Back to Journals » International Journal of Nanomedicine » Volume 20
Cross - Kingdom Dialogue of Microbial Messengers: Multi - Target Regulatory Mechanisms and Therapeutic Strategies of Gut Microbiota - Derived Extracellular Vesicles in Metabolic Diseases
Authors Huang J, Yu Y, Feng Z ![]() , Yin Y, Liu Y, Liu X, Yu R
, Yin Y, Liu Y, Liu X, Yu R ![]()
Received 20 June 2025
Accepted for publication 5 October 2025
Published 15 October 2025 Volume 2025:20 Pages 12573—12591
DOI https://doi.org/10.2147/IJN.S548624
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Xing Zhang
Jiawang Huang,* Yunfeng Yu,* Zhiying Feng, Yuman Yin, Yi Liu, Xiu Liu, Rong Yu
College of Traditional Chinese Medicine, Hunan University of Chinese Medicine, Changsha, Hunan, 410208, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Rong Yu, College of Traditional Chinese Medicine, Hunan University of Chinese Medicine, Xueshi Road 300, Changsha, Hunan, 410208, People’s Republic of China, Email [email protected] Xiu Liu, College of Traditional Chinese Medicine, Hunan University of Chinese Medicine, Xueshi Road 300, Changsha, Hunan, 410208, People’s Republic of China, Email [email protected]
Abstract: Gut microbiota-derived extracellular vesicles (GMEVs) serve as pivotal mediators of cross-communication between microorganisms and their hosts, playing a complex and central role in the onset and progression of metabolic diseases. This review systematically summarizes the biological characteristics, molecular composition, and regulatory mechanisms of GMEVs in metabolic disorders such as obesity, diabetes, and non-alcoholic fatty liver disease. GMEVs facilitate dialogue along the “gut-organ axis” by transporting active molecules such as lipopolysaccharides (LPS), flagellin, nucleic acids, and metabolic products, thereby mediating bidirectional regulation of host metabolic homeostasis. GMEVs derived from pathogenic bacteria promote metabolic inflammation and insulin resistance by compromising the intestinal barrier, activating inflammatory pathways, and inhibiting insulin signaling. Conversely, GMEVs originating from probiotics exert protective effects by enhancing tight junction functionality, modulating immune responses, and improving lipid metabolism. Notably, the dynamic fluctuations of GMEVs are closely linked to disease progression, and their microbiota-specific molecular characteristics offer novel biomarkers for the early diagnosis of metabolic diseases. Despite the challenges posed by standardization of isolation and elucidation of underlying mechanisms, the potential applications of GMEVs in the precise prevention and treatment of metabolic diseases are vast, providing a theoretical foundation for the development of novel microbiome-based intervention strategies. Future research should further elucidate the heterogeneity of GMEVs and their spatiotemporal dynamics in host interactions, facilitating the transition from fundamental research to clinical application.
Keywords: gut microbiota-derived extracellular vesicles, metabolic diseases, gut-organ axis, microbial-host interactions, precision therapy
Introduction
The gut serves as the core site for digestion and absorption within the human body, functioning as a highly intricate microbial ecosystem.1 Recent studies increasingly indicate that the gut microbiota not only maintains local homeostasis within the intestinal environment but also influences the physiological functions and disease progression of distant organs through regulatory mechanisms of the “gut-organ axis”.2 As a crucial regulator of internal environmental stability, the interplay between gut microbiota and host metabolism has emerged as a focal point in metabolic disease research. Existing evidence suggests that dysbiosis of the gut microbiota may contribute to the onset and progression of metabolic diseases such as obesity, diabetes, and its complications, as well as metabolic associated fatty liver disease (MAFLD), through mechanisms that regulate host insulin resistance, low-grade inflammatory responses, and disturbances in lipid metabolism.3,4 Given its extensive regulatory functions, the gut microbiota is regarded as a unique endocrine dynamic ecosystem, participating in the maintenance of host energy metabolic homeostasis through molecular crosstalk between microorganisms and the host.5 As research into the “gut-organ axis” deepens, the mechanisms by which gut microbiota and their metabolic products influence metabolic diseases are gradually being elucidated. Studies indicate that the gut microbiota may participate in the pathological processes of metabolic diseases by affecting intestinal barrier function, immune regulation, and the secretion of metabolites.6 However, the molecular mechanisms by which the gut microbiota “remotely controls” metabolic diseases such as obesity and diabetes remain unclear, particularly regarding the pathways of information transfer between the gut microbiota and distant tissues. In this context, gut microbiota-derived extracellular vesicles (GMEVs) have garnered significant attention as novel mediators of microbial-host interactions.
GMEVs are membrane-bound vesicles, approximately 30–300 nm in diameter, actively secreted by the gut microbiota, capable of stably carrying and delivering a wealth of bioactive molecules, including nucleic acids (such as bacterial deoxyribonucleic acid [DNA] and non-coding ribonucleic acid [RNA]), proteins (such as enzymes and virulence factors), and metabolites (such as short-chain fatty acid derivatives).7 Compared to free molecules, GMEVs possess unique biological advantages: their lipid bilayer membrane structure effectively protects their contents from degradation by the harsh intestinal environment (such as low potential of hydrogen [pH] and digestive enzymes), ensuring the intact delivery of active molecules;8,9 simultaneously, the specific adhesion molecules expressed on the surface of GMEVs confer tissue-targeting capabilities, enabling precise recognition and interaction with specific host cells.10,11 These characteristics suggest that GMEVs may play a pivotal messenger role between the gut microbiota, regarded as an “endocrine organ”, and distant target organs, thereby participating in the regulation of metabolic diseases such as obesity, diabetes, and MAFLD. In summary, this study focuses on the role of GMEVs in the relationship between gut microbiota and metabolic diseases, analyzing their biological origins, functional components, delivery mechanisms, and the influences of dietary factors. It also summarizes the regulatory mechanisms and advancements in the application of GMEVs in the treatment of metabolic diseases, aiming to provide a theoretical basis and novel intervention strategies for GMEV-targeted therapies in metabolic disorders (Figure 1).
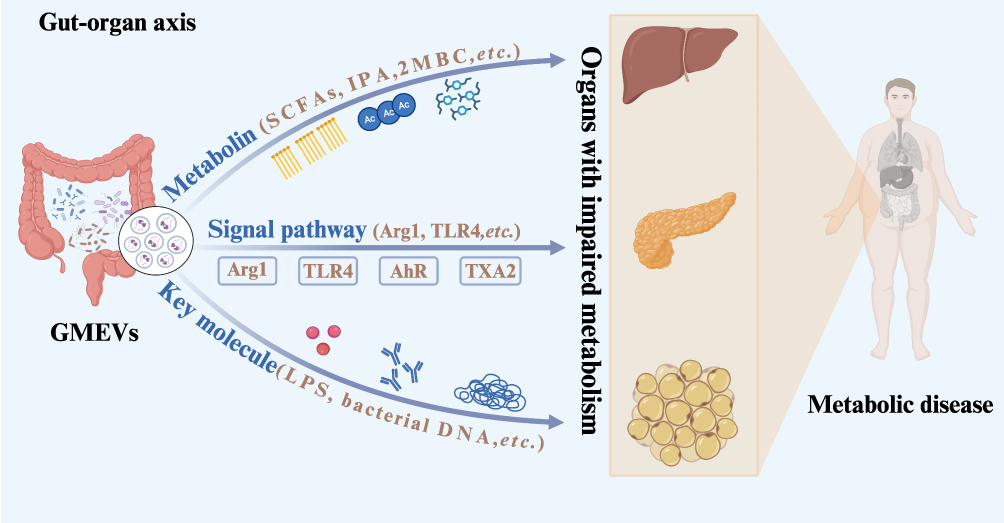 |
Figure 1 Schematic diagram of the “gut-organ axis” in gut microbiota and metabolic diseases. Abbreviations: GMEVs, Gut microbiota-derived extracellular vesicles; SCFAs, Short-Chain Fatty Acids; IPA, indolepropionic acid; 2MBC, 2-methylbutyrylcarnitine; Arg1, arginase 1; TLR4, toll-like receptor 4; LPS, lipopolysaccharides. |
Sources and Classification of GMEVs
GMEVs are nanoscale (30–300 nm) bilayer membrane structures actively secreted or passively released by gut microorganisms, playing a crucial role in microbial communication and host-microbe interactions.12,13 Based on the originating microbiota, gut microbiota-derived extracellular vesicles (EVs) can be primarily categorized into three major types: probiotics-derived EVs, pathogenic bacteria-derived EVs, and symbiotic bacteria-derived EVs (Table 1 and Figure 2).
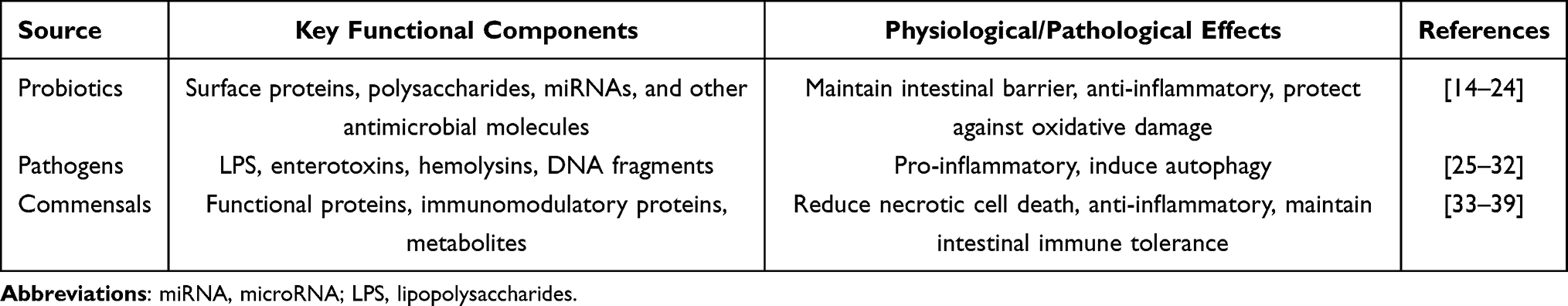 |
Table 1 Functional Comparison of GMEVs from Different Sources |
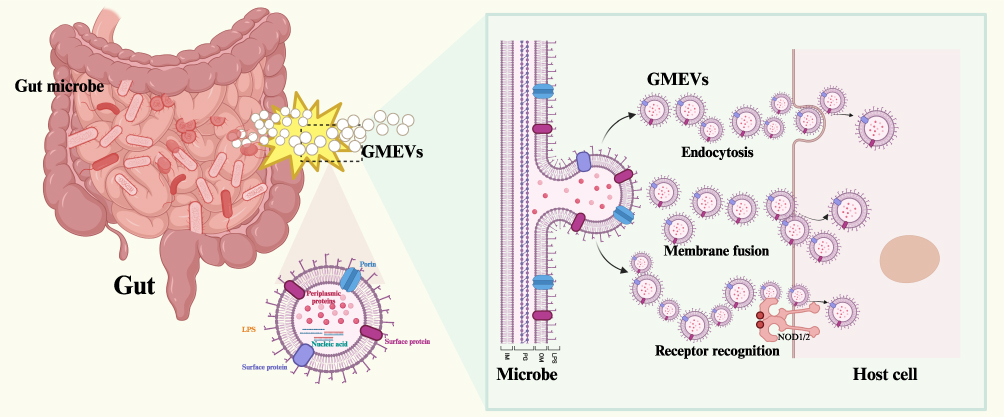 |
Figure 2 Schematic diagram of the molecular composition and delivery mechanism of GMEVs. Abbreviations: GMEVs, Gut microbiota-derived extracellular vesicles; LPS, lipopolysaccharides. |
Probiotics-Derived EVs
Probiotics-derived EVs, exemplified by Lactobacillus and Bifidobacterium, exert multifaceted regulatory roles in maintaining intestinal homeostasis and host health.14 These EVs typically carry a wealth of immunomodulatory factors, including immunogenic surface proteins and anti-inflammatory polysaccharide components,15–17 which can enhance intestinal epithelial barrier function and inhibit the colonization of pathogenic microorganisms through various mechanisms.40,41 Research has shown that extracellular vesicles derived from A. muciniphila can improve metabolic function in diabetic mice by reducing damage to the intestinal mucosal barrier and enhancing tight junction integrity.18 Furthermore, Bifidobacterium-derived EVs can carry antimicrobial substances and promote the expression of adhesion proteins that facilitate the growth of beneficial bacteria, thereby creating an environment unfavorable for pathogen colonization.42 Based on immune regulation, probiotics-derived EVs modulate host immune responses through complex signaling networks;19–21 their surface molecules interact with pattern recognition receptors on immune cells, activating the secretion of cytokines such as interleukin 22 (IL-22) and interleukin 17 (IL-17), while simultaneously inhibiting the excessive activation of pro-inflammatory signaling pathways like nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), thereby establishing an anti-inflammatory microenvironment in the gut.19–21 Notably, certain Bifidobacterium-derived EVs enhance the host cell’s antioxidant defense capabilities by activating the Nrf2 signaling pathway, mitigating oxidative stress-induced damage to the intestinal epithelium.14,22 Additionally, another class of probiotics-derived EVs from Escherichia coli Nissle 1917 regulate intestinal epithelial barrier function by upregulating the expression of tight junction proteins, thereby enhancing the integrity of intercellular connections to maintain the protective function of the gut barrier.23 Recent studies have revealed that probiotics-derived EVs possess cross-species gene regulatory capabilities; the miRNA-like molecules they carry can be taken up by host cells and mimic the functions of endogenous miRNAs, finely tuning gene expression networks by targeting specific mRNAs.24 This RNA-mediated gene silencing mechanism provides a novel molecular perspective for understanding microbial-host interactions.
Pathogenic Bacteria-Derived EVs
Pathogenic bacteria-derived EVs in the gut microbiota are primarily secreted by pathogenic strains such as Escherichia coli, Salmonella, Shigella, Staphylococcus aureus, and Bacillus cereus, acting as “molecular Trojan horses” within the intestinal microenvironment.43–45 These nanoscale vesicles are formed through the budding of the bacterial outer membrane, with their biogenesis strictly regulated by genetic factors, including EnvC-mediated outer membrane remodeling, Tol-Pal system-regulated membrane separation, and quorum sensing (QS) controlled programmed release.43,46,47 For molecular composition, these EVs represent a highly complex repository of bioactive molecules, rich in typical virulence factors such as lipopolysaccharides, enterotoxins, and hemolysins,25–27 while also carrying various antibiotic resistance genes and toxic DNA fragments.28,29 Notably, the lipopolysaccharides (LPS) on the surface of pathogenic bacteria-derived EVs exhibit immunostimulatory properties that are several times more potent than free LPS,45 capable of activating the expression of toll-like receptor 4 (TLR4) and macrophage-related markers in the host.30 Additionally, pathogenic bacteria-derived EVs can deliver a substantial amount of cytoplasmic enzymes into the host.31 These GMEVs not only inhibit autophagic flux by damaging autophagosome-lysosome fusion but also induce autophagy in the host; moreover, these pathogenic bacteria-derived EVs, laden with cytoplasmic enzymes, are more likely to activate non-classical inflammatory pathways, exacerbating the host’s inflammatory response.31,48
Symbiotic Bacteria-Derived EVs
In the gut microbiota, species such as Bacteroides serve as dominant symbiotic bacteria, and the EVs they secrete play complex and nuanced regulatory roles in maintaining host metabolic homeostasis and immune balance.48,49 These EVs are rich in unique bioactive molecules, including functional proteins, immunomodulatory proteins, and microbe-specific metabolites, which serve as important mediators of cross-talk between microorganisms and their hosts.33–36
In terms of metabolic function, symbiotic bacteria-derived EVs carry metabolites such as phosphatidylcholine, which can reduce necroptotic cell death by inhibiting the receptor-interacting protein kinase 1-receptor-interacting protein kinase 3-mixed lineage kinase domain-like protein (RIPK1-RIPK3-MLKL) signaling pathway.37 Regarding immune regulation, these EVs exhibit a distinctive dual regulatory characteristic: polysaccharide A (PSA) expressed on the surface of EVs derived from Bacteroides fragilis can mitigate inflammatory responses and potentially alter the composition of the microbiota, thereby preventing pathogen colonization.38 Additionally, these symbiotic bacteria-derived EVs effectively penetrate the intestinal barrier and are enriched with ATP-binding cassette (ABC) transporters and quorum sensing proteins. These proteins stimulate the secretion of IL-10 in co-culture systems involving splenic cells and dendritic cells with CD4+ T cells, demonstrating anti-inflammatory potential and maintaining intestinal immune tolerance.34,39 Throughout the co-evolution of hosts and microorganisms, Bacteroides-derived EVs have developed sophisticated immune evasion mechanisms. For instance, certain strains possess a capsule polysaccharide variation system on their EVs surfaces that can periodically alter their antigenic epitopes, thereby avoiding excessive clearance by the host immune system while maintaining necessary immune stimulation to “train” the host’s immune response capabilities.50,51 Disruption of this dynamic balance is associated with various diseases; for example, the proportion of immunoregulatory Bacteroides-derived EVs is significantly reduced in the intestines of patients with inflammatory bowel disease.35
With advancements in synthetic biology, GMEVs are emerging as novel therapeutic carriers due to their nanoscale structure, ability to penetrate the blood-brain barrier, low toxicity, high biocompatibility, and ease of modification. Engineered GMEVs, such as those synthesized to carry interleukin 4 (IL-4), have shown promise in promoting the repair of osteoporotic fractures.52 These studies not only deepen our understanding of the interaction mechanism between intestinal flora and host, but also provide theoretical basis and technical support for the development of precise intervention strategies based on microbial EVs.
Composition, Functional Characteristics, and Delivery Mechanisms of GMEVs
As important mediators of microbial-host interactions, GMEVs’ biological functions primarily rely on the diverse signaling molecules they carry and their sophisticated molecular delivery mechanisms. These nanoscale vesicles are rich in various bioactive substances, including lipopolysaccharides,53 flagellin proteins,54,55 nucleic acids,24 metabolites,37 and other protein components,42,56 which collectively form the functional basis of GMEVs. GMEVs interact with host cells through multiple delivery pathways, including membrane fusion, endocytosis, and receptor recognition. This precise regulatory characteristic positions them as key molecular bridges in microbial-host interactions.
LPS and Flagellin Proteins
GMEVs serve as crucial mediators in bacterial-host interactions, playing a complex and significant role in regulating host immune responses.57 These nanoscale vesicles are rich in pathogenic components such as LPS and flagellin proteins, which can efficiently deliver bacterial virulence factors to host cells through various mechanisms, triggering a series of immune responses.58 LPS, a major component of the outer membrane of Gram-negative bacteria, is abundantly present in GMEVs.59 Research indicates that GMEVs act as ideal carriers for LPS, protecting it from degradation in the external environment and ensuring its intact delivery to host target organs. This delivery process is primarily achieved through two mechanisms: first, GMEVs enter host cells via endocytosis and subsequently release LPS within early endosomes, delivering it to the cytoplasm and activating caspase-11-mediated inflammatory responses.60 Second, the LPS on the surface of GMEVs can directly bind to pattern recognition receptors on the host cell membrane, initiating a cascade of the TLR2/TLR4/NF-κB signaling pathway, which can induce insulin resistance and chronic inflammation.61,62 In the context of MAFLD, the LPS carried by GMEVs interacts with TLR4 on the host cell membrane, not only disrupting intestinal barrier function but also synergistically activating hepatic stellate cells in conjunction with circulating EVs, thereby promoting the progression of liver fibrosis.30,63 These two mechanisms collectively regulate the host’s inflammatory response and immune system function, exhibiting a dual role in maintaining intestinal homeostasis or triggering inflammatory responses. It is noteworthy that the immunomodulatory effects of GMEVs are closely related to the activity of the LPS they carry. For instance, in studies involving the pathogenic strain Escherichia coli Nissle 1917, it was found that GMEVs induced by glycine exhibited significantly reduced LPS activity, being 6–8 times lower than that of the non-induced group, yet still retained the ability to activate macrophage immune responses.64 This phenomenon suggests that the immunostimulatory effects of GMEVs depend not only on the absolute quantity of LPS but also on the overall structure and delivery efficiency of the vesicle. The biological functions of GMEVs carrying LPS in inducing host responses extend beyond local immune regulation; they can also traverse critical protective structures such as the intestinal barrier and blood-brain barrier, inducing responses in distal tissues and organ.65 Studies have shown that gut microbiota-derived GMEVs penetrate the blood-brain barrier, carrying LPS and other pro-inflammatory factors into the central nervous system, thereby activating neuroimmune cells such as microglia and triggering sustained neuroinflammatory responses, which can lead to tissue damage and organ dysfunction.66
Flagellin proteins primarily originate from motile intestinal bacteria. These bacteria deliver flagellin proteins to the cytoplasm of host immune cells, such as macrophages, via EVs. This delivery process is endocytosis-dependent but does not rely on guanosine triphosphate (GTP)-binding proteins. Once inside the cell, flagellin can strongly activate NOD-like receptor C4 (NLRC4)-mediated caspase-1 activation and IL-1β secretion, thereby triggering the classical activation pathway of the NLRC4 inflammasome.54 Research indicates that pathogen-associated molecular patterns (PAMPs) such as LPS and flagellin can help train the immune system at low concentrations, but excessive amounts can lead to inflammatory responses.67 Salmonella, for instance, utilizes a transport pathway regulated by its SPI-2 virulence island to deliver flagellin to the basolateral side of epithelial cells, activating the TLR5 inflammatory signaling pathway and promoting mucosal inflammation.68 Notably, this process does not depend on the motility of the bacteria or their direct invasion.
Nucleic Acids, Metabolites, and Other Proteins
In addition to LPS and flagellin proteins, GMEVs are also rich in key signaling molecules such as nucleic acids, metabolites, and other functional proteins. Once delivered to host cells, these nucleic acid components can specifically activate nucleotide-binding oligomerization domain-containing protein 1 (NOD1), facilitating immune crosstalk with the epithelial cells on the gastrointestinal surface and regulating the activation of innate immune receptors in the gastrointestinal mucosa.69 Recent discoveries have identified miRNA-like small RNAs, which, despite their shorter length and lack of typical hairpin structures, can exert gene-silencing effects within host cells. This phenomenon of cross-domain regulation provides new insights into the co-evolution of hosts and microorganisms. Studies have shown that analyzing the miRNA characteristics in GMEVs using machine learning-driven models can serve as effective biomarkers for disease diagnosis, prediction, and early warning.13
Regarding metabolites, dysbiosis in the gut microbiota can significantly alter amino acid levels and the balance of short-chain to long-chain fatty acids, contributing to disease pathogenesis. These findings greatly expand our understanding of the functional diversity of GMEVs.70 Notably, GMEVs derived from gut microbiota can deliver bacterial effector proteins to the central nervous system via the circulatory system or neural pathways, regulating the onset and progression of neuroinflammatory diseases.66 Additionally, GMEVs released by non-pathogenic E. coli Nissle 1917 can carry inner membrane and cytoplasmic proteins to host cells, modulating host immune functions.64 Recent studies have found that the secretion of GMEVs from probiotic gut microbiota is reduced in diabetic model rats, leading to decreased levels of functional proteins or lipid components such as Amuc-1100, P9, and phosphatidylcholine in target organs. This reduction results in impaired intestinal barrier function, suppression of glucagon-like peptide-1 (GLP-1) levels, and disruption of insulin signaling pathways, ultimately triggering a series of metabolic syndrome-related issues.71 These findings highlight the synergistic advantages of GMEVs as multi-component carriers—transporting proteins, metabolites, and genetic materials—that can target and regulate gut-liver axis functions while simultaneously intervening in gut microbiota reconstruction, metabolic reprogramming, and systemic inflammation alleviation.72 This offers new perspectives for the prevention and treatment of metabolic diseases such as obesity, MAFLD, and diabetes.
Molecular Delivery Mechanisms and Regulatory Characteristics of GMEVs
GMEVs interact with host cells through diverse molecular delivery mechanisms, including membrane fusion, endocytosis, and receptor recognition. These precisely regulated delivery processes enable GMEVs to dynamically influence the physiological and pathological states of the host. Membrane fusion is a typical delivery method for EVs derived from Gram-positive bacteria. GMEVs preferentially fuse with the liquid-ordered regions of host cell membranes, altering the fluidity, dipole potential, and compressibility of the host membrane. The efficiency of this fusion is regulated by lipid composition (eg, phosphatidylcholine and C16 acyl chains promote fusion, while cholesterol inhibits it), thereby affecting virulence transfer and host mechanism manipulation.73
In contrast, endocytosis is a more common delivery pathway. GMEVs enter host cells through clathrin-dependent endocytosis, where the DNA fragments carried by GMEVs determine the efficiency of internalization.74 Recent studies have also identified an atypical endocytic mechanism—raft-mediated uptake—dependent on membrane cholesterol microdomains. GMEVs adhere via a pathway that does not rely on α5β1 integrin but rather on lipid rafts, with subsequent endocytosis relying on cholesterol and sphingolipid components, driving actin remodeling through the recruitment of Rac1 to raft regions.75 Receptor binding is another key mechanism through which GMEVs exert their functions. GMEVs, by packaging immunostimulatory molecules such as peptidoglycan and LPS, are recognized and activated by host cell NOD-like receptors (NLRs), such as nucleotide-binding oligomerization domain containing protein 1 (NOD1), nucleotide-binding oligomerization domain containing protein 2 (NOD2), and NOD-like receptor family pyrin domain-containing 3 (NLRP3), thereby driving inflammatory responses or maintaining intestinal immune homeostasis.76 The delivery and synergistic action of these molecules enable GMEVs to precisely regulate the physiological and pathological processes of the host: under homeostatic conditions, symbiotic bacterial EVs maintain intestinal tolerance by delivering immune regulatory molecules;34 conversely, during dysbiosis, the excessive release of virulence factors from pathogenic bacterial EVs disrupts barrier functions.77 Particularly noteworthy is the spatiotemporal specificity of GMEVs’ molecular delivery. For instance, circadian rhythm disturbances alter the metabolic signaling molecules in GMEVs, reducing insulin sensitivity in adipocytes and inducing systemic inflammation, indicating that GMEVs mediate the relationship between circadian disruption and metabolic abnormalities.78 This precise regulatory mechanism allows GMEVs to dynamically adjust their biological effects according to the host’s state.
The Impact of Metabolic Risk Factors on GMEVs
The biological characteristics of GMEVs establish their unique position in the regulation of host metabolism. For one thing, the molecules carried by GMEVs exhibit strain specificity, with significant differences in composition and function among GMEVs produced by different strains, and even among strains of the same species. For another, GMEVs demonstrate environmental responsiveness and tissue tropism, with their production and composition dynamically adjusting in response to environmental factors such as diet and medication.61 Following their generation, GMEVs can target specific organs through pathways such as the portal venous system and lymphatic circulation.11 Research indicates that a high-fat diet can markedly alter the physical properties and molecular composition of GMEVs, such as increasing vesicle diameter, elevating LPS content, and reducing the levels of phospholipid fatty acids.79,80 These changes do not entirely align with alterations in microbial community composition, suggesting that diet may influence GMEV production by directly modulating bacterial membrane components. At the molecular level, the composition of fatty acids, particularly the ratio of saturated to unsaturated fatty acids, affects GMEV generation by regulating bacterial membrane fluidity. For instance, saturated fatty acids can dose-dependently promote the release of GMEVs from Bacteroides fragilis, while having no significant effect on Bacteroides thetaiotaomicron; unsaturated fatty acids, conversely, exhibit differential regulatory effects across various strains.81 Furthermore, disruptions in the body’s biological rhythms can also impact the development of metabolic diseases through GMEVs. Studies have found that reversing light cycles can lead to insulin resistance, inflammation in adipose tissue, and a reduction in regulatory T cells, with mechanisms involving gut barrier disruption, GMEV-mediated inhibition of insulin signaling, and peripheral circadian rhythm disturbances.78 The circadian regulatory characteristics of GMEVs provide a novel perspective on the temporal biological features of metabolic diseases. These findings underscore the pivotal role of GMEVs in the regulation of the diet-microbiome-host metabolism axis and suggest that they may serve as a crucial molecular bridge connecting biological rhythms with metabolic homeostasis.
The Regulatory Role of GMEVs in Metabolic Diseases
As key mediators of microbial-host interactions, GMEVs play a central regulatory role in the onset and progression of metabolic diseases. They deliver microbial-derived bioactive molecules to affect the host’s energy metabolism, immune homeostasis and endocrine function in a multi-target and multi-level manner, thereby participating in pathological processes such as insulin resistance, chronic low-grade inflammation, and lipid metabolism disorders.82,83 The gut microbiota, utilizing the cross-communication capabilities of GMEVs, can simultaneously target the host’s intestinal barrier function and the metabolic homeostasis of distant organs (Table 2), thereby forming a comprehensive regulatory network across spatial and temporal dimensions. This “local-distant” synergistic action enables GMEVs to integrate microbial signals with host physiological needs, offering a fresh perspective on the pathogenesis of metabolic diseases through the lens of “microbial-host dialogue”. It also lays a significant theoretical foundation for the development of precision therapeutic strategies based on microbiome interventions.
 |
Table 2 Dual Roles of GMEVs in Metabolic Diseases |
Targeting the Host Intestinal Barrier
GMEVs play a pivotal regulatory role in the onset and progression of metabolic diseases, with one of their core mechanisms being the targeting of the host’s intestinal barrier system to induce systemic metabolic disturbances (Figure 3). The regulation of intestinal barrier function by GMEVs manifests primarily at two levels: the physical barrier and the immune barrier. Regarding the physical barrier, GMEVs from different sources exert differential regulatory effects on the tight junction protein network. For instance, EVs derived from pathogenic bacteria encapsulate virulent toxins that can inhibit the expression of tight junction proteins such as Occludin and ZO-1, thereby compromising the intestinal epithelial barrier and activating intrinsic inflammation.87 Concurrently, pathogenic bacterial EVs may reduce the overall concentration of short-chain fatty acids in the gut microbiota, inducing inflammatory responses that damage the ileal mucosa.88 In contrast, EVs derived from probiotics or symbiotic bacteria exhibit opposing effects. The quantity of EVs originating from the probiotic A. muciniphila is significantly greater in fecal samples from healthy individuals compared to those from patients with type 2 diabetes.18 Furthermore, in a diabetic mouse model fed a high-fat diet, it was observed that EVs derived from A. muciniphila can influence intestinal permeability by modulating tight junctions.18 This enhancement of tight junction functionality not only mitigates weight gain and improves glucose tolerance but also reduces intestinal permeability in Caco-2 cells treated with LPS,18 while simultaneously upregulating the expression of Occludin to bolster intestinal barrier function.84,86 Similarly, EVs from symbiotic bacteria can effectively penetrate the intestinal barrier system, maintaining intestinal immune tolerance and indirectly enhancing tight junction integrity.34 When the gut microbiota becomes dysregulated, an increase in pathogenic bacteria leads to the destruction of the host’s intestinal barrier, resulting in the translocation of endotoxins (particularly LPS) from the gut, which contributes to metabolic endotoxemia. The circulating LPS activates inflammatory signaling in adipose tissue and the liver by binding to TLR4 receptors, serving as a significant trigger for obesity-related insulin resistance.101,102 Conversely, probiotic-derived EVs can help restore gut microbiota balance and promote the production of short-chain fatty acids, thereby improving intestinal barrier function.90 In terms of immune barrier regulation, GMEVs activate the immune cell network within the intestinal lamina propria through multiple pathways. Macrophages, as the first line of defense in intestinal immunity, are precisely regulated by GMEVs. GMEVs from probiotics can promote the M2 anti-inflammatory polarization of macrophages, enhancing the levels of tight junction proteins and reducing permeability, thereby further strengthening the intestinal barrier through dual immune-barrier regulation.89 In contrast, GMEVs from pathogenic bacteria inhibit the granulocyte-macrophage colony-stimulating factor/signal transducer and activator of transcription 5 (staT5)/Arg-1 signaling pathway, blocking the differentiation of macrophages into an anti-inflammatory phenotype and promoting their polarization towards the pro-inflammatory M1 phenotype.99 GMEVs not only recruit macrophages to the intestine, disrupting the balance of intestinal immunity, but also enhance pro-inflammatory responses by altering their functional state. Under the influence of GMEVs, dendritic cells (DCs) become activated, and through differential regulation of miRNAs, they affect T cell responses. Following stimulation by GMEVs, DCs induce naïve CD4+ T cells to secrete effector cytokines.100 Notably, the differences in Th1 and Treg-related factors are pronounced. The EVs released by DCs are enriched with co-stimulatory molecules, and the immune-related miRNAs they carry vary depending on the source of the GMEVs.100 These disparities align with the phenotypes of immune responses, indicating that GMEVs regulate adaptive immune polarization by reshaping the secretion profile of DC-derived EVs.
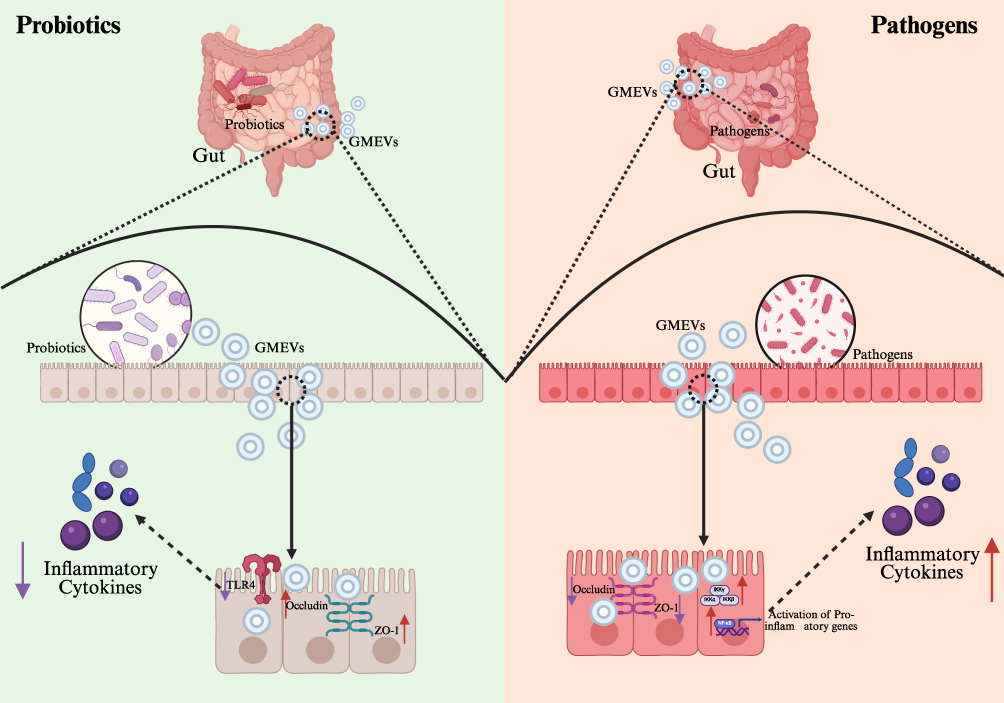 |
Figure 3 The mechanistic role of GMEVs in modulating metabolic diseases via intestinal barrier regulation. Purple arrows (↓) denote downregulation, while red arrows (↑) denote upregulation. Abbreviations: GMEVs, Gut microbiota-derived extracellular vesicles; TLR4, toll-like receptor 4; ZO-1, Zona Occludens-1. |
In the clinical translation of metabolic diseases, the barrier-regulating properties of GMEVs hold dual value: for one thing, the detection of specific GMEV subpopulations in feces could serve as early biomarkers for intestinal barrier damage; for another thing, engineered probiotic GMEVs, which overexpress intestinal barrier-stimulating factors such as Occludin, hold promise as novel therapeutic agents for repairing the intestinal barrier. Recent studies suggest that targeted delivery of protective components of GMEVs to the intestine via nanocarriers can inhibit pro-inflammatory factors, repair the mucus layer and tight junction barriers, and restore gut microbiota balance while suppressing inflammation through multiple mechanisms, thereby counteracting the destructive effects of pathogenic GMEVs.103 With the advent of single-cell sequencing and spatial transcriptomics, our understanding of the relationship among GMEVs, the intestinal barrier, and metabolic dysregulation is deepening from a tissue level to a cellular subpopulation level, providing a theoretical basis for the development of precise intervention strategies. Future research should focus on addressing the heterogeneity of GMEVs (the differences in EVs produced by the same species under varying physiological states) and establishing more humanized mouse models that closely mimic the true roles of GMEVs in human metabolic diseases.
Interactions with Distant Organs
GMEVs serve as crucial mediators of communication between gut microbiota and the host, carrying a wealth of bioactive molecules that facilitate inter-organ signaling through the circulatory system, thereby participating in the metabolic regulation of distant tissues.9 These vesicles, ranging from approximately 30 to 300 nm in diameter, not only mediate local immune regulation but also exert systemic effects on glucose and lipid metabolism and immune responses, providing a novel perspective for understanding the pathogenesis of metabolic diseases. The distal regulatory effects of GMEVs on lipid metabolism in metabolic diseases exhibit a complex “double-edged sword” effect. For one thing, GMEVs derived from pathogenic bacteria can inhibit the tyrosine phosphorylation of insulin receptor substrates, thereby diminishing the sensitivity of adipocytes to insulin.92 For another, GMEVs from probiotics demonstrate significant protective effects on lipid metabolism. Research indicates that probiotic-derived GMEVs are more effective than the bacteria themselves in ameliorating obesity, significantly reducing body weight and fat mass in mice subjected to a high-fat diet. This protective mechanism involves enhancing intestinal barrier function, reducing systemic LPS leakage, upregulating the expression of lipolysis-related genes in adipose tissue, and promoting the browning of white adipose tissue.30,84,94 Notably, the effects of these probiotic GMEVs surpass those of heat-inactivated bacterial cells, highlighting their therapeutic potential as “postbiotics”.12,86 In terms of liver metabolic regulation, GMEVs participate in the development of MAFLD through the “gut-liver axis” (Figure 4). The portal circulation positions the liver as the primary target organ for GMEV action.63 Studies have shown that pathological GMEVs produced during gut microbiota dysbiosis can promote liver injury through various mechanisms:11 increasing intestinal permeability to allow more LPS into the portal circulation,30 directly activating hepatic stellate cells to promote collagen deposition and fibrosis,84 and interfering with hepatocyte lipid metabolism.96 Particularly noteworthy is the ability of some GMEVs to traverse the pores of hepatic sinusoidal endothelial cells and enter peripheral circulation, which may represent a significant mechanism for the “dissemination” of metabolic inflammation throughout the body.30
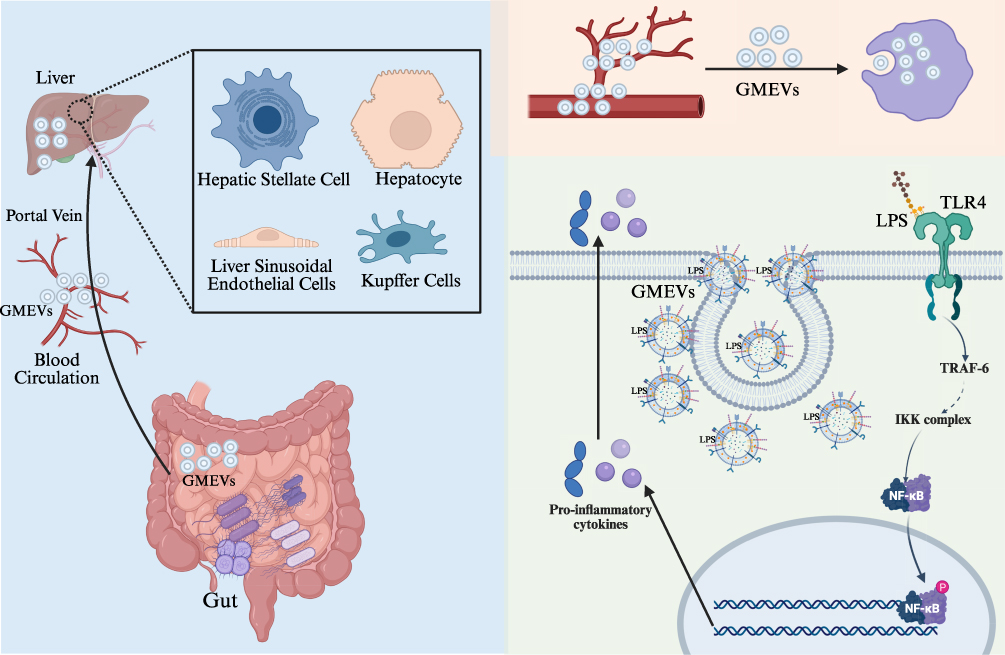 |
Figure 4 The “gut-liver axis” interaction network of GMEVs. Abbreviations: GMEVs, Gut microbiota-derived extracellular vesicles; LPS, lipopolysaccharides; TLR4, toll-like receptor 4; TRAF-6, Recombinant TNF Receptor Associated Factor 6; NF-κB, nuclear factor kappa-B. |
As key messengers linking the gut micro-environment with systemic metabolism, GMEVs play a central regulatory role in the dysregulation of glucose metabolism in metabolic diseases through their unique compositional characteristics and inter-organ delivery capabilities. Research on obese diabetic rat models has revealed that dysbiosis in diabetic rats leads to altered proportions of gut microbiota-derived GMEVs, impaired glucose tolerance, and disrupted glucose metabolism.93 This indicates that GMEVs can participate in the process of glucose metabolic dysregulation by delivering bacterial active components, exerting remote effects on multiple metabolic organs. For instance, a study involving patients with type 2 diabetes found that increased intestinal permeability in these patients facilitated the abnormal absorption of GMEVs and their contents, resulting in chronic inflammation and metabolic dysregulation.91 This process promotes the translocation of GMEVs across the intestinal barrier into the circulatory system, where they induce insulin resistance and impaired glucose tolerance through the delivery of microbial components. Additionally, other studies have reported that GMEVs from mice fed a high-fat diet can induce insulin resistance and glucose intolerance, a process characterized by the specific blockade of insulin signaling in skeletal muscle and adipose tissue, leading to impaired glucose tolerance and systemic insulin resistance, thereby remotely regulating host glucose metabolism.79 Concurrently, the virulence factors carried by GMEVs can be transferred to the liver via the circulatory system, reducing hepatic sensitivity to insulin, inhibiting glycogen synthesis, and resulting in sustained hyperglycemia, which disrupts hepatic glucose metabolic homeostasis and directly impairs insulin signaling and glycogen synthesis, thereby promoting the progression of metabolic dysregulation in diabetes.104
The gut microbiota, as a complex and dynamic ecosystem, is regarded as an “essential organ” of the human body,105 playing a significant regulatory role in modulating immune responses, including inflammation.106 GMEVs, as messengers of the gut microbiota, also play a crucial role in maintaining inflammatory balance in metabolic diseases. Research indicates that GMEVs derived from pathogenic bacteria, such as vesicles containing high levels of LPS, can promote inflammation in adipose tissue through the TLR4 signaling pathway, characterized by an increase in pro-inflammatory macrophage infiltration and a decrease in regulatory T cells (Tregs).11,63 Numerous studies have shown that the biological characteristics of GMEVs undergo significant changes under metabolic dysregulation, thereby influencing the host’s inflammatory response and immune response. For instance, it has been reported that the proportion of LPS-carrying EVs in the feces of diet-induced obese mice significantly increases, along with a higher abundance of Proteobacteria.30 These EVs can breach the intestinal barrier and enter the liver and bloodstream, leading to upregulation of TLR4 expression and activation of inflammatory factors, thereby promoting metabolic inflammation.30 The structural differences in LPS from specific gut bacteria (such as Ralstonia) affect their pro-inflammatory capacity; low-acylation LPS strains, although exhibiting weaker inflammatory activity, secrete more EVs, efficiently transporting LPS into the bloodstream and exacerbating chronic low-grade inflammation.97 Recent studies have also found that specific bacterial DNA markers in serum are significantly correlated with the degree of perirenal fat inflammation, and the EPC index established based on these markers (AUC=0.81) offers a novel approach for non-invasive assessment of adipose tissue inflammation.107 Furthermore, GMEVs play a critical role in the progression of diabetic nephropathy. A study investigating the regulatory role of GMEVs in the development of diabetic nephropathy showed that GMEVs from diabetic nephropathy rats can migrate from the gut to the renal interstitium, inducing renal inflammatory damage through the Caspase-11 inflammatory pathway.94
Notably, the host organs are not passive recipients of GMEV-mediated regulation but actively modulate the generation, release, and function of GMEVs through multiple mechanisms. Studies have reported that metabolic states such as diabetes can alter the intestinal environment, thereby influencing the gut microbiota composition and their vesicle secretion behaviors. For instance, in diabetic rats treated with bariatric/metabolic surgery, glucose tolerance and microbial richness were reversed, accompanied by changes in GMEV composition.93 Furthermore, the liver, as a major organ for the clearance and modification of GMEVs, actively participates in regulating their systemic distribution and biological effects.108 Upon entering hepatic tissue, GMEVs are actively phagocytosed by Kupffer cells, which contributes to their removal from circulation.108 Recent evidence also suggests that the host neuroendocrine system may indirectly influence the secretion profile of GMEVs via the gut-brain axis, further highlighting the complexity of the bidirectional communication network between GMEVs and the host.71,109
Therapeutic Applications of GMEVs
As “microbial messengers” connecting microorganisms with the host, GMEVs exhibit unprecedented core value in the pathogenesis and intervention strategies of metabolic diseases. These nanoscale bioactive carriers, through their unique molecular composition and precise delivery mechanism, have built a “microbiome-host dialogue system” that transcends species boundaries. This provides a new perspective for understanding the microbiome basis of metabolic diseases, and also provides a new, efficient and safe microecological intervention strategy for the treatment of metabolic diseases. Research has shown that EVs derived from gut probiotics, as natural nanocarriers, can improve metabolic disorders by repairing the intestinal barrier, regulating microbial balance, and suppressing inflammatory responses.110 They not only enhance intestinal integrity to prevent endotoxin entry into the bloodstream but also regulate systemic glucose and lipid metabolism and neuroendocrine functions through the gut-liver/brain axis, providing new strategies for the microecological treatment of metabolic diseases such as obesity and diabetes.110 Moreover, GMEVs improve intestinal barrier function and regulate the gut-liver metabolic axis, ultimately achieving reductions in blood glucose levels, increased insulin sensitivity, and alleviation of obesity.90 They intervene in the progression of metabolic diseases through mechanisms of microbial-host metabolic dialogue.90 Additionally, GMEVs effectively ameliorate ethanol-induced liver damage by enhancing hepatic antioxidant capacity and reducing blood ethanol concentrations, thereby modulating key pathological processes such as oxidative stress.111 EVs derived from Akkermansia muciniphila can significantly enhance intestinal barrier function and the integrity of the mucosal layer by delivering functional proteins and various bioactive components, while also modulating glucose and lipid metabolism. They improve symptoms of obesity and diabetes by activating GLP-1 signaling to enhance insulin sensitivity and inhibiting the NF-κB pathway to alleviate systemic low-grade inflammation and regulate blood glucose and lipid levels. Furthermore, they influence hepatic lipid metabolism and neuroinflammation through the circulatory system, demonstrating a superior effect compared to that of the bacterial cells themselves.61,86,112 Akkermansia muciniphila-derived EVs outperform live bacteria in improving metabolic dysregulation, significantly inhibiting obesity induced by a high-fat diet, reducing weight gain, metabolic organ weight, and plasma metabolic abnormalities, whereas also alleviating inflammation in the gut, adipose tissue, and liver, and effectively restoring gut microbiota balance and increasing beneficial bacterial abundance.84 Studies have shown that GMEVs from healthy mice conditioned with oral nano-drugs directly alleviate colonic inflammation and repair the mucosal barrier by reshaping gut microbiota balance, whereas also regulating purine metabolism to lower uric acid levels, demonstrating significantly greater efficacy than traditional fecal microbiota transplantation.113 The cross-boundary information transmission role of GMEVs provides novel intervention strategies for metabolic diseases such as metabolic syndrome, obesity, and diabetes, with their multi-target and low-toxicity characteristics making them highly promising therapeutic carriers (Table 3).
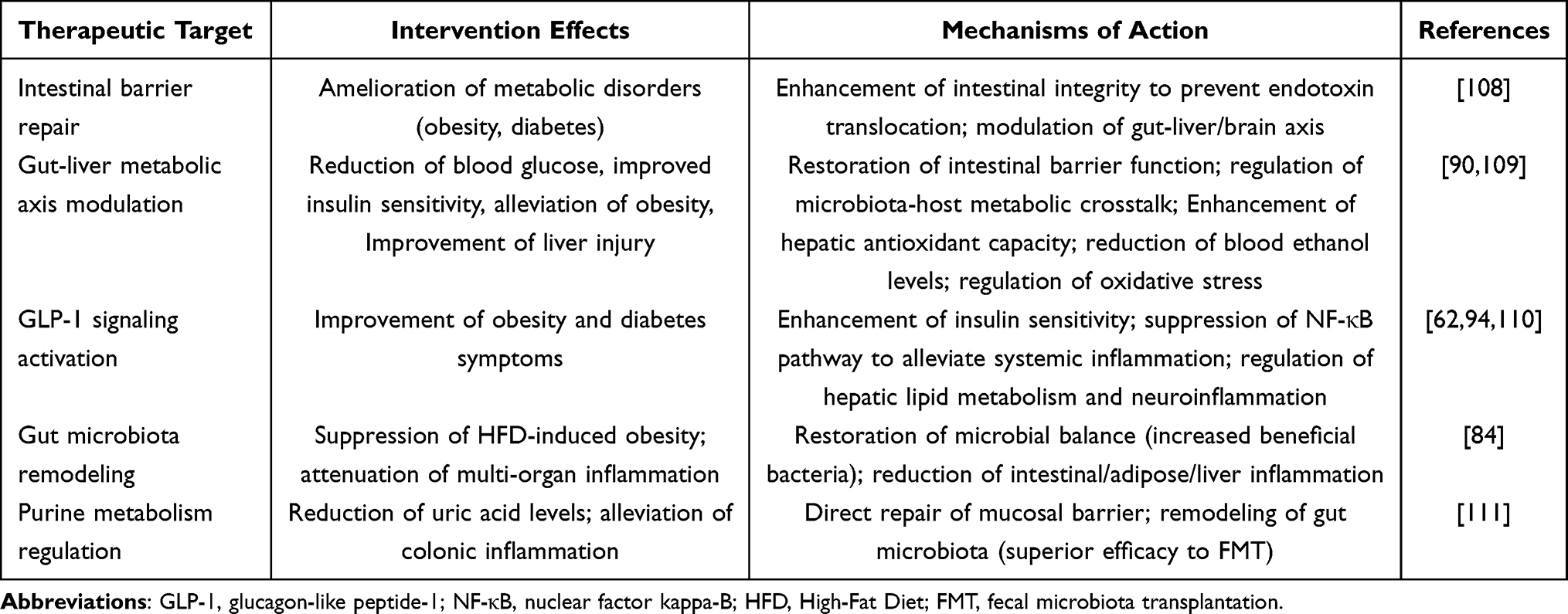 |
Table 3 Therapeutic Targets and Mechanisms of GMEVs in Metabolic Diseases |
Limitations and Future Perspectives
GMEVs, as important mediators of interactions between gut microbiota and the host, have demonstrated significant potential in the diagnosis and treatment of metabolic diseases in recent years. However, there are still several limitations in this field. For instance, the biogenesis pathways of GMEVs and their regulatory factors remain incompletely understood, particularly regarding how environmental signals (such as dietary components) are translated into molecular mechanisms that instruct vesicle loading. Additionally, the lack of standardized methods for the isolation and identification of GMEVs limits the comparability of research findings, while quality control issues in large-scale production pose major bottlenecks for clinical translation. Furthermore, the potential risks associated with long-term GMEV application (such as the induction of autoimmune responses) require more comprehensive evaluation systems. Notably, current research predominantly focuses on specific components such as LPS, while the roles of other functional molecules carried by GMEVs (eg, microbial-derived miRNAs, signaling proteins, and metabolites) in host metabolic and immune regulation remain poorly understood, thereby limiting our comprehensive understanding of the systemic functions of GMEVs. Addressing these challenges necessitates the establishment of an internationally collaborative research network, the development of next-generation analytical tools, and the formulation of unified technical standards.
Looking ahead, research on GMEVs can leverage two key advantages to develop and apply clinical diagnostic markers, providing a novel source of biomarkers for disease diagnosis (Figure 5). First, GMEVs are continuously released into the circulatory system by gut microbiota and can stably exist in clinical samples such as blood and feces. Second, they carry bioactive molecules, including proteins, nucleic acids, lipids, and metabolites from the parent bacteria, which can cross the intestinal mucosal barrier and directly or indirectly regulate the host’s metabolic and immune responses. Not only does the double-membrane structure of GMEVs effectively protect their cargo from enzymatic degradation, thereby preserving the stability of bioactive molecules such as proteins, nucleic acids, and metabolites in blood, urine, and other samples over extended periods, but their surface markers also exhibit species-specificity, facilitating traceability analysis. More importantly, although the secretion profile and molecular composition of GMEVs undergo dynamic changes in response to external factors, these alterations follow predictable and identifiable patterns that reflect characteristic features of microbiota-host interactions under specific physiological or pathological conditions. For instance, under high-fat diet or circadian rhythm disruption, although the total quantity or specific components of GMEVs may be altered, these changes are often closely associated with metabolic abnormalities. Consequently, rather than diminishing their diagnostic value, such variations provide temporal information that can be utilized for disease staging, risk stratification, and dynamic monitoring.9,114,115 Population studies have shown that the characteristics of EVs in urine can predict the risk of abdominal obesity: individuals with a high abundance of Firmicutes and a low abundance of Proteobacteria have a 2.59-fold increased risk of abdominal obesity within ten years.116 Multi-omics analyses indicate that the microbial composition of EVs in the feces, serum, and urine of diabetic patients exhibits abnormal co-occurrence patterns, which may serve as new non-invasive biomarkers for diabetes diagnosis.91 Moreover, the composition and functional characteristics of gut microbiota undergo specific changes under different disease states, and these alterations are directly reflected in the molecular profiles of GMEVs.117–120 This makes it possible to achieve early disease diagnosis by detecting specific components of GMEVs in blood or feces. In metabolic diseases such as obesity, diabetes, and MAFLD, dysregulation of gut microbiota composition and function is often accompanied by changes in the secretion profiles of GMEVs.11,94,121 These GMEVs, derived from specific sources or containing specific components, participate in disease development by influencing insulin sensitivity, lipid metabolism, and inflammatory responses. Their unique molecular characteristics also provide new biomarkers for early diagnosis and classification of diseases. These findings suggest that the microbial source characteristics or specific molecules contained within GMEVs (such as species-specific proteins, differential miRNAs, and specific metabolites) may serve as potential diagnostic indicators for metabolic diseases.11,94,121 These findings suggest that by integrating multi-timepoint sampling, dynamic monitoring, and big data analytics, it is anticipated that the compositional dynamics of GMEVs can be transformed into “dynamic biomarkers” reflecting an individual’s metabolic status, thereby further enhancing their potential for application in precision diagnosis. Future research should integrate multi-omics technologies and longitudinal cohorts to further clarify the dynamic changes of GMEVs in metabolic diseases and their sensitivity and specificity as diagnostic markers. This will promote their clinical application and achieve a qualitative leap from “understanding microorganisms” to “harnessing microbial signals”, providing new tools for the prevention and treatment of metabolic diseases.
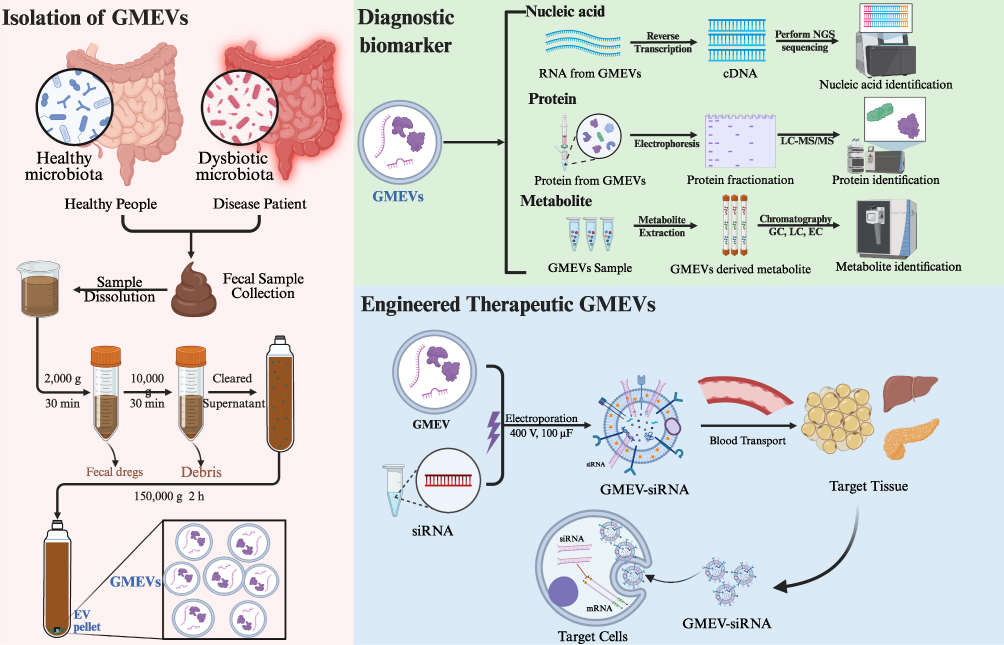 |
Figure 5 The Application Prospects of GMEVs as Diagnostic Markers and Therapeutic Vectors. Abbreviations: GMEVs, Gut microbiota-derived extracellular vesicles; siRNA, Small interfering RNA. |
Conclusion
In summary, as mediators of cross-communication between microorganisms and the host, GMEVs carry active molecules that mediate the regulation of the “gut-organ axis”. In metabolic diseases such as obesity and diabetes, they play dual roles: GMEVs derived from pathogenic bacteria exacerbate metabolic inflammation and insulin resistance by disrupting the intestinal barrier, activating the TLR4/NF-κB inflammatory pathway, and inhibiting insulin signaling. In contrast, GMEVs from probiotics and commensal bacteria exert protective effects by enhancing tight junctions, regulating immune responses, and improving glucose and lipid metabolism. Metabolic risk factors can dynamically modulate the composition and function of GMEVs, and the specific molecular characteristics provide novel biomarkers for early disease diagnosis. Despite challenges such as standardization of isolation methods, GMEVs exhibit vast potential in precision therapy, such as engineered probiotic EVs for intestinal barrier repair and as drug delivery vehicles. Future research should delve deeper into the spatiotemporal interaction patterns of GMEVs and host response mechanisms, facilitating the translation of basic research into clinical diagnostics and intervention strategies, and providing a new perspective on the “microbial-host dialogue” for the prevention and treatment of metabolic diseases.
Funding
This work was supported by grants from the guiding project of the “Academician Liu Liang Workstation” (No: 24YS003), National Natural Science Foundation of China (No: U21A2041), Hunan Provincial Natural Science Foundation Innovation Research Group Project (No: 2024JJ1007), Hunan University of Chinese Medicine Disciplinary Construction “Revealing the List and Appointing Leaders” Project (No: 22JBZ002), Hunan Provincial Postgraduate Research and Innovation Project (No:CX20251150), Scientific Research Project of Hunan Provincial Health Commission (No:20258052).
Disclosure
The authors declare that there are no conflicts of interest.
References
1. Porcari S, Fusco W, Spivak I, et al. Fine-tuning the gut ecosystem: the current landscape and outlook of artificial microbiome therapeutics. Lancet Gastroenterol Hepatol. 2024;9(5):460–475. doi:10.1016/S2468-1253(23)00357-6
2. Fukasawa N, Tsunoda J, Sunaga S, et al. The gut-organ axis: clinical aspects and immune mechanisms. Allergol Int. 2025;74(2):197–209. doi:10.1016/j.alit.2025.01.004
3. Boulangé CL, Neves AL, Chilloux J, et al. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016;8(1):42. doi:10.1186/s13073-016-0303-2
4. Tilg H, Zmora N, Adolph TE, Elinav E. The intestinal microbiota fuelling metabolic inflammation. Nat Rev Immunol. 2020;20(1):40–54. doi:10.1038/s41577-019-0198-4
5. Doğan D, Çelik T. Research trends on the gut microbiota in endocrine metabolism: a thematic and bibliometric analysis. Front Cell Infect Microbiol. 2024;14:1371727. doi:10.3389/fcimb.2024.1371727
6. Cheng G, Liu Y, Guo R, et al. Molecular mechanisms of gut microbiota in diabetic nephropathy. Diabet Res Clin Pract. 2024;213:111726. doi:10.1016/j.diabres.2024.111726
7. Melo-Marques I, Cardoso SM, Empadinhas N. Bacterial extracellular vesicles at the interface of gut microbiota and immunity. Gut Microbes. 2024;16(1):2396494. doi:10.1080/19490976.2024.2396494
8. Macia L, Nanan R, Hosseini-Beheshti E, et al. Host- and microbiota-derived extracellular vesicles, immune function, and disease development. Int J Mol Sci. 2019;21(1):107. doi:10.3390/ijms21010107
9. Taitz JJ, Tan JK, Potier-Villette C, et al. Diet, commensal microbiota-derived extracellular vesicles, and host immunity. Eur J Immunol. 2023;53(7):e2250163. doi:10.1002/eji.202250163
10. Northrop-Albrecht EJ, Taylor WR, Huang BQ, et al. Assessment of extracellular vesicle isolation methods from human stool supernatant. J Extracell Vesicles. 2022;11(4):e12208. doi:10.1002/jev2.12208
11. Fizanne L, Villard A, Benabbou N, et al. Faeces-derived extracellular vesicles participate in the onset of barrier dysfunction leading to liver diseases. J Extracell Vesicles. 2023;12(2):e12303. doi:10.1002/jev2.12303
12. Liang X, Dai N, Sheng K, et al. Gut bacterial extracellular vesicles: important players in regulating intestinal microenvironment. Gut Microbes. 2022;14(1):2134689. doi:10.1080/19490976.2022.2134689
13. Zhang Z, Liu X, Peng C, et al. Machine learning-aided identification of fecal extracellular vesicle microRNA signatures for noninvasive detection of colorectal cancer. ACS Nano. 2025;19(10):10013–10025. doi:10.1021/acsnano.4c16698
14. Jiao Q, Liu J, Zhou L, et al. Lactobacillus extracellular vesicles alleviate alcohol-induced liver injury in mice by regulating gut microbiota and activating the Nrf-2 signaling pathway. Food Funct. 2025;16(4):1284–1298. doi:10.1039/d4fo04364b
15. de Rezende Rodovalho V, da Luz B, Nicolas A, et al. Environmental conditions modulate the protein content and immunomodulatory activity of extracellular vesicles produced by the probiotic Propionibacterium freudenreichii. Appl Environ Microbiol. 2021;87(4):e02263–20. doi:10.1128/AEM.02263-20
16. Liu Y, Liu Q, Zhao L, et al. Essential role of membrane vesicles for biological activity of the bacteriocin micrococcin P1. J Extracell Vesicles. 2022;11(4):e12212. doi:10.1002/jev2.12212
17. Chernov VM, Chernova OA, Markelova MI, et al. Extracellular vesicles of probiotics: from structural and functional characteristics to practical application. Nanobiotechnol Rep. 2024;19(6):887–900. doi:10.1134/s2635167624600792
18. Chelakkot C, Choi Y, Kim DK, et al. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp Mol Med. 2018;50(2):e450. doi:10.1038/emm.2017.282
19. Rodovalho VR, da Luz B, Rabah H, et al. Extracellular vesicles produced by the probiotic propionibacterium freudenreichii CIRM-BIA 129 mitigate inflammation by modulating the NF-κB pathway. Front Microbiol. 2020;11:1544. doi:10.3389/fmicb.2020.01544
20. Li J, Shi M, Wang Y, et al. Probiotic-derived extracellular vesicles alleviate AFB1-induced intestinal injury by modulating the gut microbiota and AHR activation. J Nanobiotechnology. 2024;22(1):697. doi:10.1186/s12951-024-02979-3
21. Zhou H, Huang W, Li J, et al. Oral probiotic extracellular vesicle therapy mitigates Influenza A Virus infection via blunting IL-17 signaling. Bioact Mater. 2025;45:401–416. doi:10.1016/j.bioactmat.2024.11.016
22. Li Z, Li M, Fang X, et al. Dietary Lactobacillus johnsonii-derived extracellular vesicles ameliorate acute colitis by regulating gut microbiota and maintaining intestinal barrier homeostasis. Food Funct. 2024;15(23):11757–11779. doi:10.1039/d4fo04194a
23. Alvarez CS, Badia J, Bosch M, et al. Outer membrane vesicles and soluble factors released by probiotic Escherichia coli Nissle 1917 and commensal ECOR63 enhance barrier function by regulating expression of tight junction proteins in intestinal epithelial cells. Front Microbiol. 2016;7:1981. doi:10.3389/fmicb.2016.01981
24. Busby LB, Hoxha K, Oakes W, et al. Identification of small regulatory RNA transcripts in extracellular vesicles from lactic acid bacteria. FASEB J. 2022;36. doi:10.1096/fasebj.2022.36.s1.r5232
25. Balsalobre C, Silván JM, Berglund S, et al. Release of the type I secreted alpha-haemolysin via outer membrane vesicles from Escherichia coli. Mol Microbiol. 2006;59(1):99–112. doi:10.1111/j.1365-2958.2005.04938.x
26. Liu Q, Yi J, Liang K, et al. Outer membrane vesicles derived from Salmonella enteritidis protect against the virulent wild-type strain infection in a mouse model. J Microbiol Biotechnol. 2017;27(8):1519–1528. doi:10.4014/jmb.1705.05028
27. Buchacher T, Digruber A, Kranzler M, et al. Bacillus cereus extracellular vesicles act as shuttles for biologically active multicomponent enterotoxins. Cell Commun Signal. 2023;21(1):112. doi:10.1186/s12964-023-01132-1
28. Pérez-Cruz C, Delgado L, López-Iglesias C, et al. Outer-inner membrane vesicles naturally secreted by gram-negative pathogenic bacteria. PLoS One. 2015;10(1):e0116896. doi:10.1371/journal.pone.0116896
29. Catalan EA, Seguel-Fuentes E, Fuentes B, et al. Oral pathobiont-derived outer membrane vesicles in the oral-gut axis. Int J Mol Sci. 2024;25(20):11141. doi:10.3390/ijms252011141
30. Jain H, Kumar A, Almousa S, et al. Characterisation of LPS+ bacterial extracellular vesicles along the gut-hepatic portal vein-liver axis. J Extracell Vesicles. 2024;13(7):e12474. doi:10.1002/jev2.12474
31. Wen Y, Chen S, Jiang Z, et al. Dysregulated haemolysin promotes bacterial outer membrane vesicles-induced pyroptotic-like cell death in zebrafish. Cell Microbiol. 2019;21(6):e13010. doi:10.1111/cmi.13010
32. David L, Taieb F, Pénary M, et al. Outer membrane vesicles produced by pathogenic strains of Escherichia coli block autophagic flux and exacerbate inflammasome activation. Autophagy. 2022;18(12):2913–2925. doi:10.1080/15548627.2022.2054040
33. Liang L, Yang C, Liu L, et al. Commensal bacteria-derived extracellular vesicles suppress ulcerative colitis through regulating the macrophages polarization and remodeling the gut microbiota. Microb Cell Fact. 2022;21(1):88. doi:10.1186/s12934-022-01812-6
34. Mandelbaum N, Zhang L, Carasso S, et al. Extracellular vesicles of the Gram-positive gut symbiont Bifidobacterium longum induce immune-modulatory, anti-inflammatory effects. NPJ Biofilms Microbiomes. 2023;9(1):30. doi:10.1038/s41522-023-00400-9
35. Zheng C, Zhong Y, Xie J, et al. Bacteroides acidifaciens and its derived extracellular vesicles improve DSS-induced colitis. Front Microbiol. 2023;14:1304232. doi:10.3389/fmicb.2023.1304232
36. Liu X, Guo X, Su X, et al. Extracellular vehicles from commensal skin Malassezia restricta inhibit Staphylococcus aureus proliferation and biofilm formation. ACS Infect Dis. 2024;10(2):624–637. doi:10.1021/acsinfecdis.3c00511
37. Xing J, Niu T, Yu T, et al. Faecalibacterium prausnitzii-derived outer membrane vesicles reprogram gut microbiota metabolism to alleviate Porcine Epidemic Diarrhea Virus infection. Microbiome. 2025;13(1):90. doi:10.1186/s40168-025-02078-x
38. Shagaleeva OY, Kashatnikova DA, Kardonsky DA, et al. Bacteroides vesicles promote functional alterations in the gut microbiota composition. Microbiol Spectr. 2024;12(11):e0063624. doi:10.1128/spectrum.00636-24
39. Preet R, Islam MA, Shim J, et al. Gut commensal Bifidobacterium-derived extracellular vesicles modulate the therapeutic effects of anti-PD-1 in lung cancer. Nat Commun. 2025;16(1):3500. doi:10.1038/s41467-025-58553-4
40. Zheng T, Hao H, Liu Q, et al. Effect of extracelluar vesicles derived from Akkermansia muciniphila on intestinal barrier in colitis mice. Nutrients. 2023;15(22):4722. doi:10.3390/nu15224722
41. Chin WL, Lee BH, Hsu QY, et al. Small intestine-residing probiotics suppress neurotoxic bile acid production via extracellular vesicle-mediated inhibition of Clostridium scindens. Food Res Int. 2025;207:116049. doi:10.1016/j.foodres.2025.116049
42. Nishiyama K, Takaki T, Sugiyama M, et al. Extracellular vesicles produced by Bifidobacterium longum export mucin-binding proteins. Appl Environ Microbiol. 2020;86(19):e01464–20. doi:10.1128/AEM.01464-20
43. Roier S, Zingl FG, Cakar F, et al. A novel mechanism for the biogenesis of outer membrane vesicles in Gram-negative bacteria. Nat Commun. 2016;7:10515. doi:10.1038/ncomms10515
44. Bose S, Aggarwal S, Singh DV, et al. Extracellular vesicles: an emerging platform in gram-positive bacteria. Microb Cell. 2020;7(12):312–322. doi:10.15698/mic2020.12.737
45. Dauros-Singorenko P, Hong J, Swift S, et al. Effect of the extracellular vesicle RNA cargo from uropathogenic Escherichia coli on bladder cells. Front Mol Biosci. 2020;7:580913. doi:10.3389/fmolb.2020.580913
46. Dell’Annunziata F, Folliero V, Giugliano R, et al. Gene transfer potential of outer membrane vesicles of gram-negative bacteria. Int J Mol Sci. 2021;22(11):5985. doi:10.3390/ijms22115985
47. Abolhasani FS, Vaghefinanekaran N, Yarahmadi A, et al. Outer membrane vesicles in gram-negative bacteria and its correlation with pathogenesis. Front Immunol. 2025;16:1541636. doi:10.3389/fimmu.2025.1541636
48. Fonseca S, Carvalho AL, Miquel-Clopés A, et al. Extracellular vesicles produced by the human gut commensal bacterium Bacteroides thetaiotaomicron elicit anti-inflammatory responses from innate immune cells. Front Microbiol. 2022;13:1050271. doi:10.3389/fmicb.2022.1050271
49. Yousuf B, Mottawea W, Esmail GA, et al. Multi-omics unveils strain-specific neuroactive metabolite production linked to inflammation modulation by Bacteroides and their extracellular vesicles. Curr Res Microb Sci. 2025;8:100358. doi:10.1016/j.crmicr.2025.100358
50. Nakayama-Imaohji H, Hirota K, Yamasaki H, et al. DNA inversion regulates outer membrane vesicle production in Bacteroides fragilis. PLoS One. 2016;11(2):e0148887. doi:10.1371/journal.pone.0148887
51. Hampton CM, Guerrero-Ferreira RC, Storms RE, et al. The opportunistic pathogen Vibrio vulnificus produces outer membrane vesicles in a spatially distinct manner related to capsular polysaccharide. Front Microbiol. 2017;8:2177. doi:10.3389/fmicb.2017.02177
52. Zhou G, Zhou Q, Li R, et al. Synthetically engineered bacterial extracellular vesicles and IL-4-encapsulated hydrogels sequentially promote osteoporotic fracture repair. ACS Nano. 2025;19(16):16064–16083. doi:10.1021/acsnano.5c03106
53. Jeong GJ, Khan F, Tabassum N, et al. Bacterial extracellular vesicles: modulation of biofilm and virulence properties. Acta Biomater. 2024;178:13–23. doi:10.1016/j.actbio.2024.02.029
54. Yang J, Hwang I, Lee E, et al. Bacterial outer membrane vesicle-mediated cytosolic delivery of flagellin triggers host NLRC4 canonical inflammasome signaling. Front Immunol. 2020;11:581165. doi:10.3389/fimmu.2020.581165
55. Kurdy W, Zelenikhin PV, Yakovleva GY, et al. The proteome of extracellular membrane vesicles from Bacillus pumilus 3–19. Mol Biol. 2024;58(1):178–186. doi:10.1134/s0026893324010059
56. Zhao Y, Jiang B, Zuo S. Cucurbitacin iia alleviates colitis via promoting the release of host-derived extracellular vesicles encapsulating microRNA-30b-5p. J Inflamm Res. 2025;18:1447–1458. doi:10.2147/JIR.S500722
57. Kameli N, Becker H, Jonkers DM, et al. Investigating the immunomodulatory impact of fecal bacterial membrane vesicles and their IgA coating patterns in Crohn’s disease patients. Int J Mol Sci. 2024;25(23):13194. doi:10.3390/ijms252313194
58. Olovo CV, Wiredu Ocansey DK, Ji Y, et al. Bacterial membrane vesicles in the pathogenesis and treatment of inflammatory bowel disease. Gut Microbes. 2024;16(1):2341670. doi:10.3390/ijms252313194
59. Heo M, Park YS, Yoon H, et al. Potential of gut microbe-derived extracellular vesicles to differentiate inflammatory bowel disease patients from healthy controls. Gut Liver. 2023;17(1):108–118. doi:10.5009/gnl220081
60. Vanaja SK, Russo AJ, Behl B, et al. Bacterial outer membrane vesicles mediate cytosolic localization of LPS and Caspase-11 activation. Cell. 2016;165(5):1106–1119. doi:10.1016/j.cell.2016.04.015
61. Díez-Sainz E, Milagro FI, Riezu-Boj JI, et al. Effects of gut microbiota-derived extracellular vesicles on obesity and diabetes and their potential modulation through diet. J Physiol Biochem. 2022;78(2):485–499. doi:10.1007/s13105-021-00837-6
62. Almousa S, Kim S, Kumar A, et al. Bacterial nanovesicles as interkingdom signaling moieties mediating pain hypersensitivity. ACS Nano. 2025;19(3):3210–3225. doi:10.1021/acsnano.4c10529
63. Villard A, Boursier J, Andriantsitohaina R. Bacterial and eukaryotic extracellular vesicles and nonalcoholic fatty liver disease: new players in the gut-liver axis. Am J Physiol Gastrointest Liver Physiol. 2021;320(4):G485–G495. doi:10.1152/ajpgi.00362.2020
64. Hirayama S, Nakao R. Glycine significantly enhances bacterial membrane vesicle production: a powerful approach for isolation of LPS-reduced membrane vesicles of probiotic Escherichia coli. Microb Biotechnol. 2020;13(4):1162–1178. doi:10.1111/1751-7915.13572
65. Schwechheimer C, Kuehn MJ. Outer-membrane vesicles from Gram-negative bacteria: biogenesis and functions. Nat Rev Microbiol. 2015;13(10):605–619. doi:10.1155/2021/1490732
66. Macion A, Wyszyńska A, Godlewska R. Delivery of toxins and effectors by bacterial membrane vesicles. Toxins. 2021;13(12):845. doi:10.3390/toxins13120845
67. Tiku V, Tan MW. Host immunity and cellular responses to bacterial outer membrane vesicles. Trends Immunol. 2021;42(11):1024–1036. doi:10.1016/j.it.2021.09.006
68. Lyons S, Wang L, Casanova JE, et al. Salmonella typhimurium transcytoses flagellin via an SPI2-mediated vesicular transport pathway. J Cell Sci. 2004;117(Pt 24):5771–5780. doi:10.1242/jcs.01500
69. Gilmore WJ, Johnston EL, Bitto NJ, et al. Bacteroides fragilis outer membrane vesicles preferentially activate innate immune receptors compared to their parent bacteria. Front Immunol. 2022;13:970725. doi:10.3389/fimmu.2022.970725
70. Kim DJ, Yang J, Seo H, et al. Colorectal cancer diagnostic model utilizing metagenomic and metabolomic data of stool microbial extracellular vesicles. Sci Rep. 2020;10(1):2860. doi:10.1038/s41598-020-59529-8
71. Sundaram K, Teng Y, Mu J, et al. Outer membrane vesicles released from Garlic Exosome-like Nanoparticles (GaELNs) train gut bacteria that reverses type 2 diabetes via the gut-brain axis. Small. 2024;20(20):e2308680. doi:10.1002/smll.202308680
72. Abreo Medina A, Shi M, Wang Y, et al. Exploring extracellular vesicles: a novel approach in nonalcoholic fatty liver disease. J Agric Food Chem. 2025;73(5):2717–2731. doi:10.1021/acs.jafc.4c09209
73. Prince A, Tiwari A, Mandal T, et al. Lipid specificity of the fusion of bacterial extracellular vesicles with the host membrane. J Phys Chem B. 2024;128(34):8116–8130. doi:10.1021/acs.jpcb.4c02321
74. Cañas MA, Giménez R, Fábrega MJ, et al. Outer membrane vesicles from the probiotic Escherichia coli Nissle 1917 and the Commensal ECOR12 enter intestinal epithelial cells via clathrin-dependent endocytosis and elicit differential effects on DNA damage. PLoS One. 2016;11(8):e0160374. doi:10.1371/journal.pone.0160374
75. Tsuda K, Furuta N, Inaba H, et al. Functional analysis of alpha5beta1 integrin and lipid rafts in invasion of epithelial cells by Porphyromonas gingivalis using fluorescent beads coated with bacterial membrane vesicles. Cell Struct Funct. 2008;33(1):123–132. doi:10.1247/csf.08012
76. Johnston EL, Heras B, Kufer TA, et al. Detection of Bacterial Membrane Vesicles by NOD-Like Receptors. Int J Mol Sci. 2021;22(3):1005. doi:10.3390/ijms22031005
77. Zheng C, Zhong Y, Zhang W, et al. Chlorogenic acid ameliorates post-infectious irritable bowel syndrome by regulating extracellular vesicles of gut microbes. Adv Sci. 2023;10(28):e2302798. doi:10.1002/advs.202302798
78. Khalyfa A, Poroyko VA, Qiao Z, et al. Exosomes and metabolic function in mice exposed to alternating dark-light cycles mimicking night shift work schedules. Front Physiol. 2017;8:882. doi:10.3389/fphys.2017.00882
79. Choi Y, Kwon Y, Kim DK, et al. Gut microbe-derived extracellular vesicles induce insulin resistance, thereby impairing glucose metabolism in skeletal muscle. Sci Rep. 2015;5:15878. doi:10.1038/srep15878
80. Kumar A, Sundaram K, Mu J, et al. High-fat diet-induced upregulation of exosomal phosphatidylcholine contributes to insulin resistance. Nat Commun. 2021;12(1):213. doi:10.1038/s41467-020-20500-w
81. Mirjafari Tafti ZS, Moshiri A, Ettehad Marvasti F, et al. The effect of saturated and unsaturated fatty acids on the production of outer membrane vesicles from Bacteroides fragilis and Bacteroides thetaiotaomicron. Gastroenterol Hepatol Bed Bench. 2019;12(2):155–162.
82. Liu J, Zhang Y, Tian Y, et al. Integrative biology of extracellular vesicles in diabetes mellitus and diabetic complications. Theranostics. 2022;12(3):1342–1372. doi:10.7150/thno.65778
83. Wang Y, Luo X, Xiang X, et al. Roles of bacterial extracellular vesicles in systemic diseases. Front Microbiol. 2023;14:1258860. doi:10.3389/fmicb.2023.1258860
84. Ashrafian F, Keshavarz Azizi Raftar S, Lari A, et al. Extracellular vesicles and pasteurized cells derived from Akkermansia muciniphila protect against high-fat induced obesity in mice. Microb Cell Fact. 2021;20(1):219. doi:10.1186/s12934-021-01709-w
85. Maccioni L, Gao B, Leclercq S, et al. Intestinal permeability, microbial translocation, changes in duodenal and fecal microbiota, and their associations with alcoholic liver disease progression in humans. Gut Microbes. 2020;12(1):1782157. doi:10.1080/19490976.2020.1782157
86. Ashrafian F, Shahriary A, Behrouzi A, et al. Akkermansia muciniphila-derived extracellular vesicles as a mucosal delivery vector for amelioration of obesity in mice. Front Microbiol. 2019;10:2155. doi:10.3389/fmicb.2019.02155
87. Nie Y, Xie XQ, Zhou L, et al. Desulfovibrio fairfieldensis-derived outer membrane vesicles damage epithelial barrier and induce inflammation and pyroptosis in macrophages. Cells. 2022;12(1):89. doi:10.3390/cells12010089
88. Luo M, Li S, Yang Y, et al. Effects of salmonella outer membrane vesicles on intestinal microbiota and intestinal barrier function. Foodborne Pathog Dis. 2024;21(4):257–267. doi:10.1089/fpd.2023.0096
89. Wu Z, Song M, Fan J, et al. Maternal Lactobacillus amylovorus-derived extracellular vesicles modulate M2 macrophage polarization to promote early-life gut development in offspring. Chem Eng J. 2025;512:162704. doi:10.1016/j.cej.2025.162704
90. Shi J, Ma D, Gao S, et al. Probiotic Escherichia coli Nissle 1917-derived outer membrane vesicles modulate the intestinal microbiome and host gut-liver metabolome in obese and diabetic mice. Front Microbiol. 2023;14:1219763. doi:10.3389/fmicb.2023.1219763
91. Nah G, Park SC, Kim K, et al. Type-2 diabetics reduces spatial variation of microbiome based on extracellur vesicles from gut microbes across human body. Sci Rep. 2019;9(1):20136. doi:10.1038/s41598-019-56662-x
92. Villard A, Boursier J, Andriantsitohaina R. Microbiota-derived extracellular vesicles and metabolic syndrome. Acta Physiol. 2021;231(4):e13600. doi:10.1111/apha.13600
93. Huh YJ, Seo JY, Nam J, et al. Bariatric/metabolic surgery induces noticeable changes of microbiota and their secreting extracellular vesicle composition in the gut. Obes Surg. 2019;29(8):2470–2484. doi:10.1007/s11695-019-03852-1
94. Chen PP, Zhang JX, Li XQ, et al. Outer membrane vesicles derived from gut microbiota mediate tubulointerstitial inflammation: a potential new mechanism for diabetic kidney disease. Theranostics. 2023;13(12):3988–4003. doi:10.7150/thno.84650
95. Saad MJA, Prada PO, Santos A. Linking gut microbiota and inflammation to obesity and insulin resistance. Physiology. 2016;31(4):283–293. doi:10.1152/physiol.00041.2015
96. Qin LN, Yu YF, Ma L, et al. Intestinal bacteria-derived extracellular vesicles in metabolic dysfunction-associated steatotic liver disease: from mechanisms to therapeutics. Mol Cells. 2025;48(6):100216. doi:10.1016/j.mocell.2025.100216
97. Zhang-Sun W, Tercé F, Burcelin R, et al. Structure function relationships in three lipids A from the Ralstonia genus rising in obese patients. Biochimie. 2019;159:72–80. doi:10.1016/j.biochi.2019.01.015
98. Hu X, Wu Q, Huang L, et al. Clinical efficacy of washed microbiota transplantation on metabolic syndrome and metabolic profile of donor outer membrane vesicles. Front Nutr. 2024;11:1465499. doi:10.3389/fnut.2024.1465499
99. Tang W, Ni Z, Wei Y, et al. Extracellular vesicles of Bacteroides uniformis induce M1 macrophage polarization and aggravate gut inflammation during weaning. Mucosal Immunol. 2024;17(5):793–809. doi:10.1016/j.mucimm.2024.05.004
100. Diaz-Garrido N, Badia J, Baldomà L. Modulation of dendritic cells by microbiota extracellular vesicles influences the cytokine profile and exosome cargo. Nutrients. 2022;14(2):344. doi:10.3390/nu14020344
101. Xue L, He J, Gao N, et al. Probiotics may delay the progression of nonalcoholic fatty liver disease by restoring the gut microbiota structure and improving intestinal endotoxemia. Sci Rep. 2017;7:45176. doi:10.1038/srep45176
102. Li L, Li R, Zhu R, et al. Salvianolic acid B prevents body weight gain and regulates gut microbiota and LPS/TLR4 signaling pathway in high-fat diet-induced obese mice. Food Funct. 2020;11(10):8743–8756. doi:10.1039/d0fo01116a
103. Chen J, Wei C, Pan C, et al. Genetically engineered Bacillus subtilis-derived extracellular vesicles Alleviates Ulcerative colitis by restoring intestinal barrier and regulating gut microbiota. Chem Eng J. 2025;512:162727. doi:10.1016/j.cej.2025.162727
104. Seyama M, Yoshida K, Yoshida K, et al. Outer membrane vesicles of Porphyromonas gingivalis attenuate insulin sensitivity by delivering gingipains to the liver. Biochim Biophys Acta Mol Basis Dis. 2020;1866(6):165731. doi:10.1016/j.bbadis.2020.165731
105. Gill PA, Inniss S, Kumagai T, et al. The role of diet and gut microbiota in regulating gastrointestinal and inflammatory disease. Front Immunol. 2022;13:866059. doi:10.3389/fimmu.2022.866059
106. Martin-Gallausiaux C, Marinelli L, Blottière HM, et al. SCFA: mechanisms and functional importance in the gut. Proc Nutr Soc. 2021;80(1):37–49. doi:10.1017/S0029665120006916
107. Uemura T, Kawashima A, Jingushi K, et al. Bacterial information in serum extracellular vesicles reflects the inflammation of adherent perinephric fat. Cancer Sci. 2025;116(2):338–349. doi:10.1111/cas.16410
108. Zhu S, Xu W, Chen Z, et al. Nuciferine exerts anti-inflammatory effects in mice with non-alcoholic steatohepatitis by regulating the miR-23a-3p-SIRT1-NF-κB pathway and Akkermansia muciniphila-extracellular vesicles. Phytother Res. 2025;39(8):3508–3525. doi:10.1002/ptr.8528
109. Pei J, Zhang C, Zhang Q, et al. Probiotics alleviate chronic ethanol exposure-induced anxiety-like behavior and hippocampal neuroinflammation in male mice through gut microbiota-derived extracellular vesicles. J Nanobiotechnology. 2024;22(1):730. doi:10.1186/s12951-024-03017-y
110. Wang K, Li X, Huang K, et al. Dietary probiotic-derived extracellular vesicles as delivery systems of bioactive compounds to maintain intestinal homeostasis. Chem Eng J. 2025;505:159546. doi:10.1016/j.cej.2025.159546
111. Zhu Y, Zhang X, Huang W, et al. Protective Effect of Enterococcus faecium Against Alcohol-Induced Acute Liver Injury Via Extracellular Vesicles in Rats. Foodborne Pathog Dis. 2025. doi:10.1089/fpd.2025.0005
112. Jian H, Liu Y, Wang X, et al. Akkermansia muciniphila as a next-generation probiotic in modulating human metabolic homeostasis and disease progression: a role mediated by gut-liver-brain axes. Int J Mol Sci. 2023;24(4):3900. doi:10.3390/ijms24043900
113. Zu M, Liu G, Xu H, et al. Extracellular Vesicles from Nanomedicine-Trained Intestinal Microbiota Substitute for Fecal Microbiota Transplant in Treating Ulcerative Colitis. Adv Mater. 2024;36(41):e2409138. doi:10.1002/adma.202409138
114. Tan J, Taitz JJ, Ni D, et al. Gut microbiota-derived extracellular vesicles exhibit diurnal regulation and activate hepatic gluconeogenesis. Mol Metab. 2025;98:102180. doi:10.1016/j.molmet.2025.102180
115. Tan J, Ni D, Taitz J, et al. Dietary protein increases T-cell-independent sIgA production through changes in gut microbiota-derived extracellular vesicles. Nat Commun. 2022;13(1):4336. doi:10.1038/s41467-022-31761-y
116. Shin C, Baik I. Bacterial extracellular vesicle composition in human urine and the 10-year risk of abdominal obesity. Metab Syndr Relat Disord. 2023;21(5):233–242. doi:10.1089/met.2022.0109
117. Park KS, Lee J, Lee C, et al. Sepsis-like systemic inflammation induced by nano-sized extracellular vesicles from feces. Front Microbiol. 2018;9:1735. doi:10.3389/fmicb.2018.01735
118. Kameli N, Becker H, Welbers T, et al. Metagenomic profiling of fecal-derived bacterial membrane vesicles in Crohn’s disease patients. Cells. 2021;10(10):2795. doi:10.3390/cells10102795
119. Kameli N, Borman R, Lpez-Iglesias C, et al. Characterization of feces-derived bacterial membrane vesicles and the impact of their origin on the inflammatory response. Front Cell Infect Microbiol. 2021;11:667987. doi:10.3389/fcimb.2021.667987
120. Zhang Z, Liu X, Yang X, et al. Identification of faecal extracellular vesicles as novel biomarkers for the non-invasive diagnosis and prognosis of colorectal cancer. J Extracell Vesicles. 2023;12(1):e12300. doi:10.1002/jev2.12300
121. Rodríguez-Díaz C, Martín-Reyes F, Taminiau B, et al. The metagenomic composition and effects of fecal-microbe-derived extracellular vesicles on intestinal permeability depend on the Patient’s disease. Int J Mol Sci. 2023;24(5):4971. doi:10.3390/ijms24054971
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.