2 Herbal Preparation Ameliorate Pulmonary Fibrosis by Regulating Endoplasmic Reticulum Stress |
 |
Figure 1 ERS and the UPR. |
 |
Figure 2 The effect of ERS in different cell types on pulmonary fibrosis. |
 |
Figure 3 Mechanisms of potential target-TXNDC5. |
 |
Figure 4 Mechanisms of potential target-STING. |

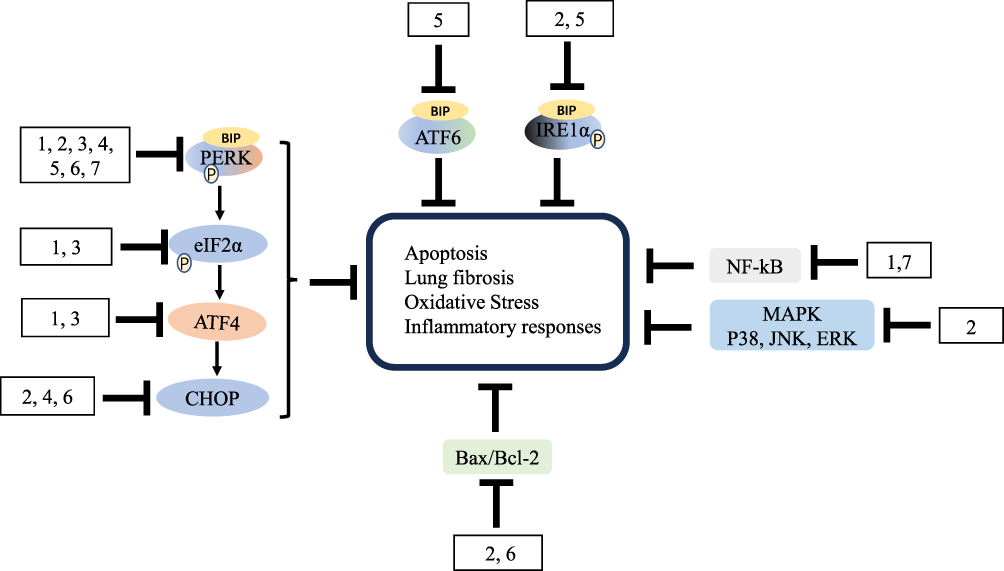
Bioactive Compounds
Naringin
Naringin is a naturally resourced flavanone glycoside mainly existed in grapefruit and citrus fruits.166 Previous studies have identified that naringin possesses diverse pharmacological activities such as anti-inflammatory and anti-oxidative stress.167,168 It has been reported that naringin shows potential benefits to withstand BLM-induced fibrosis in vivo with reducing the level of inflammatory cytokines (TNF-α, IL-6, and IL-1β), regulating oxidative stress markers (MDA, SOD, and GSH-Px) and regulating the apoptosis-related genes (Bax and Bcl-2). Moreover, it has been revealed that naringin can inhibit ERS and mitophagy-related genes (BIP, PERK, p-eIF2α, ATF4, LC-3B, p62, and Parkin), thereby activating ATF-3 and suppressing PINK1. Thus, naringin may be an up-and-coming therapeutic active ingredient for treating IPF through inhibiting ERS, decreasing apoptosis, and keeping mitochondrial homeostasis, which may be related to its modulation of ATF3/PINK1 pathway.143
Pachymic Acid
Pachymic acid is a bioactive ingredient of Poria cocos with various pharmacological properties, including anti-tumor, anti-inflammatory, antioxidant, hypoglycemic, and sedative hypnosis.169 Furthermore, pachymic acid possesses beneficial effects against fibrosis. Li et al reported that pachymic acid alleviated rat pancreatic injury and fibrosis with pancreatitis, and the inhibition of NLRP3 inflammasome is involved.170 Furthermore, in rats with BLM-induced pulmonary fibrosis, pachymic acid also showed benefits with decreasing the expressions of HYP and TGF-β1, and regulating oxidative indicators (MDA, SOD, CAT, and ROS).144 Importantly, pachymic acid treatment also could down-regulate the levels of ERS-related proteins (BIP, CHOP, caspase-9, and ATF4). Collectively, pachymic acid may alleviate BLM-induced lung fibrosis and pathological injury in vivo by inhibiting ERS and improving mitochondrial function.170
Triptolide
Triptolide, a diterpenoid triepoxide, is the active component of Tripterygium wilfordii Hook F. Triptolide has been proved to various prospective pharmacological effects, including anti-cancer, anti-tumor, anti-obesity and anti-diabetes.171 Previously, the study of triptolide on BLM-induced fibrosis in mice lung were conducted. The pulmonary function indicators such as FVC, mass FVC, and Cydn were up-regulated, and FEV0.4/FVC, RI, and RE were down-regulated in the triptolide treated groups. Moreover, triptolide can mitigate the seriousness of alveolitis and fibrosis by downregulating the abnormal expression of BIP and CHOP, indicating the inhibition ERS is involved in the triptolide against lung fibrosis.145
Tauroursodeoxycholic Acid
Tauroursodeoxycholic acid (TUDCA), a molecular chaperone, has been proved to decrease ERS via promoting protein folding and transporting.172 According to the recent research, it has been demonstrated that TUDCA mitigated non-liver diseases, for instance intestinal inflammation and neurodegenerative disorders through reducing ERS.173,174 Additionally, TUDCA has displayed a therapeutic effectiveness of lung fibrosis. Firstly, TUDCA treatment repressed ERS-related molecules/events (ATF6 and eIF2a) and subsequently alleviated paraquat-caused pulmonary fibrosis.146 Another study also demonstrated that in BLM-caused lung fibrosis, TUDCA not only prevented the BLM-induced fibrotic changes (the HYP content and histological scores), but also suppressed inflammations (total protein and leucocytes, peculiarly neutrophils), and IL-1β, caspase-11, IRE1, eIF2a, XBP-1, and CHOP.89 Then, addition to BLM-induced lung fibrosis model, TUDCA also inhibited PERK pathway-dependent ERS activation in chronic intermittent hypoxia (IH)-induced model. The levels of TGF-β1 and TSP-1 mRNAs were up-regulated after IH induction; however, these changes were reduced by administration of TUDCA. Moreover, TSP-1/TGF-β1 pathway is in relation to the protection of TUDCA against IH-stimulated lung fibrosis.147 The latest research showed that TUDCA reduced excessive cell proliferation and ECM in BLM-stimulated fibrosis model. Moreover, TUDCA prevents pulmonary TGF-β/Smad2/3-mediated EMT and fibrosis in part via inhibiting BLM-induced oxidative stress and ERS (BIP, p-PERK, p-eIF2α, ATF4, ATF6, and XBP-1s).148 Thus, TUDCA may have potential preventive and therapeutic effects for inhibiting apoptosis, oxidative stress, and fibrosis.
Engeletin
Engeletin, a flavonoid glycoside, was largely obtained from the dry rhizome of Liliaceae plant Smilax china L., shows a variety of potentially beneficial effects, for instance, inhibiting inflammation and oxidative stress.175,176 In a study, protective action of engeletin against BLM-induced lung fibrosis and TGF-β1-induced L929 cells were investigated. The results demonstrated that engeletin suppressed myofibroblast activation and ameliorated lung structure. Engeletin administration remarkably decreased the expression of collagens I and III, a-SMA, and vimentin in vivo and in vitro. RNA sequencing unveiled that PERK/ATF4 signaling pathway in relation to ERS was involved in antifibrotic actions of engeletin. Furthermore, engeletin treatment decreased the expressions of ATF4, CHOP and BIP, which was dependent on its inhibition of p-smad2/3, p-JNK, and lnc949. Overall, engeletin may be a new and promising therapeutic drug for lung fibrogenesis through suppressing ERS via lnc949-mediated TGF-β1-Smad2/3 and JNK were upstream signaling pathways.149
Spermidine
Spermidine is prevailing in living organisms as a natural polyamine.177 Spermidine exerts various activities, including antioxidant, anticancer, anti-oxidative, and anti-inflammatory.178 The actions of spermidine in BLM-induced lung fibrosis mice were also studied. BLM stimulated the upregulation of β-gal, IL-1β, TNF-α, and TGF-β1 in mice and alveolar epithelial cells, while all the changes were inhibited by spermidine therapy. BALF outcomes unveiled that spermidine obviously mitigated inflammatory reaction, including the reduction of macrophages, neutrophils, and lymphocytes. BLM-induced upregulation of ER-related proteins, for instance CHOP, GRP78, ATF6, and IRE-1 were also decreased with spermidine treatment. Thus, exogenous spermidine mitigated lung fibrosis via the downregulation of the ERS signaling pathway. Interestingly, spermidine was able to increase the LC3 I/II ratio and the autophagy-related protein (ATG7 and beclin-1), indicating autophagy may be also involved in the beneficial effects of spermidine in lung fibrosis.150
Salidroside
Salidroside, a phenylpropanoid glycoside, is the major effective ingredient discovered in all species of Rhodiola. A study has revealed that salidroside has the effect of reducing oxidative and inflammatory.179 The actions of salidroside on BLM-induced lung fibrosis in mouse have been studied, and the results showed that salidroside represented strong anti-fibrotic functions via suppressing alveolar structure injury and collagen deposition. In addition, salidroside inhibited the levels of ERS associated proteins, including ATF-4, BIP, CHOP, XBP-1, and regulating the levels of PI3K/AKT/mTOR signal proteins, including p-AKT, p-mTOR and p-p70S6K in lung tissues. Thus, perhaps salidroside is a very potential chemical constituent in alleviating BLM-induced pulmonary fibrosis.151
Ginsenoside Rb1
Ginsenoside Rb1 as one of the most prominent compounds in Panax ginseng C. A. Mey. has been proved to own various bioactivities, including decreasing oxidative stress and inflammation, balancing cell autophagy, reducing apoptosis, affecting sugar and lipid metabolism, and modulating different cytokines.180 The protective action of ginsenoside Rb1 on paraquat-induced lung fibrosis has been reported. Paraquat caused a severe respiratory failure and fibrosis in rats, and the inflammatory factors in serum were increased significantly. Otherwise, ginsenoside Rb1 administration inverted the whole biomarkers and cytokine levels, and histopathological changes induced with paraquat. Furthermore, ginsenoside Rb1 inhibited the level of BIP, MMP2 and β-catenin in lung. Thus, the outcome points out a potential part of ginsenoside Rb1 in treating paraquat-induced pulmonary fibrosis.152
Curcumin
Curcumin is an active compound extensively extracted from a member of the Zingiberaceae family. Previous researches have exhibited that curcumin holds several pharmacological actions, such as apoptosis, antiatherosclerosis, anti-inflammation, and antithrombotic activities in vitro or in vivo.181 In BLM-treated mice, the levels of α‐SMA, CCN2, and vimentin were markedly upregulated, which were reversed by an intraperitoneal injection of curcumin. Furthermore, curcumin suppressed fibroblast differentiation in BLM-treated mice lung tissues, and suppressed endothelin-1 or thrombin‐induced MAPK activation and PERK protein level in WI-38 cells. Additionally, curcumin reversed endothelin-1 or thrombin‐induced decrease of miR-19a, miR-19b, and miR-26b levels, which contributed to curcumin‐mediated suppression of CTGF generation and fibroblast differentiation.153
Isorhamnetin
Isorhamnetin is a flavonol aglycone obtained from the plant Hippophae rhamnoides L which is extensively used in traditional Chinese medicine (TCM) to the prevention and treatment of various diseases. Isorhamnetin has been proved to be effective components that exert several effects, for instance anti‑ERS, antiviral, antitumor, antioxidant, anti‑inflammatory, and neurodegenerative injury protection effects.182,183 Antifibrosis effect of isorhamnetin on mice lung fibrosis model with BLM was estimated. According to the report, isorhamnetin inhibited BLM‑induced collagen deposition, reduced collagen I and α‑SMA expression, and alleviated ERS‑mediated EMT in vivo. Furthermore, incubation of HBECs and A549 cells with TGF-β1 markedly activated EMT and ERS, and this effect was reversed by isorhamnetin via PERK pathway. Further investigations are necessary to illuminate the all-round antifibrotic effective of isorhamnetin, and identify the precise mechanism.154
Chlorogenic Acid
Chlorogenic acid, a naturally occurring non-flavonoid polyphenol, is extensively discovered in green coffee beans, teas, certain fruits, and vegetables. Researches have demonstrated that chlorogenic acid exerts antiviral, antitumor, antibacterial, and antioxidant effects.184 Importantly, chlorogenic acid (60 mg/kg) significantly suppressed BLM-promoted mesenchymal markers α-SMA and collagen I. Furthermore, chlorogenic acid represented inhibitory actions on the phosphorylation of PERK and ATF-6 in lung tissues. Interestingly, the expressions of cleaved caspase-12, caspase-9 and caspase-3 were evidently upregulated when induced by TGF-β1, which were significantly suppressed with chlorogenic acid.155
Melatonin
Melatonin as the main secretory substance of the pineal gland possesses anti-oxidant and anti-inflammatory effect.185,186 Moreover, previous research revealed the effective of melatonin in BLM-treated pulmonary fibrosis model.187,188 Melatonin clearly mitigated BLM-treated EMT and fibroblasts differentiation, as determined by its inhibition of a-SMA level. Further information discovered that melatonin markedly mitigated BLM-induced BIP upregulation and elevation of the cleaved ATF6 in the lungs. Additionally, melatonin obviously reduced the level of eIF2a, a downstream protein of the PERK pathway, as well as IRE1a phosphorylation. In brief, melatonin alleviates ERS and ERS-mediated EMT in BLM-treated lung fibrosis. Hence, melatonin may be helpful in protecting against IPF.77
Herbal Preparation
Tanreqing Injection
As is known to all, Tanreqing injection (TRQ), a Traditional Chinese Patent Medicine, is popular for the syndrome of wind-warm lung fever and phlegm-heat blocking lung. Thus, TRQ is widespread used clinically for various lung diseases, including pneumonia, COPD, and IPF.189–193 TRQ comprises five TCMs, namely Scutellaria baicalensis Georgi (Huangqin), Selenaretos thibetanus Cuvier (Xiongdanfen), Capra hircus Linnaeus (Shanyangjiao), Lonicera japonica Thunb. (Jinyinhua), and Forsythia suspensa (Thunb.) Vahl (Lianqiao).194 Previous researches have demonstrated that chemical compositions of TRQ are greater than 126 compounds, including flavonoids, phenolic acids, lignans, iridoids, amino acids, phenethyl alcohol glycosides, and steroids, and possesses a series of pharmacological effects, for instance anti-microbial, anti-inflammatory, anti-apoptotic, anti-oxidative, and anti-virus actions.190,195,196
Previously, the promising functions of TRQ in BLM-treated mice lung fibrosis have been estimated. TRQ not only improved lung edema and pulmonary function of mice with lung fibrosis (down-regulated airway resistance and up-regulated lung compliance) but also increased inflammatory responses (down-regulated the number of total cells and neutrophils in BALF, down-regulated inflammatory factors). Additionally, TRQ mitigated collagen synthesis and deposition of lung tissues as well. Moreover, TRQ relieved fibrosis via down-regulating α-SMA and up-regulating E-cadherin. Furthermore, the decreased expression of STING, p-P65, BIP, p-PERK, p-eIF2α, and ATF4 were also involved in the mechanism of TRQ treatment of IPF. However, further research should pay close attention to the potential mechanisms and the active components for TRQ treatment of IPF.40
Bushen Yifei Xiaozheng Decoction
Bushen Yifei Xiaozheng decotion (BSYF), a Chinese herbal prescription, is derived from the classic formula “Jinshui Liujun decoction”. BSYF comprises six traditional Chinese medicine, including Rehmannia glutinosa (Gaert.) Libosch. ex Fisch. et Mey. (Shudihaung), Angelica sinensis (Oliv.) Diels (Danggui), Citrus reticulata Blanco (Chenpi), Pinellia ternata (Thunb.) Ten. ex Breitenb. (Banxia), Fritillaria thunbergii Miq. (Zhebeimu), Whitmania pigra Whitman (Shuizhi). In clinical, BSYF is used to tonify lung, kidney, phlegm and eliminate disease. IPF belongs to “lump of pulmonary collateral”.197,198 Thereby, a series of studies were conducted to estimate the roles of BSYF on BLM-treated lung injury. Yan et al showed for the first time that BSYF could relieve the inflammatory reaction and inhibit deposition of extracellular matrix protein and collagen in diseased region to refrain the process of IPF rats.199 And then, systemic researches are carried out to clarify the underlying mechanisms. BSYF (12.68 g/(kg·d)) administration significantly decreased the AEC II apoptosis, and interfered the process of EMT in BLM-treated mice, and these effects of BSYF were mediated by the downregulation the abnormal expression of SP-C.156,157 Another study identified that BSYF could regulate the expression of CHOP pathway, suppress the ERS (down-regulated PERK) and inhibit the apoptosis (down-regulated Bax) of AEC II, thereby delaying the pathological changes of IPF.158 Further investigation revealed the underlying mechanism of BSYF inhibiting ERS in IPF. BSYF was given to A549 (200 μg/mL) after TGF-β1 administration. On the basis of the results presented, TGF-β1 dropped markedly the expression of SP-C and α-SMA, and the decrease in E-cadherin. Furthermore, BSYF suppressed TGF-β1-mediated EMT by inhibiting JNK signaling pathway and the BIP/IRE1 signaling pathway.159
Citrus Alkaline Extract
Citrus (Citrus reticulata Blanco) is used as a food and dietary supplement around the world.200 It is assumed that anti-fibrotic, anti-apoptosis, and anti-senescence effects of Citrus is believed to be mediated by its flavonoids and alkaloids.201–204 In Wang’s study, the effects of Citrus alkaline extract on lung fibrosis induced by BLM in rats were investigated, according to the result of the experiment, Citrus alkaline extract effectively mitigated collagen deposition, thereby ameliorating fibrosis in vivo. Furthermore, the administration of this extract was found to suppress the BLM or tunicamycin-induced upregulation of ERS biomarker (BIP and PERK), leading to a reduction of ERS levels in lung and A549 cells. What’s more, the extract treatment restrained BLM or tunicamycin-treated activation of ERS with increasing ATF-3 and PINK1 expression in vivo or in vitro. These outcomes indicate that Citrus alkaline extract acts as a promising therapeutic drug for lung fibrosis. However, further studies were needed to clarify underlying mechanisms of the whole plant extract.160
Maimendong Decoction
Maimendong Decoction (MMDD) consists of six herbs (Radix Ophiopogonis (Maimendong), Rhizoma Pinelliae (Banxia), Radix et rhizoma ginseng (Renshen), Radix glycyrrhizae (Gancao), Fructus Jujubae (Dazao), and Oryza sativa L. (Jingmi)). In clinical, MMDD is frequently used in China to treat allergic asthma, radiation pneumonitis, chronic bronchitis with lung yin deficiency, pulmonary fibrosis and other respiratory illnesses.205–207 It has been confirmed that MMDD contains various chemical constituents, such as Steroidal saponins, 4-O-Demethylophiopogonanone E, methylophiopogonanone A, and liquiritin, and possesses various pharmacological effect of anti-inflammation, anti-oxidative, and anti-apoptosis.208–210 Previously, MMDD has been confirmed to alleviate pulmonary fibrosis, improve pulmonary function (FVC), and decrease SP-C expression in BLM-induced fibrotic rats. Importantly, MMDD significantly suppressed the activation of ERS (BIP and CHOP) and cell apoptosis in AEC II. Therefore, the inhibition of ERS and apoptotic pathway in fibrotic lung tissue by MMDD may be involved in the therapeutic effects of MMDD against lung fibrosis.161
Gualou Xiebai Decoction
Gualou Xiebai Decoction (GLXB) is one of the classical prescriptions originally recorded in “Jin Kui Yao Lue” by the famous Chinese physician Zhang Zhongjing in the Han Dynasty. GLXB consists of Trichosanthes kirilowii Maxim. (Gualou) and Allium macrostemon Bunge (Xiebai) and is clinically used in treating angina pectoris and coronary heart disease.211–214 Recent studies have revealed that GLXB exerts anti-oxidative, anti-inflammatory, anti-apoptotic, and anti-fibrotic.215–217 Oral administration of GLXB increased the loss in body weight and decreased lung index by BLM-induced. Additionally, GLXB prevented lung histology injury, and relieved the severity of alveolitis and fibrosis by downregulating these abnormal expressions of p-PERK, p-IRE1α, BIP, and ATF6α. The findings suggested that GLXB might be a promising drug candidate in treating IPF.162
Yougui Drink
Yougui drink (YG) is originally recorded in “Jin Kui Shen Qi Decoction” by the famous Chinese physician Zhang Zhongjing in the Han Dynasty. YG is comprised of Rehmannia glutinosa (Gaert.) Libosch. ex Fisch. et Mey. (Shudihaung), Dioscorea polystachya Turczaninow (Shanyao), Cornus officinalis Sieb. et Zucc. (Shanzhuyu), Glycyrrhiza uralensis Sieb. et Zucc. (Shanzhuyu), Glycyrrhiza uralensis Fisch. (Zhigancao), Cinnamomum cassia (L.) D. Don (Rougui), Eucommia ulmoides Oliv. (Duzhong), Lycium chinense Miller (Gouqizi), and Aconitum carmichaelii Debeaux (Fuzi). It has been defined that YG inhibited oxidative stress, inflammatory, apoptotic and fibrotic.163,218,219 In Qiu’s experiment, the preventive and therapeutic role of YG on BLM-treated lung injury was discussed. YG administration significantly relieved dysfunction of lung and improved alveolar gas exchange function. Furthermore, YG prevented BLM-induced inflammatory infiltration and collagen deposition in lung tissues through depressing the expression of EP2 in AEC II, elevating the expression of EP3 in macrophage in lung tissues. Additionally, YG delayed lung fibrosis via modulating the levels of BIP and CHOP protein, and improving lung cell apoptosis induced by ERS.163
Danshao Huaxian Capsule
Danshao Huaxian Capsule (DSHX), a mixed preparation, is composed of five traditional Chinese herbal medicinal ingredients, including Salvia miltiorrhiza Bunge (Danshen), Paeonia lactiflora Pall (Chishao), Astragalus membranaceus (Fisch.) Bge. var. mongholicus (Bge.) Hsiao (Huangqi), Ginkgo biloba L. (Yinxingye) et al. DSHX has been used for activating blood, eliminating stasis, clearing heat and removing dampness. The ingredients of DSHX have been demonstrated to possess therapeutic actions, for instance, anti-inflammatory, anti-fibrosis, anti-oxidation.164,220 In Han’s study, the role of DSHX on BLM-treated rat pulmonary injuries was discussed. Experimental results have discovered that DSHX markedly diminished the lung HYP contents. Moreover, DSHX ameliorated oxidative stress through down-regulating the MDA level and up-regulating SOD activity. In addition, the alveolitis and fibrosis scores in the pulmonary pathology of the DSHX groups were obviously improved. Further, DSHX postponed the progression of lung fibrosis via down-regulating the level of BIP protein.164 Simultaneously, another research revealed that DSHX administration significantly relieved lung index and prevented pulmonary fibrosis by suppressing BIP and NF-κB. Therefore, these findings suggest a potential action of DSHX in treating IPF through inhibiting ERS.165
Conclusion and Future Directions
There is an ever-increasing scholarly center on the potential therapeutic targets associated with components within the endoplasmic reticulum stress (ERS) response in the treatment of idiopathic pulmonary fibrosis (IPF). Given the crucial role of the unfolded protein response (UPR) in maintaining cellular homeostasis, current research suggests that blocking or extensively inhibiting signaling through one or more branches of the ERS signaling pathway may hold promise as a treatment approach. This review article provides a comprehensive description of the actions on herbal preparations and bioactive compounds by inhibiting ERS in a lung fibrosis model, elucidating the pharmacological actions and underlying mechanisms of these agents. In addition to ERS, the roles of natural compounds have been ascribed to their anti-inflammatory, anti-oxidant, and anti-apoptotic effects, as well as their ability to activate autophagy. Various components may exhibit comparable protective roles and target similar pathways. Notably, in the process of ERS-induced fibrosis, the BIP, PERK, eIF2α, and ATF4 signaling pathways were frequently implicated. Consequently, phytochemicals hold promise as potential therapeutic agents for IPF. These studies will provide a new direction for natural products to treat IPF via clarifying the pharmacological effects and underlying mechanistic functions of these drugs. Nevertheless, there is an urgent need for further clinical trials to validate the therapeutic efficacy of these compounds against lung fibrosis.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant numbers [82304844]), the Health Profession Clinical Research Funds of Shanghai Municipal Health Commission (Grant numbers [20234Y0199]), the Natural Science Research Funds of Minhang District, Shanghai (Grant numbers [2023MHZ071]), the Scientific Research Project funded by Shanghai Municipal Science and Technology Commission (Grant numbers [21ZR1450400]), the Scientific Research Project funded by Shanghai Fifth People’s Hospital, Fudan University (Grant numbers [2020WYZD01]), and the Shanghai Municipal Health Commission clinical research project (Grant numbers [202140315]).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lee TH, Yeh CF, Lee YT, et al. Fibroblast-enriched endoplasmic reticulum protein TXNDC5 promotes pulmonary fibrosis by augmenting TGFbeta signaling through TGFBR1 stabilization. Nat Commun. 2020;11(1):4254. doi:10.1038/s41467-020-18047-x
2. Mora AL, Rojas M, Pardo A, Selman M. Emerging therapies for idiopathic pulmonary fibrosis, a progressive age-related disease. Nat Rev Drug Discov. 2017;16(11):810. doi:10.1038/nrd.2017.225
3. Raghu G, Collard HR, Egan JJ, et al. An official ATS/ERS/JRS/ALAT statement: idiopathic pulmonary fibrosis: evidence-based guidelines for diagnosis and management. Am J Respir Crit Care Med. 2011;183(6):788–824. doi:10.1164/rccm.2009-040GL
4. Martinez FJ, Collard HR, Pardo A, et al. Idiopathic pulmonary fibrosis. Nat Rev Dis Primers. 2017;3:17074. doi:10.1038/nrdp.2017.74
5. Raghu G, Weycker D, Edelsberg J, Bradford WZ, Oster G. Incidence and prevalence of idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 2006;174(7):810–816. doi:10.1164/rccm.200602-163OC
6. Schwartz DA, Helmers RA, Galvin JR, et al. Determinants of survival in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 1994;149(2 Pt 1):450–454. doi:10.1164/ajrccm.149.2.8306044
7. Gay SE, Kazerooni EA, Toews GB, et al. Idiopathic pulmonary fibrosis: predicting response to therapy and survival. Am J Respir Crit Care Med. 1998;157(4 Pt 1):1063–1072. doi:10.1164/ajrccm.157.4.9703022
8. Bjoraker JA, Ryu JH, Edwin MK, et al. Prognostic significance of histopathologic subsets in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 1998;157(1):199–203. doi:10.1164/ajrccm.157.1.9704130
9. King TE, Tooze JA, Schwarz MI, Brown KR, Cherniack RM. Predicting survival in idiopathic pulmonary fibrosis: scoring system and survival model. Am J Respir Crit Care Med. 2001;164(7):1171–1181. doi:10.1164/ajrccm.164.7.2003140
10. Park Y, Ahn C, Kim TH. Occupational and environmental risk factors of idiopathic pulmonary fibrosis: a systematic review and meta-analyses. Sci Rep. 2021;11(1):4318. doi:10.1038/s41598-021-81591-z
11. Michalski JE, Schwartz DA. Genetic Risk Factors for Idiopathic Pulmonary Fibrosis: insights into Immunopathogenesis. J Inflamm Res. 2020;13:1305–1318. doi:10.2147/JIR.S280958
12. Pardo A, Selman M. The Interplay of the Genetic Architecture, Aging, and Environmental Factors in the Pathogenesis of Idiopathic Pulmonary Fibrosis. Am J Respir Cell Mol Biol. 2021;64(2):163–172. doi:10.1165/rcmb.2020-0373PS
13. Garcia-Sancho C, Buendia-Roldan I, Fernandez-Plata MR, et al. Familial pulmonary fibrosis is the strongest risk factor for idiopathic pulmonary fibrosis. Respir Med. 2011;105(12):1902–1907. doi:10.1016/j.rmed.2011.08.022
14. Richeldi L, Collard HR, Jones MG. Idiopathic pulmonary fibrosis. Lancet. 2017;389(10082):1941–1952. doi:10.1016/S0140-6736(17)30866-8
15. Strieter RM, Mehrad B. New mechanisms of pulmonary fibrosis. Chest. 2009;136(5):1364–1370. doi:10.1378/chest.09-0510
16. Ramli I, Cheriet T, Posadino AM, et al. Potential Therapeutic Targets of Resveratrol in the Prevention and Treatment of Pulmonary Fibrosis. Front Biosci (Landmark Ed). 2023;28(9):198. doi:10.31083/j.fbl2809198
17. Mai TH, Han LW, Hsu JC, Kamath N, Pan L. Idiopathic pulmonary fibrosis therapy development: a clinical pharmacology perspective. Ther Adv Respir Dis. 2023;17:17534666231181537. doi:10.1177/17534666231181537
18. Jia Q, Lei Y, Chen S, Liu S, Wang T, Cheng Y. Circulating inflammatory cytokines and risk of idiopathic pulmonary fibrosis: a Mendelian randomization study. BMC Pulm Med. 2023;23(1):369. doi:10.1186/s12890-023-02658-3
19. Salton F, Ruaro B, Confalonieri P, Confalonieri M. Epithelial-Mesenchymal Transition: a Major Pathogenic Driver in Idiopathic Pulmonary Fibrosis? Medicina (Kaunas). 2020;56(11):608. doi:10.3390/medicina56110608
20. King TE, Pardo A, Selman M. Idiopathic pulmonary fibrosis. Lancet. 2011;378(9807):1949–1961. doi:10.1016/S0140-6736(11)60052-4
21. Princiotta MF, Finzi D, Qian SB, et al. Quantitating protein synthesis, degradation, and endogenous antigen processing. Immunity. 2003;18(3):343–354. doi:10.1016/s1074-7613(03)00051-7
22. Frakes AE, Dillin A. The UPR(ER): sensor and Coordinator of Organismal Homeostasis. Mol Cell. 2017;66(6):761–771. doi:10.1016/j.molcel.2017.05.031
23. Hetz C, Saxena S. ER stress and the unfolded protein response in neurodegeneration. Nat Rev Neurol. 2017;13(8):477–491. doi:10.1038/nrneurol.2017.99
24. Oyadomari S, Mori M. Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2004;11(4):381–389. doi:10.1038/sj.cdd.4401373
25. Wouters BG, Koritzinsky M. Hypoxia signalling through mTOR and the unfolded protein response in cancer. Nat Rev Cancer. 2008;8(11):851–864. doi:10.1038/nrc2501
26. Xu C, Bailly-Maitre B, Reed JC. Endoplasmic reticulum stress: cell life and death decisions. J Clin Invest. 2005;115(10):2656–2664. doi:10.1172/JCI26373
27. Kowalczyk M, Majsterek I, Galecki P, Talarowska M. The role of the endoplasmic reticulum stress in depression. Psychiatr Pol. 2020;54(3):499–508. doi:10.12740/PP/109130
28. Hotamisligil GS. Endoplasmic reticulum stress and atherosclerosis. Nat Med. 2010;16(4):396–399. doi:10.1038/nm0410-396
29. Tabas I. The role of endoplasmic reticulum stress in the progression of atherosclerosis. Circ Res. 2010;107(7):839–850. doi:10.1161/CIRCRESAHA.110.224766
30. Saxena S, Cabuy E, Caroni P. A role for motoneuron subtype-selective ER stress in disease manifestations of FALS mice. Nat Neurosci. 2009;12(5):627–636. doi:10.1038/nn.2297
31. Schonthal AH. Targeting endoplasmic reticulum stress for cancer therapy. Front Biosci (Schol Ed). 2012;4:412–431. doi:10.2741/276
32. Eizirik DL, Cardozo AK, Cnop M. The role for endoplasmic reticulum stress in diabetes mellitus. Endocr Rev. 2008;29(1):42–61. doi:10.1210/er.2007-0015
33. Hotamisligil GS. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell. 2010;140(6):900–917. doi:10.1016/j.cell.2010.02.034
34. Fonseca SG, Lipson KL, Urano F. Endoplasmic reticulum stress signaling in pancreatic beta-cells. Antioxid Redox Signal. 2007;9(12):2335–2344. doi:10.1089/ars.2007.1790
35. Maris M, Overbergh L, Gysemans C, et al. Deletion of C/EBP homologous protein (Chop) in C57Bl/6 mice dissociates obesity from insulin resistance. Diabetologia. 2012;55(4):1167–1178. doi:10.1007/s00125-011-2427-7
36. Ma J, Wang JQ, Ma CJ, et al. Wnt5a/Ca(2+) signaling regulates silica-induced ferroptosis in mouse macrophages by altering ER stress-mediated redox balance. Toxicology. 2023;490:153514. doi:10.1016/j.tox.2023.153514
37. Mo LH, Liu Y, Xu XJ, et al. Endoplasmic reticulum stress impairs the immune regulation property of macrophages in asthmatic patients. Clin Immunol. 2023:109639. doi:10.1016/j.clim.2023.109639
38. Sui HH, Xiao SS, Jiang SP, et al. Regorafenib induces NOX5-mediated endoplasmic reticulum stress and potentiates the anti-tumor activity of cisplatin in non-small cell lung cancer cells. Neoplasia. 2023;39:100897. doi:10.1016/j.neo.2023.100897
39. Li AM, Chen SY, Wu JJ, Li JX, Wang J. Ischemic Postconditioning Attenuates Myocardial Ischemia-Reperfusion-Induced Acute Lung Injury by Regulating Endoplasmic Reticulum Stress-Mediated Apoptosis. Braz J Cardiovasc Surg. 2023;38(1):79–87. doi:10.21470/1678-9741-2021-0043
40. Deng JL, He YQ, Sun GC, et al. Tanreqing injection protects against bleomycin-induced pulmonary fibrosis via inhibiting STING-mediated endoplasmic reticulum stress signaling pathway. J Ethnopharmacol. 2023;305:116071. doi:10.1016/j.jep.2022.116071
41. Phan THG, Paliogiannis P, Nasrallah GK, et al. Emerging cellular and molecular determinants of idiopathic pulmonary fibrosis. Cell Mol Life Sci. 2021;78(5):2031–2057. doi:10.1007/s00018-020-03693-7
42. Hsu HS, Liu CC, Lin JH, et al. Involvement of ER stress, PI3K/AKT activation, and lung fibroblast proliferation in bleomycin-induced pulmonary fibrosis. Sci Rep. 2017;7(1):14272. doi:10.1038/s41598-017-14612-5
43. Delbrel E, Soumare A, Naguez A, et al. HIF-1alpha triggers ER stress and CHOP-mediated apoptosis in alveolar epithelial cells, a key event in pulmonary fibrosis. Sci Rep. 2018;8(1):17939. doi:10.1038/s41598-018-36063-2
44. Zhang Z, Zhang L, Zhou L, Lei YL, Zhang YY, Huang CH. Redox signaling and unfolded protein response coordinate cell fate decisions under ER stress. Redox Biol. 2019;25:101047. doi:10.1016/j.redox.2018.11.005
45. Niforou K, Cheimonidou C, Trougakos IP. Molecular chaperones and proteostasis regulation during redox imbalance. Redox Biol. 2014;2:323–332. doi:10.1016/j.redox.2014.01.017
46. Chen X, Shen JS, Prywes R. The luminal domain of ATF6 senses endoplasmic reticulum (ER) stress and causes translocation of ATF6 from the ER to the Golgi. J Biol Chem. 2002;277(15):13045–13052. doi:10.1074/jbc.M110636200
47. Credle JJ, Finer-Moore JS, Papa FR, Stroud RM, Walter P. On the mechanism of sensing unfolded protein in the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2005;102(52):18773–18784. doi:10.1073/pnas.0509487102
48. Bertolotti A, Zhang Y, Hendershot LM, Harding HP, Ron D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat Cell Biol. 2000;2(6):326–332. doi:10.1038/35014014
49. Kimata Y, Oikawa D, Shimizu Y, Ishiwata-Kimata Y, Kohno K. A role for BiP as an adjustor for the endoplasmic reticulum stress-sensing protein Ire1. J Cell Biol. 2004;167(3):445–456. doi:10.1083/jcb.200405153
50. Kropski JA, Blackwell TS. Endoplasmic reticulum stress in the pathogenesis of fibrotic disease. J Clin Invest. 2018;128(1):64–73. doi:10.1172/JCI93560
51. Senft D, Ronai ZA. UPR, autophagy, and mitochondria crosstalk underlies the ER stress response. Trends Biochem Sci. 2015;40(3):141–148. doi:10.1016/j.tibs.2015.01.002
52. Romero F, Summer R. Protein Folding and the Challenges of Maintaining Endoplasmic Reticulum Proteostasis in Idiopathic Pulmonary Fibrosis. Ann Am Thorac Soc. 2017;14(Supplement_5):S410–S413. doi:10.1513/AnnalsATS.201703-207AW
53. Ye J, Rawson RB, Komuro R, et al. ER stress induces cleavage of membrane-bound ATF6 by the same proteases that process SREBPs. Mol Cell. 2000;6(6):1355–1364. doi:10.1016/s1097-2765(00)00133-7
54. Shen J, Prywes R. Dependence of site-2 protease cleavage of ATF6 on prior site-1 protease digestion is determined by the size of the luminal domain of ATF6. J Biol Chem. 2004;279(41):43046–43051. doi:10.1074/jbc.M408466200
55. Vekich JA, Belmont PJ, Thuerauf DJ, Glembotski CC. Protein disulfide isomerase-associated 6 is an ATF6-inducible ER stress response protein that protects cardiac myocytes from ischemia/reperfusion-mediated cell death. J Mol Cell Cardiol. 2012;53(2):259–267. doi:10.1016/j.yjmcc.2012.05.005
56. Liu ZW, Zhang Y, Tang ZG, et al. Matrine attenuates cardiac fibrosis by affecting ATF6 signaling pathway in diabetic cardiomyopathy. Eur J Pharmacol. 2017;804:21–30. doi:10.1016/j.ejphar.2017.03.061
57. Li BZ, Gao B, Ye LB, et al. Hepatitis B virus X protein (HBx) activates ATF6 and IRE1-XBP1 pathways of unfolded protein response. Virus Res. 2007;124(1–2):44–49. doi:10.1016/j.virusres.2006.09.011
58. Yoshida H, Matsui T, Yamamoto A, Okada T, Mori K. XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell. 2001;107(7):881–891. doi:10.1016/s0092-8674(01)00611-0
59. He SQ, Fu TT, Yu Y, et al. IRE1alpha regulates skeletal muscle regeneration through Myostatin mRNA decay. J Clin Invest. 2021. doi:10.1172/JCI143737
60. Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation. Science. 2011;334(6059):1081–1086. doi:10.1126/science.1209038
61. Mori K. Signalling pathways in the unfolded protein response: development from yeast to mammals. J Biochem. 2009;146(6):743–750. doi:10.1093/jb/mvp166
62. Calfon M, Zeng H, Urano F, et al. IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature. 2002;415(6867):92–96. doi:10.1038/415092a
63. Malhotra JD, Kaufman RJ. Endoplasmic reticulum stress and oxidative stress: a vicious cycle or a double-edged sword? Antioxid Redox Signal. 2007;9(12):2277–2293. doi:10.1089/ars.2007.1782
64. Yoshida H, Matsui T, Hosokawa N, Kaufman RJ, Nagata K, Mori K. A time-dependent phase shift in the mammalian unfolded protein response. Dev Cell. 2003;4(2):265–271. doi:10.1016/s1534-5807(03)00022-4
65. Liu CY, Schroder M, Kaufman RJ. Ligand-independent dimerization activates the stress response kinases IRE1 and PERK in the lumen of the endoplasmic reticulum. J Biol Chem. 2000;275(32):24881–24885. doi:10.1074/jbc.M004454200
66. Krishnamoorthy T, Pavitt GD, Zhang F, Dever TE, Hinnebusch AG. Tight binding of the phosphorylated alpha subunit of initiation factor 2 (eIF2alpha) to the regulatory subunits of guanine nucleotide exchange factor eIF2B is required for inhibition of translation initiation. Mol Cell Biol. 2001;21(15):5018–5030. doi:10.1128/MCB.21.15.5018-5030.2001
67. Luhr M, Torgersen ML, Szalai P, et al. The kinase PERK and the transcription factor ATF4 play distinct and essential roles in autophagy resulting from tunicamycin-induced ER stress. J Biol Chem. 2019;294(20):8197–8217. doi:10.1074/jbc.RA118.002829
68. Heydt Q, Larrue C, Saland E, et al. Oncogenic FLT3-ITD supports autophagy via ATF4 in acute myeloid leukemia. Oncogene. 2018;37(6):787–797. doi:10.1038/onc.2017.376
69. Cohen DM, Won KJ, Nguyen N, Lazar MA, Chen CS, Steger DJ. ATF4 licenses C/EBPbeta activity in human mesenchymal stem cells primed for adipogenesis. Elife. 2015;4:e06821. doi:10.7554/eLife.06821
70. Ham S, Kim JH, Kim H, Shin JY, Lee Y. ATF4-activated parkin induction contributes to deferasirox-mediated cytoprotection in Parkinson’s disease. Toxicol Res. 2023;39(2):191–199. doi:10.1007/s43188-022-00157-x
71. Griffiths A, Wang J, Song Q, Lee SM, Cordoba-Chacon J, Song Z. ATF4-mediated CD36 upregulation contributes to palmitate-induced lipotoxicity in hepatocytes. Am J Physiol Gastrointest Liver Physiol. 2023;324(5):G341–G353. doi:10.1152/ajpgi.00163.2022
72. Harding HP, Zhang Y, Zeng H, et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol Cell. 2003;11(3):619–633. doi:10.1016/s1097-2765(03)00105-9
73. Qi ZH, Chen LX. Endoplasmic Reticulum Stress and Autophagy. Adv Exp Med Biol. 2019;1206:167–177. doi:10.1007/978-981-15-0602-4_8
74. Byrne AJ, Maher TM, Lloyd CM. Pulmonary Macrophages: a New Therapeutic Pathway in Fibrosing Lung Disease? Trends Mol Med. 2016;22(4):303–316. doi:10.1016/j.molmed.2016.02.004
75. Byrne AJ, Mathie SA, Gregory LG, Lloyd CM. Pulmonary macrophages: key players in the innate defence of the airways. Thorax. 2015;70(12):1189–1196. doi:10.1136/thoraxjnl-2015-207020
76. Aziz M, Matsuda A, Yang WL, Jacob A, Wang P. Milk fat globule-epidermal growth factor-factor 8 attenuates neutrophil infiltration in acute lung injury via modulation of CXCR2. J Immunol. 2012;189(1):393–402. doi:10.4049/jimmunol.1200262
77. Zhao H, Wu QQ, Cao LF, et al. Melatonin inhibits endoplasmic reticulum stress and epithelial-mesenchymal transition during bleomycin-induced pulmonary fibrosis in mice. PLoS One. 2014;9(5):e97266. doi:10.1371/journal.pone.0097266
78. Lawson WE, Cheng DS, Degryse AL, et al. Endoplasmic reticulum stress enhances fibrotic remodeling in the lungs. Proc Natl Acad Sci U S A. 2011;108(26):10562–10567. doi:10.1073/pnas.1107559108
79. Korfei M, Ruppert C, Mahavadi P, et al. Epithelial endoplasmic reticulum stress and apoptosis in sporadic idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 2008;178(8):838–846. doi:10.1164/rccm.200802-313OC
80. Lawson WE, Crossno PF, Polosukhin VV, et al. Endoplasmic reticulum stress in alveolar epithelial cells is prominent in IPF: association with altered surfactant protein processing and herpesvirus infection. Am J Physiol Lung Cell Mol Physiol. 2008;294(6):L1119–26. doi:10.1152/ajplung.00382.2007
81. Bridges JP, Wert SE, Nogee LM, Weaver TE. Expression of a human surfactant protein C mutation associated with interstitial lung disease disrupts lung development in transgenic mice. J Biol Chem. 2003;278(52):52739–52746. doi:10.1074/jbc.M309599200
82. Mulugeta S, Maguire JA, Newitt JL, Russo SJ, Kotorashvili A, Beers MF. Misfolded BRICHOS SP-C mutant proteins induce apoptosis via caspase-4- and cytochrome c-related mechanisms. Am J Physiol Lung Cell Mol Physiol. 2007;293(3):L720–9. doi:10.1152/ajplung.00025.2007
83. Mulugeta S, Nguyen V, Russo SJ, Muniswamy M, Beers MF. A surfactant protein C precursor protein BRICHOS domain mutation causes endoplasmic reticulum stress, proteasome dysfunction, and caspase 3 activation. Am J Respir Cell Mol Biol. 2005;32(6):521–530. doi:10.1165/rcmb.2005-0009OC
84. Sitaraman S, Martin EP, Na CL, et al. Surfactant protein C mutation links postnatal type 2 cell dysfunction to adult disease. JCI Insight. 2021;6(14).
85. Rodriguez L, Tomer Y, Carson P, et al. Chronic Expression of a Clinical SFTPC Mutation Causes Murine Lung Fibrosis with Idiopathic Pulmonary Fibrosis Features. Am J Respir Cell Mol Biol. 2023;68(4):358–365. doi:10.1165/rcmb.2022-0203MA
86. Burman A, Kropski JA, Calvi CL, et al. Localized hypoxia links ER stress to lung fibrosis through induction of C/EBP homologous protein. JCI Insight. 2018;3(16).
87. Klymenko O, Huehn M, Wilhelm J, et al. Regulation and role of the ER stress transcription factor CHOP in alveolar epithelial type-II cells. J Mol Med (Berl). 2019;97(7):973–990. doi:10.1007/s00109-019-01787-9
88. Burman A, Tanjore H, Blackwell TS. Endoplasmic reticulum stress in pulmonary fibrosis. Matrix Biol. 2018;68-69:355–365. doi:10.1016/j.matbio.2018.03.015
89. Tanaka Y, Ishitsuka Y, Hayasaka M, et al. The exacerbating roles of CCAAT/enhancer-binding protein homologous protein (CHOP) in the development of bleomycin-induced pulmonary fibrosis and the preventive effects of tauroursodeoxycholic acid (TUDCA) against pulmonary fibrosis in mice. Pharmacol Res. 2015;99:52–62. doi:10.1016/j.phrs.2015.05.004
90. Yang XY, Sun W, Jing XY, Zhang Q, Huang H, Xu ZJ. C/EBP homologous protein promotes Sonic Hedgehog secretion from type II alveolar epithelial cells and activates Hedgehog signaling pathway of fibroblast in pulmonary fibrosis. Respir Res. 2022;23(1):86. doi:10.1186/s12931-022-02012-x
91. Yang XY, Sun W, Jing XY, Zhang Q, Huang H, Xu ZJ. Endoplasmic reticulum stress modulates the fate of lung resident mesenchymal stem cell to myofibroblast via C/EBP homologous protein during pulmonary fibrosis. Stem Cell Res Ther. 2022;13(1):279. doi:10.1186/s13287-022-02966-1
92. Scaffidi AK, Mutsaers SE, Moodley YP, et al. Oncostatin M stimulates proliferation, induces collagen production and inhibits apoptosis of human lung fibroblasts. Br J Pharmacol. 2002;136(5):793–801. doi:10.1038/sj.bjp.0704769
93. Huang CQ, Xiao X, Yang Y, et al. MicroRNA-101 attenuates pulmonary fibrosis by inhibiting fibroblast proliferation and activation. J Biol Chem. 2017;292(40):16420–16439. doi:10.1074/jbc.M117.805747
94. Liu HJ, Fang SC, Wang W, et al. Macrophage-derived MCPIP1 mediates silica-induced pulmonary fibrosis via autophagy. Part Fibre Toxicol. 2016;13(1):55. doi:10.1186/s12989-016-0167-z
95. Wang YN, Huang GJ, Wang ZX, Qin HP, Mo BW, Wang CM. Elongation factor-2 kinase acts downstream of p38 MAPK to regulate proliferation, apoptosis and autophagy in human lung fibroblasts. Exp Cell Res. 2018;363(2):291–298. doi:10.1016/j.yexcr.2018.01.019
96. Maiers JL, Malhi H. Endoplasmic Reticulum Stress in Metabolic Liver Diseases and Hepatic Fibrosis. Semin Liver Dis. 2019;39(2):235–248. doi:10.1055/s-0039-1681032
97. Cybulsky AV. Endoplasmic reticulum stress, the unfolded protein response and autophagy in kidney diseases. Nat Rev Nephrol. 2017;13(11):681–696. doi:10.1038/nrneph.2017.129
98. Wu B, Tang L, Kapoor M. Fibroblasts and their responses to chronic injury in pulmonary fibrosis. Semin Arthritis Rheum. 2021;51(1):310–317. doi:10.1016/j.semarthrit.2020.12.003
99. Baek HA, Kim DS, Park HS, et al. Involvement of endoplasmic reticulum stress in myofibroblastic differentiation of lung fibroblasts. Am J Respir Cell Mol Biol. 2012;46(6):731–739. doi:10.1165/rcmb.2011-0121OC
100. Qin XF, Lin XF, Liu L, et al. Macrophage-derived exosomes mediate silica-induced pulmonary fibrosis by activating fibroblast in an endoplasmic reticulum stress-dependent manner. J Cell Mol Med. 2021;25(9):4466–4477. doi:10.1111/jcmm.16524
101. Cao ZL, Xiao QL, Dai XN, et al. circHIPK2-mediated sigma-1R promotes endoplasmic reticulum stress in human pulmonary fibroblasts exposed to silica. Cell Death Dis. 2017;8(12):3212. doi:10.1038/s41419-017-0017-4
102. Cheng YS, Luo W, Li Z, et al. CircRNA-012091/PPP1R13B-mediated Lung Fibrotic Response in Silicosis via Endoplasmic Reticulum Stress and Autophagy. Am J Respir Cell Mol Biol. 2019;61(3):380–391. doi:10.1165/rcmb.2019-0017OC
103. Song M, Peng H, Guo W, et al. Cigarette Smoke Extract Promotes Human Lung Myofibroblast Differentiation by the Induction of Endoplasmic Reticulum Stress. Respiration. 2019;98(4):347–356. doi:10.1159/000502099
104. Chen X, Li C, Liu JL, He YY, Wei YG, Chen J. Inhibition of ER stress by targeting the IRE1alpha-TXNDC5 pathway alleviates crystalline silica-induced pulmonary fibrosis. Int Immunopharmacol. 2021;95:107519. doi:10.1016/j.intimp.2021.107519
105. Vannella KM, Wynn TA. Mechanisms of Organ Injury and Repair by Macrophages. Annu Rev Physiol. 2017;79:593–617. doi:10.1146/annurev-physiol-022516-034356
106. Larson-Casey JL, Deshane JS, Ryan AJ, Thannickal VJ, Carter AB. Macrophage Akt1 Kinase-Mediated Mitophagy Modulates Apoptosis Resistance and Pulmonary Fibrosis. Immunity. 2016;44(3):582–596. doi:10.1016/j.immuni.2016.01.001
107. Li GQ, Jin FQ, Du JX, He QJ, Yang B, Luo PH. Macrophage-secreted TSLP and MMP9 promote bleomycin-induced pulmonary fibrosis. Toxicol Appl Pharmacol. 2019;366:10–16. doi:10.1016/j.taap.2019.01.011
108. Young LR, Gulleman PM, Short CW, et al. Epithelial-macrophage interactions determine pulmonary fibrosis susceptibility in Hermansky-Pudlak syndrome. JCI Insight. 2016;1(17):e88947. doi:10.1172/jci.insight.88947
109. Wang Y, Zhu JH, Zhang L, et al. Role of C/EBP homologous protein and endoplasmic reticulum stress in asthma exacerbation by regulating the IL-4/signal transducer and activator of transcription 6/transcription factor EC/IL-4 receptor alpha positive feedback loop in M2 macrophages. J Allergy Clin Immunol. 2017;140(6):1550–1561 e8. doi:10.1016/j.jaci.2017.01.024
110. Yao YY, Wang Y, Zhang ZJ, et al. Chop Deficiency Protects Mice Against Bleomycin-induced Pulmonary Fibrosis by Attenuating M2 Macrophage Production. Mol Ther. 2016;24(5):915–925. doi:10.1038/mt.2016.36
111. Oh J, Riek AE, Weng S, et al. Endoplasmic reticulum stress controls M2 macrophage differentiation and foam cell formation. J Biol Chem. 2012;287(15):11629–11641. doi:10.1074/jbc.M111.338673
112. Ryan AJ, Larson-Casey JL, He C, Murthy S, Carter AB. Asbestos-induced disruption of calcium homeostasis induces endoplasmic reticulum stress in macrophages. J Biol Chem. 2014;289(48):33391–33403. doi:10.1074/jbc.M114.579870
113. Divangahi M, Chen M, Gan H, et al. Mycobacterium tuberculosis evades macrophage defenses by inhibiting plasma membrane repair. Nat Immunol. 2009;10(8):899–906. doi:10.1038/ni.1758
114. Lim YJ, Choi JA, Choi HH, et al. Endoplasmic reticulum stress pathway-mediated apoptosis in macrophages contributes to the survival of Mycobacterium tuberculosis. PLoS One. 2011;6(12):e28531. doi:10.1371/journal.pone.0028531
115. Yu X, Huang Y, Li YZ, et al. Mycobacterium tuberculosis PE_PGRS1 promotes mycobacteria intracellular survival via reducing the concentration of intracellular free Ca(2+) and suppressing endoplasmic reticulum stress. Mol Immunol. 2023;154:24–32. doi:10.1016/j.molimm.2022.12.007
116. Ayaub EA, Kolb PS, Mohammed-Ali Z, et al. GRP78 and CHOP modulate macrophage apoptosis and the development of bleomycin-induced pulmonary fibrosis. J Pathol. 2016;239(4):411–425. doi:10.1002/path.4738
117. Hu YB, Wu X, Qin XF, Wang L, Pan PH. Role of Endoplasmic Reticulum Stress in Silica-induced Apoptosis in RAW264.7 Cells. Biomed Environ Sci. 2017;30(8):591–600. doi:10.3967/bes2017.078
118. Wulff H, Castle NA. Therapeutic potential of KCa3.1 blockers: recent advances and promising trends. Expert Rev Clin Pharmacol. 2010;3(3):385–396. doi:10.1586/ecp.10.11
119. Olivan-Viguera A, Lozano-Gerona J, Lopez de Frutos L, et al. Inhibition of Intermediate-Conductance Calcium-Activated K Channel (KCa3.1) and Fibroblast Mitogenesis by alpha-Linolenic Acid and Alterations of Channel Expression in the Lysosomal Storage Disorders, Fabry Disease, and Niemann Pick C. Front Physiol. 2017;8:39. doi:10.3389/fphys.2017.00039
120. Sevelsted Moller L, Fialla AD, Schierwagen R, et al. The calcium-activated potassium channel KCa3.1 is an important modulator of hepatic injury. Sci Rep. 2016;6:28770. doi:10.1038/srep28770
121. Perera UE, Organ L, Dewage SNV, Derseh HB, Stent A, Snibson KJ. Increased Levels of ER Stress and Apoptosis in a Sheep Model for Pulmonary Fibrosis Are Alleviated by In Vivo Blockade of the KCa3.1 Ion Channel. Can Respir J. 2021;2021:6683195. doi:10.1155/2021/6683195
122. Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. 2006;24(2):179–189. doi:10.1016/j.immuni.2006.01.001
123. Simonian PL, Roark CL, Wehrmann F, et al. Th17-polarized immune response in a murine model of hypersensitivity pneumonitis and lung fibrosis. J Immunol. 2009;182(1):657–665.
124. Jia QY, Li QY, Wang Y, et al. Lung microbiome and transcriptome reveal mechanisms underlying PM2.5 induced pulmonary fibrosis. Sci Total Environ. 2022;831:154974. doi:10.1016/j.scitotenv.2022.154974
125. Duffield JS, Forbes SJ, Constandinou CM, et al. Selective depletion of macrophages reveals distinct, opposing roles during liver injury and repair. J Clin Invest. 2005;115(1):56–65. doi:10.1172/JCI22675
126. Geng JJ, Zhang K, Chen LN, et al. Enhancement of CD147 on M1 macrophages induces differentiation of Th17 cells in the lung interstitial fibrosis. Biochim Biophys Acta. 2014;1842(9):1770–1782. doi:10.1016/j.bbadis.2014.06.008
127. Dong ZX, Lu X, Yang YN, et al. IL-27 alleviates the bleomycin-induced pulmonary fibrosis by regulating the Th17 cell differentiation. BMC Pulm Med. 2015;15:13. doi:10.1186/s12890-015-0012-4
128. Segawa S, Goto D, Iizuka A, et al. The regulatory role of interferon-gamma producing gamma delta T cells via the suppression of T helper 17 cell activity in bleomycin-induced pulmonary fibrosis. Clin Exp Immunol. 2016;185(3):348–360. doi:10.1111/cei.12802
129. Brucklacher-Waldert V, Ferreira C, Stebegg M, et al. Cellular Stress in the Context of an Inflammatory Environment Supports TGF-beta-Independent T Helper-17 Differentiation. Cell Rep. 2017;19(11):2357–2370. doi:10.1016/j.celrep.2017.05.052
130. Shih YC, Chen CL, Zhang Y, et al. Endoplasmic Reticulum Protein TXNDC5 Augments Myocardial Fibrosis by Facilitating Extracellular Matrix Protein Folding and Redox-Sensitive Cardiac Fibroblast Activation. Circ Res. 2018;122(8):1052–1068. doi:10.1161/CIRCRESAHA.117.312130
131. Chen YT, Jhao PY, Hung CT, et al. Endoplasmic reticulum protein TXNDC5 promotes renal fibrosis by enforcing TGF-beta signaling in kidney fibroblasts. J Clin Invest. 2021;131(5).
132. Jonsson KL, Laustsen A, Krapp C, et al. IFI16 is required for DNA sensing in human macrophages by promoting production and function of cGAMP. Nat Commun. 2017;8:14391. doi:10.1038/ncomms14391
133. Parvatiyar K, Zhang Z, Teles RM, et al. The helicase DDX41 recognizes the bacterial secondary messengers cyclic di-GMP and cyclic di-AMP to activate a type I interferon immune response. Nat Immunol. 2012;13(12):1155–1161. doi:10.1038/ni.2460
134. Smith JA. STING, the Endoplasmic Reticulum, and Mitochondria: is Three a Crowd or a Conversation? Front Immunol. 2020;11:611347. doi:10.3389/fimmu.2020.611347
135. Zhang Y, Chen WZ, Wang Y. STING is an essential regulator of heart inflammation and fibrosis in mice with pathological cardiac hypertrophy via endoplasmic reticulum (ER) stress. Biomed Pharmacother. 2020;125:110022. doi:10.1016/j.biopha.2020.110022
136. Savigny F, Schricke C, Lacerda-Queiroz N, et al. Protective Role of the Nucleic Acid Sensor STING in Pulmonary Fibrosis. Front Immunol. 2020;11:588799. doi:10.3389/fimmu.2020.588799
137. Xiao Y, Zhao C, Tai Y, et al. STING mediates hepatocyte pyroptosis in liver fibrosis by Epigenetically activating the NLRP3 inflammasome. Redox Biol. 2023;62:102691. doi:10.1016/j.redox.2023.102691
138. Hu SY, Gao Y, Gao RF, et al. The selective STING inhibitor H-151 preserves myocardial function and ameliorates cardiac fibrosis in murine myocardial infarction. Int Immunopharmacol. 2022;107:108658. doi:10.1016/j.intimp.2022.108658
139. Zhang D, Liu YT, Zhu YZ, et al. A non-canonical cGAS-STING-PERK pathway facilitates the translational program critical for senescence and organ fibrosis. Nat Cell Biol. 2022;24(5):766–782. doi:10.1038/s41556-022-00894-z
140. Kreuter M, Bonella F, Wijsenbeek M, Maher TM, Spagnolo P. Pharmacological Treatment of Idiopathic Pulmonary Fibrosis: current Approaches, Unsolved Issues, and Future Perspectives. Biomed Res Int. 2015;2015:329481. doi:10.1155/2015/329481
141. Richeldi L, du Bois RM, Raghu G, et al. Efficacy and safety of nintedanib in idiopathic pulmonary fibrosis. N Engl J Med. 2014;370(22):2071–2082. doi:10.1056/NEJMoa1402584
142. Ma YJ, Zhang Q, Wang CX, Wu W. The efficacy and safety of pirfenidone in the treatment of HPS-related pulmonary fibrosis and Idiopathic pulmonary fibrosis: a systematic review and meta-analysis. Eur Rev Med Pharmacol Sci. 2022;26(22):8411–8424. doi:10.26355/eurrev_202211_30377
143. Wei Y, Sun L, Liu C, Li LJ. Naringin regulates endoplasmic reticulum stress and mitophagy through the ATF3/PINK1 signaling axis to alleviate pulmonary fibrosis. Naunyn Schmiedebergs Arch Pharmacol. 2023;396(6):1155–1169. doi:10.1007/s00210-023-02390-z
144. Li L, Jin R-J, Ji L. Pachymic acid ameliorates bleomycin-induced pulmonary fibrosis through inhibiting endoplasmic reticulum stress in rats. Environ Toxicol: Int J. 2023. doi:10.1002/tox.23824
145. Song QN, Li M, Li GX, et al. Triptolide improves lung function and endoplasmic reticulum stress in Bleomycin-induced pulmonary fibrosis in mice. Shi Zhen Chinese Medicine. 2022;33(12):2971–2973.
146. Omura T, Asari M, Yamamoto J, et al. Sodium tauroursodeoxycholate prevents paraquat-induced cell death by suppressing endoplasmic reticulum stress responses in human lung epithelial A549 cells. Biochem Biophys Res Commun. 2013;432(4):689–694. doi:10.1016/j.bbrc.2013.01.131
147. Shi ZH, Xu LH, Xie H, et al. Attenuation of intermittent hypoxia-induced apoptosis and fibrosis in pulmonary tissues via suppression of ER stress activation. BMC Pulm Med. 2020;20(1):92. doi:10.1186/s12890-020-1123-0
148. Tong B, Fu L, Hu B, et al. Tauroursodeoxycholic acid alleviates pulmonary endoplasmic reticulum stress and epithelial-mesenchymal transition in bleomycin-induced lung fibrosis. BMC Pulm Med. 2021;21(1):149. doi:10.1186/s12890-021-01514-6
149. Zhang JJ, Chen XQ, Chen HB, et al. Engeletin ameliorates pulmonary fibrosis through endoplasmic reticulum stress depending on lnc949-mediated TGF-beta1-Smad2/3 and JNK signalling pathways. Pharm Biol. 2020;58(1):1105–1114. doi:10.1080/13880209.2020.1834590
150. Baek AR, Hong J, Song KS, et al. Spermidine attenuates bleomycin-induced lung fibrosis by inducing autophagy and inhibiting endoplasmic reticulum stress (ERS)-induced cell death in mice. Exp Mol Med. 2020;52(12):2034–2045. doi:10.1038/s12276-020-00545-z
151. Huang L, Zou H, Zhao L, Xiong G, Hu CL. Effect and mechanism of salidroside on pulmonary fibrosis. J Guangxi Med Univ. 2020;37(08):1494–1499. doi:10.16190/j.cnki.45-1211/r.2020.08.016
152. Luo WJ, Wang CM, Ye MY. Effect and Mechanism of Ginsenoside Rb1 on Lung Function in Rats with Respiratory Failure Induced by Paraquat. J Med Res. 2020;49(02):86–91+98.
153. Chen YC, Chen BC, Huang HM, Lin SH, Lin CH. Activation of PERK in ET-1- and thrombin-induced pulmonary fibroblast differentiation: inhibitory effects of curcumin. J Cell Physiol. 2019;234(9):15977–15988. doi:10.1002/jcp.28256
154. Zheng Q, Tong M, Ou BQ, Liu CZ, Hu CP, Yang Y. Isorhamnetin protects against bleomycin-induced pulmonary fibrosis by inhibiting endoplasmic reticulum stress and epithelial-mesenchymal transition. Int J Mol Med. 2019;43(1):117–126. doi:10.3892/ijmm.2018.3965
155. Wang C, Dong J, Nie J, et al. Amelioration of bleomycin-induced pulmonary fibrosis by chlorogenic acid through endoplasmic reticulum stress inhibition. Apoptosis. 2017;22(9):1147–1156. doi:10.1007/s10495-017-1393-z
156. Gao WH. Study on the Mechanism of Bushen Yifei Xiaozheng Formula in the Intervention of ERS and Clinical Analysis of 28 Cases of IPF. Dissertation. Beijing University of Chinese Medicine; 2017.
157. Yang YX, Chai LM, Yan J, et al. Influences of Bushen Yifei Xiaozheng decoction on the distribution and expression of α-SMA and SP-C protein in IPF rat. Global Traditional Chine Med. 2017;10(06):659–663.
158. Gao WH, Chai LM, Yang YX, et al. Effects of Bushen Yifei Xiaozheng Formula on Expression of Key Molecules in CHOP Signal Pathway in Endoplasmic Reticulum Stress of IPF rat. Liaoning J Traditional Chine Med. 2016;43(09):1987–1990. doi:10.13192/j.issn.1000-1719.2016.09.068
159. He JF, Zhang Y, Tian SQ, et al. Mechanism of Bushen Yifei Xiaozheng Decotion Intervening in Pulmonary Fibrosis Type II Alveolar Epithelial Cell-Mesenchymal Transition Regulating Endoplasmic Reticulum Stress. World J Integrated Traditional Western Med. 2022;17(01):1–5. doi:10.13935/j.cnki.sjzx.220101
160. Wang ZC, Feng FC, He HL, et al. Citrus alkaline extracts prevent endoplasmic reticulum stress in type II alveolar epithelial cells to ameliorate pulmonary fibrosis via the ATF3/PINK1 pathway. Phytomedicine. 2021;89:153599. doi:10.1016/j.phymed.2021.153599
161. Shen MM, Nan YN, Zhang L, et al. Maimendong Decoction Improves Pulmonary Function in Rats With Idiopathic Pulmonary Fibrosis by Inhibiting Endoplasmic Reticulum Stress in AECIIs. Front Pharmacol. 2020;11:1262. doi:10.3389/fphar.2020.01262
162. Lin JM, Li LH. Effect of Gualou Xiebai Decoction on endoplasmic reticulum stress response in pulmonary fibrosis induced by Bleomycin. J Anhui Traditional Chine Med. 2019;38(05):71–76.
163. Qiu Y. Study on TCM Syndrome of Idiopathic Pulmonary Fibrosis and Experimental Study on Intervention of Yougui Yin in Rats with Pulmonary Fibrosis. Dissertation. Beijing University of Chinese Medicine; 2019.
164. Xie RJ, Han B, Zhang CJ, He XF, Yang T, Yang Q. The effect of endoplasmic reticulum stress and oxidative stress in bleomycin induced pulmonary fibrosis and protective effect of Danshaohuaxian capsule in this model. J Chongqing Med Univ. 2011;36(07):805–808. doi:10.13406/j.cnki.cyxb.2011.07.007
165. Han B, Xie RJ, Zhang CJ, He XF, Yang T, Yang Q. The effect of Dan Shao Hua Xian Capsule on expression of endoplasmic reticulum stress related protein GRP78 and NF-κB in bleomycin-induced pulmonary fibrosis. China J Tradition Chinese Med Pharm. 2011;26(11):2551–2554.
166. Turgut NH, Kara H, Elagoz S, Deveci K, Gungor H, Arslanbas E. The Protective Effect of Naringin against Bleomycin-Induced Pulmonary Fibrosis in Wistar Rats. Pulm Med. 2016;2016:7601393. doi:10.1155/2016/7601393
167. Zhao H, Liu MR, Liu H, Suo R, Lu CZ. Naringin protects endothelial cells from apoptosis and inflammation by regulating the Hippo-YAP Pathway. Biosci Rep. 2020;40(3).
168. Akamo AJ, Rotimi SO, Akinloye DI, et al. Naringin prevents cyclophosphamide-induced hepatotoxicity in rats by attenuating oxidative stress, fibrosis, and inflammation. Food Chem Toxicol. 2021;153:112266. doi:10.1016/j.fct.2021.112266
169. Wei CY, Wang HZ, Sun X, et al. Pharmacological profiles and therapeutic applications of pachymic acid (Review). Exp Ther Med. 2022;24(3):547. doi:10.3892/etm.2022.11484
170. Li F, Chen M, Ji JM, et al. Pachymic acid alleviates experimental pancreatic fibrosis through repressing NLRP3 inflammasome activation. Biosci Biotechnol Biochem. 2022;86(11):1497–1505. doi:10.1093/bbb/zbac114
171. Song J, He GN, Dai L. A comprehensive review on celastrol, triptolide and triptonide: insights on their pharmacological activity, toxicity, combination therapy, new dosage form and novel drug delivery routes. Biomed Pharmacother. 2023;162:114705. doi:10.1016/j.biopha.2023.114705
172. Ozcan U, Yilmaz E, Ozcan L, et al. Chemical chaperones reduce ER stress and restore glucose homeostasis in a mouse model of type 2 diabetes. Science. 2006;313(5790):1137–1140. doi:10.1126/science.1128294
173. Li P, Fu D, Sheng QF, Yu SH, Bao XQ, Lv ZB. TUDCA attenuates intestinal injury and inhibits endoplasmic reticulum stress-mediated intestinal cell apoptosis in necrotizing enterocolitis. Int Immunopharmacol. 2019;74:105665. doi:10.1016/j.intimp.2019.05.050
174. Launay N, Ruiz M, Grau L, et al. Tauroursodeoxycholic bile acid arrests axonal degeneration by inhibiting the unfolded protein response in X-linked adrenoleukodystrophy. Acta Neuropathol. 2017;133(2):283–301. doi:10.1007/s00401-016-1655-9
175. Peng XL, Yu J, Yu Q, Bian HD, Huang FP, Liang H. Binding of engeletin with bovine serum albumin: insights from spectroscopic investigations. J Fluoresc. 2012;22(1):511–519. doi:10.1007/s10895-011-0985-1
176. Ye WJ, Chen RJ, Sun W, et al. Determination and pharmacokinetics of engeletin in rat plasma by ultra-high performance liquid chromatography with tandem mass spectrometry. J Chromatogr B Analyt Technol Biomed Life Sci. 2017;1060:144–149. doi:10.1016/j.jchromb.2017.06.018
177. Pegg AE, Casero RA. Current status of the polyamine research field. Methods Mol Biol. 2011;720:3–35. doi:10.1007/978-1-61779-034-8_1
178. Madeo F, Eisenberg T, Pietrocola F, Kroemer G. Spermidine in health and disease. Science. 2018;359:6374. doi:10.1126/science.aan2788
179. Han JX, Luo LL, Wang YC, Wu SR, Kasim VV. Therapeutic potential and molecular mechanisms of salidroside in ischemic diseases. Front Pharmacol. 2022;13:974775. doi:10.3389/fphar.2022.974775
180. Lin Z, Xie RF, Zhong CH, Huang JY, Shi PY, Yao H. Recent progress (2015-2020) in the investigation of the pharmacological effects and mechanisms of ginsenoside Rb(1), a main active ingredient in Panax ginseng Meyer. J Ginseng Res. 2022;46(1):39–53. doi:10.1016/j.jgr.2021.07.008
181. Mokhtari-Zaer A, Marefati N, Atkin SL, Butler AE, Sahebkar A. The protective role of curcumin in myocardial ischemia-reperfusion injury. J Cell Physiol. 2018;234(1):214–222. doi:10.1002/jcp.26848
182. Ahn H, Lee GS. Isorhamnetin and hyperoside derived from water dropwort inhibits inflammasome activation. Phytomedicine. 2017;24:77–86. doi:10.1016/j.phymed.2016.11.019
183. Gao L, Yao R, Liu YZ, et al. Isorhamnetin protects against cardiac hypertrophy through blocking PI3K-AKT pathway. Mol Cell Biochem. 2017;429(1–2):167–177. doi:10.1007/s11010-017-2944-x
184. Rashidi R, Rezaee R, Shakeri A, Hayes AW, Karimi G. A review of the protective effects of chlorogenic acid against different chemicals. J Food Biochem. 2022;46(9):e14254. doi:10.1111/jfbc.14254
185. Lei Y, He H, Raza A, et al. Exogenous melatonin confers cold tolerance in rapeseed (Brassica napus L.) seedlings by improving antioxidants and genes expression. Plant Signal Behav. 2022;17(1):2129289. doi:10.1080/15592324.2022.2129289
186. Chen Z, Wang K, Guo JH, et al. Melatonin Maintains Homeostasis and Potentiates the Anti-inflammatory Response in Staphylococcus aureus-Induced Mastitis through microRNA-16b/YAP1. J Agric Food Chem. 2022;70(48):15255–15270. doi:10.1021/acs.jafc.2c05904
187. Genovese T, Di Paola R, Mazzon E, Muia C, Caputi AP, Cuzzocrea S. Melatonin limits lung injury in bleomycin treated mice. J Pineal Res. 2005;39(2):105–112. doi:10.1111/j.1600-079X.2005.00229.x
188. Yildirim Z, Kotuk M, Erdogan H, et al. Preventive effect of melatonin on bleomycin-induced lung fibrosis in rats. J Pineal Res. 2006;40(1):27–33. doi:10.1111/j.1600-079X.2005.00272.x
189. Chen XY, Kang FQ, Lai JQ, Deng XY, Guo XF, Liu SN. Comparative effectiveness of phlegm-heat clearing Chinese medicine injections for AECOPD: a systematic review and network meta-analysis. J Ethnopharmacol. 2022;292:115043. doi:10.1016/j.jep.2022.115043
190. He YQ, Zhou CC, Deng JL, Wang L, Chen WS. Tanreqing Inhibits LPS-Induced Acute Lung Injury In Vivo and In Vitro Through Downregulating STING Signaling Pathway. Front Pharmacol. 2021;12:746964. doi:10.3389/fphar.2021.746964
191. Yang HM, j Y, Chen SN, et al. Clinical observation of Tanreqing Injection in treating idiopathic pulmonary fibrosis complicated with infection caused by phlegm-heat blockage. Traditional Chine Med J. 2016;15(04):49–51. doi:10.14046/j.cnki.zyytb2002.2016.04.019
192. Hu YH, Yu WJ, Geng L. Study on Effect of Tanreqing Injection in Treatment of Cell in Body and Pulmonary Fibrosis Patients. Chinese Archives Traditional Chin Med. 2018;36(01):61–63. doi:10.13193/j.issn.1673-7717.2018.01.015
193. Zhong YQ, Mao B, Wang G, et al. Tanreqing injection combined with conventional Western medicine for acute exacerbations of chronic obstructive pulmonary disease: a systematic review. J Altern Complement Med. 2010;16(12):1309–1319. doi:10.1089/acm.2009.0686
194. Rivera D, Allkin R, Obon C, Alcaraz F, Verpoorte R, Heinrich M. What is in a name? The need for accurate scientific nomenclature for plants. J Ethnopharmacol. 2014;152(3):393–402. doi:10.1016/j.jep.2013.12.022
195. Wang L, Ma Y, He YQ, et al. Systematic investigation of the pharmacological mechanism of Tanreqing injection in treating respiratory diseases by UHPLC/Q-TOF-MS/MS based on multiple in-house chemical libraries coupled with network pharmacology. J Pharm Biomed Anal. 2021;202:114141. doi:10.1016/j.jpba.2021.114141
196. Yang WF, Cui KY, Tong Q, et al. Traditional Chinese Medicine Tanreqing Targets Both Cell Division and Virulence in Staphylococcus aureus. Front Cell Infect Microbiol. 2022;12:884045. doi:10.3389/fcimb.2022.884045
197. He JF, Zhang Y, Zhang D, Jin YH, Yan J. Theory and Treatment of IPF Based on the “Micro-accumulation in Lung Collaterals”. Acta Chine Medi. 2022;37(03):507–510. doi:10.16368/j.issn.1674-8999.2022.03.096
198. Chai LM, Liu J, Wang Z, Yan J. Treatment of idiopathic pulmonary fibrosis from “pulmonary collateral miniature syndrome”. Guangxi J Traditional Chine Med. 2012;35(02):44–45.
199. Yan J, Liu J, Wang Z, Chai LM. Influences of Bushenyifeixiaozheng decoction on MMPs gene expressions and pathological change in rats with idiopathic pulmonary fibrosis. Mod J Integr Traditional Chinese and Western Med. 2012;21(35):3893–3895.
200. Wu GA, Terol J, Ibanez V, et al. Genomics of the origin and evolution of Citrus. Nature. 2018;554(7692):311–316. doi:10.1038/nature25447
201. Wu Q, Zhou Y, Feng FC, Jin YH, Wang ZC, Zhou XM. Probing into the Mechanism of Alkaline Citrus Extract Promoted Apoptosis in Pulmonary Fibroblasts of Bleomycin-Induced Pulmonary Fibrosis Mice. Evid Based Complement Alternat Med. 2018;(2018):9658950. doi:10.1155/2018/9658950
202. Duan L, Guo L, Dou LL, et al. Discrimination of Citrus reticulata Blanco and Citrus reticulata ‘Chachi’ by gas chromatograph-mass spectrometry based metabolomics approach. Food Chem. 2016;212:123–127. doi:10.1016/j.foodchem.2016.05.141
203. Feng FC, Wang ZC, Li RF, et al. Citrus alkaline extracts prevent fibroblast senescence to ameliorate pulmonary fibrosis via activation of COX-2. Biomed Pharmacother. 2019;112:108669. doi:10.1016/j.biopha.2019.108669
204. Wu Q, Zhou Y, Zhou XM. Citrus Alkaline Extract Delayed the Progression of Pulmonary Fibrosis by Inhibiting p38/NF-kappaB Signaling Pathway-Induced Cell Apoptosis. Evid Based Complement Alternat Med. 2019;2019:1528586. doi:10.1155/2019/1528586
205. Lin JH, Walter P, Yen TS. Endoplasmic reticulum stress in disease pathogenesis. Annu Rev Pathol. 2008;3:399–425. doi:10.1146/annurev.pathmechdis.3.121806.151434
206. Luo M. Treatment of 80 cases of dry cough after cold with Maimendong decoction. Nei Mongol J Traditional Chin Med. 2013;32(26):114. doi:10.16040/j.cnki.cn15-1101.2013.26.048
207. Yu L. Maimendong Decoction for the Treatment of Idiopathic Pulmonary Fibrosis for 30 Cases. Chine Med Modern Distance Educ China. 2015;13(06):55–56.
208. He F, Xu BL, Chen C, et al. Methylophiopogonanone A suppresses ischemia/reperfusion-induced myocardial apoptosis in mice via activating PI3K/Akt/eNOS signaling pathway. Acta Pharmacol Sin. 2016;37(6):763–771. doi:10.1038/aps.2016.14
209. Zhao JW, Chen DS, Deng CS, Wang Q, Zhu W, Lin L. Evaluation of anti-inflammatory activity of compounds isolated from the rhizome of Ophiopogon japonicas. BMC Complement Altern Med. 2017;17(1):7. doi:10.1186/s12906-016-1539-5
210. Wu ZW, Zhao XK, Miyamoto A, et al. Effects of steroidal saponins extract from Ophiopogon japonicus root ameliorates doxorubicin-induced chronic heart failure by inhibiting oxidative stress and inflammatory response. Pharm Biol. 2019;57(1):176–183. doi:10.1080/13880209.2019.1577467
211. Ding MY, Ma WF, Wang XY, et al. A network pharmacology integrated pharmacokinetics strategy for uncovering pharmacological mechanism of compounds absorbed into the blood of Dan-Lou tablet on coronary heart disease. J Ethnopharmacol. 2019;242:112055. doi:10.1016/j.jep.2019.112055
212. Yan LL, Zhang WY, Wei XH, et al. Gualou Xiebai Decoction, a Traditional Chinese Medicine, Prevents Cardiac Reperfusion Injury of Hyperlipidemia Rat via Energy Modulation. Front Physiol. 2018;9:296. doi:10.3389/fphys.2018.00296
213. Zhang HM, Tang DL, Tong L, et al. Gualou xiebai banxia decoction inhibits NF-kappa B-dependent inflammation in myocardial ischemia-reperfusion injury in rats. J Tradit Chin Med. 2011;31(4):338–343. doi:10.1016/s0254-6272(12)60015-6
214. Zhang WY, Yu Y, Yan LL, et al. Discovery of cardio-protective constituents of Gualou Xiebai Decoction, a classical traditional Chinese medicinal formula. Phytomedicine. 2019;54:318–327. doi:10.1016/j.phymed.2018.04.047
215. Li C, Zhang WY, Yu Y, et al. Discovery of the mechanisms and major bioactive compounds responsible for the protective effects of Gualou Xiebai Decoction on coronary heart disease by network pharmacology analysis. Phytomedicine. 2019;56:261–268. doi:10.1016/j.phymed.2018.11.010
216. Aldous SJ. Cardiac biomarkers in acute myocardial infarction. Int J Cardiol. 2013;164(3):282–294. doi:10.1016/j.ijcard.2012.01.081
217. Xiang Z, Wang YZ, Liu SD. The chemical and metabolite profiles of Gualou-Xiebai-Banxia decoction, a classical traditional Chinese medicine formula, by using high-performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry and in-house software. J Ethnopharmacol. 2022;288:114994. doi:10.1016/j.jep.2022.114994
218. Wang QX. To Investigate the Mechanism of Yougui Yin Inhibiting MLO-Y4 Bone Cell Apoptosis Based on HIF-1α-Mediated Mitochondrial Autophagy. Dissertation. Shaanxi University of Chinese Medicine; 2020.
219. Jin WM, Sun ZH, Qin F. Study on the Regulation and Renal Protective effect of Youguiyin on ROS /NF-κB Signal Pathway in Rats with Gouty Nephropathy. Chinese J Integr Traditional and Western Nephrol. 2021;22(02):106–110+189.
220. Yu L, Zhao XK, Mu M, et al. Danshao Huaxian capsule attenuates hepatic fibrosis induced by CCL4 in rats by up-regulating expression of ALK2 and p-Smad1/5/8. Chongqing Med. 2016;45(35):4904–4907+4910.
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.