Back to Journals » International Journal of Nanomedicine » Volume 21
Current Research on Aloe-Derived Extracellular Vesicles in Injury Repair
Received 25 November 2025
Accepted for publication 14 February 2026
Published 20 March 2026 Volume 2026:21 584266
DOI https://doi.org/10.2147/IJN.S584266
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. RDK Misra
Yuqing Mu,1 Han Zhang2
1Shandong First Medical University, Jinan, Shandong, 250118, People’s Republic of China; 2Department of Ophthalmology, Shandong Provincial Hospital Affiliated to Shandong First Medical University, Jinan, Shandong, 250022, People’s Republic of China
Correspondence: Han Zhang, Email [email protected]
Abstract: In recent years, plant-derived extracellular vesicle–like nanoparticles have garnered significant interest as promising therapeutic agents and delivery vehicles, owing to their biocompatibility and multifaceted bioactivity. Among these, extracellular vesicles derived from Aloe species (A-EVs) have shown considerable potential in promoting tissue repair. However, a consolidated overview linking their physicochemical properties to in vivo reparative functions and clinical translatability is still lacking. This review systematically summarizes current methods for isolating and characterizing A-EVs, highlighting the technical variability that challenges standardization. Evidence is synthesized demonstrating that A-EVs facilitate injury repair through integrated mechanisms, including potent antioxidant effects via Nrf2/HO-1 pathway activation, anti-inflammatory action via macrophage polarization and NF-κB suppression, and the promotion of cellular proliferation and migration. Notably, emerging research reveals their unique capacity to induce immunogenic cell death (eg, pyroptosis) in diseased tissues, setting them apart from many other plant EV sources. Compared to EVs from other medicinal plants, A-EVs offer a distinctive combination of anthraquinone-enriched cargo, pro-healing protein profiles, and mucoadhesive properties, making them particularly suited for wound and gastrointestinal repair. Despite low immunogenicity and a natural propensity for drug encapsulation, major hurdles—such as scalable production, pharmacokinetic profiling, and rigorous safety assessment—must be overcome to advance clinical translation. By critically evaluating recent progress and existing gaps, this review clarifies the mechanistic basis for A-EVs’ reparative effects and provides a rationale for their future development as standardized, next-generation nanotherapeutics for regenerative medicine.
Keywords: aloe extracellular vesicles, injury repair, isolation and characterization, biological functions, mechanisms of action
Introduction
Extracellular vesicles (EVs) are nanoscale, lipid bilayer-enclosed particles secreted by cells that carry bioactive cargo such as proteins, lipids, and nucleic acids. They play essential roles in mediating intercellular communication across diverse biological systems. In recent years, plant-derived extracellular vesicle-like nanoparticles (PDVLNs) have emerged as a promising alternative to their mammalian counterparts. PDVLNs offer distinct advantages, including abundant sources, lower immunogenicity, scalable production from agricultural resources, and inherent biocompatibility, positioning them as attractive candidates for therapeutic development.1,2
Tissue injury repair, encompassing processes like wound healing and resolution of internal organ damage, remains a central challenge in clinical medicine. Efficient repair requires a precisely coordinated cascade of events, including inflammation control, oxidative stress mitigation, cell proliferation, migration, and extracellular matrix remodeling. While numerous therapeutic agents have been explored, many face limitations such as poor stability, low bioavailability, and off-target effects, highlighting the need for novel, targeted delivery platforms.
The genus Aloe (family Liliaceae), with a history of medicinal use spanning millennia, is a rich source of bioactive compounds. Modern research confirms that its constituents, including polysaccharides, anthraquinones (eg, aloin, aloe-emodin), flavonoids, and phenolic acids, contribute to potent antioxidant, anti-inflammatory, and immunomodulatory activities relevant to tissue repair.3–5 However, the therapeutic efficacy of crude aloe extracts can be limited by rapid degradation, inconsistent absorption, and the complex, sometimes antagonistic, mixture of compounds. This is where the unique properties of aloe-derived extracellular vesicles (A-EVs) become significant. These natural nanoparticles serve as sophisticated delivery systems, encapsulating and protecting aloe’s bioactive cargo. Thus, A-EVs represent more than a refined extract; they constitute a natural, membrane-based delivery system that enhances the stability and bioavailability of key therapeutic molecules and may facilitate targeted delivery, thereby amplifying the reparative potential of aloe beyond that of traditional extracts.6,7
Within the broader landscape of PDVLNs, A-EVs exhibit a distinctive profile. Compared to vesicles from other well-studied plants like ginger or grapefruit, A-EVs are uniquely enriched in specific anthraquinones and flavonoids, which may confer a synergistic combination of antioxidant and anti-inflammatory actions highly suited for the multiphase demands of injury repair.8,9 Moreover, A-EVs have been studied in multiple preclinical models relevant to tissue repair, such as skin wounds and colitis, providing a valuable translational foundation.10
Despite rapid progress, the field faces significant challenges that must be addressed to translate promise into practice. Key issues include a lack of standardized isolation protocols leading to variable EV preparations, incomplete understanding of their in vivo biodistribution and pharmacokinetics, and only partially elucidated molecular mechanisms of action. Furthermore, while their potential as natural nanocarriers for drug delivery is recognized, research into engineering A-EVs for enhanced targeting or combination therapies remains in its infancy.
Therefore, this review aims to: (1) systematically summarize current methods for the isolation and characterization of A-EVs, critically evaluating their strengths and limitations; (2) synthesize the evidence for their multifaceted roles in injury repair—antioxidant, pro-proliferative, anti-inflammatory, and extracellular matrix-modulating—with a focus on underlying molecular pathways such as the Nrf2/HO-1 and NF-κB axes; (3) place A-EVs in the context of other PDVLNs, discussing their comparative advantages and translational potential; and (4) identify critical knowledge gaps, technical hurdles, and future directions, including standardization efforts, mechanistic depth, and their emerging role in nanomedicine as engineered therapeutic carriers. By providing this comprehensive and critical analysis, we seek to clarify the current state of the science and chart a course for the rational development of A-EV-based therapies for tissue repair.
Isolation and Characterization of Aloe-Derived Extracellular Vesicles
Isolation Methods
Obtaining high-purity, high-quality extracellular vesicles (EVs) is fundamental for downstream experimentation. At present, differential centrifugation remains the most commonly used method for isolating aloe-derived EVs.11,12 This technique separates cellular debris, organelles, and large particles based on their sedimentation rates under gradually increasing centrifugal forces, ultimately yielding nanoscale vesicles. Differential centrifugation is simple, reproducible, and suitable for processing large sample volumes, making it widely used in early studies of plant EVs. However, it has notable limitations: ultracentrifugation is time-consuming and sensitive to operational variability; repeated centrifugation can lead to sample loss and reduced EV activity; and co-precipitation of non-vesicular proteins and other impurities may interfere with functional analyses.13,14 To address these issues, researchers have explored complementary techniques to improve purity and yield, though each approach entails trade-offs between scalability, cost, and biological fidelity.
Differential Centrifugation Combined with Ultrafiltration
Ultrafiltration (UF) is a pressure-driven separation technique based on the size-selective properties of semipermeable membranes. A patented protocol for isolating aloe-derived nanovesicles integrates differential centrifugation with ultrafiltration: sequential centrifugation at 350–500×g, 500–1500×g, 1500–2500×g, and 12,000–15,000×g first removes large impurities from aloe extracts. The resulting supernatant containing EVs is then concentrated using ultrafiltration tubes (1500–2500×g) to remove small molecules and excess water. Finally, EVs are pelleted by ultracentrifugation at 100,000–150,000×g and resuspended in 1×PBS containing protease inhibitors (1.5 μg/mL) and 25 mM trehalose. Incorporating ultrafiltration improves both purity and concentration while better preserving EV bioactivity.15 However, the dependence on single-use UF cartridges increases cost and limits scalability.16 Importantly, this hybrid approach reduces contamination from polysaccharides—a major challenge in plant EV isolation due to the high mucilage content in aloe gel—thereby enhancing compatibility with downstream omics analyses.
Polymer-Based Precipitation
Polymer precipitation relies on the ability of polymers to create a hydrophobic microenvironment around EVs, reducing their solubility. Kim et al employed polyethylene glycol (PEG) precipitation after differential centrifugation: the post-centrifugation supernatant was mixed with an equal volume of 2× PEG and incubated at 4 °C, followed by centrifugation at 1500×g. The pellet was washed with PBS and resuspended to obtain aloe-derived EVs. Among different conditions, incubation with 8% PEG for 16 hours produced the highest purity AS-EVs.6 This method is cost-effective and suitable for large sample volumes, but residual polymers may remain in the final preparation, potentially interfering with cell-based assays or mass spectrometry.17 Notably, PEG-based isolation has been shown to co-precipitate exosome-like structures along with non-vesicular nanoparticles in plant systems, raising concerns about specificity when interpreting functional data.
Tangential Flow Filtration (TFF)-Assisted Centrifugation
Tangential flow filtration (TFF) uses pressure to drive sample flow tangentially across a membrane surface, allowing selective passage of smaller components while minimizing membrane fouling.18 In one study extracting EVs from aloe peel, differential centrifugation was combined with TFF: after initial centrifugation, samples were passed through a 0.22 µm filter and concentrated using a 300-kDa TFF membrane.19 TFF enables efficient processing of large volumes and reduces clogging, but it requires specialized instrumentation, increasing both cost and operational complexity.16 Recent comparative studies suggest that TFF yields EV preparations with lower protein-to-particle ratios than ultracentrifugation alone, indicating higher purity—particularly valuable for in vivo applications where immune activation by contaminants must be minimized.20
Critical Perspective on Methodological Variability
The lack of standardized protocols for plant EV isolation remains a major barrier to cross-study comparability. Unlike mammalian EVs, plant EVs are embedded in complex matrices rich in phenolics, polysaccharides, and secondary metabolites, which complicate purification—a challenge not unique to aloe but common across the field of plant-derived extracellular vesicles (PELNs).21 Moreover, no consensus exists on optimal tissue sources (gel vs. peel), harvest timing, or storage conditions—all factors influencing EV yield and composition.22 This heterogeneity contributes to the wide size distribution reported across studies (see Table 1) and underscores the urgent need for community-wide validation of reference materials and benchmarking procedures aligned with MISEV2018/2023 guidelines.23
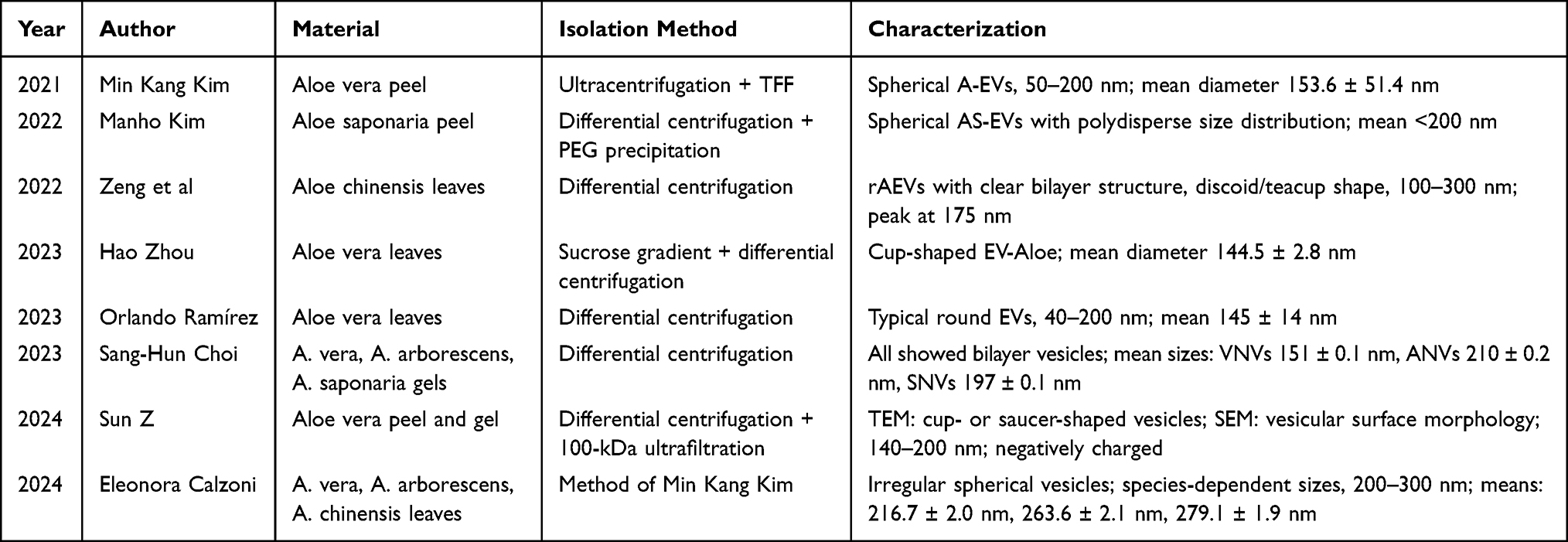 |
Table 1 Summary of Reported EV Morphology and Size from Various Aloe Species |
Characterization Techniques
According to the guidelines of the International Society for Extracellular Vesicles (ISEV), aloe-derived EVs are typically characterized in terms of morphology, particle size distribution, and molecular composition.24 Consistent with these recommendations, current studies employ multimodal approaches.
Morphology and Particle Size
Transmission electron microscopy (TEM), atomic force microscopy (AFM), and scanning electron microscopy (SEM) are essential for morphological characterization. Across multiple studies, aloe EVs isolated from different species, tissues, and centrifugal conditions consistently display classical EV morphology: irregular spherical or discoid vesicles with smooth surfaces and a phospholipid bilayer. Particle size and concentration are commonly determined using nanoparticle tracking analysis (NTA) and dynamic light scattering (DLS). Published data indicate that aloe-derived EVs generally range from 100 to 300 nm in diameter,15,20,25–28 providing a structural basis for understanding their biological behavior. After summarizing the general morphological features and size distribution of aloe-derived EVs, the specific findings reported across different studies are compiled in Table 1.
Composition Analysis and Safety Considerations
Protein composition is typically analyzed using BCA assays, SDS–PAGE, and mass spectrometry. Aloe-derived nanovesicles contain common plant vesicle-related proteins, such as histone H4, heat shock protein 70, and actin.29,30 Notably, four proteins enriched in aloe EVs—glutathione S-transferase P, serum transferrin, RNA polymerase σ factor sigE, and UTP–glucose-1-phosphate uridylyltransferase—were not detected in crude aloe extracts, suggesting active sorting mechanisms.20 Lipidomic analyses have identified multiple phospholipid species, including phosphatidylcholine and phosphatidylethanolamine, consistent with a lipid bilayer structure.20 Small-molecule profiling has revealed anthraquinone compounds such as aloin and aloe-emodin, which possess antioxidant and anti-inflammatory properties. Owing to selective encapsulation, however, their concentrations in EVs are lower than in crude plant extracts.20 This selective packaging implies a regulated biogenesis process, possibly involving ESCRT-independent pathways common in plant cells.23
Despite widespread use of aloe in traditional medicine, potential toxicity of aloe-derived EVs requires attention. Aloin, a major anthraquinone, is known to cause genotoxicity at high doses in rodent models.31 However, current evidence suggests that EV-encapsulated aloin levels are significantly reduced compared to whole extracts, potentially mitigating risk.32 No adverse effects have been reported in murine wound or colitis models treated with aloe EVs at therapeutic doses (≤50 μg/mL or ≤1×109 particles/mL).6,12,15,17–19 Nevertheless, comprehensive toxicokinetic studies—including long-term biodistribution, immunogenicity, and organ accumulation—are still lacking and represent a key knowledge gap before clinical translation.
Biological Functions and Mechanistic Insights of Aloe-Derived Extracellular Vesicles
Antioxidant Activity
Oxidative stress is a key pathological mechanism in tissue injury. Excessive reactive oxygen species (ROS) induce lipid peroxidation, protein carbonylation, and DNA damage, ultimately impairing cellular function.33 Aloe-derived EVs exhibit potent antioxidant capacity through two complementary mechanisms. First, phenolic compounds enriched in aloe EVs—including aloe-emodin and aloin—can directly scavenge ROS.5,34 Second, aloe EVs modulate intracellular oxidative stress pathways to enhance cellular defense.
Using ABTS radical scavenging assays and intracellular ROS inhibition assays, in vitro studies demonstrated that cortex-derived aloe EVs (rAEVs) clear ABTS radicals in a dose-dependent manner, reaching a scavenging efficiency of 93.17% at 30×105 mL−1. rAEVs also suppressed intracellular ROS formation, decreasing ROS fluorescence by 52.55% at a protein concentration of 50 μg/mL.21 The transcription factor Nrf2 is central to the antioxidant defense system, triggering the expression of cytoprotective enzymes such as HO-1, SOD, and CAT upon activation.35,36 Aloe peel–derived EVs (A-EVs) markedly activated the Nrf2 signaling pathway in human keratinocytes (HaCaT) and dermal fibroblasts (HDF), significantly reducing cellular ROS levels by over 40% and increasing cell viability by 25%.19 Consistently, EVs derived from several Aloe species (VNVs, ANVs, SNVs) lowered the expression of the oxidative damage marker 3-NT in the colon and small intestine of mice with acute colitis, confirming their antioxidant role in vivo.24 The multifunctional roles of A-EVs in orchestrating tissue repair, from cargo delivery to coordinated cellular responses, are summarized in Figure 1.
 |
Figure 1 Schematic overview of the isolation, composition, and multifaceted mechanisms of aloe-derived extracellular vesicles (A-EVs) in tissue injury repair. (A) Isolation workflow: Fresh aloe leaves undergo homogenization, followed by sequential differential centrifugation to remove cellular debris and large particles. The crude EV-containing supernatant is further purified using techniques such as ultrafiltration (UF), polyethylene glycol (PEG) precipitation, or tangential flow filtration (TFF) to obtain purified A-EVs. (B) Key cargo components: A-EVs are enriched with bioactive molecules including anthraquinones (eg, aloin, aloe-emodin), flavonoids, specific proteins (eg, heat shock proteins, actin), lipids, and potential nucleic acids. (C) Multimodal reparative actions: Upon delivery to injury sites, A-EVs orchestrate repair through interconnected mechanisms: 1) Antioxidant: Activating the Nrf2/HO-1 pathway to scavenge ROS. 2) Anti-inflammatory and Immunomodulation: Promoting macrophage polarization from M1 to M2 phenotype and inhibiting the NF-κB pathway. 3) Proliferation & Migration: Enhancing fibroblast and keratinocyte motility, potentially via cytoskeletal remodeling. 4) ECM Regulation: Inhibiting TGF-β1-induced myofibroblast differentiation and restoring tight junction proteins. 5) Pyroptosis Induction: Triggering Gasdermin-mediated pyroptosis in dysfunctional cells (eg, in tumor microenvironments), releasing DAMPs and cytokines (eg, IL-18) to stimulate adaptive immunity. |
Promotion of Cell Proliferation and Migration
Cell proliferation and migration are critical events governing wound repair, directly determining the rate and quality of tissue regeneration.37 Multiple studies have shown that aloe-derived EVs significantly enhance these processes. Using WST-8 assays and scratch-wound experiments, AS-EVs from Aloe saponaria were shown to increase human dermal fibroblast proliferation by 2.3-fold and migration by more than threefold at 5×109 particles/mL, exhibiting strong dose dependence. These results suggest that higher concentrations of AS-EVs may be particularly beneficial during chronic wound repair.12
Similarly, rAEVs promoted the proliferation and migration of human epidermal keratinocytes (HaCaT) and human umbilical vein endothelial cells (HUVECs), with both effects increasing in a concentration-dependent manner. At 50 μg/mL, rAEVs increased HUVEC migration to 3.9 times that of controls.6 Scratch assays by Kim et al further showed that A-EVs (1×109 particles/mL) achieved 70% scratch closure in HaCaT cells and 100% in HDFs after 24 hours.15
Although the precise mechanisms remain incompletely defined, cytoskeletal remodeling—particularly actin dynamics—is essential for cell motility. Thus, aloe EVs may enhance cell migration not only by improving the microenvironment through anti-inflammatory and antioxidant actions, but also by modulating actin cytoskeletal reorganization.3,38
Anti-Inflammatory Activity
Inflammation is a protective response essential for early wound defense, but persistent or excessive inflammation delays healing and can lead to chronic inflammatory disorders.39,40 Aloe-derived EVs exhibit notable anti-inflammatory activity via immunomodulation and inhibition of pro-inflammatory signaling pathways.
In an acute colitis mouse model, aloe nanovesicles significantly reduced the expression of inflammatory markers (p-IκB, p-NF-κB) and oxidative stress markers (COX-2, 3-NT) in colonic and intestinal tissues.18 Ramírez et al further evaluated peel-derived vesicles (AVpNVs) in LPS-stimulated RAW264.7 macrophages, THP-1 macrophages, and HaCaT cells. High-dose AVpNVs (500 particles/cell) markedly reduced IL-1β and TNF-α secretion, outperforming both low-dose EVs and the aloe flavonoids quercetin and kaempferol.18
Zhou et al demonstrated that aloe EVs are efficiently internalized by macrophages, promoting a shift from pro-inflammatory M1 (iNOS⁺) to anti-inflammatory M2 (Arg-1⁺) polarization, accompanied by decreased pro-inflammatory cytokines and increased IL-10 secretion.17
There is emerging evidence that aloe EVs may exert anti-inflammatory effects partly through crosstalk between the Nrf2 and NF-κB pathways. A-EVs activate Nrf2/HO-1 signaling under oxidative stress,15 and Nrf2 activation is known to suppress NF-κB through reduced ROS generation and competitive inhibition of transcriptional activity.41,42 Thus, aloe EVs may attenuate inflammation by indirectly limiting NF-κB nuclear translocation.
Regulation of Extracellular Matrix (ECM) Metabolism
The extracellular matrix (ECM) is essential for maintaining tissue structure and function, and its balanced remodeling is crucial for successful wound repair.43,44 Fibroblasts, which synthesize ECM components and differentiate into myofibroblasts during healing, contribute to collagen deposition and wound contraction. However, persistent myofibroblast activation can lead to excessive ECM accumulation and pathological scarring.45
Beyond promoting fibroblast proliferation and migration, aloe EVs help modulate ECM remodeling by suppressing myofibroblast differentiation. AVpNVs significantly inhibited TGF-β1–induced myofibroblast transformation in neonatal human dermal fibroblasts (HDNFs), reducing α-SMA expression.18 In gastrointestinal inflammation models, aloe nanovesicles also preserved ECM integrity by restoring tight junction proteins such as claudin-4 and ZO-1, thereby strengthening mucosal barrier function.17
Induction of Programmed Cell Death and Immune Activation
Emerging evidence positions aloe-derived EVs as regulators of programmed cell death, a critical process in removing damaged cells during injury and cancer. A landmark study demonstrated that aloe vera-derived EV-like particles, particularly those isolated by ultracentrifugation, exhibit potent anti-tumor activity in pancreatic cancer models by inducing pyroptosis, a lytic and inflammatory form of programmed cell death.46 Mechanistically, aloe vera-derived EV-like particles triggered a significant increase in intracellular ROS, which in turn activated caspases-1/3/7/9 and cleaved their downstream effector proteins, Gasdermin D and E (GSDMD/E), leading to characteristic pyroptotic cell swelling and membrane rupture.31
This pyroptosis induction has profound implications for injury repair and immune modulation. The lytic cell death releases damage-associated molecular patterns (DAMPs) and inflammatory cytokines such as IL-18, which can stimulate an adaptive immune response.47 This finding creates a novel mechanistic link between the established antioxidant (ROS-modulating) capacity of aloe EVs and their immunomodulatory functions. It suggests that in contexts like chronic wounds or tumor microenvironments, aloe EVs may not only suppress detrimental inflammation via the Nrf2/NF-κB axis but also actively remodel the tissue by eliminating dysfunctional cells and converting the immune milieu from a suppressed (“cold”) state to a more active (“hot”) one, thereby facilitating clearance and regeneration.48 The reparative effects of A-EVs, as demonstrated across a range of in vitro and in vivo injury models, are comprehensively summarized in Table 2.
 |
Table 2 Summary of Biological Activities of Aloe-Derived Extracellular Vesicles in Various Injury Models |
Comparative Advantages, Biodistribution, and Emerging Applications
Why Aloe? Unique Attributes Among Plant EV Sources
Compared to EVs from other widely studied medicinal plants (eg, ginger, grapefruit, broccoli), aloe-derived EVs offer a distinctive combination of properties relevant to healing.49 They inherit the longstanding safety profile and traditional use of Aloe vera in wound care. Their cargo is uniquely enriched with both antioxidant anthraquinones (aloin, aloe-emodin) and anti-inflammatory lipids/proteins, supporting a multi-targeted approach to repair.25,50 Furthermore, their physical stability, potentially due to membrane composition, and possible mucoadhesive properties make them suitable for topical and oral delivery.10,15 The distinctive profile of A-EVs becomes evident when compared to other well-studied plant EVs, as detailed in Table 3.
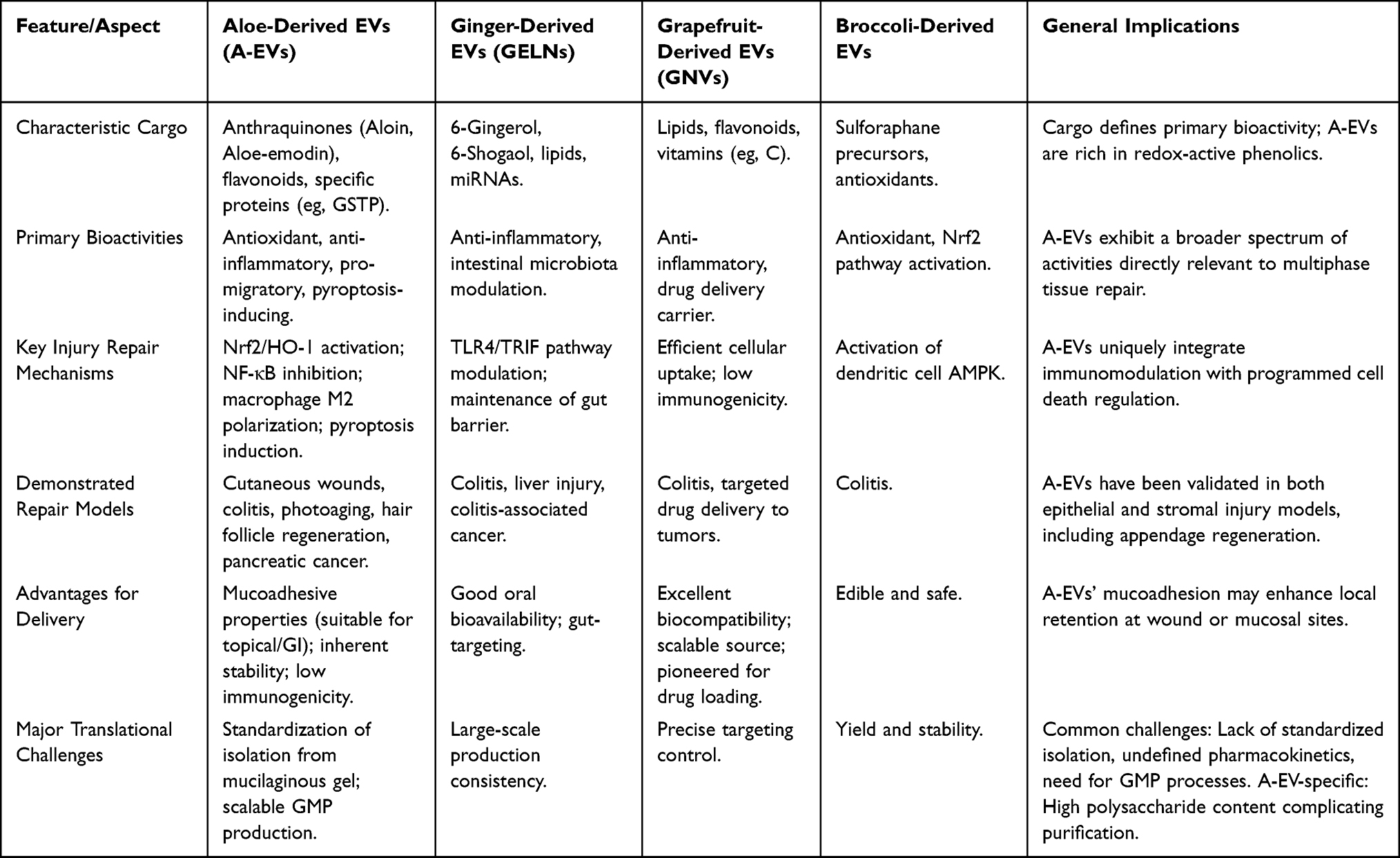 |
Table 3 Comparative Analysis of Aloe-Derived Extracellular Vesicles with Other Representative Plant-Derived EVs |
Pharmacokinetics, Biodistribution, and Therapeutic Carrier Potential
The in vivo journey of aloe EVs remains largely uncharted, a common gap in the PELNs field.51 While evidence confirms oral aloe EVs reach the gut and exert effects,17–19 their systemic distribution, cellular targeting specificity, and clearance pathways after topical or intravenous administration are unknown. Addressing this via tracking studies is crucial for clinical translation.
Concurrently, the innate biocompatibility, low immunogenicity, and nano-scale structure of aloe EVs make them compelling natural drug delivery vehicles.11,13,52 The broader PELNs field has demonstrated remarkable success in loading chemotherapeutic agents (eg, paclitaxel), nucleic acids (siRNA, miRNA), and anti-inflammatory drugs into vesicles from ginger or grapefruit, enhancing drug stability, targeting, and efficacy while reducing off-target toxicity.53 Although similar engineering of aloe EVs is in its infancy, their demonstrated stability and bioactive membrane render them an excellent candidate platform for such applications. Future directions include surface functionalization for active targeting and embedding within hydrogels for sustained release in chronic wounds.47
Conclusion and Perspectives
Current evidence demonstrates that aloe-derived extracellular vesicles (A-EVs) exert therapeutic effects across multiple injury models. They accelerate full-thickness wound closure in mice (from 42.42% in controls to 75.29%), mitigate UV-induced fibroblast damage during photoaging, and reduce body weight loss and intestinal injury in DSS-induced acute colitis models.10,12,19 Notably, A-EVs also promote regeneration of damaged hair follicles, suggesting a capacity not only for structural repair but also for functional restoration. Beyond their intrinsic bioactivity, A-EVs exhibit distinct advantages over other plant-derived EVs: their natural enrichment in redox-modulating anthraquinones (eg, aloin, aloe-emodin), cytoskeleton-regulating proteins, and mucoadhesive surface properties endow them with a unique ability to simultaneously resolve oxidative stress, modulate immune responses, and enhance epithelial cell migration—key requirements in complex, non-healing wounds such as diabetic ulcers or radiation-induced tissue damage.41,50 This multifunctionality positions A-EVs not merely as alternatives to synthetic nanocarriers, but as irreplaceable biological platforms for regenerative applications where coordinated signaling is essential. Nevertheless, the path from bench to bedside remains obstructed by critical translational gaps. First, the lack of consensus on isolation methods (eg, differential ultracentrifugation versus size-exclusion chromatography) results in heterogeneous preparations with variable cargo and bioactivity, undermining reproducibility.16,40 Second, scalable, GMP-compliant production of A-EVs from agricultural sources has not yet been demonstrated; batch consistency, endotoxin control, and long-term stability under storage conditions remain unresolved.45 Third, while preclinical data are compelling, pharmacokinetic profiles, tissue biodistribution, and chronic toxicity in higher mammals are largely unknown—essential prerequisites for regulatory approval. To realize the clinical potential of A-EVs, we propose a focused research roadmap: (i) establish standardized protocols for A-EV isolation, characterization, and potency assays aligned with MISEV guidelines; (ii) deconvolute the contribution of individual cargo components (eg, via proteomic/lipidomic profiling coupled with functional knockdown studies) to identify minimal effective signatures; (iii) engineer A-EVs for enhanced targeting (eg, surface modification for homing to inflamed endothelium) or combination therapy (eg, co-loading with growth factors or siRNA); and (iv) prioritize validation in clinically relevant models of chronic wounds, inflammatory bowel disease, and mucosal injury—conditions with high unmet medical need and limited therapeutic options. In summary, A-EVs represent a convergence of traditional herbal wisdom and modern nanomedicine. With strategic investment in standardization, mechanistic depth, and translational validation, they hold realistic promise as next-generation biologics for regenerative medicine—not as panaceas, but as precisely defined, naturally derived therapeutics tailored to the dynamic demands of tissue repair.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Gao C, Chen Y, Wen X, et al. Plant-derived exosome-like nanoparticles in tissue repair and regeneration. J Mat Chem B. 2025;13(7):2254–11. doi:10.1039/d4tb02394c
2. Yi Q, Xu Z, Thakur A, et al. Current understanding of plant-derived exosome-like nanoparticles in regulating the inflammatory response and immune system microenvironment. Pharmacol Res. 2023;190:106733. doi:10.1016/j.phrs.2023.106733
3. Catalano A, Ceramella J, Iacopetta D, et al. Aloe vera—An extensive review focused on recent studies. Foods. 2024;13(13):2155. doi:10.3390/foods13132155
4. Khaldoune K, Fdil N, Ait Ali M. Exploring Aloe vera: a comprehensive review on extraction, chemical composition, biological effects, and its utilization in the synthesis of metallic nanoparticles. Biocatal Agric Biotechnol. 2024;57:103052. doi:10.1016/j.bcab.2024.103052
5. Kumar S, Kalita S, Basumatary IB, et al. Recent advances in therapeutic and biological activities of Aloe vera. Biocatal Agric Biotechnol. 2024;57:103084. doi:10.1016/j.bcab.2024.103084
6. Kim M, Park JH. Isolation of aloe saponaria-derived extracellular vesicles and investigation of their potential for chronic wound healing. Pharmaceutics. 2022;14(9):1905. doi:10.3390/pharmaceutics14091905
7. Wu W, Zhang B, Wang W, et al. Plant-derived exosome-like nanovesicles in chronic wound healing. Int J Nanomed. 2024;19:11293–11303. doi:10.2147/IJN.S485441
8. Froldi G, Baronchelli F, Marin E, Grison M. Antiglycation activity and HT-29 cellular uptake of aloe-emodin, aloin, and aloe arborescens leaf extracts. Molecules. 2019;24(11):2128. doi:10.3390/molecules24112128
9. Kim M, Jang H, Kim W, Kim D, Park JH. Therapeutic applications of plant-derived extracellular vesicles as antioxidants for oxidative stress-related diseases. Antioxidants. 2023;12(6):1286. doi:10.3390/antiox12061286
10. Barzin M, Bagheri AM, Ohadi M, Abhaji AM, Salarpour S, Dehghannoudeh G. Application of plant-derived exosome-like nanoparticles in drug delivery. Pharm Dev Technol. 2023;28(5):383–402. doi:10.1080/10837450.2023.2202242
11. Zhang Y, Bi J, Huang J, et al. Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int J Nanomed. 2020;15:6917–6934. doi:10.2147/IJN.S264498
12. Davidson SM, Boulanger CM, Aikawa E, et al. Methods for the identification and characterization of extracellular vesicles in cardiovascular studies: from exosomes to microvesicles. Cardiovasc Res. 2023;119(1):45–63. doi:10.1093/cvr/cvac031
13. Cvjetkovic A, Lötvall J, Lässer C. The influence of rotor type and centrifugation time on the yield and purity of extracellular vesicles. J Extracell Vesicles. 2014;3(1):23111. doi:10.3402/jev.v3.23111
14. Ramesh D, Shaju MC, Kumar A, et al. Biological, pathological, and multifaceted therapeutic functions of exosomes to target cancer. Oncol Res. 2024;32(1):73–94. doi:10.32604/or.2023.030401
15. Jiangsu University; The First People’s Hospital of Kunshan. Aloe exosome-like nanovesicles capable of delaying skin photoaging, extraction method, and application: CN202411445126.X;2025.
16. Mukerjee N, Bhattacharya A, Maitra S, et al. Exosome isolation and characterization for advanced diagnostic and therapeutic applications. Mater Today Bio. 2025;31:101613. doi:10.1016/j.mtbio.2025.101613
17. Janouskova O, Herma R, Semeradtova A, et al. Conventional and nonconventional sources of exosomes—Isolation methods and influence on their downstream biomedical application. Front Mol Biosci. 2022;9:846650. doi:10.3389/fmolb.2022.846650
18. Heinemann ML, Vykoukal J. Sequential filtration: a gentle method for the isolation of functional extracellular vesicles. In: Methods in Molecular Biology. Humana Press; 2017:33–41. doi:10.1007/978-1-4939-7253-1_4
19. Kim MK, Choi YC, Cho SH, et al. The antioxidant effect of small extracellular vesicles derived from Aloe vera peels for wound healing. Tissue Eng and Regener Med. 2021;18(4):561–571. doi:10.1007/s13770-021-00367-8
20. Calzoni E, Bertoldi A, Cesaretti A, et al. Aloe extracellular vesicles as carriers of photoinducible metabolites exhibiting cellular phototoxicity. Cells. 2024;13(22):1845. doi:10.3390/cells13221845
21. Fu H, Yong S, Song Y, et al. Advances in plant-derived extracellular vesicles: implications for apple-derived EVs. Plants. 2025;14(22):3425. doi:10.3390/plants14223425
22. Xu JY, Xiao YL, Yu ZL. From biomedical mechanisms to clinical applications: research progress in plant-derived vesicles for cancer therapy. Int J Nanomed. 2026;21:1–25. doi:10.2147/IJN.S545633
23. Luisotti L, Germelli L, Piccarducci R, Giacomelli C, Marchetti L, Martini C. Extracellular vesicles as vehicles for small non-coding RNA therapeutics: standardization challenges for clinical translation. Extracell Vesicles Circ Nucleic Acids. 2025;6(3):403. doi:10.20517/evcna.2025.33
24. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
25. Zeng L, Wang H, Shi W, et al. Wound repair effects based on Aloe cortex-derived exosome-like nanovesicles. J Fujian Med Univ. 2022;56(6):489–497. doi:10.3969/j.issn.1672-4194.2022.06.005
26. Zhou H, Peng K, Wang J, et al. Aloe-derived vesicles enable macrophage reprogramming to regulate the inflammatory immune environment. Front Bioeng Biotechnol. 2023;11:1339941. doi:10.3389/fbioe.2023.1339941
27. Ramírez O, Pomareda F, Olivares B, et al. Aloe vera peel-derived nanovesicles display anti-inflammatory properties and prevent myofibroblast differentiation. Phytomedicine. 2024;122:155108. doi:10.1016/j.phymed.2023.155108
28. Choi SH, Eom JY, Kim HJ, et al. Aloe-derived nanovesicles attenuate inflammation and enhance tight junction proteins for acute colitis treatment. Biomater Sci. 2023;11(16):5490–5501. doi:10.1039/d3bm00591g
29. Nemati M, Singh B, Mir RA, et al. Plant-derived extracellular vesicles: a novel nanomedicine approach with advantages and challenges. Cell Commun Signaling. 2022;20(1):69. doi:10.1186/s12964-022-00889-1
30. Deng F, Miller J. A review on protein markers of exosomes from different bio-resources and the antibodies used for characterization. J Histotechnol. 2019;42(4):226–239. doi:10.1080/01478885.2019.1646984
31. Zeng L, Wang H, Shi W, et al. Aloe-derived nanovesicle as a functional carrier for indocyanine green encapsulation and phototherapy. J Nanobiotechnol. 2021;19(1):439. doi:10.1186/s12951-021-01195-7
32. Cao W, Peng S, Yao Y, et al. A nanofibrous membrane loaded with doxycycline and printed with conductive hydrogel strips promotes diabetic wound healing in vivo. Acta Biomater. 2022;152:60–73. doi:10.1016/j.actbio.2022.08.048
33. Velázquez-López RA, Torres-Gregorio M, García-Rojas V, et al. In vitro study of the biological activity of Aloe barbadensis Miller and its physicochemical characterization. Plant Foods Human Nutr. 2025;80(1):55. doi:10.1007/s11130-025-01299-7
34. Kahroba H, Davatgaran-Taghipour Y. Exosomal Nrf2: from antioxidant and anti-inflammatory response to wound healing and tissue regeneration in age-related diseases. Biochimie. 2020;171–172:103–109. doi:10.1016/j.biochi.2020.06.005
35. Quatrana A, Petrillo S, Torda C, et al. Redox homeostasis and inflammation in fibroblasts of patients with Friedreich ataxia: a possible cross-talk. Front Mol Neurosci. 2025;18:1571402. doi:10.3389/fnmol.2025.1571402
36. Franco JJ, Atieh Y, Bryan CD, et al. Cellular crowding influences extrusion and proliferation to facilitate epithelial tissue repair. Mol Biol Cell. 2019;30(16):1939–1950. doi:10.1091/mbc.E19-02-0106
37. Phimnuan P, Dirand Z, Tissot M, et al. Beneficial effects of a blended fibroin/Aloe gel extract film on the biomolecular mechanism(s) via the MAPK/ERK pathway relating to diabetic wound healing. ACS Omega. 2023;8(7):6813–6824. doi:10.1021/acsomega.2c07507
38. Gunaydin-Akyildiz A, Yanikoglu RS, Gulec M, et al. Emodin and aloe-emodin regulate skin cell migration through the MAP kinase pathway and affect Caenorhabditis elegans thermotolerance. BMC Mol Cell Biol. 2023;24(1):23. doi:10.1186/s12860-023-00486-1
39. Moon HH, Kreis NN, Friemel A, et al. Mitotic centromere-associated kinesin (MCAK/KIF2C) regulates cell migration and invasion by modulating microtubule dynamics and focal adhesion turnover. Cancers. 2021;13(15):3734. doi:10.3390/cancers13153734
40. Fäßler F, Javoor G, Schur FK. Deciphering the molecular mechanisms of actin cytoskeleton regulation in cell migration using cryo-EM. Biochem Soc Trans. 2023;51(1):87–99. doi:10.1042/BST20220221
41. Liu XG, Huang GH, Huang J. The role and research progress of immune inflammation in skin wounds. Mod Immunol. 2018;38(2):165–169.
42. Xie HL, Shi HP. Current status and progress of inflammation research. e-J Tumor Metabol Nutr. 2024;11(1):1–7.
43. Pérez S, Rius-Pérez S. Macrophage polarization and reprogramming in acute inflammation: a redox perspective. Antioxidants. 2022;11(7):1334. doi:10.3390/antiox11071334
44. Guo X, Chen Y, Steinmetz R, et al. NFκB promotes oxidative stress-induced necrosis and ischemia–reperfusion injury through the NRF2-ARE pathway. Circulat Res. 2019;125(9):e106–e107. doi:10.1161/CIRCRESAHA.119.315248
45. Cai F, Chen W, Zhao R, et al. Mechanisms of Nrf2 and NF-κB pathways in diabetic wounds and potential treatment strategies. Mol Biol Rep. 2023;50(6):5977–5988. doi:10.1007/s11033-023-08099-0
46. Ceballos-Santa MC, Gaborski TR, Schulze S, Wuertz-Kozak K. Targeting fibrotic scars with extracellular vesicles extracted from mature aloe vera: enhanced antioxidant, anti-inflammatory, and antifibrotic activity through M2 macrophage polarization and myofibroblasts inhibition. bioRxiv. 2025. doi:10.1101/2025.06.17.660207
47. Zeng M, Liu M, Tao X, Yin X, Shen C, Wang X. Emerging trends in the application of extracellular vesicles as novel oral delivery vehicles for therapeutics in inflammatory diseases. Int J Nanomed. 2024;19:8573–8601. doi:10.2147/IJN.S475532
48. Bai M, Li Z, Shi T, et al. Exogenous extracellular vesicles as emerging platforms in translational medicine. BIO Integr. 2025;6(1):973. doi:10.15212/bioi-2025-0122
49. Luo YX. Extracellular matrix and tissue regeneration and repair. Chin J Tissue Eng Res. 2021;25(11):1785–1790. doi:10.12307/2021.248
50. Preußer C, Salander DJ, Szymanski W, et al. Isolation Defines Identity: functional Consequences of Extracellular Vesicle Purification Strategies. Adv Healthcare Mater. 2026;e04684. doi:10.1002/adhm.202504684
51. Kyriakopoulou K, Piperigkou Z, Tzaferi K, et al. Trends in extracellular matrix biology. Mol Biol Rep. 2023;50(1):853–863. doi:10.1007/s11033-022-07931-y
52. Jiang D, Guo R, Machens HG, et al. Diversity of fibroblasts and their roles in wound healing. Cold Spring Harbor Perspect Biol. 2023;15(3):a044472. doi:10.1101/cshperspect.a044472
53. Zeng L, Shi W, Wang H, et al. Codelivery of π–π stacked dual anticancer drugs based on Aloe-derived nanovesicles for breast cancer therapy. ACS Appl Mater Interfaces. 2022;14(24):27686–27702. doi:10.1021/acsami.2c06546
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.