Back to Journals » Clinical Interventions in Aging » Volume 14
Current and emerging biomarkers of frailty in the elderly
Authors Al Saedi A ![]() , Feehan J
, Feehan J ![]() , Phu S
, Phu S ![]() , Duque G
, Duque G ![]()
Received 16 November 2018
Accepted for publication 18 January 2019
Published 19 February 2019 Volume 2019:14 Pages 389—398
DOI https://doi.org/10.2147/CIA.S168687
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Richard Walker
Ahmed Al Saedi,1,2 Jack Feehan,1,2 Steven Phu,1,2 Gustavo Duque1,2
1Australian Institute for Musculoskeletal Science (AIMSS), University of Melbourne and Western Health, St Albans, Melbourne, VIC, Australia; 2Department of Medicine, Western Health, Melbourne Medical School, University of Melbourne, St Albans, Melbourne, VIC, Australia
Abstract: The term “frailty” is used to describe a subset of older adults who appear weaker and more vulnerable than their age-matched counterparts, despite having similar comorbidities, demography, sex, and age. The diagnosis of frailty is usually clinical and based on specific criteria, which are sometimes inconsistent. Therefore, there is an increasing need to identify and validate robust biomarkers for this condition. In this review, we summarize current evidence on the validity and practicality of the most commonly used biomarkers for frailty, while also comparing them with new upcoming strategies to identify this condition.
Keywords: frailty, biomarkers, COP cells, mesenchymal stem cells, disability, elderly
Introduction
For several years, medical practitioners have used the term “frailty” to describe a subset of older adults that appear weaker and more vulnerable than their age-matched counterparts, despite having similar comorbidities, demography, sex, and age.1 With regard to the clinical diagnosis of frailty, the operational definition proposed by Fried et al2 is based on five criteria. The presence of one or two criteria was defined as “prefrailty” and three or more criteria as frailty. Although this definition remains the most solid parameter used in clinical practice,3,4 some components of this phenotype are more relevant than others, thus limiting its applicability in multiple clinical settings.5
To overcome the limitations of these criteria, several groups have proposed new diagnostic criteria. Among these proposed diagnostic tools, the Frailty Index proposed by Rockwood et al6 has received growing acceptance as an alternative method of identifying frail older persons.7 This index is a count of 70 clinical deficits and includes the presence and severity of current diseases, ability in the activities of daily living, and physical and neurological signs on clinical examination. In addition, each deficit is scored according to severity or frequency of the problem. Despite both Fried et al’s and Rockwood et al’s criteria identifying prefrailty and frailty in the clinical setting, their validity to predict and recognize progression from robust or prefrail to frail is still limited.
The lack of a unique operational definition for frailty and the complex underlying pathophysiology make the development of biomarkers for this condition extremely challenging. Indeed, the current definitional ambiguities of frailty impact the accuracy, specificity, and sensitivity of individual biomarkers proposed so far.8 The most popular circulating markers are those related to the inflammatory response (eg, C-reactive protein [CRP], IL6, and tumor necrosis factor alpha [TNFα]). Other proposed biomarkers include clinical parameters (eg, hemoglobin, glomerular filtration rate, and albumin), hormones (eg, dehydroepiandrosterone [DHEA] sulfate, testosterone, insulin-like growth factor-1 [IGF1] and vitamin D [VitD]), products of oxidative damage (eg, advanced glycation end products, protein carbonyls, and oxidized low-density lipoproteins), or antioxidants (eg, sirtuins and α-tocopherol).8–10
However, the major limitation of these biomarkers is that most are also considered biological markers of aging;10 therefore, alterations in serum levels could also be associated with the aging process per se, independently of the presence of frailty. In addition, most of these proposed biomarkers are able to capture only single aspects of the conditions, are weak predictors of disease progression (eg, from prefrail to frail), or are poorly associated with clinically meaningful outcomes (ie, disability).
In this review, we summarize the present evidence on current and upcoming biomarkers for frailty (Figure 1) and their advantages, disadvantages, validity, and predictive value. As life expectancy increases worldwide, the prevalence and clinical consequences of frailty will increase rapidly. Therefore, early diagnosis of frailty and timely interventions to prevent poor outcomes (eg, disability, falls, fractures) is crucial.
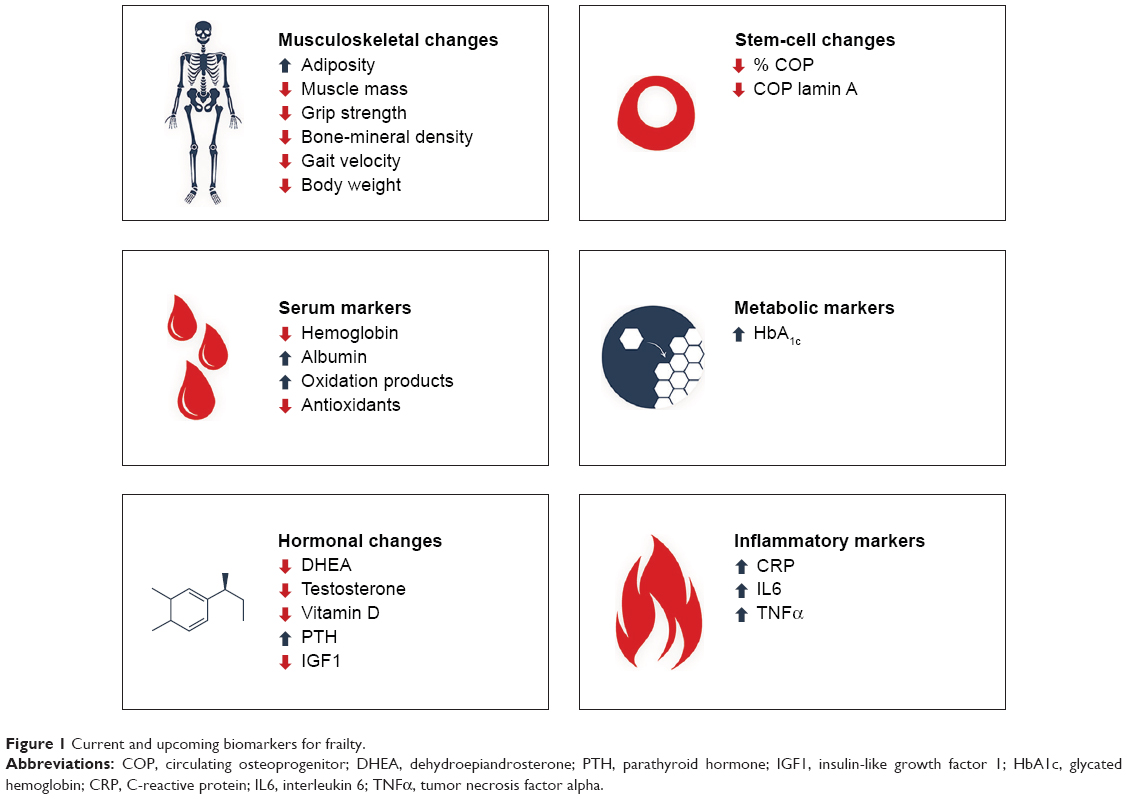 | Figure 1 Current and upcoming biomarkers for frailty. |
Frailty and inflammation: cause and effect, or two sides of the same pathological coin?
Inflammation has been associated with the aging process, purported as a key physiological mechanism in the gradual decline over the life span.11 It follows logically that an acceleration of this low-grade inflammatory activity could provide an underlying cause of frailty syndrome. This hypothesis has driven research into several common inflammatory markers and their relationship to frailty for use as biomarkers to aid in diagnosis and track progression of the syndrome. Most of the research in this area has centered on IL6, CRP, and TNFα.
IL6 and TNFα are cytokines closely involved with the acute-phase inflammatory response produced by a wide range of human cells, including macrophages, myocytes, and adipocytes. Increasing levels of these cytokines have been shown to be associated with frailty by several authors,12–19 making them attractive targets for screening and monitoring of the condition. Increased levels of IL6 and TNFα have also been demonstrated to correlate with a number of important components of frailty, including decreased muscle mass,20 grip strength,21,22 and bone mass,23 as well as depression.24 This systemic role of these inflammatory cytokines in multiple components of the frailty phenotype strengthens their case for inclusion as key biomarkers. Similarly, CRP is another target of interest for those seeking to identify a serum measure of the inflammatory process involved in frailty. A by-product of the acute-phase reaction, CRP levels increase in the presence of inflammation. Similarly to IL6 and TNFα, increased levels of CRP have been shown to correlate with frailty14,25–28 and other conditions closely linked to it, such as osteoporosis.29 These markers are simple to measure and correlate not only with frailty but with negative patient outcomes and comorbidities, providing insight to the systemic effects of the syndrome. However, inflammatory biomarkers have been shown to be unable to predict the transition of patients from robust to prefrail and frail.30–32
While IL6, TNFα, and CRP have been used in clinical trials as potential biomarkers of existing frailty, their usefulness in early identification, risk stratification, and prevention has been shown to be limited. These inflammatory markers are aspecific, meaning that they give little understanding of the underlying processes involved in the development of frailty or insight into a specific patient’s condition. This means that while they provide some diagnostic value, their future role in the ongoing management of a patient is questionable, as they cannot be used to measure such variables as muscle or bone degeneration, metabolic or hormonal decline, or progression toward disability. Additionally, all three are increased in a vast range of inflammatory, neoplastic, or infectious conditions, leaving them unable to provide reliable diagnostic information on their own, and as these markers have been shown to increase with age,33 clear cutoff points are required for accurate use; however, these have not yet been defined. In addition, confounding factors and inconsistent results in prospective studies dampen their ability to allow for screening and prevention of the progression into frailty. This is critically limiting, due to the low likelihood of progression from an established frail state back into a robust one, making their eventual utilization as a screening tool unlikely.
Hormonal changes: not just for teenagers
As the human body ages, a range of key hormonal changes occur, resulting in functional deterioration across a range of connected physiological systems. Much like the markers of inflammation, changes in the endocrine system provide a logical basis for the decline into frailty. Due to the phenotypic changes of frailty being so commonly characterized by its musculoskeletal profile, much of the research drawing links with changes in the endocrine system has focused on hormones with established effects on the bone and muscular systems. These include testosterone and its precursor DHEA, parathyroid hormone (PTH), VitD, and IGF1.
The human gonadal hormones are vital anabolic factors in musculoskeletal physiology, and their age-linked decline is well understood. As the development of skeletal fragility is closely associated with the decline in these hormones, as well as being a key part of the manifestation of frailty, their monitoring can provide key information on patient functional decline. A decrease in testosterone levels has been associated with frailty in both men34,35 and women.35,36 Indeed, testosterone-replacement therapy has been shown to have a beneficial effect on the physical manifestations of frailty, increasing muscle and bone mass,37 though some have reported no overall effect on function.38,39 While this implies a functional relationship between the two, some other studies have cast doubt on the relationship, finding no correlation,40 or only in women.36 Additionally, there is a small amount of evidence to suggest that testosterone could be used to screen for and predict frailty in men.34
In an attempt to find a more consistent measure between sexes, DHEA has also been examined in frail patients. As DHEA is the precursor to both testosterone and estrogen, its circulating level is more similar in men and women than testosterone, though its role in pathology between sexes has still been shown to differ.41 As seen in testosterone, lower levels of DHEA are associated with increasing rates of frailty,16,42 and its supplementation can improve musculoskeletal symptoms of the condition.43,44 Another study in older women found that low levels in either hormone had no significant correlation with frailty, but accumulation of multiple anabolic deficiencies was strongly predictive for it.45 While these two androgens appear to have a weak correlation with frailty, it is difficult to conclude whether it is a parallel manifestation of the same condition or a causative factor for the syndrome. Additionally, differences in biology between men and women make them more difficult to research and apply clinically.
Hyperparathyroidism is a known factor in musculoskeletal fragility,46 suggesting a possible link to frailty that has been explored with variable results. Some studies have shown increased levels of PTH to correlate with frailty;47,48 however, other studies have found that there was no relationship,49,50 casting doubt on its ability to predict and diagnose frailty. Despite unclear evidence for a role in the diagnosis or prediction of frailty, PTH does appear to have the capacity for identification of mortality and morbidity in frailty. It has been shown to be an independent predictor of time to fall in frail men and women in aged-care facilities, something of major concern in these patients.51 It has also been demonstrated that even subtle elevations in PTH levels are correlated with 2-year all-cause mortality in very frail patients with poor prognosis.52 While further research is required to examine the relationship between PTH and frailty and fully explore its ability to predict morbidity and mortality, it does hold some promise of utility in managing these complex patients.
In an increasingly sedentary population, VitD deficiency is becoming highly prevalent and is attracting significant attention from clinicians and researchers. Considered a hormone more than a vitamin, circulating levels of VitD are well established as a vital element in a wide range of physiological processes, including musculoskeletal development and maintenance, mood regulation, and autoimmune disease, among others. Low levels of VitD have been shown to correlate with increased rates of frailty,31,53–55 as well as correlating strongly with the transition into frailty and prefrailty.31,56 Indeed, VitD supplementation could cause a reversal from prefrail to robust, but not from frail to prefrail, providing some evidence of a mechanistic relationship between the two.57 Despite these promising findings, VitD is a challenging biomarker, due to its highly variable normal range. Wide variation is seen in circulating VitD levels among seasons, geographical areas, and racial groups, limiting its clinical usability. Additionally, there is widespread dissent over what are considered healthy and unhealthy circulating levels of the hormone. These issues make the identification of a specific diagnostic value of VitD deficiency in frailty a daunting task.
IGF1 is a hormone with key functions in anabolic processes in a wide range of body tissues, and deficiency is associated with a host of manifestations, many of which are shared by frailty. This makes it a logical target for investigation in frailty, as both a causative factor and a screening tool. Low levels of IGF1 have been correlated with higher incidence of frailty16,58 or such components of frailty as sarcopenia;59 however, other studies refute this connection.31 The relationship between IGF1 and frailty, while logical, requires further analysis to elucidate fully. It is possible that the decreases in IGF1 are due to other physiological changes in the frail patient. For example, it has been shown that increases in TNFα have reciprocal decreases in IGF1,60 potentially a relationship reflected in frailty.
Hormonal biomarkers have several major strengths over other types of serum analysis in the diagnosis and monitoring of frailty. Hormone-function tests are simple, already in widespread use, and have a well-evidenced series of parameters describing normal and abnormal results. Additionally, they give some indication of an underlying mechanism for the condition, while at the same time assisting with a diagnosis. Endocrine dysregulation has a well-documented series of physiological effects, so treatment strategies can be applied to a specific patient’s frailty presentation. Some hormonal biomarkers appear to have some capacity for screening and prediction of a transition into prefrailty and frailty, providing essential early intervention and treatment. However, before they can be translated effectively into clinical use, further research is required to confirm their relationships to the syndrome, establish effective cutoff points for all patient demographics, and identify their ability to predict onset and transition through the different stages of frailty.
Glucose dysregulation: not as sweet as it seems
Metabolic dysregulation is becoming an issue of increasing concern in healthcare systems globally. As increasing numbers of people become diabetic, the burden of its associated complications become harder to bear. While most commonly associated with cardiovascular and neurological complications, increasing evidence has begun to identify the onset of frailty as a new sequela of the disease. This knowledge has prompted investigation into the markers of metabolism to inform and predict the onset of frailty in older adults, as well as to identify new avenues to prevent its onset. The most promising of these is glycated hemoglobin (HbA1c). HbA1c is a commonly employed measure in diabetes management, used to estimate the average plasma-glucose level over the previous 3 months. As plasma-glucose concentrations increase, there is a reciprocal rise in the amount that binds to hemoglobin, providing a way to measure how much blood sugar a red blood cell has encountered over its 3-month life span. HbA1c has been shown a number of times to indicate risk of frailty, with increased levels correlating with greater incidence of the syndrome.61–64 HbA1c is a simple laboratory test that is done regularly in geriatric and general practice settings, and it is not subject to the fluctuations associated with many other tests of glucose regulation, strengthening the case for its use as a frailty biomarker. However, it has not yet been shown to have the capacity to predict the transition from robust to prefrail or prefrail to frail, making it unable to be used as a screening tool. Additionally, it has not been shown to correlate with poorer frailty outcomes or disability. Indeed, much lower HbA1c levels have been associated with increased falls in the frail elderly,65 confusing the picture on the relationship between glycemic control and outcomes in these patients.
Aging blood: circulating hallmarks of frailty
A range of commonly used serum markers have been evaluated for potential relationships with frailty that could be exploited to assist in early diagnosis and intervention. One hypothesis on the pathogenesis of frailty suggests a systemic dysregulation of protein synthesis and metabolism as part of the underlying process resulting in its presentation. Serum-protein levels are well understood and offer a simple insight into the functional status of the patient’s hematological system. While a wide range of serum markers have been investigated, hemoglobin, glomerular filtration rate, and albumin levels are particularly noteworthy as potential indicators of frailty.
A decrease in the level of circulating hemoglobin,66–69 albumin,19,70,71 and low glomerular filtration rate72 have been shown by several authors to correlate with the incidence of frailty. These markers also correlate symptomatically with some of the key symptoms of frailty, such as exhaustion and muscle weakness, and likely contribute to the poor outcomes of patients with the condition. While these biomarkers may be well correlated with the incidence of frailty, their relationship with several comorbidities common in the elderly makes their use more challenging as a standalone indicator of frailty.
Researchers have also hypothesized that the onset of frailty may stem from an accelerated accumulation of cellular damage through oxidation processes. This has led to several studies investigating both increased levels of oxidative markers and lower antioxidant concentrations in identifying the disease. Indicators of oxidation, such as advanced glycation end products, protein carbonyls, oxidized lipoproteins, and antioxidant deficiencies, have been associated with frailty,18,73–75 and a number of its components, such as sarcopenia76,77 and decreased gait speed.78–80
While these markers have shown promise in utilization, they all suffer similar flaws and challenges on the path to clinical use. The major challenge associated with their use is that the changes associated with frailty are also largely associated with the normal aging process. This blurs the line between a normal and abnormal result for a patient of a given age, making clear cutoff points to indicate the onset or progression of disease challenging to determine. They are also associated with very common comorbidities, leading to a wide range of confounding variables. Additionally, their individual associations with frailty are also typically weak, likely reflecting the complex multisystem pathophysiology underpinning the condition. While this has prompted many to include a diagnostic panel of markers with stronger associations,10,81 more research is required to explore the utility of this approach fully. These flaws continue to drive research into better methods of predicting and diagnosing frailty, with a growing body of research moving into new fields of biological research.
Circulating osteoprogenitor cells: new biomarkers for frailty
Mesenchymal stem cells (MSCs) are providing new avenues for cell-based therapy in the renewal of damaged or diseased tissue, due to their capacity to maintain multipotency into various cell types and proliferate broadly in vitro.82,83 Although resident MSCs, which are found in almost all tissues when they undergo a repair process and renewal, are well documented and intensively studied,84 the role of circulating MSCs is still yet to be explored. Since circulating MSCs were initially identified by Friedenstein et al 40 years ago,85 very few researchers have brought to attention and characterized these cells in terms of their biological properties, adherence, and osteogenic potential.86,87 Resident MSCs obtained from bone marrow by aspiration are typically low in number and lack superiority, eg, studies have shown that the quality of MSCs obtained from the spine and iliac crest present with reduced capacity of differentiation and function when compared to MSCs obtained from other regions, such as the femur.88,89 This has been the rationale behind several studies that have isolated MSCs from various tissue types, such as umbilical cord blood,90 adipose tissue,91 peripheral blood,92 liver, and amniotic fluid.93 Although these MSCs isolated from differing tissues share some common identification markers, their differentiation abilities and gene-expression profiles vary greatly.
In the past decade, a group of stem-cell-like populations has been found in the circulation, known as circulating osteoprogenitor (COP) cells,94 which are considered surrogates of the marrow stem-cell population. These MSC-like cells have been studied extensively in ex vivo systems and can be isolated from the blood mononuclear cell population through their plastic adherence capacity and differentiation potential.94–96 Systemic infusion studies have shown that MSCs have limited capacity to cross the endothelial barrier, with poor migration to uninjured skeletal tissue. This is in contrast to COP cells,97 which are fluid-phase, allowing them to cross the endothelial barrier.98
There are several types of COP cells, based on their adherence in culture and their differentiation profile,94,99 thus suggesting that these cells represent different developmental stages from hematopoietic/stromal cells in bone marrow. This reasoning has allowed COP cells to become a potentially feasible alternative to other sources of stem cells and potentially the first step in the development of new biomarkers for musculoskeletal disorders. Due to their strong mesenchymal potential, quantification of this population of COP cells should ensure a biological connection between this cell population and the most common biological and/or physical hallmarks of frailty.
Little was known about the normal reference range for COP cells in healthy conditions. To identify this reference range, we completed the COP Study.100 This was a cross-sectional study undertaken in 144 healthy volunteers in Western Sydney (20- to 90-year-olds, ten male and ten female subjects per decade). In this healthy population, we found an average percentage of COP cells (%COP) of 0.42 of the peripheral blood mononuclear cell population. In addition, we identified an age- and sex-independent reference range of %COP of 0.1%–3.8%. These results suggested that %COP may be compared to other components of the blood count (eg, neutrophils), which remain stable under healthy conditions, but are affected by disease, thus indicating changes in the bone marrow in response to stressors. For example, high %COP cells have been associated with calcific aortic stenosis101 and nonhereditary heterotopic calcification.102 In these cases, very high %COP in the circulation was associated with excessive endothelial and cartilage growth and calcification.
However, understanding the biological impact of low levels of COP cells remained unknown. Therefore, in the Nepean Osteoporosis and Frailty (NOF) study,103 we tested the hypothesis that low %COP levels were associated with frailty and disability. In this study, a random sample of community-dwelling and residential aged-care volunteers aged 65 years and older (mean age 82.8 years; n=77; 70% female; 27 robust, 23 prefrail, and 27 frail) was recruited, with frailty status determined using the Fried et al and Rockwood et al models. We found lower %COP to be associated with frailty, disability, and poor physical performance in participants with %COP in the lower quartile more likely to be frail (OR 2.65, 95% CI 2.72–3.15). Interestingly, when compared with serum IL6 – one of the most widely used biomarkers for frailty8 – %COP showed a stronger correlation with the clinical indicators of frailty (Fried et al’s and Rockwood et al’s) and with physical performance and disability. In addition, preliminary analysis from NOF showed sensitivity of 93% and specificity of 89% of %COP for the diagnosis of frailty.
Lamin A and COP cells: a robust biomarker for frailty
The nuclear lamina is an assemblage of intermediate filament proteins that lies between the inner nuclear membrane and the nucleoplasm. Given its location, it serves many functions, including the regulation of gene expression, mobilizing a repressive/active chromatin environment, communication with the cytoskeleton by nuclear import and export, and sequestration of transcriptional factors across the nucleus.104–107 The lamina consists of a network of proteins called lamins, with three major lamins identified – A, B, and C, with lamin A/C involved in cell differentation.
In aging MSCs, the nuclear lamina is believed to play an important role, assisting in their differentiation into myocytes and osteoblasts, while also regulating osteoblast function and survival.108 Lamin A is an intermediate filament of the lamina and plays a vital role in cell survival, replication, differentiation, and structural support to the nucleus.105 Interestingly, patients deficient in lamin A or with mutations in LMNA suffer from a group of diseases known as laminopathies, which also include Hutchinson–Gilford progeria syndrome,109 all of which affect bone and muscle.104 Furthermore, lamin A deficiency affects osteoblast differentiation and induces skeletal muscle abnormalities. Our team has shown that lamin A knockdown in MSCs inhibits osteoblastogenesis and decreases mineralization in vitro.110 In addition, low levels of lamin A were associated with high levels of fat infiltration of muscle and bone in a mouse model of progeria.111
Overall, lamin A plays an important role in MSC and musculoskeletal disease. As such, there is a potential impact on the aging process, in particular the progression from healthy aging to frailty and disability. Based on this current knowledge, we assessed lamin A expression in COP cells and hypothesized that lamin A COP had a strong association with frailty. As part of the COP Study, we initially quantified the normal reference range of lamin A COP for a healthy population, finding that, as with %COP, it remains stable in healthy aging individuals.103 We then analyzed the samples obtained from participants in the NOF Study, assessing lamin A COP in our three subpopulations.112 As an additional control, we used buccal swabs of epithelial cells to determine whether potential changes in lamin A expression were exclusive of COP cells as cells of mesenchymal origin.
Lamin A expression in buccal cells showed steady levels at all ages and no association with either disability or frailty. In contrast, low lamin A COP was strongly associated with frailty. Frail individuals showed 60% lower lamin A COP compared to robust subjects (95% CI −36 to −74%, P<0.001) and 62% lower levels compared to prefrail (95% CI −40 to −76%, P<0.001). The association between low lamin A COP and frailty was stronger than %COP alone. Despite this promising evidence supporting the use of %COP and lamin A-COP as biomarkers for frailty. further longitudinal studies assessing their sensitivity, specificity, and predictive value are still required.
Biomarkers of frailty: future prospects
With the advancement of scientific technology, analytical methods once too difficult and expensive to apply to many issues facing global medicine have been simplified and are increasingly being applied to aging and frailty. Currently, research on biomarkers indicating frailty is moving into the realm of genetic and proteomic research in the hunt for a unifying system of prediction and diagnosis of the condition.
Changes in the proteome of patients have been hypothesized as a unifying pathophysiological process underpinning accelerated progression into frailty. Researchers have begun to identify strong proteomic indicators of frailty in a range of targets, including angiotensin, antithrombin, haptoglobin, transferrin, and fibrinogen, all of which correlate with hallmark symptoms of the syndrome.113,114 Others have ventured deeper still, attempting to identify a genetic cause for the aberrations in protein expression. Genetic dysregulation provides a comprehensive explanation for the phenotypic changes seen in the development of frailty. However, identifying the specific genes involved remains a daunting task. It has been suggested that exploration of the genetic regulation of inflammation, metabolism, calcium homeostasis, cellular apoptosis, and neuromuscular interaction should be examined to aid in explaining the typical changes seen in frailty.115 This wide-spanning approach to identification of suitable genetic targets has been mirrored in the European FRAILOMIC initiative, in which a vast range of genetic and proteomic markers are to be examined and matched to the more classical serum and clinical biomarkers, in order to find the best correlations and predictors of onset and transitions from robust to disabled.116 In a similar manner, others are investigating circulating miRNA targets, with the hypothesis that they may drive the changes in protein and gene expression. miRNA has been suggested as the biological mechanism behind midlife physical decline117 and this has driven exploration into the area, with researchers finding a number of targets seemingly linked to the development of frailty.118,119
Conclusion
Frailty presents an additional burden on older adults, with greater levels of weakness and inactivity observed when compared to age-matched counterparts, even when matched for comorbidities, demographics, sex, and age.6 The development and validation of clinical criteria have aided the diagnosis of frailty in research and clinical settings. These diagnostic tools, although clinically plausible, are not strictly associated with the biological changes of frailty, and may fail to distinguish between individuals who are frail or prefrail from normal elderly people. Therefore, a comprehensive diagnostic tool or test for frailty with biological elucidations is still missing.120
As life expectancies increase worldwide, the incidence and clinical consequences of frailty will increase rapidly. Therefore, early diagnosis of frailty is pivotal to prevent disability and adverse outcomes. Given the lack of robust biomarkers for frailty, there is a growing necessity to undertake research aimed at identifying novel, practical, and reliable assessments. This will thus reduce the burden on the healthcare system through fewer and less intensive services and provide the opportunity for older persons to stay healthy and independent in the community for a longer period.
Disclosure
The authors report no conflicts of interest in this work.
References
Bergman H, Ferrucci L, Guralnik J, et al. Frailty: an emerging research and clinical paradigm – issues and controversies. J Gerontol A Biol Sci Med Sci. 2007;62(7):731–737. | ||
Fried LP, Tangen CM, Walston J, et al; Cardiovascular Health Study Collaborative Research Group. Frailty in older adults: evidence for a phenotype. J Gerontol A Biol Sci Med Sci. 2001;56(3):M146–M157. | ||
Walston J, Hadley EC, Ferrucci L, et al. Research agenda for frailty in older adults: toward a better understanding of physiology and etiology: summary from the American geriatrics Society/National Institute on Aging Research conference on frailty in older adults. J Am Geriatr Soc. 2006;54(6):991–1001. | ||
Pritchard JM, Kennedy CC, Karampatos S, et al. Measuring frailty in clinical practice: a comparison of physical frailty assessment methods in a geriatric out-patient clinic. BMC Geriatr. 2017;17(1):264. | ||
Walston J, Buta B, Xue QL. Frailty screening and interventions: considerations for clinical practice. Clin Geriatr Med. 2018;34(1):25–38. | ||
Rockwood K, Song X, MacKnight C, et al. A global clinical measure of fitness and frailty in elderly people. CMAJ. 2005;173(5):489–495. | ||
Gill TM, Gahbauer EA, Allore HG, Han L. Transitions between frailty states among community-living older persons. Arch Intern Med. 2006;166(4):418–423. | ||
Calvani R, Marini F, Cesari M, et al. Biomarkers for physical frailty and sarcopenia: state of the science and future developments. J Cachexia Sarcopenia Muscle. 2015;6(4):278–286. | ||
Cannizzo ES, Clement CC, Sahu R, Follo C, Santambrogio L. Oxidative stress, inflamm-aging and immunosenescence. J Proteomics. 2011;74(11):2313–2323. | ||
Mitnitski A, Collerton J, Martin-Ruiz C, et al. Age-related frailty and its association with biological markers of ageing. BMC Med. 2015;13(1):161. | ||
Franceschi C, Garagnani P, Parini P, Giuliani C, Santoro A. Inflammaging: a new immune-metabolic viewpoint for age-related diseases. Nat Rev Endocrinol. 2018;14(10):576–590. | ||
Leng SX, Xue QL, Tian J, Walston JD, Fried LP. Inflammation and frailty in older women. J Am Geriatr Soc. 2007;55(6):864–871. | ||
Leng S, Chaves P, Koenig K, Walston J. Serum interleukin-6 and hemoglobin as physiological correlates in the geriatric syndrome of frailty: a pilot study. J Am Geriatr Soc. 2002;50(7):1268–1271. | ||
Boxer RS, Dauser DA, Walsh SJ, Hager WD, Kenny AM. The association between vitamin D and inflammation with the 6-minute walk and frailty in patients with heart failure. J Am Geriatr Soc. 2008;56(3):454–461. | ||
Soysal P, Stubbs B, Lucato P, et al. Inflammation and frailty in the elderly: a systematic review and meta-analysis. Ageing Res Rev. 2016;31:1–8. | ||
Leng SX, Cappola AR, Andersen RE, et al. Serum levels of insulin-like growth factor-I (IGF-I) and dehydroepiandrosterone sulfate (DHEA-S), and their relationships with serum interleukin-6, in the geriatric syndrome of frailty. Aging Clin Exp Res. 2004;16(2):153–157. | ||
Darvin K, Randolph A, Ovalles S, et al. Plasma protein biomarkers of the geriatric syndrome of frailty. J Gerontol A Biol Sci Med Sci. 2014;69(2):182–186. | ||
Liu CK, Lyass A, Larson MG, et al. Biomarkers of oxidative stress are associated with frailty: the Framingham offspring study. Age (Dordr). 2016;38(1):1. | ||
Hubbard RE, O’Mahony MS, Savva GM, Calver BL, Woodhouse KW. Inflammation and frailty measures in older people. J Cell Mol Med. 2009;13(9B):3103–3109. | ||
Visser M, Pahor M, Taaffe DR, et al. Relationship of interleukin-6 and tumor necrosis factor-alpha with muscle mass and muscle strength in elderly men and women: the Health ABC study. J Gerontol A Biol Sci Med Sci. 2002;57(5):M326–M332. | ||
Bautmans I, Gorus E, Njemini R, Mets T. Handgrip performance in relation to self-perceived fatigue, physical functioning and circulating IL-6 in elderly persons without inflammation. BMC Geriatr. 2007;7(1):5. | ||
Kilgour AH, Firth C, Harrison R, et al. Seropositivity for CMV and IL-6 levels are associated with grip strength and muscle size in the elderly. Immun Ageing. 2013;10(1):33. | ||
Ding C, Parameswaran V, Udayan R, Burgess J, Jones G. Circulating levels of inflammatory markers predict change in bone mineral density and resorption in older adults: a longitudinal study. J Clin Endocrinol Metab. 2008;93(5):1952–1958. | ||
Maes M, Bosmans E, De Jongh R, Kenis G, Vandoolaeghe E, Neels H. Increased serum IL-6 and IL-1 receptor antagonist concentrations in major depression and treatment resistant depression. Cytokine. 1997;9(11):853–858. | ||
Barzilay JI, Blaum C, Moore T, et al. Insulin resistance and inflammation as precursors of frailty: the Cardiovascular Health study. Arch Intern Med. 2007;167(7):635–641. | ||
Carcaillon L, García-García FJ, Tresguerres JA, Gutiérrez Avila G, Kireev R, Rodríguez-Mañas L. Higher levels of endogenous estradiol are associated with frailty in postmenopausal women from the toledo study for healthy aging. J Clin Endocrinol Metab. 2012;97(8):2898–2906. | ||
Fontana L, Addante F, Copetti M, et al. Identification of a metabolic signature for multidimensional impairment and mortality risk in hospitalized older patients. Aging Cell. 2013;12(3):459–466. | ||
Saum KU, Dieffenbach AK, Jansen EH, et al. Association between oxidative stress and frailty in an elderly German population: results from the ESTHER cohort study. Gerontology. 2015;61(5):407–415. | ||
Koh JM, Khang YH, Jung CH, et al. Higher circulating hsCRP levels are associated with lower bone mineral density in healthy pre- and postmenopausal women: evidence for a link between systemic inflammation and osteoporosis. Osteoporos Int. 2005;16(10):1263–1271. | ||
Baylis D, Bartlett DB, Syddall HE, et al. Immune-endocrine biomarkers as predictors of frailty and mortality: a 10-year longitudinal study in community-dwelling older people. Age. 2013;35(3):963–971. | ||
Puts MT, Visser M, Twisk JW, Deeg DJ, Lips P. Endocrine and inflammatory markers as predictors of frailty. Clin Endocrinol (Oxf). 2005;63(4):403–411. | ||
Reiner AP, Aragaki AK, Gray SL, et al. Inflammation and thrombosis biomarkers and incident frailty in postmenopausal women. Am J Med. 2009;122(10):947–954. | ||
Ferrucci L, Corsi A, Lauretani F, et al. The origins of age-related proinflammatory state. Blood. 2005;105(6):2294–2299. | ||
Hyde Z, Flicker L, Almeida OP, et al. Low free testosterone predicts frailty in older men: the Health in Men Study. J Clin Endocrinol Metab. 2010;95(7):3165–3172. | ||
Wu IC, Lin XZ, Liu PF, Tsai WL, Shiesh SC. Low serum testosterone and frailty in older men and women. Maturitas. 2010;67(4):348–352. | ||
Carcaillon L, Blanco C, Alonso-Bouzón C, Alfaro-Acha A, Garcia-García FJ, Rodriguez-Mañas L. Sex differences in the association between serum levels of testosterone and frailty in an elderly population: the Toledo study for healthy aging. PLoS One. 2012;7(3):e32401. | ||
Srinivas-Shankar U, Roberts SA, Connolly MJ, et al. Effects of testosterone on muscle strength, physical function, body composition, and quality of life in intermediate-frail and frail elderly men: a randomized, double-blind, placebo-controlled study. J Clin Endocrinol Metab. 2010;95(2):639–650. | ||
Kenny AM, Kleppinger A, Annis K, et al. Effects of transdermal testosterone on bone and muscle in older men with low bioavailable testosterone levels, low bone mass, and physical frailty. J Am Geriatr Soc. 2010;58(6):1134–1143. | ||
Storer TW, Woodhouse L, Magliano L, et al. Changes in muscle mass, muscle strength, and power but not physical function are related to testosterone dose in healthy older men. J Am Geriatr Soc. 2008;56(11):1991–1999. | ||
Mohr BA, Bhasin S, Kupelian V, Araujo AB, O’Donnell AB, McKinlay JB. Testosterone, sex hormone-binding globulin, and frailty in older men. J Am Geriatr Soc. 2007;55(4):548–555. | ||
Goldman N, Glei DA. Sex differences in the relationship between DHEAS and health. Exp Gerontol. 2007;42(10):979–987. | ||
Voznesensky M, Walsh S, Dauser D, Brindisi J, Kenny AM. The association between dehydroepiandosterone and frailty in older men and women. Age Ageing. 2009;38(4):401–406. | ||
Muller M, van den Beld AW, van der Schouw YT, Grobbee DE, Lamberts SW. Effects of dehydroepiandrosterone and atamestane supplementation on frailty in elderly men. J Clin Endocrinol Metab. 2006;91(10):3988–3991. | ||
Kenny AM, Boxer RS, Kleppinger A, Brindisi J, Feinn R, Burleson JA. Dehydroepiandrosterone combined with exercise improves muscle strength and physical function in frail older women. J Am Geriatr Soc. 2010;58(9):1707–1714. | ||
Cappola AR, Xue QL, Fried LP. Multiple hormonal deficiencies in anabolic hormones are found in frail older women: the women’s health and aging studies. J Gerontol A Biol Sci Med Sci. 2009;64A(2):243–248. | ||
Suriyaarachchi P, Gomez F, Curcio CL, et al. High parathyroid hormone levels are associated with osteosarcopenia in older individuals with a history of falling. Maturitas. 2018;113:21–25. | ||
Murthy L, Dreyer P, Suriyaarachchi P, et al. Association between high levels of parathyroid hormone and frailty: the Nepean osteoporosis and frailty (nof) study. J Frailty Aging. 2018;7(4):253–257. | ||
Tajar A, Lee DM, Pye SR, et al. The association of frailty with serum 25-hydroxyvitamin D and parathyroid hormone levels in older European men. Age Ageing. 2013;42(3):352–359. | ||
Swiecicka A, Lunt M, Ahern T, et al. Nonandrogenic anabolic hormones predict risk of frailty: European male Ageing Study prospective data. J Clin Endocrinol Metab. 2017;102(8):2798–2806. | ||
Liu LK, Lee WJ, Chen LY, et al. Association between frailty, osteoporosis, falls and hip fractures among community-dwelling people aged 50 years and older in Taiwan: results from I-Lan longitudinal aging study. PLoS One. 2015;10(9):e0136968. | ||
Sambrook PN, Chen JS, March LM, et al. Serum parathyroid hormone predicts time to fall independent of vitamin D status in a frail elderly population. J Clin Endocrinol Metab. 2004;89(4):1572–1576. | ||
Björkman M, Sorva A, Tilvis R. Parathyroid hormone as a mortality predictor in frail aged inpatients. Gerontology. 2009;55(6):601–606. | ||
Ensrud KE, Blackwell TL, Cauley JA, et al; Osteoporotic Fractures in Men Study Group. Circulating 25-hydroxyvitamin D levels and frailty in older men: the osteoporotic fractures in Men Study. J Am Geriatr Soc. 2011;59(1):101–106. | ||
Hirani V, Naganathan V, Cumming RG, et al. Associations between frailty and serum 25-hydroxyvitamin D and 1,25-dihydroxyvitamin D concentrations in older Australian men: the Concord Health and ageing in Men Project. J Gerontol A Biol Sci Med Sci. 2013;68(9):1112–1121. | ||
Alvarez-Ríos AI, Guerrero JM, García-García FJ, et al. Associations between frailty and serum N-terminal propeptide of type I procollagen and 25-hydroxyvitamin D in older Spanish women: the Toledo study for healthy aging. Exp Gerontol. 2015;69:79–84. | ||
Vogt S, Decke S, de Las Heras Gala T, et al. Prospective association of vitamin D with frailty status and all-cause mortality in older adults: results from the KORA-Age study. Prev Med. 2015;73:40–46. | ||
Shardell M, D’Adamo C, Alley DE, et al. Serum 25-hydroxyvitamin D, transitions between frailty states, and mortality in older adults: the Invecchiare in Chianti study. J Am Geriatr Soc. 2012;60(2):256–264. | ||
Mohamad MI, Khater MS. Evaluation of insulin like growth factor-1 (IGF-1) level and its impact on muscle and bone mineral density in frail elderly male. Arch Gerontol Geriatr. 2015;60(1):124–127. | ||
Onder G, Liperoti R, Russo A, et al. Body mass index, free insulin-like growth factor I, and physical function among older adults: results from the ilSIRENTE study. Am J Physiol Endocrinol Metab. 2006;291(4):E829–E834. | ||
Heemskerk VH, Daemen MA, Buurman WA. Insulin-like growth factor-1 (IGF-1) and growth hormone (GH) in immunity and inflammation. Cytokine Growth Factor Rev. 1999;10(1):5–14. | ||
Blaum CS, Xue QL, Tian J, Semba RD, Fried LP, Walston J. Is hyperglycemia associated with frailty status in older women? J Am Geriatr Soc. 2009;57(5):840–847. | ||
Kalyani RR, Tian J, Xue QL, et al. Hyperglycemia and incidence of frailty and lower extremity mobility limitations in older women. J Am Geriatr Soc. 2012;60(9):1701–1707. | ||
García-Esquinas E, Graciani A, Guallar-Castillón P, López-García E, Rodríguez-Mañas L, Rodríguez-Artalejo F. Diabetes and risk of frailty and its potential mechanisms: a prospective cohort study of older adults. J Am Med Dir Assoc. 2015;16(9):748–754. | ||
Abdelhafiz AH, Rodríguez-Mañas L, Morley JE, Sinclair AJ. Hypoglycemia in older people – a less well recognized risk factor for frailty. Aging Dis. 2015;6(2):156. | ||
Nelson JM, Dufraux K, Cook PF. The relationship between glycemic control and falls in older adults. J Am Geriatr Soc. 2007;55(12):2041–2044. | ||
Khandelwal D, Goel A, Kumar U, Gulati V, Narang R, Dey AB. Frailty is associated with longer hospital stay and increased mortality in hospitalized older patients. J Nutr Health Aging. 2012;16(8):732–735. | ||
Ramsay SE, Arianayagam DS, Whincup PH, et al. Cardiovascular risk profile and frailty in a population-based study of older British men. Heart. 2015;101(8):616–622. | ||
Sanchis J, Núñez E, Ruiz V, et al. Usefulness of clinical data and biomarkers for the identification of frailty after acute coronary syndromes. Can J Cardiol. 2015;31(12):1462–1468. | ||
Palmer K, Vetrano DL, Marengoni A, et al. The relationship between anaemia and frailty: a systematic review and meta-analysis of observational studies. J Nutr Health Aging. 2018;22(8):965–974. | ||
Vanitallie TB. Frailty in the elderly: contributions of sarcopenia and visceral protein depletion. Metabolism. 2003;52(10 Suppl 2):22–26. | ||
Hazzard WR. Depressed albumin and high-density lipoprotein cholesterol: signposts along the final common pathway of frailty. J Am Geriatr Soc. 2001;49(9):1253–1254. | ||
Dalrymple LS, Katz R, Rifkin DE, et al. Kidney function and prevalent and incident frailty. Clin J Am Soc Nephrol. 2013;8(12):2091–2099. | ||
Inglés M, Gambini J, Carnicero JA, et al. Oxidative stress is related to frailty, not to age or sex, in a geriatric population: lipid and protein oxidation as biomarkers of frailty. J Am Geriatr Soc. 2014;62(7):1324–1328. | ||
Soysal P, Isik AT, Carvalho AF, et al. Oxidative stress and frailty: a systematic review and synthesis of the best evidence. Maturitas. 2017;99:66–72. | ||
Saum KU, Dieffenbach AK, Jansen EH, et al. Association between oxidative stress and frailty in an elderly German population: results from the ESTHER cohort study. Gerontology. 2015;61(5):407–415. | ||
Dalal M, Ferrucci L, Sun K, Beck J, Fried LP, Semba RD. Elevated serum advanced glycation end products and poor grip strength in older community-dwelling women. J Gerontol A Biol Sci Med Sci. 2009;64A(1):132–137. | ||
Semba RD, Blaum C, Guralnik JM, Moncrief DT, Ricks MO, Fried LP. Carotenoid and vitamin E status are associated with indicators of sarcopenia among older women living in the community. Aging Clin Exp Res. 2003;15(6):482–487. | ||
Semba RD, Ferrucci L, Sun K, et al. Oxidative stress and severe walking disability among older women. Am J Med. 2007;120(12):1084–1089. | ||
Cesari M, Kritchevsky SB, Nicklas BJ, et al. Lipoprotein peroxidation and mobility limitation: results from the health, aging, and body composition study. Arch Intern Med. 2005;165(18):2148–2154. | ||
Alipanah N, Varadhan R, Sun K, Ferrucci L, Fried LP, Semba RD. Low serum carotenoids are associated with a decline in walking speed in older women. J Nutr Health Aging. 2009;13(3):170–175. | ||
Lara J, Cooper R, Nissan J, et al. A proposed panel of biomarkers of healthy Ageing. BMC Med. 2015;13(1):222. | ||
Ankrum JA, Ong JF, Karp JM. Mesenchymal stem cells: immune evasive, not immune privileged. Nat Biotechnol. 2014;32(3):252–260. | ||
Bianco P. “Mesenchymal” stem cells. Annu Rev Cell Dev Biol. 2014;30:677–704. | ||
Sreenivas SD, Rao AS, Satyavani SS, Reddy BH, Vasudevan S, Vasudevan S, Sreenivas A. Where will the stem cells lead us? prospects for dentistry in the 21 century. 2011;15(3):199–204. | ||
Friedenstein AJ, Chailakhjan RK, Lalykina KS. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Proliferation. 1970;3(4):393–403. | ||
Suda RK, Billings PC, Egan KP, et al. Circulating osteogenic precursor cells in heterotopic bone formation. Stem Cells. 2009;27(9):2209–2219. | ||
Eghbali-Fatourechi GZ, Lamsam J, Fraser D, Nagel D, Riggs BL, Khosla S. Circulating osteoblast-lineage cells in humans. N Engl J Med. 2005;352(19):1959–1966. | ||
Clines GA. Prospects for osteoprogenitor stem cells in fracture repair and osteoporosis. Curr Opin Organ Transplant. 2010;15(1):73–78. | ||
Mohanty ST, Bellantuono I. Intra-femoral injection of human mesenchymal stem cells. Methods Mol Biol. 2013;976:131–141. | ||
Rosada C, Justesen J, Melsvik D, Ebbesen P, Kassem M. The human umbilical cord blood: a potential source for osteoblast progenitor cells. Calcif Tissue Int. 2003;72(2):135–142. | ||
Zuk PA, Zhu M, Mizuno H, et al. Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue Eng. 2001;7(2):211–228. | ||
Kuznetsov SA, Mankani MH, Gronthos S, Satomura K, Bianco P, Robey PG. Circulating skeletal stem cells. J Cell Biol. 2001;153(5):1133–1140. | ||
Campagnoli C, Roberts IA, Kumar S, Bennett PR, Bellantuono I, Fisk NM. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood. 2001;98(8):2396–2402. | ||
Feehan J, Nurgali K, Apostolopoulos V, Al Saedi A, Duque G. Circulating osteogenic precursor cells: building bone from blood. EBioMedicine. 2019;39:603–611. | ||
Gronthos S, Graves SE, Ohta S, Simmons PJ. The STRO-1+ fraction of adult human bone marrow contains the osteogenic precursors. Blood. 1994;84(12):4164–4173. | ||
Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for cellular therapy position statement. Cytotherapy. 2006;8(4):315–317. | ||
George AL, Boulanger CA, Anderson LH, Cagnet S, Brisken C, Smith GH. In vivo reprogramming of non-mammary cells to an epithelial cell fate is independent of amphiregulin signaling. J Cell Sci. 2017;130(12):2018–2025. | ||
Tang PX. [Adjunctive reflections on history of hematopoietic stem cell study – editorial]. Zhongguo. 2005;13(5):723–732. Chinese. | ||
Rohban R, Pieber TR. Mesenchymal stem and progenitor cells in regeneration: tissue specificity and regenerative potential. Stem Cells Int. 2017;2017:5173732. | ||
Gunawardene P, Al Saedi A, Singh L, et al. Age, gender, and percentage of circulating osteoprogenitor (COP) cells: the cop study. Exp Gerontol. 2017;96:68–72. | ||
Urbich C, Dimmeler S. Endothelial progenitor cells: characterization and role in vascular biology. Circ Res. 2004;95(4):343–353. | ||
Egan KP, Duque G, Keenan MA, Pignolo RJ. Circulating osteogentic precursor cells in non-hereditary heterotopic ossification. Bone. 2018;109:61–64. | ||
Gunawardene P, Bermeo S, Vidal C, et al. Association between circulating osteogenic progenitor cells and disability and frailty in older persons: the Nepean osteoporosis and frailty study. J Gerontol A Biol Sci Med Sci. 2016;71(9):1124–30. | ||
Gerace L, Huber MD. Nuclear lamina at the crossroads of the cytoplasm and nucleus. J Cachexia Sarcopenia Muscle. 2012;177(1):24–31. | ||
Worman HJ, Bonne G. “Laminopathies”: a wide spectrum of human diseases. Exp Cell Res. 2007;313(10):2121–2133. | ||
Prokocimer M, Davidovich M, Nissim-Rafinia M, et al. Nuclear lamins: key regulators of nuclear structure and activities. J Cell Mol Med. 2009;13(6):1059–1085. | ||
Goldberg M, Harel A, Gruenbaum Y. The nuclear lamina: molecular organization and interaction with chromatin. Crit Rev Eukar Gene Expr. 1999;9(3–4):285–293. | ||
Vidal C, Bermeo S, Fatkin D, Duque G. Role of the nuclear envelope in the pathogenesis of age-related bone loss and osteoporosis. Bonekey Rep. 2012;1(5):62. | ||
Dutta S, Bhattacharyya M, Sengupta K. Implications and assessment of the elastic behavior of lamins in laminopathies. Cells. 2016;5(4):37. | ||
Akter R, Rivas D, Geneau G, Drissi H, Duque G. Effect of lamin A/C knockdown on osteoblast differentiation and function. J Bone Miner Res. 2009;24(2):283–293. | ||
Tong J, Li W, Vidal C, Yeo LS, Fatkin D, Duque G. Lamin A/C deficiency is associated with fat infiltration of muscle and bone. Mech Ageing Dev. 2011;132(11–12):552–559. | ||
Al Saedi A, Gunawardene P, Bermeo S, et al. Lamin A expression in circulating osteoprogenitors as a potential biomarker for frailty: the Nepean osteoporosis and frailty (nof) study. Exp Gerontol. 2018;102:69–75. | ||
Shamsi KS, Pierce A, Ashton AS, Halade DG, Richardson A, Espinoza SE. Proteomic screening of glycoproteins in human plasma for frailty biomarkers. J Gerontol A Biol Sci Med Sci. 2012;67(8):853–864. | ||
Lin CH, Liao CC, Huang CH, et al. Proteomics analysis to identify and characterize the biomarkers and physical activities of Non-Frail and frail older adults. Int J Med Sci. 2017;14(3):231–239. | ||
Cardoso AL, Fernandes A, Aguilar-Pimentel JA, et al. Towards frailty biomarkers: candidates from genes and pathways regulated in aging and age-related diseases. Ageing. 2018;47:214–277. | ||
Erusalimsky JD, Grillari J, Grune T, et al; FRAILOMIC Consortium. In Search of ‘Omics’-Based Biomarkers to Predict Risk of Frailty and Its Consequences in Older Individuals: The FRAILOMIC Initiative. Gerontology. 2016;62(2):182–190. | ||
Wang E, Microrna WE. MicroRNA, the putative molecular control for mid-life decline. Ageing Res Rev. 2007;6(1):1–11. | ||
Ipson BR, Fletcher MB, Espinoza SE, Fisher AL. Identifying Exosome-Derived microRNAs as candidate biomarkers of frailty. J Frailty Aging. 2018;7(2):100–103. | ||
Rusanova I, Diaz-Casado ME, Fernández-Ortiz M, et al. Analysis of plasma microRNAs as predictors and biomarkers of aging and frailty in humans. Oxid Med Cell Longev. 2018;2018:7671850. | ||
Walston J, Buta B, Xue QL. Frailty screening and interventions: considerations for clinical practice. Clin Geriatr Med. 2018;34(1):25–38. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.