Back to Journals » International Journal of Women's Health » Volume 17
Development and Application of Tumor Organoids: An Emerging Platform for Gynecological Cancers
Authors Hu Y, Chen W, Zhang X, Wu Y, Du F, Wang J, He M, Sun W
Received 12 September 2025
Accepted for publication 16 December 2025
Published 30 December 2025 Volume 2025:17 Pages 5761—5773
DOI https://doi.org/10.2147/IJWH.S567162
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Matteo Frigerio
Yu Hu,1,* Wei Chen,2,* Xiaojing Zhang,3,* Ying Wu,4 Furong Du,5 Jingrui Wang,5 Meijun He,4 Wei Sun6
1Laboratory Department, the People’s Hospital of Chuxiong Yi Autonomous Prefecture & the Fourth Affiliated Hospital of Dali University, Chuxiong, Yunnan, 675000, People’s Republic of China; 2Department of Gynecology, Clinical Oncology School of Fujian Medical University, Fujian Cancer Hospital, Fuzhou, Fujian, 350014, People’s Republic of China; 3Reproductive Health Research Institute, Henan Academy of Innovations in Medical Science, Zhengzhou, Henan, 450000, People’s Republic of China; 4Department of Obstetrics and Gynecology, the 920th Hospital of Joint Logistics Support Force, Kunming, Yunnan, 650032, People’s Republic of China; 5Department of Medicine, Kingbio Medical Co., Ltd., Chongqing, 401123, People’s Republic of China; 6Department of General Internal Medicine, Shanxi Province Cancer Hospital/Shanxi Hospital Affiliated to Cancer Hospital, Chinese Academy of Medical Sciences/Cancer Hospital Affiliated to Shanxi Medical University, Taiyuan, Shanxi, 030013, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Wei Sun, Department of General Internal Medicine, Shanxi Province Cancer Hospital/Shanxi Hospital Affiliated to Cancer Hospital, Chinese Academy of Medical Sciences/Cancer Hospital Affiliated to Shanxi Medical University, Taiyuan, Shanxi, 030013, People’s Republic of China, Email [email protected] Meijun He, Department of Obstetrics and Gynecology, the 920th Hospital of Joint Logistics Support Force, Kunming, Yunnan, 650032, People’s Republic of China, Email [email protected]
Abstract: Gynecological cancers pose great health threats to women, in which ovarian cancer (OC), cervical cancer (CC) and endometrial cancer (EC) are most common. Conventional treatment modalities for gynecological cancers, such as surgery, chemotherapy and radiotherapy, are usually affected by multiple factors like drug resistance and recurrence. Organoids, a three-dimensional culture system derived from stem cells, have been demonstrated to recapitulate the biological structure and function of human organs and gradually used in the treatment of various cancer types, including gynecological cancers. The organoids established from gynecological cancers have patient tumor-dependent morphology and disease characteristics while retaining the tumor’s marker expression and genomic profiling, and present heterogeneous inter- and intra-patient drug responses, offering novel insights into drug response of the individual patients. This review provides an overview of recent advancements in the development and application of organoids from gynecological cancers, promoting the understanding of the mechanism underlying tumorigenesis, drug screening prediction and personalized treatment application in gynecological cancers.
Keywords: gynecological cancers, tumor organoids, drug screening, molecular mechanism, precision medicine
Introduction
Gynecological cancers are the unregulated growth of neoplastic cells arising in the ovaries, uterus, cervix, fallopian tubes, vulva and vagina, among which ovarian cancer (OC), cervical cancer (CC) and endometrial cancer (EC) are most common.1 According to the estimates of the International Agency for Research on Cancer, the incidence of gynecological cancers is increasing year by year, posing a serious threat to women’s lives and health.2 Currently, comprehensive approaches mainly based on surgery, chemotherapy, radiotherapy and targeted therapy are commonly used for gynecological cancers.3,4 Nevertheless, for gynecological cancers, complete surgical resection is challenging due to the unique anatomical relationship, consequently resulting in high recurrence rates. Although chemotherapy plays a crucial role in improving surgical cure rates and reducing recurrence rates, some patients show lowly sensitive to chemotherapeutic agents or develop resistance, limiting the survival improvement.5 Additionally, the efficacy of radiotherapy is also compromised in the cases of poor anatomical conditions, extensive pelvic adhesions or retroperitoneal lymph node metastasis.6,7 Therefore, it is very necessary to seek more effective and precise treatment methods to improve the survival outcomes of patients with gynecological cancers.
Conventional cancer cell lines and patient-derived xenograft (PDX) models play important roles in drug screening, but they both have certain limitations. Cancer cell lines cannot simulate the tumor microenvironment (TME) of tumor growth during culture, thus losing the physiological characteristics and heterogeneity of tumor cells.8 Although PDX models can fully recapitulate the basic process of the tumor interaction with in vivo environment, they are time-consuming, expensive and may undergo mouse-specific tumor evolution rendering them unable to reflect the pathogenic process of patients.9,10 Organoids, a three-dimensional (3D) culture system derived from stem cells, have been demonstrated to recapitulate the biological structure and function of human organs. They can be established from different sources of stem cells, such as induced pluripotent stem cells (iPSCs), adult stem cells (ASCs) and embryonic stem cells (ESCs), and each of them had unique advantages in cancer research (Figure 1). To investigate the mechanisms of tumorigenesis, screen drugs, or perform functional genomics studies, PSC and ESC-derived organoids are usually genetically engineered to simulate oncogenic mutations.11 In precision oncology, ASC-derived organoids are valuable due to their capability of maintaining genomic integrity and TME of the original tumor. Moreover, the recently emerging integrated technology of organoids with TME further promotes the organoid capability of capturing tumor-immune interactions and predicting the treatment response.12 Hence, the organoid system as a powerful model shows promise in understanding tumor biology and guiding personalized treatment.
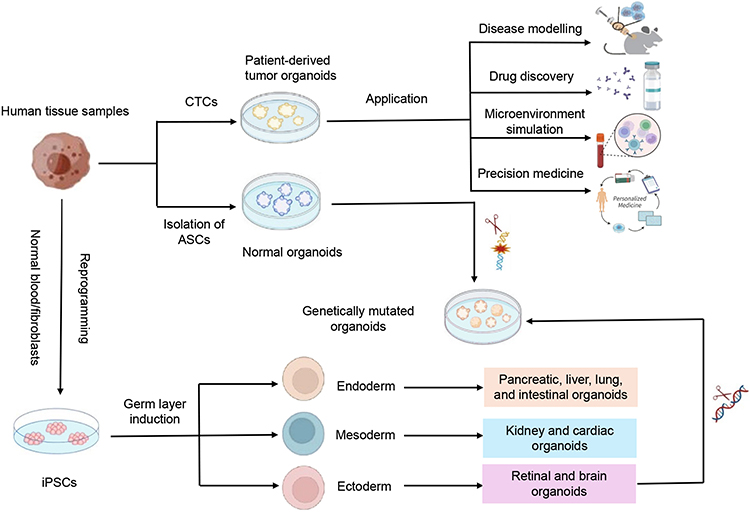 |
Figure 1 Application of organoids derived from adult stem cells (ASCs) and pluripotent stem cells (PSCs). |
Over the past few years, the patient-derived tumor organoids (PDTOs) have been gradually used in the treatment of various cancer types, including gynecological cancers.13–16 By establishing patient-derived OC organoids, Maenhoudt et al found that these organoids could not only have patient tumor-dependent morphology and disease characteristics but also retain the tumor’s marker expression and genomic profiling.17 de Witte et al further demonstrated that OC organoids showed heterogeneous inter- and intra-patient drug responses while mimicking the clinical response, offering novel insights into drug response of the individual patient with OC.16 Importantly, the ovarian organoid-based drug sensitivity testing was closely associated with the patient clinical response, which promoted the personalized screening of drugs and application of precision medicine.18 Overall, PDTOs show a promising prospect in both basic and clinical studies of gynecological cancers. Here, we reviewed the culture techniques and application strategies of organoids from gynecological cancers to enhance the realization of precision medicine in gynecological cancers.
Culture Techniques for Gynecological Cancers
Currently, there are three common approaches for establishing PDTOs, including Matrigel culture, air-liquid interface (ALI) culture and microfluidic culture, in which Matrigel culture is most typical and frequent. The dissociated glandular epithelial cells or cell clusters are resuspended in extracellular matrix (ECM) hydrogel, such as basement membrane extract (BME) or Matrigel, and then seeded into 24-well or 48-well plates to form domes. Finally, an appropriate volume of organoid culture medium is added to submerge the domes for cultivation.19 Compared with the traditional 2D culture model, ALI and microfluidic culture techniques are innovative as they can mimic the natural growth environment of tumor cells in a 3D space, which is closer to the real situation in vivo.20 Then, the application of these culture techniques in gynecological cancers was described.
Matrigel Culture
As a scaffold for cell growth, the ECM not only provides a site for cell attachment, growth, and differentiation but also simulates the natural environment for cell growth. It effectively promotes the formation of organoids and ensures the maintenance of their physiological functions. Rinehart et al first developed a method for in vitro culture of human endometrial glandular epithelium, in which a serum-free culture medium combined with Matrigel was used as a coating for cell culture plates, allowing the cells to grow in a monolayer and then spontaneously forming glandular or organoid structures.21 Subsequently, the culture method for EC organoids was further improved. Maru et al cultured the dissociated tumor cells on the solidified BME overnight and then covered the living cells with another layer of basement membrane extract, which significantly increased the success rate of organoid culture from 45% to 90%.22 In the study made by Boretto et al, the effect of the medium components on the growth of EC organoids was observed, and the results showed that RSPO1, a WNT signaling enhancer, was an indispensable factor for organoid growth. Meanwhile, a long-term in vitro proliferation of EC organoids was also achieved through adjustment of the medium components.23
Kopper et al proposed an effective culture protocol for OC organoids. They first collected tissues from primary tumors or metastatic lesions, dissociated the tumor cells, and then suspended these cells in the basement membrane extract.24 By coating and supplementing the culture medium, they successfully established OC organoid models. Notably, hydrocortisone, forskolin, and heregulin β-1 were added to the culture medium, which significantly improved the formation efficiency of OC organoids. Additionally, Maru et al first established a PDTO model from cervical clear cell carcinoma, in which patient-derived cells were seeded on the solidified Matrigel and cultured in serum-free medium, resulting in the formation of small or large solid structures with multiple buds.25 However, the organoids established by Matrigel culture only preserve epithelial components and lack stromal components.26 Therefore, it is necessary to establish a co-culture system of organoids with fibroblasts or immune cells to study the interactions between different types of cells.26,27
ALI Culture
Unlike the Matrigel culture, the ALI culture retains not only epithelial cells but also stromal cells and immune cells, allowing for accurately recapitulating the stem cell niche in the organoid culture system and facilitating the investigation of in vitro interactions between tumor and immune cells in the tumor microenvironment.28,29 Without the need for exogenous growth factor supplementation, Li et al successfully established the robust organoids with both epithelial and mesenchymal components from the small intestine, colon, and stomach using the ALI culture method that was realized by forming inner and outer culture dishes through a transwell chamber.30 Afterwards, the ALI method was extended to culture clinical tumor samples as the PDTOs. In the environment of the ALI culture, fibroblasts and various immune cell populations, such as T cells, B cells and macrophages, from primary tumor tissues could continuously proliferate for several weeks, while retaining the diversity of T cell clones in the original tumor.29
Currently, the ALI culture has been gradually applied in various tumors, including renal cell carcinoma and colorectal cancer,31,32 but its application in gynecological cancers remains in the initial stage. Dong et al developed a perfusion air culture (PAC) system that could provide a continuous and controllable oxygen medium and drug supply, adaptable for assessing drug responses in a tissue-specific microenvironment.33 In this study, fresh human ovarian tumor tissues were alternatively cultured on a self-made PAC system. It was observed that cisplatin treatment could induce increased cleavage of caspase-3 and PD-L1 expression, highlighting the heterogeneity of responses to drugs in different patients.33 In another study, novel endometrial assembloids were developed by combining endometrial epithelial cells and stromal cells based on an improved matrix and ALI culture method, which not only recapitulated the anatomical structure of the human endometrium, cellular composition, and hormone-induced menstrual cycle changes but also exhibited the gene expression profiles and dynamic ciliation.34 These groundbreaking studies provide new perspectives for subsequent research and lay a solid foundation for expanding future therapeutic strategies.
Microfluidic Culture
Microfluidics is a technology that manipulates fluids in the volume range from nanoliters to microliters through micron-scale channel networks.35 By dynamically replenishing the culture medium, microfluidic technology enables precise control of organoids and provides dynamic physical conditions, allowing organoids to be cultured uniformly in large quantities while maintaining a complex microenvironment.36 The high-throughput microfluidic platform can realize the repeatable and mass production of homogeneous organoids, thus facilitating high-throughput drug screening. In a previous study, a microfluidic platform termed EVATAR was created after reproductive tract tissues and peripheral organs were integrated, which represented a powerful tool that allowed organ-organ integration of hormonal signaling as a phenocopy of menstrual cycle and pregnancy-like endocrine loops, with great potential in drug discovery and toxicology studies.37 To date, however, EVATAR is only used in the non-malignant reproductive tract. In the future, it will be interesting to investigate its application in gynecological malignancies.
Application of Organoids from Gynecological Cancers
Compared with conventional cancer cell lines, PDTOs can preserve the histological, genetic and molecular characteristics of the original tumors more accurately, thus allowing for a more profound understanding of the molecular mechanisms underlying gynecological cancers (Figure 2). As a promising platform for drug screening and personalized medication evaluation, PDTOs offer a closer approximation to clinical conditions, promoting their application in research and therapeutic development.
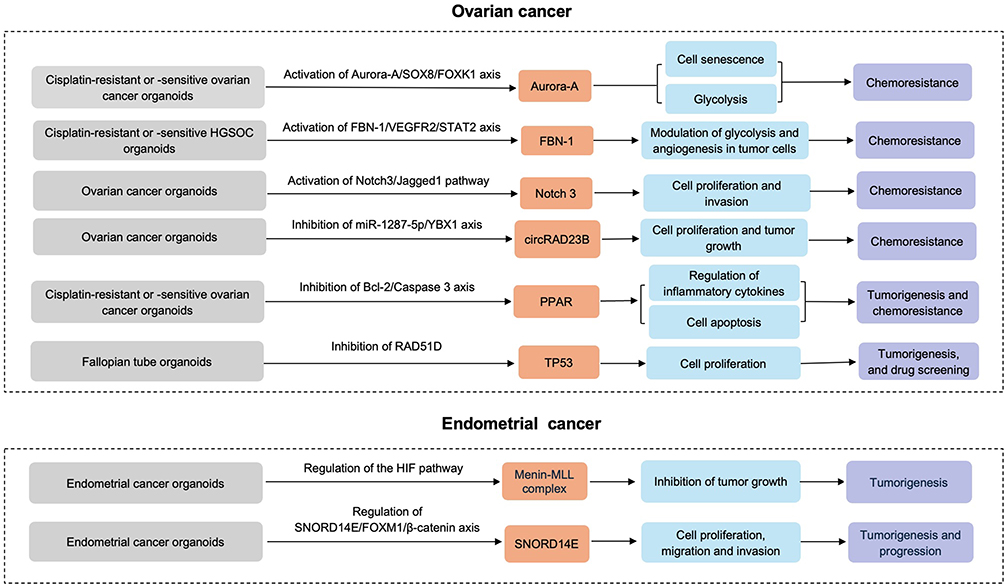 |
Figure 2 Molecular mechanisms underlying ovarian cancer and endometrial cancer based on the organoids. |
OC
OC is one of the most lethal gynecological malignancies, with over 240000 new cases diagnosed annually and a 5-year survival rate of less than 45%.38 Due to the presence of high heterogeneity in OC, accurate assessment of tumor heterogeneity is particularly crucial for predicting drug resistance and exploring effective therapeutic approaches.
OC organoids, a 3D dynamic tumor model, can be established from tumor tissues, pleural effusions or ascites of OC patients, consequently recapitulating the tumor clonal heterogeneity, tumor microenvironment, cell-cell and cell-matrix interactions. It has been demonstrated that OC organoids have a high concordance with original tumors in histologic, genomic and transcriptomic features.39 Regarding the replication of primary tumor characteristics and treatment responses, PDTOs from ovarian clear cell carcinoma were superior to conventional 2D cell lines, and they could reflect clinical drug resistance.40 Notably, strong spheroid and tumor-forming capabilities and higher drug resistance were shown in the iOVCAR-3-OSKM model that could develop into organoids to simulate the poor prognostic features of OC while maintaining the ovarian lineage differentiation abilities, providing a potential strategy for personalized treatment in OC.41
Drug Screening
Over the past few years, numerous studies have focused on establishing and applying OC organoid models to advance the clinical treatment.17,42–45 Kopper et al established 56 organoids from different pathological subtypes of OC, with a success rate of up to 65%.24 Meanwhile, they also demonstrated that OC organoids could be used for drug screening and recapitulate the responses of different tumor subtypes to the standard platinum-based chemotherapy. Phan et al developed a small-scale high-throughput organoid screening system by seeding cells around the rim of the wells, and successfully cultured organoids from 2 ovarian and 1 peritoneal high-grade serous carcinomas and 1 carcinosarcoma of the ovary, laying the foundation for high-throughput drug screening.42 By developing expandable OC organoids, Nanki et al performed drug sensitivity testing on 23 FDA-approved drugs and found that these organoid models had great potential in screening personalized therapeutic drugs.43 Additionally, short-term organoid culture from high-grade serous ovarian carcinoma (HGSOC) malignant effusions can also be used as a potential platform for drug sensitivity testing.44
OC organoids can not only retain the patient tumor-dependent morphology and disease characteristics but also capture the original tumor marker expression and mutational landscape. In a previous study, HGSOC organoids showed tumor-specific sensitivity to clinical chemotherapeutic drugs in HGSOC.17 Moreover, a functional defect in homologous recombination (HR) in the HGSOC organoids was associated with the sensitivity of poly-ADP ribose polymerase inhibitors (PARPi), regardless of DNA repair gene mutational status.45 Importantly, genomic analysis combined with organoid functional testing allowed for the identification of targetable DNA damage repair defects, offering a rapid means of evaluating targetable defects and more appropriate treatment options for OC.
Currently, combination therapy is frequently used for the treatment of OC. By assessing the ability of OC organoids to predict clinical drug response and functional consequences of tumor heterogeneity, de Witte et al demonstrated that OC organoids could faithfully recapitulate the patient response to neoadjuvant carboplatin/paclitaxel combination therapy, and inter- and intra-patient heterogeneity of drug response to both chemotherapeutic and targeted drugs was present in organoids,16 making the organoid model more closer to the clinical application.
Due to lack of stromal components, PDTOs are limited in studying the effect of tumor microenvironment on tumor behaviors. Several studies have shown the potential of PDTOs in studying the mechanisms of immunotherapy efficacy and resistance when being cocultured with immune cells.16,17,29,45–47 Nevertheless, the value of adding immune checkpoint inhibitors, such as such as cytotoxic T lymphocyte associated antigen-4 (CTLA-4) and programmed death-1 (PD-1)/programmed death ligand-1 (PD-L1) inhibitors, to standard treatment remains unclear, and is being emphasized in ongoing clinical trials for OC. Combining CTLA-4 and PD-1 inhibitors may promote the immunotherapy efficacy in epithelial ovarian cancer, and the relevant studies on combination of immune checkpoint inhibitors with other emerging treatment methods are underway.48 Notably, there is no solid evidence regarding the co-culture of PDTOs with immune cells in predicting immunotherapy efficacy. In the future, the co-culture technology of PDTOs with immune cells may be attempted to predict the immunotherapy efficacy in epithelial ovarian cancer.
Mechanisms Underlying Drug Resistance and Tumorigenesis
The advent of organoid technology has revolutionized cancer research, particularly for gynecologic cancers. Due to maintenance of the genetic and phenotypic features of the original tumors, PDTOs have been an ideal model to study the tumor cell interactions with the microenvironment, essential for understanding the potential mechanisms underlying drug resistance, tumorigenesis, progression, and metastasis.
Based on OC organoids, Aurora-A/SOX8/FOXK1 signaling axis was identified to enhance cisplatin chemoresistance through inhibition of cell senescence and induction of glucose metabolism.49 Wang et al found that the expression of FBN1 was significantly increased in cisplatin-resistant OC organoids and tissues, while knockout of FBN1 could decrease glycolysis, damage angiogenesis, and enhance the sensitivity to cisplatin.50 Through modulation of glycolysis and angiogenesis, FBN-1/VEGFR2/STAT2 signaling axis facilitated cisplatin chemoresistance in OC organoids. These findings suggest a novel mechanism of cisplatin resistance and offering potential therapeutic targets to overcome chemoresistance in OC.
Akt-activated endothelial cells could form resistant vascular spheroids when cocultured with OC cells. During vascular spheroids, FGF2, Pentraxin 3, PD-ECGF and TIMP-1 were all involved, and Notch3/Jagged1 pathway was also identified in the interactions between OC cells and endothelial cells, which was associated with the proliferation of OC cells and peritoneal invasion.51 Based on OC organoids, circRAD23B was found to facilitate proliferation and decrease sensitivity to carboplatin, consistent with in vivo findings.52 Meanwhile, the upregulation of miR-1287-5p could partially reverse the pro-proliferation and carboplatin resistance mediated by circRAD23B. Moreover, there is also a study suggesting that the expression of SGK1 and VEGFA was increased in cisplatin-resistant and cisplatin-sensitive OC organoids, respectively.53 Interestingly, the model genes were mainly enriched in the PPAR signaling pathway, a pivotal regulator in chemoresistance and tumorigenesis. By establishing organoids from human fallopian tube epithelium, Dai et al observed that TP53 deletion could salvage the adverse effects of RAD51D deletion on the proliferation, stemness, senescence, and apoptosis in fallopian tube epithelial organoids.54 Except for downregulating cilia assembly, development and motility, TP53 and RAD51D co-deletion also upregulated various HGSOC-associated pathways, including the IL-17 signaling pathway, thereby enhancing the sensitivity to platinum, PARPi and cell cycle-associated medication.54 The findings above highlight the application of RAD51D-mutant fallopian tube epithelial organoids in the early detection of tumorigenesis, mechanistic exploration, and drug screening.
CC
CC is globally the fourth most common malignancy in individuals assigned female at birth, with 85% of cases occurring in developing countries.55 With the widespread use of human papillomavirus (HPV) vaccines, precancerous screening and timely surgical treatment, the incidence and mortality of CC have been decreased significantly, but for metastatic and recurrent CC it is still difficult to be cured completely. Currently, there is a lack of biomarkers that can accurately evaluate the therapeutic effect of cervical cancer. Therefore, there is an urgent need for a new clinical model to predict the treatment response based on tumor characteristics.
Drug Screening
CC organoids can be successfully established using Pap smear-derived cervical tumor tissues, which can not only form the tumors but also faithfully retain the key features of the original tumors, indicating the tumorigenic capability of the organoid models.56 Maru et al first cultured the organoids from a patient with cervical clear cell carcinoma (cCCC) and demonstrated that the cCCC organoids resembled the original cCCC components in histology, immunostaining profile, and genome-wide copy number variations.25 Notably, these organoids were sensitive to major chemotherapeutic drugs and MET inhibitors. By constructing the organoids from 4 patients with different types of cervical cancer, Seol et al found that the organoids could preserve the major elements of the original tumors, such as DNA copy number and mutational profiles, and KMT2C mutations in different types of CC organoids were identified, indicating a latent therapeutic target.26 In addition, 7 drugs with growth inhibition in CC organoids were selected from 171 FDA-approved drugs. Importantly, squamous cell carcinoma and villoglandular carcinoma showed sensitive to radiotherapy, while adenocarcinoma and large-cell neuroendocrine carcinoma were resistant to radiotherapy.25 These results further unveil that PDTOs from CC may serve as a useful platform for drug screening and radiation sensitivity testing. Di Fonte et al performed drug sensitivity testing based on CC organoids and revealed that propranolol, a β-blocker, could enhance the therapeutic effect of trabectedin, an oncogene transcription inhibitor, in the treatment of CC.57 Kusakabe et al established HPV18-positive small cell carcinoma of the uterine cervix (SCCC) organoids, and new therapeutic targets were identified using transcriptomics and genomics. Drug sensitivity testing revealed strong anti-cancer effects of a KRAS pathway inhibitor on SCCC organoids, which were also validated in the PDX model.27 Therefore, establishment of PDTOs can be effectively used for drug sensitivity testing, promoting the development of personalized treatment in cervical cancer.
Radiation therapy is one of the main treatment modalities for CC, but patients with CC exhibit varying degrees of radiosensitivity. The measurement of radiation sensitivity using organoids provides a new possibility for predicting the efficacy of tumor radiotherapy and exploring the mechanisms of radiation resistance. In a previous study, the survival outcomes of patients with squamous mesenchymal subtype were worse than those with other 2 molecular subtypes. Based on CC organoids, they demonstrated that an increase in mesenchymal components in cancer cells and elevated glycolysis levels enhanced radiation resistance, thereby reducing radiotherapy efficacy.58 Huang et al compared the radiation sensitivity of 14 independent CC organoids under different radiation doses and observed differences in radiosensitivity among CC organoids derived from different patients, which was further verified by transplanting the organoids into immunodeficient mice.59 Notably, the efficacy of chemotherapy-sensitized radiotherapy was also determined using the PDTOs from 2 patients with advanced cervical cancer, and consistent treatment response with experimental prediction was obtained in the subsequent clinical follow-up.
The immune system plays a crucial role in the process of tumor malignant transformation. The interaction between HPV-infected cervical cells and the immune microenvironment influences the progression of cervical cancer. The main effector cells of anti-tumor immunity include natural killer cells and T cells, and the co-culture system of organoids and immune cells provides a new platform for immunotherapy.60 To simulate individual responses to adoptive T cell therapy (ACT), Huang et al cocultured the expanded tumor-infiltrating lymphocytes with paired organoids. By observing the specificity of CC organoids from different patients, they demonstrated that TTN mutations might be a potential predictive factor for ACT in CC.59 Furthermore, Dong et al also constructed a coculture system of γδT cells with CC organoids, which could be used to validate the immunotherapeutic effect of γδT cells on cervical cancer, offering a novel in vitro model to optimize the patient-specific immunotherapy in CC.61
Mechanisms Underlying Tumorigenesis
By developing an effective method for organoid culture of gynecological cancer, Maru et al observed that the organoids established could preserve various characteristics of the original tumor, such as histological features, mutation profiles and intra-tumoral heterogeneity, which was helpful for in-depth understanding of the pathogenesis of gynecological cancers.22 Lohmussaar et al generated the organoids from cervical squamous cell carcinoma and adenocarcinoma and found that these organoids highly resembled the original tumor tissue in transcriptomic characteristics, including the tumor genomic features mediated by HPV genes, indicating that CC organoids may serve as an effective platform for studying sexually transmitted infections and the occurrence and development of cervical cancer.56
HPV, a circular DNA virus, has over 100 genotypes, in which HPV 16/18 is most aggressive.62 These genotypes are conductive to cervical tumorigenesis after integrating with the host genome. HPV-related CC involves a multi-stage carcinogenic process. HPV viral oncoproteins E6 and E7 inhibit the tumor suppressor proteins p53 and Rb in host cells, respectively (Figure 3). The integration of HPV-DNA into the human genome triggers genetic alterations, such as oncogene amplification and chromosomal rearrangement. The accumulation of host cell DNA changes induced by persistent HPV infection play a crucial role in the formation of invasive CC.63 Toyohara et al established an HPV18-transfected organoid system of the cervical transformation zone. Through RNA sequencing and functional experiments, they identified the NPM3 gene as a new therapeutic target for HPV-related cervical adenocarcinoma.64 In another study, CC organoids were cocultured with peripheral blood immune cells (PBMCs) activated by HPV antigenic peptides, the results indicated that the cocultured organoids responded to immunized PBMCs differently, highlighting the feasibility of organoids as a credible model for studying virus-specific T-cell responses and screening therapeutic HPV vaccines.65
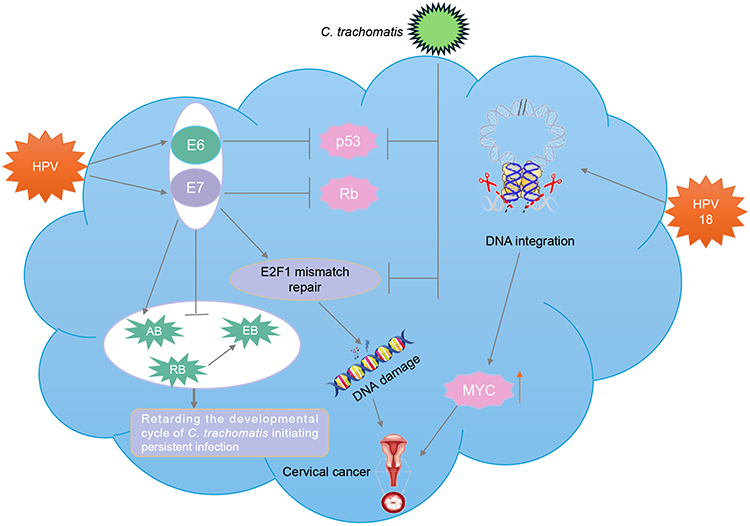 |
Figure 3 The pathogenic mechanism of cervical cancer based on the organoids co-infected with HPV and C. trachomatis. HPV16 E6/E7 viral oncoproteins impede the tumor suppressor proteins p53 and Rb, respectively. E2F1 target genes can be significantly upregulated by HPV infection, while downregulated by C. trachomatis infection. Additionally, (C) trachomatis infection can also result in p53 degradation. During co-infection, HPV E6 and E7 can increase the number of aberrant chlamydial developmental forms, while suppressing the redifferentiation of non-infectious replicative reticulate bodies (RB) into non-replicative elementary bodies (EB), thereby retarding the developmental life cycle of C. trachomatis initiating persistent infection. |
Recently, cervical organoids have been gradually used for studying the integration of HPV18 DNA into the host cell genome and the effect of HPV co-infected with Chlamydia trachomatis (C. trachomatis). In a previous study, RNA-seq analysis of the organoids from HPV18-positive SCCC revealed integration of HPV18 into chromosome 8 at 8q24.21 could upregulate the expression of the proto-oncogene MYC, indicating that integration of HPV18 into chromosome 8 may induce DNA modifications in the host cell genes, consequently enhancing the expression of the proto-oncogene MYC and triggering the development of CC.27 Based on the organoids co-infected with HPV and C. trachomatis, Koster et al found that this co-infection model could induce the unique cell reprogramming in the host, in which multiple genes were upregulated or upregulated. Surprisingly, C. trachomatis inhibited the HPV-induced mechanisms that preserved cellular and genome integrity, involving mismatch repair in the stem cells.66 These findings unveil that HPV E6/E7 co-infected with C. trachomatis may be unfavorable for cellular and genomic stability, potentially contributing to the progression of cervical cancer.
EC
The incidence and mortality of endometrial cancer, a malignancy of the inner epithelial lining of the uterus, are rising worldwide. Historically, EC is classified into type I (correlation with unopposed estrogen stimulation, containing more common low-grade cells with a favorable prognosis) and type II (not estrogen driven, containing less common high-grade cells with an unfavorable prognosis).67 Currently, the treatment options for advanced EC are still limited. Emergence of EC organoids, a preclinical model, provides novel insights into understanding the pathological status of EC and developing effective and personalized treatment options.
Drug Screening
EC organoids can not only recapitulate important histological, genetic, and molecular characteristics of the original tumor, but also faithfully reflect the mutational profiling of intra-tumoral heterogeneity.22,23 Patient-derived EC organoids, when subcutaneously injected or orthotopically transplanted into immunocompromised mice, can simulate the physiological TME.23 These xenografted organoids replicate the histology and biomarkers of endometrial cancer, including dysregulated hormone receptor expression, malignant behaviors like metastasis and invasion, thus providing a potential platform for in vivo drug screening.
Wang et al performed an unbiased drug screening on driver-defined EC organoids in mice and indicated that MI-136, an inhibitor of the menin-MLL complex, could strongly inhibit the organoid growth possibly through regulation of the HIF pathway based on the screening of a small molecule library targeting epigenetic factors, which was further validated in the PDTOs.68 This study suggest that MI-136 may serve as a novel therapeutic target for EC. Another high-throughput drug screening based on mouse EC organoids was performed to explore the responses of drugs targeting different pathways, such as endocrine and hormone pathways, and protein tyrosine kinase pathway, validating the heterogeneity in drug responses among EC organoids with different genetic mutations.69 By culturing EC organoids from 15 samples, Girda et al observed that the STAT3 transcription factor inhibitor BBI608 significantly suppressed the organoid growth, highlighting the importance of stemness programing in organoid growth; Inhibitory effects exerted by different growth factor receptor tyrosine kinase inhibitors in several organoids suggested variability in growth factor signaling pathways.70 Strikingly, fulvestrant suppressed the growth of four organoids, indicating the importance of estrogen receptor signaling in certain PDTOs. Additionally, 16 PDX organoids were established from multiple EC subtypes. The results showed that PDX organoids with high or medium FGFR2 expression were markedly sensitive to the FGFR inhibitor BGJ398, and presented significant inhibition on the tumor growth and survival extension after treatment with FGFR inhibitors.71 These studies all demonstrate the great prospects of patient-derived EC organoids in drug sensitivity testing and personalized treatment.
Mechanisms Underlying Tumorigenesis
In a previous study, MI-136 was verified to be effective whether in tumor organoids or orthotopic models, and the HIF pathway was the most significantly downregulated pathway via MI-136 treatment. Importantly, MI-136 significantly inhibited the growth of patient-derived EC organoids. Thus, MI-136 contributes to EC treatment by modulating the HIF pathway.68 SNORD14E overexpressed in EC tissues was associated with poor prognosis of patients. In vitro experiments, SNORD14E could facilitate proliferation, migration and invasion while suppressing EC cell apoptosis. Notably, antisense oligonucleotides targeting SNORD14E could inhibit the proliferation in patient-derived high-risk EC organoids. Mechanistically, SNORD14E promoted EC progression by increasing the expression of FOXM1 malignant subtypes (FOXM1b and FOXM1c) and inducing the nuclear accumulation of β-catenin.72 Collectively, PDTOs provide a new research platform for EC development, progression, invasion, and recurrence.
For high-grade EC, there are lack of preclinical models for studying cancer biology, such as cancer-immune cell interactions. By establishing an endometrial biobank comprising patient-derived organoids, Chung et al found that high-grade EC organoids pre-treated with IFNg or Tazemetostat exhibited a significant immune cell co-localization and immune cell-mediated apoptosis during the co-culture period, highlighting the utility of the co-culture model in studying cancer-immune cell interactions.73 Additionally, based on the co-culture model of PDTOs with macrophages, the effects of estrogen-related receptor α (ERRα) on the migration ability of different subtypes of macrophages were validated in EC.74 In combination with high-throughput assays, such as proteomics, lipidomics and coculture systems, ERRα functions were identified as a bridge between EC metabolism and immunity, paving the way for developing novel therapies targeting cancer metabolism and immunotherapy.
Limitations and Prospects
PDTOs have shown great promise in drug screening and tumor biological research, particularly in gynecological cancers. Nevertheless, there still faces several challenges. The primary challenge for PDTOs lies in accurately recapitulating the complexity of TME. Currently, PDTOs cannot fully replicate the supporting structures required by tumors, such as blood vessels, immune cells, stroma, and nerves, thereby limiting their ability to accurately capture the multifactorial nature of tumor progression and treatment response, especially in the context of immunotherapy. Although the cocultured organoid systems have been developed to mimic the TME complexity through introduction of immune cells and specific interleukins, they are still at the early stage of optimization. Second, PDTOs cannot faithfully reflect the process of pharmacokinetics and pharmacodynamics like drug metabolism, absorption, distribution and excretion, which are indispensable for the assessment of in vivo drug efficacy and safety. Third, the tumor tissues used for PDTO culture are limited, especially for the rare cancers such as vulvar and vaginal cancers, limiting the development of comprehensive organoid systems for these melanomas. In addition, the establishment of PDTOs is costly, mainly depending on raw materials such as multiple cytokines and Matrigel.75
At present, most PDTO models use Matrigel or collagen as supporting materials to establish 3D structures, but the unknown extracellular components in these extracellular matrices may alter the original tumor heterogeneity, thus affecting the stability and repeatability of experimental results. With the continuous development of PDTO models and gene editing technologies, the application of gene manipulation at the PDTO level holds broad prospects in the study of gynecological cancers. The study made by Godbole et al revealed extracellular vesicles as a potential delivery platform for gene editing therapies in epithelial OC, offering a new delivery strategy for gene manipulation at the PDTO level.76 Based on the cell-free DNA methylation, a liquid biopsy assay was developed to assess the risk of HGSOC, providing insights into identification of the potential molecular markers for gene manipulation at the PDTO level.77 Notably, gene manipulation at the PDTO level still faces huge technical challenges. For example, the success rate, stability, and repeatability of PDTOs need to be further studied and improved; the safety and effectiveness of gene manipulation need to be further verified. Therefore, except for introduction of automated operations, intelligent monitoring and precise control, it is urgent to develop more advanced algorithms and technical means to allow for high-throughput and highly homogeneous generation of PDTOs.78
With continuous improvement of the organoid technology, PDTOs will more accurately capture the TME, creating an unprecedented platform for studying the molecular mechanisms driving tumorigenesis, progression, and metastasis in gynecological cancers. Importantly, novel therapeutic targets that can guide the targeted therapy, especially for gynecological cancers can be discovered through the organoid-based identification of key signaling pathways, favorable for modification of treatment options to improve the patient clinical outcomes and minimize adverse reactions.
Conclusion
PDTOs established from gynecological cancers can highly retain histological, genetic, and molecular characteristics of the original tumor while preserving the tumor heterogeneity, thereby promoting the development and utilization of precise medicine. Under the circumstance of understanding the mechanism underlying tumorigenesis and drug screening of female reproductive system malignancies, PDTOs facilitate the optimization of treatment regiments, making great contributions to the prognostic improvement. Despite presence of significant advantages in biomedical research, the organoid technology still requires for optimizing the culture protocols, such as integration of stromal components into tumor parenchymal cells. With continuous improvement of the organoid technology, PDTOs will more accurately capture the TME, creating a powerful platform for tumor biological research, treatment response, and personalized treatment.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
Yu Hu, Wei Chen, and Xiaojing Zhang are co-first authors for this study. The authors report no conflicts of interest in this work.
References
1. Campo L, Zhang C, Breuer EK. EMT-inducing molecular factors in gynecological cancers. Biomed Res Int. 2015;2015:420891. doi:10.1155/2015/420891
2. Bray F, Laversanne M, Sung H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024;74(3):229–263. doi:10.3322/caac.21834
3. Perelli F, Mattei A, Scambia G, et al. Editorial: methods in gynecological oncology. Front Oncol. 2023;13:1167088. doi:10.3389/fonc.2023.1167088
4. Virarkar M, Bhosale PR. Editorial: women in gynecological oncology 2021. Front Oncol. 2023;13:1220467. doi:10.3389/fonc.2023.1220467
5. Richardson DL, Eskander RN, O’Malley DM. Advances in ovarian cancer care and unmet treatment needs for patients with platinum resistance. JAMA Oncol. 2023;9(6):851–859. doi:10.1001/jamaoncol.2023.0197
6. Baldewpersad Tewarie N, van Driel WJ, van Ham MAPC, et al. An overview of clinical quality registries (CQRs) on gynecological oncology worldwide. Eur J Surg Oncol. 2022;48(10):2094–2103. doi:10.1016/j.ejso.2022.06.020
7. Sznurkowski JJ, Rys J, Kowalik A, et al. The polish society of gynecological oncology guidelines for the diagnosis and treatment of endometrial carcinoma (2023). J Clin Med. 2023;12(4):1480. doi:10.3390/jcm12041480
8. Magré L, Verstegen MMA, Buschow S, et al. Emerging organoid-immune co-culture models for cancer research: from oncoimmunology to personalized immunotherapies. J Immunother Cancer. 2023;11:e006290. doi:10.1136/jitc-2022-006290
9. Saito R, Kobayashi T, Kashima S, et al. Faithful preclinical mouse models for better translation to bedside in the field of immuno-oncology. Int J Clin Oncol. 2020;25:831–841. doi:10.1007/s10147-019-01520-z
10. Ben-David U, Ha G, Tseng YY, et al. Patient-derived xenografts undergo mouse-specific tumor evolution. Nat Genet. 2017;49(11):1567–1575. doi:10.1038/ng.3967
11. Izzo R, Bevivino G, De Simone V, et al. Knockdown of Smad7 with a specific antisense oligonucleotide attenuates colitis and colitis-driven colonic fibrosis in mice. Inflamm Bowel Dis. 2018;24(6):1213–1224. doi:10.1093/ibd/izy062
12. Polak R, Zhang ET, Kuo CJ. Cancer organoids 2.0: modelling the complexity of the tumour immune microenvironment. Nat Rev Cancer. 2024;24(8):523–539. doi:10.1038/s41568-024-00706-6
13. Chen P, Zhang X, Ding R, et al. Patient-derived organoids can guide personalized-therapies for patients with advanced breast cancer. Adv Sci. 2021;8(22):e2101176. doi:10.1002/advs.202101176
14. Jiang Y, Sun X, Song X, et al. Patient-derived bladder cancer organoid model to predict sensitivity and feasibility of tailored precision therapy. Curr Urol. 2023;17(4):221–228. doi:10.1097/CU9.0000000000000219
15. Tiriac H, Belleau P, Engle DD, et al. Organoid profiling identifies common responders to chemotherapy in pancreatic cancer. Cancer Discov. 2018;8(9):1112–1129. doi:10.1158/2159-8290.CD-18-0349
16. de Witte CJ, Espejo Valle-Inclan J, Hami N, et al. Patient-derived ovarian cancer organoids mimic clinical response and exhibit heterogeneous inter- and intrapatient drug responses. Cell Rep. 2020;31(11):107762. doi:10.1016/j.celrep.2020.107762
17. Maenhoudt N, Defraye C, Boretto M, et al. Developing organoids from ovarian cancer as experimental and preclinical models. Stem Cell Reports. 2020;14(4):717–729. doi:10.1016/j.stemcr.2020.03.004
18. Chen L-Y, Chou Y-T, Liew P-L, et al. In vitro drug testing using patient-derived ovarian cancer organoids. J Ovarian Res. 2024;17(1):194. doi:10.1186/s13048-024-01520-2
19. Driehuis E, Kretzschmar K, Clevers H. Establishment of patient-derived cancer organoids for drug-screening applications. Nat Protoc. 2020;15(10):3380–3409. doi:10.1038/s41596-020-0379-4
20. Sun C-P, Lan H-R, Fang X-L, et al. Organoid models for precision cancer immunotherapy. Front Immunol. 2022;13:770465. doi:10.3389/fimmu.2022.770465
21. Rinehart CA, Lyn-Cook BD, Kaufman DG. Gland formation from human endometrial epithelial cells in vitro. Vitro Cell Dev Biol. 1988;24(10):1037–1041. doi:10.1007/BF02620878
22. Maru Y, Tanaka N, Itami M, et al. Efficient use of patient-derived organoids as a preclinical model for gynecologic tumors. Gynecol Oncol. 2019;154(1):189–198. doi:10.1016/j.ygyno.2019.05.005
23. Boretto M, Maenhoudt N, Luo X, et al. Patient-derived organoids from endometrial disease capture clinical heterogeneity and are amenable to drug screening. Nat Cell Biol. 2019;21(8):1041–1051. doi:10.1038/s41556-019-0360-z
24. Kopper O, de Witte CJ, Lõhmussaar K, et al. An organoid platform for ovarian cancer captures intra- and interpatient heterogeneity. Nat Med. 2019;25(5):838–849. doi:10.1038/s41591-019-0422-6
25. Maru Y, Tanaka N, Ebisawa K, et al. Establishment and characterization of patient-derived organoids from a young patient with cervical clear cell carcinoma. Cancer Sci. 2019;110(9):2992–3005. doi:10.1111/cas.14119
26. Seol HS, Oh JH, Choi E, et al. Preclinical investigation of patient-derived cervical cancer organoids for precision medicine. J Gynecol Oncol. 2023;34(3):e35. doi:10.3802/jgo.2023.34.e35
27. Kusakabe M, Taguchi A, Tanikawa M, et al. Application of organoid culture from HPV18 -positive small cell carcinoma of the uterine cervix for precision medicine. Cancer Med. 2023;12(7):8476–8489. doi:10.1002/cam4.5588
28. Rauth S, Karmakar S, Batra SK, et al. Recent advances in organoid development and applications in disease modeling. Biochim Biophys Acta Rev Cancer. 2021;1875(2):188527. doi:10.1016/j.bbcan.2021.188527
29. Neal JT, Li X, Zhu J, et al. Organoid modeling of the tumor immune microenvironment. Cell. 2018;175(7):1972–1988.e16. doi:10.1016/j.cell.2018.11.021
30. Li X, Nadauld L, Ootani A, et al. Oncogenic transformation of diverse gastrointestinal tissues in primary organoid culture. Nat Med. 2014;20(7):769–777. doi:10.1038/nm.3585
31. Esser LK, Branchi V, Leonardelli S, et al. Cultivation of clear cell renal cell carcinoma patient-derived organoids in an air-liquid interface system as a tool for studying individualized therapy. Front Oncol. 2020;10:1775. doi:10.3389/fonc.2020.01775
32. Elbadawy M, Usui T, Yamawaki H, et al. Development of an experimental model for analyzing drug resistance in colorectal cancer. Cancers. 2018;10(6):164. doi:10.3390/cancers10060164
33. Dong M, Böpple K, Thiel J, et al. Perfusion air culture of precision-cut tumor slices: an ex vivo system to evaluate individual drug response under controlled culture conditions. Cells. 2023;12(5):807. doi:10.3390/cells12050807
34. Tian J, Yang J, Chen T, et al. Generation of human endometrial assembloids with a luminal epithelium using air–liquid interface culture methods. Adv Sci. 2023;10(30):e2301868. doi:10.1002/advs.202301868
35. Kumar AS, Venkatesalu S, Dilliyappan S, et al. Microfluidics as diagnostic tools. Clin Chim Acta. 2024;556:117841. doi:10.1016/j.cca.2024.117841
36. Kim SK, Kim YH, Park S, et al. Organoid engineering with microfluidics and biomaterials for liver, lung disease, and cancer modeling. Acta Biomater. 2021;132:37–51. doi:10.1016/j.actbio.2021.03.002
37. Xiao S, Coppeta JR, Rogers HB, et al. A microfluidic culture model of the human reproductive tract and 28-day menstrual cycle. Nat Commun. 2017;8(1):14584. doi:10.1038/ncomms14584
38. Qu J, Kalyani FS, Liu L, et al. Tumor organoids: synergistic applications, current challenges, and future prospects in cancer therapy. Cancer Commun. 2021;41:1331–1353. doi:10.1002/cac2.12224
39. Senkowski W, Gall-Mas L, Falco MM, et al. A platform for efficient establishment and drug-response profiling of high-grade serous ovarian cancer organoids. Dev Cell. 2023;58(12):1106–1121.e7. doi:10.1016/j.devcel.2023.04.012
40. Thorel L, Morice P-M, Paysant H, et al. Comparative analysis of response to treatments and molecular features of tumor-derived organoids versus cell lines and PDX derived from the same ovarian clear cell carcinoma. J Exp Clin Cancer Res. 2023;42(1):260. doi:10.1186/s13046-023-02809-8
41. Chen Y-A, Lu C-Y, Cheng W-F, et al. An experimental model for ovarian cancer: propagation of ovarian cancer initiating cells and generation of ovarian cancer organoids. BMC Cancer. 2022;22(1):967. doi:10.1186/s12885-022-10042-3
42. Phan N, Hong JJ, Tofig B, et al. A simple high-throughput approach identifies actionable drug sensitivities in patient-derived tumor organoids. Commun Biol. 2019;2(1):78. doi:10.1038/s42003-019-0305-x
43. Nanki Y, Chiyoda T, Hirasawa A, et al. Patient-derived ovarian cancer organoids capture the genomic profiles of primary tumours applicable for drug sensitivity and resistance testing. Sci Rep. 2020;10(1):12581. doi:10.1038/s41598-020-69488-9
44. Chen H, Gotimer K, De Souza C, et al. Short-term organoid culture for drug sensitivity testing of high-grade serous carcinoma. Gynecol Oncol. 2020;157(3):783–792. doi:10.1016/j.ygyno.2020.03.026
45. Hill SJ, Decker B, Roberts EA, et al. Prediction of DNA repair inhibitor response in short-term patient-derived ovarian cancer organoids. Cancer Discov. 2018;8(11):1404–1421. doi:10.1158/2159-8290.CD-18-0474
46. Dijkstra KK, Cattaneo CM, Weeber F, et al. Generation of tumor-reactive T Cells by co-culture of peripheral blood lymphocytes and tumor organoids. Cell. 2018;174(6):1586–1598.e12. doi:10.1016/j.cell.2018.07.009
47. Sui Q, Zhang X, Chen C, et al. Inflammation promotes resistance to immune checkpoint inhibitors in high microsatellite instability colorectal cancer. Nat Commun. 2022;13(1):7316. doi:10.1038/s41467-022-35096-6
48. Bogani G, Moore KN, Ray-Coquard I, et al. Incorporating immune checkpoint inhibitors in epithelial ovarian cancer. Gynecol Oncol. 2025;193:30–40. doi:10.1016/j.ygyno.2024.12.011
49. Sun H, Wang H, Wang X, et al. Aurora-A/SOX8/FOXK1 signaling axis promotes chemoresistance via suppression of cell senescence and induction of glucose metabolism in ovarian cancer organoids and cells. Theranostics. 2020;10(15):6928–6945. doi:10.7150/thno.43811
50. Wang Z, Chen W, Zuo L, et al. The Fibrillin-1/VEGFR2/STAT2 signaling axis promotes chemoresistance via modulating glycolysis and angiogenesis in ovarian cancer organoids and cells. Cancer Commun. 2022;42(3):245–265. doi:10.1002/cac2.12274
51. Hoarau-Véchot J, Blot-Dupin M, Pauly L, et al. Akt-activated endothelium increases cancer cell proliferation and resistance to treatment in ovarian cancer cell organoids. Int J Mol Sci. 2022;23(22):14173. doi:10.3390/ijms232214173
52. Wang H, Zhang Y, Miao H, et al. CircRAD23B promotes proliferation and carboplatin resistance in ovarian cancer cell lines and organoids. Cancer Cell Int. 2024;24(1):42. doi:10.1186/s12935-024-03228-1
53. Cai X, Li Y, Zheng J, et al. Modeling of senescence-related chemoresistance in ovarian cancer using data analysis and patient-derived organoids. Front Oncol. 2024;13:1291559. doi:10.3389/fonc.2023.1291559
54. Dai Y, Xu J, Gong X, et al. Human fallopian tube-derived organoids with TP53 and RAD51D mutations recapitulate an early stage high-grade serous ovarian cancer phenotype in vitro. Int J Mol Sci. 2024;25:886. doi:10.3390/ijms25020886
55. Abu-Rustum NR, Yashar CM, Arend R, et al. NCCN guidelines® insights: cervical cancer, version 1.2024. J Natl Compr Canc Netw. 2023;21(12):1224–1233. doi:10.6004/jnccn.2023.0062
56. Lõhmussaar K, Oka R, Espejo Valle-Inclan J, et al. Patient-derived organoids model cervical tissue dynamics and viral oncogenesis in cervical cancer. Cell Stem Cell. 2021;28(8):1380–1396.e6. doi:10.1016/j.stem.2021.03.012
57. Di Fonte R, Strippoli S, Garofoli M, et al. Cervical cancer benefits from trabectedin combination with the β-blocker propranolol: in vitro and ex vivo evaluations in patient-derived organoids. Front Cell Dev Biol. 2023;11:1178316. doi:10.3389/fcell.2023.1178316
58. Zhang J, Rashmi R, Inkman M, et al. Integrating imaging and RNA-seq improves outcome prediction in cervical cancer. J Clin Invest. 2021;131(5):e139232. doi:10.1172/JCI139232
59. Huang H, Pan Y, Huang J, et al. Patient-derived organoids as personalized avatars and a potential immunotherapy model in cervical cancer. iScience. 2023;26(11):108198. doi:10.1016/j.isci.2023.108198
60. Cattaneo CM, Dijkstra KK, Fanchi LF, et al. Tumor organoid–T-cell coculture systems. Nat Protoc. 2020;15(1):15–39. doi:10.1038/s41596-019-0232-9
61. Dong J, Holthaus D, Peters C, et al. γδ T cell-mediated cytotoxicity against patient-derived healthy and cancer cervical organoids. Front Immunol. 2023;14:1281646. doi:10.3389/fimmu.2023.1281646
62. Bhatla N, Singhal S. Primary HPV screening for cervical cancer. Best Pract Res Clin Obstet Gynaecol. 2020;65:98–108. doi:10.1016/j.bpobgyn.2020.02.008
63. Hu Z, Zhu D, Wang W, et al. Genome-wide profiling of HPV integration in cervical cancer identifies clustered genomic hot spots and a potential microhomology-mediated integration mechanism. Nat Genet. 2015;47(2):158–163. doi:10.1038/ng.3178
64. Toyohara Y, Taguchi A, Ishii Y, et al. Identification of target cells of human papillomavirus 18 using squamocolumnar junction organoids. Cancer Sci. 2024;115(1):125–138. doi:10.1111/cas.15988
65. Hu B, Wang R, Wu D, et al. A promising new model: establishment of patient-derived organoid models covering HPV-related cervical pre-cancerous lesions and their cancers. Adv Sci. 2024;11:e2302340.
66. Koster S, Gurumurthy RK, Kumar N, et al. Modelling chlamydia and HPV co-infection in patient-derived ectocervix organoids reveals distinct cellular reprogramming. Nat Commun. 2022;13(1):1030. doi:10.1038/s41467-022-28569-1
67. Makker V, MacKay H, Ray-Coquard I, et al. Endometrial cancer. Nat Rev Dis Primers. 2021;7(1):88. doi:10.1038/s41572-021-00324-8
68. Chen J, Zhao L, Peng H, et al. An organoid-based drug screening identified a menin-MLL inhibitor for endometrial cancer through regulating the HIF pathway. Cancer Gene Ther. 2021;28(1–2):112–125. doi:10.1038/s41417-020-0190-y
69. Chen J, Dai S, Zhao L, et al. A new type of endometrial cancer models in mice revealing the functional roles of genetic drivers and exploring their susceptibilities. Adv Sci. 2023;10:e2300383.
70. Girda E, Huang EC, Leiserowitz GS, et al. The use of endometrial cancer patient-derived organoid culture for drug sensitivity testing is feasible. Int J Gynecol Cancer. 2017;27:1701–1707. doi:10.1097/IGC.0000000000001061
71. Sengal AT, Bonazzi V, Smith D, et al. Endometrial cancer PDX-derived organoids (PDXOs) and PDXs with FGFR2c isoform expression are sensitive to FGFR inhibition. Npj Precision Oncol. 2023;7(1):127. doi:10.1038/s41698-023-00478-6
72. Chen X, Liu X, Li Q-H, et al. A patient-derived organoid-based study identified an ASO targeting SNORD14E for endometrial cancer through reducing aberrant FOXM1 expression and β-catenin nuclear accumulation. J Exp Clin Cancer Res. 2023;42(1):230. doi:10.1186/s13046-023-02801-2
73. Chung C, Nizam A, Goldberg GL, et al. Organoid models for high-grade endometrial cancer to dissect tumor immune microenvironments. J Immunol. 2022;208(Supplement_1):
74. Ma J, Mao X, Ren Y, et al. Antagonism of estrogen-related receptor-α inhibits mitochondrial oxidative phosphorylation and reduces M2 macrophage infiltration in endometrial cancer. J Immunother Cancer. 2025;13(9):e012521. doi:10.1136/jitc-2025-012521
75. Marx V. Closing in on cancer heterogeneity with organoids. Nat Methods. 2024;21(4):551–554. doi:10.1038/s41592-024-02231-8
76. Godbole N, Quinn A, Carrion F, Pelosi E, Salomon C. Extracellular vesicles as a potential delivery platform for CRISPR-Cas based therapy in epithelial ovarian cancer. Semin Cancer Biol. 2023;96:64–81. doi:10.1016/j.semcancer.2023.10.002
77. Buckley DN, Lewinger JP, Gooden G, et al. OvaPrint—a cell-free DNA methylation liquid biopsy for the risk assessment of high-grade serous ovarian cancer. Clin Cancer Res. 2023;29(24):5196–5206. doi:10.1158/1078-0432.CCR-23-1197
78. Shi H, Kowalczewski A, Vu D, et al. Organoid intelligence: integration of organoid technology and artificial intelligence in the new era of in vitro models. Med Novel Technol Devices. 2024;21:100276. doi:10.1016/j.medntd.2023.100276
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.