Back to Journals » International Journal of Nanomedicine » Volume 20
CD42-Enriched Extracellular Vesicles Contribute to Increased Platelet Aggregation and Possibly Organ Damage in Patients with Burn Injury Complicated by Sepsis
Authors Schiavello M ![]() , Vizio B, Bosco O, Mariano F
, Vizio B, Bosco O, Mariano F ![]() , Bruno S
, Bruno S ![]() , Pensa A, Cagna Vallino P, Dini C, Montrucchio G, Lupia E
, Pensa A, Cagna Vallino P, Dini C, Montrucchio G, Lupia E
Received 30 May 2025
Accepted for publication 27 September 2025
Published 21 October 2025 Volume 2025:20 Pages 12733—12750
DOI https://doi.org/10.2147/IJN.S543857
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. RDK Misra
Martina Schiavello,1 Barbara Vizio,1,* Ornella Bosco,1,* Filippo Mariano,1 Stefania Bruno,1 Anna Pensa,2 Paolo Cagna Vallino,1,3 Chiara Dini,1 Giuseppe Montrucchio,1 Enrico Lupia1
1Department of Medical Science, University of Turin, Turin, Italy; 2Burn Centre and Plastic Surgery, Città della Salute e della Scienza di Torino, University Hospital, Turin, Italy; 3School of Specialization in Emergency Medicine, University of Turin, Turin, Italy
*These authors contributed equally to this work
Correspondence: Martina Schiavello, Department of Medical Sciences, University of Turin, via Genova 3, Turin, Italy, Email [email protected]
Background: Sepsis is the leading cause of death in patients with burn injury, in whom it represents a real diagnostic challenge. Here, we studied extracellular vesicles (EVs) released during sepsis in burn patients by multiplexed phenotyping and explored their role in promoting platelet aggregation.
Methods: We enrolled 33 burn patients, 23 with (Burn Septic Patients – BSP) and 10 without sepsis (Burn Non-Septic Patients – BnSP), and 10 healthy subjects (HS). EVs, isolated by ultracentrifugation or precipitation-based methods, were characterized by Nanoparticle Tracking Analysis, Transmission Electron Microscopy, and flow cytometry, and their surface antigens studied by bead-based multiplex flow cytometry. Platelet aggregation was studied in platelet-rich plasma using light-transmission aggregometry.
Results: EVs from BSP expressed a specific pattern of epitopes distinct from those from BnSP and HS. Specifically, EVs from BSP showed an increase in CD42a expression compared to BnSP-(p< 0.05) and HS-(p< 0.0001) derived EVs. Moreover, CD42a-EVs expression increased according to sepsis severity in BSP. In vitro, EVs from BSP, but not from HS, primed platelet aggregation, an effect reduced by an anti-CD42 neutralizing monoclonal antibody.
Conclusion: Our results suggest the potential of CD42a-enriched EVs as diagnostic and prognostic markers of sepsis in burn patients and their role in increasing platelet activation in these patients.
Keywords: extracellular vesicles, platelet aggregation, sepsis
Background
Severe burn is one of the most life-threatening injuries, associated with substantial morbidity and mortality.1 As a consequence of burn injury, patients have skin and other tissue vulnerable to bacterial invasion and this dramatically increases the subsequent risk of sepsis.1 Sepsis is, indeed, the major cause of death in these patients after the first 24 hours from burn injury, representing the final cause of death in 50–60% of non-survivors.2,3 Despite significant recent advances in the knowledge of sepsis pathophysiology, precise and timely detection of sepsis is often a challenge due to its various and often non-specific clinical signs.4 This is especially true in burn patients, who are at higher risk of sepsis development than the general critical-care patient population.3,5–7
One of the most important pathophysiologic mechanisms implicated in the development of organ damage in patients with sepsis is the profound derangement of the delicate balance between pro-coagulant and anti-coagulant mediators within the vasculature.8 During sepsis, several pro-inflammatory cytokines promote the generation and release of pro-coagulant factors that disrupt vascular homeostasis and may finally lead to establish a pro-coagulant state.9 Subsequent thrombosis and vascular leaking are implicated in hypo-perfusion of multiple organs and the onset of multiple organ dysfunction syndrome.9
Extracellular vesicles (EVs) are particles released by all cell types and covered by a lipid bilayer.10 The increasing interest in EVs is based on their bioactive cargo components and mediators, which include cytokines, enzymes, proteins, and microRNAs, which can be delivered to target cells in a paracrine fashion or through the circulation.11,12 EVs thus may play a key role in cell-to-cell communication, cross-organ signaling, and tissue homeostasis.10,13 Recent studies have demonstrated that circulating EV populations and their content change during sepsis,14–18 and that this could be associated with the severity of organ failure.19 In addition, the possible involvement of EVs as markers and mediators of thrombo-inflammation in sepsis has been recently described by our group and others.20,21 It has been also shown that EV surface antigens reflect their cellular origin and may help to discover novel cellular communication mechanisms in several diseases,22,23 including trauma.24
Burn injury not only initiates a complex systemic inflammatory response but also deeply impairs wound healing, a process regulated by cellular cross-talk and EV-mediated signaling.25 Previous studies have highlighted the contribution of EVs in tissue repair and angiogenesis.25,26 On the contrary, little is known about the influence of EVs on platelet activation and the development of systemic complications in burn patients complicated by sepsis, which we specifically intended to address in our study.
Here, we studied plasma-derived EVs released during sepsis in burn patients, and performed a multiplexed phenotyping, with the aim of better describing their cellular origin. Moreover, we evaluated the effects of EVs isolated from plasma of burn patients with sepsis on in vitro platelet aggregation, hypothesizing their potential involvement in sustaining thrombo-inflammation in these patients.
Methods
Patients and Case Adjudication
Patient enrollment was conducted in the Burn Unit of the “Città della Salute e della Scienza di Torino” University Hospital – CTO Site, Turin, Italy, between November 2020 and September 2024. We enrolled 33 patients with burn injury with Total Body Surface Area (TBSA) comprised between 10% and 50%. Among these, 23 patients developed sepsis during their hospital stay (Burn Septic Patients – BSP), whereas 10 burn patients showed no evidence of infection at the time of enrollment and for the following 72 hours (Burn Non-Septic Patients – BnSP). Ten healthy subjects (HS) were also enrolled as an additional control group. All patients gave informed consent for the participation to the study (signed written consent form was waived due to safety protocols), and all data were immediately de-identified.
The diagnosis of sepsis was confirmed before blood collection by an experienced intensivist, blinded to the goals of the study, according to the criteria defined by Sepsis-3 guidelines,27 modified as follows: increase in the Sequential Organ Failure Assessment (SOFA) score of ≥ 2 points plus blood culture positivity. The severity of organ dysfunction was estimated by using the SOFA score,28 calculated at the time of blood collection. The probability of survival was calculated in each patient using the revised Baux score at the beginning of treatment.29 Diagnosis and treatment of patients were managed by a multidisciplinary team on the basis of the guidelines in place at the time.27,30,31
BSP entered the study within 24 hours from their symptom onset. Patients with burn injury who were enrolled as BnSP were monitored after the enrollment in the study for the appearance of signs of infection, positivization of blood culture and deterioration of SOFA score in the following days, and kept in the study only if they did not match the criteria for the diagnosis of sepsis during the 72 hours following venipuncture.
Exclusion criteria were: i) age < 18 years; ii) TBSA <10% and >50%; iii) cancer (active or recent history); iv) autoimmune or chronic inflammatory disease; v) SARS-CoV-2 infection; vi) presence of other diseases that affect blood coagulation or disseminated intravascular coagulation (DIC); vii) platelet count <100 × 109/L.
The study was approved by the Ethics Committee of “Città della Salute e della Scienza di Torino” University Hospital, Turin, Italy (n. CS2/815) and conducted according to the ethical standards of the Declaration of Helsinki and its later amendments. Written informed consent was obtained from all participants, and their samples were anonymized and securely stored for analysis.
Plasma EV Isolation
Ultracentrifugation-Based Method for Quantification and Phenotyping Experiments
Blood was collected into EDTA-containing tubes using a central venous catheter or, in healthy subjects, by clean venipuncture of a peripheral vein using a 21-gauge butterfly infusion set.
Within 1 hour from blood collection, blood was centrifuged for 10 min at 1600 g at 4°C, to eliminate cellular components; then, an additional centrifugation for 30 min at 5000 g at 4°C was performed to further purify the plasma, eliminating apoptotic bodies and platelets. Plasma samples were then aliquoted and stored at −80°C.
Pre-analytical factors for sample handling and storage complied with the recommendations of the International Society for Extracellular Vesicles.32,33
In quantification and phenotyping experiments, EVs were isolated from 10 mL of plasma by ultracentrifugation for 2 hours at 100,000 g at 4°C (Beckham Coulter Optima L-100K, Brea, CA). After discarding the supernatant, EV pellets were resuspended in particle-free PBS, and again ultracentrifugated for 2 hours at 100,000 g at 4°C to reduce protein contaminants. Finally, EVs were resuspended in 150 µL of particle-free PBS with 1% dimethyl sulfoxide (DMSO, Sigma Aldrich). Single-use aliquots were stored at −80°C and used within 3 months from isolation.
Precipitation-Based Method for Functional Experiments
To prepare EVs used in in vitro functional experiments, 1 mL of plasma underwent the following precipitation procedure for EV isolation. Plasma samples were initially submitted to two centrifugations for 20 min at 3000 g at 4°C. Then, a protamine (Sigma, St. Louis, MO, USA)/Polyethylene glycol (35,000; Merck KGaA, Darmstandt, Germany) precipitation solution (0.2 g PEG 35,000 and 1 mg protamine chloride/mL) was added to the plasma samples (1 volume precipitation solution:4 volume sample).34 After overnight incubation at 4°C, the mixture was centrifuged for 30 min at 1500 g at 4°C and the pellet was resuspended in 150 µL of particle-free PBS. Single-use aliquots were stored at −80°C and used within 3 months from isolation.
Nanoparticle Tracking Analysis (NTA)
Detectable sample concentration was approximated using NanoSight LM10 instrument (Malvern Panalytical, Malvern, UK) equipped with a 405 nm laser. Particles isolated by ultracentrifugation were diluted 1:1000 in particle-free PBS and three 30 second videos were captured per sample. The minimum expected particle size, minimum track length, and biomedical light unit setting were set to automatic, and the detection threshold was set to 4. The particle concentration and the distribution graph of the particle size were determined per sample by averaging the results obtained from the analysis of the three independent videos obtained. For data analysis, Nanoparticle Tracking Analysis NTA v2.3 software (Malvern Panalytical) was used, using a camera level of 14.
The concentration of particles per plasma mL prepared from blood taken from HS or patients was calculated considering the starting plasma volume (10 mL) and the final volume of particle-free PBS buffer in which purified particles were resuspended (150 µL) according to the formula provided below. The correction for the 1:1000 dilution of samples before acquisition by NanoSight was applied directly by the NTA software.

Transmission Electron Microscopy (TEM)
TEM was performed on EVs isolated by ultracentrifugation and placed on 200 mesh nickel formvar carbon-coated grids (Electron Microscopy Science, Hatfield, PA, USA). After an adhesion step of 20 minutes, the grids were incubated with 2.5% glutaraldehyde containing 2% sucrose and extensively washed in distilled water. Finally, EVs were negatively stained with NanoVan (Nanoprobes, Yaphank, NK, USA) and acquired with a Jeol JEM 1010 electron microscope (Jeol, Tokyo, Japan).
Flow Cytometry Analysis
Purified EVs were resuspended in 150 µL of particle-free PBS, separated into four aliquots of 20 µL and labelled with either a phycoerythrin (PE; Invitrogen)-conjugated mouse anti-human IgG1 monoclonal antibody against the tetraspanin CD9 (clone eBioSN4, C3-3A2) or a PE-Cyanine7 (Invitrogen)-conjugated mouse anti-human IgG1 monoclonal antibody against the tetraspanin CD63 (clone H5C6) at a volume of 5 µL/test in a final volume of 100 µL diluted in particle-free PBS, for 1 hour at room temperature (RT) in the dark. Mouse IgG1 kappa isotype controls lacking specificity for CD9 and CD63 (P3.6.2.8.1, PE or PE-Cyanine7, eBioscience) were used to measure the signal from non-specific flow cytometry interactions. The Attune NxT Small Particle Side-Scatter Filter (488/10) (Invitrogen, ThermoFisher Scientific) was installed to enable SSC resolution at the scale required to visualise nanoparticles in Attune NxT Acoustic Focusing Flow Cytometer system (ThermoFisher Scientific, MA, US).
For fluorochrome-conjugated antibody compensation, the AbC Total Antibody Compensation Bead Kit (Life Technologies) was used.
Gates were established using the Flow Cytometry Sub-Micron Particle Size Reference Kit (Life Technologies), which provides a set of green fluorescent microsphere suspensions (with a nominal diameter of 100, 200, 500 and 1000 nm) to serve as reliable size references for flow cytometry (Supplementary Figure 1). Sub-Micron Particle Size Reference calibration beads were run before every session for internal calibration, according to a standardized protocol for evaluation of EVs using attune NxT system.35
The acquisition was performed at the flow rate of 25 µL/min and the set options were set on 20,000 events. Control experiments for flow cytometric characterization were performed according to MIFlowCyt-EV instructions.36 A buffer-only control of 0.1 µm-filtered PBS was recorded at the same flow cytometer acquisition settings as all other samples, including triggering threshold, voltages and flow rate. The buffer-only control had a count of ≈100 events/s. Samples were serially diluted six times, with 150 µL of sample added to 150 µL of PBS and measured. The 3rd, 4th, 5th and 6th dilutions showed a linear decrease between dilution factor and measured particle count over one minute (data not shown). Stained samples were treated with 0.1% Triton X-100 for 5 min at RT to test the lability of CD9-PE and CD63-PE stained events. CD9-PE and CD63-PE positive events decreased 85% upon treatment with 0.1% Triton X-100 for 5 min.
Data was analyzed with Attune NxT 3.2 Software.
Multiplex Bead-Based EV Flow Cytometry Assay
In order to phenotype EVs isolated from the plasma of patients and HS, we used a multiplex bead-based EV flow cytometry assay (MACSPlex Exosome Kit, Miltenyi Biotec GmbH, Bergish Gladbach, Germany) according to the protocol provided by the manufacturer and made available in a previously published paper performing a systematic evaluation of the assay.37,38 The principle of the assay is based on the use of polystyrene beads (4.8 µm in diameter). Each bead population is coupled to different antibodies that recognize EV surface antigens and are used to capture antigen-positive EVs. Internal isotype controls are included to monitor any potential unspecific binding. Capture beads are distinguished from each other by their respective hard-dyed fluorescence characteristics detected in PE versus FITC channels, and bead-bound EVs are detected by using APC-conjugated detection antibodies against CD9, CD63 and CD81 (EV markers) and subsequent quantification of APC fluorescence. The kit includes 39 different capture populations mainly directed against a panel of surface antigens relevant for immuno-inflammatory research. EV inputs were based on particle numbers obtained using NTA (assay input dose for control and patients: 3 × 109 NTA-based particles) and diluted with MACSPlex buffer to a final volume of 120 µL and incubated overnight with MACSPlex Exosome Capture Beads, protected from light on shaking at RT. After a washing step, beads-EV complexes were labeled with APC-conjugated detection antibodies against CD9, CD63 and CD81 for 1 hour on an orbital shaker at RT protected from light and then washed twice. As negative control, bead-EV solution without the APC-tetraspanin antibodies underwent the same procedure than EV samples to determine non-specific signals. Finally, samples were analyzed using a CytoFLEX Instrument (Beckham Coulter, Brea, CA, USA).
Single beads were gated to exclude doublets and non-bead events, and FITC and PE voltage were adapted to optimize the discrimination of the 39 bead subsets (Supplementary Figure 2).
Processing of the raw APC-median fluorescence intensity of all 39 capture bead populations was performed based on the indication provided by the guidelines.39 Briefly, background correction was performed by subtracting the respective MFI APC values of matched control sample (buffer only), as well as the fluorescent intensity of the corresponding isotype controls (mIgG1, REA). For each sample APC-median fluorescence intensity (MFI) was normalized by the average MFI of CD9-CD63-CD81 (EV levels), and expressed as normalized MFI (nMFI).
Platelet Aggregation Experiments
Platelet aggregation was evaluated in vitro in 3.8% trisodium citrate-anticoagulated platelet-rich plasma (PRP) as previously described,40 at 37°C with constant stirring at 1000 rpm in a lumi-aggregometer (Chronolog, Havertown, PA). PRP was prepared by centrifugation (15 min at 180xg) of anticoagulated blood obtained from healthy donors by clean venipuncture using a 19-gauge butterfly infusion set, without venous stasis. In preliminary experiments, PRP was incubated with increasing volumes (5µL, 10µL and 20µL) of EV concentrate for 10 minutes at 37°C, the following experiments were performed using 5 µL of EV concentrate. When evaluating priming activity, epinephrine (EPI) or adenosine-diphosphate (ADP) (Helena Laboratories, Beaumont, TX) was added as secondary agonist. For each experiment, the agonist concentration that induced the minimum measurable aggregation was determined: EPI and ADP dose ranges were 0.05–0.2 µm/L and 0.1–1.0 µm/L, respectively. The priming index (PI) was calculated as the response to plasma and agonist together, divided by the sum of individual responses elicited by EVs and agonists.
In separate experiments, designed to inhibit CD42, EVs were incubated with 20 µg/mL of anti-CD42b antibody (anti-GPIbα antibody, Clone 6B4, courtesy of Prof. SF De Meyer, University of KU Leuven Kulak, Belgium) for 10 min at 37°C. The mixture of EVs and anti-CD42b antibody was then added to PRP and further incubated for 10 min at 37°C. Finally, the secondary agonist (EPI or ADP) was added and platelet aggregation was evaluated.
Data Analysis
Data are presented as median (range) or mean ± standard error of mean (SEM), according to data distribution as assessed by the Shapiro–Wilk test. Comparison between groups was carried out by Kruskal–Wallis one-way analysis of variance on ranks followed by Dunn’s multiple comparison tests, and unpaired or paired Student’s t-test, as appropriate.
A p-value < 0.05 was considered significant.
All statistical analyses were performed using GraphPad Prism 9.0 software for Windows and Macintosh (GraphPad Software, La Jolla, CA, USA).
Results
Patient Characteristics
We enrolled 33 patients with burn injury, 23 with (BSP) and 10 without sepsis (BnSP), and 10 healthy subjects (HS). All clinical characteristics and laboratory findings at enrollment for burn patients and healthy subjects are listed in Table 1.
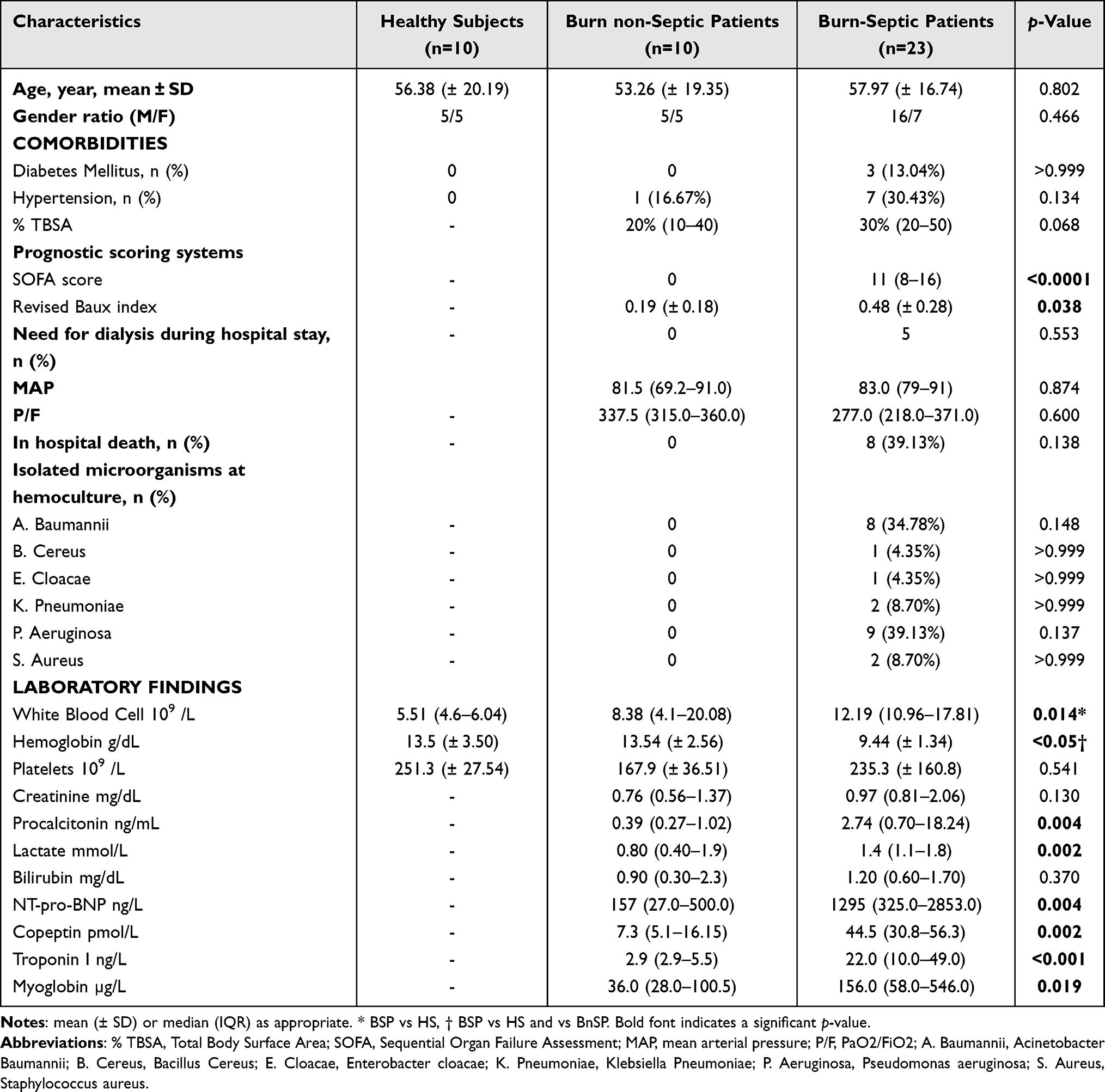 |
Table 1 Demographic and Clinical Characteristics of Patients and Healthy Subjects |
The clinical characteristics were similar in burn patients without (BnSP) vs those with sepsis (BSP), except for the higher SOFA score (p<0.0001), and for the increased Revised Baux Index (p=0.038) in BSP compared to BnSP (Table 1).
The intra-hospital mortality in BSP was about 39% (eight out of 23), whereas none of the BnSP died during hospitalization (Table 1).
White blood cell count was higher in BSP than in HS, whereas hemoglobin concentration was lower in BSP than BnSP and HS (Table 1). In addition, the concentrations of several biomarkers (procalcitonin, NT-pro-BNP, copeptin, troponin I, and myoglobin) were also higher in BSP than in BnSP (Table 1).
Characterization of EVs
Particles isolated from the plasma of HS, BnSP and BSP were first characterized morphologically using TEM (Supplementary Figure 3a–c). EVs displayed a round, cup-shape morphology and were approximately 100 nm in diameter. Further analysis with Nanoparticle tracking analysis (NTA) determined that the particle size distribution was about 100–200 nm (Supplementary Figure 3d–f).
The analysis of flow cytometry experiments demonstrates that particles expressed the EV-specific markers CD9 and CD63 (Supplementary Figure 3g–l).
Sepsis Significantly Alters the Phenotype of Circulating EVs in Burn Patients
NTA showed that the number of particles per mL of plasma was not significantly different in BSP compared to BnSP, whereas it was higher in BSP than in HS (p<0.001). On the contrary, particle concentration did not significantly differ between BnSP and HS (Supplementary Figure 4a). In addition, no differences in particle diameter were found in the three study groups (Supplementary Figure 4b). Of note, we used the NTA analysis to obtain a rough estimate of the number of particles utilized in the following experiments aimed to phenotype EVs isolated from patients and HS. Next, we used the expression of CD9, CD63 and CD81, determined by MACSPlex and flow cytometry, expressed as nMFI, as a measure of EV concentration in the three study groups. We found that the nMFI of all three markers, CD9, CD63 and CD81, was significantly higher in BSP compared with HS (CD9 p=0.004; CD63 p=0.005; CD81 p=0.008) (Supplementary Figure 4c). Furthermore, CD63 expression was also increased in BSP compared to BnSP (p=0.02) (Supplementary Figure 4c).
To investigate the cellular origin of EVs and the potential pathophysiologic role of specific EV surface markers, we performed a multiplex flow cytometry analysis combined with bead-based immune-capture, which allowed to simultaneously analyze the expression of 39 surface proteins by EVs (Supplementary Figure 2b). The average MFI for CD9-CD63-CD81 was used as an internal normalizer of fluorescence levels for all 37 markers to enable the comparison between different samples and experiments and to exclude non-specific binding due to the presence of small debris.
The expression of EV surface markers in HS, BnSP and BSP is shown in Figure 1. In each group, the most abundantly expressed EV surface epitopes were CD42a (platelet/megakaryocytes membrane glycoprotein IX), CD41b (platelet/megakaryocytes membrane glycoprotein II-b), CD62p (p-selectin, vascular endothelial cells and platelet/megakaryocytes membrane protein), CD29 (B1-integrin, platelet, leukocytes, mesenchymal stem cells), and CD31 (PECAM-1, monocytes, platelet, granulocytes, endothelial cell, lymphocytes subsets, and epithelial cells) (Figure 1).
 |
Figure 1 EV epitopes profiling in healthy subjects, burn non-septic patients and burn septic patients. Multiplex flow cytometric analysis of 34 EV surface epitopes in HS (n=10), BnSP (n=10) and BSP (n=23). APC-median fluorescence intensity (MFI) was corrected by subtracting the respective value of blank and isotype control, normalized by the average MFI of CD9-CD63-CD81 (EVs levels), and expressed as normalized MFI (nMFI). Abbreviations: HS, healthy subjects; BnSP, burn non-septic patients; BSP, burn septic patients. Notes: Bar graphs show mean (±SEM). |
When we studied the differential expression of these markers in the three study groups, we found that CD42a expression was increased in BSP compared with BnSP (p=0.03) and HS (p<0.0001) (Figure 2a), whereas CD41b expression was increased only in BSP compared with HS (p=0.02) (Figure 2b). On the contrary, the expression of other platelet and endothelial markers, such as CD29, CD62p, and CD31, did not differ between burn patients, either with or without sepsis, and HS (Figure 2c, e and f).
 |
Figure 2 EVs platelet and endothelial markers expression in healthy subjects, burn non-septic patients and burn septic patients. Expression of CD42a (a), CD41b (b), CD29 (c), CD62p (e), and CD31 (f) in HS (n=10), BnSP (n=10) and BSP (n=23) by multiplex flow cytometric analysis. ROC curve of CD42a-EVs (d) in the discrimination of BSP vs BnSP. Abbreviations: HS, healthy subjects; BnSP, burn non-septic patients; BSP, burn septic patients. Notes: *p<0.05; ****p<0.0001. |
Receiver-operating characteristic curve (ROC) analysis showed that CD42a expression as determined by flow cytometry had a good accuracy for the diagnosis of sepsis in burn patients (Figure 2d), with an AUC of 0.947 [(95% CI, 0.855–1.000), p=0.003], and a specificity of 100% and a sensitivity of 82% based on an nMFI threshold value of 3.340.
CD42a-Enriched EVs Correlate with SOFA Score Severity in Burn Patients with Sepsis
We stratify BSP based on the severity of organ damage in three groups based on SOFA score (Table 2). Detailed clinical characteristics and laboratory findings of the three patient groups so defined are listed in Table 2.
 |
Table 2 Demographic and Clinical Characteristics of Burn Patients with Sepsis Stratified According to SOFA Score |
CD42a-enriched EVs were significantly more abundant in BSP with the highest SOFA score (≥ 11) compared to those with SOFA score 4–6 (p= 0.008) or 7–10 (p=0.014) (Figure 3a). On the contrary, no difference in CD41b expression between these three groups of patients was found (Figure 3b).
 |
Figure 3 CD42a- and CD41b-EVs in stratified patients for SOFA score and %TBSA. nMFI of CD42a (a) and CD41b (b) in stratified patients for SOFA score: 4–6 (n=3), 7–10 (n=8), ≥ 11 (n=12). nMFI of CD42a (c) and CD41b (d) in stratified patients for %TBSA: <30 (n= 10), ≥30 (n= 13). Abbreviations: nMFI, normalized median fluorescence intensity; TBSA, Total Body Surface Area; SOFA, Sequential Organ Failure Assessment. Notes: Bar graphs show mean (± SEM). *p<0.05; **p<0.01. |
We also stratified BSP based on %TBSA (median value cut off=30%; p<0.001) (Figure 3c), but no significant differences were found in EV expression of CD42a and CD41b between these two groups of patients (Figure 3c and d).
In vitro Effect of EVs Isolated From BSP Plasma Samples on Platelet Aggregation
Based on the expression epitope profile, we hypothesized that EVs could be involved in platelet aggregation. To determine the response of naïve platelets to EVs isolated from plasma of BSP, we tested in vitro their effect on platelet aggregation in PRP from healthy donors.
EVs isolated from plasma of burn patients did not show any direct effect on platelet aggregation in PRP from healthy donors at any of the volumes tested (5 µL, 10 µL and 20 µL) (not shown). However, they were able to prime platelet aggregation in PRP when a secondary agonist was subsequently added. In particular, EVs isolated from plasma of BSP significantly enhanced the aggregatory response induced in PRP from healthy donors when a secondary agonist, EPI (Figure 4a) or ADP (Figure 4b), was added. On the contrary, EVs isolated from plasma of HS did not show any priming effect on platelet aggregation in PRP from healthy donors (Figure 4a and b).
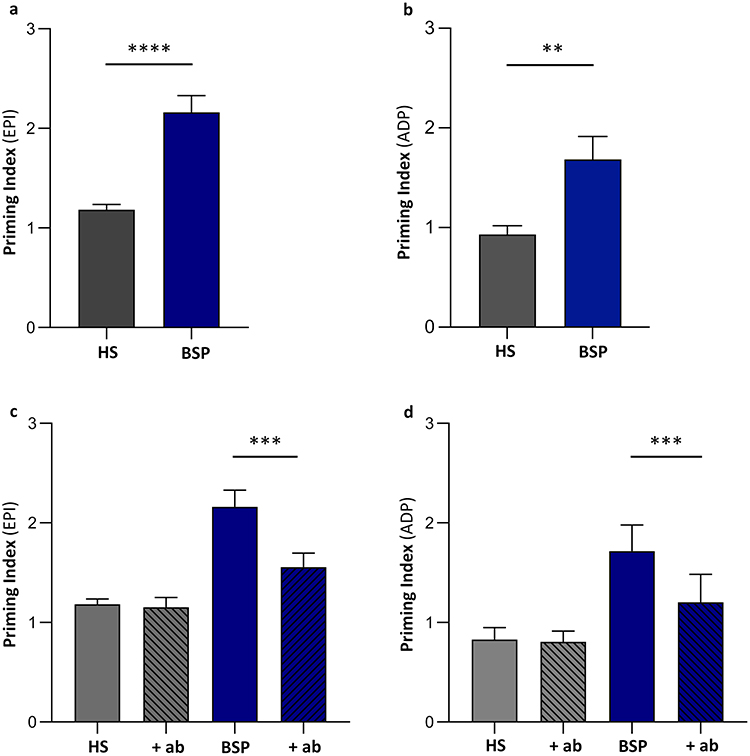 |
Figure 4 In vitro effects of EVs on platelet aggregation. EVs isolated from plasma of BSP or HS (5 µL) were added to platelet-rich plasma (PRP) from healthy donors as described in the Methods section. When evaluating priming activity, epinephrine (EPI) or adenosine-diphosphate (ADP) was added as secondary agonist. EVs from plasma of BSP significantly increase platelet aggregation with both agonists (a and b) compared to those derived from plasma of HS. The pre-incubation of BSP-derived EVs with a neutralizing antibody anti-CD42 (+ab) (c and d) reduced this priming effect on platelet aggregation. Abbreviations: HS, healthy subjects; BSP, burn septic patients. Notes: Bar graphs show mean (± SEM). **p<0.01; ***p<0.001; ****p<0.0001. |
To test whether the observed priming effect was related to different concentrations of EVs present in the different samples, we tested different volumes (5 µL, 10 µL and 20 µL) of EV preparation from HS and BSP and we found no significantly dose-depending effect on platelet aggregation (Supplementary Figure 5a and b). In particular, EVs from healthy subjects, even at the volume of 20 µL (containing approximately 5.08 × 109 particles/mL) and of 5 µL (containing approximately 1.27 × 109 particles/mL), were not able to induce any priming effect (Supplementary Figure 5a). Moreover, increasing the volume of EVs isolated from BSP up to 20 µL (containing approximately 7.19 × 109 particles/mL) did not show any additional effect on the priming induced by 5 µL (containing approximately 1.80 × 109 particles/mL) (Supplementary Figure 5b), further supporting the concept that the functional effect observed on platelet aggregation was due to the phenotype of EVs, rather than to their concentration.
Role of CD42-Enriched EVs in the Priming Activity Exerted by EVs Isolated From Plasma of BSP on Platelet Aggregation
In order to investigate whether elevated circulating levels of CD42a-enriched EVs may contribute to the enhanced platelet aggregation induced by EVs isolated from BSP, we studied in vitro the effect of a neutralizing antibody directed against CD42 on this effect.
In preliminary experiments, we verified that the blocking antibody anti-CD42 does not affect per se platelet aggregation in PRP (data no shown).
Pre-incubation of EVs isolated from plasma of BSP with the anti-CD42-antibody reduced the priming effect observed in PRP from healthy donors when EPI (Figure 4c) or ADP (Figure 4d) were used as secondary agonists. Conversely, the pre-incubation of EVs isolated from plasma of HS with the anti-CD42-antibody did not show any effect (Figure 4c and d).
Discussion
The occurrence of sepsis represents one of the most critical complications in patients with burn injury, largely accounting for the dramatically high mortality in these patients.9 This high lethality is also related to the difficulty in the diagnosis and the consequent delay in the treatment.1,4 Therefore, considerable research efforts have been directed toward searching for new diagnostic parameters that could reflect injury patterns and identifying reliable biomarkers, which could reflect and predict sepsis pattern severity and guide therapeutic choices in burn patients.
Although previous studies have investigated the role of EVs in septic patients,13–15,19 little is known about the role of EVs in the complex scenario of sepsis in burn patients. A few preliminary studies focused on EV characterization only in the first phases of burn injury,41 whereas others completely ignored the insurgence of sepsis.42
A wide area of research is currently devoted to identify signature biomarkers of sepsis useful for diagnostic and prognostic purposes. EV characterization, including the study of both specific surface markers and miRNA EV-cargo, is an area that has been receiving huge interest in this contest. Several EV subtypes have been already associated with various aspects of sepsis pathobiology, such as modulation of inflammation, lymphocyte apoptosis, coagulation, and organ dysfunction.43
Aim of our study was to characterize plasma-derived EVs released during sepsis in burn patients and to investigate their potential involvement in increased platelet activation and thrombo-inflammation in these patients.
The first major finding of our study consists in providing a comprehensive characterization of EV surface epitopes using a standardized multiplex flow cytometric assay,37,38,44 aiming to identify a molecular signature diagnostic of sepsis in patients with burn injury.
The results obtained show that the EVs released upon sepsis insurgence in burn patients are mainly derived from platelets and endothelial cells. In particular, two platelet-associated markers, namely CD42a and CD41b, resulted increase in burn patients with sepsis, and CD42a seems able to discriminate burn patients with sepsis from those without infection. Notably, when we looked at the potential diagnostic performance by ROC curve analysis, CD42a showed a good diagnostic accuracy in discriminating BSP from BnSP. Furthermore, CD42a expression was increased in BSP compared with HS and BnSP. Of note, we did not detect any difference in CD42a expression based on %TBSA in burn patients, suggesting CD42a-enriched EVs as a potential marker of sepsis in these patients, but not of burn severity. In addition, we have also found that CD42a-enriched EVs were significantly more abundant in BSP with the highest SOFA score (≥ 11) compared to those with lower SOFA score (4–7 and 8–10). This suggests that the levels of CD42a-enriched EVs may be useful in helping stratifying burn patients by severity.
In addition to CD42a, EVs isolated from burn patients with sepsis showed a membrane enrichment of other antigens, such as CD62p (p-selectin) and CD146 (tissue factor), on their surface, analogously to what had been described for parent cells45 that mediated pro-coagulant effects.
Previous studies had explored the utility of circulating EVs as biomarkers in order to early differentiate patients with sepsis from those with infection46 or patients with septic shock from critically ill patients without infection.15
While previous studies on EV markers in septic patients focused on a single marker,19 or on few cell type-specific markers,47 in our study we characterized, for the first time, a comprehensive panel of surface markers, which allowed us to identify CD42a as the marker that achieved the highest diagnostic accuracy in discriminating septic from non-septic burn patients. Our results are in line with and expand those of previous studies on different disease,48–51 where EV biomarker profiling is emerging as an important general approach for precision medicine and personalized treatment.52
In addition to their potential role as biomarkers, EVs are receiving increased attention as mediators of cell to cell communication and effectors of disease in sepsis.53,54
In previous studies, EVs have been already shown to be implicated in pathogenic mechanisms important to sustain sepsis hypercoagulability. For instance, in a preliminary study, EVs have been shown to exert pro-coagulant activity in septic patients with disseminating intravascular coagulation (DIC) through increased tissue factor-EV expression.55 Another study reported higher levels of platelet-derived EVs (PEVs) in HIV-infected patients.56 Moreover, viral or bacterial infections have been shown to stimulate platelets to secrete more PEVs.57 In a completely different clinical setting, Kafian S et al demonstrated that EVs from patients with acute coronary syndrome (ACS) enhance platelet aggregation, hypothesizing that blocking EV-formation may reduce platelet hyperactivity in patients with ACS and clopidogrel non-responsiveness.58 Since enhanced platelet activation and aggregation occur in burn patients, especially after sepsis development, suggesting that activated platelets amplify the inflammatory reactions and favor the insurgence of organ damage in these pathological conditions,59 the characterization of EVs as novel mediators involved in these phenomena may help to identify new therapeutic targets allowing for early intervention during the hypercoagulability phase of sepsis.60,61
Moving from these bases, we also aimed to investigate whether CD42-enriched EVs, isolated from plasma of burn patients with sepsis, may be involved in stimulating platelet aggregation. The results we obtained demonstrated that EVs isolated from plasma of burn patients with sepsis are not able per se to stimulate platelet aggregation in PRP from healthy donors, but they enhance the effect of different agonists (EPI and ADP) on platelet aggregation. In addition, the blockade of CD42 expressed on EV surface using a specific monoclonal antibody significantly reduced this effect. Therefore, in light of our and other’s results,62–64 EVs may be considered among the mediators involved in increased platelet activation, potentially contributing to thrombus formation and thrombo-inflammation in sepsis.
Until now, nothing was known about the pathophysiologic functions of CD42-enriched EVs in patients with sepsis/septic shock. It is known that GPIbα (CD42b), together with GPIbβ and GPIX (CD42a), forms the glycoprotein Ib-IX complex, which is a platelet receptor fundamental for platelet recruitment during hemostasis.65 When vasculature is exposed to abnormally high shear stress, platelets and endothelial cells secrete von Willebrand Factor (vWF); next, platelet GPIbα receptors spontaneously associate with circulating plasma vWF, causing platelet activation and aggregation, thereby inducing the formation of platelet thrombi that may have severe pathological consequences.66 Given the dual role of vWF-GPIbα in both thrombosis and inflammation,67 the vWF-GPIbα axis has received great attention, particularly in the setting of ischemic stroke,68 leading to hypothesize that blocking the vWF-GPIbα axis could be an interesting novel treatment strategy in stroke. Our results suggest that the high concentration of CD42-expressing EVs present in burn patients with sepsis may have a pathogenic role in this condition by facilitating platelet aggregation and elucidates a new pathogenic pathway that may lead to increased platelet aggregation in sepsis. Analogously, others demonstrated that the functional delivery of GPIbα from PEVs supports monocyte recruitment in systemic inflammation,69 describing a novel thrombo-inflammatory pathway in which EVs transfer a platelet adhesion receptor to monocytes.
In our study, we focused on sepsis development and diagnosis in a very heterogeneous and complex patient population, but at high risk of developing sepsis, such as the burn patient. To improve the generalizability of our results, it will be very important to expand and confirm our results in other patient populations, including patients with sepsis originating from different sites. Since platelet activation and consequent increased thombo-inflammation are a pathogenic mechanism widely described in sepsis patients, it can be hypothesized that the increased expression of CD42a by platelets we observed in burn patients upon sepsis development may also have a role in patients affected by sepsis of different origin. However, at this stage of our knowledge, we have neither experimental nor clinical data to support this hypothesis.
An apparent contradictory result of our study, since thrombocytopenia has been correlated with poor outcome in extensive burns and it may be considered a sensitive marker of sepsis,70,71 is that BSP had a platelet count within the normal range. However, we expressly excluded from our study those burn patients who matched the diagnostic criteria for DIC or had a platelet count < 100 × 109/L. Thrombocytopenia, while a relevant sepsis biomarker, introduces heterogeneity in EV analysis due to altered platelet activation, consumption and fragmentation.72,73 This exclusion criterion, together with the relatively small number of patients enrolled, may explain our failure to detect thrombocytopenia in burned septic patients.
In our study, we used two different methods, either by ultracentrifugation or charge precipitation to isolate EVs from plasma samples. We acknowledge that the use of different EV isolation methods for phenotypic and functional analyses may have resulted in the assessment of non-identical EV subpopulations. However, since there is no single optimal EV separation method, the choice of the isolation method should be based on the downstream applications and scientific questions addressed.33 It is well known that ultracentrifugation leads to the co-purification of EVs with lipoproteins and soluble proteins,32 hampering the downstream analyses of isolated EVs, such as their count and functional analysis. For this reason, in our study, we used ultracentrifugation for phenotype analysis, whereas we prefer to use a charge-precipitation based method for the functional assay on platelet aggregation.
Finally, it must be accounted that the use of NTA to measure the concentration of EVs in complex biofluids, like plasma, should be interpreted with caution due to counting of co-isolates such as lipoproteins and large protein complexes.33 Therefore, we decided to normalize the number of particles quantified by NTA in the different samples studied by MACSPlex assay, as previously described by others.74,75
Our study had some major limitations. First, we studied a relatively small number of patients, especially in the BnSP groups. This was due to the difficulty in enrolling this type of patients since the occurrence of infectious complications in burn patients is extremely frequent. Another potential limitation resides in the aforementioned difficulty in correctly diagnosing sepsis in burn patients, as burn patients exhibit an excessive inflammatory response syndrome throughout their hospitalization4 and well-established criteria specific for the diagnosis of sepsis in burn patients are not available.4 Moreover, our study was conducted in a single university center, which may limit the generalizability of our data. Therefore, our results need to be confirmed in larger multi-institutional randomized trials. As already anticipated, future studies would also need to include patients with sepsis originated from other sites as well as enrolled at different time-points. Another technical limitation inherent to the use of the multiplex bead-based assay to determine the expression of different markers on EV surface needs to be taken into account. While this method is very useful, indeed, for understanding the general repertoire of EV surface marker expression and is highly valuable for many researchers in the EV field in different experimental contexts, multiplex analysis itself is semi-quantitative and the change in signal intensity can occur for a variety of reasons, included sample preparation and the quality of EV sample. Finally, in functional platelet experiments, EVs were added based on sample volume rather than exact particle count normalization, thus not allowing to definitely rule-out a dose-dependence effect. However, the control experiments performed, as detailed in the Methods section, suggest that the functional effects observed on platelet aggregation were due to EV phenotype rather than their concentrations.
Conclusions
Our results, based on a complex and extensive characterization of EVs isolated from patient plasma samples, suggest the potential of CD42a-enriched EVs both as diagnostic biomarkers of sepsis in burn patients and as pathogenic mediators of increased platelet activation in this patient population. Taken together, our proof-of-concept findings establish an important step forward in understanding the contribution of EVs in enhancing platelet activation in burn patients complicated by sepsis, thus potentially sustaining their role as mediators of thrombo-inflammation and organ damage development in sepsis. Since sepsis in burn patients is correlated with poor prognosis and low survival, our results may be crucial on the process of detecting novel diagnostic sepsis biomarkers and identifying innovative therapeutic targets potentially able to prevent and counteract organ damage development.
Data Sharing Statement
Deidentified participant data will be made available on a collaborative basis upon reasonable request with publication. Data and research materials used in this study are available upon request to qualified researcher for purposes of replication, further analysis, and academic collaboration.
Acknowledgments
We would like to thank all the clinicians and staff members of the Burn Center of CTO Hospital and Molinette Hospital – A.O.U. Città della Salute e della Scienza di Torino, in particular Prof. Maurizio Stella, Dr Daniela Risso, Dr Alberto Sciarrillo, Dr Emanuele Pivetta, Prof. Fulvio Morello and Dr Loris Del Mastro, for their efforts in enrolling patients. We would like to thank Prof. Simon F. De Meyer (Laboratory for Thrombosis Research, KU Leuven Campus Kulak, Kortrijk, Belgium) for giving us the anti-GPIbα antibody, Clone 6B4. We would like to thank Prof. Giovanni Camussi and Dr Maria Chiara Deregibus for their support and analysis of extracellular vesicles using transmission electron microscopy. We would also like to thank the Department of Medical Sciences – University of Turin – for the BioRender License provided to E.L.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Ministero dell’Università e della Ricerca scientifica e Tecnologica (MURST) ex 60% to EL and GM and Research Projects of National Relevance (PRIN) prot. 2022BPNY3E to EL.
Disclosure
The authors declare no conflict of interest exists.
References
1. Jeschke MG, van Baar ME, Choudhry MA, Chung KK, Gibran NS. Burn injury. Nat Rev Dis Primer. 2020;6:11.
2. Williams FN, Herndon DN, Hawkins HK, et al. The leading causes of death after burn injury in a single pediatric burn center. Crit Care BMC. 2009;13(6):R183. doi:10.1186/cc8170.
3. Jeschke MG, Pinto R, Nathens AB, et al. Inflammation and the Host Response to Injury Collaborative Research Program. Morbidity and survival probability in burn patients in modern burn care. Crit Care Med. 2015;43(4):808–815. doi:10.1097/CCM.0000000000000790.
4. Boehm D, Menke H. Sepsis in Burns-Lessons Learnt from Developments in the Management of Septic Shock. Med Kaunas Lith. 2021;58:26.
5. Mann EA, Baun MM, Meininger JC, Wade CE. Comparison of mortality associated with sepsis in the burn, trauma, and general intensive care unit patient: a systematic review of the literature. Shock Augusta Ga. 2012;37(1):4–16. doi:10.1097/SHK.0b013e318237d6bf
6. Knuth CM, Rehou S, Barayan D, Jeschke MG. EVALUATING SEPSIS CRITERIA IN DETECTING ALTERATIONS IN CLINICAL, METABOLIC, AND INFLAMMATORY PARAMETERS IN BURN PATIENTS. Shock. 2022;58(2):103. doi:10.1097/SHK.0000000000001965
7. Boehm D, Menke H. Sepsis in Burns—Lessons Learnt from Developments in the Management of Septic Shock. Medicina. 2022;58(1):26. doi:10.3390/medicina58010026.
8. Burns JP. Septic shock in the pediatric patient: pathogenesis and novel treatments. Pediatr Emerg Care. 2003;19(2):112–115. doi:10.1097/00006565-200304000-00013
9. Zhang P, Zou B, Liou Y-C, Huang C. The pathogenesis and diagnosis of sepsis post burn injury. Burns Trauma. 2021;9:tkaa047. doi:10.1093/burnst/tkaa047
10. van Niel G, Carter DRF, Clayton A, Lambert DW, Raposo G, Vader P. Challenges and directions in studying cell–cell communication by extracellular vesicles. Nat Rev Mol Cell Biol 2022; 23: 369–382.
11. Zhou E, Li Y, Wu F, et al. Circulating extracellular vesicles are effective biomarkers for predicting response to cancer therapy. eBioMedicine. 2021;67:1.
12. Ciferri MC, Quarto R. Extracellular Vesicles as Biomarkers and Therapeutic Tools: from Pre-Clinical to Clinical Applications. Biology. 2021;10:359.
13. Weber B, Franz N, Marzi I, Henrich D, Leppik L. Extracellular vesicles as mediators and markers of acute organ injury: current concepts. Eur J Trauma Emerg Surg. 2022;48(3):1525–1544. doi:10.1007/s00068-021-01607-1
14. Real JM, Ferreira LRP, Esteves GH, et al. Exosomes from patients with septic shock convey miRNAs related to inflammation and cell cycle regulation: new signaling pathways in sepsis? Crit Care. 2018;22(1):68. doi:10.1186/s13054-018-2003-3.
15. Dakhlallah DA, Wisler J, Gencheva M, et al. Circulating extracellular vesicle content reveals de novo DNA methyltransferase expression as a molecular method to predict septic shock. J Extracell Vesicles. 2019;8:1669881. doi:10.1080/20013078.2019.1669881.
16. Shin B, Lee JY, Yoo H, Park J, Jeon K. Prognostic implication of downregulated exosomal miRNAs in patients with sepsis: a cross-sectional study with bioinformatics analysis. J Intensive Care. 2023;11(1):35. doi:10.1186/s40560-023-00683-2.
17. O’Dea KP, Porter JR, Tirlapur N, Katbeh U, Singh S, Handy JM. Circulating Microvesicles Are Elevated Acutely following Major Burns Injury and Associated with Clinical Severity. PLoS One. 2016;11(12):e0167801. doi:10.1371/journal.pone.0167801.
18. Schiavello M, Bosco O, Vizio B, et al. Profiling of miRNAs Contained in Circulating Extracellular Vesicles and Associated with Sepsis Development in Burn Patients: a Proof-of-Concept Study. Int J Mol Sci. 2025;26:1844.
19. Im Y, Yoo H, Ko R-E, Lee JY, Park J, Jeon K. Exosomal CD63 in critically ill patients with sepsis. Sci Rep. 2021;11(1):20300. doi:10.1038/s41598-021-99777-w
20. Schiavello M, Vizio B, Bosco O, et al. Extracellular Vesicles: new Players in the Mechanisms of Sepsis- and COVID-19-Related Thromboinflammation. Int J Mol Sci. 2023;24(3):1920. doi:10.3390/ijms24031920
21. Li Y, Wang W, Zhang B, Li L, Zhou D. A bibliometric analysis of exosomes in sepsis from 2004 to 2022. Medicine. 2023;102(31):e34613. doi:10.1097/MD.0000000000034613
22. Al-Koussa H, AlZaim I, El-Sabban ME. Pathophysiology of Coagulation and Emerging Roles for Extracellular Vesicles in Coagulation Cascades and Disorders. J Clin Med. 2022;11:4932.
23. Plaschke K, Brenner T, Fiedler MO, et al. Extracellular Vesicles as Possible Plasma Markers and Mediators in Patients with Sepsis-Associated Delirium—A Pilot Study. Int J Mol Sci. 2023;24:15781.
24. Kuravi SJ, Yates CM, Harrison P, et al. Changes in the pattern of plasma extracellular vesicles after severe trauma. PLoS One. 2017;12(8):e0183640. doi:10.1371/journal.pone.0183640
25. Qi X, Li Y, Xiang Y, et al. Hyperthermia-enhanced immunoregulation hydrogel for oxygenation and ROS neutralization in diabetic foot ulcers. Cell Biomater. 2025;1:2.
26. Cheng S, Wang H, Pan X, et al. Dendritic Hydrogels with Robust Inherent Antibacterial Properties for Promoting Bacteria-Infected Wound Healing. ACS Appl Mater Interfaces. 2022;14(9):11144–11155. doi:10.1021/acsami.1c25014
27. Singer M, Deutschman CS, Seymour CW, et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA. 2016;315(8):801–810. doi:10.1001/jama.2016.0287
28. Vincent JL, Moreno R, Takala J, et al. The SOFA (Sepsis-related Organ Failure Assessment) score to describe organ dysfunction/failure. On behalf of the Working Group on Sepsis-Related Problems of the European Society of Intensive Care Med. Intensive Care Med. 1996;22(7):707–710. doi:10.1007/BF01709751
29. Lam NN, Hung NT, Duc NM. Prognosis value of revised Baux score among burn patients in developing country. Int J Burns Trauma. 2021;11(3):197–201.
30. Greenhalgh DG, Hill DM, Burmeister DM, et al. Surviving Sepsis After Burn Campaign. Burns J Int Soc Burn Inj. 2023;2023:1.
31. Evans L, Rhodes A, Alhazzani W, et al. Surviving sepsis campaign: international guidelines for management of sepsis and septic shock 2021. Intensive Care Med. 2021;47(11):1181–1247. doi:10.1007/s00134-021-06506-y
32. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi:10.1080/20013078.2018.1535750.
33. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
34. Deregibus MC, Figliolini F, D’antico S, et al. Charge-based precipitation of extracellular vesicles. Int J Mol Med. 2016;38(5):1359–1366. doi:10.3892/ijmm.2016.2759.
35. Lugo-Gavidia LM, Burger D, Bosio E, et al. A standarized protocol for evaluation of large extracellular vesicles using the attuneTM NXT system. J Immunol Methods. 2021;499:113170. doi:10.1016/j.jim.2021.113170
36. Welsh JA, Van Der Pol E, Arkesteijn GJA, et al. MIFlowCyt-EV: a framework for standardized reporting of extracellular vesicle flow cytometry experiments. J Extracell Vesicles. 2020;9(1):1713526. doi:10.1080/20013078.2020.1713526.
37. d’Alessandro M, Soccio P, Bergantini L, et al. Extracellular Vesicle Surface Signatures in IPF Patients: a Multiplex Bead-Based Flow Cytometry Approach. Cells. 2021;10:1045.
38. Ekström K, Crescitelli R, Pétursson HI, Johansson J, Lässer C, Olofsson Bagge R. Characterization of surface markers on extracellular vesicles isolated from lymphatic exudate from patients with breast cancer. BMC Cancer. 2022;22(1):50. doi:10.1186/s12885-021-08870-w.
39. Welsh JA, Killingsworth B, Kepley J, et al. MPAPASS software enables stitched multiplex, multidimensional EV repertoire analysis and a standard framework for reporting bead-based assays. Cell Rep Methods. 2022;2.:1
40. Lupia E, Capuano M, Vizio B, et al. Thrombopoietin participates in platelet activation in COVID-19 patients. eBioMedicine. 2022;85:2.
41. Coleman LG, Maile R, Jones SW, Cairns BA, Crews FT. HMGB1/IL-1β complexes in plasma microvesicles modulate immune responses to burn injury. PLoS One. 2018;13:e0195335. doi:10.1371/journal.pone.0195335.
42. Maile R, Willis ML, Herring LE, et al. Burn Injury Induces Proinflammatory Plasma Extracellular Vesicles That Associate with Length of Hospital Stay in Women: CRP and SAA1 as Potential Prognostic Indicators. Int J Mol Sci. 2021;22:10083.
43. Burgelman M, Vandendriessche C, Vandenbroucke RE. Extracellular Vesicles: a Double-Edged Sword in Sepsis. Pharmaceuticals. 2021;14:829.
44. Li L, Görgens A, Mussack V, et al. Description and optimization of a multiplex bead-based flow cytometry method (MBFCM) to characterize extracellular vesicles in serum samples from patients with hematological malignancies. Cancer Gene Ther. 2022;29(11):1600–1615. doi:10.1038/s41417-022-00466-1
45. Mosad E, Elsayh KI, Eltayeb AA. Tissue factor pathway inhibitor and P-selectin as markers of sepsis-induced non-overt disseminated intravascular coagulopathy. Clin Appl Thromb. 2011;17:80–87.
46. Lambden S, Laterre PF, Levy MM, Francois B. The SOFA score—development, utility and challenges of accurate assessment in clinical trials. Crit Care. 2019;23(1):374. doi:10.1186/s13054-019-2663-7
47. Fendl B, Weiss R, Eichhorn T, et al. Extracellular vesicles are associated with C-reactive protein in sepsis. Sci Rep. 2021;11(1):6996. doi:10.1038/s41598-021-86489-4
48. Burrello J, Bolis S, Balbi C, et al. An extracellular vesicle epitope profile is associated with acute myocardial infarction. J Cell Mol Med. 2020;24(17):9945–9957. doi:10.1111/jcmm.15594
49. O’Toole HJ, Lowe NM, Arun V, et al. Plasma-derived extracellular vesicles (EVs) as biomarkers of sepsis in burn patients via label-free Raman spectroscopy. J Extracell Vesicles. 2024;13(9):e12506. doi:10.1002/jev2.12506
50. Lässer C, Kishino Y, Park K-S, et al. Immune-Associated Proteins Are Enriched in Lung Tissue-Derived Extracellular Vesicles during Allergen-Induced Eosinophilic Airway Inflammation. Int J Mol Sci. 2021;22(9):4718. doi:10.3390/ijms22094718
51. Shephard AP, Mbengue M, Alraies A, et al. Stroma-derived extracellular vesicle mRNA signatures inform histological nature of prostate cancer. J Extracell Vesicles. 2021;10(12):e12150. doi:10.1002/jev2.12150
52. Beetler DJ, Di Florio DN, Bruno KA, et al. Extracellular vesicles as personalized medicine. Mol Aspects Med. 2023;91:101155. doi:10.1016/j.mam.2022.101155
53. Zhang Y, Meng H, Ma R, et al. Circulating microparticles, blood cells, and endothelium induce procoagulant activity in sepsis through phosphatidylserine exposure. Shock Augusta Ga. 2016;45(3):299–307. doi:10.1097/SHK.0000000000000509
54. Mooberry MJ, Bradford R, Hobl EL, Lin FC, Jilma B, Key NS. Procoagulant microparticles promote coagulation in a factor XI-dependent manner in human endotoxemia. J Thromb Haemost. 2016;14(5):1031–1042. doi:10.1111/jth.13285
55. Meng S, Kang K, Fei D, et al. Preliminary study of microparticle coagulation properties in septic patients with disseminated intravascular coagulation. J Int Med Res. 2021;49(5):3000605211014094. doi:10.1177/03000605211014094
56. Boilard E, Paré G, Rousseau M, et al. Influenza virus H1N1 activates platelets through FcγRIIA signaling and thrombin generation. Blood. 2014;123(18):2854–2863. doi:10.1182/blood-2013-07-515536.
57. Goubran H, Seghatchian J, Sabry W, Ragab G, Burnouf T. Platelet and extracellular vesicles in COVID-19 infection and its vaccines. Transfus Apher Sci. 2022;61(3):103459. doi:10.1016/j.transci.2022.103459
58. Kafian S, Wallén H, Samad BA, Mobarrez F. Microvesicles from patients with acute coronary syndrome enhance platelet aggregation. Scand J Clin Lab Invest. 2019;79(7):507–512. doi:10.1080/00365513.2019.1663554
59. Lupia E, Bosco O, Mariano F, et al. Elevated thrombopoietin in plasma of burned patients without and with sepsis enhances platelet activation. J Thromb Haemost. 2009;7(6):1000–1008. doi:10.1111/j.1538-7836.2009.03348.x
60. Iba T, Helms J, Connors JM, Levy JH. The pathophysiology, diagnosis, and management of sepsis-associated disseminated intravascular coagulation. J Intensive Care. 2023;11(1):24. doi:10.1186/s40560-023-00672-5
61. Tsantes AG, Parastatidou S, Tsantes EA, et al. Sepsis-Induced Coagulopathy: an Update on Pathophysiology, Biomarkers, and Current Guidelines. Life. 2023;13:350.
62. Klaihmon P, Phongpao K, Kheansaard W, et al. Microparticles from splenectomized β-thalassemia/HbE patients play roles on procoagulant activities with thrombotic potential. Ann Hematol. 2017;96(2):189–198. doi:10.1007/s00277-016-2885-6
63. Garabet L, Ghanima W, Hellum M, et al. Increased microvesicle-associated thrombin generation in patients with immune thrombocytopenia after initiation of thrombopoietin receptor agonists. Platelets. 2020;31(3):322–328. doi:10.1080/09537104.2019.1639655
64. Puhm F, Boilard E, Machlus KR. Platelet Extracellular Vesicles: beyond the Blood. Arterioscler Thromb Vasc Biol. 2021;41:87–96.
65. Bendas G, Schlesinger M. The GPIb-IX complex on platelets: insight into its novel physiological functions affecting immune surveillance, hepatic thrombopoietin generation, platelet clearance and its relevance for cancer development and metastasis. Exp Hematol Oncol. 2022;11(1):19. doi:10.1186/s40164-022-00273-2
66. Mody NA, King MR. Platelet Adhesive Dynamics. Part II: high Shear-Induced Transient Aggregation via GPIbα-vWF-GPIbα Bridging. Biophys J. 2008;95(5):2556–2574. doi:10.1529/biophysj.107.128520
67. Manz XD, Bogaard HJ, Aman J. Regulation of VWF (Von Willebrand Factor) in Inflammatory Thrombosis. Arterioscler Thromb Vasc Biol. 2022;42(11):1307–1320. doi:10.1161/ATVBAHA.122.318179
68. Denorme F, Vanhoorelbeke K, De Meyer SF. von Willebrand Factor and Platelet Glycoprotein Ib: a Thromboinflammatory Axis in Stroke. Front Immunol. 2019;10:2884. doi:10.3389/fimmu.2019.02884
69. Chimen M, Evryviadou A, Box CL, et al. Appropriation of GPIbα from platelet-derived extracellular vesicles supports monocyte recruitment in systemic inflammation. Haematologica. 2020;105(5):1248–1261. doi:10.3324/haematol.2018.215145
70. Bordeanu-Diaconescu E-M, Grosu-Bularda A, Grama S, et al. Diagnostic and Prognostic Value of Thrombocytopenia in Severe Burn Injuries. Diagnostics. 2024;14(6):582. doi:10.3390/diagnostics14060582
71. Vardon-Bounes F, Ruiz S, Gratacap M-P, Garcia C, Payrastre B, Minville V. Platelets Are Critical Key Players in Sepsis. Int J Mol Sci. 2019;20(14):3494. doi:10.3390/ijms20143494
72. Setarehaseman A, Mohammadi A, Maitta RW. Thrombocytopenia in Sepsis. Life. 2025;15:274.
73. Eustes AS, Dayal S. The Role of Platelet-Derived Extracellular Vesicles in Immune-Mediated Thrombosis. Int J Mol Sci. 2022;23(14):7837. doi:10.3390/ijms23147837
74. Lia G, Di Vito C, Bruno S, et al. Extracellular Vesicles as Biomarkers of Acute Graft-vs.-Host Disease After Haploidentical Stem Cell Transplantation and Post-Transplant Cyclophosphamide. Front Immunol. 2022;12:1.
75. Wiklander OPB, Bostancioglu RB, Welsh JA, et al. Systematic Methodological Evaluation of a Multiplex Bead-Based Flow Cytometry Assay for Detection of Extracellular Vesicle Surface Signatures. Front Immunol. 2018;9:5.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.