Back to Journals » International Journal of Nanomedicine » Volume 20
Development of a Ferritin-Based Nano-Allergen and Its Immunological Effects in vitro and in vivo
Authors Zhou D, Ren Y, Zhou Y, Liao Y, Cheng Q, Chen J, Yuan C, Zeng D, Cui Y ![]()
Received 5 April 2025
Accepted for publication 19 July 2025
Published 30 July 2025 Volume 2025:20 Pages 9505—9516
DOI https://doi.org/10.2147/IJN.S527210
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sachin Mali
Dongmei Zhou,1,* Yaning Ren,1,* Ying Zhou,2 Yuanfen Liao,1 Qi Cheng,1 Jinni Chen,3 Cunyin Yuan,1 Dan Zeng,4 Yubao Cui1
1Clinical Research Center, The Affiliated Wuxi People’s Hospital of Nanjing Medical University, Wuxi, 214023, People’s Republic of China; 2Department of Pediatrics Laboratory, The Affiliated Children´s Hospital of Jiangnan University, Wuxi, 214023, People’s Republic of China; 3Department of Respiratory, Hainan Women and Children’s Medical Center, Affiliated Pediatrics Clinical College of Hainan Medical University, Haikou, 570100, People’s Republic of China; 4Department of Allergy, Chongqing General Hospital, Chongqing University, Chongqing, 401147, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yubao Cui; Dan Zeng, Email [email protected]; [email protected]
Introduction: We aimed to develop recombinant allergens on the nanoscale by using self-assembly property of ferritin and to investigate their immune response and protective effect.
Methods: The cDNA encoding Ftn-Blo t 2 was synthesized and cloned into pET-21a (+) vector, then expressed in E. coli BL21 (DE3). Recombinant proteins (rBlo t 2, rFtn, and rFtn-Blo t 2) were purified and their immunoreactivity confirmed for immunoreactivity by IgE-ELISA and Western blot. In vitro, the immunogenicity of rFtn-Blo t 2 was assessed using BEAS-2B human bronchial epithelial cells by measuring epithelial cytokine responses. In vivo, a murine airway inflammation model was established via sensitization and challenge with crude B. tropicalis extract, followed by treatment with rFtn-Blo t 2 to investigate its effects on airway inflammation, Th1/Th2 cytokine profiles, and allergen-specific antibody production.
Results: The average diameter of rFtn and rFtn-Blo t 2 particles are (11.24 ± 1.31) nm and (16.00 ± 1.59) nm. The IgE- binding rates of rFtn-Blo t 2 and rBlo t 2 were 62.5% (15/24) and 58.3% (12/24), respectively. The expressions of IL-25, IL-33, and TSLP increased by 3.29, 3.43, and 2.22 folds after the addition of rFtn-Blo t 2 in co-culture with BEAS-2B. Mice challenged with rBlo t 2 had lower levels of cytokines IL-13 and IL-4 but higher levels of IFN-γ and TGF-β in alveolar lavage fluid compared to controls (p < 0.05). HE staining showed that compared with rBlo t 2 group, the inflammation level in lung tissue of rFtn-Blo t 2 group was reduced.
Discussion: A nano-scale allergen was successfully developed, providing a concept, a platform, and a paradigm for the construction of nano-scale allergens.
Keywords: Blomia tropicalis, nanometer, ferritin, allergic disease, Blo t 2
Introduction
House dust mites (HDMs) are a major source of indoor allergens and are regarded as the most prevalent and widespread indoor inhalation allergens globally.1 The most significant HDM species are Dermatophagoides pteronyssinus, D. farinae, Euroglyphus maynei, and Blomia tropicalis.2 An epidemiological survey of 1671 patients with asthma in 17 cities in East Africa was conducted by skin pricking test. The results showed the participants commonly reacted to house dust mites (66%), Blomia tropicalis (62%), and the German cockroach (52%).3 Additionally, the epidemiology of allergic sensitization in atopic children and adolescents in Curitiba, Southern Brazil revealed that 70.7% of patients with asthma and rhinitis tested positive for B. tropicalis.4 Furthermore, a study of 3,351 cases with allergic rhinitis in Shenzhen China identified positive for airborne allergens, while the positive rate of B. tropicalis (2231, 66.6%) was the top 1 inhalation allergen.5 Consequently, B. tropicalis is recognized as a crucial sensitizing mite species in China.
Currently, allergen-specific immunotherapy (AIT) is the only treatment that induces long-term responses against IgE-mediated allergies.6,7 Allergic patients commonly receive treatment with crude mite extracts.8,9 These extracts are mixtures containing multiple allergenic components, though often only one or a few of these components are relevant to the patient’s specific allergy. Additionally, crude mite extracts suffer from a lack of standardization, while the main components are insufficient, the type or content of components is unstable, and the immunogenicity of some components is low.10 Therefore, the use of crude mite extract in the clinical treatment process may lead to multiple serious and even life-threatening side effects, necessitating more intricate and careful management of these adverse reactions.11
With the rapid advancements in molecular biology technology, recombinant allergens come into being. These allergens offer several advantages over crude mite extracts, including higher purity, enhanced safety, optimal dosing, and ease of standardization, allowing them to completely replace traditional natural allergens in the clinical diagnosis and treatment of type I hypersensitivity diseases.12 However, the high purity and small molecular weight of recombinant allergens often result in poor immunogenicity.13 To counter this issue, adjuvants are employed, which are substances that boost the immunogenic strength of an antigen and amplify the specific immune response to an antigen, thereby eliciting a higher and longer term immune response in the body.14 Therefore, we can combine adjuvants and recombinant allergens to achieve an effective treatment for allergic reactions.
In recent years, nanotechnology has become increasingly prevalent in lesion detection, targeted therapy, drug carriers, and more. Nanoparticles offer the following advantages: Firstly, nanoparticles can protect antigens from rapid degradation and prolong the antigen release time, thereby inducing a long-lasting immune response. In contrast, free antigens (those not bound to the delivery system) degrade quickly after vaccination, leading to a relatively short-lived immune response.15 Secondly, nanoparticles can enhance antigen uptake and presentation by antigen-presenting cells (APC), with efficacy largely dependent on the type and size of nanoparticles.16 Additionally, 10–100 nm nanoparticles can easily navigate through the extracellular matrix, reach the lymph nodes, and are taken up by dendritic cells within the nodes. Nanotechnology thus opens up new avenues for developing novel allergen vaccines.17
Ferritin is a kind of nanomaterial that naturally exists in living organisms including animals, plants, and microorganisms, which plays a crucial role in stabilizing the internal environment of the body and supporting the immune system.18,19 It uniquely self-assembles into a cage-like structure composed of 24 subunits, with its N-terminal exposed, allowing for the fusion and display of foreign proteins. This feature makes ferritin an innovative platform for antigen presentation and immune response modulation.20–22 Wenjun Wang et al (2020) conducted ferritin loading experiments on hepatitis B virus capsid protein (preS1) and found that it induced high titer antibodies against preS1, showcasing its potential for disease prevention and treatment.23 Nie Jet al (2024) developed an intranasal multiepitope influenza nanovaccine using the Helicobacter pylori ferritin nanoplatform, and found that mucosal immunity induced by the multiepitope nanovaccine lasts for 6 months.24 Weidenbacher PA (2023) also reported a ferritin-based COVID-19 nanoparticle vaccine that elicited robust, enduring, and broad-spectrum neutralizing antisera in non-human primates.25 Building on these advances, this study constructed a ferritin-based self-assembled nanoscale fusion protein of B. tropicalis allergen Blo t 2 (rFtn-Blo t 2) to investigate its immunogenicity and therapeutic potential. We compared its effects with native rBlo t 2 on epithelial cell activation in vitro and in a murine airway inflammation model in vivo. Additionally, we assessed its ability to modulate Th1/Th2 immune responses, allergen-specific antibody production, and epithelial cytokine secretion, aiming to provide insights into the feasibility of ferritin-based nanoallergens for allergen-specific immunotherapy.
Materials and Methods
Construction of Expression Plasmids of pET21a (+) -Ftn-Blo t 2
Based on our previous work with recombinant allergen,26–28 we designed a ferritin-allergen fusion protein to enhance immunogenicity for potential allergen-specific immunotherapy. Total RNA of B. tropicalis was isolated and reverse-transcribed, followed by amplification of Blo t 2 cDNA (GenBank No. DQ677253) using RT-PCR with primers containing engineered restriction sites: the forward primer (5ʹ-CGCGCGGCAGCCATATGGGTGATGTCAAATTTACCGAT-3ʹ) incorporated an Nde I site (underlined) containing the ATG start codon, while the reverse primer (5ʹ-GACGGAGCTCGAATTCTTAATCGACAACCTCGGTGTTG-3ʹ) included an EcoR I site to enable downstream in-frame cloning. The Blo t 2 sequence was fused to the C-terminus of ferritin (GenBank No. NZ_CP006577.1) to create the Ftn-Blo t 2 fusion gene. For optimal protein expression, individual components were cloned into pET28a(+) vectors (pET28a(+)-Ftn and pET28a(+)-Blo t 2) to leverage its N-terminal His-tag and high expression efficiency, while the fusion construct was cloned into pET21a(+) (pET21a(+)-Ftn-Blo t 2) to utilize its C-terminal His-tag configuration that prevents interference with ferritin’s self-assembly domains. All constructs (detailed in Supplementary Figure 1) were verified by Sanger sequencing using T7 primers.
Protein Expression and Purification
The pET28a (+)-Blo t 2, pET28a (+)-Ftn, and pET21a (+)-Ftn-Blo t 2 were transformed into E.Coli BL21(DE3) receptive cells and expressed under isopropyl β-D-thiogalactoside (IPTG) induction. The recombinant proteins were collected and purified through GE Hitrap Talon Crude (Cytiva, No. 28953766) and Talon Superflow (No. 28-9537-66) column chromatography. The recombinant protein was identified through sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The purified recombinant protein was filtered with a 0.22-μm aseptic filter. Next, a protein concentration detection kit (No. P0010S; Beyotime) was used for protein quantification. The endotoxin content of our recombinant protein was determined to be <0.5 EU/μg using the ToxinSensor™ Chromogenic LAL Endotoxin Assay Kit (No. L00350; GenScript).
Characterizing Recombinant Proteins by Transmission Electron Microscope
A carbon-coated copper grid was treated with a plasma cleaner for 30 seconds to enhance surface hydrophilicity. Then, 10 μL of the recombinant protein sample was applied to the grid and incubated for 5 minutes at room temperature. Excess liquid was gently removed from the edge using filter paper. Subsequently, 5 μL of 1% (w/v) phosphotungstic acid was gradually added for negative staining over a period of 3 minutes. After removing the staining solution, the grid was air-dried. The structural morphology of the recombinant protein was visualized using transmission electron microscopy (TEM). Each sample was observed in triplicate to ensure reproducibility. Particle size analysis was performed using ImageJ software (version 1.8.0.112).
Identifying Ferritin/Allergen Fusion Protein by IgE-ELISA
Peripheral blood samples were collected from 29 children with allergic asthma (21 boys and 8 girls) and 6 healthy controls (5 boys and 1 girl) at the Hainan Women and Children’s Medical Center. This study involving human participants complied with the principles of the Declaration of Helsinki and was approved by the Human Ethics Committee of the Hainan Women and Children’s Medical Center (Haikou, China) under approval number HNWCMC-2022-44. Clinical characteristics of the serum donors are summarized in Supplementary Table 1. The serum characteristics are listed in Supplementary Table 1. The diagnosis of allergic asthma was based on the guidelines of the Chinese Medical Association guidelines.29 Serum allergen-specific IgE levels were determined using the AllergyScreen Test Kit (Mediwiss Analytic, Moers, Germany), with a B. tropicalis-specific IgE level > 0.35 IU/mL considered positive. According to our previous studies,9,26,100 μL of recombinant protein at a concentration of 5 μg/mL was added to 96-well ELISA plate and incubated overnight at 4 °C for coating. The wells were then blocked with 5% (w/v) skimmed milk in PBST for 2 hours at room temperature, followed by three washes with PBST. Subsequently, 100 μL of diluted serum samples (1:5 dilution), either from B. tropicalis-sensitized or non-sensitized individuals, were added to the wells and incubated overnight at 4 °C. After washing, HRP-conjugated anti-human IgE antibody was added and incubated at 37 °C for 2 hours. The plates were then washed three times, and 100 μL of TMB substrate was added for color development for 30 minutes at room temperature. The reaction was stopped by adding stop solution, and the optical density (OD) was measured at 450 nm using a microplate reader.
Identifying Ferritin/Allergen Fusion Protein by IgE-Western Blotting
Recombinant proteins were separated by SDS-PAGE and transferred onto PVDF membranes. The membranes were blocked and then incubated overnight at 4 °C with sera from B. tropicalis-sensitized or non-sensitized individuals, diluted 1:20. After washing with TBST, membranes were incubated with HRP-conjugated anti-human IgE antibody at room temperature for 2 hours, followed by three washes. ECL substrate was applied for 5 seconds, and signal detection was performed using a chemiluminescence imaging system (Tanon 5200, Shanghai, China).
Human Alveolar Epithelial Cells Exposed with Ferritin/Allergen Fusion Protein
BEAS-2B alveolar epithelial cells (Hunan Fenghui Biotechnology Co., Ltd., Hunan, China) were inoculated into four wells of a 6-well plate at a density of 15 × 104/well. Wells were divided into four groups and treated with PBS, rBlo t 2 (20 μg/mL), rFtn-Blo t 2 (20 μg/mL), and rFtn (20 μg/mL), respectively. After 24h, total RNA was extracted by FastPure Cell/Tissue Total RNA Isolation Kit V2 (vazyme, Cat# RC112-01), and reverse-transcribed into cDNA. Quantitative real-time PCR (qPCR) was performed on a QuantStudio 5 Real-Time PCR System (Thermo Fisher Scientific) using ChamQ Universal SYBR qPCR Master Mix (vazyme, Cat# Q711-02), according to the manufacturer’s instructions. Samples were analyzed in triplicate and normalized to GAPDH gene expression. The qPCR primers for specific target genes were designed based on the reported sequences and synthesized by Shenggong Bioengineering LTD. See Table 1 for a list of the oligonucleotide sequences.
 |
Table 1 Primer Sequences Used in Real-Time PCR |
BALB/c Mice Challenged with Ferritin/Allergen Fusion Protein and Evaluation
Following established protocols,30 we evaluated the immunogenicity of rFtn, rBlo t 2, and the fusion protein rFtn-Blo t 2 in mice. Female BALB/c mice (6–8 weeks old, purchased from Shanghai Laboratory Animal Center, SLAC) were housed under specific pathogen-free (SPF) conditions. All animal experiments were conducted in strict accordance with the Guide for the Care and Use of Laboratory Animals (9th edition, 2023, National Research Council) and the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines. The experimental protocol was approved by the Animal Ethics Committee of The Affiliated Wuxi People’s Hospital of Nanjing Medical University (Approval No. 42, 2022).
Twenty mice were randomly divided into the control group, rFtn group, rBlo t 2 group, and rFtn-Blo t 2 group with 5 mice in each group. On days 0, 7, 14, and 21, BALB/c mice were intraperitoneally injected with 50 μg B. tropicalis crude extract adsorbed on 2 mg Al(OH)3 to sensitize them. On days 28, 32, and 38, the mice were injected subcutaneously with 100 μg rFtn, 100 μg rBlo t 2, and 100 μg rFtn-Blo t 2 protein adsorbed on 2mg of Al (OH)3. After two more weeks, 50 μg B. tropicalis crude extract was given by nasal drip once a day for 6 days. Control mice were injected with PBS. On day 58, the mice were euthanized by cervical dislocation.
Serum-Specific Antibodies and Cytokines were Detected by ELISA
Peripheral blood of mice was collected by eyeball blood sampling, centrifuged at 3000 rpm for 10 min, and the supernatant was transferred to a new EP tube and stored at −80 ° C. 100 μL of recombinant protein with a concentration of 5 μg/mL was added to 96-well plates and incubated at 4°C overnight. On the next day, 100 μL 5% skim milk was added and closed at 37°C for 2 hours, and 100 μL animal serum was added at a 1:100 dilution ratio and incubated at 4°C overnight. HRP labeled IgG1, IgG2a, and IgE antibodies were added and incubated at 37°C for 2 h. 100 μL TMB was added and incubated for 30 min and 50 μL termination solution was added. The OD value was determined using a microplate reader at 450 nm. Serum cytokines IFN-γ, IL-4, IL-13, and TGF-β were detected by ELISA kit (GEMIC, China).
BALF Cellularity Analysis
The mice’s lungs were partially cleaned with 1 mL precooled PBS, and after three repetitions, the alveolar lavage fluid was obtained. Following centrifugation at 4000 rpm for 10 minutes, the precipitate was resuspended by adding 200 μL PBS. Differential cell counts were performed according to standard protocols.31
Histological Analysis
The fixed lungs were cut into 4–6 μm sections, placed on glass slides, and stained with hematoxylin (Thermo Fisher Scientific, Cat# 7221) and eosin (Thermo Fisher Scientific, Cat# 7111) (H&E). The bronchial wall thickness was quantified using ImageJ (version: V1.8.0.112). The inflammation score of lung tissue was assessed as described previously,32 based on the extent of inflammatory cell infiltration observed under light microscopy. The scoring criteria were as follows: Score 1: minimal infiltration affecting <10% of total lung tissue area; Score 2: mild infiltration affecting 10–25%; Score 3: moderate infiltration affecting 25–50%; Score 4: severe infiltration affecting >50% of the lung tissue area. No samples were assigned a score of 0.
Statistical Analysis
Statistical analysis was performed using GraphPad Prism 9 (GraphPad Software Inc., La Jolla, USA). All data were expressed as mean ± standard deviation (SD). For comparisons between two groups, unpaired two-tailed Student’s t-test was used. For multiple group comparisons, one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test was performed. A value of p < 0.05 was considered statistically significant.
Results
Construction of Expression Plasmids of pET21a(+) -Ftn-Blo t 2 and Its Expression and Identification
SDS-PAGE analysis revealed that the relative molecular weights of rFtn-Blo t 2, rFtn, and rBlo t 2 were approximately 35 kDa (Figure 1A), 20 kDa (Figure 1B), and 15 kDa (Figure 1C), respectively. The theoretical molecular weights, calculated based on their amino acid sequences using the ExPASy ProtParam tool (https://web.expasy.org/protparam/), were 34.95 kDa, 19.56 kDa, and 14.5 kDa, respectively, which are consistent with the observed SDS-PAGE results. Electron microscopy demonstrated that the self-assembled particles exhibit a uniform nanostructure with a high density of nanoscale particles within the field of view. Size analysis of the rFtn particles ranged from 8 to 15 nm, with an average diameter of (11.24±1.31) nm (Figure 1D and E). In contrast, rFtn-Blo t 2 particles varied from 13 to 20 nm, averaging at (16.00±1.59) nm (Figure 1F and G). Particles with sizes significantly outside these ranges were rare and were excluded from the statistical analysis; such particles likely represent minor populations due to incomplete assembly, aggregation, or imaging artifacts. The increased particle size of rFtn-Blo t 2 compared to the rFtn monocarrier protein indicates a successful conjugation of the two proteins.
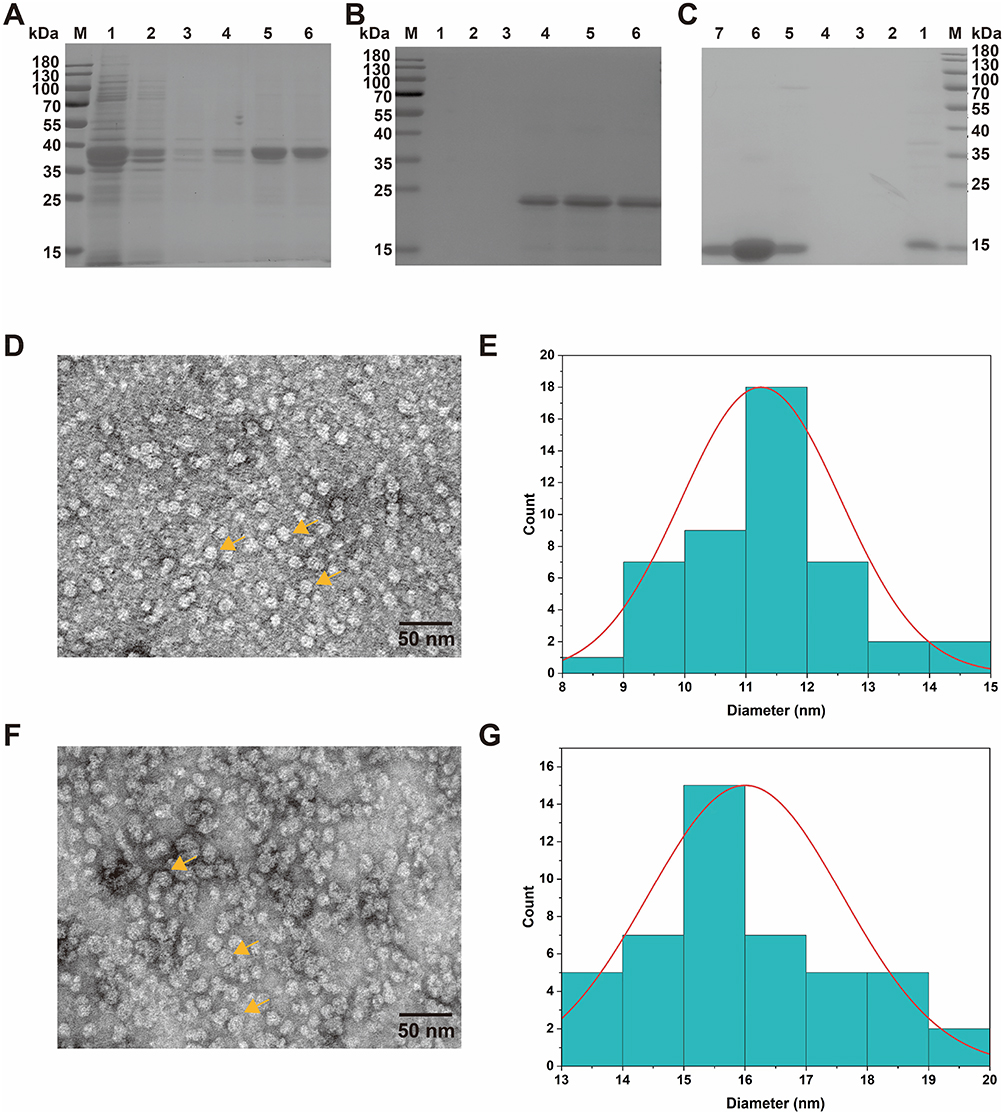 |
Figure 1 Expression of pET21a-rFtn-Blo t 2 in Rosetta2 (DE3) pLysS and purification. (A) SDS-PAGE identified rFtn-Blo t 2 purification. Lane M, protein marker; Lane1-6 for a stock solution, 10, 50, 100, 200, and 300 mM imidazole elutions. (B) SDS-PAGE identified rFtn purification. Lane M, protein marker; Lane1-6 for 20, 50, 100, 200, 300, and 500 mM imidazole elutions. (C) SDS-PAGE identified rBlo t 2 purification. Lane M, protein marker; Lane1-6 for 10, 20, 50, 100, 250, 300, and 500 mM imidazole elutions. (D) Image of rFtn transmission electron microscopy. (E) rFtn particle size analysis. (F) Image of rFtn-Blo t 2 transmission electron microscopy. (G) rFtn-Blo t 2 particle size analysis. |
Immunoreactivity of Ferritin/Allergen Fusion Protein
IgE-Western blotting analysis was conducted using 10 sera from children allergic to Blomia tropicalis. Among them, 6 sera (60%) showed specific IgE binding to rBlo t 2, while 8 sera (80%) reacted with the fusion protein rFtn-Blo t 2 (Figure 2A and B). IgE-ELISA was subsequently performed using 24 B. tropicalis-positive sera and 4 non-allergic control sera. The results showed that 14 out of 24 sera (58.3%) were positive for IgE binding to rBlo t 2, while 15 out of 24 (62.5%) were positive for rFtn-Blo t 2 (Figure 2C).
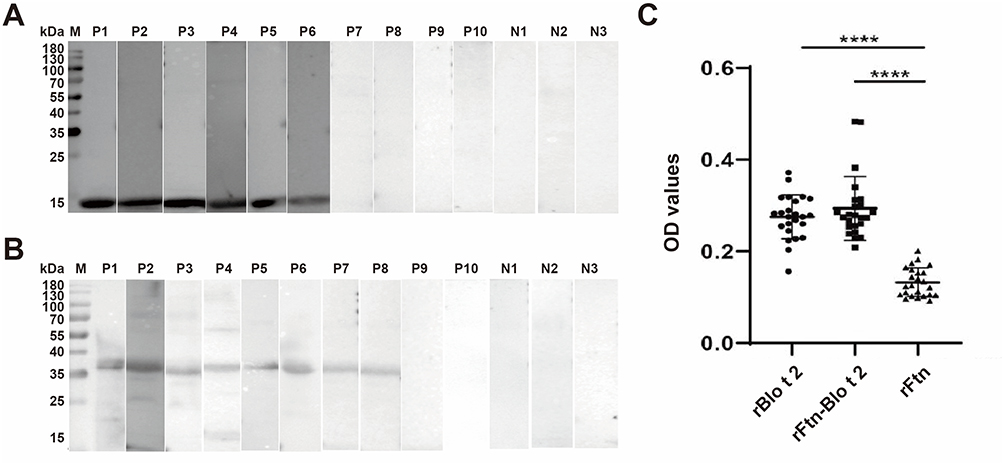 |
Figure 2 IgE binding reactivity of recombinant protein with IgE-Western blotting and IgE-ELISA. (A) IgE-Western blotting indicated IgE binding to rBlo t 2 in 60% (6/10) of B. tropicalis-positive sera from allergic children. Lane M, protein marker; Lane P1–10, patients’ sera; Lane N1, 2, 3, control sera. (B) IgE-Western blotting indicated IgE binding to rFtn-Blo t 2 in 80% (8/10) of B. tropicalis-positive sera from allergic children. Lane M, protein marker; Lane P1–10, patients’ sera; Lane N1, 2, 3, control sera. (C) IgE-ELISA indicated IgE binding to rBlo t 2 in 14/24 (58.33%), rFtn-Blo t 2 in 15/24 (62.5%) of B. tropicalis-positive sera from allergic children (**** p < 0.0001 vs the IgE-binding rate of rFtn in Blomia tropicalis-positive sera). |
rFtn-Blo t 2-Induced Inflammatory Cytokine Expression in BEAS-2B Cells
Compared to the control group, the expression levels of IL-25, IL-33, and TSLP were significantly increased after stimulation with rBlo t 2 and rFtn-Blo t 2, whereas the expression of cytokines was not changed by rFtn (Figure 3). We also found that rFtn-Blo t 2 demonstrated a more potent capacity to induce cytokine secretion than rBlo t 2.
 |
Figure 3 rFtn-Blo t 2 induced the production of IL-25 (A), IL-33 (B), and TSLP (C) in BEAS-2B. Expression levels of indicated genes were measured with qRT-PCR. Data are represented in terms of mean ± standard error of the mean (SEM) of three separate experiments. *p < 0.05, **p < 0.01, and ***p < 0.001. |
Identification of the Immunogenicity of rFtn-Blo t 2 in Mice
To assess the potential of rFtn-Blo t 2 to induce lung inflammation, mice were immunized with this allergen through a series of intraperitoneal and subcutaneous injections, followed by nasal exposure (Figure 4A). Throughout the process, no animal fatalities occurred. On day 58, mice were euthanized, and body and lung tissue weights were recorded. There were no significant differences in either body or lung tissue weights across the groups. The body weights of mice in each group were recorded as (20.35±0.16) g, (20.68±0.16) g (P= 0.0105), (20.33±0.17) g (P= 0.8387), (20.57±0.31) g (P= 0.1919), respectively. While the lung tissue weights were (0.52±0.02) g, (0.51±0.02) g (P= 0.5253), (0.51±0.01) g (P= 0.2476), (0.52±0.02) g (P= 0.8882), respectively (Figure 4B and C). After the rFtn-Blo t 2 intervention, the signs of airway wall thickening and inflammatory cell infiltration around the bronchus were alleviated (Figure 4D–F). Although not statistically significant, the total cell count in the rFtn-Blo t 2 group showed a 32% decrease compared to the PBS group, and neutrophils decreased by 44%. When compared to the rBlo t 2 group, the total cell count and neutrophils were reduced by 2.26×109/mL and 2.32×109/mL, respectively (Figure 4G). These results indicate a trend toward reduced airway inflammation following rFtn-Blo t 2 intervention.
 |
Figure 4 Therapeutic effect of rFtn-Blo t 2 in asthmatic mice. (A) Sensitizing and stimulating regimen in mice. (B) Body weight of the mice. (C) Lung weight of the mice. (D) HE staining of mouse lung tissue. (E) Quantitative analysis of total wall thickness of trachea in each group. (F) Inflammation score of lung tissue in mice. (G) Total cell, Neutrophils, and Lymphocyte counts in BALFs. (H) Expression of sIgE, sIgG1 and sIgG2a in mouse serum. (I) The ratio of IgG2a/IgG1 in mouse serum. Note: An IgG2a/IgG1 ratio higher than 1 indicates a biased Th1 immune response. (J) Expression of IL-13, IL-4, IFN-γ, and TGF-β in mouse serum. Note: * P<0.05, ** P<0.01, *** P<0.001, **** P<0.0001, n=5. |
Following the rFtn-Blo t 2 intervention, the serum IgE antibody level of the rFtn-Blo t 2 group decreased by 47% compared to the PBS group and by 31% Compared to the rBlot 2 group. Additionally, the induction of IgG subtypes showed that IgG2a levels were significantly increased by 31% in the rFtn-Blo t 2 group and by 35% in the rBlo t 2 group relative to PBS controls (Figure 4H–I). Furthermore, we calculated the IgG2a/IgG1 ratios to assess the bias of the immune response. The rFtn-Blo t 2 group showed a markedly higher IgG2a/IgG1 ratio compared to the PBS and rFtn groups, suggesting a shift toward a Th1-type response (Figure 4I). This finding complements the elevated IFN-γ levels observed in lung tissues and supports the immunomodulatory potential of the rFtn-Blo t 2 construct.
To further explore the cytokine profile in vivo, we assessed the levels of representative type 2 cytokines (IL-4, IL-13) and type 1 cytokines (IFN-γ, TGF-β) in the lungs of allergen-challenged mice. The results showed that IL-13 levels were significantly reduced in the rFtn-Blo t 2 group compared to the PBS group (P < 0.001), and IL-4 levels exhibited a downward trend. Conversely, the expression levels of IFN-γ and TGF-β exhibited a moderate but non-significant increase (P = 0.1347 and P = 0.0711, respectively), indicating a possible trend toward immune modulation. These findings further support the immunomodulatory potential of the fusion protein (Figure 4J).
Discussion
Blomia tropicalis is a predominant domestic mite species, especially abundant in tropical and subtropical regions worldwide, contributing significantly to the global prevalence of mite-related allergic diseases, particularly in China.27 The WHO/IUIS Allergen Nomenclature Sub-Committee has classified 26 allergen components of B. tropicalis, with Blo t 5 and Blo t 21 being the primary allergens.33,34 Blo t 2, a member of the MD-2-related lipid recognition (ML) protein family, is involved in lipid recognition and metabolism.35 An earlier report by Reginald K et al found that approximately 34% of adult dust-mite-sensitized individuals tested in Singapore exhibited specific IgE-reactivity to Blo t 2.35 Similarly, Mondol E et al reported a 54.3% prevalence of sensitization to Blo t 2 among 286 asthma patients.36 Consistently, in our study, over 50% of B. tropicalis-positive sera showed binding to both rBlo t 2 and rFtn-Blo t 2, confirming the immunoreactivity of these recombinant proteins.
Airway epithelial cells, as the first line of defense against inhaled allergens, are key regulators of immune responses and airway inflammation.37 Beyond their structural role, these cells are active participants in orchestrating immune responses by secreting key epithelial-derived cytokines, including thymic stromal lymphopoietin (TSLP), interleukin-25 (IL-25), and interleukin-33 (IL-33). These cytokines are pivotal in initiating and amplifying type 2 immune responses by activating downstream innate lymphoid cells (ILC2s) and promoting Th2 cell differentiation, thus driving allergic inflammation and airway remodeling.38,39 In this study, we observed that stimulation of bronchial epithelial BEAS-2B cells with the recombinant fusion protein rFtn-Blo t 2 significantly upregulated the expression of TSLP, IL-25, and IL-33 compared to rBlo t 2 alone, suggesting an enhanced pro-inflammatory stimulus by the nanoparticle-conjugated allergen. This heightened cytokine release may reflect the increased immunogenicity and cellular interaction potential of the ferritin-based nanoallergen, which could facilitate antigen presentation and immune cell recruitment in vivo. The amplified epithelial cytokine milieu is likely to contribute to the downstream recruitment and activation of eosinophils, mast cells, and other effector cells, key mediators in allergic airway disease pathogenesis. These in vitro findings provide mechanistic insight into how the nanoallergen may modulate epithelial-immune crosstalk and highlight its potential to influence airway inflammatory responses.
Allergic airway inflammation is driven predominantly by a Th2-skewed immune response, characterized by elevated production of IL-4, IL-5, and IL-13, and increased allergen-specific IgE, leading to eosinophilic inflammation, goblet cell metaplasia, and airway hyperresponsiveness.40–43 In our mouse model, exposure to crude B. tropicalis extract induced typical Th2-dominated pathology, evidenced by enhanced IL-13 levels, serum IgE production, and inflammatory cell infiltration in the bronchoalveolar lavage fluid. However, following rFtn-Blo t 2 intervention, IL-13 levels were significantly suppressed, and IL-4 expression showed a decreasing trend, suggesting that the nanoallergen may partially mitigate Th2-driven inflammation.
To assess whether this suppression of Th2 markers was accompanied by a shift toward Th1 immunity, we evaluated serum IgG subtypes and lung cytokines. In murine models, allergen-specific IgG2a production is considered a surrogate marker for Th1 polarization, as it is promoted by IFN-γ, while IgG1 is driven by IL-4.44–46 To assess whether this suppression of Th2 markers was accompanied by a shift toward Th1 immunity, we evaluated serum IgG subtypes and lung cytokines. In murine models, allergen-specific IgG2a production is considered a surrogate marker for Th1 polarization, as it is promoted by IFN-γ, while IgG1 is driven by IL-4.
It is worth noting that Th1 cytokines alone are not sufficient to determine therapeutic efficacy; functional improvements, such as reduction in airway hyperreactivity or restoration of epithelial integrity, are essential endpoints in defining clinical relevance. Due to equipment limitations, pulmonary function was not assessed in this study, which constrains our ability to definitively attribute a therapeutic role to rFtn-Blo t 2. Nonetheless, the immunological profiles observed—reduced IL-13, decreased IgE, increased IgG2a—represent a promising immunological signature that supports further investigation of rFtn-Blo t 2 as a potential candidate for allergen-specific immunotherapy.
We acknowledge several limitations in our study. The relatively small sample size may limit the statistical power to detect subtle but meaningful changes in cytokine levels. In addition, the lack of direct functional assessments, such as pulmonary function testing, precludes firm conclusions regarding clinical efficacy. These constraints will be addressed in future studies by expanding the animal cohort, extending observation periods, and incorporating comprehensive physiological and immunological assays. Nevertheless, the immunological trends observed—reduced IL-13, elevated IgG2a, and suppressed IgE—support the potential of rFtn-Blo t 2 as a novel candidate for allergen-specific immunotherapy. The ferritin-based nanoallergen platform may offer a strategic advantage in enhancing antigen delivery and modulating immune responses.
Conclusion
Our findings suggest that rFtn-Blo t 2 modulates airway inflammation by suppressing Th2 responses and promoting features associated with Th1 polarization. These results provide initial support for its potential application in allergen-specific immunotherapy, warranting further investigation in larger and functionally oriented studies.
Ethics Approval
This study was approved by the Human Ethics Committee of Hainan Women and Children’s Medical Center (Haikou, China) under approval number HNWCMC-2022-44. Informed consent was obtained from the parents or next of kin of the participants prior to the commencement of the study. Animal experimental procedures were approved by the Animal Ethic Committee at The Affiliated Wuxi People’s Hospital of Nanjing Medical University (Approval no. 2022042).
Acknowledgments
This work was supported by National Natural Sciences Foundation of China (NSFC81971511 and NSFC82100038), the Taihu Lake talent plan (Top-Level, no. 2020THRC-GD-7), the 333 project of Jiangsu Province in 2022 (ZUZHIBU 202221001).
Disclosure
The authors declare no competing interests in relation to this work.
References
1. Valenta R, Campana R, Focke-Tejkl M, Niederberger V. Vaccine development for allergen-specific immunotherapy based on recombinant allergens and synthetic allergen peptides: lessons from the past and novel mechanisms of action for the future. J Allergy Clin Immunol. 2016;137(2):351–357. doi:10.1016/j.jaci.2015.12.1299
2. Cardona-Villa R, Uribe-Garcia S, Calvo-Betancur VD, Cantillo JF, Fernandez-Caldas E. Efficacy and safety of subcutaneous immunotherapy with a mixture of glutaraldehyde-modified extracts of Dermatophagoides pteronyssinus, Dermatophagoides farinae, and Blomia tropicalis. World Allergy Organ J. 2022;15(9):100692. doi:10.1016/j.waojou.2022.100692
3. Kwizera R, Wadda V, Mugenyi L, et al. Skin prick reactivity among asthmatics in East Africa. World Allergy Organ J. 2020;13(6):100130. doi:10.1016/j.waojou.2020.100130
4. Elizabeth Maria Mercer Mourao, Nelson Augusto Rosario Conjunctival Provocation Test With Blomia tropicalis. Front Allergy. 2021;2:673462. doi:10.3389/falgy.2021.673462
5. Zhang N, Wu Y, Zhang Q, Wei Z, Liu Y. Analysis of inhalation allergen of patients with allergic rhinitis in Shenzhen. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2022;36(6):467–472. doi:10.13201/j.issn.2096-7993.2022.06.012
6. Huang HJ, Sarzsinszky E, Vrtala S. House dust mite allergy: the importance of house dust mite allergens for diagnosis and immunotherapy. Mol Immunol. 2023;158:54–67. doi:10.1016/j.molimm.2023.04.008
7. Asoudeh Moghanloo S, Forouzanfar M, Jafarinia M, Fazlollahi MR, Kardar GA. Allergen-specific immunotherapy by recombinant Der P1 allergen-derived peptide-based vaccine in an allergic mouse model. Immun Inflamm Dis. 2023;11(6):e878. doi:10.1002/iid3.878
8. Zhou Y, Li L, Yu Z, et al. Dermatophagoides pteronyssinus allergen Der p 22: cloning, expression, IgE-binding in asthmatic children, and immunogenicity. Pediatric Allergy Immunol. 2022;33(8):e13835. doi:10.1111/pai.13835
9. Zhou D, Ren Y, Zhou Y, et al. Expression, purification, and activity of novel allergen Tyr p 31 from Tyrophagus putrescentiae. Int J Biol Macromol. 2024;258(Pt 1):128856. doi:10.1016/j.ijbiomac.2023.128856
10. Kucuksezer UC, Ozdemir C, Cevhertas L, Ogulur I, Akdis M, Akdis CA. Mechanisms of allergen-specific immunotherapy and allergen tolerance. Allergol Int. 2020;69(4):549–560. doi:10.1016/j.alit.2020.08.002
11. Marth K, Focke-Tejkl M, Lupinek C, Valenta R, Niederberger V. Allergen, Peptides, Recombinant Allergens and Hypoallergens for Allergen-Specific Immunotherapy. Curr Treat Options Allergy. 2014;1(1):91–106. doi:10.1007/s40521-013-0006-5
12. Makatsori M, Pfaar O, Lleonart R, Calderon MA. Recombinant allergen immunotherapy: clinical evidence of efficacy--a review. Curr Allergy Asthma Rep. 2013;13(4):371–380. doi:10.1007/s11882-013-0359-7
13. Scholl I, Boltz-Nitulescu G, Jensen-Jarolim E. Review of novel particulate antigen delivery systems with special focus on treatment of type I allergy. J Control Release. 2005;104(1):1–27. doi:10.1016/j.jconrel.2004.12.020
14. Fierens K, Kool M. The mechanism of adjuvanticity of aluminium-containing formulas. Curr Pharm Des. 2012;18(16):2305–2313. doi:10.2174/138161212800166004
15. Schulze K, Olive C, Ebensen T, Guzman CA. Intranasal vaccination with SfbI or M protein-derived peptides conjugated to diphtheria toxoid confers protective immunity against a lethal challenge with Streptococcus pyogenes. Vaccine. 2006;24(35–36):6088–6095. doi:10.1016/j.vaccine.2006.05.024
16. Dhanasooraj D, Kumar RA, Mundayoor S. Vaccine delivery system for tuberculosis based on nano-sized hepatitis B virus core protein particles. Int J Nanomed. 2013;8:835–843. doi:10.2147/IJN.S40238
17. Kalkanidis M, Pietersz GA, Xiang SD, et al. Methods for nano-particle based vaccine formulation and evaluation of their immunogenicity. Methods. 2006;40(1):20–29. doi:10.1016/j.ymeth.2006.05.018
18. Pantopoulos K, Porwal SK, Tartakoff A, Devireddy L. Mechanisms of mammalian iron homeostasis. Biochemistry. 2012;51(29):5705–5724. doi:10.1021/bi300752r
19. Chen H, Tan X, Han X, et al. Ferritin nanocage based delivery vehicles: from single-, co- to compartmentalized- encapsulation of bioactive or nutraceutical compounds. Biotechnol Adv. 2022;61:108037. doi:10.1016/j.biotechadv.2022.108037
20. Jacob SI, Khogeer B, Bampos N, Sheppard T, Schwartz R, Lowe CR. Development and Application of Synthetic Affinity Ligands for the Purification of Ferritin-Based Influenza Antigens. Bioconjug Chem. 2017;28(7):1931–1943. doi:10.1021/acs.bioconjchem.7b00253
21. Zhang N, Yu X, Xie J, Xu H. New Insights into the Role of Ferritin in Iron Homeostasis and Neurodegenerative Diseases. Mol Neurobiol. 2021;58(6):2812–2823. doi:10.1007/s12035-020-02277-7
22. Zhu Y, Zhu Y, Cao T, et al. Ferritin-based nanomedicine for disease treatment. Med Rev. 2023;3(1):49–74. doi:10.1515/mr-2023-0001
23. Wang W, Zhou X, Bian Y, et al. Dual-targeting nanoparticle vaccine elicits a therapeutic antibody response against chronic hepatitis B. Nat Nanotechnol. 2020;15(5):406–416. doi:10.1038/s41565-020-0648-y
24. Nie J, Zhou Y, Ding F, et al. Self-adjuvant multiepitope nanovaccine based on ferritin induced long-lasting and effective mucosal immunity against H3N2 and H1N1 viruses in mice. Int J Biol Macromol. 2024;259(Pt 1):129259. doi:10.1016/j.ijbiomac.2024.129259
25. Weidenbacher PA, Sanyal M, Friedland N, et al. A ferritin-based COVID-19 nanoparticle vaccine that elicits robust, durable, broad-spectrum neutralizing antisera in non-human primates. Nat Commun. 2023;14(1):2149. doi:10.1038/s41467-023-37417-9
26. Zhou Y, Zhu K, Li Q, et al. Immunobiological properties and structure analysis of group 13 allergen from Blomia tropicalis and its IgE-mediated cross-reactivity. Int J Biol Macromol. 2024;254(Pt 3):127788. doi:10.1016/j.ijbiomac.2023.127788
27. Luo W, Zhang J, Zheng X, et al. Identification of rBlo t 41 with a chitin-binding type-2 domain: a novel major allergen from Blomia tropicalis. Int J Biol Macromol. 2024;262(Pt 1):129972. doi:10.1016/j.ijbiomac.2024.129972
28. Wang N, Zhou Y, Wu M, Zhu H, Cui Y. Expression, epitope prediction and IgE-binding of the Tyrophagus putrescentiae group 13 allergen. Exp Ther Med. 2020;20(5):21. doi:10.3892/etm.2020.9149
29. Respiratory Allergy Group of Chinese Society of A, Asthma Group of Chinese Thoracic Society CMA. Chinese guidelines for the diagnosis and treatment of allergic asthma (2019, the first edition). Zhonghua Nei Ke Za Zhi. 2019;58(9):636–655. doi:10.3760/cma.j.issn.0578-1426.2019.09.004.
30. Peng X, Liang Y, Yin Y, Liao H, Li L. Development of a hollow mesoporous silica nanoparticles vaccine to protect against house dust mite induced allergic inflammation. Int J Pharm. 2018;549(1–2):115–123. doi:10.1016/j.ijpharm.2018.07.047
31. Kalidhindi RSR, Ambhore NS, Sathish V. Cellular and Biochemical Analysis of Bronchoalveolar Lavage Fluid from Murine Lungs. Methods Mol Biol. 2021;2223:201–215. doi:10.1007/978-1-0716-1001-5_15
32. Wang D, Li W, Albasha N, et al. Long-term exposure to house dust mites accelerates lung cancer development in mice. J Exp Clin Cancer Res. 2023;42(1):26. doi:10.1186/s13046-022-02587-9
33. Kuo IC, Cheong N, Trakultivakorn M, Lee BW, Chua KY. An extensive study of human IgE cross-reactivity of Blo t 5 and Der p 5. J Allergy Clin Immunol. 2003;111(3):603–609. doi:10.1067/mai.2003.167
34. Gao YF, Wang de Y, Ong TC, Tay SL, Yap KH, Chew FT. Identification and characterization of a novel allergen from Blomia tropicalis: blo t 21. J Allergy Clin Immunol. 2007;120(1):105–112. doi:10.1016/j.jaci.2007.02.032
35. Reginald K, Pang SL, Chew FT. Blo t 2: group 2 allergen from the dust mite Blomia tropicalis. Sci Rep. 2019;9(1):12239. doi:10.1038/s41598-019-48688-y
36. Mondol E, Donado K, Regino R, et al. The Allergenic Activity of Blo t 2, a Blomia tropicalis IgE-Binding Molecule. Int J Mol Sci. 2023;24(6). doi:10.3390/ijms24065543
37. Liu C, Huang XL, Liang JP, et al. Serum‑derived exosomes from house dust mite‑sensitized Guinea pigs contribute to inflammation in BEAS‑2B cells via the TLR4‑NF‑kappaB pathway. Mol Med Rep. 2021;24(5). doi:10.3892/mmr.2021.12387
38. Roan F, Obata-Ninomiya K, Ziegler SF. Epithelial cell-derived cytokines: more than just signaling the alarm. J Clin Invest. 2019;129(4):1441–1451. doi:10.1172/JCI124606
39. Ito T, Liu YJ, Arima K. Cellular and molecular mechanisms of TSLP function in human allergic disorders--TSLP programs the “Th2 code” in dendritic cells. Allergol Int. 2012;61(1):35–43. doi:10.2332/allergolint.11-RAI-0376
40. Larche M, Robinson DS, Kay AB. The role of T lymphocytes in the pathogenesis of asthma. J Allergy Clin Immunol. 2003;111(3):450–463. doi:10.1067/mai.2003.169
41. Shang Q, Yu X, Sun Q, Li H, Sun C, Liu L. Polysaccharides regulate Th1/Th2 balance: a new strategy for tumor immunotherapy. Biomed Pharmacother. 2024;170:115976. doi:10.1016/j.biopha.2023.115976
42. Butcher MJ, Zhu J. Recent advances in understanding the Th1/Th2 effector choice. Fac Rev. 2021;10:30. doi:10.12703/r/10-30
43. Jacquet A. Innate immune responses in house dust mite allergy. ISRN Allergy. 2013;2013:735031. doi:10.1155/2013/735031
44. Durham SR, Shamji MH. Allergen immunotherapy: past, present and future. Nat Rev Immunol. 2023;23(5):317–328. doi:10.1038/s41577-022-00786-1
45. da Silva ES, de Santana MBR, Silveira EF, et al. The hybrid protein BTH2 suppresses allergic airway inflammation in a murine model of HDM-specific immunotherapy. Clin Exp Allergy. 2023;53(8):821–832. doi:10.1111/cea.14293
46. Allam JP, Novak N. Immunological mechanisms of allergen-specific immunotherapy. Hautarzt. 2017;68(4):265–270. doi:10.1007/s00105-017-3961-0
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.