Back to Journals » Drug Design, Development and Therapy » Volume 19
Decoding the Crosstalk Between Heat Shock Proteins and Phytochemicals in Thyroid Nodule Regression: Mechanisms and Therapeutic Implications
Received 16 July 2025
Accepted for publication 22 November 2025
Published 13 December 2025 Volume 2025:19 Pages 11093—11105
DOI https://doi.org/10.2147/DDDT.S554080
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Ruina Zhang, Juanqin Lv
Department of Endocrinology, The Third Affiliated Hospital of Gansu University of Chinese Medicine, Baiyin, Gansu, People’s Republic of China
Correspondence: Juanqin Lv, Department of Endocrinology, The Third Affiliated Hospital of Gansu University of Chinese Medicine, Baiyin, Gansu, People’s Republic of China, Email [email protected]
Abstract: Thyroid nodules represent one of the most prevalent endocrine disorders, affecting up to 60% of adults worldwide, with diverse biological behaviors ranging from benign hyperplastic nodules to malignant thyroid carcinoma. Heat shock proteins (HSPs), especially HSP70 and HSP90, act as key molecular chaperones involved in cellular proliferation, stress tolerance, and survival, thereby contributing to both benign nodule persistence and malignant transformation. In parallel, phytochemicals such as curcumin, epigallocatechin gallate (EGCG), resveratrol, and quercetin have emerged as bioactive modulators capable of influencing HSP expression and activity. This review decodes the molecular crosstalk between HSPs and phytochemicals, elucidating how these natural compounds regulate signaling cascades including NF-κB, MAPK, and PI3K/Akt to mediate apoptosis, autophagy, and oxidative stress balance. Particular emphasis is placed on differentiating their therapeutic implications for benign nodule regression versus thyroid cancer inhibition. Preclinical evidence demonstrates that phytochemicals can downregulate HSPs, enhance apoptotic signaling, and sensitize tumor cells to therapy; however, clinical translation remains limited due to low bioavailability, off-target effects, and dosage optimization challenges. Addressing these pharmacokinetic and delivery barriers—through nanotechnology and precision-medicine-based approaches—may unlock new integrative strategies for thyroid disease management. Collectively, this review provides a mechanistic and translational framework highlighting HSP–phytochemical interactions as potential molecular targets for improving therapeutic outcomes in both benign and malignant thyroid pathologies.
Keywords: HSPs, phytochemicals, thyroid nodules, apoptosis and autophagy, signaling pathway modulation
Introduction
Thyroid nodules are among the most common endocrine disorders, with studies estimating their prevalence at approximately 50–60% in the adult population, although only a small percentage (5–15%) are malignant.1 Recent large-scale epidemiological studies have confirmed that the global prevalence of thyroid nodules continues to rise, largely due to increased use of high-resolution ultrasonography, while also emphasizing demographic and geographic variability in risk.2–4 The diagnosis and clinical management of thyroid nodules present significant challenges, given the heterogeneity in nodule characteristics and their potential for malignancy. Despite advancements in diagnostic imaging and fine-needle aspiration biopsy (FNAB), distinguishing benign from malignant nodules remains difficult in many cases, necessitating more effective therapeutic strategies.5 Recent diagnostic studies highlight that while FNAB remains the gold standard, limitations persist, especially for indeterminate lesions, where novel imaging modalities and molecular classifiers are increasingly investigated.6–8Current treatments for thyroid nodules, ranging from active surveillance to surgical interventions, fail to address the underlying molecular mechanisms driving nodule growth and regression, highlighting the need for more targeted, molecularly driven therapies.9 Clinically, this distinction is crucial: benign hyperplastic nodules are typically managed with the aim of size reduction or symptom relief, whereas malignant tumors require oncologic control and prevention of progression. HSPs are a family of highly conserved molecular chaperones that assist in the proper folding of nascent polypeptides and the refolding or degradation of denatured proteins under stress conditions. They are classified into several families, including HSP70, HSP90, and HSP27, with each playing crucial roles in cellular homeostasis and stress responses.10 HSPs are implicated in various pathological conditions, including cancer, due to their ability to regulate critical cellular processes such as protein stability, apoptosis, and cellular survival. Specifically, HSPs are known to contribute to the development and progression of malignancies, including thyroid cancer, by facilitating tumor cell survival under oxidative stress and other unfavorable conditions.11 In thyroid nodules, the overexpression of certain HSPs, such as HSP70 and HSP90, has been associated with increased cell proliferation, resistance to apoptosis, and overall tumorigenic potential.12 These findings indicate that while HSPs are central to malignant transformation, they may also contribute to the persistence of benign nodular disease, though the therapeutic endpoints differ. Given their central role in cellular stress responses, HSPs are considered potential therapeutic targets for cancer treatment.
Phytochemicals, naturally occurring bioactive compounds found in plants, have garnered significant attention for their potential therapeutic effects in various diseases, including cancer. Many phytochemicals, such as curcumin, EGCG, resveratrol, and quercetin, have demonstrated anti-inflammatory, antioxidant, and anti-cancer properties, making them attractive candidates for adjunctive therapies in cancer management.13–15 Recent studies have highlighted the potential of these compounds to modulate the expression and activity of HSPs, thereby influencing cancer cell survival, apoptosis, and tumor progression. For example, curcumin has been shown to downregulate HSP90 in several cancer models, inducing cell cycle arrest and promoting apoptosis.16 Similarly, EGCG has demonstrated the ability to interact with HSP70, inhibiting its chaperone function and sensitizing cancer cells to apoptosis.17 Thus, phytochemicals should be evaluated not only as modulators of HSP activity in malignant thyroid tumors but also in the context of benign nodule regression.
This review aims to explore the crosstalk between heat shock proteins and phytochemicals in the context of thyroid nodule regression. We critically examine the implications for benign versus malignant nodules, synthesize mechanistic and translational evidence, and highlight key challenges such as bioavailability and dosing. Importantly, we ask: can phytochemicals act as clinically relevant modulators of HSPs, and what therapeutic strategies—including novel delivery systems and biomarker-driven patient selection—are required for successful translation.
Molecular Insights Into Thyroid Nodules and HSPs
Mechanisms of Thyroid Nodule Formation and Progression
Thyroid nodules are clinically characterized by the presence of discrete lesions within the thyroid gland. These nodules may be benign, hyperplastic, or malignant, with benign nodules making up the majority of cases.18 The molecular mechanisms underlying thyroid nodule formation and progression are complex, involving various factors such as genetic mutations, environmental influences, and dysregulated signaling pathways. In benign or hyperplastic nodules, non-neoplastic processes such as iodine imbalance, oxidative stress, and TSH overstimulation contribute to focal cellular hyperplasia and colloid accumulation, leading to nodule formation without oncogenic transformation. These benign mechanisms differ fundamentally from malignant transformation.19 Commonly observed mutations in thyroid cancer include those in the BRAF, RAS, and RET proto-oncogenes, which drive abnormal cellular proliferation and growth.20 Additionally, alterations in the phosphatidylinositol 3-kinase (PI3K)/Akt and mitogen-activated protein kinase (MAPK) pathways play significant roles in thyroid carcinogenesis.21
The progression from benign to malignant lesions is rare but involves additional genetic and epigenetic alterations that enhance proliferative signaling, genomic instability, and angiogenesis. The progression from benign to malignant thyroid nodules is associated with a series of biological events, including increased cellular proliferation, resistance to apoptosis, and enhanced angiogenesis.22 The tumor microenvironment in thyroid nodules is often hypoxic, contributing to an increased oxidative stress, which further supports tumor cell survival through the upregulation of various protective proteins, including HSPs.23 However, while such stress responses in benign nodules may serve protective or adaptive functions, in malignant thyroid tumors they facilitate immune evasion, therapeutic resistance, and sustained proliferation. Thus, differentiating the biological role of HSPs in benign versus malignant contexts is crucial for understanding disease progression and therapeutic targeting.
Role of Heat Shock Proteins in Thyroid Nodules
Heat shock proteins, particularly HSP70 and HSP90, have been implicated in thyroid nodule biology. These chaperones function to maintain protein stability and facilitate the proper folding of newly synthesized proteins, preventing aggregation under stressful conditions such as hypoxia, oxidative stress, and DNA damage.23 In benign thyroid nodules, mild cellular stress and oxidative imbalance can induce moderate upregulation of HSPs, which acts as a protective response to maintain thyroid follicular cell integrity and prevent apoptosis.24 In thyroid cancer cells, HSPs not only protect against stress-induced cell death but also regulate several processes involved in tumor progression, including cell cycle progression, apoptosis, and resistance to chemotherapeutic agents.25
HSP70, for example, is known to interact with several client proteins involved in cell survival and apoptosis, such as the tumor suppressor p53. In thyroid cancer, HSP70 has been found to inhibit p53-mediated apoptosis, thus promoting cell survival.26 Similarly, HSP90 plays a critical role in stabilizing and activating various oncoproteins, including steroid hormone receptors, protein kinases, and transcription factors. In thyroid carcinoma, the overexpression of HSP90 contributes to tumorigenesis by stabilizing key oncogenic proteins like RET/PTC and BRAF, thereby enhancing their signaling capabilities and promoting cell proliferation and survival.27 Moreover, the expression levels of HSPs in thyroid nodules correlate with the degree of malignancy. Studies have shown that HSP90 is significantly overexpressed in malignant thyroid tumors compared to benign nodules, suggesting its potential role as a prognostic marker.17 The overexpression of HSPs in malignant thyroid nodules is thought to be a protective mechanism that helps tumor cells survive under adverse conditions such as nutrient deprivation, hypoxia, and chemotherapy. By contrast, in benign or hyperplastic nodules, elevated HSP expression generally reflects adaptive cytoprotection rather than oncogenic transformation, highlighting the context-dependent nature of HSP function in thyroid pathology.28
HSPs as Therapeutic Targets in Thyroid Nodules
Given the critical roles of HSPs in thyroid nodule progression, they represent promising therapeutic targets. In benign thyroid nodules, mild inhibition or modulation of HSP activity may promote nodule regression by restoring apoptotic balance and reducing aberrant proliferative signaling, while in malignant thyroid lesions, more potent inhibition of HSP70 and HSP90 is required to suppress oncogenic signaling pathways.29 Inhibitors of HSPs, particularly HSP90 and HSP70, are being explored as potential anti-cancer agents. Small-molecule HSP90 inhibitors, such as 17-AAG (17-Allylamino-17-demethoxygeldanamycin), have demonstrated efficacy in preclinical thyroid cancer models by promoting the degradation of key oncogenic client proteins, leading to reduced tumor growth and enhanced sensitivity to chemotherapy.30 Similarly, HSP70 inhibitors have been shown to sensitize thyroid cancer cells to apoptotic stimuli, making them a potential adjuvant therapy in combination with other treatment modalities.12
However, the clinical application of HSP inhibitors faces several challenges, including the need to develop selective inhibitors that target specific HSP isoforms without affecting normal cellular functions. Moreover, the high expression of HSPs in normal tissues poses a risk of toxicity, underscoring the importance of targeting HSPs specifically in the tumor microenvironment.31 Additionally, achieving therapeutic selectivity remains a key barrier, as complete HSP inhibition can disrupt normal proteostasis. Therefore, future strategies may focus on partial modulation or tissue-specific inhibition to preserve physiological stress responses while limiting tumor-promoting activity.
Emerging evidence suggests that combining conventional HSP inhibitors with natural phytochemicals could provide a safer and more physiologically compatible means of modulating HSP activity. Phytochemicals such as curcumin or EGCG have been shown to downregulate HSPs in tumor cells at sub-toxic concentrations, suggesting a synergistic potential with pharmacologic inhibitors. Therefore, strategies that combine HSP inhibitors with phytochemicals, which may modulate HSP expression and activity in a more selective manner, are gaining attention as potentially effective therapeutic approaches.
Interplay Between HSPs and Phytochemicals
Key Phytochemicals Targeting Heat Shock Proteins
Phytochemicals, derived from various plant sources, have attracted considerable attention due to their potential therapeutic effects, particularly in cancer treatment. Many phytochemicals have been shown to interact with HSPs, either by modulating their expression or directly inhibiting their activity. Among the most studied phytochemicals are curcumin, EGCG, quercetin, and resveratrol. These compounds have demonstrated the ability to target HSPs in various cancers, including thyroid cancer. In addition, preclinical studies suggest that mild modulation of HSPs by certain phytochemicals can also contribute to protective antioxidant responses in benign thyroid hyperplasia, indicating context-dependent biological effects.
Curcumin, the active compound in turmeric, has been widely studied for its ability to modulate multiple cellular pathways, including the inhibition of HSP90. In thyroid cancer cells, curcumin has been shown to reduce HSP90 expression, leading to the degradation of oncogenic client proteins such as BRAF and RET, thereby inhibiting tumor growth and promoting apoptosis.13 At lower concentrations or in non-malignant thyroid tissues, curcumin may also act indirectly through Nrf2 activation and oxidative stress reduction, helping restore cellular redox balance rather than inducing apoptosis. Similarly, EGCG, a major polyphenol in green tea, interacts with HSP70 and has been found to inhibit its chaperone activity, thereby sensitizing thyroid cancer cells to oxidative stress and apoptotic stimuli.17 EGCG also displays a dual role: while suppressing tumor-associated HSPs, it enhances stress tolerance in normal thyroid follicular cells through AMPK-dependent signaling, which may explain its differential cytoprotective versus cytotoxic outcomes. Quercetin, a flavonoid with known antioxidant properties, has also been shown to downregulate HSP90 in cancer cells, leading to increased sensitivity to chemotherapy and reduced cell proliferation.32 Notably, quercetin may synergize with other phytochemicals such as resveratrol to stabilize HSP70 under physiological stress while downregulating it in tumor contexts, highlighting a fine-tuned regulatory balance. Resveratrol, a polyphenolic compound found in grapes, also exerts its effects by modulating HSP expression, particularly HSP70, thereby enhancing the apoptotic effects in cancer cells.33 Beyond its anti-tumor role, resveratrol has been shown to promote thyroid cell autophagy and improve mitochondrial function in oxidative-stress models, supporting its relevance to benign nodular regression. The ability of these phytochemicals to modulate HSPs highlights their potential as complementary therapeutic agents in the treatment of thyroid nodules, particularly in overcoming the protective roles of HSPs in tumor cells. Collectively, these compounds appear to exert concentration- and context-dependent regulation of HSPs—cytoprotective in benign lesions and pro-apoptotic in malignant ones—providing a mechanistic basis for dual therapeutic potential. These key interactions are summarized in Table 1.
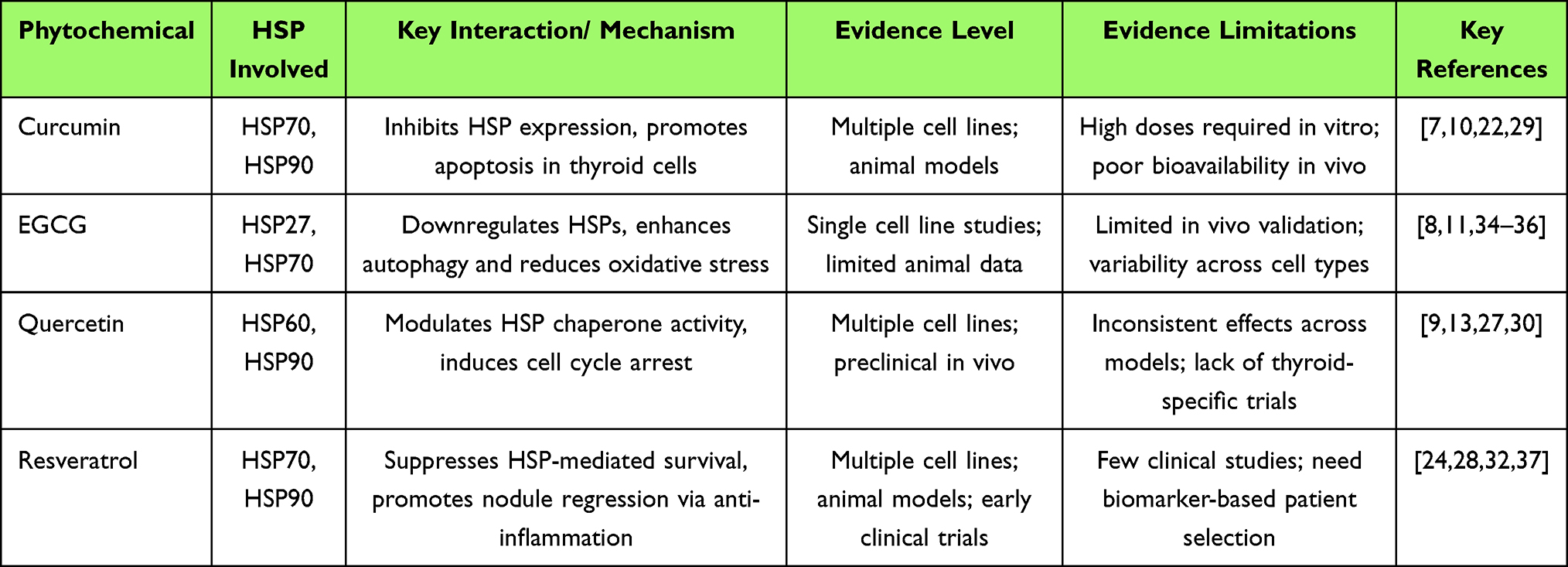 |
Table 1 Key Interactions Between Phytochemicals and Heat Shock Proteins in Thyroid Nodule Regression: Mechanisms and Evidence Levels |
Regulatory Mechanisms of Phytochemicals on HSP Expression
Phytochemicals exert their effects on HSPs through multiple regulatory mechanisms, both at the transcriptional and post-translational levels. Many of these compounds act by influencing the expression of heat shock factors (HSFs), the primary transcription factors that regulate HSP gene expression. For instance, curcumin has been shown to activate HSF1, a key regulator of HSP expression, thereby promoting the upregulation of HSP70 under stress conditions, which contributes to the protection of cells from damage.38 However, curcumin also exhibits the ability to inhibit HSP90 expression, indicating its dual role in regulating HSPs depending on cellular context.39 This bidirectional modulation reflects a concentration-dependent balance between cytoprotection in normal cells and anti-proliferative activity in tumor cells.40
EGCG, on the other hand, influences HSP expression by modulating the NF-κB pathway, which is involved in inflammation and cellular stress responses. By inhibiting NF-κB activation, EGCG reduces the expression of stress-induced HSPs, thus sensitizing cancer cells to apoptosis.17 At physiological levels, EGCG can also indirectly stabilize HSP70 in benign thyroid cells by reducing oxidative stress, thereby maintaining protein homeostasis and preventing DNA damage. Quercetin has been reported to regulate HSP expression through the PI3K/Akt signaling pathway, which is crucial for cell survival. In thyroid cancer models, quercetin downregulates HSP90 expression, contributing to decreased cell proliferation and enhanced cell death.34 Quercetin-mediated inhibition of PI3K/Akt can also intersect with NF-κB suppression, forming a synergistic network that amplifies the suppression of pro-survival HSPs in malignant cells. Additionally, resveratrol modulates HSP expression through the Sirt1/AMPK pathway, a key regulator of cellular metabolism and stress responses, leading to the reduction of HSP70 and promotion of apoptotic cell death in thyroid cancer cells.41 Activation of Sirt1/AMPK also promotes autophagic clearance of misfolded proteins, linking HSP modulation to improved proteostasis in both benign and malignant thyroid contexts. Collectively, these phytochemicals act on convergent signaling hubs—NF-κB, MAPK, PI3K/Akt, and Sirt1/AMPK—that intersect at the transcriptional regulation of HSF1 and downstream HSP70/HSP90 expression. The integrated effect is context-dependent: maintaining cellular homeostasis under moderate stress in benign nodules, and triggering apoptosis or autophagy under oncogenic stress in malignant lesions. These phytochemicals’ ability to influence multiple cellular signaling pathways underlines their therapeutic potential in regulating HSPs, which are critical for the progression and regression of thyroid nodules.42 As illustrated in Figure 1, phytochemicals such as curcumin and EGCG modulate HSP70 and HSP90 via NF-κB and MAPK pathways, inhibiting apoptosis, regulating cell cycle, inducing autophagy, and reducing oxidative stress in thyroid nodule cells.
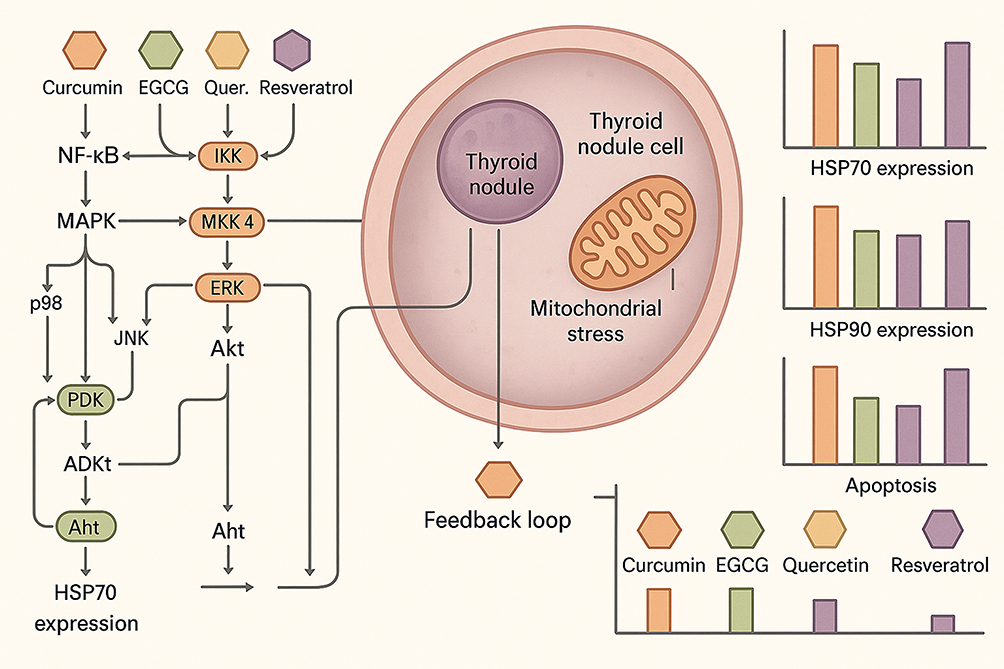 |
Figure 1 Crosstalk Mechanisms Between Heat Shock Proteins and Phytochemicals in Thyroid Nodule Regression. Schematic overview of the crosstalk between heat shock proteins (HSP70, HSP90) and phytochemicals (curcumin, EGCG, quercetin, resveratrol) in thyroid nodule regression, illustrating modulation of NF-κB, MAPK, and PI3K/Akt pathways leading to apoptosis induction, autophagy enhancement, and oxidative stress reduction. |
Effects on Cellular Pathways Relevant to Thyroid Nodules
The modulation of HSPs by phytochemicals has significant downstream effects on cellular pathways involved in thyroid nodule progression and regression. One of the most critical pathways influenced by HSPs is apoptosis, a process that is often dysregulated in cancer cells, allowing for uncontrolled cell survival. Phytochemicals such as curcumin and EGCG have been shown to enhance apoptosis in thyroid cancer cells by downregulating HSPs and upregulating pro-apoptotic proteins like Bax and caspases.43 In benign thyroid nodules, a milder activation of apoptotic signaling may contribute to nodule shrinkage by selectively eliminating hyperplastic cells without triggering extensive tissue damage, suggesting dose- and context-specific regulation.
Moreover, HSPs play a crucial role in the regulation of autophagy, a cellular process involved in the degradation of damaged cellular components. Dysregulated autophagy can contribute to cancer cell survival by preventing the accumulation of damaged proteins. Phytochemicals like quercetin and resveratrol have been found to induce autophagy in thyroid cancer cells by modulating HSP expression, thus promoting the clearance of damaged proteins and enhancing cell death.37 Importantly, recent evidence indicates that apoptosis and autophagy are not isolated outcomes but interconnected responses coordinated through HSP70/HSP90 modulation; their balance determines whether cells undergo adaptive recovery (in benign nodules) or programmed death (in malignancies). In addition to apoptosis and autophagy, HSPs also influence the oxidative stress response, which is critical for tumor progression. Phytochemicals such as resveratrol and curcumin reduce oxidative stress in thyroid cancer cells by modulating HSPs and other cellular antioxidants, thereby enhancing the therapeutic efficacy of existing cancer treatments.44 This antioxidant regulation may also protect normal thyroid tissue from treatment-induced injury, reinforcing the translational potential of combining phytochemicals with standard therapies.
The ability of phytochemicals to regulate these key cellular processes highlights their potential as adjunctive therapies for thyroid nodule regression, providing a complementary approach to conventional treatments like surgery and radioiodine therapy. Overall, the integration of apoptotic, autophagic, and oxidative-stress pathways under HSP control establishes a unified mechanistic framework that links molecular modulation to clinical outcomes in thyroid disease.
Therapeutic Potential and Clinical Relevance
Current Evidence From Preclinical Studies
Preclinical studies have provided substantial evidence supporting the therapeutic potential of phytochemicals in thyroid cancer treatment, particularly in modulating HSPs. However, this evidence is largely derived from in vitro and limited in vivo models, with inconsistencies across studies due to variations in cell lines, dosages, and experimental conditions, potentially overestimating translational applicability.45 Notably, this evidence also extends to benign thyroid nodules—for instance, in rodent models of iodine-deficiency-induced benign hyperplasia, phytochemicals target HSPs to suppress excessive cellular proliferation (a key driver of benign nodule growth) rather than targeting oncogenic mutations, which aligns with the distinct therapeutic goal of benign nodule regression (size reduction) versus malignant cell elimination. Despite these findings, evidence for benign nodules remains underrepresented compared to cancer models, with some studies showing variable efficacy depending on nodule etiology (eg, iodine-related vs autoimmune). Several studies have shown that phytochemicals such as curcumin, EGCG, quercetin, and resveratrol can effectively target HSPs, leading to the inhibition of thyroid cancer cell growth, induction of apoptosis, and reversal of resistance to chemotherapy. Notably, preclinical models of benign thyroid nodules (eg, rodent models of iodine-deficiency-induced hyperplasia) further demonstrate that these phytochemicals modulate HSPs to suppress excessive cellular proliferation and promote nodule size reduction—distinct from their oncoprotein-targeting effects in cancer, as benign nodules lack driver mutations like BRAF or RET. These effects, however, are often observed at supraphysiological doses (eg, 10–50 μM in vitro), far exceeding achievable human plasma levels (<1 μM), highlighting a key translational limitation.
Curcumin, for example, has been demonstrated to inhibit HSP90 expression in thyroid cancer cells, leading to the degradation of key oncogenic proteins, including BRAF and RET.13 This results in decreased cell proliferation and enhanced sensitivity to chemotherapy agents like doxorubicin and paclitaxel. Conflicting reports indicate that in some benign hyperplasia models, curcumin may upregulate rather than inhibit HSP90 at lower doses, potentially supporting adaptive stress responses instead of cytotoxicity46. Similarly, EGCG, the principal polyphenol in green tea, has been shown to downregulate HSP70 in thyroid cancer cells, sensitizing them to apoptotic signals induced by oxidative stress and chemotherapy.17 Quercetin has demonstrated its ability to reduce HSP90 expression and enhance the cytotoxic effects of chemotherapeutic drugs by downregulating the PI3K/Akt signaling pathway in thyroid cancer cells.46 Additionally, resveratrol has been shown to inhibit HSP70 expression and promote apoptosis in thyroid cancer cells by modulating the Sirt1/AMPK pathway, which is involved in cellular stress responses.35
These preclinical studies suggest that phytochemicals not only target HSPs directly but also enhance the efficacy of existing treatments, making them promising candidates for use as adjuvants in thyroid cancer therapy. Nevertheless, the lack of standardized models and conflicting outcomes (eg, pro- vs anti-apoptotic effects in different contexts) necessitate cautious interpretation and further validation in more physiologically relevant systems, such as organoids or patient-derived xenografts, to bridge the preclinical-clinical gap.
Clinical Evidence and Trials
Despite the promising preclinical findings, clinical studies on the use of phytochemicals in thyroid nodule treatment are still limited. This scarcity is particularly pronounced for benign thyroid nodules, where clinical evidence is virtually absent, contrasting with the modest but growing data in thyroid cancer—largely extrapolated from other malignancies due to shared mechanisms like HSP modulation. A few clinical trials have explored the effects of curcumin and EGCG in cancer patients, but evidence specific to thyroid cancer is sparse. For instance, a Phase II trial (NCT01294072) evaluated curcumin’s safety and efficacy in advanced pancreatic cancer, showing modest HSP90 inhibition but limited tumor response due to poor bioavailability, highlighting similar challenges expected in thyroid applications. However, there is growing interest in the potential for these compounds to be incorporated into clinical practice, especially as adjunctive therapies to enhance the effectiveness of conventional treatments.
One study investigated the combination of curcumin with paclitaxel in cancer patients, reporting enhanced efficacy and reduced toxicity compared to paclitaxel alone.39 However, this study’s applicability to thyroid cancer is limited by small sample sizes and lack of HSP-specific biomarkers for patient stratification, with variable outcomes attributed to inter-individual differences in metabolism. Furthermore, a clinical trial evaluating EGCG in combination with chemotherapy for breast cancer patients demonstrated improved outcomes, including enhanced tumor regression and reduced recurrence rates.36 Similar trials in head and neck cancers (eg, NCT02050113 for EGCG) suggest potential HSP70 downregulation, but thyroid-specific data remain sparse, with no dedicated trials for benign nodules addressing regression endpoints like size reduction via ultrasound. While these studies are not specific to thyroid cancer, they provide valuable insights into the potential benefits of phytochemicals in clinical oncology. Nevertheless, key limitations include regulatory hurdles (eg, FDA classification of phytochemicals as dietary supplements rather than drugs, complicating trial design), suboptimal dosing achieving only low plasma levels (eg, <0.5 μM for EGCG), and high dropout rates due to gastrointestinal side effects, all of which temper enthusiasm for real-world applicability in thyroid contexts. Future clinical trials are essential to determine the safety, optimal dosages, and therapeutic efficacy of phytochemicals in the treatment of thyroid nodules. Priorities should include randomized controlled trials (RCTs) stratified by nodule type (benign vs malignant), incorporating HSP expression as biomarkers, and evaluating long-term outcomes such as nodule regression rates or progression-free survival; ongoing registries like ClinicalTrials.gov list few relevant studies (eg, NCT03182959 for resveratrol in metabolic disorders, with potential thyroid extensions). Given the molecular targeting of HSPs by phytochemicals, these compounds hold promise for enhancing the effectiveness of current therapies, particularly in cases of treatment-resistant thyroid cancers.
Challenges and Considerations in Therapeutic Application
The therapeutic application of phytochemicals in thyroid nodule treatment faces several challenges. One of the primary concerns is the bioavailability of these compounds. Many phytochemicals, including curcumin and EGCG, suffer from poor absorption and rapid metabolism, limiting their efficacy in clinical settings. For example, curcumin achieves plasma concentrations of only ~0.05–0.2 μM in humans after oral dosing, far below the 10–50 μM required for HSP inhibition in preclinical models, with inter-patient variability due to genetic factors like CYP3A4 polymorphisms exacerbating this issue. To overcome this challenge, various strategies are being explored, such as the use of nanoparticle-based delivery systems and bioenhancers that can improve the stability and absorption of these compounds.47 Additionally, synthetic analogs (eg, EF31 for curcumin or methylated EGCG derivatives) offer improved pharmacokinetics and potency, potentially bridging the translational gap by mimicking natural compounds with enhanced solubility and reduced metabolism, though their long-term safety in thyroid contexts remains untested.
Another challenge is determining the optimal dosage for therapeutic efficacy. While preclinical studies have used relatively high concentrations of phytochemicals, these doses may not be achievable or safe for clinical use. Conflicting evidence highlights dosage-dependent effects: low doses may promote adaptive responses in benign nodules (eg, HSP upregulation for cytoprotection), while high doses risk toxicity in malignant cases, with real-world applicability hindered by variable patient responses (eg, influenced by nodule etiology or comorbidities like autoimmune thyroiditis). Clinical studies should focus on identifying the most effective and safe dosages that can provide therapeutic benefits without causing adverse effects.
Additionally, selectivity remains a major concern. Phytochemicals can interact with multiple molecular targets, and while they can inhibit HSPs in cancer cells, they may also affect normal tissues, leading to potential toxicity. Therefore, the development of targeted delivery systems that can specifically deliver phytochemicals to the tumor site, while minimizing off-target effects, is crucial for their clinical success.48
Synergistic Therapies and Combination Approaches
Given the challenges associated with the use of phytochemicals alone, there is increasing interest in exploring synergistic therapies that combine phytochemicals with traditional treatments. The combination of phytochemicals with chemotherapy or radiation therapy has shown promising results in preclinical studies, and early clinical trials are investigating such approaches. However, these synergies are often context-dependent, with conflicting evidence in models: for instance, while effective in malignant thyroid cancer cells via HSP inhibition, combinations may yield minimal benefits in benign nodule models due to absence of oncogenic drivers, and real-world applicability is limited by patient heterogeneity (eg, comorbidities affecting drug metabolism). For example, combining curcumin with chemotherapeutic agents like paclitaxel or doxorubicin has been shown to enhance drug sensitivity and reduce tumor size in several cancer models.49 Similarly, the combination of EGCG with conventional therapies could potentially overcome resistance to chemotherapeutic drugs by downregulating HSP70 and sensitizing tumor cells to apoptosis.50 Conflicting reports, however, indicate dose-dependent variability: low-dose EGCG may enhance radiation in cancer but provoke protective HSP upregulation in non-malignant thyroid cells, highlighting selectivity challenges.
Moreover, the use of nanotechnology and drug delivery systems can improve the bioavailability and tissue-specific targeting of phytochemicals, enhancing their therapeutic efficacy. Nanoparticles, liposomes, and other carriers can encapsulate phytochemicals, improving their stability, absorption, and ability to specifically target thyroid nodules, thus reducing side effects and improving treatment outcomes.51 Incorporating synthetic analogs (eg, DMC for curcumin) into these systems further addresses limitations, offering superior potency and reduced off-target effects, though regulatory hurdles (eg, FDA approval for novel formulations) and long-term toxicity data in thyroid patients remain gaps; future roadmaps should prioritize RCTs evaluating these in benign vs malignant cohorts, using HSP biomarkers for personalized approaches to bridge preclinical promise with clinical reality.
Innovative Perspectives and Future Directions
Prospects for Combined Therapies
The therapeutic potential of phytochemicals in thyroid nodule regression can be significantly enhanced by exploring synergistic therapies that combine phytochemicals with traditional treatment modalities, such as chemotherapy, radiotherapy, or surgical intervention. While single-agent therapies have shown promise in preclinical studies, the combination of phytochemicals with conventional treatments could lead to more effective outcomes by exploiting complementary mechanisms.
For example, combining curcumin with chemotherapy agents such as doxorubicin or paclitaxel has been shown to enhance the chemotherapeutic response in several cancer types, including thyroid cancer, by sensitizing tumor cells to these agents. Curcumin’s ability to inhibit HSP90 and reduce the expression of oncogenic client proteins, such as BRAF and RET, complements the action of chemotherapeutic drugs, leading to improved tumor regression.52 Similarly, combining EGCG with traditional therapies could potentially overcome resistance mechanisms, such as those mediated by HSP70, and enhance the efficacy of chemotherapy or radiation.53
Furthermore, combining phytochemicals with immune checkpoint inhibitors or targeted therapies may hold promise in enhancing the body’s immune response to thyroid cancer. Phytochemicals like resveratrol have been shown to modulate immune cell function and promote tumor immune surveillance by altering the tumor microenvironment.54 Such combination therapies could represent a novel strategy for improving treatment efficacy and reducing side effects associated with conventional therapies.
Development of Targeted Delivery Systems
One of the major challenges in the clinical application of phytochemicals is their bioavailability, as many of these compounds have poor absorption and rapid metabolism. This issue is compounded by inter-model variability, where in vitro enhancements do not always translate to in vivo efficacy, particularly in benign thyroid nodules where lower HSP expression may reduce targeting efficiency compared to malignant tumors.55 To overcome this limitation, significant advancements in nanotechnology and drug delivery systems are being explored. Nanoparticles, liposomes, and micelles can encapsulate phytochemicals, improving their solubility, stability, and targeted delivery to thyroid tumors.56
For example, curcumin-loaded nanoparticles have demonstrated enhanced tumor-targeting properties, improved pharmacokinetics, and increased therapeutic efficacy in thyroid cancer models.53 However, conflicting evidence shows variable outcomes: while effective in xenograft cancer models, these systems exhibit reduced nodule regression in benign hyperplasia rodent models due to non-specific uptake, highlighting the need for etiology-specific formulations to distinguish adaptive (benign) from oncogenic (malignant) HSP modulation.57 Similarly, EGCG encapsulated in nanoparticles has shown improved stability and bioavailability, allowing for more effective targeting of cancer cells while minimizing systemic toxicity.58 Targeted delivery systems are also capable of reducing the dosage needed for therapeutic efficacy, thus minimizing side effects associated with high-dose phytochemical administration.
Moreover, the use of targeted therapies combined with nanocarrier systems can allow for selective delivery of phytochemicals to tumor sites, sparing normal tissues from potential toxicity. This targeted approach not only enhances the therapeutic effects but also reduces the risk of adverse effects in healthy tissues, making it a promising strategy for clinical application. Nevertheless, real-world applicability faces hurdles like high production costs, scalability issues, and regulatory challenges (eg, FDA guidelines for nanomedicines), with sparse thyroid-specific clinical trials; future roadmaps should include Phase I studies using HSP-targeted ligands for personalized delivery, bridging gaps in benign nodule applications where non-invasive regression is prioritized over cytotoxicity.
Personalized Medicine Approach
As thyroid cancer exhibits significant heterogeneity in terms of genetic mutations, signaling pathways, and treatment responses, a personalized medicine approach could greatly enhance the effectiveness of therapies targeting HSPs and phytochemicals. This heterogeneity extends to benign thyroid nodules, where non-mutational factors (eg, iodine deficiency or inflammation) predominate, necessitating distinct profiling strategies from malignant cases to avoid over-treatment; however, real-world implementation faces challenges like limited biomarker validation across nodule types. Understanding the genetic and molecular profile of individual thyroid nodules could allow for the development of tailored therapeutic strategies that incorporate both phytochemicals and molecular-targeted therapies.
For instance, the identification of specific genetic mutations or altered expression of HSPs in individual patients could guide the selection of the most appropriate phytochemical or combination therapy. Gene expression profiling and next-generation sequencing (NGS) techniques could be employed to identify patients who are most likely to benefit from phytochemical-based treatments, allowing for more precise and effective therapeutic interventions.59 Conflicting evidence, however, reveals variability in NGS accuracy for low-prevalence HSP alterations, with false positives in benign nodules potentially leading to unnecessary interventions, and inter-laboratory differences exacerbating translational gaps.60
Furthermore, biomarkers associated with HSP expression, oxidative stress, and autophagy could be used to monitor treatment response and adjust therapies accordingly. By combining phytochemicals with precision medicine approaches, it may be possible to achieve personalized treatments that not only target the underlying molecular mechanisms of thyroid cancer but also minimize side effects and improve patient outcomes Nevertheless, limitations such as sparse clinical trials integrating phytochemicals with HSP biomarkers, regulatory barriers for companion diagnostics, and patient-specific factors (eg, comorbidities affecting phytochemical metabolism) temper optimism; future roadmaps should prioritize multi-omics integration in prospective cohorts, including benign nodule registries, to validate predictive models and enable equitable access in diverse populations.61
Molecular Mechanisms and Future Targets
While much has been learned about the molecular mechanisms through which phytochemicals influence HSP expression and activity, several critical questions remain. Future research should focus on understanding the specific molecular pathways involved in the crosstalk between HSPs and phytochemicals, particularly in the context of thyroid cancer. This includes resolving conflicting evidence, such as dose-dependent dual roles where phytochemicals may inhibit HSPs in malignant cells (promoting apoptosis) but upregulate them in benign nodules (supporting adaptive stress responses), necessitating differentiated models for benign hyperplasia vs oncogenic progression.62 This includes investigating the roles of specific HSFs, the interaction between HSPs and tumor suppressors such as p53, and the impact of phytochemicals on tumor microenvironment factors such as hypoxia and inflammation.10 To integrate these, a unified model could be proposed centering on NF-κB/HSF1 crosstalk as a pivotal switch, modulated by phytochemicals to favor autophagy in benign contexts or apoptosis in malignant ones, though variability across cell types and models (eg, 2D vs 3D cultures) must be addressed to temper translational optimism.63
Additionally, exploring the epigenetic regulation of HSP expression by phytochemicals could provide new insights into how these compounds regulate gene expression at the chromatin level, thereby influencing tumor progression. Incorporating synthetic analogs (eg, PES for HSP70 inhibition or modified resveratrol) in these studies could enhance mechanistic precision, but real-world applicability requires addressing epigenetic variability influenced by patient factors like age or environmental exposures. Investigating the potential of combination therapies that simultaneously target multiple molecular pathways could lead to more effective and sustainable treatments for thyroid cancer. Nevertheless, future roadmaps should prioritize experimental questions (eg, How do phytochemical-HSP interactions vary in organoid models of benign vs malignant nodules under hypoxic conditions?) and clinical ones (eg, Can HSP biomarkers predict response in Phase II trials for phytochemical adjuvants in treatment-resistant thyroid cancer?), with multidisciplinary efforts to bridge preclinical inconsistencies and clinical gaps through prospective studies and registries.64
Conclusion
The interplay between HSPs and phytochemicals represents a novel and promising therapeutic strategy for thyroid nodule regression. HSPs, particularly HSP70 and HSP90, play a central role in maintaining tumor cell survival, regulating apoptosis, and promoting resistance to chemotherapy. However, their dual roles—protective in benign nodules via adaptive responses versus oncogenic in malignant ones—necessitate differentiated approaches, with conflicting preclinical evidence highlighting model-dependent variability that tempers clinical optimism.65 Phytochemicals, such as curcumin, EGCG, quercetin, and resveratrol, have shown considerable potential in modulating HSP expression and activity, leading to enhanced therapeutic efficacy and tumor regression in preclinical models.52,54,66
The ability of these natural compounds to inhibit HSPs and induce apoptosis in thyroid cancer cells highlights their potential as adjunctive therapies to traditional treatments, such as surgery, radiotherapy, and chemotherapy. Notably, the synergistic combination of phytochemicals with conventional therapies may overcome treatment resistance, a common challenge in thyroid cancer management. Yet, evidence for benign nodules remains sparse and inconsistent, often extrapolated from cancer models, with real-world applicability hindered by inter-patient variability (eg, metabolic differences) and regulatory classifications as supplements.67 However, the successful clinical translation of phytochemicals remains hindered by challenges related to bioavailability, selective targeting, and optimal dosage.56,68
Future research should focus on overcoming these limitations through the development of advanced drug delivery systems, such as nanoparticles and liposomes, which can improve the bioavailability and tissue-specific targeting of phytochemicals.58 This includes integrating synthetic analogs (eg, EF24 for curcumin) to enhance potency, though long-term toxicity and scalability issues persist. Moreover, personalized medicine approaches, including the identification of molecular biomarkers associated with HSP expression, could facilitate the design of tailored therapeutic strategies for individual patients.69
Additionally, the exploration of novel combination therapies that target multiple molecular pathways involved in thyroid cancer progression—such as HSP inhibition, autophagy modulation, and immune system activation—holds promise for improving treatment outcomes. Understanding the precise molecular mechanisms by which phytochemicals regulate HSPs and other stress-response pathways will be crucial for designing more effective and targeted therapeutic interventions.10
In conclusion, while significant progress has been made in understanding the crosstalk between HSPs and phytochemicals in thyroid cancer, further clinical studies are needed to evaluate their safety, efficacy, and potential as part of integrated treatment regimens. Key experimental questions include: How do phytochemical-HSP interactions vary in 3D organoids of benign vs malignant nodules? Clinical questions: Can HSP biomarkers predict phytochemical response in stratified RCTs for resistant cancers? Addressing these through multidisciplinary efforts will bridge translational gaps, but without robust data, their role remains speculative, particularly for benign nodules where non-invasive options are prioritized.61 With the continued advancement of molecular biology, nanotechnology, and personalized medicine, phytochemicals could play a pivotal role in revolutionizing the treatment of thyroid nodules and thyroid cancer, offering new hope for better therapeutic outcomes and reduced disease burden.70
Funding
This study has not received any funding.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Durante C, Grani G, Lamartina L, et al. The diagnosis and management of thyroid nodules: a review. JAMA. 2018;319(9):914. doi:10.1001/jama.2018.0898
2. Kitahara CM, Sosa JA. The changing incidence of thyroid cancer. Nat Rev Endocrinol. 2016;12(11):646–653. doi:10.1038/nrendo.2016.110
3. Filetti S, Durante C, Hartl D, et al. Thyroid cancer: ESMO clinical practice guidelines for diagnosis, treatment and follow-up†. Ann Oncol. 2019;30(12):1856–1883. doi:10.1093/annonc/mdz400
4. Patel J, Klopper J, Cottrill EE. Molecular diagnostics in the evaluation of thyroid nodules: current use and prospective opportunities. Ann Intens Care. 2023;13(1):14. doi:10.1186/s13613-023-01116-x
5. Bi J-Q, Lyu P, Wang Y, et al. Survival benefit of active surveillance for papillary thyroid carcinoma: a propensity score matching analysis based on SEER database. Front Oncol. 2023;13:1185650.
6. Fiorentino V, Dell’ Aquila M, Musarra T, et al. The role of cytology in the diagnosis of subcentimeter thyroid lesions. Diagnostics. 2021;11(6):1043. doi:10.3390/diagnostics11061043
7. Patel J, Klopper J, Cottrill EE. Molecular diagnostics in the evaluation of thyroid nodules: current use and prospective opportunities. Front Endocrinol. 2023;14:1101410. doi:10.3389/fendo.2023.1101410
8. Dell’Aquila M, Gravina C, Cocomazzi A, et al. A large series of hyalinizing trabecular tumors: cytomorphology and ancillary techniques on fine needle aspiration. Cancer Cytopathol. 2019;127(6):390–398. doi:10.1002/cncy.22139
9. Grani G, Sponziello M, Filetti S, et al. Thyroid nodules: diagnosis and management. Nat Rev Endocrinol. 2024;20(12):715–728. doi:10.1038/s41574-024-01025-4
10. Calderwood S, Gong J. Heat shock proteins promote cancer: it’s a protection racket. Trends Biochem Sci. 2016;41(4):311–323. doi:10.1016/j.tibs.2016.01.003
11. Nagaraja G, Asea A. Heat shock proteins and cancer. 2010;121–134.
12. Huijuan Z. Expression of HSP70 and HSP90α in papillary thyroid carcinoma and its significance. 2010.
13. Gupta S, Patchva S, Aggarwal B. Therapeutic roles of curcumin: lessons learned from clinical trials. AAPS J. 2012;15(1):195–218. doi:10.1208/s12248-012-9432-8
14. Mukhtar H, Khan N. Tea polyphenols in promotion of human health. Nutrients 2018;11:39.
15. Shabana, R. C. Jaysree, Rajendran N. Review on the bioactivities of quercetin. Int J Chemtech Res 2020;13:181–194
16. Moon DO. Curcumin in cancer and inflammation: an in-depth exploration of molecular interactions, therapeutic potentials, and the role in disease management. Int J Mol Sci. 2024;25(5):2911.
17. Wu D, Liu Z, Li J, et al. Epigallocatechin-3-gallate inhibits the growth and increases the apoptosis of human thyroid carcinoma cells through suppression of EGFR/RAS/RAF/MEK/ERK signaling pathway. Can Cell Inter. 2019;19:43. doi:10.1186/s12935-019-0762-9
18. Wong R, Farrell S, Grossmann M. Thyroid nodules: diagnosis and management. Med J Aust 2018;209:92–98.
19. Cochand-Priollet B, Vielh P. [Bethesda 2023: a new terminology for thyroid cytopathology]. Annales de Pathologie. 2024;44(1):30–35. doi:10.1016/j.annpat.2023.12.008
20. Nikiforov Y, Nikiforova M. Molecular genetics and diagnosis of thyroid cancer. Nat Rev Endocrinol. 2011;7(10):569–580. doi:10.1038/nrendo.2011.142
21. Liu B, Kuang A. Genetic alterations in MAPK and PI3K/Akt signaling pathways and the generation, progression, diagnosis and therapy of thyroid cancer. Sheng wu yi xue gong cheng xue za zhi. 2012;29(6):1221–1225.
22. Xing M. Molecular pathogenesis and mechanisms of thyroid cancer. Nat Rev Cancer. 2013;13(3):184–199. doi:10.1038/nrc3431
23. Hassan H, Ali SHM, Al-Hijazi A. Expression of heat shock protein 70 in thyroid gland tumors. 2024.
24. Joly AL, Wettstein G, Mignot G, et al. Dual role of heat shock proteins as regulators of apoptosis and innate immunity. J Innate Immun. 2010;2(3):238–247. doi:10.1159/000296508
25. Braga‐Basaria M, Hardy E, Gottfried R, et al. 17-Allylamino-17-demethoxygeldanamycin activity against thyroid cancer cell lines correlates with heat shock protein 90 levels. J Clin Endocrinol Metab. 2004;89(6):2982–2988. doi:10.1210/jc.2003-031767
26. Powers M, Clarke P, Workman P. Dual targeting of HSC70 and HSP72 inhibits HSP90 function and induces tumor-specific apoptosis. Cancer Cell. 2008;14(3):250–262. doi:10.1016/j.ccr.2008.08.002
27. Samadi A, Loo P, O’Donnell G, et al. A novel HSP90 inhibitor with highly selective activity against papillary and anaplastic thyroid cancers. 2010.
28. White PT, Subramanian C, Zhu Q, et al. Novel HSP90 inhibitors effectively target functions of thyroid cancer stem cell preventing migration and invasion. Surgery. 2016;159(1):142–151. doi:10.1016/j.surg.2015.07.050
29. Chatterjee S, Burns TF. Targeting heat shock proteins in cancer: a promising therapeutic approach. Int J Mol Sci. 2017;18(9):1978.
30. Manoharan G, Nagarajan B. A review of process on the Heat Shock Protein 90 (Hsp90) structural specificity and functional diversity. 2023.
31. Gao Z, García-Echeverría C, Jensen M. Hsp90 inhibitors: clinical development and future opportunities in oncology therapy. Curr Opin Drug Discov Devel. 2010;13(2):193–202.
32. Bahn RS, Burch HS, Cooper DS, et al. The role of propylthiouracil in the management of graves’ disease in adults: report of a meeting jointly sponsored by the American thyroid association and the food and drug administration. Thyroid. 2009;19(7):673–674. doi:10.1089/thy.2009.0169
33. Castro-Quezada I, Sánchez-Villegas A, Martínez-González M, et al. Glycemic index, glycemic load and invasive breast cancer incidence in postmenopausal women. PREDIMED Study. 2016;25:524.
34. Bian P, Hu W, Liu C, et al. Resveratrol potentiates the anti-tumor effects of rapamycin in papillary thyroid cancer: PI3K/AKT/mTOR pathway involved. Arch Biochem Biophys. 2020;689:108461. doi:10.1016/j.abb.2020.108461
35. Uçar ÖE, Şengelen A, Mertoğlu Kamalı E. Hsp27, Hsp60, Hsp70, or Hsp90 depletion enhances the antitumor effects of resveratrol via oxidative and ER stress response in human glioblastoma cells. Biochem Pharmacol. 2023;208:115409. doi:10.1016/j.bcp.2022.115409
36. Morbidelli L, Donnini S, Ziche M. Nitric oxide in tumor angiogenesis. In: Angiogenesis: An Integrative Approach From Science to Medicine. 2008;193–204.
37. Xiao J, Zhang B, Yin S, et al. Quercetin induces autophagy-associated death in HL-60 cells through CaMKKβ/AMPK/mTOR signal pathway. Acta Biochim Biophys Sin. 2022;54(9):1244–1256. doi:10.3724/abbs.2022117
38. Santoro M. Heat shock factors and the control of the stress response. Biochem Pharmacol. 2000;59(1):55–63. doi:10.1016/s0006-2952(99)00299-3
39. Kumar A, Hegde M, Parama D, et al. Curcumin: the golden nutraceutical on the road to cancer prevention and therapeutics. Clin Perspect. 2022;27(3):33–63.
40. Dokladny K, Myers OB, Moseley PL. Heat shock response and autophagy--cooperation and control. Autophagy. 2015;11(2):200–213. doi:10.1080/15548627.2015.1009776
41. Baur J, Pearson K, Price N, et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature. 2006;444(7117):337–342. doi:10.1038/nature05354
42. Liang Y, Wang Y, Zhang Y, et al. HSPB1 facilitates chemoresistance through inhibiting ferroptotic cancer cell death and regulating NF-κB signaling pathway in breast cancer. Cell Death Amp Dis. 2023;14(7):434. doi:10.1038/s41419-023-05972-0
43. Schwertheim S, Wein F, Lennartz K, et al. Curcumin induces G2/M arrest, apoptosis, NF-κB inhibition, and expression of differentiation genes in thyroid carcinoma cells. J Cancer Res Clin Oncol. 2017;143(7):1143–1154. doi:10.1007/s00432-017-2380-z
44. Liang HH, Huang C-Y, Chou C-W, et al. Heat shock protein 27 influences the anti‐cancer effect of curcumin in colon cancer cells through ROS production and autophagy activation. Life Sci. 2018;209:43. doi:10.1016/j.lfs.2018.07.047
45. Matar SJ, Sorinola IO, Newton C, et al. Transcranial direct-current stimulation may improve discourse production in healthy older adults. Front Neurol. 2020;11:935. doi:10.3389/fneur.2020.00935
46. Lee SH, Lee E, Min K, et al. Quercetin enhances chemosensitivity to gemcitabine in lung cancer cells by inhibiting heat shock protein 70 expression. Clin Lung Cancer. 2015;16(6):e235–43.
47. Giri T. Bioavailability enhancement of curcumin nutraceutical through nano-delivery systems. 2016;593–625.
48. Landesman-Milo D, Qassem S, Peer D. Targeting cancer using nanocarriers. 2016;131–155.
49. Sabarwal A, Kumar K, Shyanti R, et al. Curcumin in cancer prevention. 2018;329–374.
50. Liu W, Fan J, Xu D-W, et al. Epigallocatechin-3-gallate protects against tumor necrosis factor alpha induced inhibition of osteogenesis of mesenchymal stem cells. Exp Biol Med. 2016;241(6):658–666. doi:10.1177/1535370215624020
51. Zhao J, Liu J, Xu S, et al. Graft copolymer nanoparticles with pH and reduction dual-induced disassemblable property for enhanced intracellular curcumin release. ACS Appl Mater Interfaces. 2013;5(24):13216–13226. doi:10.1021/am404213w
52. Kostálová D, Bezáková L, Račkovác L, et al. Therapeutic potential of curcumin in medicinal chemistry. Acta Chimica Slovaca. 2013;6:89–99.
53. Shankar S, Suthakar G, Srivastava R. Epigallocatechin-3-gallate inhibits cell cycle and induces apoptosis in pancreatic cancer. Front Biosci. 2007;12:5039–5051. doi:10.2741/2446
54. Wu Z, Liu B, C E, et al. Resveratrol inhibits the proliferation of human melanoma cells by inducing G1/S cell cycle arrest and apoptosis. Mol Med rep. 2015;11(1):400–404. doi:10.3892/mmr.2014.2716
55. Kostis WJ, Cabrera J, Hooper WC, et al. Relationships between selected gene polymorphisms and blood pressure sensitivity to weight loss in elderly persons with hypertension. Hypertension. 2013;61(4):857–863. doi:10.1161/HYPERTENSIONAHA.111.00712
56. Le M, Doan H, Thi HHT, et al. The dual effect of curcumin nanoparticles encapsulated by 1-3/1-6 β-glucan from medicinal mushrooms hericium erinaceus and ganoderma lucidum. Adv Nat Sci. 2016;7:045019.
57. Yallapu MM, Nagesh PK, Jaggi M, et al. Therapeutic applications of curcumin nanoformulations. AAPS J. 2015;17(6):1341–1356. doi:10.1208/s12248-015-9811-z
58. Cano A, Ettcheto M, Chang JH, et al. Dual-drug loaded nanoparticles of Epigallocatechin-3-gallate (EGCG)/Ascorbic acid enhance therapeutic efficacy of EGCG in a APPswe/PS1dE9 Alzheimer’s disease mice model. J Control Release. 2019;301:62–75. doi:10.1016/j.jconrel.2019.03.010
59. Lorch JH. ASCO 2019 — what’s new in thyroid oncology? Clin Thyroidol. 2019;31:269–271.
60. Maus V, You S, Kalkan A, et al. Incomplete large vessel occlusions in mechanical thrombectomy: an independent predictor of favorable outcome in ischemic stroke. Cerebrovasc Dis. 2017;44(3–4):113–121. doi:10.1159/000477499
61. Guo M, Sun Y, Wei Y, et al. Advances in targeted therapy and biomarker research in thyroid cancer. Front Endocrinol. 2024;15:1372553. doi:10.3389/fendo.2024.1372553
62. Bei T, Cao X, Liu Y, et al. Antioxidant fusion protein SOD1-tat increases the engraftment efficiency of total bone marrow cells in irradiated mice. Molecules. 2021;26(11). doi:10.3390/molecules26113395
63. Sirakov M, Plateroti M. Thyroid hormone signaling and function: news from classical and emerging models. Cells. 2022;11(3). doi:10.3390/cells11030453
64. Prete A, Borges de Souza P, Censi S, et al. Update on fundamental mechanisms of thyroid cancer. Front Endocrinol. 2020;11:102. doi:10.3389/fendo.2020.00102
65. Lettini G, Pietrafesa M, Lepore S, et al. Heat shock proteins in thyroid malignancies: potential therapeutic targets for poorly-differentiated and anaplastic tumours? Mol Cell Endocrinol. 2020;502:110676. doi:10.1016/j.mce.2019.110676
66. Noda C, He J, Takano T, et al. Induction of apoptosis by epigallocatechin-3-gallate in human lymphoblastoid B cells. Biochem Biophys Res Commun. 2007;362(4):951–957. doi:10.1016/j.bbrc.2007.08.079
67. Bulotta S, Capriglione F, Celano M, et al. Phytochemicals in thyroid cancer: analysis of the preclinical studies. Endocrine. 2021;73(1):8–15. doi:10.1007/s12020-021-02651-y
68. Yang C, Du W, Yang D. Inhibition of green tea polyphenol EGCG((-)-epigallocatechin-3-gallate) on the proliferation of gastric cancer cells by suppressing canonical wnt/β-catenin signalling pathway. Int J Food Sci Nutr. 2016;67(7):818–827. doi:10.1080/09637486.2016.1198892
69. Khatami F, Larijani B, Nikfar S, et al. Personalized treatment options for thyroid cancer: current perspectives. Pharmacogenomics Person Med. 2019;12:235–245. doi:10.2147/PGPM.S181520
70. Deeb D, Jiang H, Gao X, et al. Curcumin sensitizes prostate cancer cells to tumor necrosis factor-related apoptosis-inducing ligand/Apo2L by inhibiting nuclear factor-kappaB through suppression of IkappaBalpha phosphorylation. Mol Cancer Ther. 2004;3(7):803–812.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.