Back to Journals » Journal of Asthma and Allergy » Volume 19
Differential Metabolites Before and After Allergen Specific Immunotherapy in Children with Allergic Asthma
Authors Yue Y ![]() , Sang M
, Sang M ![]() , Zhang S
, Zhang S ![]() , Hu S, Zhang Y, Zhang S
, Hu S, Zhang Y, Zhang S
Received 4 November 2025
Accepted for publication 24 March 2026
Published 3 April 2026 Volume 2026:19 578876
DOI https://doi.org/10.2147/JAA.S578876
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 7
Editor who approved publication: Dr Luis Garcia-Marcos
Yinyan Yue,* Menghong Sang,* Shuyan Zhang, Shengang Hu, Yinan Zhang, Suqin Zhang
Department of Pediatrics, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Suqin Zhang, Email [email protected]
Background: Asthma is the most common chronic respiratory disease in children. Although conventional treatments such as inhaled corticosteroids and long-acting β 2-receptor agonists can effectively control symptoms, some children still face the problem of frequent attacks and drug side effects. As a potential etiological treatment, allergen-specific immunotherapy (AIT) aims to induce immune tolerance by gradually increasing exposure to specific allergens, thereby relieving asthma symptoms in the long term. This study aimed to investigate the differential metabolites in children with allergic asthma before and after subcutaneous allergen-specific immunotherapy (SCIT) using a non-targeted metabolomics approach, to gain deeper insight into the potential mechanisms of AIT and its systemic effects.
Methods: A total of 30 children were enrolled, including 15 healthy controls and 15 children with asthma (prior to AIT and after one year of AIT). Serum samples were analyzed using LC-MS to identify differential metabolites and their associated metabolic pathways.
Results: AIT could reverse the lung function of patients with allergic asthma. There were six pathways that had difference among the control group, pre-AIT group and post-AIT group. These six pathways were the PPAR signaling pathway (impact = 0.6000), the GABAergic synapse (impact =0.5882), the alanine, aspartate and glutamate metabolism (impact =0.4290), the central carbon metabolism in cancer (impact =0.3019), the arginine biosynthesis (impact =0.2956) and the linoleic acid metabolism (impact =0.2368). There were 24 metabolites that had difference among these three groups. During them, there were 5 metabolites (9,12,13-TriHOME; 9,10-Epoxyoctadecenoic acid; Isocitrate; L-Arginine; Succinic acid semialdehyde) of the pre-AIT patients reversed by AIT compared to the control group. The levels of the 5 metabolites were increased of the pre-AIT compared to the control group. After one year of usage of AIT, the levels of the 5 metabolites were decreased. Comparing of the pre-AIT group and post-AIT group revealed additional pathways: the intestinal immune network for IgA production (impact = 0.5) and aldosterone-regulated sodium reabsorption (impact = 0.4). Key metabolites included all-trans-retinoic acid, cortisol, and cortisone.
Conclusion: This study demonstrates for the first time the impact of one-year AIT on the metabolic profile of children with allergic asthma receiving long-term inhaled glucocorticoid therapy. Children with asthma mainly exhibited six pathways which including 24 metabolites disturbances compared to healthy peers, 5 of which were reversed following AIT. AIT may synergistically enhance the therapeutic effects of glucocorticoids while also introducing novel regulatory mechanisms. The persistence of these metabolic changes over a one-year period supports their validity as stable therapeutic outcomes rather than transient fluctuations.
Keywords: allergen specific immunotherapy, metabolite, allergic asthma, GABAergic synapse, inhaled-glucocorticoid
Introduction
Asthma is a chronic inflammatory disease of airway hyperresponsiveness accompanied by reversible airflow limitation,1 often presenting with symptoms such as wheezing, coughing, shortness of breath, and chest tightness.2 Asthma affects millions of people worldwide,3 particularly children, making it one of the most common chronic diseases in pediatrics.4 The pathogenesis of asthma is complex and involves interactions among genetic, epigenetic, and environmental factors. Treatment options for allergic asthma include allergen avoidance, pharmacotherapy, and allergen-specific immunotherapy (AIT). Although conventional treatments such as inhaled corticosteroids (ICS) and long-acting beta-agonists effectively control symptoms in most patients,5 some individuals still experience frequent acute exacerbations, medication side effects, and long-term drug dependency.6,7 Especially in pediatric treatment, due to side effects, many individuals discontinue treatment during the course of therapy. Therefore, it is crucial to find alternative or adjunctive therapies that can reduce medication use while achieving long-term symptom improvement.
Previous studies have shown that metabolic changes in asthma involve multiple pathways, such as cellular energy and hypoxia,8,9 lipid metabolism and oxidative stress,10 and immune/inflammatory pathways,11 and so on. Therefore, further research on asthma metabolism, particularly studies focused on reversing abnormal metabolic pathways in asthma, has garnered increased attention.
Allergen-specific immunotherapy (AIT), especially for has gained attention as a potential therapeutic approach. The goal of AIT is to induce allergen-specific immune tolerance,12 and it is currently the only disease-modifying therapy available for allergic conditions.13,14 AIT can be administered via subcutaneous immunotherapy or sublingual immunotherapy. Studies have shown that AIT, particularly house dust mite-specific AIT, has positive effects in improving symptoms of allergic asthma in children and reducing the need for medication; however, significant heterogeneity exists among different studies.13 The precise metabolic and systemic mechanisms underlying AIT remain unclear. Therefore, further understanding of AIT holds important clinical significance.
However, the precise metabolic and systemic mechanisms of AIT remain unclear. Metabolomics offers a comprehensive view of small-molecule changes under physiological or pathological conditions. Therefore, using a non-targeted metabolomics approach, we investigated the metabolic alterations in children with asthma before and after AIT to better understand the biological mechanisms associated with the treatment.
Material and Methods
Study Population
A total of 30 children were enrolled and divided into three groups: a control group (15 healthy children), a pre-AIT group, and a post-AIT group. Data for the pre-AIT and post-AIT groups were obtained from 15 children with asthma, with samples collected before and after one year of AIT. All asthmatic children were diagnosed according to standard pediatric asthma criteria,15 and dust mites were confirmed as the primary allergen. Inclusion Criteria: Control group participants aged 5–16 years were healthy children undergoing health examinations during the same period as the children with asthma. Inclusion criteria for children with asthma were as follows: (1) Children aged 5–16 years who meet the diagnostic criteria for bronchial asthma as outlined in the Guidelines for the Diagnosis and Optimal Management of Asthma in Children (Patient and Public Version, 2025);15 (2) Positive skin prick test or serum-specific IgE indicating sensitization to house dust mites (Dermatophagoides pteronyssinus or Dermatophagoides farinae); (3) Clinically stable condition at the time of enrollment, with no acute asthma exacerbation in the preceding four weeks; (4) Children with asthma inadequately controlled by inhaled corticosteroids. All children voluntarily participated in the study, and informed consent was obtained from their guardians. Exclusion Criteria: (1) Children with other chronic pulmonary diseases (eg., bronchiectasis, cystic fibrosis, or interstitial lung disease). (2) Presence of acute respiratory infection or systemic infection within four weeks prior to sampling. (3) History of autoimmune diseases, metabolic disorders, or malignancy. (4) Long-term systemic corticosteroid or immunosuppressant use within three months prior to enrollment. (5) Incomplete clinical information or poor compliance during follow-up.
The Usage of AIT
AIT was administered by subcutaneous injection of standardized dust mite extract (Alutard SQ, Denmark) for one year. Serum samples of participants were collected of the three groups: control group, pre-AIT group, and post-AIT group. All samples were taken with the informed consent of the children’s families. All the 15 children with allergic asthma receiving long-term inhaled glucocorticoid therapy. During the course of AIT, as the dose of AIT increases over time, the dosage of inhaled corticosteroids was not reduced. This study was approved by the Ethics Committee of The First Affiliated Hospital of Zhengzhou University (2022-KY-0876-002).
Sample Preparation and LC–MS Analysis
Serum was isolated by centrifugation at 1500 g for 10 min at 4°C and stored at −80°C. LC–MS analysis was performed on a Thermo Q Exactive Plus system coupled to an Ultimate 3000 UHPLC system. Chromatographic separation was achieved on an ACQUITY UPLC HSS T3 column (150 × 2.1 mm, 1.8 µm) maintained at 40°C. The flow rate was 0.25 mL/min, and the injection volume was 2 µL. For positive ion mode analysis (ESI+), the mobile phases were (C) 0.1% formic acid in acetonitrile and (D) 0.1% formic acid in water. For negative ion mode analysis (ESI-), the mobile phases were (A) acetonitrile and (B) 5 mM ammonium formate. The linear gradient for both modes was: 0–1 min, 2% C/A; 1–9 min, 2–50% C/A; 9–12 min, 50–98% C/A; 12–13.5 min, 98% C/A; 13.5–14 min, 98–2% C/A; and 14–20 min (ESI+) or 14–17 min (ESI-), 2% C/A.16
Mass spectrometric detection was conducted using a Q Exactive Plus mass spectrometer with an electrospray ionization (ESI) source operating in full MS-ddMS2 mode (data-dependent acquisition). Key parameters included: sheath gas pressure, 30 arb; aux gas flow, 10 arb; spray voltage, ±3.5 kV; capillary temperature, 325°C; MS1 scan range, m/z 81–1000; MS1 resolution, 70,000 FWHM; MS/MS resolution, 17,500 FWHM; normalized collision energy, 30%; and a dynamic exclusion time set to automatic.17
Data Analysis
Data Processing and Multivariate Analysis
Raw data were converted to mzXML format using MSConvert (ProteoWizard)18 and processed with XCMS19 for feature detection, retention time correction, and alignment. Metabolites were identified by matching accurate mass (<30 ppm) and MS/MS data against the HMDB,20 MassBank,21 LipidMaps,22 mzCloud,23 and KEGG24 databases. Data normalization was performed using robust LOESS signal correction (QC-RLSC).25 Features with a relative standard deviation (RSD) >30% in quality control (QC) samples were excluded.
Multivariate analysis was performed using the Ropls package.26 Following data scaling, principal component analysis (PCA), partial least-squares discriminant analysis (PLS-DA), and orthogonal PLS-DA (OPLS-DA) models were constructed. Model validity was assessed through permutation tests, with model performance evaluated by R2X, R2Y, and Q2 values. Discriminating metabolites were selected based on a variable importance in projection (VIP) score > 1 from the OPLS-DA model, in conjunction with a fold change (FC) analysis and a P value < 0.05 (derived from subsequent univariate analysis).
Pathway Analysis
Metabolic pathway analysis was performed using MetaboAnalyst, integrating pathway enrichment analysis with topology analysis. Differential metabolites were mapped onto KEGG pathways for biological interpretation using the KEGG Mapper tool. For comparisons involving three or more groups, one-way ANOVA (FDR-corrected, P < 0.05) was initially applied. For metabolites with significant ANOVA results, Tukey’s HSD post-hoc test was used for pairwise comparisons, with the criteria of q < 0.05 and |log2FC| ≥ 1 to define statistically significant differential metabolites.
Statistical Analysis
Data were analyzed by SPSS version 27.0 software. The Kolmogorov–Smirnov test was used to analyze the normality of continuous variables. For normally distributed data, an independent t-test was employed to compare differences between continuous variables; for non-normally distributed data, the Mann–Whitney U-test was used. Categorical data analysis and comparisons between the two groups were conducted using the chi-squared test or Fisher’s exact test. Paired t-tests were performed to compare data obtained before and after AIT in patients. The results were presented as the mean ± standard deviation for numerical variables for categorical data. P value <0.05 indicate statistically significant difference.
Results
General Characteristics of Participants
A total of 30 children were included in this study, comprising 15 healthy controls and 15 children with asthma (before AIT and one year after AIT). The control group consisted of 9 males and 6 females, while the pre-AIT group comprised 12 males and 3 females. There was no statistically significant difference in gender distribution between the two groups (P=0.427). The mean age of the control group was (10.73±2.187) years, and that of the experimental group was (9.40±1.957) years, with no statistically significant difference between the groups (P=0.089). And there was no difference between the control group and the post-AIT group (P=0.663).
Lung Function of the Patients Before AIT and One year After AIT
We collected lung function data, including FEV1, FEV1/FVC, PEF, PEF25, PEF50, PEF75 and MMEF from children with asthma before and after one year of AIT and performed paired comparisons. The results showed statistically significant differences between the groups (Table 1).
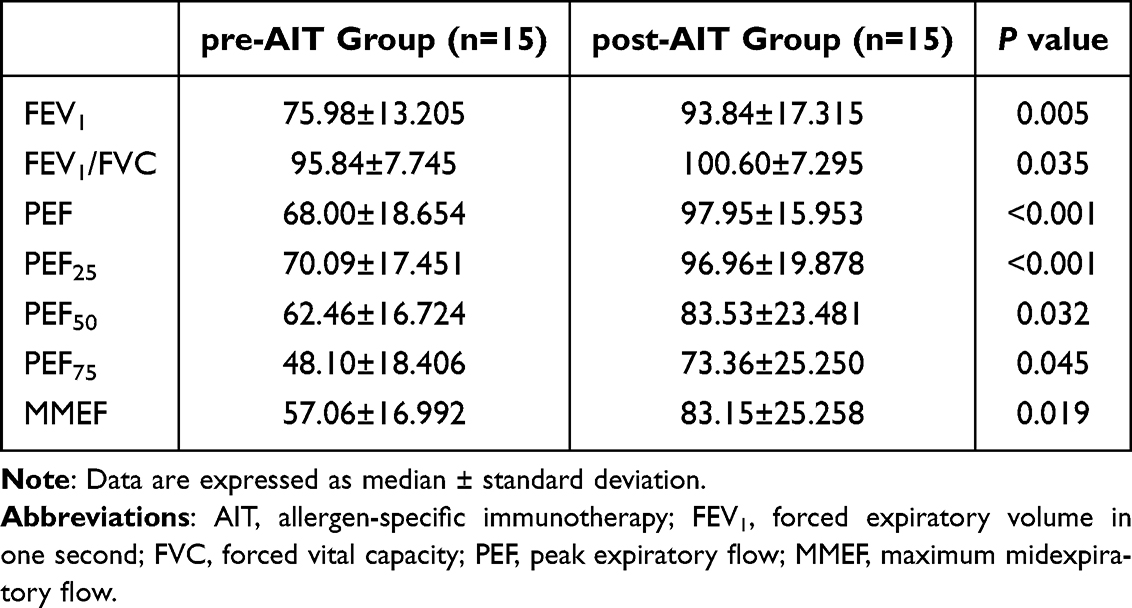 |
Table 1 Lung Function of Patients Before AIT and One year After AIT |
Data Examination
Base peak chromatograms (BPCs) of all three groups—the control group, pre-AIT group, and post-AIT group—showed similar retention time patterns, indicating good reproducibility and reliable results (Figure 1A). Principal component analysis (PCA) and orthogonal partial least squares discriminant analysis (OPLS-DA) confirmed clear separation among the three groups, and the separation observed in these figures demonstrated the reliability and high quality of the results (Figure 1B–D).
 |
Figure 1 The examination of data. (A) Typical BPC of representative samples. (B) PCA score plot of QC samples. (C) Relative standard deviation of quality assurance of QC samples. (D) OPLS-DA score plot. Abbreviations: BPC, base peak chromatograms; OPLS-DA, partial least-square discriminant analysis; PCA, principal component analysis; QC, quality control. |
Pathway Analysis of Differential Metabolites
Through searching the KEGG database, several key metabolic pathways associated with the differential metabolites were identified. A bubble plot was constructed using the pathway impact values and P-values derived from the hypergeometric distribution test, where a higher impact value and a smaller P-value indicated a greater contribution of the pathway.
Figure 2 presents a bubble plot illustrating the impact factors of metabolic pathways among the three groups: the control group, the pre-AIT group, and the post-AIT group. Six pathways were identified as significantly different across the groups. They were the PPAR signaling pathway (impact = 0.6000), the GABAergic synapse (impact = 0.5882), the alanine, aspartate and glutamate metabolism (impact =0.4290), the central carbon metabolism in cancer (impact =0.3019), the arginine biosynthesis (impact =0.2956) and the linoleic acid metabolism (impact =0.2368). Our results revealed that 24 metabolites in the pre-AIT group were significantly altered compared to the control group (Figure 3). Furthermore, these six pathways do not function in isolation; rather, they are closely interconnected through metabolites such as glutamate, tricarboxylic acid (TCA) cycle intermediates, and fatty acids. This suggests that asthma represents a systemic metabolic disorder rather than an abnormality confined to a single target. Among the 24 differentially expressed metabolites, five (9,12,13-TriHOME; 9,10-Epoxyoctadecenoic acid; Isocitrate; L-Arginine; Succinic acid semialdehyde) were found to be modulated by AIT, showing reversal toward control levels following treatment. Specifically, the levels of these five metabolites were elevated in the pre-AIT group relative to the control group, and administration of AIT reversed this trend. This reversal suggests that these metabolites may be regulated by a common upstream mechanism, with effects persisting for at least one-year post-treatment (Figure 3).
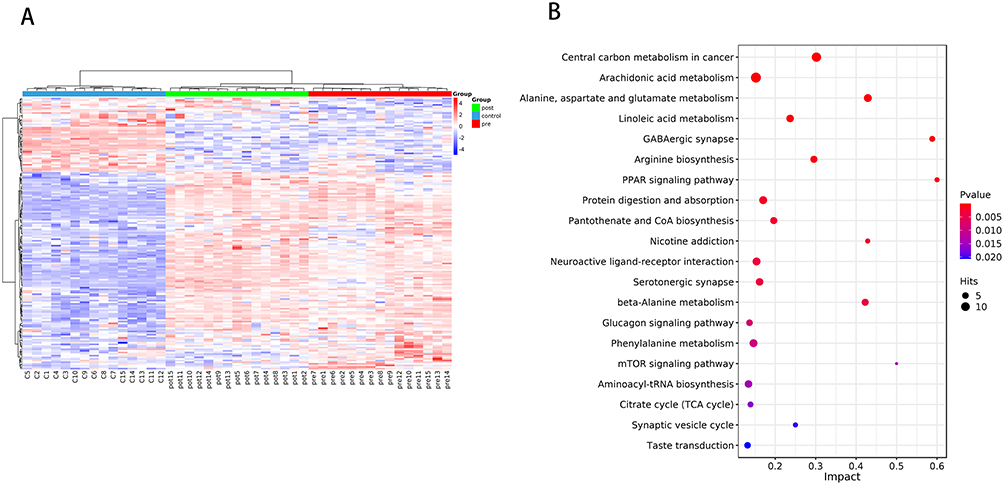 |
Figure 2 Comparisons of control group, pre-AIT group and post-AIT group. (A) Hierarchical clustering diagram of relative quantitative values of metabolites of the three groups. (B) Bubble plot of metabolic pathway influencing factors of the three groups. Each point represents a metabolic pathway, with the horizontal axis representing the impact values enriched into different metabolic pathways and the vertical axis representing enriched pathways. The dots represent the number of corresponding metabolic molecules on the pathway. Abbreviation: AIT, allergen-specific immunotherapy. |
 |
Figure 3 Comparison of the 24 metabolites among the three groups—control group, pre-AIT group, and post-AIT group. Central carbon metabolism in cancer: Citric acid (J); Fructose 1,6-bisphosphate (K); Fumaric acid (L); Isocitrate (N); L-Arginine (O); L-Aspartic acid (P); L-Glutamic acid (Q); L-Glutamine (R); L-Leucine (S); L-Methionine (T); L-Valine (U); Succinic acid (X). Alanine, aspartate and glutamate metabolism: Citric acid (J); Fumaric acid (L); gamma-Aminobutyric acid (M); L-Aspartic acid (P); L-Glutamic acid (Q); L-Glutamine (R); Succinic acid semialdehyde (W); Succinic acid (X). Linoleic acid metabolism: 9(S)-HPODE (A); 9,10-DHOME (B); 9,10-Epoxyoctadecenoic acid (C); 9-OxoODE (D); 9,12,13-TriHOME (E); 13-L-Hydroperoxylinoleic acid (H); 13S-hydroxyoctadecadienoic acid (I). GABAergic synapse: gamma-Aminobutyric acid (M); L-Glutamic acid (Q); L-Glutamine (R); Succinic acid (X). Arginine biosynthesis: Fumaric acid (L); L-Arginine (O); L-Aspartic acid (P); L-Glutamic acid (Q); L-Glutamine (R); N-acetylglutamic acid (V). PPAR signaling pathway: 12-Keto-tetrahydro-leukotriene B4 (G); 13S-hydroxyoctadecadienoic acid (I); 9-cis-Retinoic acid (F). |
The isocitrate level in the pre-AIT group was 3.54-fold higher than that in the control group (P<0.001); following one year of AIT, it decreased to 1.71-fold above the control level. The elevation of isocitrate serves as a compensatory marker of mitochondrial dysfunction, and its reversal following AIT suggests relief of metabolic blockade in the TCA cycle and restoration of mitochondrial oxidative phosphorylation capacity.
The L-arginine level in the pre-AIT group was 1.35-fold higher than that in the control group; after one year of AIT, it decreased to 0.95 times the control level. The increase in L-arginine is a typical manifestation of the “arginine paradox,” and its normalization following AIT indicates inhibition of the arginase pathway, redirecting arginine metabolism toward nitric oxide (NO) production and thereby restoring airway dilatory function.
The succinate semialdehyde level in the pre-AIT group was 28.15-fold higher than that in the control group (P <0.01); after one year of AIT, it decreased to 0.09 times the control level. The accumulation of succinic acid semialdehyde reflects impaired GABA degradation; its attenuation after AIT suggests restored succinic semialdehyde dehydrogenase (SSADH) activity, leading to normalized GABA metabolism and improved regulation of airway smooth muscle tone and immune cell function.
The 9,10-epoxyoctadecanoic acid level in the pre-AIT group was 118.03-fold higher than that in the control group (P <0.0001); following one year of AIT, it decreased to 102.31-fold above the control level (P <0.0001). The increase in 9,10-EpoME indicates abnormal activation of the cytochrome P450 (CYP) epoxygenase pathway, and its reversal suggests that AIT suppresses this overactivation and restores homeostasis of epoxy fatty acid metabolism.
The 9,12,13-trihydroxyoctadecanoic acid level in the pre-AIT group was 90.00-fold higher than that in the control group (P <0.0001); following one year of AIT, it decreased to 24.44-fold above the control level (P <0.0001). The elevation of 9,12,13-trihydroxyoctadecanoic acid provides direct evidence of oxidative stress in asthma, and its regression following AIT indicates restoration of the oxidative–antioxidative balance, protecting the airway epithelium and vascular endothelium from lipid peroxidation injury.
Although these metabolites belong to distinct pathways, they may be driven by common underlying factors—namely, oxidative stress and mitochondrial dysfunction.
Comparison between the pre-AIT and post-AIT groups identified additional pathways, including the intestinal immune network for IgA production (impact = 0.5) and aldosterone-regulated sodium reabsorption (impact = 0.4) (Figure 4). Key metabolites involved in these pathways included all-trans-retinoic acid, cortisol, and cortisone (Figure 5).
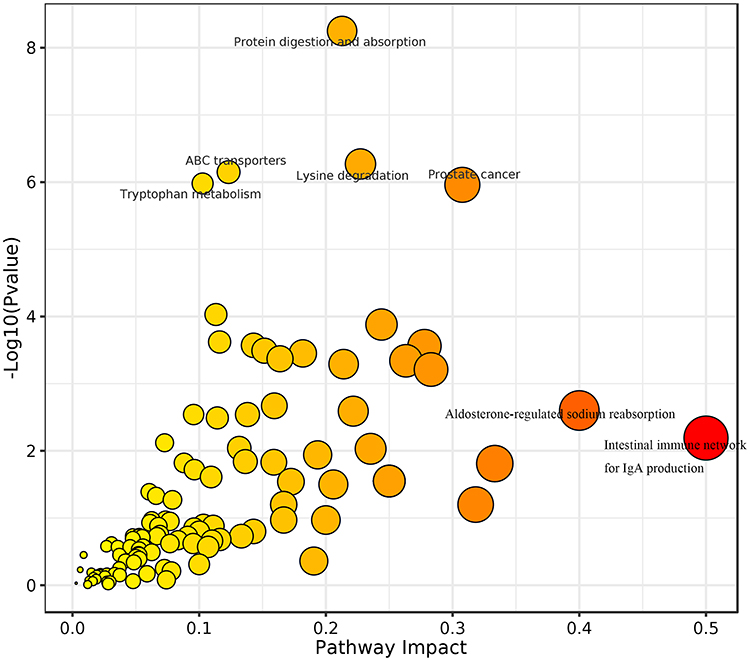 |
Figure 4 Bubble plot of metabolic pathway influencing factors of the pre-AIT group and post-AIT. Each point represents a metabolic pathway, with the horizontal axis representing the -log10 (P). The P-value represents the P-value from the hypergeometric distribution test. A smaller P-value indicates a more significant impact of the detected differential metabolites on the pathway. The larger the impact value, the larger the dot, meaning a higher value signifies a greater contribution of the detected metabolites under that pathway, resulting in a more substantial influence on the metabolic pathway. The color is related to the P-value: darker colors correspond to smaller P-values, while lighter colors correspond to larger P-values. |
 |
Figure 5 Comparison of the 3 metabolites between the pre-AIT group and post-AIT group. Intestinal immune network for IgA production: all-trans-retinoic acid (A); Aldosterone-regulated sodium reabsorption: cortisol (B), cortisone (C). |
Discussion
Our study reports, for the first time, the impact of one-year AIT intervention on the metabolic profile of asthma in patients receiving long-term glucocorticoid treatment. The within-subject design allowed us to track the dynamic metabolic evolution of the same cohort from a “disease + hormonal baseline state” to a “post-treatment state after one year,” thereby providing deeper insight into the long-term disease-modifying effects of AIT compared to conventional cross-sectional analyses.
Previous metabolomics studies have revealed that metabolic alterations in asthma predominantly involve pathways related to cellular energy metabolism and hypoxia,8,9 lipid metabolism and oxidative stress,10 immune and inflammatory responses,11 amino acid metabolism,11, steroid and nitrogen metabolism, glutamate–glutamine metabolism,9 as well as bile acid biosynthesis and vitamin metabolism.27 Consistently, our findings identified significant changes in six major metabolic pathways in asthmatic children, encompassing lipid metabolism, energy metabolism, neurotransmitter precursors, and signaling molecules. Notably, several metabolites were shared across these pathways. Among the six pathways, the PPAR signaling pathway and the GABAergic synapse pathway exerted the most significant influence.
PPARs belong to the ligand-activated nuclear receptor superfamily and comprise three subtypes: PPARα, PPARβ, and PPARγ. Leukotriene B4, retinoic acid, and 9-hydroxyoctadecadienoic acid are natural ligands of PPARs.28 Among these, PPARγ is the most widely studied subtype and functions as a master transcriptional regulator of adipogenesis. Accumulating evidence indicates that PPARγ also modulates lymphocyte responses, including those of B cells, innate lymphoid cells (ILCs), and CD4⁺ T helper cells. Studies have shown that PPARγ is selectively expressed in Th2 cells and certain Treg subsets, and its expression is upregulated in epithelial cells following IL-4 stimulation.29,30 ILC2s, which lack T-cell receptors but produce Th2-type cytokines, are enriched in allergic diseases and also express high levels of PPARγ.31 In our study, elevated levels of three PPAR ligands were observed, suggesting that the PPAR pathway may be activated in children with asthma, consistent with previous findings.
Regarding the GABAergic synapse pathway, asthmatic children exhibited decreased L-glutamine levels, along with increased levels of L-glutamic acid, succinic acid, and γ-aminobutyric acid (GABA) compared to healthy controls. GABA, a major inhibitory neurotransmitter in the central nervous system, acts as an excitatory mediator in airway epithelial cells. GABA receptor expression increases following allergen exposure, and pulmonary neuroendocrine cells (PNECs)—the primary source of GABA in the airways—promote goblet cell hyperplasia and mucus secretion via GABA signaling.32,33 A previous study reported significantly reduced serum glutaminase activity in patients with asthma relative to healthy individuals.9 Given that glutaminase catalyzes the conversion of L-glutamine to L-glutamic acid and ammonia, the observed reduction in L-glutamine may be attributable to diminished enzyme activity. However, a separate metabolomics study from Korea reported elevated levels of both glutamine and glutamate in asthma patients, suggesting that discrepancies may arise from population differences or methodological variations, warranting further investigation with larger sample sizes.8
Elevated succinic acid levels have been previously reported in adults with asthma and were similarly observed in our pediatric cohort, consistent with findings indicating that succinate expression varies among children with healthy, controlled, and uncontrolled asthma.34,35
Allergen-specific immunotherapy (AIT) is the only disease-modifying intervention recommended by the World Allergy Organization (WAO).14 The usage of AIT is to induce allergen-specific immune tolerance and prevents allergic disease progression.13 AIT can be given via a subcutaneous immunotherapy (SCIT) or sublingual immunotherapy (SLIT) route. Allergen-specific immunotherapy showed a positive effect in improving the symptoms of childhood asthma and reducing the need for medication, but there was significant heterogeneity among different studies.36 Our results showed that the level of the 5 metabolites (9,12,13-TriHOME; 9,10-Epoxyoctadecenoic acid; Isocitrate; L-Arginine; Succinic acid semialdehyde) could be reversed by AIT after one year which were increased of the pre-AIT group compared to the control group. These five regulatory metabolites belong to four dimensions: energy metabolism, amino acid metabolism, neurotransmitter metabolism, and lipid metabolism. Also, their common upstream are oxidative stress and mitochondrial dysfunction, which are the core characteristics of asthma pathophysiology. These abnormal states were significantly reversed after one year, suggesting that the investigational drug not only controlled symptoms but also potentially repaired the metabolic dysregulation of asthma at its root. This “metabolic repair” effect represents an ideal characteristic of disease-modifying therapy. It is worth noting that the model group has already been treated with glucocorticoids, while the post-AIT group has been treated with research drugs on top of glucocorticoids for one year. Therefore, the 5 metabolite callbacks observed by us are additional effects of studying drugs in the context of long-term treatment with glucocorticoids. According to literature reports, glucocorticoids themselves can regulate the metabolism of arginine and linoleic acid,37 but there is no evidence of their effects on the GABAergic pathway. In this study, the callback of succinic acid semialdehyde (GABAergic pathway) and the further decrease of arginine and linoleic acid metabolites suggest that the investigational drugs may synergistically exert their effects by enhancing the known effects of glucocorticoids and introducing new regulatory pathways. Under a one-year time frame, the continued existence of these effects eliminates the possibility of short-term fluctuations and supports their reliability as true therapeutic effects.
Notably, significant changes were also observed in two other pathways—intestinal immune network for IgA production and aldosterone-regulated sodium reabsorption—involving all-trans-retinoic acid, cortisol, and cortisone. All-trans-retinoic acid showed no significant difference from controls before or after AIT, suggesting that this pathway may not be central to asthma pathogenesis.
Protective IgA levels are often reduced in the airways of individuals with asthma. Retinoic acid, a metabolite of vitamin A, may promote IgA synthesis.38 Importantly, all-trans-retinoic acid also exerts corticosteroid-like anti-inflammatory effects and can attenuate airway hyperresponsiveness and remodeling.39 In our study, all-trans-retinoic acid levels decreased following AIT, which may reflect treatment-related metabolic adaptation; however, post-treatment levels did not differ significantly from those in controls. Cortisol and cortisone, endogenous steroid metabolites, were reduced prior to AIT and increased after treatment to levels comparable to those in healthy children. Steroid metabolism in asthma is known to fluctuate with disease activity—adrenal glucocorticoid concentrations rise during acute exacerbations and subsequently decline, returning to levels similar to controls during remission.40 Long-term systemic corticosteroid therapy can further suppress endogenous steroid metabolites. The normalization of cortisol and cortisone observed after AIT in our study may be attributable to improved disease control and reduced use of inhaled corticosteroids, although this relationship requires further validation.
There are certain limitations to this study: firstly, the lack of a control group that only uses glucocorticoids for 1 year makes it impossible for us to fully distinguish between the drug effects studied and the long-term cumulative effects of glucocorticoids. However, literature reports that the metabolic regulatory effects of glucocorticoids typically reach a plateau in the short term, so further changes after one year are more likely to be attributed to the investigational drug. Secondly, the one-year time span may have mixed factors such as season and environment, but the self-controlled before and after control design to some extent controlled for these variables. Future research could establish a long-term treatment group with glucocorticoid monotherapy and use denser time point sampling (such as 3 months, 6 months, 12 months) to accurately depict the trajectory of metabolite changes over treatment time.
Conclusion
This study demonstrates for the first time the impact of one-year AIT on the metabolic profile of children with allergic asthma receiving long-term inhaled glucocorticoid therapy. Children with asthma mainly exhibited six pathways which including 24 metabolites disturbances compared to healthy peers, 5 of which were reversed following AIT. AIT may synergistically enhance the therapeutic effects of glucocorticoids while also introducing novel regulatory mechanisms. The persistence of these metabolic changes over a one-year period supports their validity as stable therapeutic outcomes rather than transient fluctuations.
Data Sharing Statement
The dataset used and/or analyzed during the current study is available upon reasonable request from the corresponding author.
Ethics Approval and Informed Consent
This study was conducted in accordance with the ethical principles outlined in the Declaration of Helsinki. Additionally, the study was approved by the Ethics Committee of The First Affiliated Hospital of Zhengzhou University (2022-KY-0876-002). All data collected were anonymized and securely stored.
Consent for Publication
All authors have read and approved the final version of this manuscript and consent to its publication in the Journal of Asthma and Allergy. Each author confirms that the manuscript represents original work that has not been published previously and is not under consideration for publication elsewhere. All authors have reviewed the contents of the paper in full and agree to be accountable for all aspects of the work.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Natural Science Foundation of Henan Province (222300420334,232300420255) and Henan Provincial Science and Technology Research Project of China (LHGJ20200380).
Disclosure
The authors have no conflicts of interest to declare in this work.
References
1. Kaya AB, Cihangir CK, Çetin HM, Kaya SB, Erdoğanaras F. The effects of urban areas, neighborhood and housing on urban health: a systematic review and meta-analysis on asthma. Cities. 2023;143:1–13. doi:10.1016/j.cities.2023.104583
2. Al-Muhsen S, Johnson JR, Hamid Q. Remodeling in asthma. J Allergy Clin Immunol. 2011;128(3):451–62;quiz63–4. PubMed PMID: 21636119. doi:10.1016/j.jaci.2011.04.047
3. Mak HWL, Ng DCY. Spatial and socio-classification of traffic pollutant emissions and associated mortality rates in high-density hong kong via improved data analytic approaches. Int J Environ Res Public Health. 2021;18(12):6532. doi:10.3390/ijerph18126532
4. de Benedictis FM, Attanasi M. Asthma in childhood. Eur Respir Rev. 2016;25(139):41–47. PubMed PMID: 26929420; PubMed Central PMCID: PMCPMC9487667. doi:10.1183/16000617.0082-2015
5. Takaku Y, Nakagome K, Kobayashi T, et al. Changes in airway inflammation and hyperresponsiveness after inhaled corticosteroid cessation in allergic asthma. Int Arch Allergy Immunol. 2010;152(Suppl 1):41–46. PubMed PMID: 20523062. doi:10.1159/000312124
6. Shipp CL, Gergen PJ, Gern JE, Matsui EC, Guilbert TW. Asthma management in children. J Allergy Clin Immunol Pract. 2023;11(1):9–18. PubMed PMID: 36334702. doi:10.1016/j.jaip.2022.10.031
7. Haktanir Abul M, Phipatanakul W. Severe asthma in children: evaluation and management. Allergol Int. 2019;68(2):150–157. PubMed PMID: 30648539. doi:10.1016/j.alit.2018.11.007
8. Chang C, Guo ZG, He B, Yao WZ. Metabolic alterations in the sera of Chinese patients with mild persistent asthma: a GC-MS-based metabolomics analysis. Acta Pharmacol Sin. 2015;36(11):1356–1366. PubMed PMID: 26526201; PubMed Central PMCID: PMCPMC4635323. doi:10.1038/aps.2015.102
9. Jung J, Kim SH, Lee HS, et al. Serum metabolomics reveals pathways and biomarkers associated with asthma pathogenesis. Clin Exp Allergy. 2013;43(4):425–433. PubMed PMID: 23517038. doi:10.1111/cea.12089
10. Bian X, Sun B, Zheng P, Li N, Wu JL. Derivatization enhanced separation and sensitivity of long chain-free fatty acids: application to asthma using targeted and non-targeted liquid chromatography-mass spectrometry approach. Anal Chim Acta. 2017;989:59–70. PubMed PMID: 28915943. doi:10.1016/j.aca.2017.08.009
11. Comhair SA, McDunn J, Bennett C, Fettig J, Erzurum SC, Kalhan SC. Metabolomic endotype of asthma. J Immunol. 2015;195(2):643–650. PubMed PMID: 26048149; PubMed Central PMCID: PMCPMC4490949. doi:10.4049/jimmunol.1500736
12. Akar-Ghibril N, Phipatanakul W. The indoor environment and childhood asthma. Curr Allergy Asthma Rep. 2020;20(9):43. PubMed PMID: 32548675. doi:10.1007/s11882-020-00941-5
13. Burks AW, Calderon MA, Casale T, et al. Update on allergy immunotherapy: American Academy of Allergy, Asthma & Immunology/European Academy of Allergy and Clinical Immunology/PRACTALL consensus report. J Allergy Clin Immunol. 2013;131(5):1288–96.e3. PubMed PMID: 23498595. doi:10.1016/j.jaci.2013.01.049
14. Zissler UM, Jakwerth CA, Guerth F, et al. Allergen-specific immunotherapy induces the suppressive secretoglobin 1A1 in cells of the lower airways. Allergy. 2021;76(8):2461–2474. PubMed PMID: 33528894. doi:10.1111/all.14756
15. Subspecialty Group of Respiratory, the Society of Pediatrics, Chinese Medical Association; Editorial Board, Chinese Journal of Pediatrics; China Medicine Education Association Committee on Pediatrics. [Guidelines for the diagnosis and optimal management of asthma in children (2025)]. Zhonghua Er Ke Za Zhi. 2025;63(4):324–337. PubMed PMID: 40090909. doi:10.3760/cma.j.cn112140-20250124-00074 PubMed PMID: 40090909
16. Zelena E, Dunn WB, Broadhurst D, et al. Development of a robust and repeatable UPLC-MS method for the long-term metabolomic study of human serum. Anal Chem. 2009;81(4):1357–1364. PubMed PMID: 19170513. doi:10.1021/ac8019366
17. Want EJ, Masson P, Michopoulos F, et al. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat Protoc. 2013;8(1):17–32. PubMed PMID: 23222455. doi:10.1038/nprot.2012.135
18. Smith CA, Want EJ, O’Maille G, Abagyan R, Siuzdak G. XCMS: processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal Chem. 2006;78(3):779–787. PubMed PMID: 16448051. doi:10.1021/ac051437y
19. Navarro-Reig M, Jaumot J, García-Reiriz A, Tauler R. Evaluation of changes induced in rice metabolome by Cd and Cu exposure using LC-MS with XCMS and MCR-ALS data analysis strategies. Anal Bioanal Chem. 2015;407(29):8835–8847. PubMed PMID: 26403240. doi:10.1007/s00216-015-9042-2
20. Wishart DS, Tzur D, Knox C, et al. HMDB: the Human Metabolome Database. Nucleic Acids Res. 2007;35(Database issue):D521–6. PubMed PMID: 17202168; PubMed Central PMCID: PMCPMC1899095. doi:10.1093/nar/gkl923
21. Horai H, Arita M, Kanaya S, et al. MassBank: a public repository for sharing mass spectral data for life sciences. J Mass Spectrom. 2010;45(7):703–714. PubMed PMID: 20623627. doi:10.1002/jms.1777
22. Sud M, Fahy E, Cotter D, et al. LMSD: LIPID MAPS structure database. Nucleic Acids Res. 2007;35(Database issue):D527–32. PubMed PMID: 17098933; PubMed Central PMCID: PMCPMC1669719. doi:10.1093/nar/gkl838
23. Abdelrazig S, Safo L, Rance GA, et al. Metabolic characterisation of Magnetospirillum gryphiswaldense MSR-1 using LC-MS-based metabolite profiling. RSC Adv. 2020;10(54):32548–32560. PubMed PMID: 35516490; PubMed Central PMCID: PMCPMC9056635. doi:10.1039/d0ra05326k
24. Ogata H, Goto S, Sato K, Fujibuchi W, Bono H, Kanehisa M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999;27(1):29–34. PubMed PMID: 9847135; PubMed Central PMCID: PMCPMC148090. doi:10.1093/nar/27.1.29
25. Gagnebin Y, Tonoli D, Lescuyer P, et al. Metabolomic analysis of urine samples by UHPLC-QTOF-MS: impact of normalization strategies. Anal Chim Acta. 2017;955:27–35. PubMed PMID: 28088278. doi:10.1016/j.aca.2016.12.029
26. Thévenot EA, Roux A, Xu Y, Ezan E, Junot C. Analysis of the human adult urinary metabolome variations with age, body mass index, and gender by implementing a comprehensive workflow for univariate and OPLS statistical analyses. J Proteome Res. 2015;14(8):3322–3335. PubMed PMID: 26088811. doi:10.1021/acs.jproteome.5b00354
27. Kuruvilla ME, Lee FE, Lee GB. Understanding asthma phenotypes, endotypes, and mechanisms of disease. Clin Rev Allergy Immunol. 2019;56(2):219–233. PubMed PMID: 30206782; PubMed Central PMCID: PMCPMC6411459. doi:10.1007/s12016-018-8712-1
28. Nuclear Receptors Nomenclature Committee. A unified nomenclature system for the nuclear receptor superfamily. Cell. 1999;97(2):161–163. PubMed PMID: 10219237. doi:10.1016/s0092-8674(00)80726-6
29. Stark JM, Coquet JM, Tibbitt CA. The role of PPAR-γ in allergic disease. Curr Allergy Asthma Rep. 2021;21(11):45. PubMed PMID: 34697644; PubMed Central PMCID: PMCPMC8545719. doi:10.1007/s11882-021-01022-x
30. Winkler C, Hochdörfer T, Israelsson E, et al. Activation of group 2 innate lymphoid cells after allergen challenge in asthmatic patients. J Allergy Clin Immunol. 2019;144(1):61–9.e7. PubMed PMID: 30731124. doi:10.1016/j.jaci.2019.01.027
31. Sinha A, Krishnan V, Sethi T, et al. Metabolomic signatures in nuclear magnetic resonance spectra of exhaled breath condensate identify asthma. Eur Respir J. 2012;39(2):500–502. PubMed PMID: 22298617. doi:10.1183/09031936.00047711
32. Sui P, Wiesner DL, Xu J, et al. Pulmonary neuroendocrine cells amplify allergic asthma responses. Science. 2018;360(6393). PubMed PMID: 29599193; PubMed Central PMCID: PMCPMC6387886. doi:10.1126/science.aan8546
33. van Heerden D, van Binnendijk RS, Tromp SAM, Savelkoul HFJ, van Neerven RJJ, den Hartog G. Asthma-associated long TSLP inhibits the production of IgA. Int J Mol Sci. 2021;22(7):3592. PubMed PMID: 33808333; PubMed Central PMCID: PMCPMC8036615. doi:10.3390/ijms22073592
34. Saude EJ, Skappak CD, Regush S, et al. Metabolomic profiling of asthma: diagnostic utility of urine nuclear magnetic resonance spectroscopy. J Allergy Clin Immunol. 2011;127(3):757–64.e1–6. PubMed PMID: 21377043. doi:10.1016/j.jaci.2010.12.1077
35. Barrios J, Patel KR, Aven L, et al. Early life allergen-induced mucus overproduction requires augmented neural stimulation of pulmonary neuroendocrine cell secretion. FASEB j. 2017;31(9):4117–4128. PubMed PMID: 28566470; PubMed Central PMCID: PMCPMC5572694. doi:10.1096/fj.201700115R
36. Wu K, Wu S, Wang L. Efficacy evaluation of allergen-specific immunotherapy in children with asthma: a systematic review and meta-analysis. BMC Pulm Med. 2025;25(1):293. PubMed PMID: 40604821; PubMed Central PMCID: PMCPMC12218942. doi:10.1186/s12890-025-03763-1
37. Xu Z, Wu Y, Zhao X, Zhou H. Integrating nontargeted metabolomics and RNA sequencing of dexamethasone-treated and untreated asthmatic mice reveals changes of amino acids and aminoacyl-tRNA in group 2 innate lymphoid cells. Int J Biol Macromol. 2024;283(Pt 1):137630. PubMed PMID: 39547613. doi:10.1016/j.ijbiomac.2024.137630
38. Fang H, Jin H, Wang H. Effect of all-trans retinoic acid on airway inflammation in asthmatic rats and its mechanism. J Huazhong Univ Sci Technolog Med Sci. 2004;24(3):229–232. PubMed PMID: 15315333. doi:10.1007/bf02831997
39. Nomura S, Fujitaka M, Sakura N, Ueda K. Adrenocortical function in asthmatic children: low levels of adrenocortical hormones in children with persistent attacks. Eur J Pediatr. 1997;156(4):323–328. PubMed PMID: 9128821. doi:10.1007/s004310050610
40. Reinke SN, Gallart-Ayala H, Gómez C, et al. Metabolomics analysis identifies different metabotypes of asthma severity. Eur Respir J. 2017;49(3):1601740. PubMed PMID: 28356371; PubMed Central PMCID: PMCPMC5399350. doi:10.1183/13993003.01740-2016
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.