Back to Journals » Vascular Health and Risk Management » Volume 17
Characterizing and Profiling microRNAs in Dogs Undergoing Induced Ischemic Brain Stroke After Middle Cerebral Artery Occlusion Under Fluoroscopic Guidance
Authors Alawneh KZ ![]() , Raffee La
, Raffee La ![]() , Alshehabat MAM, Haddad H
, Alshehabat MAM, Haddad H ![]() , Jaradat SA
, Jaradat SA ![]()
Received 17 May 2021
Accepted for publication 27 August 2021
Published 8 September 2021 Volume 2021:17 Pages 543—550
DOI https://doi.org/10.2147/VHRM.S317861
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Harry Struijker-Boudier
Khaled Z Alawneh,1 Liqaa A Raffee,2 Musa Ahmed Mohammed Alshehabat,3 Hazem Haddad,4 Saied A Jaradat5
1Department of Diagnostic Radiology and Nuclear Medicine, Faculty of Medicine, Jordan University of Science and Technology, Irbid, Jordan; 2Department of Accident and Emergency Medicine, Faculty of Medicine, Jordan University of Science and Technology, Irbid, Jordan; 3Department of Clinical Veterinary Medical Sciences, Faculty of Veterinary Medicine, Jordan University of Science and Technology, Irbid, Jordan; 4Haya Biotechnology Center, Jordan University of Science and Technology, Irbid, 22110, Jordan; 5Princess Haya Biotechnology Center, Jordan University of Science and Technology, Irbid, 22110, Jordan
Correspondence: Liqaa A Raffee
Department of Accident and Emergency Medicine, Faculty of Medicine, Jordan University of Science and Technology, Irbid, Jordan
Email [email protected]
Purpose: Ischemic stroke of the brain is still considered one of the most common causes of disability and death in developed and developing countries in human beings despite advances in medicine and technology. This study was conducted to characterize and profile tens of induced biomarkers (microRNAs) after experimentally inducing regional ischemic stroke of the brain by occluding the middle cerebral artery under fluoroscopic guidance using an autologous blood clot.
Patient and Methods: A total of six healthy dogs were recruited for this study. The microRNAs were profiled in the blood and urine before and after occluding the middle cerebral artery using genetic techniques.
Results: The very highly expressed genes were comprised within cluster A, followed by cluster D in both 24 and 48-hour brain samples. Clusters B and C revealed down-regulated genes, while miRNAs remained up-regulated in the 24-hour samples merely in cluster F. Upregulated genes at 48 hours of reperfusion were included in cluster E. On the other hand, changes were observed after a day on the cluster G genes. Exclusive upregulation was notified after 2 days due to the changes in mIR-138. The normalized gene expression in the test sample is witnessed through Fold-Change, which divides the control sample’s normalized gene expression. Moreover, fold-change has emerged as a significant approach for representing fold-regulation.
Conclusion: The microRNAs expression in blood and urine may have a potential role in the diagnosis, prognosis, and assessment of therapy associated with cerebral artery occlusion under fluoroscopic guidance.
Keywords: dogs, fluoroscopic induced ischemic brain, microRNAs, middle cerebral artery stroke
Introduction
Ischemic stroke of the brain is considered one of the challenging causes of short- or long-term neurological disability and one of the most frequent causes of mortality in human medicine worldwide. The most commonly implicated ischemic stroke in human medicine includes embolic or thrombotic obstruction of major cerebral arteries, such as the middle cerebral artery.1,2 It has been estimated in 2008 that ischemic stroke did account for an estimated cost of over 65 billion USD. The confirmed incidence of stroke has been estimated to be approximately about 10% and this is on the rise due to many factors. In veterinary medicine, the incidence and prevalence rates are undocumented, but this disease identity is becoming more recognized with advanced development and widespread availability of more advanced diagnostic imaging techniques, such as contrast magnetic resonance imaging.
The exact etiology and pathogenesis of brain stroke are still not fully understood. There are many risk factors associated with stroke, including age, hypertension, diabetes mellitus, and obesity. The clinical manifestation and variability of clinical signs associated with ischemic stroke of the cerebrum depends on many factors but largely on the location of ischemia, the severity of the ischemia, complexity of the ischemia, age, presence of cancer, and potentially concurrent diseases.1,3–6
This study has been proposed to induce ischemic stroke of the cerebrum by selectively and mechanically occluding the middle cerebral artery (MCA). Selective occlusion of the MCA has been suggested to induce focal-type ischemia.7 This model mimics the pathophysiology of ischemic stroke in humans. Occlusion of the MCA was performed using autologous blood clots under aseptic technique through the right femoral arterial access under fluoroscopy guidance. Fluoroscopy is a real-time dynamic X-ray machine that will help in navigating the location of the MCA, thus allowing to occlude the described artery.
A group of regulatory aspects is widely represented through microRNAs for facilitating cells to furnish multifaceted gene expression cascades in biological procedures.8 MicroRNAs have been found in many tissues, including blood and body fluids, such as cerebral spinal fluids, in stable forms that are protected from endogenous RNase activity. Characterizing and profiling microRNAs is considered an innovative technique to identify biomarkers associated with many diseases. For instance, microRNAs (miR 206, miR 574-5p) have been proven to be a powerful tool in the diagnosis and treatment of many cardiac diseases including coronary artery diseases.8 Researchers have pinpointed other significant microRNAs (miR-125b-2*−27a*miR-210, −422a, −488, and −627) that were considered to be of significant values in patients with cardiovascular and ischemic conditions.
Here, the main aim is to characterize blood microRNAs before and after experimental induction of cerebral ischemia. The secondary objectives of this study include profiling urine microRNAs in dogs undergoing experimental induction of cerebral stroke by occluding the middle cerebral artery under fluoroscopic guidance.
To the authors' knowledge, this is considered the first publishable research effort to experimentally induce acute cerebral ischemia in dogs by characterizing and profiling potential microRNAs biomarkers in blood and urine and concurrently measuring cardiac troponin in this group of dogs. Knowledge of the biomarkers that can potentially be of clinical significance in patients with acute cerebral ischemia can be a novel approach to accurately utilize in diagnosis and evaluate the effectiveness of treatment, thereby lowering mortality and morbidity in such group of patients.
Patient and Methods
in veterinary care were followed, and that the client/owner provided informed consent for their pet/animal to be used in this study. A total of 6 healthy middle-aged dogs (1–5 years) were recruited for this study. The dogs were obtained from local breeders. Dogs underwent complete physical examination, complete blood count (platelet counts, white and red blood cell counts), serum biochemistry panel (urea, creatinine, liver enzymes [ALT, ALP, GGT], glucose, albumin, globulin) to evaluate the function of internal organs to ensure general health status. Only healthy dogs were included in this study. Dogs with historical or clinical signs suggestive of internal organ dysfunction were excluded from the study. Obese and aggressive dogs were not considered for this project. Selected animals were vaccinated against canine distemper, adenovirus, parvovirus, Leptospira species and rabies [Vanguard® and Rabies® and Leptospira] at the time of admission to the hosting facility. In addition, a deworming regimen using Ascaten® (1 tab/5kg) was started and repeated in 2 weeks. Dogs were housed at the hosting facility for 14 days before the start of the experimental design. Each dog was given Collar ID and was housed separately in cages and provided with daily dry food, and ad-lib source of water. The dogs were fed commercially available food (Benson™). The main purpose of this pre-experimental housing was to give ample time for dogs to adjust and acclimatize to the new environment and to observe any developing abnormally physical or behavioral manifestation that could not be identified at the time of selection.
Selected dogs were anesthetized using intravenous Propofol (4–6 mg/kg). Selected dogs underwent arterial catheterization of the right femoral artery under strict aseptic technique and under fluoroscopic guidance to navigate around to selectively occlude the middle cerebral artery. Measuring microRNAs and data analysis: Assay Kits were used to purify microRNA and total RNA from the collected urine and serum samples according to the manufacturer’s instructions and were stored aliquoted in RNase/DNase-free water at –80°C. The total RNA and microRNA quality and quantity were checked using both bio-analyzer 2100 (Agilent) and Nano-DropTM 1000 Spectrophotometer (Thermo Scientific). The TaqMan Human MicroRNA Assay Set (Applied Biosystems) consisting of Sanger database unique assays to quantify Stroke microRNAs and 10 controls (Z30 and nine different SNORs/RNUs) were used to screen a set of 20 strokes and 20 normal specimens for differentially expressed microRNAs.
The reliance of TaqMan MicroRNA Reverse Transcription assay is on a particular hairpin structure throughout cDNA synthesis and mature microRNA particular detection probes. These aspects preclude the identification of precursor microRNAs. To be precise, pooled analysis for different RT primers was done based on the suggestions given for 30 min in a speed vacuum centrifuge and resuspended for an ultimate concentration of 50 nM each. The final volume was reverse-transcribed through 25 to 50 ng of total RNA sample aliquots using the TaqMan MicroRNA Reverse Transcription Kit.
Blood was collected from the jugular or cephalic vein to profile the microRNAs. Descriptive statistics were performed for each of the listed variables. Data were analyzed with one-way ANOVA, using GraphPad Prism 5.0 software (GraphPad Prism, San Diego, CA) and GenEx software (www.multid.se) or REST V3. The significant difference was set at a P value less than 0.05. Relative quantification of microRNA expression was calculated using the comparative Ct method (Applied Biosystems User Bulletin N2 (P/N 4303859)). For proper comparison of microRNA, expression data were normalized with appropriate reference genes using geNorm algorithms. The microRNAs were serially evaluated: one analysis before the arterial collusion and 3 analyses post occlusions (1-hour post, 4 hours post, 1-day s post).
The project has no special safety considerations. Dogs determined to fulfill the criteria of the study were housed at the Veterinary Health Center at Jordan University of Science and Technology. Individual animal subjects were housed in separate and clean cages with free access to water and a standard feeding program. Dogs were fully vaccinated to eliminate any potential zoonotic hazards. A portable fluoroscopy machine was housed in the lead radiology room for the study duration at the veterinary teaching hospital. Urine samples were collected regarding microRNA.
Results
A total of 6 middle-aged dogs were screened based on brain and blood samples subjected to microRNA. Very few microRNA transcripts were identified in the urine and blood, and the majority of the transcripts were identified in the brains of dogs for 24 and 48 hours, respectively. Hierarchical clustering of the microRNAs identified in the brain revealed 7 different clusters. The very highly expressed genes were comprised within cluster A, followed by cluster D in both 24 and 48-hour brain samples. Clusters B and C revealed the down-regulated genes while miRNAs remained up-regulated in the 24-hour samples merely in cluster F. Upregulated genes at 48 hours of reperfusion were included in cluster E. On the other hand, changes were observed after a day on the cluster G genes. Exclusive upregulation was notified after 2 days due to the changes in mIR-138.
Tables 1 and 2 show the list of genes and plate-position selected for both groups in serum and urine profile, respectively. The selection was made after elevation in serum to increase expression from 2 h to 24 h.
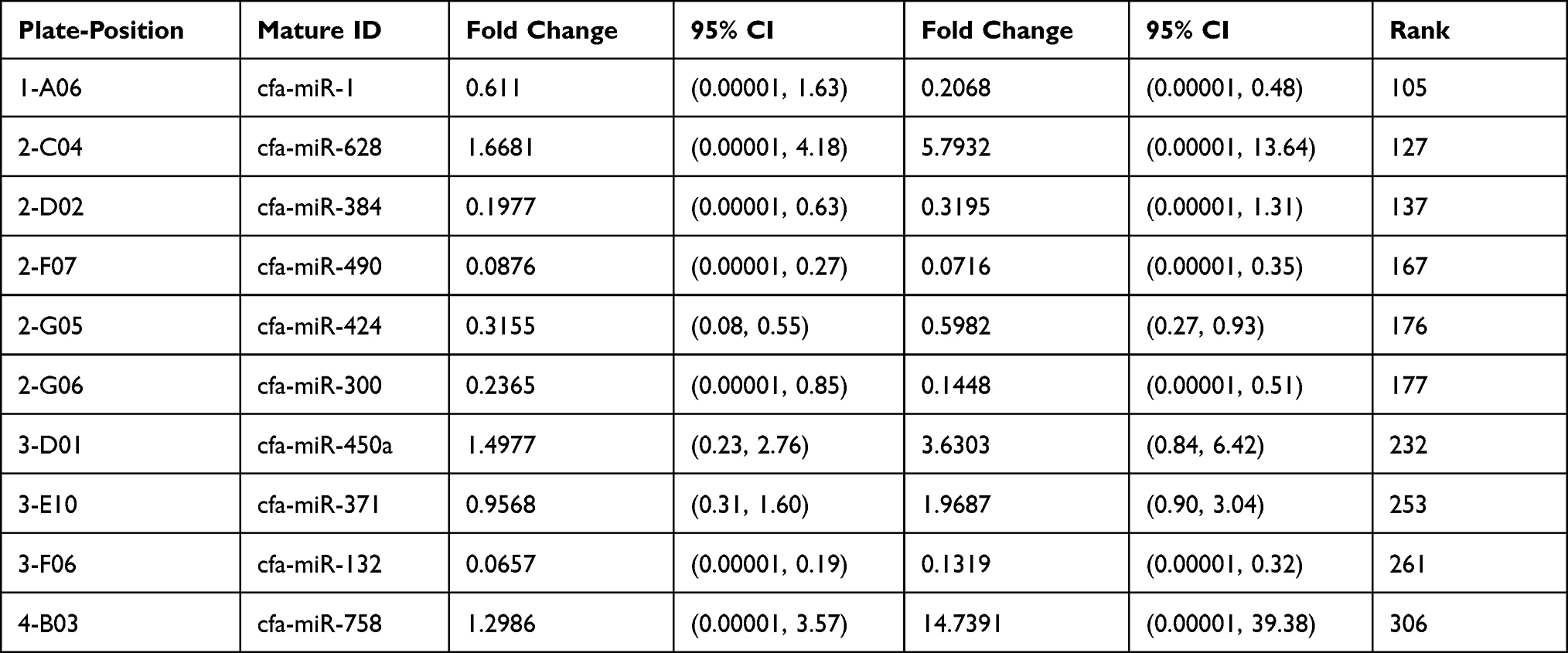 |
Table 1 List of Genes and Plate-Position for Both Groups in Our Data (Serum Groups) |
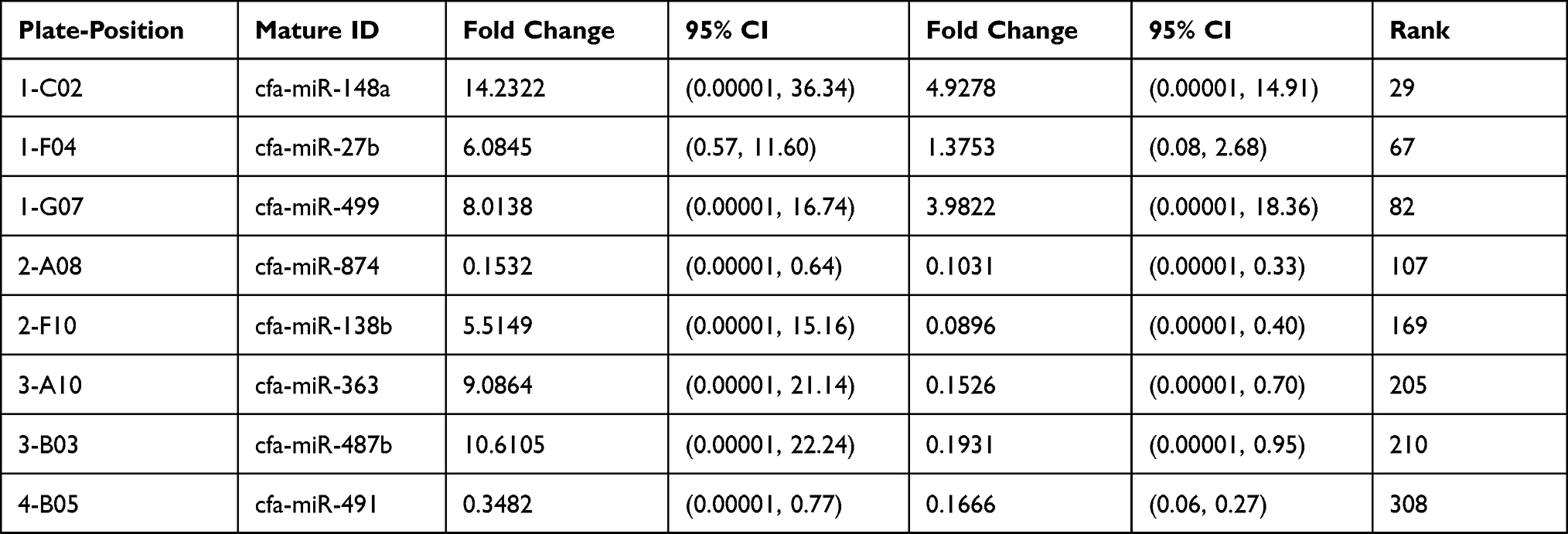 |
Table 2 List of Genes and Plate-Position for Both Groups in Our Data (Urine Groups) |
Fold Change and Fold Regulation (Serum Profile)
The normalized gene expression in the test sample is witnessed through Fold-Change, which divides the control sample’s normalized gene expression. Moreover, fold-change has emerged as a significant approach for representing fold-regulation, as shown in Tables 3 and 4. The values are shown to a greater extent in the fold-change, indicating a positive or an up-regulation, whereas the fold-regulation is similar to the fold change. As shown in Figures 1 and 2, a negative or down-regulation is represented through fold-change values less than 1, where both urine and serum profiles showed a negative inverse of the fold-change.
 |
Table 3 Fold Regulation for Serum Groups |
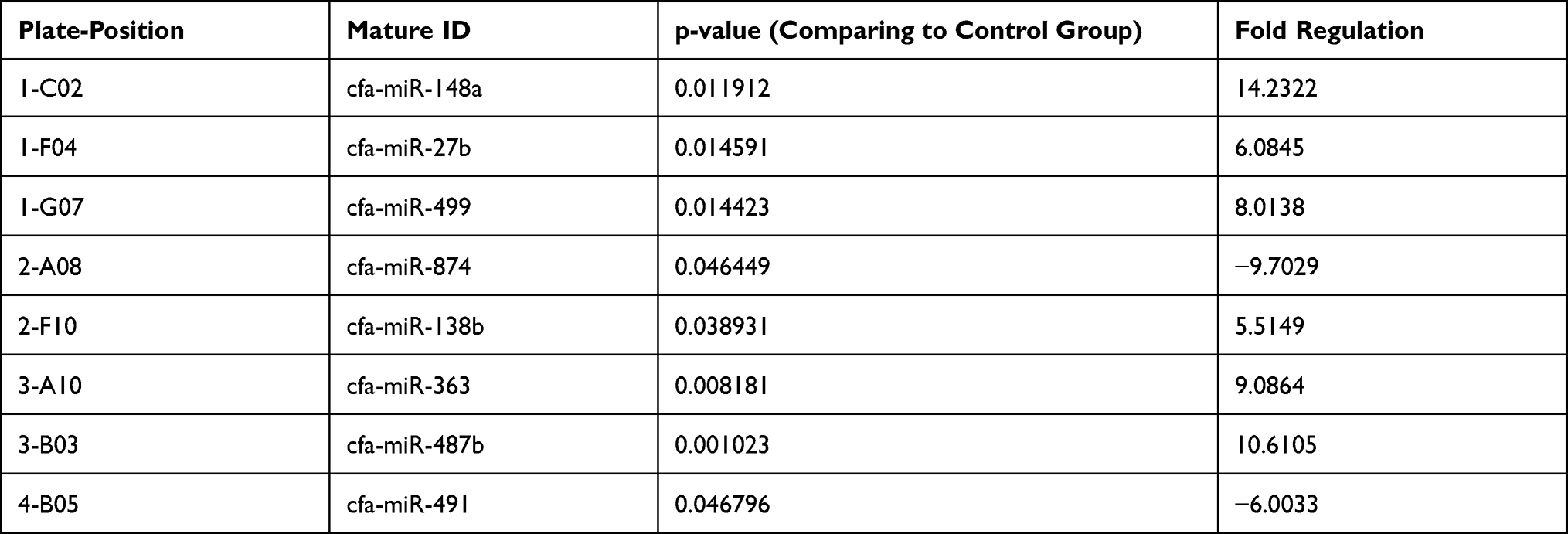 |
Table 4 Fold Regulation for Urine Groups |
 |
Figure 1 Fold change regulation: (A) Serum profile; (B) Urine profile. |
 |
Figure 2 Magnitude of gene expression: (A) Serum profile; (B) Urine profile. |
Discussion
The role of miRNAs in many disease disorders has been studied in recent years. Many miRNAs have important roles in the cell cycle, hematopoiesis, and cellular metabolic control.9 Several in vitro10–12 and in vivo (human)13,14 investigations have demonstrated that miRNAs are temporally modulated not only in the brain but also in the peripheral blood during the course of and notably during the reperfusion phase of a cerebral ischemic event. Differentially expressed miRNAs have been linked to inflammation, endothelial/vascular function, erythropoiesis, angiogenesis, brain function, and hypoxia in people who have had a stroke.13–15 Therefore, we conducted this study to experimentally induce acute cerebral ischemia in dogs to characterize and profiling potential microRNAs biomarkers in blood and urine samples.
When we look at the microRNA of urine, the sample elevates after 2 hours and then decreases expression at 24 hours. On the contrary, the serum elevated to increase expression from 2 hours to 24 hours. The urine will be a tremendous indication in the case model and the performance of the molecular pathology circulation differs in serum and urine. At this point, the microRNA circulated in serum, an effect of molecular pathology in urine after 2 hours, and then the return to normal. On the other hand, the microRNA serum will elevate and reflect the destruction (Dog Model) appearing to increase the expression of miRNA.
A multifaceted cascade of biochemical and physiologic events was triggered by cerebral ischemia. It was assumed that these events were mediated in part by variations in molecular procedures including translation and transcription. In this regard, Lu et al16 detected 12 functional categories, contributing to the temporal modifications in gene expression after transient microRNAs in dogs. To date, there were no reports available on the microRNA microarray profiling of the ischemic brain. Nonetheless, the roles of particular microRNAs have been explained in several reports regarding neurogenesis, neural cell specification, neurodevelopmental function, and neuronal differentiation.17 The expression of 114 miRNAs in ischemic brain samples was identified. Out of these, 82 transcripts and 106 transcripts were detected in the 48-hour and 24-hour reperfusion brain samples, respectively.18
The data have revealed that microRNAs were actively regulated and that their expression pattern was modified with reperfusion time, indicating temporal expression throughout the ischemic injury.19 The majority of the reports have indicated that prolonged reperfusion time was increased with neuronal damage, and this damage can be more extreme as compared to the ischemic injury itself.20 Different upregulations were further observed at 24 and 48 hours of reperfusion for miR-138 and −145, respectively.21 The developmental regulation of miR-138 has been confined to different neuronal populations in the midbrain, cerebrum, and cerebellum of dogs, while miR-145 has been recognized as a particular miRNA that was down-regulated in cancers.22
A total of 32 unique transcripts were represented in the 24-hour brain samples while only 8 unique detectable miRNAs were found in the 48-hour brain samples. A high expression was identified for miR-290 at 24 hours and elevated by 5 times at the 48-hour reperfusion time. The highest expression was revealed by the miR-292-5p after ischemic injury in the brain at 24-hour reperfusion, and elevated even more, by 1.5 times, at 48-hour reperfusion. The majority of the genes, as per DNA microarray data, revealed that miR-290 was downregulated. In brain samples at 24-hour reperfusion, miR-214 was highly expressed but was reduced in the 48-hour reperfused brain.
Because ischemic stroke in people is a diverse illness with complicated pathophysiology, it is impossible to mimic all characteristics of human stroke in a single animal model. One of the models that most accurately simulate human ischemic stroke is the transient or permanent middle cerebral artery blockage (MCAo). Furthermore, the infarcts in this model are consistent and repeatable. Therefore, the MCAo model is considered for this study. The MCAo model offers several benefits including its close resemblance to human ischemic stroke, which is frequently caused by MCAo and has a penumbra similar to that of human stroke. The MCAo model is also notable for its enormous infarct size and good repeatability. Reperfusion and, as a result, the length of ischemia may be carefully controlled. Furthermore, the process is simple to carry out and does not take long. The MCAo model is thought to be appropriate for replicating ischemic stroke and associated neuronal cell death, cerebral inflammation, and BBB damage, as well as providing acceptable behavior test outcomes.23,24
Given the described study design, the dogs are expected to finish the experimental period. The findings will, for the first time, estimate and profile the induced biomarkers potentially associated with experimentally induced cerebral ischemia achieved by non-invasively occluding the middle cerebral artery utilizing advanced interventional neuroradiology techniques. Findings to be obtained from this study may potentially be translated into human medicine that will help in the diagnosis treatment and evaluation of treatment intervention.
Conclusion
This study has presented the profiling of miRNA throughout ischemia and reperfusion in a dog model and has detected different regulation patterns for seven clusters of miRNA. This study has further associated the outcomes with appropriate DNA microarray data and has detected some of the miRNAs that control the expression of 4 genes identified to be essential for the development of cerebral ischemia. The findings might be beneficial to seek additional studies on the probable use of microRNAs as biomarkers for stroke.
Acknowledgment
The authors are very thankful to all the associated personnel for any reference that contributed to/for this research.
Disclosure
This research is funded by the Deanship of research at Jordan University of Science and Technology. The authors declare no competing interest.
References
1. Bacigaluppi M, Comi G, Hermann DM. Animal models of ischemic stroke. Part one: modeling risk factors. Open Neurol J. 2010;4:26–33.
2. Sonderer J, Katan Kahles M. Aetiological blood biomarkers of ischaemic stroke. Swiss Med Wkly. 2015;145:w14138.
3. Tibæk M, Dehlendorff C, Jørgensen HS, et al. Increasing incidence of hospitalization for stroke and transient ischemic attack in young adults: a Registry-Based Study. J Am Heart Assoc. 2016;5(5):e003158. doi:10.1161/JAHA.115.003158
4. Chen HB, Li ZH, Hong H, et al. The clinical and imaging features of 118 patients with spontaneous cerebral arterial dissections presenting with ischemic stroke. Zhonghua Nei Ke Za Zhi. 2016;55:366–371.
5. Navi BB, Reiner AS, Kamel H, et al. OC-02 - risk of arterial thromboembolism in patients with breast cancer. Thromb Res. 2016;140:S169. doi:10.1016/S0049-3848(16)30119-0
6. Jeon J-H, Jung H-W, Jang H-M, et al. Canine model of ischemic stroke with permanent middle cerebral artery occlusion: clinical features, magnetic resonance imaging, histopathology, and immunohistochemistry. J Vet Sci. 2015;16(1):75–85. doi:10.4142/jvs.2015.16.1.75
7. Zhou J, Shao G, Chen X, et al. microRNA 206 and microRNA 574-5p are highly expression in coronary artery disease. Biosci Rep. 2016;36(1). doi:10.1042/BSR20150206
8. Song H-S, Back J-H, Jin D-K, et al. Cardiac troponin T elevation after stroke: relationships between elevated serum troponin T, stroke location, and prognosis. J Clin Neurol Seoul Korea. 2008;4(2):75–83. doi:10.3988/jcn.2008.4.2.75
9. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi:10.1016/S0092-8674(04)00045-5
10. Liu DZ, Tian Y, Ander BP, et al. Brain and blood microRNA expression profiling of ischemic stroke, intracerebral hemorrhage, and kainate seizures. J Cereb Blood Flow Metab. 2010;30(1):92–101. doi:10.1038/jcbfm.2009.186
11. Jeyaseelan K, Lim KY, Armugam A. MicroRNA expression in the blood and brain of rats subjected to transient focal ischemia by middle cerebral artery occlusion. Stroke. 2008;39(3):959–966. doi:10.1161/STROKEAHA.107.500736
12. Dharap A, Bowen K, Place R, et al. Transient focal ischemia induces extensive temporal changes in rat cerebral microRNAome. J Cereb Blood Flow Metab. 2009;29(4):675–687. doi:10.1038/jcbfm.2008.157
13. Tan JR, Tan KS, Koo YX, et al. Blood microRNAs in low or no risk ischemic stroke patients. Int J Mol Sci. 2013;14(1):2072. doi:10.3390/ijms14012072
14. Tan KS, Armugam A, Sepramaniam S, et al. Expression profile of microRNAs in young stroke patients. PLoS One. 2009;4(11):e7689. doi:10.1371/journal.pone.0007689
15. Jickling GC, Ander BP, Zhan X, et al. microRNA expression in peripheral blood cells following acute ischemic stroke and their predicted gene targets. PLoS One. 2014;9(6):e99283. doi:10.1371/journal.pone.0099283
16. Lu XC, Williams AJ, Yao C, et al. Microarray analysis of acute and delayed gene expression profile in rats after focal ischemic brain injury and reperfusion. J Neurosci Res. 2004;77(6):843–857. doi:10.1002/jnr.20218
17. Beuvink I, Kolb FA, Budach W, et al. A novel microarray approach reveals new tissue-specific signatures of known and predicted mammalian microRNAs. Nucleic Acids Res. 2007;35(7):e52. doi:10.1093/nar/gkl1118
18. Schratt GM, Tuebing F, Nigh EA, et al. A brain-specific microRNA regulates dendritic spine development. Nature. 2006;439(7074):283–289. doi:10.1038/nature04367
19. Lagos-Quintana M, Rauhut R, Yalcin A, et al. Identification of tissue-specific microRNAs from mouse. Curr Biol. 2002;12(9):735–739. doi:10.1016/S0960-9822(02)00809-6
20. Sempere LF, Freemantle S, Pitha-Rowe I, et al. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation. Genome Biol. 2004;5(3):R13. doi:10.1186/gb-2004-5-3-r13
21. Wulczyn FG, Smirnova L, Rybak A, et al. Post-transcriptional regulation of the let-7 microRNA during neural cell specification. FASEB J. 2007;21(2):415–426. doi:10.1096/fj.06-6130com
22. Yanaihara N, Caplen N, Bowman E, et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell. 2006;9(3):189–198. doi:10.1016/j.ccr.2006.01.025
23. Fluri F, Schuhmann MK, Kleinschnitz C. Animal models of ischemic stroke and their application in clinical research. Drug Des Devel Ther. 2015;9:3445–3454.
24. Howells DW, Porritt MJ, Rewell SS. Different strokes for different folks: the rich diversity of animal models of focal cerebral ischemia. J Cereb Blood Flow Metab. 2010;30(8):1412–1431. doi:10.1038/jcbfm.2010.66
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.