Back to Journals » International Journal of Nanomedicine » Volume 21
Black Phosphorus Nanomaterials and the Senescent Osteoimmune Microenvironment: Mechanisms, Opportunities, Challenges, and Future Outlook
Authors Wang JW, Xun JJ, Zhao FF
Received 23 October 2025
Accepted for publication 26 January 2026
Published 2 February 2026 Volume 2026:21 576356
DOI https://doi.org/10.2147/IJN.S576356
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Eng San Thian
Jia-Wen Wang, Jian-Jun Xun, Fei-Fei Zhao
Department of Orthopedics, The Fourth Hospital of Hebei Medical University, Shijiazhuang, Hebei, 050011, People’s Republic of China
Correspondence: Fei-Fei Zhao, Department of Orthopedics, The Fourth Hospital of Hebei Medical University, Shijiazhuang, Hebei, 050011, People’s Republic of China, Email [email protected] Jian-Jun Xun, Department of Orthopedics, The Fourth Hospital of Hebei Medical University, Shijiazhuang, Hebei, 050011, People’s Republic of China, Email [email protected]
Abstract: The pathogenesis of senile osteoporosis involves immune cell imbalance, inflammaging, and dysregulation of the RANKL/OPG bone–immune axis, collectively defining the concept of immunoporosis. These interrelated processes mutually reinforce one another, leading to a 2– 3-fold prolongation of bone healing time, while conventional single-target therapies fail to achieve coordinated regulation of bone regeneration and the immune microenvironment. Black phosphorus (BP) nanomaterials, as an emerging class of biomaterials, represent a paradigm shift from “passive scaffolds” to “active immuno–bone synergistic regulators.” BP exerts multifunctional effects by restoring macrophage M1/M2 polarization balance, scavenging reactive oxygen species (ROS) to disrupt inflammatory feedback loops, and modulating the RANKL/OPG axis, thereby promoting a transition from a pro-inflammatory, destructive state to an anti-inflammatory, reparative phenotype. Experimental evidence indicates that BP can reduce pro-inflammatory cytokine expression by approximately 60% and achieve bone defect bridging rates of up to 93%. However, the clinical translation of BP remains challenged by the complexity of aging-related immune mechanisms, insufficient long-term safety data, and unclear translational pathways. This Perspective systematically discusses the regulatory mechanisms of BP in the aged osteoimmune microenvironment, the current limitations, and future research directions.
Keywords: black phosphorus nanomaterials, immunoporosis, inflammaging, RANKL/OPG axis, osteoimmune regulation
Introduction
With the rapid progression of global population aging, senile osteoporosis has emerged as a major cause of fractures, disability, and mortality in older adults.1,2 Among individuals aged 65 years and older, the lifetime risk of osteoporotic fractures is estimated at 40–50% in women and 13–22% in men,3 with fracture incidence continuing to rise with advancing age.4 This condition imposes substantial medical, economic, and societal burdens.5 Beyond traditional paradigms of bone mass loss and microarchitectural deterioration, the pathogenesis of senile osteoporosis is increasingly recognized as an osteoimmune disorder characterized by immune cell imbalance, inflammaging, and dysregulation of the RANKL/OPG bone–immune axis. These interconnected mechanisms synergistically disrupt osteoblast–osteoclast homeostasis and impair bone remodeling, rendering fracture-induced bone defects particularly difficult to heal in elderly individuals.6–12
Currently available anti-osteoporotic agents, including bisphosphonates, denosumab, and teriparatide, predominantly act on single molecular targets and lack interventions tailored to aging-specific osteoimmune mechanisms. As a result, they are insufficient to modulate the complex immune–bone crosstalk required for effective bone regeneration and immune microenvironment remodeling in older populations.13–16 Moreover, anabolic parathyroid hormone analogs such as teriparatide and abaloparatide are restricted to treatment durations of less than two years and carry potential tumorigenic risks.17 These limitations underscore the urgent need for therapeutic strategies capable of simultaneously improving bone quantity, microstructural integrity, and aging-associated osteoimmune dysfunction to address refractory bone defects in senile osteoporosis.
To overcome the shortcomings of conventional therapies and achieve coordinated regulation of the immune–bone system, recent research has explored various bone repair nanomaterials, including MXenes, bioactive glass, and cerium oxide nanozymes. MXenes exhibit favorable electrical conductivity and hydrophilicity and can modulate immune responses by regulating T-cell N-glycosylation and reducing ROS levels; however, their application in tissue regeneration remains in its infancy.18–20 Bioactive glass demonstrates good osteoconductivity and can promote macrophage polarization toward the M2 phenotype through the release of ions such as Se, Sr, and Zn, yet its dense pore architecture and low specific surface area limit both mechanical performance and bioactivity.21–23 Cerium oxide nanozymes possess strong antioxidant capacity and can enhance osteogenic differentiation,24,25 but their long-term in vivo metabolism remains insufficiently characterized, and they lack direct bone mineralization-promoting effects.26 These constraints highlight the need to identify more effective and versatile nanomaterials.
In this context, black phosphorus nanomaterials have attracted increasing attention. Since their initial report in 2014,27 BP-based materials have been rapidly translated into biomedical applications,28 evolving from photothermal osteogenic scaffolds29–31 to active osteoimmune immunomodulatory regulators.32,33 Compared with other nanomaterials, BP not only promotes osteogenic differentiation and bone regeneration but also exerts immunomodulatory effects by regulating macrophage polarization, suppressing excessive inflammation, and improving the osteoimmune microenvironment through multi-target synergistic mechanisms. These properties enable BP to address aging-specific pathophysiological and immunological alterations in a coordinated manner, thereby enhancing bone repair and overall skeletal health.32–34 Nevertheless, challenges such as poor environmental stability, difficulty in precisely controlling degradation kinetics, and limited long-term safety data continue to impede its clinical translation.
This Perspective highlights BP as a representative paradigm shift from “passive scaffolds” to “active immuno–bone synergistic regulation” and focuses on the following key issues (Figure 1):
- How do immunosenescence, inflammaging, and dysregulation of the bone–immune axis mutually reinforce each other in the aged osteoimmune microenvironment to form a difficult-to-reverse pathological cycle? What other mechanisms are involved, and what are their specific roles?
- How do black phosphorus nanomaterials reverse pathological alterations in the aged bone microenvironment through immune remodeling, antioxidant effects, and osteoimmune crosstalk?
- What are the key bottlenecks in immune mechanism elucidation, long-term safety, and clinical translation when applying black phosphorus nanomaterials in elderly patients?
- How can AI-assisted design, aged animal models, and long-term safety evaluation facilitate the clinical translation of black phosphorus nanomaterials in the future?
 |
Figure 1 Mechanistic overview of black phosphorus nanomaterials in remodeling the aged osteo-immune microenvironment. Abbreviations: ROS, reactive oxygen species; BP, black phosphorus; RANKL, receptor activator of nuclear factor-κB ligand; OPG, osteoprotegerin; TNF-α, tumor necrosis factor-α; PO43−, phosphate ion. Notes: The left panel illustrates the pathological features of the aged osteo-immune microenvironment, including excessive reactive oxygen species (ROS) accumulation, predominance of M1 macrophages, and an increased RANKL/OPG ratio that drives osteoclast hyperactivity. The central panel highlights black phosphorus (BP) nanomaterials as an active immuno-osteogenic modulator, exerting therapeutic effects through three coordinated mechanisms: ROS scavenging, induction of macrophage polarization from the pro-inflammatory M1 phenotype toward the reparative M2 phenotype, and restoration of RANKL/OPG axis homeostasis. Concurrently, BP releases PO43− ions to directly support bone mineralization. The right panel depicts the reprogrammed regenerative microenvironment following BP intervention, characterized by M2 macrophage dominance, enhanced osteoblast activity, and dense trabecular architecture. Notably, BP treatment reduces TNF-α expression by approximately 60% and achieves a bone continuity formation rate of up to 93%.Symbol legend: ↑, upregulation/increase; ↓, downregulation/decrease; →, transition or causative relationship. |
Pathological Basis of the Aged Osteo-Immune Microenvironment
Osteoporosis is highly prevalent in older adults (≥65 years), and bone regenerative capacity is markedly compromised. This phenomenon is primarily driven by immune cell imbalance, inflammaging, and dysregulation of the bone–immune axis within the osteo-immune microenvironment. These factors interact synergistically, forming a complex pathological network. As a consequence, bone healing time in elderly patients is prolonged by approximately 2–3 fold compared with younger individuals, accompanied by a significantly increased incidence of nonunion and pronounced reductions in bone volume fraction (BV/TV) and bone mineral density (BMD).35–37
At the cellular level, profound immune dysregulation characterizes the aged bone microenvironment. Macrophage polarization is severely disrupted, with the bone marrow M1/M2 macrophage ratio markedly elevated, reaching an average of 22.1 ± 16.0.38 Pro-inflammatory M1 macrophages continuously secrete TNF-α and IL-6, which directly suppress osteoblast function39 and upregulate RANKL expression, thereby promoting osteoclastogenesis and exacerbating osteoporosis.7 In parallel, neutrophils in aged individuals exhibit enhanced formation of neutrophil extracellular traps (NETs). Excessive NET accumulation intensifies local inflammation and tissue damage, further accelerating bone destruction.11,40 The balance between regulatory T cells (Treg) and T helper 17 (Th17) cells is also disrupted, with reduced Treg numbers or impaired function, decreased Foxp3 expression, and a relative expansion of Th17 cells. This shift results in elevated IL-17 levels,41,42 which contribute to bone mineral density loss and inflammatory bone damage.43 Aberrant activation of the PI3K/Akt and STAT3 signaling pathways, commonly observed in aging, further skews T-cell differentiation toward Th17 dominance and amplifies bone-destructive processes.44,45
Inflammaging represents another central pathological feature, with excessive reactive oxygen species (ROS) accumulation as a key driver. Aging is associated with mitochondrial dysfunction, diminished activity of antioxidant defense systems (SOD, CAT, and GPx), and progressive ROS accumulation.46 These alterations lead to sustained NF-κB activation47 and facilitate TXNIP–NLRP3 interaction, triggering caspase-1 activation and promoting the maturation and release of IL-1β and IL-18.48–52 Together, these events establish a self-amplifying inflammatory cascade—“ROS → NF-κB → TXNIP/NLRP3 → IL-1β/IL-18 → further ROS generation”—which exacerbates oxidative stress and tissue injury.53,54 Aging also enhances expression of the senescence-associated secretory phenotype (SASP), comprising pro-inflammatory cytokines, chemokines, growth factors, and matrix-remodeling enzymes,55,56 thereby inducing a “senescence contagion” effect.57 Immune cells recruited by SASP are functionally compromised and fail to efficiently clear senescent cells; instead, they aggravate local inflammation, suppress osteogenic differentiation of mesenchymal stem cells (MSCs), and promote osteoclastogenesis.58–60 Moreover, aging-induced mitochondrial damage, genomic instability, and impaired autophagy result in cytoplasmic DNA accumulation,61 activating the cGAS–STING pathway and driving type I interferon production, NF-κB–dependent inflammation,62 and SASP amplification.63 Importantly, cGAS–STING signaling interacts with the NLRP3 inflammasome, forming a positive feedback loop that further enhances ROS production.64 This pathway also increases Toll-like receptor sensitivity to damage-associated molecular patterns (DAMPs) while attenuating responses to pathogen-associated molecular patterns (PAMPs),54 leading to persistent expression of inflammatory mediators such as IL-6, IL-1β, and IL-23. In concert with TGF-β, these factors promote differentiation of CD4⁺ precursor cells into Th17 cells, thereby aggravating immune dysregulation.42
The RANKL/OPG bone–immune axis is likewise disrupted in aging. The balance between RANKL and its endogenous antagonist osteoprotegerin (OPG) critically determines osteoclast differentiation and bone resorption intensity.39 Pro-inflammatory cytokines, including TNF-α, IL-6, and IL-17, strongly induce RANKL expression while suppressing OPG production.65,66 Under aging or chronic inflammatory conditions, sustained elevation of these cytokines significantly increases the RANKL/OPG ratio, intensifying bone resorption.67 TNF-α further amplifies this effect by promoting TRAF3 degradation and enhancing RANKL-induced osteoclastogenesis.68 This imbalance is reinforced by bidirectional interactions between immune and bone cells: M1 macrophage–derived TNF-α and IL-6 persistently inhibit osteoblast activity,39,69 whereas increased Th17 cells and IL-17 levels drive inflammatory bone destruction and immunosenescence-associated degenerative changes.70,71 Concurrently, aging-associated decline in MSC immunosuppressive capacity leads to excessive IL-6 production,72 positioning senescent MSCs as potent amplifiers of inflammation. Through paracrine signaling, aged MSCs suppress osteogenic differentiation of neighboring MSCs and exacerbate bone marrow inflammation.59,60 They may also activate pro-inflammatory programs that impair hematopoietic stem and progenitor cell clonogenicity,73 collectively contributing to compromised bone homeostasis and potentially insufficient bone perfusion.
In addition to these core mechanisms, the aged bone microenvironment exhibits other notable pathological features. Mechanotransduction is impaired by age-related alterations in lacunar morphology and degeneration of the lacunar–canalicular network,74,75 leading to reduced YAP/TAZ activity and Piezo1 channel dysfunction, and ultimately diminishing osteocyte mechanosensitivity and skeletal responsiveness to mechanical loading.76,77 Epigenetically, aged MSCs display global DNA hypomethylation alongside increased promoter-specific methylation78 and abnormal histone modifications, including altered SETD2/H3K36me3 levels,79,80 which collectively compromise osteogenic potential. From a proteostatic perspective, aging is associated with autophagic dysfunction and markedly reduced autophagy in bone tissue,81,82 disrupting proteostasis networks83 and resulting in impaired osteoblast mineralization and delayed bone regeneration.84 Collectively, these alterations define the distinctive features of the aged osteo-immune microenvironment, with immune imbalance, inflammaging, and bone–immune axis dysregulation remaining the dominant pathological drivers.
Table 1 provides a systematic overview of the pathological basis of the aged bone–immune microenvironment.
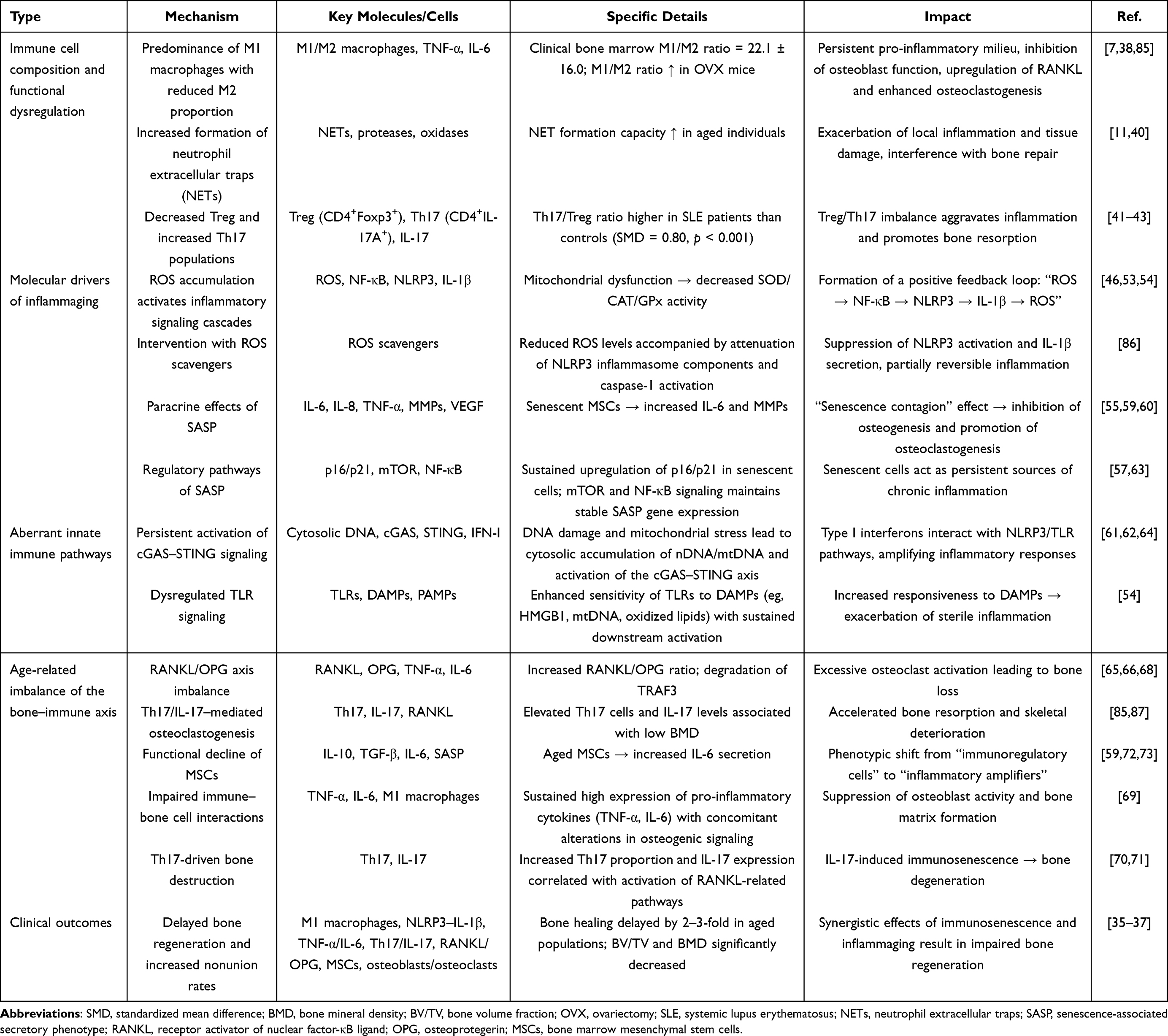 |
Table 1 Pathological Foundations of the Aged Osteo-Immune Microenvironment |
Mechanisms and Experimental Evidence Underlying the Regulation of the Aged Osteo-Immune Microenvironment by Black Phosphorus Nanomaterials
Targeting the three major pathological features of the aged osteo-immune microenvironment, black phosphorus (BP) nanomaterials demonstrate multifaceted and highly specific regulatory effects.
At the level of immune cell imbalance, BP effectively reverses macrophage polarization dysregulation in aged bone marrow. Both in vitro and in vivo studies show that PLGA/BP scaffolds significantly downregulate M1-associated markers, including TNF-α, IL-6, and IL-1β, while upregulating M2-associated markers such as IL-10, TGF-β, and CD206, resulting in a marked reduction in the M1/M2 ratio.32,33,39 By lowering oxidative stress, BP attenuates M1-polarizing signals, whereas its degradation products further promote M2-related gene expression through pathways such as STAT6, reinforcing this shift in macrophage phenotype.33 Moreover, BP indirectly restores Treg/Th17 balance by enhancing M2 polarization and suppressing inflammatory signaling, thereby reducing IL-17 and other pro-inflammatory mediators and inhibiting osteoclast activity.32,33,56,88,89 Collectively, these actions reprogram the bone microenvironment from a pro-inflammatory, destructive state toward an anti-inflammatory, reparative phenotype.
Beyond immune cell imbalance, BP plays a pivotal role in mitigating inflammaging. Through efficient scavenging of reactive oxygen species (ROS), improvement of mitochondrial function, and attenuation of inflammatory injury, BP counteracts age-related chronic inflammation.47,90 Mechanistically, BP interrupts the ROS–TXNIP–NLRP3 signaling cascade, suppressing NLRP3 activation,48,91 inflammasome-related cytokine release,49,92,93 and pathways associated with the senescence-associated secretory phenotype (SASP). Consequently, the secretion of pro-inflammatory cytokines such as IL-6, IL-8, and TNF-α, as well as matrix metalloproteinases (MMPs), is markedly reduced, leading to an overall improvement in the inflammatory milieu.56,94–98 In parallel, BP-mediated modulation of the immune microenvironment indirectly restrains excessive activation of the cGAS–STING pathway, further alleviating chronic inflammation associated with immunosenescence.99–102
Within the RANKL/OPG osteo-immune axis, BP exerts dual immunomodulatory and osteogenic effects. By downregulating pro-inflammatory cytokines such as TNF-α and IL-6 and promoting M2 macrophage polarization, BP indirectly reduces RANKL expression and suppresses osteoclast differentiation, thereby correcting osteo-immune imbalance.32,33 Concurrently, BP facilitates osteogenic differentiation of bone marrow mesenchymal stem cells (BMSCs), activates key osteogenic pathways including PI3K–AKT,32,33 and induces immunoreparative factors such as IL-10 and TGF-β,103 enabling a critical transition from tissue destruction to regeneration. Additionally, the controlled degradation of BP results in sustained release of phosphate ions (PO43−), which directly enhances bone matrix mineralization29 and activates osteogenic regulators such as Runx2 and BMP-2, thereby driving BMSC differentiation toward osteoblast lineages and achieving comprehensive modulation of the osteo-immune axis.104,105
Consistent with these mechanistic insights, BP has shown compelling therapeutic efficacy in experimental models. In an aged rat vascularized bone regeneration model, Wu et al reported that BP–GelMA hydrogel scaffolds reduced TNF-α expression by 60%.56 In a titanium implant study, Ma et al demonstrated that BP–HA-coated implants increased the bone formation area by 47% compared with pure hydroxyapatite coatings.106 From an immunomodulatory perspective, Jing et al summarized evidence that BP-based strategies increased bone mineral density by 32% while restoring the macrophage M2/M1 polarization ratio.107 In a mouse model of osteoarthritis, Lu et al observed a reduction exceeding 70% in IL-17A expression.47 Notably, Wu et al further demonstrated in an aged rat nonunion model that a BP–IL-4 co-delivery system, employing an “immunomodulation-first, osteogenesis-later” strategy, achieved a bone continuity formation rate of 93%, compared with 40% in controls.108 Given that the degradation kinetics of BP are tunable,109 Cai et al reported that AI-assisted, personalized 3D-printed BP scaffolds shortened the healing duration of aged bone nonunion by 35% in vivo,110,111 highlighting a promising translational avenue for improving bone injury and fracture repair in elderly populations.
Table 2 provides a comprehensive overview of the key regulatory mechanisms and representative experimental data supporting the role of black phosphorus nanomaterials in the immunosenescent bone microenvironment.
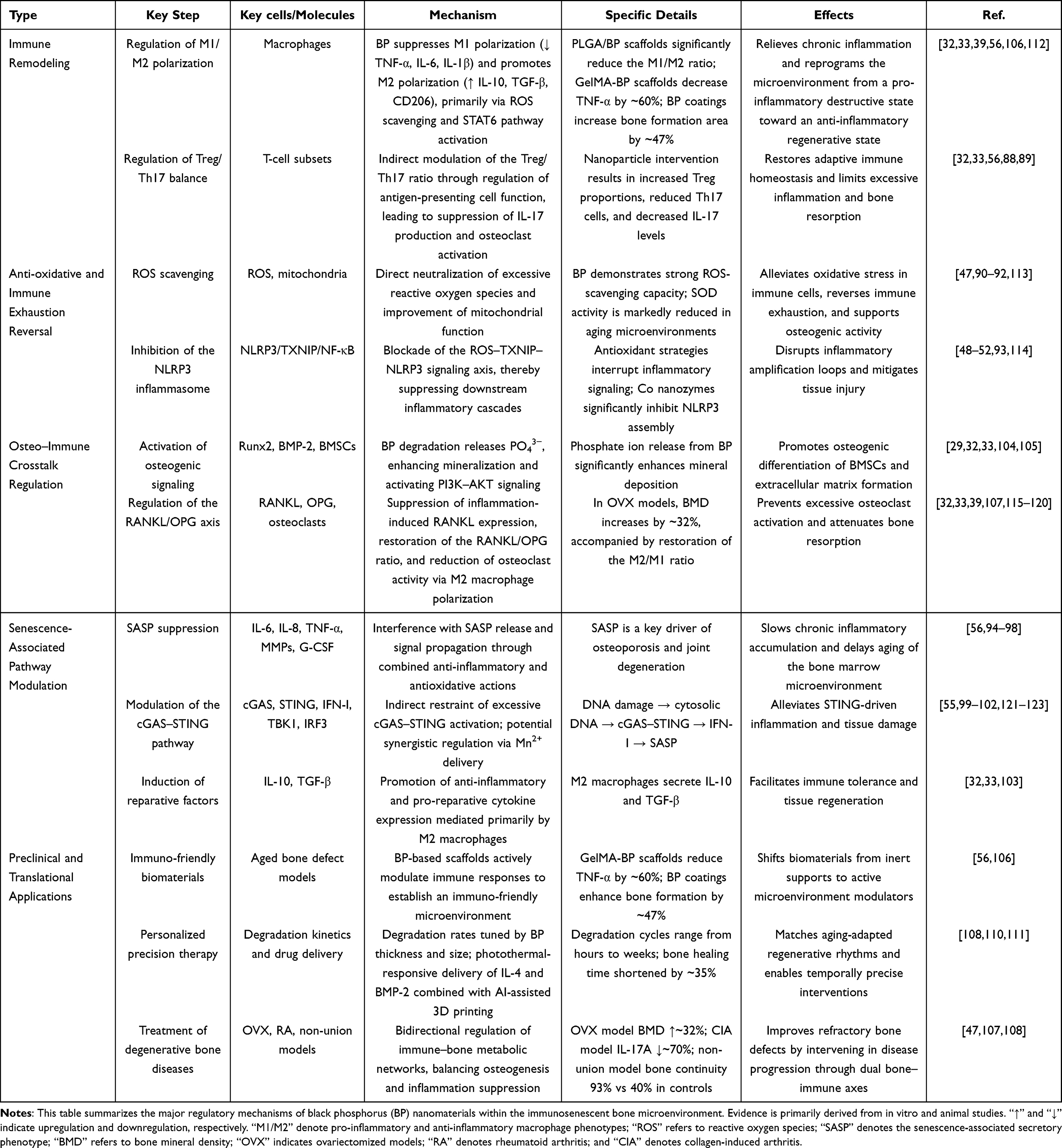 |
Table 2 Core Regulatory Mechanisms of Black Phosphorus (BP) Nanomaterials in the Immunosenescent Bone Microenvironment |
Challenges and Limitations
Despite its promising potential, the clinical application of black phosphorus (BP) remains constrained by three major challenges: the complexity of immune mechanisms within the aged osteoimmune microenvironment, the lack of long-term safety data, and unclear pathways for clinical translation (Table 3).
 |
Table 3 Key Challenges Associated with Black Phosphorus (BP)-Based Therapy |
Complexity of Immune Mechanisms in the Elderly
Current evidence is largely derived from animal models, with a notable absence of human studies. In particular, the regulatory effects of BP on immune cells such as macrophages and T cells within the aged osteoimmune microenvironment have not been systematically elucidated.33,124,125 Moreover, aging is associated with a reduced number of bone marrow mesenchymal stem cells (BMSCs), diminished proliferative capacity, and a pronounced tendency toward adipogenic differentiation.56 These aging-specific features substantially limit mechanistic interpretation and accurate prediction of BP therapeutic efficacy in elderly populations.
Lack of Long-Term Safety Data
Age-related declines in metabolic capacity may predispose elderly individuals to long-term BP accumulation in vivo. Although existing studies generally report favorable biocompatibility, they are predominantly based on short-term observations.107,132 In contrast, data on long-term in vivo degradation, toxicity, immune responses, and biodistribution remain limited, particularly in aged animal models.126–129 Accordingly, extended-duration (>1 year) toxicological and immunological investigations are urgently required to comprehensively characterize the safety profile of BP-based interventions in elderly populations.
Unclear Clinical Translation Pathways
Most available studies employ rodent models,32,47 whose physiological characteristics differ substantially from those of humans, thereby limiting direct clinical extrapolation. Furthermore, practical challenges, including difficulties in large-scale BP production, poor physicochemical stability, significant batch-to-batch variability, and insufficient standardization and controllability, continue to impede clinical translation.49,128,130,131
Future Perspectives
Over the next 5–10 years, increasing emphasis will be placed on elucidating BP–immune cell interactions, particularly involving macrophages and T cells, within the context of the aged osteoimmune microenvironment.33,124,125 Advanced spatial transcriptomics will be applied to resolve the spatial organization, signaling pathways, and interaction networks among immune cells, bone cells, and vascular cells,133,134 enabling construction of a three-dimensional BP-regulated “immune–bone–vascular” interaction atlas. In parallel, an “aged osteoimmune multi-omics database” will be established by integrating genomic, transcriptomic, proteomic, and metabolomic responses of elderly individuals under different BP-based interventions. This approach will facilitate identification of key biomarkers governing BP-mediated immunomodulatory efficacy.56 On this basis, rational surface modification strategies can be developed to fine-tune BP-induced immune responses, thereby supporting bone regeneration therapies in elderly patients.
During the same period, long-term (>1 year) in vivo toxicological and immunological evaluations, together with the establishment of aged or large-animal models,107,126–129,132 will enable more comprehensive safety assessments. Concurrently, standardized synthesis and characterization protocols will be implemented to enhance large-scale BP production and address issues of poor stability and pronounced batch-to-batch variability, ultimately accelerating clinical translation.32,47,49,128,130,131
Artificial intelligence (AI) is also expected to play an increasingly central role in BP research, spanning material design, therapeutic optimization, and safety evaluation. In intelligent nanomaterial development, AI-assisted machine learning and molecular modeling will support high-throughput screening and structural prediction, enabling optimization of BP nanoparticle size, surface modification, and drug-loading efficiency to improve stability, biocompatibility, and overall safety.135 At the therapeutic level, AI algorithms will be used to predict drug release kinetics, targeting performance, and in vivo distribution of BP-based delivery systems, thereby enabling intelligent responsiveness to the tumor microenvironment. Such strategies have the potential to enhance bone regeneration while achieving precise drug delivery and improved anticancer efficacy.135,136 In addition, AI-driven analyses of long-term follow-up and toxicological data will support early identification of potential safety risks, facilitate toxicity prediction and risk management, optimize clinical trial design, and ultimately improve clinical safety and therapeutic outcomes.90,135 Collectively, these advances are expected to accelerate the clinical translation and implementation of BP-based therapies.
Looking ahead, further investigations will focus on layer-dependent immune response mechanisms of BP in the aged bone microenvironment. Systematic comparisons of macrophage polarization induced by BP with different layer numbers will be conducted to establish structure–activity relationships linking layer number, particle size, and immune response.34,131 In parallel, continued attention will be given to interactions between BP and aging hallmarks, including its effects on mechanotransduction pathways in aged osteocytes and its antioxidant capacity to modulate epigenetic states of aged mesenchymal stem cells and autophagy–proteostasis networks, enabling more quantitative and precise evaluations.75,79,83 Finally, machine learning approaches may be employed to construct temporal coupling models between BP degradation kinetics and dynamic changes in the aged osteoimmune microenvironment. When integrated with immunosenescence indicators in elderly patients, these models could support individualized response prediction systems, advancing precision therapy and maximizing the beneficial effects of BP.110,135
Table 3 summarizes the three major challenges associated with BP-based therapies and the corresponding strategic solutions.
Data Sharing Statement
The present study did not involve the generation or analysis of any datasets.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors declare no competing interests.
References
1. Dudinskaya E, Brailova N, Kuznetsova V, et al. Osteoporosis in the elderly. Osteoporosis and Bone Diseases. 2020;22(3):34–16. doi:10.14341/osteo12352
2. Zanker J, Duque G. Osteoporosis in older persons: old and new players. J Am Geriatr Soc. 2019;67(4):831–840. doi:10.1111/jgs.15716
3. Cauley JA. Osteoporosis: fracture epidemiology update 2016. Curr Opin Rheumatol. 2017;29(2):150–156. doi:10.1097/BOR.0000000000000365
4. Meng S, Tong M, Yu Y, et al. The prevalence of osteoporotic fractures in the elderly in China: a systematic review and meta-analysis. J Orthop Surg Res. 2023;18(1):536. doi:10.1186/s13018-023-04030-x
5. Bouvard B, Annweiler C, Legrand E. Osteoporosis in older adults. Joint Bone Spine. 2021;88(3):105135. doi:10.1016/j.jbspin.2021.105135
6. Frase D, Lee C, Nachiappan C, et al. The inflammatory contribution of b-lymphocytes and neutrophils in progression to osteoporosis. Cells. 2023;12(13):1744. doi:10.3390/cells12131744
7. Kushioka J, Chow SK, Toya M, et al. Bone regeneration in inflammation with aging and cell-based immunomodulatory therapy. Inflamm Regen. 2023;43(1):29. doi:10.1186/s41232-023-00279-1
8. Lencel P, Magne D. Inflammaging: the driving force in osteoporosis? Med Hypotheses. 2011;76(3):317–321. doi:10.1016/j.mehy.2010.09.023
9. Li C, Xiao Y, Sun Y, et al. Senescent immune cells release grancalcin to promote skeletal aging. Cell Metab. 2021;33(10):1957–1973.e6. doi:10.1016/j.cmet.2021.08.009
10. Saxena Y, Routh S, Mukhopadhaya A. Immunoporosis: role of innate immune cells in osteoporosis. Front Immunol. 2021;687037. doi:10.3389/fimmu.2021.687037
11. Zhang W, Gao R, Rong X, et al. Immunoporosis: role of immune system in the pathophysiology of different types of osteoporosis[J]. Front Endocrinol. 2022;13:965258. doi:10.3389/fendo.2022.965258
12. Zhou F, Wang Z, Zhang G, et al. Immunosenescence and inflammaging: conspiracies against alveolar bone turnover. Oral Dis. 2024;30(4):1806–1817. doi:10.1111/odi.14642
13. Chen Y, Jia L, Han T, et al. Osteoporosis treatment: current drugs and future developments. Front Pharmacol. 2024;15:1456796. doi:10.3389/fphar.2024.1456796
14. He C, He P, Ou Y, et al. Rectifying the crosstalk between the skeletal and immune systems improves osteoporosis treatment by core-shell nanocapsules. ACS Nano. 2025;19(5):5549–5567. doi:10.1021/acsnano.4c14728
15. Li Q, Tian C, Liu X, et al. Anti-inflammatory and antioxidant traditional Chinese Medicine in treatment and prevention of osteoporosis. Front Pharmacol. 2023;14:1203767. doi:10.3389/fphar.2023.1203767
16. Luo Y, Liu H, Chen M, et al. Immunomodulatory nanomedicine for osteoporosis: current practices and emerging prospects. Acta Biomater. 2024;179:13–35. doi:10.1016/j.actbio.2024.03.011
17. Reid IR, Billington EO. Drug therapy for osteoporosis in older adults[J]. Lancet. 2022;399(10329):1080–1092. doi:10.1016/S0140-6736(21)02646-5
18. Wan S, Chen Y, Huang C, et al. Scalable ultrastrong MXene films with superior osteogenesis. Nature. 2024;634(8036):1103–1110. doi:10.1038/s41586-024-08067-8
19. Yu F, Zhao X, Zhang S, et al. Regulation of T Cell Glycosylation by MXene/β-TCP Nanocomposite for Enhanced Mandibular Bone Regeneration. Adv Healthc Mater. 2025;14(6):e2404015. doi:10.1002/adhm.202404015
20. Zhang S, Wang L, Feng Z, et al. Engineered mxene biomaterials for regenerative medicine. ACS Nano. 2025;19(10):9590–9635. doi:10.1021/acsnano.4c16136
21. Chen D, Liang Z, Su Z, et al. Selenium-doped mesoporous bioactive glass regulates macrophage metabolism and polarization by scavenging ros and promotes bone regeneration in vivo. ACS Appl Mater Interfaces. 2023;15(29):34378–34396. doi:10.1021/acsami.3c03446
22. Qiu H, Xiong H, Zheng J, et al. Sr-incorporated bioactive glass remodels the immunological microenvironment by enhancing the mitochondrial function of macrophage via the pi3k/akt/mtor signaling pathway. ACS Biomater Sci Eng. 2024;10(6):3923–3934. doi:10.1021/acsbiomaterials.4c00228
23. Zhao M, Yang Q, Zhang S, et al. Enhancing bone regeneration with a novel bioactive glass-functionalized polyetheretherketone scaffold by regulating the immune microenvironment. Smart Materials in Medicine. 2023. doi:10.1016/j.smaim.2023.09.002
24. Li J, Wen J, Li B, et al. Valence state manipulation of cerium oxide nanoparticles on a titanium surface for modulating cell fate and bone formation. Adv Sci. 2018;5(2):1700678. doi:10.1002/advs.201700678
25. Xiang J, Li J, He J, et al. Cerium oxide nanoparticle modified scaffold interface enhances vascularization of bone grafts by activating calcium channel of mesenchymal stem cells. ACS Appl Mater Interfaces. 2016;8(7):4489–4499. doi:10.1021/acsami.6b00158
26. Wei F, Neal CJ, Sakthivel TS, et al. A novel approach for the prevention of ionizing radiation-induced bone loss using a designer multifunctional cerium oxide nanozyme. Bioact Mater. 2023;21:547–565. doi:10.1016/j.bioactmat.2022.09.011
27. Li L, Yu Y, Ye GJ, et al. Black phosphorus field-effect transistors. Nat Nanotechnol. 2014;9(5):372–377. doi:10.1038/nnano.2014.35
28. Chen W, Ouyang J, Liu H, et al. Black phosphorus nanosheet-based drug delivery system for synergistic photodynamic/photothermal/chemotherapy of cancer. Adv Mater. 2017;29(5). doi:10.1002/adma.201603864
29. Huang K, Wu J, Gu Z. Black phosphorus hydrogel scaffolds enhance bone regeneration via a sustained supply of calcium-free phosphorus. ACS Appl Mater Interfaces. 2019;11(3):2908–2916. doi:10.1021/acsami.8b21179
30. Tong L, Liao Q, Zhao Y, et al. Near-infrared light control of bone regeneration with biodegradable photothermal osteoimplant[J]. Biomaterials. 2019;193:1–11. doi:10.1016/j.biomaterials.2018.12.008
31. Wang Z, Zhao J, Tang W, et al. Multifunctional nanoengineered hydrogels consisting of black phosphorus nanosheets upregulate bone formation. Small. 2019;15(41):e1901560. doi:10.1002/smll.201901560
32. Long J, Yao Z, Zhang W, et al. Regulation of osteoimmune microenvironment and osteogenesis by 3d-printed plag/black phosphorus scaffolds for bone regeneration. Adv Sci. 2023;10(28):e2302539. doi:10.1002/advs.202302539
33. Zou Y, Xie Q, Lin J, et al. Immunomodulatory effects and mechanisms of two-dimensional black phosphorus on macrophage polarization and bone regeneration. Int J Nanomedicine. 2025;20:4337–4355. doi:10.2147/IJN.S508309
34. Cheng L, Cai Z, Zhao J, et al. Black phosphorus-based 2D materials for bone therapy. Bioact Mater. 2020;5(4):1026–1043. doi:10.1016/j.bioactmat.2020.06.007
35. Li D, Li X, Zhang J, et al. The immunomodulatory effect of IL-4 accelerates bone substitute material-mediated osteogenesis in aged rats via NLRP3 inflammasome inhibition. Front Immunol. 2023;14:1121549. doi:10.3389/fimmu.2023.1121549
36. Newman H, Shih YV, Varghese S. Resolution of inflammation in bone regeneration: from understandings to therapeutic applications. Biomaterials. 2021;277:121114. doi:10.1016/j.biomaterials.2021.121114
37. Tchouto MN, Bucher CH, Mess A, et al. Pronounced impairment of B cell differentiation during bone regeneration in adult immune experienced mice. Front Immunol. 2025;16:1511902. doi:10.3389/fimmu.2025.1511902
38. Butler JJ, Dankert JF, Keller LE, et al. Assessment of the monocyte subpopulations and m1/m2 macrophage ratio in concentrated bone marrow aspirate. Cartilage. 2024:19476035241304308. doi:10.1177/19476035241304308.
39. Qiu M, Tulufu N, Tang G, et al. Black phosphorus accelerates bone regeneration based on immunoregulation. Adv Sci. 2024;11(1):e2304824. doi:10.1002/advs.202304824
40. Ortmann W, Kolaczkowska E. Age is the work of art? Impact of neutrophil and organism age on neutrophil extracellular trap formation. Cell Tissue Res. 2018;371(3):473–488. doi:10.1007/s00441-017-2751-4
41. Schmitt V, Rink L, Uciechowski P. The Th17/Treg balance is disturbed during aging. Exp Gerontol. 2013;48(12):1379–1386. doi:10.1016/j.exger.2013.09.003
42. Zhu L, Hua F, Ding W, et al. The correlation between the Th17/Treg cell balance and bone health. Immun Ageing. 2020;17:30. doi:10.1186/s12979-020-00202-z
43. Huang J, Li X, Zhu Q, et al. Imbalance of Th17 cells, Treg cells and associated cytokines in patients with systemic lupus erythematosus: a meta-analysis[J]. Front Immunol. 2024;15:1425847. doi:10.3389/fimmu.2024.1425847
44. Geng L, Tang X, Wang S, et al. Reduced let-7f in bone marrow-derived mesenchymal stem cells triggers treg/th17 imbalance in patients with systemic lupus erythematosus. Front Immunol. 2020;11:233. doi:10.3389/fimmu.2020.00233
45. Lan H, Qiu W, Wu J, et al. Formononetin reverses Treg/Th17 imbalance in immune-mediated bone marrow failure mice by regulating the PI3K/Akt signaling pathway. Chin Med. 2024;19(1):55. doi:10.1186/s13020-024-00919-9
46. Salminen A, Ojala J, Kaarniranta K, et al. Mitochondrial dysfunction and oxidative stress activate inflammasomes: impact on the aging process and age-related diseases. Cell Mol Life Sci. 2012;69(18):2999–3013. doi:10.1007/s00018-012-0962-0
47. Lu H, Wei J, Liu K, et al. Radical-scavenging and subchondral bone-regenerating nanomedicine for osteoarthritis treatment. ACS Nano. 2023;17(6):6131–6146. doi:10.1021/acsnano.3c01789
48. Abais JM, Xia M, Zhang Y, et al. Redox regulation of NLRP3 inflammasomes: ROS as trigger or effector?. Antioxid Redox Signal. 2015;22(13):1111–1129. doi:10.1089/ars.2014.5994
49. An D, Fu J, Xie Z, et al. Progress in the therapeutic applications of polymer-decorated black phosphorus and black phosphorus analog nanomaterials in biomedicine. J Mater Chem B. 2020;8(32):7076–7120. doi:10.1039/d0tb00824a
50. Russell-Guzmán J, Silva LA, Cadagan C, et al. Activation of the ROS/TXNIP/NLRP3 pathway disrupts insulin-dependent glucose uptake in skeletal muscle of insulin-resistant obese mice. Free Radic Biol Med. 2024;222:187–198. doi:10.1016/j.freeradbiomed.2024.06.011
51. Xu X, Zhang L, Ye X, et al. Nrf2/ARE pathway inhibits ROS-induced NLRP3 inflammasome activation in BV2 cells after cerebral ischemia reperfusion. Inflamm Res. 2018;67(1):57–65. doi:10.1007/s00011-017-1095-6
52. Yin Y, Zhou Z, Liu W, et al. Vascular endothelial cells senescence is associated with NOD-like receptor family pyrin domain-containing 3 (NLRP3) inflammasome activation via reactive oxygen species (ROS)/thioredoxin-interacting protein (TXNIP) pathway. Int J Biochem Cell Biol. 2017;84:22–34. doi:10.1016/j.biocel.2017.01.001
53. Dominic A, Le N, Takahashi M. Loop between NLRP3 inflammasome and reactive oxygen species. Antioxid Redox Signal. 2022;36(10–12):784–796. doi:10.1089/ars.2020.8257
54. Ramos-Tovar E, Muriel P. Molecular mechanisms that link oxidative stress, inflammation, and fibrosis in the liver. Antioxidants. 2020;9(12). doi:10.3390/antiox9121279
55. Wang B, Han J, Elisseeff JH, et al. The senescence-associated secretory phenotype and its physiological and pathological implications. Nat Rev Mol Cell Biol. 2024;25(12):958–978. doi:10.1038/s41580-024-00727-x
56. Wu T, Ren M, Li Y, et al. Bioelectrically reprogramming hydrogels rejuvenate vascularized bone regeneration in senescence. Adv Healthc Mater. 2025;14(6):e2403837. doi:10.1002/adhm.202403837
57. Lopes-Paciencia S, Saint-Germain E, Rowell M, et al. The senescence-associated secretory phenotype and its regulation. Cytokine. 2019;117:15–22. doi:10.1016/j.cyto.2019.01.013
58. Cuollo L, Antonangeli F, Santoni A, et al. The Senescence-Associated Secretory Phenotype (SASP) in the challenging future of cancer therapy and age-related diseases. Biology. 2020;9(12). doi:10.3390/biology9120485
59. Bai J, Wang Y, Wang J, et al. Irradiation-induced senescence of bone marrow mesenchymal stem cells aggravates osteogenic differentiation dysfunction via paracrine signalin. Am J Physiol Cell Physiol. 2020;318(5):C1005–C1017. doi:10.1152/ajpcell.00520.2019
60. Xu L, Wang Y, Wang J, et al. Radiation-induced osteocyte senescence alters bone marrow mesenchymal stem cell differentiation potential via paracrine signaling. Int J Mol Sci. 2021;22(17). doi:10.3390/ijms22179323
61. Decout A, Katz JD, Venkatraman S, et al. The cGAS-STING pathway as a therapeutic target in inflammatory diseases. Nat Rev Immunol. 2021;21(9):548–569. doi:10.1038/s41577-021-00524-z
62. Aarreberg LD, Esser-Nobis K, Driscoll C, et al. Interleukin-1β Induces mtDNA release to activate innate immune signaling via cGAS-STING. Mol Cell. 2019;74(4):801–815.e6. doi:10.1016/j.molcel.2019.02.038
63. Song J, Bae Y. CK2 down-regulation increases the expression of senescence-associated secretory phenotype factors through nf-κb activation. Int J Mol Sci. 2021;22(1). doi:10.3390/ijms22010406
64. Schmitz CRR, Maurmann RM, Guma FTCR, et al. cGAS-STING pathway as a potential trigger of immunosenescence and inflammaging. Front Immunol. 2023;14:1132653. doi:10.3389/fimmu.2023.1132653
65. Kitaura H, Marahleh A, Ohori F, et al. Osteocyte-related cytokines regulate osteoclast formation and bone resorption. Int J Mol Sci. 2020;21(14). doi:10.3390/ijms21145169
66. Zha L, He L, Liang Y, et al. TNF-α contributes to postmenopausal osteoporosis by synergistically promoting RANKL-induced osteoclast formation. Biomed Pharmacother. 2018;102:369–374. doi:10.1016/j.biopha.2018.03.080
67. Weitzmann MN. The Role of Inflammatory Cytokines, the RANKL/OPG Axis, and the Immunoskeletal Interface in physiological bone turnover and osteoporosis. Scientifica (Cairo). 2013;2013:125705. doi:10.1155/2013/125705
68. Yao Z, Lei W, Duan R, et al. RANKL cytokine enhances TNF-induced osteoclastogenesis independently of TNF receptor associated factor (TRAF) 6 by degrading TRAF3 in osteoclast precursors. J Biol Chem. 2017;292(24):10169–10179. doi:10.1074/jbc.M116.771816
69. Qu X, Xu G, Hou X, et al. M1 Macrophage-Derived Interleukin-6 promotes the osteogenic differentiation of ligamentum flavum cells. Spine. 2022;47(15):E527–E535. doi:10.1097/BRS.0000000000004319
70. Pöllinger B, Junt T, Metzler B, et al. Th17 cells, not IL-17+ γδ T cells, drive arthritic bone destruction in mice and humans. J Immunol. 2011;186(4):2602–2612. doi:10.4049/jimmunol.1003370
71. Faust HJ, Zhang H, Han J, et al. IL-17 and immunologically induced senescence regulate response to injury in osteoarthritis. J Clin Invest. 2020;130(10):5493–5507. doi:10.1172/JCI134091
72. Peng X, Zhou X, Yin Y, et al. Inflammatory microenvironment accelerates bone marrow mesenchymal stem cell aging. Front Bioeng Biotechnol. 2022;10:870324. doi:10.3389/fbioe.2022.870324
73. Gnani D, Crippa S, Volpe LD, et al. An early-senescence state in aged mesenchymal stromal cells contributes to hematopoietic stem and progenitor cell clonogenic impairment through the activation of a pro-inflammatory program. Aging Cell. 2019;18(3):e12933. doi:10.1111/acel.12933
74. Cui J, Shibata Y, Zhu T, et al. Osteocytes in bone aging: advances, challenges, and future perspectives. Ageing Res Rev. 2022;77:101608. doi:10.1016/j.arr.2022.101608
75. Hemmatian H, Bakker AD, Klein-Nulend J, et al. Aging, osteocytes, and mechanotransduction. Curr Osteoporos Rep. 2017;15(5):401–411. doi:10.1007/s11914-017-0402-z
76. Hu YJ, Wu X, Wang F, et al. Piezo1-mediated mechanotransduction controls osteocyte maturation and dendrite development via a YAP-CCN-Src signaling axis. Nat Commun. 2025;16(1):10859. doi:10.1038/s41467-025-65636-9
77. Kwon Y. YAP/TAZ as molecular targets in skeletal muscle atrophy and osteoporosis. Aging Dis. 2024. doi:10.14336/AD.2024.0306
78. Cakouros D, Gronthos S.The Changing Epigenetic Landscape of Mesenchymal Stem/Stromal Cells During Aging.Bone.2020;Vol. 137.115440. doi:10.1016/j.bone.2020.115440
79. Cakouros D, Gronthos S. Epigenetic regulators of mesenchymal stem/stromal cell lineage determination. Curr Osteoporos Rep. 2020;18(5):597–605. doi:10.1007/s11914-020-00616-0
80. Smith N, Shirazi S, Cakouros D, et al. Impact of environmental and epigenetic changes on mesenchymal stem cells during aging. Int J Mol Sci. 2023;24(7). doi:10.3390/ijms24076499
81. Xu Y, Sun B, Wang H, et al. Autophagy regulates age-related delayed jawbone regeneration and decreased osteoblast osteogenesis by degrading FABP3. FASEB J. 2024;38(14):e23824. doi:10.1096/fj.202400549RR
82. Yin X, Zhou C, Li J, et al. Autophagy in bone homeostasis and the onset of osteoporosis. Bone Res. 2019;7:28. doi:10.1038/s41413-019-0058-7
83. Ma C, Yu R, Li J, et al. Targeting proteostasis network in osteoporosis: pathological mechanisms and therapeutic implications. Ageing Res Rev. 2023;90:102024. doi:10.1016/j.arr.2023.102024
84. Nollet M, Santucci-Darmanin S, Breuil V, et al. Autophagy in osteoblasts is involved in mineralization and bone homeostasis. Autophagy. 2014;10(11):1965–1977. doi:10.4161/auto.36182
85. Dou C, Ding N, Zhao C, et al. Estrogen Deficiency-Mediated M2 Macrophage Osteoclastogenesis Contributes to M1/M2 ratio alteration in ovariectomized osteoporotic mice. J Bone Miner Res. 2018;33(5):899–908. doi:10.1002/jbmr.3364
86. Sarkar S, Malovic E, Harishchandra DS, et al. Mitochondrial impairment in microglia amplifies NLRP3 inflammasome proinflammatory signaling in cell culture and animal models of Parkinson’s disease. NPJ Parkinsons Dis. 2017;3:30. doi:10.1038/s41531-017-0032-2
87. Tan J, Dai A, Pan L, et al. Inflamm-aging-related cytokines of il-17 and ifn-γ accelerate osteoclastogenesis and periodontal destruction. J Immunol Res. 2021;2021:9919024. doi:10.1155/2021/9919024
88. Li J, Li M, Zhang C, et al. Active targeting microemulsion-based thermosensitive hydrogel against periodontitis by reconstructing Th17/Treg homeostasis via regulating ROS-macrophages polarization cascade[J]. Int J Pharm. 2024;659:124263. doi:10.1016/j.ijpharm.2024.124263
89. Yang X, Zhou F, Yuan P, et al. T cell-depleting nanoparticles ameliorate bone loss by reducing activated T cells and regulating the Treg/Th17 balance. Bioact Mater. 2021;6(10)):3150–3163. doi:10.1016/j.bioactmat.2021.02.034
90. Kong N, Ji X, Wang J, et al. ROS-mediated selective killing effect of black phosphorus: mechanistic understanding and its guidance for safe biomedical applications. Nano Lett. 2020;20(5):3943–3955. doi:10.1021/acs.nanolett.0c01098
91. Minutoli L, Puzzolo D, Rinaldi M, et al. ROS-Mediated NLRP3 inflammasome activation in brain, heart, kidney, and testis ischemia/reperfusion injury. Oxid Med Cell Longev. 2016;2016:2183026. doi:10.1155/2016/2183026
92. Pagliaro P, Penna C. Inhibitors of NLRP3 inflammasome in ischemic heart disease: focus on functional and redox aspects. Antioxidants. 2023;12(7). doi:10.3390/antiox12071396
93. Wan L, Bai X, Zhou Q, et al. The advanced glycation end-products (AGEs)/ROS/NLRP3 inflammasome axis contributes to delayed diabetic corneal wound healing and nerve regeneration. Int J Biol Sci. 2022;18(2):809–825. doi:10.7150/ijbs.63219
94. Alavimanesh S, Jazi NN, Choubani M, et al. Cellular senescence in the tumor with a bone niche microenvironment: friend or foe? Clin Exp Med. 2025;25(1):44. doi:10.1007/s10238-025-01564-8
95. Chandra A, Rajawat J. Skeletal aging and osteoporosis: mechanisms and therapeutics. Int J Mol Sci. 2021;22(7). doi:10.3390/ijms22073553
96. Fang C, Liu B, Wan M. “Bone-SASP” in Skeletal Aging[J]. Calcif Tissue Int. 2023;113(1):68–82. doi:10.1007/s00223-023-01100-4
97. Farr JN, Khosla S. Cellular senescence in bone. Bone. 2019;121:121–133. doi:10.1016/j.bone.2019.01.015
98. Zhu R, Wan H, Yang H, et al. The role of senescence-associated secretory phenotype in bone loss. Front Cell Dev Biol. 2022;10:841612. doi:10.3389/fcell.2022.841612
99. Chen X, Xu Z, Li T, et al. Nanomaterial-encapsulated STING agonists for immune modulation in cancer therapy. Biomark Res. 2024;12(1):2. doi:10.1186/s40364-023-00551-z
100. Lawrence M, Goyal A, Pathak S, et al. Cellular senescence and inflammaging in the bone: pathways, genetics, anti-aging strategies and interventions. Int J Mol Sci. 2024;25(13). doi:10.3390/ijms25137411
101. Loo TM, Miyata K, Tanaka Y, et al. Cellular senescence and senescence-associated secretory phenotype via the cGAS-STING signaling pathway in cancer. Cancer Sci. 2020;111(2):304–311. doi:10.1111/cas.14266
102. Ohtani N. The roles and mechanisms of senescence-associated secretory phenotype (SASP): can it be controlled by senolysis? Inflamm Regen. 2022;42(1):11. doi:10.1186/s41232-022-00197-8
103. Li Q, Wang R, Xue J, et al. ZIF-8-modified black phosphorus nanosheets incorporated into injectable dual-component hydrogels for enhanced photothermal antibacterial and osteogenic activities. ACS Appl Mater Interfaces. 2024;16(25):32058–32077. doi:10.1021/acsami.4c05298
104. Li Z, Zhang X, Ouyang J, et al. Ca2+-supplying black phosphorus-based scaffolds fabricated with microfluidic technology for osteogenesis. Bioact Mater. 2021;6(11):4053–4064. doi:10.1016/j.bioactmat.2021.04.014
105. Lu N, Wang X, Shi W, et al. Black phosphorus nanoparticles promote osteogenic differentiation of emscs through upregulated TG2 expression. Nanoscale Res Lett. 2021;16(1):154. doi:10.1186/s11671-021-03610-2
106. Ma S, Sun R, Wang Y, et al. Improving osseointegration and antimicrobial properties of titanium implants with black phosphorus nanosheets-hydroxyapatite composite coatings for vascularized bone regeneration. J Biomed Mater Res B Appl Biomater. 2024;112(4):e35403. doi:10.1002/jbm.b.35403
107. Jing X, Xiong Z, Lin Z, et al. The application of black phosphorus nanomaterials in bone tissue engineering. Pharmaceutics. 2022;14(12). doi:10.3390/pharmaceutics14122634
108. Wu A, Ma G, Chen Y, et al. Improved Black Phosphorus Nanocomposite Hydrogel for Bone Defect Repairing: mechanisms for Advancing Osteogenesis. Adv Healthc Mater. 2025;14(7):e2404934. doi:10.1002/adhm.202404934
109. Xu H, Liu X, Park S, et al. Size-dependent osteogenesis of black phosphorus in nanocomposite hydrogel scaffolds[J]. J Biomed Mater Res A. 2022;110(8):1488–1498. doi:10.1002/jbm.a.37382
110. Cai Z, Chen Z, Tang Y, et al. Bioinspired bone seed 3d-printed scaffold via trapping black phosphorus nanosheet for bone regeneration. Small Sci. 2024;4(6):2300357. doi:10.1002/smsc.202300357
111. Wu N, Li J, Li X, et al. 3D printed biopolymer/black phosphorus nanoscaffolds for bone implants: a review. Int J Biol Macromol. 2024;279(Pt 3):135227. doi:10.1016/j.ijbiomac.2024.135227
112. Song Q, Zhang Y, Hu H, et al. Multifunctional hydrogel with synergistic reactive oxygen species scavenging and macrophage polarization-induced osteo-immunomodulation for enhanced bone regeneration. ACS Appl Mater Interfaces. 2025;17(27):38985–39001. doi:10.1021/acsami.5c08737
113. Iyer SS, He Q, Janczy JR, et al. Mitochondrial cardiolipin is required for Nlrp3 inflammasome activation. Immunity. 2013;39(2):311–323. doi:10.1016/j.immuni.2013.08.001
114. Chen Z, Chen P, Zhu Y, et al. 2D cobalt oxyhydroxide nanozymes inhibit inflammation by targeting the NLRP3 inflammasome. Advanced Functional Materials. 2023. doi:10.1002/adfm.202214693
115. Boyce BF, Xing L. The RANKL/RANK/OPG pathway. Curr Osteoporos Rep. 2007;5(3):98–104. doi:10.1007/s11914-007-0024-y
116. Boyce BF, Xing L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch Biochem Biophys. 2008;473(2):139–146. doi:10.1016/j.abb.2008.03.018
117. D DL-O, Barrena-Blázquez S, Jiménez-álvarez L, et al. The RANK-RANKL-OPG System: a multifaceted regulator of homeostasis, immunity, and cancer. Medicina. 2023;59(10). doi:10.3390/medicina59101752
118. Ono T, Hayashi M, Sasaki F, et al. RANKL biology: bone metabolism, the immune system, and beyond. Inflamm Regen. 2020;40:2. doi:10.1186/s41232-019-0111-3
119. Takegahara N, Kim H, Choi Y. RANKL biology. Bone. 2022;159:116353. doi:10.1016/j.bone.2022.116353
120. Udagawa N, Koide M, Nakamura M, et al. Osteoclast differentiation by RANKL and OPG signaling pathways. J Bone Miner Metab. 2021;39(1):19–26. doi:10.1007/s00774-020-01162-6
121. Gao M, Xie Y, Lei K, et al. A Manganese Phosphate Nanocluster Activates the cGAS‐STING Pathway for Enhanced Cancer Immunotherapy. Advanced Therapeutics. 2021;4. doi:10.1002/adtp.202100065
122. Ling K, Zheng J, Jiang X, et al. Mn2+/CpG oligodeoxynucleotides codecorated black phosphorus nanosheet platform for enhanced antitumor potency in multimodal therapy. ACS Nano. 2024;18(4):2841–2860. doi:10.1021/acsnano.3c07123
123. Zhou L, Huang Y, Wu Y, et al. Nanoparticle targeting cGAS-STING signaling in disease therapy. Nano Research. 2024. doi:10.1007/s12274-024-6714-x
124. Dong W, Wang H, Liu H, et al. Potential of black phosphorus in immune-based therapeutic strategies. Bioinorg Chem Appl. 2022;2022:3790097. doi:10.1155/2022/3790097
125. Zhuang C, Sun R, Zhang Y, et al. Treatment of rheumatoid arthritis based on the inherent bioactivity of black phosphorus nanosheets. Aging Dis. 2024;16(3):1652–1673. doi:10.14336/AD.2024.0319
126. Anju S, Ashtami J, Mohanan PV. Black phosphorus, a prospective graphene substitute for biomedical applications. Mater Sci Eng C Mater Biol Appl. 2019;97:978–993. doi:10.1016/j.msec.2018.12.146
127. Qian X, Gu Z, Chen Y. Two-dimensional black phosphorus nanosheets for theranostic nanomedicine. Materials Horizons. 2017;4:800–816. doi:10.1039/c7mh00305f
128. Smith LS, Haidari H, Amsalu A, et al. Black phosphorus nanoflakes: an emerging nanomaterial for clinical wound management and biomedical applications. Int J Mol Sci. 2024;25(23). doi:10.3390/ijms252312824
129. Xie H, Chen Z, Zhang N, et al. A review of recent advances in the stability, efficacy, and biosafety of black phosphorus-based drug delivery. Journal of Materials Science. 2024. doi:10.1007/s10853-024-09850-8
130. Liu W, Dong A, Wang B, et al. Current advances in black phosphorus-based drug delivery systems for cancer therapy. Adv Sci. 2021;8(5):2003033. doi:10.1002/advs.202003033
131. Luo M, Fan T, Zhou Y, et al. 2D Black Phosphorus–Based Biomedical Applications[J]. Advanced Functional Materials. 2019. doi:10.1002/adfm.201808306
132. Sun T, Li C, Luan J, et al. Black phosphorus for bone regeneration: mechanisms involved and influencing factors. Mater Today Bio. 2024;28:101211. doi:10.1016/j.mtbio.2024.101211
133. Ferreira RM, Sabo AR, Winfree S, et al. Integration of spatial and single-cell transcriptomics localizes epithelial cell-immune cross-talk in kidney injury. JCI Insight. 2021;6(12). doi:10.1172/jci.insight.147703
134. Xiao X, Juan C, Drennon T, et al. Spatial transcriptomic interrogation of the murine bone marrow signaling landscape. Bone Res. 2023;11(1):59. doi:10.1038/s41413-023-00298-1
135. Sun L, Liu H, Ye Y, et al. Smart nanoparticles for cancer therapy. Signal Transduct Target Ther. 2023;8(1)):418. doi:10.1038/s41392-023-01642-x
136. Choi JR, Yong KW, Choi JY, et al. Black phosphorus and its biomedical applications. Theranostics. 2018;8(4):1005–1026. doi:10.7150/thno.22573
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.