Back to Journals » International Journal of Women's Health » Volume 17
Causal Links Between Cerebrospinal Fluid Metabolites and Postpartum Depression: A Bidirectional Mendelian Randomization Study
Authors Long Y ![]() , Liu H, Ban R, Xu Z, Xu T
, Liu H, Ban R, Xu Z, Xu T ![]()
Received 16 May 2025
Accepted for publication 27 August 2025
Published 5 September 2025 Volume 2025:17 Pages 2919—2928
DOI https://doi.org/10.2147/IJWH.S540467
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Matteo Frigerio
Yanxi Long,1,2 Hui Liu,1,2 Rumeng Ban,1,2 Zifeng Xu,1,2 Tao Xu1,2
1Department of Anesthesiology, The International Peace Maternity and Child Health Hospital, School of Medicine, Shanghai Jiao Tong University, No. 910, Henshan Road, Shanghai, 200030, People’s Republic of China; 2Shanghai Key Laboratory of Embryo Original Diseases, Shanghai, People’s Republic of China
Correspondence: Zifeng Xu, Department of Anesthesiology, The International Peace Maternity and Child Health Hospital, School of Medicine, Shanghai Jiao Tong University, Henshan Road 910, Shanghai, People’s Republic of China, Email [email protected] Tao Xu, Department of Anesthesiology, The International Peace Maternity and Child Health Hospital, School of Medicine, Shanghai Jiao Tong University, Henshan Road 910, Shanghai, People’s Republic of China, Email [email protected]
Background: Postpartum depression (PPD) significantly affects maternal health and infant development, yet its underlying neurochemical mechanisms remain poorly understood. While observational studies have linked cerebrospinal fluid (CSF) metabolites with mood disorders, causal relationships are difficult to establish due to confounding environmental factors.
Methods: We applied bidirectional two-sample Mendelian randomization (MR) to explore causal associations between 338 CSF metabolites and PPD. Genetic instruments were obtained from a metabolome-wide GWAS (n=291), and PPD summary statistics were derived from the FinnGen study (7604 PPD cases; 59,601 controls). Forward MR assessed how CSF metabolites affect PPD risk, while reverse MR evaluated the impact of PPD on metabolite levels. The primary analysis used inverse-variance weighting (IVW), supported by MR-Egger, weighted median/mode, and sensitivity tests including Cochran’s Q and MR-PRESSO.
Results: Forward MR identified 10 CSF metabolites with causal effects on PPD. Protective metabolites included guanosine, argininosuccinate, maleate, N6-methyllysine, and homocarnosine. In contrast, increased levels of N-acetyl-isoputreanine, glucose, benzoate, gluconate, and arachidonate were associated with higher PPD risk. Reverse MR showed that PPD may causally lower CSF maleate levels, suggesting mitochondrial involvement in disease progression. All findings were robust to sensitivity analyses, with no evidence of pleiotropy.
Conclusion: This study is the first to use bidirectional MR to investigate the causal role of CSF metabolites in PPD. Our results highlight potential metabolic drivers and feedback mechanisms in PPD, especially the reciprocal link with maleate. These findings offer new insights into PPD’s neurobiology and suggest novel targets for early detection and may inform novel therapeutic strategies in maternal mental health.
Plain Language Summary: Postpartum depression (PPD) is a serious condition that affects both the mother’s mental health and the baby’s development. Some certain chemicals in cerebrospinal fluid (CSF) may be linked to depression, but it’s hard to tell whether those chemicals actually cause the depression or are just a result of it. In this study, researchers used a method called Mendelian randomization, which uses genetic data to understand cause-and-effect relationships. We looked at genetic information from thousands of people to find out whether specific substances found in the fluid around the brain play a role in PPD. The study found that 10 different CSF chemicals may affect the risk of developing PPD. Some, like guanosine and maleate, seem to help protect against depression. Others, like glucose and benzoate, may increase the risk. Interestingly, they also found that PPD itself may lower levels of maleate, which could be a sign of problems in the cells’ energy production. These findings suggest that certain CSF chemicals could be both causes and consequences of PPD. This opens the door to developing new ways to detect or treat PPD earlier, perhaps by targeting these chemicals.
Keywords: postpartum depression, cerebrospinal fluid metabolites, mendelian randomization
Introduction
Postpartum depression (PPD), affecting 10–20% of women globally, represents a critical public health burden due to its profound impact on maternal-infant bonding, infant neurodevelopment, and maternal mortality risk.1,2 Notably, prevalence disparities exist between developing (15%) and developed nations (12%), underscoring socioeconomic and biological complexities in its etiology.3 Despite advances in understanding hormonal and psychosocial contributors, the neurobiological mechanisms underlying PPD remain poorly defined, particularly regarding the role of cerebrospinal fluid (CSF) metabolites-direct biochemical proxies of brain physiology.4,5
Recent studies highlight the CSF as a dynamic mediator of brain-environment interactions, reflecting metabolic and inflammatory changes tied to neuropsychiatric disorders.6,7 For instance, CSF metabolites such as kynurenine pathway (KP) intermediates-kynurenine, kynurenic acid (KynA), and quinolinic acid (QA)-are implicated in major depressive disorder (MDD) through their dual roles in neuroprotection and excitotoxicity.8 A Swedish population-based study found that subtle CSF pleocytosis (2–10 cells/μL) is associated with reduced survival rates in individuals with Parkinson’s disease.9 The concentrations of amyloid-β peptide 1–42 and total tau protein in the CSF are recognized as established diagnostic biomarkers for Alzheimer’s disease.10 In PPD, hormonal fluctuations during the perinatal period, including rapid declines in estradiol and progesterone, may dysregulate neurotransmitter systems, as evidenced by altered CSF levels of 5-hydroxyindoleacetic acid and γ-aminobutyric acid (GABA) in pregnancy.11 These shifts potentially prime vulnerability to mood disturbances postpartum, yet causal relationships remain speculative due to confounding factors in observational studies.12
Mendelian randomization (MR), a genetic instrumental variable approach, offers a robust framework to infer causality by leveraging genetic variants associated with exposures (eg, CSF metabolites) to assess their effects on outcomes (eg, PPD) while minimizing reverse causation and environmental confounding.13 Applying this method to PPD could resolve longstanding debates—for example, whether CSF KP metabolites are drivers or consequences of depressive pathophysiology. Existing MR studies on PPD have focused on gut microbiota and blood metabolites,14,15 but CSF-centric analyses are absent, despite the compartment’s direct relevance to brain function. Critically, CSF metabolomics in depression reveal discordances with peripheral blood. While plasma KynA/QA ratios are reduced in MDD, CSF levels show no such association, suggesting central-peripheral metabolic decoupling.16 Similarly, CSF prolactin and oxytocin during pregnancy correlate weakly with plasma levels, highlighting the CSF’s unique role in mediating neuroendocrine adaptations.11 These findings underscore the necessity of CSF-focused investigations to elucidate PPD-specific mechanisms. This CSF-focused approach offers critical advantages for three reasons. First, cerebrospinal fluid (CSF) directly interfaces with brain interstitial fluid, capturing real-time neurochemical changes unaltered by the blood-brain barrier’s (BBB) selective permeability.17 In contrast, peripheral blood metabolites—though readily accessible—reflect systemic physiology and serve as poor surrogates for CNS-specific processes, particularly for polar molecules whose BBB transit is compromised.18 Second, Perinatal neuroendocrine adaptations (eg, oxytocin fluctuations, HPA axis remodeling) manifest uniquely in CSF, as demonstrated by the weak correlation between CSF and plasma oxytocin levels during pregnancy.11 Third, Although Mendelian randomization (MR) studies of blood and gut microbiota metabolites reveal systemic associations,14,15 their indirect nature precludes direct insight into central nervous system (CNS)-specific mechanisms in postpartum depression (PPD). For instance, gut-derived metabolites such as short-chain fatty acids (SCFAs) undergo extensive metabolism in peripheral tissues (including liver, adipose, skeletal muscle and pancreas) and blood-brain barrier (BBB) filtration before reaching neural targets, dissociating their peripheral signatures from neuropathology.19,20 Furthermore, interventions like repetitive transcranial magnetic stimulation (rTMS) modulate CSF metabolites such as cystine (oxidative stress marker) and creatinine (energy metabolism indicator), linking metabolic shifts to therapeutic efficacy in treatment-resistant depression.21 Such evidence posits CSF metabolites as both biomarkers and mechanistic bridges in PPD.
This study employs bidirectional MR to investigate causal relationships between CSF metabolites and PPD using genome-wide association study (GWAS) data from European-ancestry cohorts. By analyzing genetic instruments for 338 CSF metabolites and PPD risk (FinnGen database), we address two hypotheses: (1) CSF metabolites causally influence PPD susceptibility, and (2) PPD reciprocally alters CSF metabolic profiles. Critically, bidirectional MR is uniquely valuable here for disentangling temporal relationships: it simultaneously identifies potential metabolic drivers of PPD and metabolic consequences of established disease (eg, downstream effects reflecting neuroadaptive or pathological feedback loops). This distinction is essential for clinical translation—while causal metabolites may represent therapeutic targets, disease-altered metabolites could serve as diagnostic biomarkers or indicators of reversible neurochemical sequelae. Our approach adheres to STROBE-MR guidelines, ensuring rigorous IV selection and sensitivity analyses to mitigate pleiotropy.22
This work fills a critical gap in PPD research. First, it identifies actionable metabolic targets for prevention. Second, it clarifies whether PPD-associated metabolic changes are reversible sequelae or fixed risk factors. Finally, by integrating CSF and genetic data, we advance precision psychiatry, offering pathways to subtype PPD based on metabolic signatures and optimize interventions like rTMS or hormonal therapies.
Materials and Methods
Study Design Overview
The schematic framework of this bidirectional MR analysis is illustrated in Figure 1. We employed a two-sample bidirectional MR approach to investigate the putative causal relationships between CSF metabolites and PPD. To ensure robust causal inference, our study adhered to three core MR assumptions: (1) genetic instruments must strongly associate with the exposure; (2) instruments must remain independent of confounders influencing the exposure-outcome relationship; and (3) instruments must affect the outcome exclusively via the exposure, with no alternative pathways.
 |
Figure 1 Flowchart of Mendelian randomization study revealing causality between cerebrospinal fluid metabolites and the risk of postpartum depression. |
Data Sources
The genetic instruments for CSF metabolites were derived from a metabolome-wide GWAS conducted by Panyard et al7 which comprehensively profiled 338 metabolites in a cohort of 291 individuals of European ancestry. Although the sample size (n=291) is modest, this represents the largest available CSF metabolome GWAS to date. Crucially, the validity of MR analyses using this dataset is supported by: (1) successful application of similar-scale CSF metabolomic GWAS in published MR studies examining neurological outcomes;23–25(2) stringent genetic instrument selection (F-statistic >10), effectively minimizing weak instrument bias; (3) demonstrated biological plausibility of identified metabolite-trait associations in prior publications7,16 and (4) the high-depth metabolic quantification (>338 metabolites) compensates for sample size limitations by capturing comprehensive neurochemical profiles. Among these, 299 metabolites were chemically characterized and systematically categorized into eight principal metabolic pathways: amino acids, carbohydrates, cofactors and vitamins, energy substrates, lipids, nucleotides, peptides, and xenobiotic metabolism. The remaining 39 metabolites remain unidentified due to unresolved chemical properties. The GWAS summary statistics, encompassing these findings, are publicly accessible through the GWAS Catalog (accession numbers GCST90025999 to GCST90026336) and can be retrieved from the European Bioinformatics Institute (EBI) repository at http://ftp.ebi.ac.uk/pub/databases/gwas/summary_statistics/. This dataset provides a high-resolution genetic map of CSF metabolites, offering a valuable resource for investigating their causal relationships with complex traits.
The genetic association data for PPD were obtained from the FinnGen R8 public database (https://www.finngen.fi/). The dataset included 7604 PPD cases and 59,601 control individuals of European ancestry. PPD was defined using the International Classification of Diseases, 10th Revision (ICD-10) codes F32 (depressive episode), F33 (recurrent depressive disorder), and F53.0 (mental and behavioral disorders associated with the puerperium, not elsewhere classified). No additional ethical approval was required for this study, as all participants provided informed consent, and the data collection procedures complied with local ethical standards.
Selection of IVs
In this study, instrumental variables (IVs) were carefully selected from publicly available GWAS data to ensure the validity of our MR analysis. For CSF metabolites as the exposure, single-nucleotide polymorphisms (SNPs) were identified using a genome-wide significance threshold of P-value<1×10−5. In the reverse mendelian analysis, for PPD as the exposure, a stricter threshold of P-value<5×10−6 was applied, as the standard threshold of P-value<5×10−8 resulted in an insufficient number of SNPs for robust analysis. This adjustment was made to balance statistical rigor with the need for an adequate number of genetic instruments. To ensure the independence of the selected SNPs and minimize potential bias from linkage disequilibrium (LD), we applied a clumping procedure with an LD threshold of R2 <0.001 and a clumping distance of >10,000 kb between SNPs. Palindromic SNPs were excluded to avoid ambiguity in allele coding or strand orientation. During harmonization, all SNPs were aligned to the human genome reference sequence, and ambiguous or duplicated variants were removed to ensure consistency across datasets. The strength of the selected IVs was assessed using the F-statistic, calculated as: F = Beta2/Se2. SNPs with an F-statistic <10 were excluded to mitigate weak instrument bias. This rigorous selection process ensured that the IVs used in our MR analysis were robust and reliable, strengthening the validity of our findings.
Mendelian Randomization Analysis
We conducted bidirectional MR analyses to investigate the causal relationships between CSF metabolites and PPD. For the forward MR analysis, genetic instruments (SNPs) robustly associated with CSF metabolites were selected as IVs to estimate their causal effects on PPD. Multiple MR methods were applied, including the inverse-variance weighted (IVW) model (primary method), MR-Egger regression, weighted median, weighted mode, and simple mode models. To ensure robustness, only results with consistent odds ratio (OR) directions across all five methods were retained. Statistical significance was defined as P-value<0.05. To examine potential reverse causation, reverse MR analysis was performed using SNPs significantly associated with PPD as IVs to assess their effects on CSF metabolite levels. The same suite of MR methods (IVW, MR-Egger, weighted median, etc). and consistency criteria (directionality and P-value<0.05) were applied.
Sensitivity Analysis
To evaluate potential biases arising from genetic confounding, we performed rigorous sensitivity assessments. Heterogeneity among instrumental SNPs was quantified using Cochran’s Q statistic, with P-values below 0.05 threshold suggesting significant variability. The MR-Egger intercept test and MR-PRESSO global tests (P-value>0.05 indicating acceptable levels) were employed to detect horizontal pleiotropy. Identified outlier SNPs were subsequently removed for sensitivity reanalysis. Finally, leave-one-out analyses systematically evaluated each variant’s disproportionate impact by sequentially excluding individual SNPs from the full dataset.
Statistical Analysis
Statistical analyses were implemented in the R language environment (version 4.2.1). The “TwoSampleMR” package generated exposure-outcome effect estimates, with “MR-PRESSO” implementing diagnostic evaluations for horizontal pleiotropy and heterogeneity. Effect sizes were reported as odds ratios (OR) with 95% confidence intervals (CI) calculated via inverse-variance weighted methods. A conservative statistical significance threshold of P-value<0.05 (two-tailed) was applied throughout the investigation.
Results
Investigation of the Causal Effect of CSF Metabolites on PPD Risk
Using a two-sample bidirectional MR framework, we identified genetic instruments for 338 CSF metabolites by applying stringent criteria (P-value<1×10−5), yielding 452 independent SNPs with robust instrument strength (Table S1 and Figure S1). At a nominal significance threshold (P-value<0.05), IVW regression revealed causal associations between 10 CSF metabolites and PPD risk. Notably, elevated CSF levels of guanosine (OR: 0.93; 95% CI: 0.87–1.00), argininosuccinate (OR:0.96; 95% CI: 0.93–0.99), maleate (OR: 0.96; 95% CI: 0.92–1.00), N6-methyllysine (OR: 0.97; 95% CI: 0.94–0.99) and homocarnosine (OR: 0.98; 95% CI: 0.96–1.00) exhibited protective effects against PPD. Conversely, higher concentrations of N-acetyl-isoputreanine (OR: 1.22; 95% CI: 1.01–1.46), glucose (OR: 1.15; 95% CI: 1.01–1.30), benzoate (OR: 1.07; 95% CI: 1.04–1.11), gluconate (OR: 1.07; 95% CI: 1.02–1.13), and arachidonate (20:4n6) (OR: 1.04; 95% CI: 1.00–1.08) were associated with increased PPD susceptibility (Figure 2, Table S2, and Figure S2). For enhanced clarity, Table 1 summarizes the direction, effect size, and significance of all 10 CSF metabolites with causal effects on PPD risk.
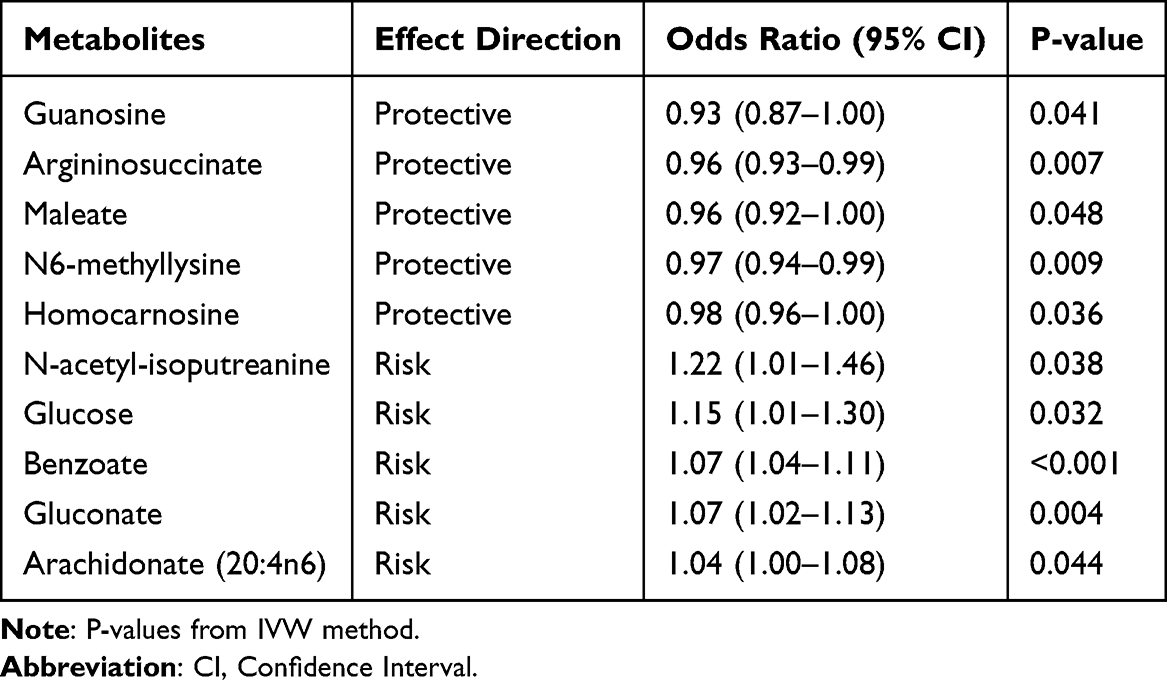 |
Table 1 Causal Effects of CSF Metabolites on PPD Risk |
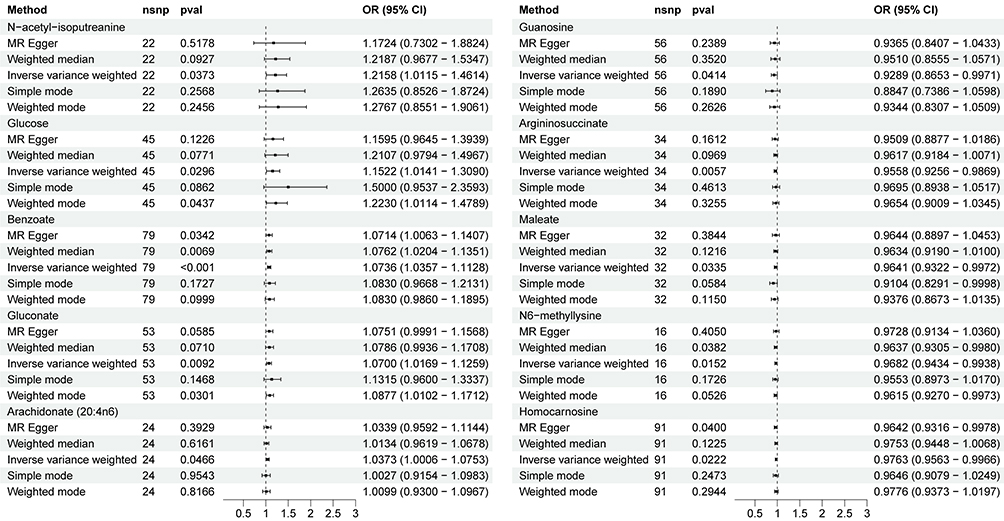 |
Figure 2 Associations between genetically predicted cerebrospinal fluid metabolites on the risk of postpartum depression. |
Exploration of the Causal Effect of PPD Risk on CSF Metabolites
To assess reverse causality, we performed bidirectional MR using PPD as the exposure and CSF metabolites as outcomes. Genetic instruments for PPD were selected at a genome-wide significance threshold (P-value<5×10−6), with 22 independent SNPs retained (Table S3 and Figure S3). Causal effects were estimated via IVW method. Reverse MR analysis identified a robust negative causal effect of PPD risk on CSF maleate levels (P-value=0.0007; OR: 0.69; 95% CI: 0.56–0.85; Figure 3), suggesting that PPD pathogenesis may dysregulate central metabolic pathways. Full results, including metabolite-specific effect estimates (Figure 3, Table S4, and Figure S4). These findings implicate CSF metabolic remodeling as a plausible mechanistic link between PPD and neurochemical homeostasis, with potential therapeutic implications for targeting maleate-associated pathways.
 |
Figure 3 Associations between genetically predicted postpartum depression on cerebrospinal fluid metabolites. |
Sensitivity Analysis
Heterogeneity testing, utilizing the Cochran’s Q test, revealed no significant variability among the included SNPs. The MR-Egger regression intercept provided no evidence of horizontal pleiotropy between CSF metabolites and PPD, thereby validating the exclusion restriction assumption and indicating robust causal relationships. Furthermore, the MR-PRESSO global test did not identify any outliers (Table S5). The funnel plot analysis showed that the data points for individual SNPs were approximately symmetrically distributed, suggesting minimal influence from potential biases (Figures S5 and S6). Leave-one-out sensitivity analysis demonstrated that the estimated effect size remained stable when each SNP was sequentially excluded, indicating that no single SNP significantly impacted the MR estimates, thus confirming the stability of the causal relationship (Figures S7 and S8).
Discussion
Our bidirectional MR analysis provides novel insights into the causal interplay between cerebrospinal fluid CSF metabolites and PPD risk, advancing the mechanistic understanding of this debilitating disorder. The findings align with and extend prior neurobiological and metabolomic research on PPD pathogenesis, as highlighted in the literature.26,27 Below, we contextualize these results within existing literature, propose biological mechanisms, and discuss clinical implications.
Due to direct brain contact, CSF better reflects central nervous system metabolic changes than peripheral samples restricted by the blood-brain barrier.28 The protective association of elevated CSF guanosine with reduced PPD risk resonates with emerging evidence linking purine metabolism to mood regulation. Guanosine, a purine nucleoside, modulates glutamatergic and adenosinergic signaling, both implicated in neuroinflammation and synaptic plasticity.29 Reduced guanosine levels have been linked to depressive phenotypes in animal models, potentially due to impaired neurotrophic support and increased oxidative stress.21,30 Our results suggest that guanosine’s neuroprotective properties, potentially via anti-inflammatory or neurotrophic mechanisms, could mitigate PPD susceptibility. Similarly, the inverse association of argininosuccinate and homocarnosine with PPD risk underscores the role of urea cycle and GABAergic dysregulation in perinatal mood disorders.31,32 Argininosuccinate is a precursor in the urea cycle and arginine biosynthesis, influencing NO production-a mediator of cerebral blood flow and neuroinflammation.33,34 Dysregulated NO signaling has been implicated in perinatal mood disorders, where excessive NO may exacerbate oxidative damage.35 Thus, higher argininosuccinate levels may buffer against PPD by maintaining NO homeostasis. Homocarnosine, a GABA-derived dipeptide, reflects GABAergic tone, which is critical for stress resilience.36 Homocarnosine enhances GABA availability, counteracting the postpartum decline in neurosteroids like allopregnanolone, which are critical for mood stability.35 Also, Braga et al found that female Carns1-/- mice exhibited depression-like behaviors with higher plasma corticosterone levels in the forced swimming test, indicating a role for homocarnosine deficiency in depressive phenotypes. This aligns with clinical trials showing GABAergic agents (eg, brexanolone) effectively treat PPD.37 Conversely, elevated CSF glucose and arachidonate were associated with increased PPD risk, implicating metabolic and inflammatory pathways. It had been proved that higher CSF glucose levels were significantly associated with depressive symptoms at baseline, suggesting a potential link between elevated CSF glucose and depression in first-episode psychosis patients.38 Previous study found that arachidonate metabolism via sEH/NF-κB pathways may drive postpartum depression by promoting neuroinflammation. Shenqi Jieyu Formula alleviates PPD by inhibiting sEH, enhancing anti-inflammatory EETs, and suppressing NF-κB-mediated cytokine release.39 The positive association between arachidonate (20:4n6) and PPD underscores that, a precursor of pro-inflammatory eicosanoids, may amplify cytokine-driven depressive symptoms. The robust reverse causal effect of PPD on CSF maleate suggests a feedback loop wherein PPD exacerbates metabolic perturbations. Maleate, an intermediate in the citric acid cycle, is critical for cellular energy production.40 Its depletion may reflect mitochondrial dysfunction, a hallmark of depression.41,42 This finding parallels studies showing that chronic stress reduces tricarboxylic acid (TCA) cycle metabolites, impairing neuronal energy metabolism.43 The bidirectional relationship highlights PPD’s enduring metabolic consequences, potentially perpetuating a vicious cycle of neurochemical imbalance.
Clinically, our identification of CSF metabolites could serve as promising diagnostic and therapeutic targets. For instance, boosting purine metabolism via guanosine supplementation or enhancing GABAergic signaling through homocarnosine modulation could be explored as novel interventions. Conversely, targeting glucose metabolism or arachidonate-derived inflammation may mitigate PPD risk. The bidirectional MR framework also underscores the need for early intervention. Since PPD may reduce CSF maleate, a marker of mitochondrial health, prompt treatment could prevent metabolic deterioration and improve long-term outcomes.
While our bidirectional MR approach provides robust causal inference, several considerations merit attention. First, genetic heterogeneity across different ethnic group could limit the generalizability of the results. Second, the cerebrospinal fluid metabolomics data (n=291) lacked sex stratification despite postpartum depression’s female-specific etiology. This precludes analysis of hormonal interactions or sex-dimorphic metabolic pathways. Third, our MR approach identifies putative causal relationships but cannot replace biochemical validation of the implicated metabolites (eg, guanosine, maleate, homocarnosine). Without orthogonal confirmation through targeted metabolomics in clinical PPD cohorts, animal models, or in vitro systems, the clinical utility of these biomarkers remains speculative. Finally, though sensitivity analyses (MR-Egger, MR-PRESSO) mitigated horizontal pleiotropy, residual confounding cannot be fully excluded. The modest CSF-GWAS sample size constrained instrument strength, despite all instruments satisfying F > 10 thresholds.
Future studies should validate these findings in diverse cohorts and integrate multi-omics data (eg, proteomics, epigenetics) to unravel mechanistic pathways. Longitudinal assessments of CSF metabolites during pregnancy and postpartum could clarify temporal dynamics.
Conclusion
This bidirectional MR analysis bridges genetic epidemiology and neurochemical research, revealing CSF metabolites as both contributors to and consequences of PPD. By elucidating causal pathways involving purine, GABAergic, and inflammatory systems, our findings advance the quest for precision therapeutics in perinatal psychiatry. Future work should translate these insights into targeted interventions, ultimately improving maternal and infant well-being.
Data Sharing Statement
The GWAS summary statistics for endometritis are available on the IEU Open GWAS database (https://gwas.mrcieu.ac.uk/).
Ethics Approval and Consent to Participate
All data utilized in this Mendelian randomization study were derived from publicly available, de-identified genome-wide association study (GWAS) summary statistics on cerebrospinal fluid metabolites and postpartum depression. These original datasets had obtained prior ethical approvals and participant informed consent through their respective consortiums. This study is exempt from institutional ethics review under Article 32 of China’s Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (effective February 18, 2023), specifically meeting the following criteria: (1) Item 1: Research based on public databases that does not involve the identification or use of personal information. (2) Item 2: Analysis of collected data that does not involve the collection of new data or direct intervention in participants. As this work exclusively involved secondary analysis of aggregated genetic summary data without access to individual-level information, no additional ethical approval was required by our institutional review board.
Acknowledgments
We would like to thank the IEU Open GWAS database for providing the dataset.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by grants from the International Peace Maternity and Child Health Hospital (Award Number: IPMCH2022CR-07); the Bethune Charitable Foundation (Award Number: ezmr2022-23).
Disclosure
The authors report no conflicts of interest in this work.
References
1. O’Hara MW, McCabe JE. Postpartum depression: current status and future directions. Annu Rev Clin Psychol. 2013;9:379–407. doi:10.1146/annurev-clinpsy-050212-185612
2. Gavin NI, Gaynes BN, Lohr KN, Meltzer-Brody S, Gartlehner G, Swinson T. Perinatal depression: a systematic review of prevalence and incidence. Obstet Gynecol. 2005;106(5 Pt 1):1071–1083. doi:10.1097/01.AOG.0000183597.31630.db
3. Liu X, Wang S, Wang G. Prevalence and Risk Factors of Postpartum Depression in Women: a Systematic Review and Meta-analysis. J Clin Nurs. 2022;31(19–20):2665–2677. doi:10.1111/jocn.16121
4. Brummelte S, Galea LA. Postpartum depression: etiology, treatment and consequences for maternal care. Horm Behav. 2016;77:153–166. doi:10.1016/j.yhbeh.2015.08.008
5. Yim IS, Tanner Stapleton LR, Guardino CM, Hahn-Holbrook J, Dunkel Schetter C. Biological and psychosocial predictors of postpartum depression: systematic review and call for integration. Annu Rev Clin Psychol. 2015;11:99–137. doi:10.1146/annurev-clinpsy-101414-020426
6. Neumann ID, Landgraf R. Balance of brain oxytocin and vasopressin: implications for anxiety, depression, and social behaviors. Trends Neurosci. 2012;35(11):649–659. doi:10.1016/j.tins.2012.08.004
7. Panyard DJ, Kim KM, Darst BF, et al. Cerebrospinal fluid metabolomics identifies 19 brain-related phenotype associations. Commun Biol. 2021;4(1):63. doi:10.1038/s42003-020-01583-z
8. Paul ER, Schwieler L, Erhardt S, et al. Peripheral and central kynurenine pathway abnormalities in major depression. Brain Behav Immun. 2022;101:136–145. doi:10.1016/j.bbi.2022.01.002
9. Bäckström D, Granåsen G, Domellöf ME, et al. Early predictors of mortality in parkinsonism and Parkinson disease: a population-based study. Neurology. 2018;91(22):e2045–e2056. doi:10.1212/WNL.0000000000006576
10. Jack CR, Albert MS, Knopman DS, et al. Introduction to the recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement. 2011;7(3):257–262. doi:10.1016/j.jalz.2011.03.004
11. Altemus M, Fong J, Yang R, Damast S, Luine V, Ferguson D. Changes in cerebrospinal fluid neurochemistry during pregnancy. Biol Psychiatry. 2004;56(6):386–392. doi:10.1016/j.biopsych.2004.06.002
12. Borgsted C, Høgh S, Høgsted ES, et al. The role of central serotonergic markers and estradiol changes in perinatal mental health. Acta Psychiatr Scand. 2022;146(4):357–369. doi:10.1111/acps.13461
13. Bowden J, Holmes MV. Meta-analysis and Mendelian randomization: a review. Res Synth Methods. 2019;10(4):486–496. doi:10.1002/jrsm.1346
14. Gao Z, Zhou R, Chen Z, et al. Genetic prediction of blood metabolites mediating the relationship between gut microbiota and postpartum depression: a mendelian randomization study. J Psychiatr Res. 2025;181:614–622. doi:10.1016/j.jpsychires.2024.12.025
15. Li H, Meng H, Dang C, et al. Exploring potential causal relationships between gut microbiota, inflammatory factors, and postpartum depression: a Mendelian randomization analysis. BMC Pregnancy Childbirth. 2025;25(1):177. doi:10.1186/s12884-025-07304-w
16. Savitz J. Blood versus cerebrospinal fluid: kynurenine pathway metabolites in depression. Brain Behav Immun. 2022;101:333–334. doi:10.1016/j.bbi.2022.01.021
17. Ogawa S, Tsuchimine S, Kunugi H. Cerebrospinal fluid monoamine metabolite concentrations in depressive disorder: a meta-analysis of historic evidence. J Psychiatr Res. 2018;105:137–146. doi:10.1016/j.jpsychires.2018.08.028
18. Hashimoto K. Metabolomics of major depressive disorder and bipolar disorder: overview and future perspective. Adv Clin Chem. 2018;84:81–99. doi:10.1016/bs.acc.2017.12.005
19. Frampton J, Murphy KG, Frost G, Chambers ES. Short-chain fatty acids as potential regulators of skeletal muscle metabolism and function. Nat Metab. 2020;2(9):840–848. doi:10.1038/s42255-020-0188-7
20. Harris E. What to know about monkeypox. JAMA. 2022;327(23):2278–2279. doi:10.1001/jama.2022.9499
21. Tateishi H, Setoyama D, Kato TA, et al. Changes in the metabolites of cerebrospinal fluid induced by rTMS in treatment-resistant depression: a pilot study. Psychiatry Res. 2022;313:114636. doi:10.1016/j.psychres.2022.114636
22. Emdin CA, Khera AV, Kathiresan S. Mendelian randomization. JAMA. 2017;318(19):1925–1926. doi:10.1001/jama.2017.17219
23. Jie J, Gong Y, Hu H, Liu S. The role of cerebrospinal fluid metabolites in mediating the impact of lipids on late-onset Alzheimer’s disease: a two-step mendelian randomization analysis. J Transl Med. 2024;22(1):1077. doi:10.1186/s12967-024-05796-2
24. Liao J, Jiang L, Qin Y, Hu J, Tang Z. Causal association between cerebrospinal fluid metabolites and Parkinson’s disease: a two-sample bidirectional mendelian randomization study. Behav Brain Res. 2025;482:115426. doi:10.1016/j.bbr.2025.115426
25. Zhao Z, Xing N, Hou L. Cerebrospinal fluid metabolites as potential biomarkers for epilepsy: insights from genome-wide association studies. Epilepsia Open. 2025;10(1):233–242. doi:10.1002/epi4.13101
26. Worthen RJ, Beurel E. Inflammatory and neurodegenerative pathophysiology implicated in postpartum depression. Neurobiol Dis. 2022;165:105646. doi:10.1016/j.nbd.2022.105646
27. Li S, Yang Z, Yao M, et al. Exploration for biomarkers of postpartum depression based on metabolomics: a systematic review. J Affect Disord. 2022;317:298–306. doi:10.1016/j.jad.2022.08.043
28. Orts-Del’Immagine A, Wyart C. Cerebrospinal-fluid-contacting neurons. Curr Biol. 2017;27(22):R1198–R1200. doi:10.1016/j.cub.2017.09.017
29. Kaddurah-Daouk R, Yuan P, Boyle SH, et al. Cerebrospinal fluid metabolome in mood disorders-remission state has a unique metabolic profile. Sci Rep. 2012;2:667. doi:10.1038/srep00667
30. Camargo A, Bettio L, Rosa PB, Rosa JM, Altê GA, Rodrigues A. The antidepressant-like effect of guanosine involves the modulation of adenosine A(1) and A(2A) receptors. Purinergic Sig. 2023;19(2):387–399. doi:10.1007/s11302-022-09898-8
31. Zhao C, Gammie SC. Glutamate, GABA, and glutamine are synchronously upregulated in the mouse lateral septum during the postpartum period. Brain Res. 2014;1591:53–62. doi:10.1016/j.brainres.2014.10.023
32. Enns GM, O’Brien WE, Kobayashi K, Shinzawa H, Pellegrino JE. Postpartum “psychosis” in mild argininosuccinate synthetase deficiency. Obstet Gynecol. 2005;105(5 Pt 2):1244–1246. doi:10.1097/01.AOG.0000157769.90230.24
33. Yan J, Kuzhiumparambil U, Bandodkar A, Bandodkar S, Dale RC, Fu S. Cerebrospinal fluid metabolites in tryptophan-kynurenine and nitric oxide pathways: biomarkers for acute neuroinflammation. Dev Med Child Neurol. 2021;63(5):552–559. doi:10.1111/dmcn.14774
34. Erez A, Nagamani SC, Shchelochkov OA, et al. Requirement of argininosuccinate lyase for systemic nitric oxide production. Nat Med. 2011;17(12):1619–1626. doi:10.1038/nm.2544
35. Payne JL, Maguire J. Pathophysiological mechanisms implicated in postpartum depression. Front Neuroendocrinol. 2019;52:165–180. doi:10.1016/j.yfrne.2018.12.001
36. Almeida FB, Nin MS, Barros H. The role of allopregnanolone in depressive-like behaviors: focus on neurotrophic proteins. Neurobiol Stress. 2020;12:100218. doi:10.1016/j.ynstr.2020.100218
37. Stewart DE, Vigod SN. Postpartum depression: pathophysiology, treatment, and emerging therapeutics. Annu Rev Med. 2019;70:183–196. doi:10.1146/annurev-med-041217-011106
38. Giné-Servén E, Martinez-Ramirez M, Boix-Quintana E, et al. Routine cerebrospinal fluid parameters as biomarkers in first-episode psychosis: a prospective observational study. Prog Neuropsychopharmacol Biol Psychiatry. 2022;112:110424. doi:10.1016/j.pnpbp.2021.110424
39. Jingya L, Song L, Lu L, Zhang Q, Zhang W. Effect of Shenqi Jieyu formula on inflammatory response pathway in hippocampus of postpartum depression rats. Heliyon. 2024;10(9):e29978. doi:10.1016/j.heliyon.2024.e29978
40. Molina-Jijón E, Aparicio-Trejo OE, Rodríguez-Muñoz R, et al. The nephroprotection exerted by curcumin in maleate-induced renal damage is associated with decreased mitochondrial fission and autophagy. Biofactors. 2016;42(6):686–702. doi:10.1002/biof.1313
41. Cao B, Xiao Y, Liu D. Associations of methylmalonic acid and depressive symptoms with mortality: a population-based study. Transl Psychiatry. 2024;14(1):297. doi:10.1038/s41398-024-03015-6
42. Gorman-Sandler E, Robertson B, Crawford J, et al. Gestational stress decreases postpartum mitochondrial respiration in the prefrontal cortex of female rats. Neurobiol Stress. 2023;26:100563. doi:10.1016/j.ynstr.2023.100563
43. Wang C, Cui C, Xu P, et al. Targeting PDK2 rescues stress-induced impaired brain energy metabolism. Mol Psychiatry. 2023;28(10):4138–4150. doi:10.1038/s41380-023-02098-9
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.